
production and characterization of fuel briquette from mango

Food Chemistry 130 (2012) 58–66
Contents lists available at ScienceDirect
Food Chemistry
journal homepage: www.elsevier .com/locate / foodchem
Geographic variation in the flavour volatiles of Alphonso mango
Ram S. Kulkarni a, Hemangi G. Chidley a, Keshav H. Pujari b, Ashok P. Giri a, Vidya S. Gupta a,⇑a Plant Molecular Biology Unit, Division of Biochemical Sciences, National Chemical Laboratory, Pune 411 008, Indiab Dr. Balasaheb Sawant Konkan Krishi Vidyapeeth, Dapoli 415 712, India
a r t i c l e i n f o
Article history:Received 1 February 2011Received in revised form 12 May 2011Accepted 27 June 2011Available online 1 July 2011
Keywords:MangoPhenotypic plasticityRipeningVolatiles
0308-8146/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.foodchem.2011.06.053
⇑ Corresponding author. Tel.: +91 20 25902247; faxE-mail address: [email protected] (V.S. Gupta).
a b s t r a c t
Alphonso, one of the most popular cultivars of mango in India is known to exhibit geographic variation inthe flavour of ripe fruits. To get chemical insight into this difference, volatiles were studied in the ripeningfruits of Alphonso mangoes from three cultivation locations in India. Ripe fruits from Deogad had lowercontent of mono- and sesquiterpenes and higher content of lactones and furanones as compared to thefruits from Dapoli; whereas fruits from Vengurle had average quantities of these chemicals in comparisonwith Deogad and Dapoli fruits. This variation was clearly reflected as separate clustering of the localitiesin the Principal Component Analysis. The localities were indistinguishable from each other in terms ofraw fruit volatiles. This study exemplifies a case of phenotypic plasticity; since the plants chosen wereclonally propagated, such geographic variation in the volatiles can be attributed to varied abiotic condi-tions at these three localities.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
India stands first in the list of mango producing (39.1% of theworld production of 34.9 MMT) and exporting (23% of the worldexport of 1.2 MMT) countries, which contributes significantly toits foreign income and agro-rural development (http://fao-stat.fao.org/site/291/default.aspx). India is known as the countryof origin of mango and is blessed with thousands of cultivars of thisnational fruit (Salvi & Gunjate, 1988). Alphonso is the most popularmango cultivar, which is favoured mainly because of its highlyattractive flavour. Apart from this, Alphonso possesses many sig-nificant attributes such as attractive colour, ample sweetness,low fibre-containing pulp and long shelf life. Ripe Alphonso fruitsare popularly used in processed and canned foods. Similarly, theraw fruits of Alphonso are also used in food products, such as pick-les, tarts, curries and salads (Bawa, 2007).
Despite having so many virtues, cultivation of Alphonso in dif-ferent localities in India does not result in same quality of fruits.Because of this fact, Alphonso cultivation is concentrated in a700-km long, narrow coastal belt of western India, the Konkan re-gion. Even within this region, the fruits show conspicuous variationin their taste and flavour. Throughout Konkan, the growing prac-tice and post-harvest treatment of these mangoes is the same,wherein, after harvesting, the fruits are packed in hay and areimmediately transported to market. It takes about 15 days afterharvest for complete ripening. In spite of such consistent cultiva-tion and harvesting practices, the flavour of the ripe mangoes dif-
ll rights reserved.
: +91 20 25902648.
fers between the localities of cultivation. Because of this, Alphonsofruits from a specific region (Deogad) of Konkan are preferred forfresh table use as well as for processed food products, resultingin high market value for the fruits from this region.
Till now, effect of geographic location on mango fruit qualityhas been shown in Australian cultivars (Hassan, Irving, Dann,Coates, & Hofman, 2009; Hofman, Smith, Meiburg, & Giles, 1997),but these studies were not focused on the flavour volatiles, whichis one of the most important functional phenotypes of the fruits. Tounderstand chemical basis behind the flavour variation observed inAlphonso mango, we initiated a study of the volatiles through thedevelopmental and ripening stages of Alphonso fruits. As the firstpart, we studied the fruits from the middle Konkan region, Deogad,which is known to produce the best quality Alphonso. This analysisindicated that Alphonso fruits are rich in mono and sesquiterpenesand that the ripening stages are characterised by the unique pres-ence of lactones and furanones (Pandit, Kulkarni, et al., 2009). Inthe present study, we have extended this work towards the volatileanalysis of ripening stages of the Alphonso from three differentcultivation localities, each from north, middle and south Konkan.
2. Materials and methods
2.1. Plant material
Mature raw fruits of Alphonso mango were collected from theorchards of Konkan Krishi Vidyapeeth at Dapoli (N17�450
E73�110) and Deogad (N16�310 E73�200) and from a private orchardat Vengurle (N15�510 E73�390). For each of the three localities, fourplants were selected and from each of these plants at least 20 fruits

R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66 59
were used for the analysis to justify the statistical validity of theanalysis. These randomly chosen plants from the orchard were dis-tantly located (at least 30 m away) from each other and the fruitswere harvested from the branches at the periphery and at theheight range of 2–5 m to have the fruits exposed to the similarintensity of light. This practice was followed for all the plants fromall the three localities. After harvesting, fruits were put in the hay,carried to the laboratory and allowed to ripen at ambient temper-ature. Conventionally, ripening of Alphonso is determined based onthe period in terms of days after harvesting, change in skin colour(green to yellow-orange), smell (raw leafy to sweet fruity) andsoftness to touch. According to these indices the fruits of 15 daysafter harvest (15 DAH) are considered completely ripe (Bandyo-padhyay & Gholap, 1973). Moreover, our previous volatile analysisof ripening stages of Alphonso from one location, Deogad, has indi-cated 15 DAH as the completely ripe stage (Pandit, Kulkarni, et al.,2009). To span this whole period of ripening, at an interval of everyfive days, fruits were cut, immediately frozen in liquid nitrogenand stored at �80 �C until use, giving rise to four stages of ripen-ing: 0, 5, 10 and 15 DAH for each tree from each of the threelocalities.
2.2. Extraction of volatiles
Volatiles were extracted from the fruits at various ripeningstages employing the method which has been previously opti-mised with dichloromethane as a solvent in our laboratory (Pandit,Chidley, et al., 2009; Pandit, Kulkarni, et al., 2009). Briefly, extrac-tion was carried out from 2 g fruit mesocarp for 1 h in 10 mldichloromethane having 40 lg nonyl acetate as an internal stan-dard. Supernatant was decanted and dehydrated with sodium sul-phate. After carefully concentrating to 1 ml in a rotary evaporatorat ambient temperature (ca. 25 �C), the extracts were stored at�20 �C overnight to precipitate the fats which were removed onthe next day by centrifugation at 16,000g at 4 �C for 10 min. Theextracts were finally concentrated in a similar way to 50 ll.
2.3. GC–MS and GC-FID analysis
Gas chromatographic analysis was carried out on Clarus 500(Perkin Elmer, Waltham, MA) instrument with Rtx-5MS (RestekCorporation, Bellefonte, PA) capillary column (30 m � 0.32 mmi.d. � 0.25 lm film thickness). Column temperatures were pro-grammed from 40 �C for 5 min, raised to 180 �C at 5 �C/min fol-lowed by an increase till 280 �C at the rate of 20 �C/min and heldat 280 �C for 5 min. The rest of the chromatographic and massspectrometric conditions were the same as described earlier (Pan-dit, Kulkarni, et al., 2009). Compounds were identified by compar-ing acquired mass spectra with those of authentic standards andwith the help of the Wiley registry of mass spectral data (8th edi-tion). Identification was confirmed by comparison of the obtainedretention indices with those of the authentic standards and withthose reported in the literature. Quantification was carried out byinternal standard method, where concentrations of different vola-tiles were normalised with that of nonyl acetate.
2.4. Statistical analyses
For each stage of the ripening a minimum of two fruits of eachof the four trees from each of the three localities was used for twoindependent extractions while each extract was independentlyanalysed twice on the GC. Both ANOVA and principal componentanalysis were carried out using Systat statistical software (Version11, Richmond, CA). Fisher’s LSD test was carried out (p 6 0.05) byANOVA to compare the quantity of each compound and each class
of compounds with respect to various stages and cultivationlocalities.
3. Results
3.1. Detection of volatiles
Thirty-one volatile compounds were detected, out of which 20were quantitated in the fruits of four ripening stages from thethree different localities (Table 1). Compounds were classified intofour classes, viz., monoterpenes (13 compounds), sesquiterpenes (5compounds), lactones (8 compounds), furanones (2 compounds)and others (3 compounds). Monoterpenes were found in the high-est concentration (2.55 to 374 lg g�1) in all the 48 tissues ana-lysed. (Z)–Ocimene, one of the monoterpenes, showeddominance throughout the range of the tissues studied (Table 1).Ten compounds, viz., 2-hexanol, camphene, b-pinene, a-methylbu-tyrolactone, c-valerolactone, b-terpinene, 4-carene, ocimene epox-ide, b-cyclocitral and b-ionone which were detected in trace levelsonly in some tissues were not quantified. Furaneol, one of the mostimportant flavour compounds in mango identified in our previousstudies (Pandit, Chidley, et al., 2009; Pandit, Kulkarni, et al., 2009)could not be quantified because of its inconsistent detection.
3.2. Ripening related changes
All the 12 mono- and sesquiterpenes were detected in all thefour ripening stages. For all the three localities, the highest concen-tration of total monoterpenes was found in the ripe fruits (Fig. 1).For Dapoli, all the monoterpenes except p-cymene showed thehighest quantity in the ripe fruit. In the case of Deogad fruits,although the highest quantity of all except p-cymene was detectedin the ripe fruit, none of the compounds differed statistically be-tween the ripening stages for their quantity. For Vengurle, all themonoterpenes, except p-cymene and limonene showed signifi-cantly higher quantity in the ripe fruit as compared to the otherstages (Table 1).
Out of the five sesquiterpenes detected, b-caryophyllene andhumulene showed statistically higher values in the ripe fruit thanany other stage for all the three localities. The pattern of appear-ance of the other three sesquiterpenes, longicyclene, isolongifoleneand longifolene was inconsistent; each of them showed dominanceat different stages through the localities but the difference be-tween the stages was significant only for Dapoli (Table 1).
Lactones and mesifuran were detected as ripening-relatedcompounds and their trend of appearance through the ripeningstages was similar for all three localities. They were completely ab-sent in raw and 5 DAH tissues and the highest quantity for each ofthese ripening-related compounds was observed in the ripe fruit(Table 1). In addition to lactones and furanones, five unidentifiedripening-related compounds were detected; out of which one wasquantified (Table 1, Supplementary Fig. 1), whereas four other werenot quantified because of their low amounts (Supplementary Fig. 2).
3.3. Cultivation locality wise variation
Qualitatively the volatile composition of the fruits from all thethree sampling localities selected for the present study was simi-lar; there were only quantitative differences in the appearancesof the volatiles.
3.3.1. 15 DAHQuantity of total monoterpenes in the ripe Dapoli fruits
(201–374 lg g�1) was significantly higher than those of Deogad(57–187 lg g�1) and Vengurle (73–282 lg g�1). A similar trend

Tabl
e1
Vol
atile
com
posi
tion
( lg
g�1
tiss
ue)
ofri
peni
ngfr
uits
(DA
H:
days
afte
rha
rves
t)of
Alp
hons
om
ango
from
thre
ecu
ltiv
atio
nlo
calit
ies,
Dap
oli,
Deo
gad
and
Ven
gurl
ein
Indi
a.V
alue
ssh
own
are
aver
age
offo
urpl
ants
sam
pled
from
each
loca
lity
and
thos
efo
llow
edby
±sy
mbo
lind
icat
est
anda
rder
ror
ofm
easu
rem
ents
.Com
poun
dsha
vebe
enor
dere
dac
cord
ing
toth
eir
rete
ntio
nin
dice
s.D
iffe
renc
ebe
twee
nth
elo
calit
ies
was
sign
ifica
nt(p6
0.05
)if
the
lett
ers
follo
win
gth
equ
anti
tyof
the
com
poun
dsar
edi
ffer
ent
for
apa
rtic
ular
stag
e.
Com
pou
nds
0D
AH
5D
AH
10D
AH
15D
AH
Dap
oli
Deo
gad
Ven
gurl
eD
apol
iD
eoga
dV
engu
rle
Dap
oli
Deo
gad
Ven
gurl
eD
apol
iD
eoga
dV
engu
rle
a-P
inen
ea0.
14±
0.05
0.09
±0.
030.
06±
0.01
0.15
±0.
050.
07±
0.01
0.09
±0.
020.
05±
0.03
0.11
±0.
030.
06±
00.
3±
0.08
b0.
14±
0.02
a0.
23±
0.02
ab
c-B
uty
rola
cton
en
.d.
n.d
.n
.d.
n.d
.n
.d.
n.d
.0.
52±
0.09
0.72
±0.
210.
34±
0.15
1.52
±0.
39a
5.5
±1.
69b
2.07
±0.
66a
Myr
cen
ea0.
84±
0.42
0.43
±0.
130.
51±
0.13
0.89
±0.
370.
36±
0.03
0.54
±0.
080.
4±
0.15
0.49
±0.
170.
45±
0.12
2.18
±0.
32b
0.71
±0.
12a
1.17
±0.
28a
p-C
ymen
ea0.
13±
0.05
0.04
±0.
030.
15±
0.08
00.
04±
0.04
0.04
±0.
020.
04±
0.02
0.06
±0.
060.
02±
0.01
0.07
±0.
040.
02±
0.02
0.03
±0.
03Li
mon
enea
0.19
±0.
040.
13±
0.03
0.18
±0.
040.
13±
0.06
0.14
±0.
050.
22±
0.03
0.11
±0.
020.
15±
0.01
0.11
±0.
020.
3±
0.03
b0.
18±
0.02
a0.
21±
0.02
a
(Z)-
Oci
men
e11
6±
63.2
58.1
±18
.362
.9±
17.2
116
±48
.942
.1±
3.76
81.1
±18
.553
.5±
21.7
68.6
±25
.958
.4±
14.4
273
±39
.5b
104
±26
.4a
157
±40
.8a
(E)-
Oci
men
e5.
1±
2.69
2.58
±0.
82.
88±
0.8
5.59
±2.
432.
05±
0.2
2.93
±0.
42.
41±
0.96
2.84
±0.
962.
65±
0.67
11.5
±2.
77b
4.12
±0.
85a
6.74
±1.
63ab
Mes
ifu
ran
an
.d.
n.d
.n
.d.
n.d
.n
.d.
n.d
.0.
51±
0.14
1.54
±0.
590.
38±
0.23
3.38
±0.
93a
11.4
±2.
17b
12.0
±5.
83b
c-H
exal
acto
nea
n.d
.n
.d.
n.d
.n
.d.
n.d
.n
.d.
1.33
±0.
16ab
1.81
±0.
43b
0.52
±0.
23a
2.64
±0.
43a
4.57
±0.
53b
2.29
±0.
42a
d-H
exal
acto
nea
n.d
.n
.d.
n.d
.n
.d.
n.d
.n
.d.
0.88
±0.
231.
22±
0.46
0.44
±0.
231.
92±
0.53
a7.
48±
1.78
b2.
58±
0.71
a
Allo
-oci
men
e1.
45±
0.69
0.74
±0.
230.
76±
0.22
1.79
±0.
710.
66±
0.1
0.72
±0.
150.
73±
0.3
0.99
±0.
290.
88±
0.21
3.17
±0.
36b
1.23
±0.
25a
1.72
±0.
57a
c-O
ctal
acto
nea
n.d
.n
.d.
n.d
.n
.d.
n.d
.n
.d.
2.08
±0.
372.
54±
0.7
0.95
±0.
515.
34±
1.22
5.74
±0.
523.
9±
0.92
d-O
ctal
acto
ne
n.d
.n
.d.
n.d
.n
.d.
n.d
.n
.d.
1.04
±0.
13ab
1.58
±0.
36b
0.42
±0.
13a
1.7
±0.
28a
2.86
±0.
16b
1.39
±0.
29a
Lon
gicy
clen
e1.
83±
0.3
0.73
±0.
631.
79±
0.97
0.28
±0.
131.
62±
1.06
0.7
±0.
350.
45±
0.12
1.14
±0.
690.
48±
0.2
1.18
±0.
470.
62±
0.36
0.84
±0.
55Is
olon
gifo
len
ea0.
65±
0.12
0.25
±0.
11.
14±
0.9
0.23
±0.
050.
6±
0.15
0.36
±0.
140.
47±
0.09
1.06
±0.
660.
54±
0.03
1.38
±0.
680.
6±
0.26
0.77
±0.
42Lo
ngi
fole
nea
6.36
±2.
532.
28±
2.06
5.44
±2.
031.
2±
14.
64±
2.74
1.26
±0.
681.
68±
0.72
1.9
±1.
072.
1±
1.26
3.03
±0.
791.
59±
0.76
1.97
±1.
35b
-Car
yoph
ylle
nea
2.43
±1.
141.
75±
0.67
0.99
±0.
263
±1.
211.
5±
0.18
0.87
±0.
221.
5±
0.53
1.38
±0.
460.
9±
0.15
4.15
±0.
66b
2.11
±0.
51a
2.48
±0.
69ab
Hu
mu
len
ea1.
36±
0.59
0.98
±0.
380.
55±
0.16
1.58
±0.
610.
87±
0.1
0.51
±0.
110.
77±
0.29
0.72
±0.
250.
43±
0.06
2.37
±0.
4b1.
17±
0.32
a1.
25±
0.36
ab
c-D
ecal
acto
nea
n.d
.n
.d.
n.d
.n
.d.
n.d
.n
.d.
0.08
±0.
040.
07±
0.01
0.07
±0.
010.
2±
0.01
0.23
±0.
060.
22±
0.07
Un
iden
tifi
edn
.d.
n.d
.n
.d.
n.d
.n
.d.
n.d
.0.
59±
0.2
0.7
±0.
280.
85±
0.18
0.36
±0.
11a
2.85
±1.
08b
0.6
±0.
23a
n.d
.:n
otde
tect
ed.
aTh
ese
com
pou
nds
wer
eid
enti
fied
byco
mpa
rin
gth
eir
mas
ssp
ectr
um
and
Kov
at’s
inde
xw
ith
thos
eof
auth
enti
cex
tern
alst
anda
rds;
the
rest
ofth
eco
mpo
un
dsw
ere
iden
tifi
edby
com
pari
ng
thei
rm
ass
spec
tru
man
dK
ovat
’sin
dex
wit
hth
ose
repo
rted
inth
eli
tera
ture
.
60 R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66
was observed when the compounds were considered individu-ally. Levels of all seven monoterpenes were significantly higherfor Dapoli fruits as compared to Deogad (Fig. 1). Dapoli fruitsalso had higher content of all the monoterpenes when comparedwith Vengurle, and the difference was significant for myrcene,limonene, (Z)-ocimene and allo-ocimene (Table 1). Vengurlefruits had higher levels of all the monoterpenes compared toDeogad.
Similar to monoterpenes, total and individual sesquiterpeneswere present in decreasing quantities from Dapoli (7.4–14 lg g�1) to Vengurle (2.8–15 lg g�1) to Deogad (1.5–9.1 lg g�1). Nevertheless, the difference was insignificant withrespect to total sesquiterpenes content (Fig. 1). Similarly, eachof the sesquiterpenes, when considered independently, waspresent in decreasing amount from Dapoli to Vengurle to Deo-gad and the higher amounts of b-caryophyllene and humulenein Dapoli than Deogad were statistically significant (Table 1).
The level of total lactones was significantly higher in Deogad(19–26 lg g�1) as compared to Dapoli (8–19 lg g�1) and Veng-urle (7–19 lg g�1) (Fig. 1). When considered individually, allthe lactones occurred in higher quantity in Deogad fruits ascompared to Dapoli and Vengurle and the difference was signif-icant for four (c-butyrolactone, c-hexalactone, d-hexalactoneand d-octalactone) of the six lactones (Table 1). The quantityof total and individual lactones was statistically similar forDapoli and Vengurle (Fig. 1). Mesifuran was the only furanonequantitated in the present study; its quantity was higher in Deo-gad and Vengurle than Dapoli.
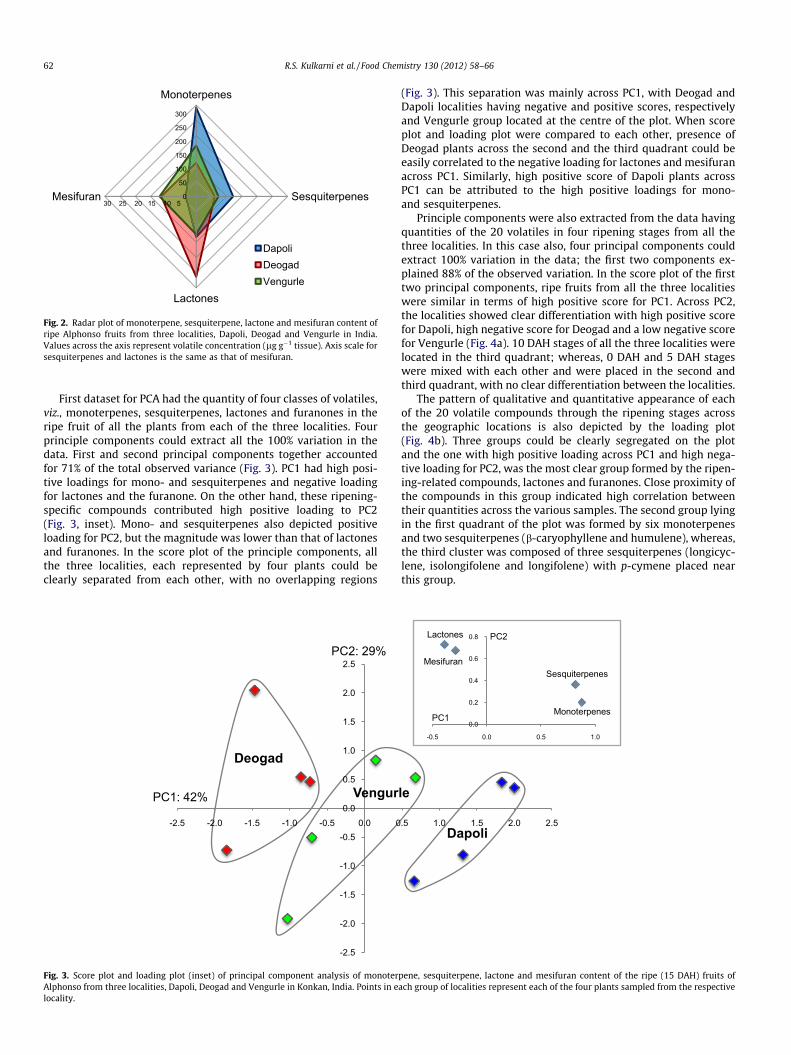
When the quantity of each of the abovementioned fourchemical classes was compared to each other on a radar plot(Fig. 2), Deogad fruits were clearly seen as lactone and mesifu-ran dominant as compared to Dapoli fruits, which could be seenas mono- and sesquiterpene dominant. Vengurle region wassimilar to Deogad in terms of mesifuran content and to Dapoliin terms of lactone content. On the scale of terpenes, Vengurlefruits showed average quantities.
3.3.2. 10 DAHWhen 10 DAH stage of each of the three localities was com-
pared with each other, Deogad fruits had slightly higher level ofthe mono- and sesquiterpenes (32–153 and 2.8–11 lg g�1) ascompared to Dapoli (2.6–105 and 1.4–8.8 lg g�1) and Vengurle(29–101 and 2.2–8.5 lg g�1), but the difference was insignifi-cant (Fig. 1). Also, all the individual monoterpenes and two ses-quiterpenes, longicyclene and isolongifolene were present in thehighest quantity in Deogad fruits (Table 1). Significantly higheramount of total lactones was detected in Deogad (3.3–13 lg g�1) in comparison with Vengurle (0.3–5.5 lg g�1) butnot in comparison with Dapoli (3.3–7.7 lg g�1) (Fig. 1). Whenconsidered individually, c-hexalactone and d-octalactone weredetected at higher levels in Deogad compared to the other twolocalities (Table 1). Deogad fruits contained higher amount ofmesifuran than that of Dapoli and Vengurle and the differencebetween Deogad and Vengurle was significant at p < 0.06.
3.3.3. 5 DAHTotal monoterpene content of Dapoli fruits (22–220 lg g�1)
was higher than that of Deogad (34–51 lg g�1) and Vengurle(41–133 lg g�1) and sesquiterpene content of Deogad fruits(2.8–19 lg g�1) was slightly higher than that of Dapoli (1.4–11.8 lg g�1) and Vengurle (2.1–5.8 lg g�1), but the differencewas insignificant (Fig. 1). When considered individually, fiveout of seven monoterpenes along with the two sesquiterpeneswere higher in Dapoli and the rest of the three sesquiterpeneswere higher in Deogad; in this case also the difference wasinsignificant (Table 1).

Fig. 1. Concentration (lg g�1 tissue) of various volatile classes in the ripening fruits of Alphonso from three different localities, Dapoli, Deogad and Vengurle in Konkan, India.Lactones and furanones were not detected in 0 DAH and 5 DAH fruits. Bars having different letters on the top indicate significantly different (p 6 0.05) values.
R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66 61
3.3.4. 0 DAHSix of the seven monoterpenes and four of the five sesquiterpenes
along with total mono- and sesquiterpenes were higher in Dapoli(38.2–320 and 5.5–21 lg g�1), as compared to Deogad (4.4–90 and0.6–17 lg g�1) and Vengurle (47–117 and 1.3–18 lg g�1); however,the difference was insignificant (Table 1, Fig. 1).
3.4. Principal component analysis
To analyse the complex data of quantitative distribution of vol-atiles across various geographic locations through various ripeningstages, PCA was carried out by treating the data in differentcombinations.
Monoterpenes
Sesquiterpenes
Lactones
Mesifuran
DapoliDeogadVengurle
300
250
200
150
100
50
030 25 20 15 10 5
Fig. 2. Radar plot of monoterpene, sesquiterpene, lactone and mesifuran content ofripe Alphonso fruits from three localities, Dapoli, Deogad and Vengurle in India.Values across the axis represent volatile concentration (lg g�1 tissue). Axis scale forsesquiterpenes and lactones is the same as that of mesifuran.
62 R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66
First dataset for PCA had the quantity of four classes of volatiles,viz., monoterpenes, sesquiterpenes, lactones and furanones in theripe fruit of all the plants from each of the three localities. Fourprinciple components could extract all the 100% variation in thedata. First and second principal components together accountedfor 71% of the total observed variance (Fig. 3). PC1 had high posi-tive loadings for mono- and sesquiterpenes and negative loadingfor lactones and the furanone. On the other hand, these ripening-specific compounds contributed high positive loading to PC2(Fig. 3, inset). Mono- and sesquiterpenes also depicted positiveloading for PC2, but the magnitude was lower than that of lactonesand furanones. In the score plot of the principle components, allthe three localities, each represented by four plants could beclearly separated from each other, with no overlapping regions
Vengur
-2.5
-2.0
-1.5
-1.0
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
-2.5 -2.0 -1.5 -1.0 -0.5 0.0 0
PC1: 42%
PC2: 29%
Deogad
Fig. 3. Score plot and loading plot (inset) of principal component analysis of monoterAlphonso from three localities, Dapoli, Deogad and Vengurle in Konkan, India. Points in elocality.
(Fig. 3). This separation was mainly across PC1, with Deogad andDapoli localities having negative and positive scores, respectivelyand Vengurle group located at the centre of the plot. When scoreplot and loading plot were compared to each other, presence ofDeogad plants across the second and the third quadrant could beeasily correlated to the negative loading for lactones and mesifuranacross PC1. Similarly, high positive score of Dapoli plants acrossPC1 can be attributed to the high positive loadings for mono-and sesquiterpenes.
Principle components were also extracted from the data havingquantities of the 20 volatiles in four ripening stages from all thethree localities. In this case also, four principal components couldextract 100% variation in the data; the first two components ex-plained 88% of the observed variation. In the score plot of the firsttwo principal components, ripe fruits from all the three localitieswere similar in terms of high positive score for PC1. Across PC2,the localities showed clear differentiation with high positive scorefor Dapoli, high negative score for Deogad and a low negative scorefor Vengurle (Fig. 4a). 10 DAH stages of all the three localities werelocated in the third quadrant; whereas, 0 DAH and 5 DAH stageswere mixed with each other and were placed in the second andthird quadrant, with no clear differentiation between the localities.
The pattern of qualitative and quantitative appearance of eachof the 20 volatile compounds through the ripening stages acrossthe geographic locations is also depicted by the loading plot(Fig. 4b). Three groups could be clearly segregated on the plotand the one with high positive loading across PC1 and high nega-tive loading for PC2, was the most clear group formed by the ripen-ing-related compounds, lactones and furanones. Close proximity ofthe compounds in this group indicated high correlation betweentheir quantities across the various samples. The second group lyingin the first quadrant of the plot was formed by six monoterpenesand two sesquiterpenes (b-caryophyllene and humulene), whereas,the third cluster was composed of three sesquiterpenes (longicyc-lene, isolongifolene and longifolene) with p-cymene placed nearthis group.
0.0
0.2
0.4
0.6
0.8
-0.5 0.0 0.5 1.0
Sesquiterpenes
Monoterpenes
Mesifuran
Lactones
PC1
PC2
le
.5 1.0 1.5 2.0 2.5Dapoli
pene, sesquiterpene, lactone and mesifuran content of the ripe (15 DAH) fruits ofach group of localities represent each of the four plants sampled from the respective

Fig. 4. Score plot (a) and loading plot (b) of principal component analysis for quantity of each of the 20 volatiles detected in the four ripening stages of the Alphonso fruitsfrom three localities, Dapoli, Deogad and Vengurle in India (DAH: days after harvest). Average compound quantities of four plants were considered for each stage from eachlocality.
R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66 63
3.5. Variation in terms of flavour
In the present data, out of 20 volatiles quantified in the ripe fruit(15 DAH), odour units were calculated for 12 compounds for whichodour detection threshold values were available in the literature
(Supplementary Table 1). Three compounds having the lowestodour detection threshold were mesifuran, a-pinene and c-oct-alactone. When the odour detection thresholds of the compoundswere correlated with their concentration in the Alphonso meso-carp, for all the three localities, the highest odour units were

Fig. 5. Odour unit value ratios of mesifuran/(Z)-ocimene (a) and c-octalactone/(Z)-ocimene (b) in ripe Alphonso fruits from three localities, Dapoli, Deogad andVengurle in India. Left-side end of the bars indicates the plant having the lowestvalue, whereas right-side end indicates the plant having the highest value amongthe four plants sampled from respective localities.
64 R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66
contributed by mesifuran, followed by (Z)-ocimene and c-octalac-tone. Since (Z)-ocimene has different odour character than mesifu-ran and c-octalactone, ratios of its odour units were calculatedwith these compounds. Deogad fruits had high ratio of mesifu-ran/(Z)-ocimene and c-octalactone/(Z)-ocimene as compared toDapoli; whereas, for Vengurle, these ratios were in between thosefor Deogad and Dapoli (Fig. 5).
4. Discussion
Flavour is one of the most important characteristics of fruits andvegetables and decides acceptability and preference of theseimportant food items by humans. For plants, flavour forms one ofthe most important phenotypes of the fruits that plays a vital func-tional role by attracting seed disseminators and repelling patho-gens, thereby increasing shelf-life of the fruits (Lanciotti et al.,2004). Qualitative and quantitative nature of the flavour dependson various parameters, including intrinsic factors, such as genotypeand physiological state of the plant as well as extrinsic factors, suchas abiotic and biotic conditions (Dudareva, Negre, Nagegowda, &Orlova, 2006; Pandit, Chidley, et al., 2009; Vallat, Gu, & Dorn,2005). Alphonso mangoes which are highly rich in flavour volatilesand which are thought to vary in their flavour with cultivationlocality form one of the best systems to study regulation of flavourchemistry.
4.1. Nature of the geographic variation in volatiles
To have a crop of consistent quality, Alphonso mango trees arepropagated by an asexual propagation method, grafting; hencethey are genetically identical. To analyse the geographic variationin the flavour volatiles of Alphonso mango, which is thus likelyto be the effect of only extrinsic conditions, such as variable geo-climatic factors, four plants representing four biological replicateswere analysed from each locality. The gas chromatographic analy-
sis of the mesocarp tissue revealed that fruits from all the threecultivation localities are rich in monoterpenes and ripe fruits arecharacterised by the unique presence of lactones and furanonesin them. These results are consistent with our previous study (Pan-dit, Chidley, et al., 2009; Pandit, Kulkarni, et al., 2009) and other re-ports which have also shown (Z)-ocimene as the dominantcompound in the volatile blend of Alphonso (Idstein & Schreier,1985). Concentration of mono- and sesquiterpenes detected (2.6–374 lg g�1 and 0.6–20.8 lg g�1, respectively) was quite low ascompared to our previous study (Pandit, Kulkarni, et al., 2009) inwhich whole fruit (except endocarp) was analysed for the volatilecomposition, as compared to only the mesocarp in the presentstudy. This could be because of the fact that Alphonso exocarp con-tains much higher concentration of terpenes than mesocarp(unpublished data). For all the three localities, higher amounts oftotal and individual monoterpenes were detected in the ripe fruitsas compared to the other stages. This also is consistent with an ear-lier report on the fruits from Deogad region (Pandit, Kulkarni, et al.,2009). Furaneol which has been reported earlier as an importantflavour constituent (Pandit, Chidley, et al., 2009; Pandit, Kulkarni,et al., 2009) could not be quantified because of its inconsistentdetection, which can be attributed to its instability at high operat-ing temperatures during GC (Siegmund, Bagdonaite, & Leitner,2010). Because of this fact, furaneol is more commonly analysedby techniques such as HPLC (Perez, Olias, Sanz, & Olias, 1996).
Principle component analysis (PCA) is a powerful tool to reducedimensionality in multivariate data. Through PCA, the multidimen-sional data is converted into the data which can be represented byminimum number of variables, still retaining variability in the ori-ginal data (Ringner, 2008). For the present data also, PCA turnedout to be an efficient method of extracting variability in the vola-tiles through the ripening stages of Alphonso from multiple local-ities. One of the principle components (PC) of the PCA of quantitiesof compounds in the ripening stages from three localities repre-sented transition of the fruits towards the ripe stage with synthesisof the new compounds (lactones and furanones) and increase inquantities of the already-present volatiles (terpenes). The otherPC underlined clear discrimination between localities for the vola-tiles in the ripe fruits. On the other hand, both of the above PCssupported absence of grouping of localities for the unripe stages.In the loading plot of the same PCA, terpene volatiles formed twodistinct clusters. This points towards the possibility of presenceof at least two types of terpene synthases in Alphonso fruits. Eachof them might be regulated in a specific manner, giving rise to auniquely separate profile for their products through the ripeningstages for different localities. Furthermore, similar profile of allthe monoterpenes through the set of tissues studied as suggestedby clustering of all the monoterpenes (except p-cymene) in a singlegroup, further indicates the possibility of regulation of their bio-genesis at a common upstream step such as geranyl pyrophosphatesynthase which provides direct precursor for monoterpenes.
4.2. Geographic variation in terms of flavour
Comparative quantitative analysis of volatile compounds withina single tissue and between different tissue samples can be veryinformative in the perspective of plant biochemistry. For example,it can shed light on the most active pathways among an array ofbiosynthetic routes of volatiles or can give a hint towards the path-ways triggered during physiological processes, such as ripening(Pech, Bouzayen, & Latche, 2008). Nevertheless, when volatilesare studied from tissues such as edible fruit, the analysis cannotbe considered functional without dissecting the relative impor-tance of these compounds in the human’s viewpoint in terms ofthe flavour. Because of differential interactions of various volatilecompounds with their receptors in the human olfactory system,

R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66 65
these chemicals are sensed in different magnitudes, as well as eachof them has its own characteristic response. The minimum concen-tration at which any volatile compound can be detected is called itsodour detection threshold and the relative aroma contribution ofeach compound or odour units is calculated by dividing concentra-tion of the compound by its odour detection threshold (Goldstein,2009).
Among the present set of compounds, mesifuran, (Z)-ocimeneand c-octalactone had the highest odour units and thus they werethe major contributors to the Alphonso flavour. Out of these, mes-ifuran has coumarin-like fruity odour, (Z)-ocimene has citrus-liketerpenic odour, whereas c-octalactone has coconut-like creamyodour. Thus, having a high ratio of mesifuran/(Z)-ocimene and c-octalactone/(Z)-ocimene, Deogad fruits can be described as rela-tively ‘‘pleasant’’ and ‘‘fruity’’ in terms of the flavour as comparedto Dapoli fruits which can be said to be more ‘‘terpenic’’ becauseof the low ratios of the above mentioned compounds (Fig. 5).
4.3. Source of the geographic variation
Plants are one of the most dynamic entities among the livingorganisms that adapt themselves according to the surrounding bio-tic as well as abiotic environment. Such an ability of plants to showphenotypic changes in different environments, especially underdifferential abiotic conditions, is termed as phenotypic plasticity(Majetic, Raguso, & Ashman, 2009). Till now, existence of geo-graphic variation or phenotypic plasticity in terms of volatileshas been shown mainly in the floral scents (Dotterl, Wolfe, & Jur-gens, 2005; Knudsen, 2002; Svensson et al., 2005) and in the essen-tial oils in the vegetative tissue (Adams, 2001; Neffati et al., 2009).However, in most of these cases, plants belonged to the naturalpopulations and hence they can show some genetic variation,which along with the varied environmental conditions could ac-count for the observed variation in the volatile composition (Maj-etic et al., 2009). Since Alphonso plants are genetically identicalbecause of their propagation by grafting, the difference observedin the volatiles at different cultivation localities is likely to be reg-ulated by external environmental factors. Furthermore, cultivationpractices and the post-harvest treatment to the Alphonso fruitsthroughout the broad cultivation region, Konkan, are the same.Therefore, the influence of the external factors is probably exertedduring fruit development and in terms of flavour variation; this ef-fect is reflected 15 days after the mature fruit has been detachedfrom the plant.
Fruit ripening is one of the most important processes in the lifecycle of the trees. During this phase, fruits get their functionaldimension by becoming attractive to seed dispersing agents, whichis achieved by increase in sweetness and nutritive value and syn-thesis of colour and flavour compounds. In the case of Alphonsomango, changes in the flavour volatiles during fruit ripening werevery clear. Furanones and lactones were completely absent in theraw fruits and they were synthesized in increasing amounts duringfruit ripening till 15 DAH stage. Although ripening was also charac-terised by the increase in the concentration of terpenes, these com-pounds were already present in the raw fruits as well as in thevegetative tissue, such as leaves, as shown previously (Pandit,Kulkarni, et al., 2009). In the light of these observations, there ap-pears to be a different molecular mechanism for the regulation ofbiosynthesis of terpenes and lactones, which can also account forthe differential behaviour of these two classes of volatiles amongdifferent geographic locations.
In climacteric fruits, most of the ripening-related changes areinduced by ethylene. Indeed, in some of the mango cultivars, eth-ylene production was also observed prior to attaining harvestmaturity (Cua & Lizada, 1990), pointing towards developmentalcontrol of ripening. Both developmental and climacteric ethylene
production are regulated at the rate-limiting step in its biosyn-thetic pathway catalysed by 1-aminocyclopropane-1-carboxylatesynthase (ACC synthase), which has been shown to be the control-ler of ripening-related changes (Argueso, Hansen, & Kieber, 2007).In addition, in tomato (Vrebalov et al., 2002) and banana (Elitzur,Vrebalov, Giovannoni, Goldschmidt, & Friedman, 2010; Liu et al.,2009) it was shown that MADS-box transcription factors play a vi-tal role in developmental control of ripening-related changes. Thisinvolvement of MADS transcription factors in ripening is thoughtto be through their regulatory action on endogenous ethylene pro-duction (Elitzur et al., 2010) which further induces expression offlavour and other ripening-related genes (Klee, 2004).
In case of Alphonso mangoes, no variation in terms of volatilecomposition was observed among the localities at non-ripe stagesand the clear difference was seen at the ripe stage. It is possiblethat the geographic difference might also exist at the non-ripestages of the fruits in terms of the levels of precursors of the vola-tiles and during the process of ripening this difference is trans-formed into the variation observed in the volatiles. Although, it isdifficult to pinpoint the exact environmental factors that areresponsible for such phenotypic plasticity, there are certain knownconditions that vary between sampling localities selected for thepresent study. For example, the plantation region at Deogad is hillyand the soil is lateritic and rocky and thus has much lower mois-ture retention capacity, as compared to Dapoli and Vengurle(Burondkar, Bhingarde, Kore, & Powar, 2000; and personal observa-tion). It was also shown that at Deogad, heat units required forcomplete maturation of the fruits are gained much more rapidlyas compared to Vengurle (Burondkar et al., 2000). It can be hypoth-esized that such differential environmental clues at different geo-graphic locations are picked up by the upstream regulatoryswitches such as MADS-box transcription factors during fruitdevelopment and through their action on a series of downstreammediators, including ripening related agents, the final visible effectis brought about at the ripe stage.
In our previous study, we have studied expression profiles ofvarious flavour genes through the developmental and ripeningstages of Alphonso from Deogad which indicated a ripening-re-lated pattern of expression of many genes (Pandit et al., 2010). Itwould be interesting to study expression of these flavour genesalong with the other ripening-related genes across the geographiclocations to shed light on the downstream players in flavourregulation.
Acknowledgements
R.S.K. and H.G.C. thank CSIR for research fellowships. Authorsthank Bhushan Nabar, who provided the fruits from Vengurlelocality. Dr BSKKV staff is acknowledged for the field support.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.foodchem.2011.06.053.
References
Adams, R. P. (2001). Geographic variation in leaf essential oils and RAPDs ofJuniperus polycarpos K. Koch in central Asia. Biochemical Systematics and Ecology,29(6), 609–619.
Argueso, C. T., Hansen, M., & Kieber, J. J. (2007). Regulation of ethylene biosynthesis.Journal of Plant Growth Regulation, 26(2), 92–105.
Bandyopadhyay, C., & Gholap, A. S. (1973). Changes in fatty acids in ripening mangopulp (variety Alphonso). Journal of Agricultural and Food Chemistry, 21(3),496–497.
Bawa, A. S. (2007). Historical developments in food science and technology – Indianperspective. Journal of Food Science and Technology – Mysore, 44(6), 553–564.

66 R.S. Kulkarni et al. / Food Chemistry 130 (2012) 58–66
Burondkar, M. M., Bhingarde, R. T., Kore, V. N., & Powar, A. G. (2000). Estimation ofheat units as maturity indices for different mango varieties in Konkan region ofMaharshtra. In Proceedings of the sixth international symposium on mango (Vols.1 and 2(509), pp. 297–299).
Cua, A. U., & Lizada, M. C. C. (1990). Ethylene production in the Carabo mango(Mangifera india L.) fruit during maturation and ripening. In Symposium ontropical fruit in international trade (Vol. 269, pp. 169–179).
Dotterl, S., Wolfe, L. M., & Jurgens, A. (2005). Qualitative and quantitative analyses offlower scent in Silene latifolia. Phytochemistry, 66(2), 203–213.
Dudareva, N., Negre, F., Nagegowda, D. A., & Orlova, I. (2006). Plant volatiles: Recentadvances and future perspectives. Critical Reviews in Plant Sciences, 25(5),417–440.
Elitzur, T., Vrebalov, J., Giovannoni, J. J., Goldschmidt, E. E., & Friedman, H. (2010).The regulation of MADS-box gene expression during ripening of banana andtheir regulatory interaction with ethylene. Journal of Experimental Botany, 61(5),1523–1535.
Goldstein, E. B. (2009). The chemical senses. Sensation and perception (pp. 355–377).Cengage Learning.
Hassan, M. K., Irving, D. E., Dann, E. K., Coates, L. M., & Hofman, P. J. (2009). Sapproperties and alk(en)ylresorcinol concentrations in Australian-grownmangoes. Annals of Applied Biology, 154(3), 419–427.
Hofman, P. J., Smith, L. G., Meiburg, G. F., & Giles, J. E. (1997). Production localityaffects mango fruit quality. Australian Journal of Experimental Agriculture, 37(7),801–808.
Idstein, H., & Schreier, P. (1985). Volatile constituents of Alphonso mango(Mangifera indica). Phytochemistry, 24(10), 2313–2316.
Klee, H. J. (2004). Ethylene signal transduction. Moving beyond Arabidopsis. PlantPhysiology, 135(2), 660–667.
Knudsen, J. T. (2002). Variation in floral scent composition within and betweenpopulations of Geonoma macrostachys (Arecaceae) in the western Amazon.American Journal of Botany, 89(11), 1772–1778.
Lanciotti, R., Gianotti, A., Patrignani, F., Belletti, N., Guerzoni, M. E., & Gardini, F.(2004). Use of natural aroma compounds to improve shelf-life and safety ofminimally processed fruits. Trends in Food Science & Technology, 15(3–4),201–208.
Liu, J. H., Xu, B. Y., Hu, L. F., Li, M. Y., Su, W., Wu, J., et al. (2009). Involvement of abanana MADS-box transcription factor gene in ethylene-induced fruit ripening.Plant Cell Reports, 28(1), 103–111.
Majetic, C. J., Raguso, R. A., & Ashman, T.-L. (2009). Sources of floral scent variation:Can environment define floral scent phenotype? Plant Signalling and Behaviour,4(2), 129–131.
Neffati, A., Hennequin, D., Basset, B., Chekir-Ghedira, L., Ghedira, K., Barillier, D.,et al. (2009). Influence of growth phase and geographic origin on the essentialoil composition of Pituranthos chloranthus from Tunisia. Natural ProductCommunications, 4(11), 1585–1594.
Pandit, S. S., Chidley, H. G., Kulkarni, R. S., Pujari, K. H., Giri, A. P., & Gupta, V. S.(2009). Cultivar relationships in mango based on fruit volatile profiles. FoodChemistry, 114(1), 363–372.
Pandit, S. S., Kulkarni, R. S., Chidley, H. G., Giri, A. P., Pujari, K. H., Kollner, T. G., et al.(2009). Changes in volatile composition during fruit development and ripeningof ‘Alphonso’ mango. Journal of the Science of Food and Agriculture, 89(12),2071–2081.
Pandit, S. S., Kulkarni, R. S., Giri, A. P., Koellner, T. G., Degenhardt, J., Gershenzon, J.,et al. (2010). Expression profiling of various genes during the fruit developmentand ripening of mango. Plant Physiology and Biochemistry (Paris), 48(6),426–433.
Pech, J. C., Bouzayen, M., & Latche, A. (2008). Climacteric fruit ripening: Ethylene-dependent and independent regulation of ripening pathways in melon fruit.Plant Science, 175(1–2), 114–120.
Perez, A. G., Olias, R., Sanz, C., & Olias, J. M. (1996). Furanones in strawberries:Evolution during ripening and postharvest shelf life. Journal of Agricultural andFood Chemistry, 44(11), 3620–3624.
Ringner, M. (2008). What is principal component analysis? Nature Biotechnology,26(3), 303–304.
Salvi, M. J., & Gunjate, R. T. (1988). Mango breeding work in the Konkan region ofMaharashtra state. Acta Horticulturae, 231, 100–102.
Siegmund, B., Bagdonaite, K., & Leitner, E. (2010). Furaneol� and mesifuran instrawberries – An analytical challenge. In I. Blank, M. Wüst, & C. Yeretzian(Eds.), Expression of multidisciplinary flavour science (pp. 537–540). WinterthurZHAW Züricher Hochschule für Angewandte Wissenschaften.
Svensson, G. P., Hickman, M. O., Bartram, S., Boland, W., Pellmyr, O., & Raguso, R. A.(2005). Chemistry and geographic variation of floral scent in Yucca filamentosa(Agavaceae). American Journal of Botany, 92(10), 1624–1631.
Vallat, A., Gu, H. N., & Dorn, S. (2005). How rainfall, relative humidity andtemperature influence volatile emissions from apple trees in situ.Phytochemistry, 66(13), 1540–1550.
Vrebalov, J., Ruezinsky, D., Padmanabhan, V., White, R., Medrano, D., Drake, R., et al.(2002). A MADS-box gene necessary for fruit ripening at the tomato ripening-inhibitor (Rin) locus. Science, 296(5566), 343–346.
Copyright © 2022 FDOKUMEN




















