
Electrophysiological and Behavioral Responses to Chocolate Volatiles in Both Sexes of the Pyralid...
14
Electrophysiological and Behavioral Responses of a Cuban Population of the Sweet Potato Weevil to its Sex Pheromone Tania Sureda & Carmen Quero & Ma. Pilar Bosch & Rubén Avilés & Francisco Coll & Michel Renou & Ángel Guerrero Received: 7 February 2006 / Revised: 2 June 2006 / Accepted: 11 June 2006 / Published online: 15 September 2006 # Springer Science + Business Media, Inc. 2006 Abstract The sex pheromone of feral sweet potato weevils Cylas formicarius elegantulus from Cuba was found, via solid-phase microextraction analysis, to be identical to (Z)-3- dodecenyl (E)-2-butenoate, a previously reported compound. Females emitted 20 pg pheromone d −1 . In scanning electron microscopy studies carried out on the male antenna, we identified several types of sensilla: sensilla trichoidea of type 1 (ST1) as long hairs (100–150 μm), sensilla trichoidea of type 2 as short hairs (50–60 μm), sensilla basiconica of type 1 as thick pegs (20–25 μm), sensilla basiconica of type 2 as curved pegs (10– 15 μm), and sensilla basiconica of type 3 as thin and straight short pegs (15–20 μm). The same types were observed in female antennae but ST1 were far less abundant than in males. Sensilla chaetica were also found on the flagellum subsegments in both sexes. In electrophysiological tests, the crotonate function in the pheromone structure proved to be critical for activity since regular depolarizations (0.6–0.8 mV) were obtained with puffs on 1 μg of the attractant, but not with puffs of the formate, acetate, propionate, or butyrate analogue of the pheromone. In a double dual-choice olfactometer, males showed maximum activity between the 4th and the 8th hr of scotophase at a dose of 50–1000 ng of pheromone. In field tests, a correlation between the contents of the Z,E isomer in the pheromone formulation with activity was noticed, and baits containing this isomer of J Chem Ecol (2006) 32: 2177–2190 DOI 10.1007/s10886-006-9138-4 T. Sureda : C. Quero : M. P. Bosch : Á. Guerrero (*) Department of Biological Organic Chemistry, Institute of Chemical and Environmental Research (CSIC), Jordi Girona 18-26, 08034 Barcelona, Spain e-mail: [email protected] R. Avilés INIFAT, Av. Rancho Boyeros, Santiago de las Vegas, Cuba F. Coll Centro de Estudios de Productos Naturales, Facultad de Química, Universidad de la Habana, Zapata y G, Vedado, 10400 La Habana, Cuba M. Renou INRA, Unité de Phytopharmacie et Médiateurs Chimiques, Route de St. Cyr, 78026 Versailles Cédex, France
Transcript of Electrophysiological and Behavioral Responses to Chocolate Volatiles in Both Sexes of the Pyralid...

Electrophysiological and Behavioral Responsesof a Cuban Population of the Sweet PotatoWeevil to its Sex Pheromone
Tania Sureda & Carmen Quero & Ma. Pilar Bosch &
Rubén Avilés & Francisco Coll & Michel Renou &
Ángel Guerrero
Received: 7 February 2006 / Revised: 2 June 2006 /Accepted: 11 June 2006 / Published online: 15 September 2006# Springer Science + Business Media, Inc. 2006
Abstract The sex pheromone of feral sweet potato weevils Cylas formicarius elegantulusfrom Cuba was found, via solid-phase microextraction analysis, to be identical to (Z)-3-dodecenyl (E)-2-butenoate, a previously reported compound. Females emitted 20 pgpheromone d−1. In scanning electron microscopy studies carried out on the male antenna,we identified several types of sensilla: sensilla trichoidea of type 1 (ST1) as long hairs(100–150 μm), sensilla trichoidea of type 2 as short hairs (50–60 μm), sensilla basiconicaof type 1 as thick pegs (20–25 μm), sensilla basiconica of type 2 as curved pegs (10–15 μm), and sensilla basiconica of type 3 as thin and straight short pegs (15–20 μm). Thesame types were observed in female antennae but ST1 were far less abundant than in males.Sensilla chaetica were also found on the flagellum subsegments in both sexes. Inelectrophysiological tests, the crotonate function in the pheromone structure proved to becritical for activity since regular depolarizations (0.6–0.8 mV) were obtained with puffs on1 μg of the attractant, but not with puffs of the formate, acetate, propionate, or butyrateanalogue of the pheromone. In a double dual-choice olfactometer, males showed maximumactivity between the 4th and the 8th hr of scotophase at a dose of 50–1000 ng ofpheromone. In field tests, a correlation between the contents of the Z,E isomer in thepheromone formulation with activity was noticed, and baits containing this isomer of
J Chem Ecol (2006) 32: 2177–2190DOI 10.1007/s10886-006-9138-4
T. Sureda : C. Quero :M. P. Bosch :Á. Guerrero (*)Department of Biological Organic Chemistry,Institute of Chemical and Environmental Research (CSIC),Jordi Girona 18-26, 08034 Barcelona, Spaine-mail: [email protected]
R. AvilésINIFAT, Av. Rancho Boyeros, Santiago de las Vegas, Cuba
F. CollCentro de Estudios de Productos Naturales, Facultad de Química,Universidad de la Habana, Zapata y G, Vedado, 10400 La Habana, Cuba
M. RenouINRA, Unité de Phytopharmacie et Médiateurs Chimiques,Route de St. Cyr, 78026 Versailles Cédex, France

stereomeric purity >94% showed the highest attractivity. The presence of 5% of the Z,Zisomer in the lure did not induce any synergistic or inhibitory effect, and the alcoholprecursor of the pheromone was inactive. The results show that use of a stereomericallypure pheromone may not be necessary in pest control strategies.
Keywords Sex pheromone . Identification . Synthesis . Sweet potato weevil .
Cylas formicarius elegantulus . Electrophysiological activity .
Olfactometer bioassay . Field tests
Introduction
Sweet potato, Ipomoea batatas (L.) Lam., is one of the seven most important food cropsworldwide, with an estimated annual production of 115 million metric tons (FAO, 1984). Itis grown in more than 100 countries with 98% of the world production proceeding fromdeveloping countries (Jansson and Raman, 1991). One of the most important constraintsthat limit sweet potato production worldwide are the pre- and postharvest losses from insectfeeding, especially the sweet potato weevils Cylas formicarius (Fabricius), C. puncticollis(Boheman), and C. brunneus (Fabricius) (Jansson and Raman, 1991). C. formicarius [=C. f.elegantulus (Summers)] abounds in more than 50 countries in tropical and subtropicalregions and is the most devastating pest of sweet potato worldwide (Chalfant et al., 1990).In response to insect feeding, sweet potato tissue produces toxic sesquiterpenes that inducean extremely bitter taste in the roots, rendering it unfit for consumption (Akazawa et al.,1960; Sato et al., 1981). For this reason, even low population densities may causedevastating crop losses, i.e., yield losses of between 60% and 100% in many developingcountries. Owing to the cryptic character of these weevils, insects are difficult to controlwith conventional insecticides, and growers in many countries have tried to reducepopulations primarily by cultural practices and, more recently, through biological control(Jansson, 1991).
The sex pheromone of C. f. elegantulus was identified by Heath et al. (1986, 1988) as(Z)-3-dodecenyl (E)-2-butenoate (1) from a laboratory-reared colony. In this paper, wepresent the pheromone analysis of feral insects from Cuba, the first scanning electronmicroscopy (SEM) studies on insect antenna, new electrophysiological and behavioralresponses to the pheromone in a double dual-choice olfactometer, and activity of severalpheromone formulations in the field.
Methods and Materials
Insects
Weevils were collected in infested sweet potato fields near the Instituto de InvestigacionesFundamentales (INIFAT; Santiago de las Vegas, Cuba). They were sexed, fed withcommercial sweet potato, and sent to Barcelona, Spain, for pheromone analysis andbiological and electrophysiological activity. They were kept in a climatic chamber on a 16:8L/D photoperiod.
Solid Phase Microextraction
Commercial fibers of polydimethylsiloxane (7 and 100 μm thickness) (Supelco Inc.,Bellefonte, PA, USA) were previously conditioned in a Carlo Erba 6000 gas chromatograph
2178 J Chem Ecol (2006) 32: 2177–2190

at 250°C injector temperature for 1 hr. The fiber was introduced into a Teflon-coated screw-top glass vial containing batches of 26–90 virgin females. Volatiles emitted by the insectswere adsorbed for 24 hr at room temperature and desorbed in the injector port of aThermoQuest gas chromatograph coupled to a Trace MS ThermoQuest mass selectivedetector. The gas chromatography (GC) conditions used were injection in splitless mode onan HP-5 MS capillary column (30 m × 0.25 mm ID × 0.25 μm) at 60°C for 5 min, and thenprogrammed to 230°C at 5°C/min. The GC-mass spectrometry (MS) was run under SIMmode of ions of m/z 166, 138, 96, 87, and 69.
Chemicals
Synthesis of pheromone 1 was carried out by esterification of (Z)-3-dodecen-1-ol (2), whichwas prepared from MOM-protected 3-butyn-1-ol (3) (Scheme 1). A solution of 4-methoxymethoxy-1-butyne (3) (0.5 g, 4.3 mmol) in anh. tetrahydrofuran (THF) (3.5 ml)was cooled to −20°C. Then, a freshly prepared 0.5 M solution of LDA (8.8 ml, 4.3 ml) wasslowly added. The solution was stirred for 10 min at −20°C and 1 hr at room temperature.A solution of 1-bromooctane (0.83 g, 4.3 mmol) in a mixture of anh. THF (2 ml) and anh.3,4,5,6-tetrahydro-2-(1H)-pyrimidinone (DMPU) (3 ml) was added, and the mixture stirredfor 3 hr. THF was evaporated under vacuum, and the residue was poured over ice/water.Organic material was extracted with hexane, washed with water, and dried (Na2SO4). Afterevaporation of the solvent, the residue was purified by flash column chromatography overneutral alumina III to obtain the condensation product 4 (0.6 g, 65%) in pure form. 1HNMR δ 4.65 (s, 2H, OCH2O), 3.61 (t, J = 6.9 Hz, 2H, CH2O), 3.37 (s, 3H, CH3O), 2.46 (tt,J = 6.9, J′ = 2.4 Hz, 2H, C ≡ CCH2), 2.14 (tt, J = 6.9, J′ = 2.4 Hz, 2H, CH2C ≡ C), 1.46 (m,2H, CH2CH2C ≡ C), 1.26 (br, 10H, 5CH2), 0.87 (t, J = 6.7 Hz, 3H, CH3) ppm. 13C NMR δ95.5, 80.9, 77.3, 65.8, 54.5, 31.1, 28.5, 28.4, 28.3, 28.1, 21.9, 19.6, 17.9, 13.8 ppm. MS(EI) m/z (%): 226 (M+, 1). Elem. Anal. Calcd. for C14H26O2: C, 74.28, H, 11.58; Found: C,74.22, H, 11.56. A mixture of compound 4 (0.211 g, 0.93 mmol), MeOH (2 ml), and 12 MHCl (0.21 ml) was stirred at room temperature for 24 hr. The solvent was evaporated and
Scheme 1 Synthesis of pheromone 1 by esterification of (Z)-3-dodecen-1-ol (2)
J Chem Ecol (2006) 32: 2177–2190 2179

the residue was extracted with CH2Cl2 and washed with NaHCO3 sat solution. Usualworkup and purification by flash column chromatography over neutral alumina IIIfurnished compound 5 (0.153 g, 91%). IR ν 3340, 2220 cm−1. 1H NMR δ 3.67 (dt, J =J′ = 6.3 Hz, 2H, CH2O), 2.43 (tt, J = 6.3, J′ = 2.4 Hz, 2H, C ≡ CCH2), 2.15 (tt, J = 7, J′ =2.4 Hz, 2H, CH2C ≡C), 1.81 (t, J = 6.3 Hz, 1H, OH), 1.48 (m, 2H, CH2CH2C ≡ C), 1.26 (br,10H, 5 CH2), 0.88 (t, J = 6.75 Hz, 3H, CH3) ppm. 13C NMR δ 82.7, 76.2, 61.3, 31.8, 29.2,29.1, 28.9, 28.8, 23.1, 22.6, 18.7, 14.05 ppm. MS (EI) m/z (%): 182 (M+, 2). Alkynol 5(0.59 g, 3 mmol) was hydrogenated with Lindlar (Pd-CaCO3) catalyst (20 mg) in hexane(3 ml) and two drops of quinoline at atmospheric pressure and room temperature. Thereaction was monitored by GC with an HP-5 capillary column (0.33 μm, 25 m × 0.20 mmID). The crude was filtered through Celite, and after usual workup, compound 2 (0.54 g,91%) was obtained with a stereomeric purity Z,E 99/1. IR ν 3334 cm−1. 1H NMR δ 5.56(m, 1H, HC=), 5.35 (m, 1H, = CH), 3.64 (t, J = 6.5 Hz, 2H, CH2O), 2.33 (c, J = 6.5 Hz, 2H,=CCH2), 2.05 (c, J = 6.6 Hz, 2H, CH2C=), 1.70 (s, 1 H, OH), 1.26 (br, 12H, 6CH2), 0.87(t, 3H, J = 6.75 Hz, CH3) ppm. 13C NMR δ 133.6, 124.9, 62.3, 31.9, 30.8, 29.7, 29.5, 29.3,29.2, 27.4, 22.7, 14.1 ppm. MS (EI) m/z (%): 184 (M+, 0.4), 166 (M+-H2O, 12). A mixtureof crotonyl chloride (0.19 ml, 2 mmol) and alcohol 2 (0.3 g, 1.6 mmol) was dissolved inCH2Cl2 (3 ml) and cooled to 0°C. Pyridine (0.16 ml, 2 mmol) in CH2Cl2 (3 ml) was addeddropwise to the solution, and the mixture was stirred at 0°C for 1 hr (Heath et al., 1986).Usual workup and purification over flash column chromatography on silica gel yieldedcompound 1 (0.23 g, 57%), as a mixture of isomers (3Z)-1 and (3E)-1 with the Estereochemistry of the double bond at the acid side and the isomer containing the twodouble bonds with Z stereochemistry in 94:1:5 ratio, respectively. IR ν 1724, 1660,968 cm−1. 1H NMR δ 6.97 (dq, J = 15.6, J′ = 6.9 Hz, 1H, OCCH = CH), 5.84 (dq, J = 15.6,J′ = 1.75 Hz, 1H, OCCH = CH), 5.50 (m, 1H, HC=), 5.35 (m, 1H, =CH), 4.12 (t, J =6.75 Hz, 2H, CH2O), 2.40 (m, 2H, =CCH2), 2.03 (m, 2H, CH2C=), 1.87 (dd, J = 6.9, J′ =1.75 Hz, 3H, =CCH3), 1.26 (br, 12H, 6CH2), 0.88 (t, J = 7.0 Hz, 3H, CH3) ppm. 13C NMRδ 166.6, 144.6, 132.9, 124.3, 122.7, 63.7, 31.9, 29.6, 29.5, 29.3, 27.3, 26.9, 22.7, 18.0,14.1 ppm; MS (EI) m/z (%): 166 (M+-CH3 – CH = CH – COOH, 76).
For field tests, samples of 3-dodecenyl (E)-2-butenoate were prepared as mixtures of(3Z)- and (3E)-dodecenyl crotonate in several ratios through Wittig reaction of 3-hydroxypro-pyltriphenylphosphorane with nonanal, or by reaction of methylenetriphenylphosphorane withethylene oxide followed by metallation and Wittig reaction with nonanal (Maryanoff et al.,1985).
Scanning Electron Microscopy
The head of a freshly sacrificed weevil was cut, fixed on a stub with conductive glue, air-dried, and metalized. Observations were performed on a Jeol JSM 840 SEM at 5 eV.
Electrophysiological Activity
Electroantennogram (EAG) responses were recorded under standard techniques by using apair of glass microelectrodes filled with Ringer’s saline (Guerrero et al., 1986). The tip ofthe antenna club was cut off, the reference electrode was introduced into the base of thehead, and the recording electrode was slipped into the open tip of the antenna. A flow ofhumidified air (900 ml/min) was continuously directed over the antenna, and olfactorystimuli were delivered by puffing air (300 ml/min) with a CS-1 stimulus generator (200 ms
2180 J Chem Ecol (2006) 32: 2177–2190

duration; Syntech, Hilversum, The Netherlands) through a Pasteur pipette directly into themain stream of the air. The pipette contained a small filter paper on which several amounts(0.001–1 μg) of the pheromone were deposited. All experiments were performed with thesynthetic pheromone 1 having the highest stereochemical purity (mixture of isomers Z,Eand E,E in a 98:2 ratio). Control puffs (pure air) were intercalated between two consecutivepheromone stimuli to determine the real depolarizations to the attractant. Responses fromthe antenna were amplified (1000×), filtered (DC to 1 kHz) with an IDC 02 interface(Syntech), digitized on a PC, and analyzed with the EAG 2.6 program (Syntech).
Olfactometer Bioassay
For behavioral studies, a dual-choice olfactometer consisting of two methacrylate-basedplates of 20 × 20 cm adjusted to each other by four plastic screws was built. The upper platecontained a hole (diam 5.5 cm) in the center that allowed us to suck out the air of the systemvia an ordinary vacuum pump. The vacuum allowed volatiles from the vials to reach the testinsects, which had been initially placed in groups of 4–5 in the center of the chamber. Each ofthe four olfactometer sides had an outlet with a Tygon® hose adjusted to a glass vial. Each vialcontained a piece of paper impregnated with a bait (pheromone) or blank (hexane) so that thetwo baited vials were surrounded by two control vials in an alternate form. The number ofinsects that reached each arm of the olfactometer was counted, and the percentage of re-sponses with regard to the total number of insects was calculated. The apparatus was rotated90° after each assay so that the vials occupied all four possible positions in each assay. Byrotating the system, any photokinetic side effect induced by indirect light coming into thechamber was avoided. The experiments were performed in the dark, and the chamber wasthoroughly cleaned with 96% ethanol after every assay. To establish the optimum response ofmales, 10 insects were released and their responses to 10 ng pheromone recorded at differenttimes of the scotophase. Eight replicates were done for each assay. Responses of males of the6th hr into the scotophase to several pheromone doses (0.1 to 10,000 ng) were also recorded,and 10 replicates with 10 insects/replicate were run for each dose. For comparison, the activityof a commercial sample (50 ng) was also determined (12 replicates with 10 insects/replicate).
Field Tests
Field experiments were conducted in 1-ha infested sweet potato fields at the Cuban Institute ofTropical Agricultural Research (Santiago de las Vegas, Cuba) (July–August 2000–2001). Thetraps were made with the materials available in the country. Two thick 50-cm-long iron barssupported an open square iron box at an approx. height of 45 cm from the floor. Two oppositesides of the box were covered with a plastic sheet impregnated with Tanglefoot®. The roof of thetrap was cardboard covered with aluminum foil, from which a thin wire was suspended with arubber septum (Aldrich, Milwaukee, WI, USA) loaded with 1 mg of the formulation. In thelower part of the trap and ca. 10 cm from the ground, another plastic sheet impregnated withTanglefoot® helped to increase trap catch. The distance between the traps was 15 m. The insectscaught were counted daily for 1 mo, and the traps were rotated once a week. Traps wererandomly placed within the plots, and 3 replicates per formulation were considered. Septaloaded with hexane served as controls. For the tests, formulations were designated in terms ofthe stereochemical purity of the pheromone used; and thus, for instance, a formulation Z,E/E,E95:5 refers to a bait containing a mixture of the two isomers (Z,E)-1 and (E,E)-1 (the lettersrefer in this order to the stereochemistry of the double bond at the alcohol side and the acidside, respectively) in 95:5 ratio.
J Chem Ecol (2006) 32: 2177–2190 2181
Statistical Analysis
Behavioral data were analyzed by the χ2 homogeneity test (P < 0.05). EAG depolarizationswere analyzed by Student’s t-test (P < 0.05). Field data were transformed (√x + 0.5) andanalyzed for significance via Student’s t-test (P < 0.05).
Results and Discussion
(Z)-3-Dodecenol (2) has been documented as a candidate trail-following semiochemical forseveral termite species (Argenti et al., 1994) as well as the major component of the trail-following pheromone of the fungus-growing termiteMacrotermes annandalei (Peppuy et al.,2001). In an attempt to develop a simple procedure of preparation of (Z)-3-dodecenol (2) thatcould be adapted and scaled up in Cuba for larger treatments, a similar procedure to thatdescribed by Lo et al. (1992) was developed. In our case, we protected 3-butyn-1-ol as thecorresponding MOM ether (3) instead of the tetrahydropyranyl (THP) derivative (Lo et al.,1992) since MOM compounds are simpler and provide NMR spectra that are easier tointerpret than THP ethers. Therefore, MOM-protected butynol 3 was alkylated with 1-bromooctane in THF–DMPU in the presence of LDA [the alkylation reaction with sodamideas base in ammonia (Lo et al., 1992) failed in our hands] to provide acetylenic derivative 4 in65% yield. When MeLi or n-BuLi was used as base, 1-dodecen-3-yne was obtained as by-
Natural extract
31.0 31.5 32.0 32.5 33.0
32.09
Retention time (min.)
Rel
ativ
e In
tens
ity (
%)
100
m/z60 80 100 120 140 160
10069.00
96.00
87.00 138.00 166.00 Rel
ativ
e In
tens
ity (
%)
Synthetic pheromone
10.0 20.0 30.0 40.0 50.0
32.15100
Retention time (min.)
Rel
ativ
e In
tens
ity (
%)
m/z60 80 100 120 140 160
100 69.00
96.00
87.00 138.00 166.00
Rel
ativ
e In
tens
ity (
%)
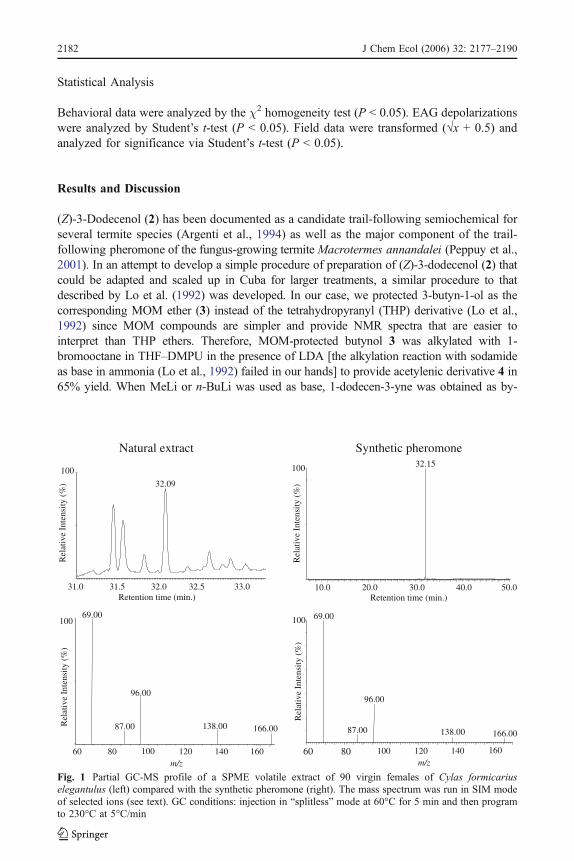
Fig. 1 Partial GC-MS profile of a SPME volatile extract of 90 virgin females of Cylas formicariuselegantulus (left) compared with the synthetic pheromone (right). The mass spectrum was run in SIM modeof selected ions (see text). GC conditions: injection in “splitless” mode at 60°C for 5 min and then programto 230°C at 5°C/min
2182 J Chem Ecol (2006) 32: 2177–2190

product in 10 – 30% yield. This compound has not hitherto been described in such a process,but a similar case was reported when the THP-protected 1-butynol was subjected to alkylationfor the synthesis of the sex pheromone of the tomato pest Scrobipalpuloides absoluta (Svátoset al., 1993). Hydrolysis of the MOM group followed by partial hydrogenation with Lindlarcatalyst/quinoline furnished dodecenol 2 in good overall yield (53.8%) and excellent
Fig. 2 Morphology of Cylas formicarius elegantulus antennae. (a) SEM micrograph of a male head. (b)SEM micrograph of a female head. (c) SEM view of a distal subsegment of a female antenna. (d) SEM viewof a distal subsegment of a male antenna. (e) High magnification of the club surface of a male antennashowing different types of sensilla. ST1: Sensilla trichoidea type 1; ST2: sensilla trichoidea type 2; SB1:sensilla basiconica type 1; SB2: sensilla basiconica type 2; SB3: sensilla basiconica type 3
J Chem Ecol (2006) 32: 2177–2190 2183

stereochemistry (Z/E 99:1). Esterification of alcohol 2 with crotonyl chloride/pyridine inCH2Cl2 (Heath et al., 1986) afforded the expected pheromone 1 in 57% yield.
Characterization of the Pheromone
Because of the expected low contents/release of pheromone in sweet potato weevil femalesof other origins (Heath et al., 1986), solid-phase microextraction (SPME) was consideredfor identifying the pheromone. The first report of the utilization of SPME in pheromoneanalysis was in monitoring the diel rhythm of emission of airborne volatiles from the sugarcane weevil Metamasius hemipterus (Malosse et al., 1995). Since then, other examples ofapplications have been reported (Frérot et al., 1997; Rochat et al., 2000; Peppuy et al.,2001).
In our case, analysis of an extract of volatiles from 90 virgin females, adsorbed for 24 hron a 100 μM fiber, by GC-MS under SIM conditions of ions of m/z 166, 138, 96, 87, 69confirmed the presence of the pheromone 1 when compared with an authentic sample(Fig. 1). The selected ions had been previously noted as major or characteristic fragments inthe ms of the synthetic pheromone (Lo et al., 1992). The amount of pheromone present inthe volatile extract was estimated as 1.8 ng, i.e., 20 pg of pheromone emitted per femaleevery 24 hr. Trials to detect the pheromone by rubbing the syringe over the abdomen ofvirgin females were unsuccessful. Likewise, an SPME extract of 30 female volatiles gaveno response when using the male antenna as detector [GC-electroantennographic detection
Fig. 3 Dose-response depolarizations of Cylas formicarius elegantulus male antennae to several amounts ofpheromone 1. Error bars are standard errors. Bars followed by the same letter are not significantly different(Student’s t-test, P < 0.05)
2184 J Chem Ecol (2006) 32: 2177–2190

(EAD)], probably as a result of the high threshold and low EAG amplitude shown by thisinsect.
Electrophysiology
Male and female antennae comprise a scape, a pedicel, and a flagellum with ninesubsegments. The first eight subsegments of the flagellum are short, whereas the last one isconsiderably larger. This last distal subsegment is club shaped in females and elongated inmales. The following types of sensilla were found on the male distal flagellum segment:sensilla trichoidea type 1 (ST1) as long hairs of 100–150 μm; sensilla trichoidea type 2(ST2) as short hairs of 50–60 μm; sensilla basiconica type 1 (SB1) as thick pegs of 20–25 μm; sensilla basiconica type 2 (SB2) as curved pegs of 10–15 μm, and sensillabasiconica type 3 (SB3) as thin and straight short pegs of 15–20 μm (Fig. 2). The sametypes were observed on female antennae, but ST1 are less abundant than on male antennae.Sensilla chaetica were also found on the first eight flagellum subsegments in both sexes.Although we did not run single sensillum recordings to confirm that sensilla trichoideahouse the olfactory receptor neurons for pheromone detection, we may assume that ST1 areprobably the most likely sites for pheromone detection for the following reasons: (1) thepheromone is emitted by females; (2) only males are attracted by the pheromone both in thelaboratory and in the field; (3) male antennae show sensitivity to the pheromone on EAGbut not female antennae; and (4) ST1 are more abundant in male antennae.
EAG responses of a male antenna to the synthetic pheromone 1 were generally small, asexpected from such a type of antenna, but typical depolarizations of 0.6–0.8 mV werenevertheless obtained with puffs on 1 μg of pheromone. A dose–response plot with puffson 0.001–1 μg of pheromone is shown (Fig. 3), with higher doses inducing higher
Fig. 4 Responses of Cylas formicarius elegantulus males to 10 ng of synthetic pheromone 1 at differenttimes of the scotophase compared with control (hexane). Bars followed by the same letter are notsignificantly different (χ2 homogeneity test, P < 0.05)
J Chem Ecol (2006) 32: 2177–2190 2185

depolarizations as expected. Female antennae did not show any response even at a 100-μgdose.
To determine the specificity of the crotonate ester on the antennal response, the formate,acetate, propionate, and butyrate esters of (Z)-3-dodecenol were prepared by standardmethods (not shown), and their possible electrophysiological activity was tested. None ofthe analogues elicited EAG responses from male antennae, which demonstrates the key roleplayed by the crotonate function on pheromone activity. In this regard, negligible attractantactivity in the field by the butyrate ester has also been reported (Heath et al., 1991).
Behavior
To assess the behavioral response of males to the synthetic pheromone, we developed adouble dual-choice olfactometer based on a four-armed bioassay designed by Vet et al.(1983) to measure olfactory responses of hymenopterous parasitoids. Preliminary experi-ments showed that the insects made the best choice between pheromone and blank vialsafter exposure for 3 min. We found that the difference in response to pheromone andcontrol generally increased with the time of the scotophase, being highest at the 8th hr (70%of insects walked to the pheromone vials vs. 30% that moved towards control vials)(Fig. 4). Nevertheless, the responses were significantly different from the 4th hr, indicatingthat the maximum activity of males was between the 4th and the 8th hr of scotophase.
As regards the effect of pheromone dose, low amounts of pheromone (0.1–1 ng) inducedno behavioral preference for any particular arm of the olfactometer. However, higher doses,particularly 50–1000 ng, elicited significantly higher responses (70 – 75%) on malesorienting to the two sources (Fig. 5). Higher doses (10,000 ng) inhibited male response. The
Fig. 5 Responses of Cylas formicarius elegantulus males to several pheromone doses at the 6th hr of thescotophase compared with control (hexane). Bars followed by the same letter are not significantly different(χ2 homogeneity test, P < 0.05)
2186 J Chem Ecol (2006) 32: 2177–2190

Fig. 6 Number of Cylas formicarius elegantulus males caught in traps baited with different pheromoneformulations. Bars containing the same letter are not significantly different according to the Student’s t-test(P < 0.05). Tests were performed in infested sweet potato fields in Santiago de las Vegas (Cuba) (July–August 2000, 2001)
J Chem Ecol (2006) 32: 2177–2190 2187

commercial pheromone (Pherobank, Wageningen, The Netherlands) induced males at thesame level of preference (70%) vs. control as our synthetic material (results not shown).
One drawback of the system could be the confined space in which insects are exposed tostimuli. This can be offset by using a sufficient number of individuals and replicates toobtain statistically significant responses. In addition, although in our olfactometer insectshad to choose between two pheromone vials and two blanks (50% choice); in principle, oursystem could also allow them to discriminate between one, two, or three odorssimultaneously. Moreover, the setup could determine the effect of the stimulus (attractionor repellency).
Field Tests
A number of reports on the attractiveness of the pheromone in the field have been described(Talekar and Lee, 1989; Jansson et al., 1990, 1992; Mason et al., 1990; Yasuda et al., 1992;Pillai et al., 1994). Here, we studied the effect of the stereomeric purity of the pheromone.We found in the first-year experiments that the formulation of the highest stereochemicalpurity was the most effective. The commercial sample (mixture of isomers Z,E and E,E in a98:2 ratio) caught more males than the formulation with the same mixture of isomers in a92:8 ratio, which in turn was more active than formulations with a blend of isomers in ca.40:60 ratio (Fig. 6).
In the following year, although formulations containing the Z,E isomer in the range 94 –98% were as active as the commercial pheromone, the bait containing a blend of the Z,Eand E,E isomers in a 40:60 ratio was again inferior. In both experiments, as notedpreviously by other authors, a clear correlation between the content of the Z,E isomer withactivity was apparent. The pheromone sample containing 5% of the Z,Z isomer induced nosynergistic or inhibitory effect, and the alcohol precursor of the pheromone was inactive.These results contrast with those of Jansson et al. (1992) who reported that pheromonepurities between 75% and 98% did not consistently affect weevil collections in traps,although some significant effects of purity were observed when trap counts were lower. Inline with our results, Yasuda et al. (1992) reported that the activity of the syntheticpheromone was not inhibited by the addition of up to 10% of the E,E isomer. We suggestthat the use of a stereochemically pure pheromone may not be necessary for future pestcontrol strategies, and that formulations containing the active Z,E isomer with 94–95%stereomeric purity may be preferable owing to their easier availability and lower cost.
In summary, the sex pheromone of feral sweet potato weevil of a Cuban population hasbeen confirmed as (Z)-3-dodecenyl (E)-2-butenoate, as previously reported from alaboratory-reared colony (Heath et al., 1986). The synthetic compound elicited electro-physiological and behavioral activity in the laboratory and was highly attractive in the field,highlighting its potential utilization for pest control on the island. In this context, it is worthnoting that the utilization of the pheromone in an IPM strategy in India has resulted in aconsiderable reduction of damage, leading to an increased production of marketable tubersby 53% (Pillai et al., 1993).
Acknowledgments The authors acknowledge M.E.C. for a predoctoral fellowship to T.S., SubdirecciónGeneral de Cooperación Internacional for a cooperative project with the University of La Habana, andCICYT (projects AGL 2003-06599-C02-01, PTR 1995-0656-OP) and Generalitat de Catalunya (2001SGR-00342) for financial support.
2188 J Chem Ecol (2006) 32: 2177–2190

References
AKAZAWA, T. L., URITANI, L., and KUBOTA, H. 1960. Isolation of ipomeamarone and two coumarinderivatives from sweet potato roots injured by the weevil Cylas formicarius elegantulus. Arch. Biochem.Biophys. 88:150–156.
ARGENTI, L., BELLINA, F., CARPITA, A., ROSSI, E., and ROSSI, R. 1994. Trail-following in termites:Stereoselective synthesis of (Z)-3-dodecen-1-ol, (3Z,6Z)-3,6-dodecadien-1-ol and (3Z,6Z,9Z)-3,6,9-dodecatrien-1-ol. Synth. Commun. 24:2281–2297.
CHALFANT, R. B., JANSSON, R. K., SEAL, D. R., and SCHALK, J. M. 1990. Ecology and management of sweetpotato insects. Annu. Rev. Entomol. 35:157–180.
FAO 1984. Food Production Yearbook. Food and Agriculture Organization, Rome.FRÉROT, B., MALOSSE, C., and CAIN, A. H. 1997. Solid-phase micro extraction (SPME): A new tool in
pheromone identification in Lepidoptera. J. High Resolut. Chromatogr. 20:340–342.GUERRERO, A., MURGÓ, R., and MARTORELL, X. 1986. An improved electroantennogram apparatus with a
new automatic sample injection system. Physiol. Entomol. 11:273–277.HEATH, R. R., COFFELT, J. A., SONNET, P. E., PROSHOLD, F. I., DUEBEN, B., and TUMLINSON, J. H. 1986.
Identification of sex pheromone produced by female sweetpotato weevil, Cylas formicarius elegantulus(Summers). J. Chem. Ecol. 12:1489–1503.
HEATH, R. R., COFFELT, J. A., PROSHOLD, F. I., SONNET, P. E., and TUMLINSON, J. H. 1988. (Z)-3-Dodecen-1-ol (E)-2-butenoate and its use in monitoring and controlling the sweetpotato weevil. US Patent4,732,756.
HEATH, R. R., COFFELT, J. A., PROSHOLD, F. I., JANSSON, R. K., and SONNET, P. E. 1991. Sex pheromone ofCylas formicarius: History and implications of chemistry in weevil management, pp. 79–97, in R. K.Jansson and K. V. Raman (eds.). Sweet Potato Pest Management. Oxford, New Delhi.
JANSSON, R. K., PROSHOLD, F. I., MASON, L. J., HEATH, R. R., and LECRONE, S. H. 1990. Monitoringsweetpotato weevil (Coleoptera: Curculionidae) with sex pheromone: Effects of dosage and age of septa.Trop. Pest Manage. 36:263–269.
JANSSON, R. K. 1991. Biological control of Cylas spp., pp. 169–201, in R. K. Jansson and K. V. Raman(eds.). Sweet Potato Pest Management. Oxford & IBH Publishing, New Delhi.
JANSSON, R. K. and RAMAN, K. V. (eds.). 1991. Sweet Potato Pest Management, pp. 1–457. Oxford & IBHPublishing, New Delhi.
JANSSON, R. K., MASON, L. J., HEATH, R. R., SORENSEN, K. A., HAMMOND, A. M., and ROBINSON, J. V.1992. Pheromone-trap monitoring system for sweet potato weevil (Coleoptera: Apionidae): Effects ofpheromone source and purity. Trop. Pest Manage. 38:196–200.
LO, C.-C., HUNG, M.-D., and LIU, C.-J. 1992. Replacement of carcinogenic alkylating agent ethylene oxidein the synthesis of (Z)-3-dodecen-1-yl (E)-2-butenoate, sex pheromone of sweet-potato weevil, Cylasformicarius elegantulus (Summers) and Cylas formicarius formicarius (F.). J. Chem. Ecol. 18:95–103.
MALOSSE, C., RAMIREZ-LUCAS, P., ROCHAT, D., and MORIN, J. 1995. Solid-phase microextraction, analternative method for the study of airborne insect pheromones (Metamasius hemipterus, Coleoptera,Curculionidae). J. High Resolut. Chromatogr. 18:669–670.
MARYANOFF, B. E., REITZ, A. B., and DUHL-EMSWILER, A. 1985. Stereochemistry of the Wittig reaction.Effect of nucleophilic groups in the phosphonium ylide. J. Am. Chem. Soc. 107:217–226.
MASON, L. J., JANSSON, R. K., and HEATH, R. R. 1990. Sampling rate of male sweetpotato weevils (Cylasformicarius elegantulus) (Summers) (Coleoptera: Curculionidae) to pheromone traps: Influence ofpheromone dosage and lure age. J. Chem. Ecol. 16:2493–2502.
PEPPUY, A., ROBERT, A., SEMON, E., GINIES, C., LETTERE, M., BONNARD, O., and BORDEREAU, C. 2001.(Z)-Dodec-3-en-1-ol, a novel termite trail pheromone identified after solid phase microextraction fromMacrotermes annandalei. J. Insect Physiol. 47:445–453.
PILLAI, K. S., RAJAMMA, P., and PALANISWAMI, M. S. 1993. New technique in the control of sweet potatoweevil using synthetic sex pheromone in India. Int. J. Pest Manag. 39:84–89.
PILLAI, K. S., PALANISWAMI, M. S., RAJAMMA, P., PREMKUMAR, T., and JAYAPRAKAS, C. A. 1994. Effect ofsynthetic sex pheromone on the activity of the sweet potato weevil. Entomon 19:119–123.
ROCHAT, D., RAMIREZ-LUCAS, P., MALOSSE, C., ALDANA, R., KAKUL, T., and MORIN, J.-P. 2000. Role ofsolid-phase microextraction of highly volatile pheromones of two Rhinoceros beetles Scapanes australisand Strategus aloeus (Coleoptera, Scarabeidae, Dynastinae). J. Chromatogr. A 885:433–444.
SATO, K., URITANI, L., and SAITO, T. 1981. Characterization of the terpene-inducing factor isolated from thelarvae of the sweet potato weevil Cylas formicarius Fabricius (Coleoptera: Brenthidae). Appl. Entomol.Zool. 16:103–112.
J Chem Ecol (2006) 32: 2177–2190 2189

SVÁTOS, A., ATTYGALLE, A. B., JHAM, G. N., FRIGHETTO, R. T. S., VILELA, E. F., SAMAN, D., andMEINWALD, J. 1993. Sex pheromone of tomato pest Scrobipalpuloides absoluta (Lepidoptera:Gelechiidae). J. Chem. Ecol. 19:735–750.
TALEKAR, N. S. and LEE, S. T. 1989. Studies on the utilization of a female sex pheromone for themanagement of sweet potato weevil, Cylas formicarius formicarius (Coleoptera: Curculionidae). Bull.Inst. Zool. Acad. Sin. 28:281–288.
VET, L. E. M., VAN LENTEREN, J. C., HEYMANS, M., and MEELIS, E. 1983. An airflow olfactometer formeasuring olfactory responses of hymenopterous parasitoids and other small insects. Physiol. Entomol.8:97–106.
YASUDA, K., SUGIE, H., and HEATH, R. R. 1992. Field evaluation of synthetic sex-attractant pheromone ofthe sweet-potato weevil Cylas formicarius Fabricius (Coleoptera: Brentidae). Nip. Oyo Dob. Kon. Gak.36:81–87. CA 117:207135 (Agrochem. Bioreg.).
2190 J Chem Ecol (2006) 32: 2177–2190




















