
Out of the Andes: patterns of diversification in clearwing butterflies

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers,academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research.
Temporal Occurrence of Two Morpho Butterflies (Lepidoptera:Nymphalidae): Influence of Weather and Food ResourcesAuthor(s): Geraldo Freire, Jr., André Rangel Nascimento , Ivan KonstantinovMalinov , and Ivone R. DinizSource: Environmental Entomology, 43(2):274-282. 2014.Published By: Entomological Society of AmericaURL: http://www.bioone.org/doi/full/10.1603/EN12352
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicatesyour acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

COMMUNITY AND ECOSYSTEM ECOLOGY
Temporal Occurrence of Two Morpho Butterflies (Lepidoptera:Nymphalidae): Influence of Weather and Food Resources
GERALDO FREIRE, JR.,1,2 ANDRE RANGEL NASCIMENTO,3 IVAN KONSTANTINOV MALINOV,4
AND IVONE R. DINIZ5
Environ. Entomol. 43(2): 274!282 (2014); DOI: http://dx.doi.org/10.1603/EN12352
ABSTRACT The seasonality of fruit-feeding butterßies is very well known. However, few studieshave analyzed the inßuence of climatic variables and resource availability on the temporal distribu-tions of butterßies. Morpho helenor achillides (C. Felder and R. Felder 1867) and Morpho menelauscoeruleus (Perry 1810) (Nymphalidae) were used as models to investigate the inßuences of climaticfactors and food resources on the temporal distribution of these Morphinae butterßies. Thesebutterßies were collected weekly from January 2005 to December 2006 in the Parque Nacional deBrasõlia (PNB). In total, 408 individuals were collected, including 274 of M. helenor and 134 of M.menelaus. The relative abundance of the two species was similar in 2005 (n! 220) and 2006 (n! 188).Of the variables considered, only the relative humidity and resource availability measured in termsof phenology of zoochorous fruits of herbaceous plants explained a large proportion of the variationin the abundance of these butterßies. Both of the explanatory variables were positively associated withthe total abundance of individuals and with the abundances ofM. helenor andM.menelaus consideredseparately.Thephenologyof anemochorous fruitswasnegativelyassociatedwithbutterßyabundance.The temporal distribution of the butterßies was better predicted by the phenology of the zoochorousfruits of herbaceous plants than by the climatic predictors.
KEY WORDS fruit-feeding Lepidoptera, plant phenology, gallery forest, seasonality
The term “temporal window” denotes the optimumperiod for resource availability, climatic conditions,and enemy-free space (Hunter and Lechowicz 1992).Previous studies conducted in the Cerrado biome in-dicate that the beginning of the wet season and thetransition between the wet and dry seasons are theperiods associated with the highest abundance of Lep-idoptera (Pinheiro et al. 2002, Silva et al. 2011a). Theabundance of the Nymphalidae of the Atlantic Foresthas been found to peak from September to March(Ribeiro et al. 2010). Climatic variation, natural ene-mies, and the availability of food resources are thethree most important predictors of lepidopteran abun-dance, including nymphalid abundance (Pinheiro andOrtiz 1992; Morais et al. 1999; Ribeiro et al. 2010, 2012;Silva et al. 2011a).
Body size is another important predictor of thetemporal distribution of the Nymphalidae because thesubfamilies of this taxon, which include larger species,are more prone to occur during a narrow time periodthan those dominated by smaller species (Ribeiro andFreitas 2011). A possible explanation of this differencecould be that larger butterßies require a higher energyintake to complete their life cycle (Garcõa-Barros2006). Therefore, larger species are more vulnerableto environmental constraints and stochastic events(Uehara-Prado et al. 2007). As a result, the temporaloccurrence of the larger species is more highly syn-chronous with periods of optimal food resources thanis the temporal occurrence of the smaller species (Ri-beiro and Freitas 2011).
Although the temporal distribution of these butter-ßies has been known for decades (DeVries et al. 1997,DeVries and Walla 1999, Brown and Freitas 2000), fewstudies have investigated how climatic factors (Brownand Freitas 2000, Ribeiro et al. 2010) and resourceavailability (Hamer et al. 2006) are associated withnymphalidsÕ temporal distributions. We know of noprevious studies that have quantitatively investigatedthe inßuence of food resources on the temporal oc-currence of these fruit-feeding butterßy species, es-pecially in the Cerrado biome.
The main objectives of the current study were: 1)to determine the temporal distribution of two speciesof fruit-feeding butterßies, Morpho helenor achillides
1 Programa de Pos-graduacao em Ecologia, Instituto de CienciasBiologicas, Universidade de Brasõlia!UnB.Universidade de Brasõlia;Campus Universitario Darcy Ribeiro-Bloco E, 1" Andar; Asa Norte,Brasõlia!DF, CEP. 70910-900, Brasil.
2 Corresponding author, e-mail: [email protected] Programa de Pos Graduacao em Ecologia e Recursos Naturais,
Universidade Federal de Sao Carlos-UFSCar; Rodovia WashingtonLuiz Km 235, Caixa Postal 676; Sao Carlos-SP, CEP. 13565-905, Brasil.
4 Centro de Estudos da Caatinga - CECAT, ICM-Bio; SCEN Trecho,Edifõcio Sede do Ibama, Bloco C, Brasõlia!DF, CEP.: 70818-900, Brasil.
5 Departamento de Zoologia, Instituto de Ciencias Biologicas, Uni-versidade de Brasõlia-UnB.Campus Universitario Darcy Ribeiro-BlocoE, 1" Andar; Asa Norte, Brasõlia!DF, CEP. 70910-900, Brasil.
0046-225X/14/0274!0282$04.00/0 ! 2014 Entomological Society of America

(C. Felder & R. Felder, 1867) and Morpho menelauscoeruleus (Perry, 1810) (Nymphalidae) (hereafter de-nominated as Morpho helenor and Morpho menelaus,respectively), in the gallery forest within the Cerradoand 2) to investigate how climatic factors (tempera-ture, relative humidity, rainfall, and wind speed) andthe fruit phenology of woody and herbaceous plantsare related to the temporal distribution of M. helenorandM.menelaus. If energy intake is important for largebutterßies (Garcõa-Barros 2006), our prediction is thatthe temporal distribution of both populations will bemore closely synchronized with fruit phenology thanwith climatic factors.
Materials and Methods
This study was conducted from January 2005 toDecember 2006 in a gallery forest located in ParqueNacional de Brasõlia (PNB), Brasõlia, Brazil (15" 42#56$S; 47" 55#22$ W). PNB is an important reserve of Cer-rado biome, composed of different types of Cerradovegetation and encompassing 30.566 ha of protectedarea. The climate in Cerrado domain is strongly char-acterized by dry winter and wet summer seasons; theprecipitation ranges between 1,400 and 1,500 mm an-nually, and this strong seasonality in climate functionsas a main determinant of phenological patterns ob-served in plants (Batalha et al. 1997, Batalha and Man-tovani 2000, Batalha and Martins 2004) and insects(Wolda 1988, Pinheiro et al. 2002, Silva et al. 2011).Gallery forest is a narrow strip of tropical forests thatoccur alongside rivers and streams (Oliveira-Filho etal. 1990). In central Brazil, the gallery forests are pre-dominantly surrounded by a matrix of Brazilian sa-vanna Cerrado (Fel"li 1995).
There is extensive literature about fruit phenologyof Cerrado biome plants, with the "rst studies datingfrom almost 30 yr ago (Mantovani and Martins 1988,Warming 1892, Miranda 1995, Batalha et al. 1997,Batalha and Mantovani 2000, Batalha and Martins2004, Silva et al. 2011b). Based on these 30 yr of studies,the phenological patterns are considered very consis-tent over the years at the Cerrado biome, which pos-sesses well marked dry and wet seasons. All of thosestudies indicate that the dry season months, a periodof high wind incidence, is the period of anemo- andautochorous dispersion. Zoochorous species, on thecontrary, fruited especially during the wettest periodof the year, when ßeshy fruits can be kept attractivefor a longer time (Mantovani and Martins 1988, Mi-randa 1995, Batalha et al. 1997, Batalha and Mantovani2000, Batalha and Martins 2004, Silva et al. 2011b).
In general, both gallery forest and Cerrado are verysimilar in their phenological patterns, despite soilmoisture variation; there are no signi"cant differencesbetween their peaks of fruit productions (Silva et al.2011b). Given the long period in which the fruit phe-nology has been studied in Cerrado biome (at least 30yr of data), added to the constancy of the resultsprovided by them, we agree with previous studies thatassumed the seasonal pattern of fruit phenology atCerrado matrix and gallery forest to be very constant
between years (Silva et al. 2011b), thus justifying theuse of current literature data as a good surrogate toassess the phenology of fruits available to Morphopopulations.
Information on the fruit phenology of woody andherbaceous plants was obtained in three steps. First,we focused on compiling information from a plantspecies list obtained from a previous forest inventoryat the study site (gallery forest of PNB; Guarino andWalter 2005; Silva-Junior and Sarmento 2009). Then,we searched for information on fruit phenologies (pe-riod of year) and types of fruits (zoochorous: animaldispersed and anemochorous: wind dispersed; Batalhaand Mantovani 2000, Munhoz and Fel"li 2005, Lenzaand Klink 2006) for those plant species previouslylisted at the PNB. Finally, with the list of plants andphenological information, we counted the number ofplant species with mature fruits each month. Thesemonthly values of plant species with zoochorous oranemochorous fruits were considered as two differentvariables. The maturation period of zoochorous fruitswas considered because they represent the principalfood source for both butterßy populations consideredin this study (DeVries et al. 1997). The phenology ofanemochorous fruits was considered as an additionalpredictor of the variation in the relative abundance ofthese butterßies.
All individuals of M. helenor and M. menelaus wereactively captured on 2-km transects (5 m in width)within the gallery forest. The collections were con-ducted once weekly over a 7-h period (10 a.m. to5 p.m.) and were performed by a single person(I.K.M.) throughout the study. Monthly, four ran-domly selected transects were used to search for but-terßies. All butterßies captured received an individualnumber(permanent inkpen)andwere released in thevicinity of the location from where they were origi-nally captured. Thus, the butterßies captured duringthe 4 wk represent the monthly total abundance ofeach species separately.
The genus Morpho is represented by 29 speciesbelonging to Nymphalidae (Lamas 2004, Cassilde et al.2012), recognized by their fruit-feeding behavior(DeVries et al. 1997) in the wet season (Ribeiro andFreitas 2011). These butterßies are endemics of Cen-tral and South America (Cassilde et al. 2012) and arean old acquaintance in the history of biology (Spix andMartius 1824, Darwin 1845, Agassiz and Agassiz 1868),mainly because of their blue, shiny colors that origi-nate from a prismatic phenomenon in their wing scales(Berthier et al. 2006, Weatherspoon et al. 2008). Dif-ferent aspects of their biology have been studied, suchas seasonality (Young 1975, Ribeiro and Freitas 2011,Grøtan et al. 2012), wing morphology and ßight be-havior (DeVries et al. 2010), phylogeny (Penz andDeVries 2002, Cassilde et al. 2012, Penz et al. 2012),geographical ranges (Young and Muyshondt 1972,Thomas 1991), and feeding behavior and biology(Young 1975, Constantino 1997, Constantino andCorredor 2004, Molleman et al. 2005, Guerra!Serrudoand e Ledezma!Arias 2008, Sourakov et al. 2012). Thespecies M. menelaus have a wide geographical range
April 2014 FREIRE ET AL.: TEMPORAL INFLUENCES OF TWO Morpho BUTTERFLIES 275

from north of Venezuela to south of Brazil (Favretto2012, Penz et al. 2012), their caterpillars are gre-garious (Guerra-Serrudo and Ledezma-Arias 2008)and polyphagous, feeding on Erythroxylum P. Browne(Erythroxylaceae), Inga Miller, Machaerium Persoon(Fabaceae), and Ormosia Jackson (Papilionaceae)(Ackery 1988, Constantino 1997, Guerra-Serrudo andLedezma-Arias 2008). The geographical range of M.helenor is wider thanM. menelaus, ranging from Mex-ico to the south of Brazil (Ritter et al. 2011, Penz et al.2012), their caterpillars are folivorous and polypha-gous, found on Erythrina L., Dalbergia L.F., Lon-chocarpus Kunth, Machaerium, Mucuna Adanson,Platymiscium Vogel, Pterocarpus Jacquin, SwartziaSchreber (Fabaceae), and Paragonia Bureau ex K.Schumann (Bignoniaceae) (Ackery 1988, Urich andEmmel 1991, Constantino and Corredor 2004). Amongthese host plants, the Erythroxylum, Dalbergia, andMachaerium were available in PNB (Roveratti 2008).Both Morpho species have understory ßight habits(DeVries et al. 2010), and they feed in rotting fruitsdropped on the ground.
Daily (7 a.m. to 6 p.m.) weather information (tem-perature, relative air humidity, rainfall, and windspeed) was obtained from the Embrapa Cerrados me-teorological station (15" 35#30$ S; 47" 42#30$ W), andthe monthly mean value of each variable was consid-ered for the statistical analysis. This meteorologicalstation is located inside the PNB %4 km from thelocation where the monthly transects were estab-lished.
A StudentÕs t-test was used to compare the relativespecies abundances and the differences between thetwo years studied (2005 and 2006). Circular analyseswere used to assess the temporal distribution of M.helenor and M. menelaus and to determine the de-gree of temporal concentration of these butterßiesthroughout the year (Zar 2010). In these analyses, themonth of capture was transformed to degrees, and themean vector (!) and its length (r) were then mea-sured. The length of the mean vector ranges from 0 to1 and describes the temporal concentration of thespecies over the year. Values near 0 indicate that theprobability of capture in each month of the year isthe same, whereas values close to 1 indicate that allcaptures were concentrated close to the same monthof the year.
A generalized linear model (GLM) with a Poissonerror distribution was used to investigate the inßuenceof weather (temperature medianÑTmed; relative airhumidityÑHumid; rainfallÑPrec.; and wind speedÑWind) and fruit phenology (number of herbaceousÑZooherb; woodyÑZoowood plants with maturezoochorous; and anemochorousÑane fruits) on therelative abundance ofM. helenor andM.menelaus.TheAkaike information criteria (AIC) is a classical mea-sure of the "t of a model. AIC is useful because itexplicitly penalizes any superßuous parameter in themodel, by adding 2 (P & 1) to the deviance, where Pis the number of parameters in a model. When com-paring two models, the smaller values indicate a better"t. If the difference between two AIC values is '2, the
lower AIC is a signi"cant improvement in parsimony(Logan 2010). We used AIC to verify the informationcontent of each statistical model.
First, we constructed exploratory models consider-ing, separately, each of the seven environmental vari-ables (weather and fruit phenology), as predictors ofbutterßy relative abundance and for each species sep-arately. Then, we formulated a second model, nowconsidering the three most important predictors,based on variance explanation values (air humidityand zoochorical herbaceous plant richness) and bio-logical meaning (zoochorical woody plant richness)and applied the hierarchical partitioning of variancetest, to verify the proportion of variance explainedindependently and jointly by each variable (Chevanand Sutherland 1991, Mac Nally 2000). This methodallows identi"cation of variables whose independentcorrelation with response variable is high, in contrastto those variables that have small independent effectsbut have a high correlation with the response variable,resulting from joint correlation with other predictors(Logan 2010). These analyses produce the indepen-dent contribution values (I) of each predictor variableand its conjoint contribution in presence of all otherpredictors (J; Chevan and Sutherland 1991). In addi-tion, we use a randomization procedure (100 times)on the independent contributions (I) of each predic-tor variable to evaluate whether the magnitude of avariableÕs contribution is great enough to be consid-ered as important. Then, the randomized outcomeswere used to calculate the Z-scores for each predictorvariable and the signi"cance was tested (Z " 1.65 atthe 95% level; Mac Nally 2002, Logan 2010).
The number of individuals captured in 2005 (n !220) and 2006 (n! 188) was similar (t! 0.36; df ! 43;P! 0.71; Fig. 2). For this reason, the abundance datafor both years were lumped together to facilitate theinterpretation of the results.
The correlation between two or more predictorvariables is known as multicollinearity and that couldbe a problem in model "tting. Some measures liketolerance analysis and variance inßation factor (VIF)could be used to diagnose this problem. Tolerance(1 ( r2) is a measure of a degree of collinearity andvalues lesser than 0.2 should be analyzed with con-cern, and values )0.1 indicate strong collinearity be-tween predictors. VIFs are the inverse of tolerance,and thus values '5 indicate collinearity (Logan 2010).
These measures were calculated, separately, to allweather and phenological predictors to verify the col-linearity between them. The values of tolerance forweather (tmed: 0.54; Prec. 0.31; wind: 0.67; and airhumid: 0.35) and phenological (ane.: 0.76; zooherb:0.49; and zoowood: 0.51) and the VIF values (tmed:1.85; Prec.: 3.31; wind: 1.48; air humid: 2.8; ane.: 1.3;zooherb: 2.02; and zoowood:1.93) indicate small ef-fects of those variables on the model performance.Thus, all predictor variables were considered in thestatistical models.
The statistical models were constructed using Rstatistical software (R Development Core Team2013), and a signi"cance level of P # 0.05 was used.
276 ENVIRONMENTAL ENTOMOLOGY Vol. 43, no. 2

The distribution of the residuals was graphically eval-uated to verify that each model satis"ed the assump-tions of normality and homoscedasticity. The circularstatistics were calculated using the software Oriana4.01 (Kovach Computing, Aberystwyth, United King-dom).
Results
The climatic diagram (Fig. 1) showed that the wetseason goes from October to April, and the dry seasonsfrom May to September. In total, 408 individuals werecaptured and subsequently released during this study.The speciesM. helenor (n! 274) was more abundantthanM.menelaus (n! 134; t! 2.18; df ! 22; P! 0.03),representing 67 and 33% of the total, respectively.
Bothbutterßieswereunevenlydistributed through-out the year, with two peaks of relative abundance.The "rst peak occurred in March!April (n! 141), thesecond in November!December (n! 125; Fig. 2). The
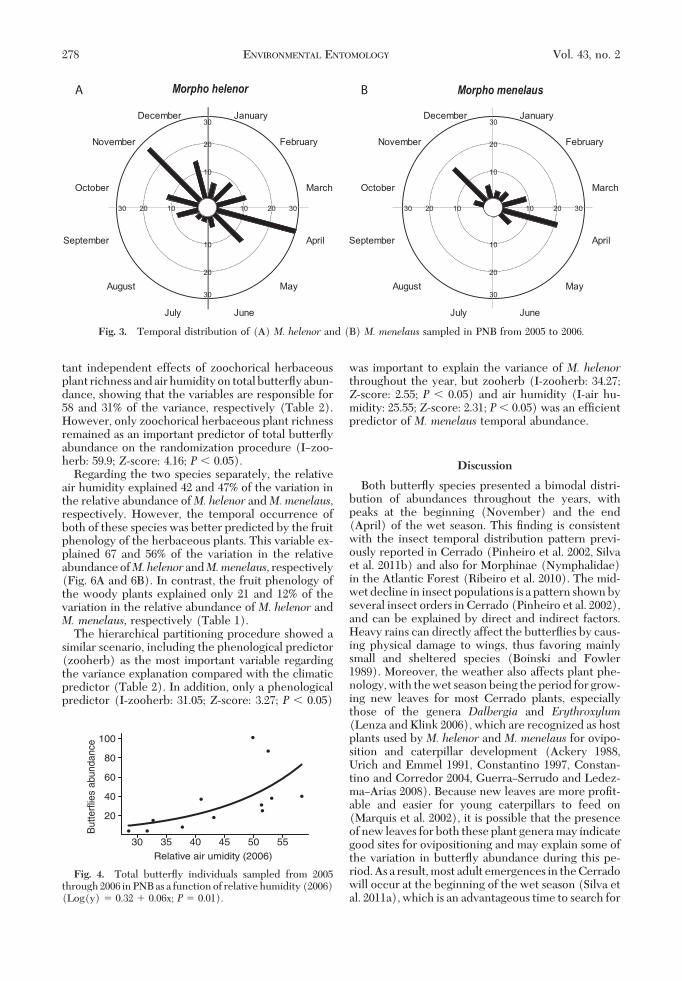
length of the mean vector indicates that the distribu-tion ofM. menelaus (Fig. 3B) is more concentrated inthese periods (r! 0.87) thanM. helenor (r! 0.41; Fig.3A).
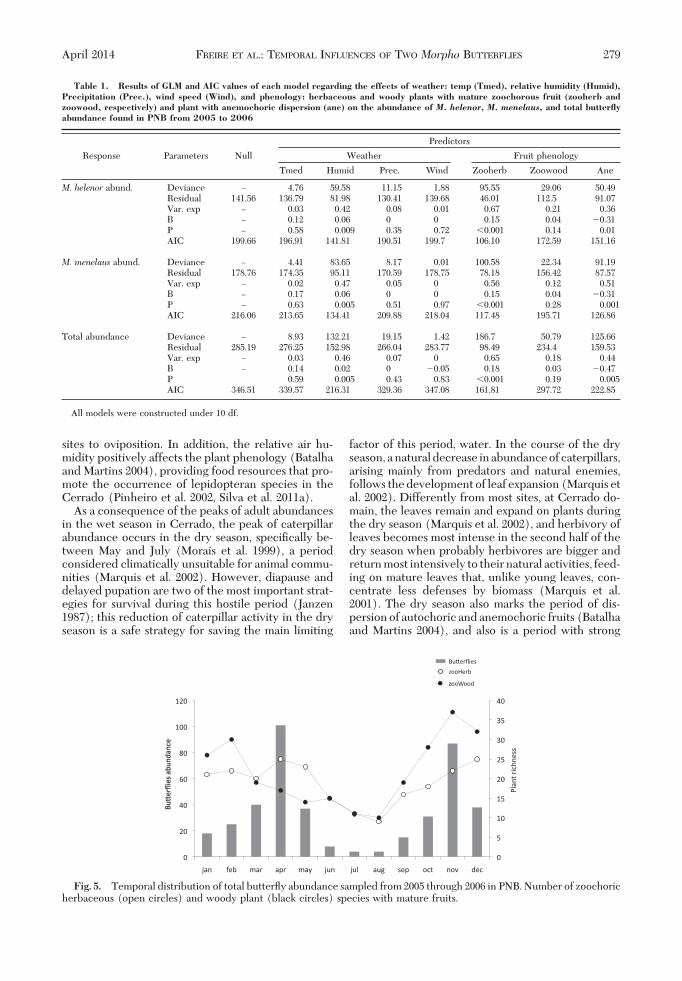
Analyzing separately all predictor variables, the rel-ative air humidity explained %46% of the variation intotal butterßy abundance and was the only climaticpredictor associated with the total number of butter-ßies captured (log(y) ! 0.32 & 0.06x; P! 0.01); thus,more individuals were captured during wet months(Fig. 4). The richness of herbaceous plants with ma-ture zoochorous fruits and the richness of anemo-choric plants explained 65 and 44% of the variation inthe total butterßy abundance, respectively (Table 1).In contrast, the number of woody plants withzoochorous fruits was not an effective predictor ofbutterßy temporal distribution. This variable ex-plained only 18% of the variation in the temporalabundance of the butterßies (Fig. 5). In a similar way,the hierarchical partitioning analyses indicate impor-
0
10
20
30
40
50
0
20
40
60
80
100
300
C mm
J F M A M J J A S O N D
Fig. 1. Climatic diagram constructed from data obtained from 2005 and 2006 at the Climatic Station of Embrapa Cerrados,Brasõlia, DF, Brazil (15" 35#30$ S, 47" 42#30# W, 1,000 m a.s.l.). The gray line is referent to monthly average temperature; theblack line is referent to monthly average precipitation. Mean annual temperature ! 22.4"C, total annual precipitation ! 1,201mm.
Fig. 2. Number of individuals of M. helenor and M. menelaus sampled in PNB from 2005 through 2006.
April 2014 FREIRE ET AL.: TEMPORAL INFLUENCES OF TWO Morpho BUTTERFLIES 277
tant independent effects of zoochorical herbaceousplant richness andairhumidityon totalbutterßyabun-dance, showing that the variables are responsible for58 and 31% of the variance, respectively (Table 2).However, only zoochorical herbaceous plant richnessremained as an important predictor of total butterßyabundance on the randomization procedure (I!zoo-herb: 59.9; Z-score: 4.16; P ) 0.05).
Regarding the two species separately, the relativeair humidity explained 42 and 47% of the variation inthe relative abundance ofM. helenor andM. menelaus,respectively. However, the temporal occurrence ofboth of these species was better predicted by the fruitphenology of the herbaceous plants. This variable ex-plained 67 and 56% of the variation in the relativeabundance ofM.helenor andM.menelaus, respectively(Fig. 6A and 6B). In contrast, the fruit phenology ofthe woody plants explained only 21 and 12% of thevariation in the relative abundance of M. helenor andM. menelaus, respectively (Table 1).
The hierarchical partitioning procedure showed asimilar scenario, including the phenological predictor(zooherb) as the most important variable regardingthe variance explanation compared with the climaticpredictor (Table 2). In addition, only a phenologicalpredictor (I-zooherb: 31.05; Z-score: 3.27; P ) 0.05)
was important to explain the variance of M. helenorthroughout the year, but zooherb (I-zooherb: 34.27;Z-score: 2.55; P ) 0.05) and air humidity (I-air hu-midity: 25.55; Z-score: 2.31; P) 0.05) was an ef"cientpredictor of M. menelaus temporal abundance.
Discussion
Both butterßy species presented a bimodal distri-bution of abundances throughout the years, withpeaks at the beginning (November) and the end(April) of the wet season. This "nding is consistentwith the insect temporal distribution pattern previ-ously reported in Cerrado (Pinheiro et al. 2002, Silvaet al. 2011b) and also for Morphinae (Nymphalidae)in the Atlantic Forest (Ribeiro et al. 2010). The mid-wet decline in insect populations is a pattern shown byseveral insect orders in Cerrado (Pinheiro et al. 2002),and can be explained by direct and indirect factors.Heavy rains can directly affect the butterßies by caus-ing physical damage to wings, thus favoring mainlysmall and sheltered species (Boinski and Fowler1989). Moreover, the weather also affects plant phe-nology, with the wet season being the period for grow-ing new leaves for most Cerrado plants, especiallythose of the genera Dalbergia and Erythroxylum(Lenza and Klink 2006), which are recognized as hostplants used by M. helenor and M. menelaus for ovipo-sition and caterpillar development (Ackery 1988,Urich and Emmel 1991, Constantino 1997, Constan-tino and Corredor 2004, Guerra!Serrudo and Ledez-ma!Arias 2008). Because new leaves are more pro"t-able and easier for young caterpillars to feed on(Marquis et al. 2002), it is possible that the presenceof new leaves for both these plant genera may indicategood sites for ovipositioning and may explain some ofthe variation in butterßy abundance during this pe-riod. As a result, most adult emergences in the Cerradowill occur at the beginning of the wet season (Silva etal. 2011a), which is an advantageous time to search for
30 35 40 45 50 55
20
40
60
80
100
Relative air umidity (2006)
But
terfl
ies
abun
danc
e
Fig. 4. Total butterßy individuals sampled from 2005through 2006 in PNB as a function of relative humidity (2006)(Log(y) ! 0.32 & 0.06x; P ! 0.01).
Morpho helenor
January
February
March
April
May
JuneJuly
August
September
October
November
December
30 30
30
30
20 20
20
20
10 10
10
10
A
January
February
March
April
May
JuneJuly
August
September
October
November
December
30 30
30
30
20 20
20
20
10 10
10
10
B Morpho menelaus
Fig. 3. Temporal distribution of (A) M. helenor and (B) M. menelaus sampled in PNB from 2005 to 2006.
278 ENVIRONMENTAL ENTOMOLOGY Vol. 43, no. 2
sites to oviposition. In addition, the relative air hu-midity positively affects the plant phenology (Batalhaand Martins 2004), providing food resources that pro-mote the occurrence of lepidopteran species in theCerrado (Pinheiro et al. 2002, Silva et al. 2011a).
As a consequence of the peaks of adult abundancesin the wet season in Cerrado, the peak of caterpillarabundance occurs in the dry season, speci"cally be-tween May and July (Morais et al. 1999), a periodconsidered climatically unsuitable for animal commu-nities (Marquis et al. 2002). However, diapause anddelayed pupation are two of the most important strat-egies for survival during this hostile period (Janzen1987); this reduction of caterpillar activity in the dryseason is a safe strategy for saving the main limiting
factor of this period, water. In the course of the dryseason, anaturaldecrease inabundanceofcaterpillars,arising mainly from predators and natural enemies,follows the development of leaf expansion (Marquis etal. 2002). Differently from most sites, at Cerrado do-main, the leaves remain and expand on plants duringthe dry season (Marquis et al. 2002), and herbivory ofleaves becomes most intense in the second half of thedry season when probably herbivores are bigger andreturn most intensively to their natural activities, feed-ing on mature leaves that, unlike young leaves, con-centrate less defenses by biomass (Marquis et al.2001). The dry season also marks the period of dis-persion of autochoric and anemochoric fruits (Batalhaand Martins 2004), and also is a period with strong
Table 1. Results of GLM and AIC values of each model regarding the effects of weather: temp (Tmed), relative humidity (Humid),Precipitation (Prec.), wind speed (Wind), and phenology: herbaceous and woody plants with mature zoochorous fruit (zooherb andzoowood, respectively) and plant with anemochoric dispersion (ane) on the abundance of M. helenor, M. menelaus, and total butterflyabundance found in PNB from 2005 to 2006
Response Parameters Null
Predictors
Weather Fruit phenology
Tmed Humid Prec. Wind Zooherb Zoowood Ane
M. helenor abund. Deviance ! 4.76 59.58 11.15 1.88 95.55 29.06 50.49Residual 141.56 136.79 81.98 130.41 139.68 46.01 112.5 91.07Var. exp ! 0.03 0.42 0.08 0.01 0.67 0.21 0.36* ! 0.12 0.06 0 0 0.15 0.04 (0.31P ! 0.58 0.009 0.38 0.72 )0.001 0.14 0.01AIC 199.66 196.91 141.81 190.51 199.7 106.10 172.59 151.16
M. menelaus abund. Deviance ! 4.41 83.65 8.17 0.01 100.58 22.34 91.19Residual 178.76 174.35 95.11 170.59 178.75 78.18 156.42 87.57Var. exp ! 0.02 0.47 0.05 0 0.56 0.12 0.51* ! 0.17 0.06 0 0 0.15 0.04 (0.31P ! 0.63 0.005 0.51 0.97 )0.001 0.28 0.001AIC 216.06 213.65 134.41 209.88 218.04 117.48 195.71 126.86
Total abundance Deviance ! 8.93 132.21 19.15 1.42 186.7 50.79 125.66Residual 285.19 276.25 152.98 266.04 283.77 98.49 234.4 159.53Var. exp ! 0.03 0.46 0.07 0 0.65 0.18 0.44* ! 0.14 0.02 0 (0.05 0.18 0.03 (0.47P 0.59 0.005 0.43 0.83 )0.001 0.19 0.005AIC 346.51 339.57 216.31 329.36 347.08 161.81 297.72 222.85
All models were constructed under 10 df.
Fig. 5. Temporal distribution of total butterßy abundance sampled from 2005 through 2006 in PNB. Number of zoochoricherbaceous (open circles) and woody plant (black circles) species with mature fruits.
April 2014 FREIRE ET AL.: TEMPORAL INFLUENCES OF TWO Morpho BUTTERFLIES 279

climatic constraint to adult butterßies (Wolda 1988,Silva et al. 2011a). Conversely, the wet season marksthe period of ripening of zoochoric fruit (Batalha andMantovani 2000), the principal food resource for thespecies considered in this study, and correspondswith the peak of adult occurrence of these Morphi-nae species.Morpho species show a clear vertical strati"cation in
ßight behavior. The type of ßight behavior and itsheight has strong phylogenetic relationships (Penzand DeVries 2002, DeVries et al. 2010). Wing shape isan important feature that determines whetherMorphospecies perform gliding ßights, patrolling over the can-opy, or ßapping ßights in the understory (DeVries etal. 2010). Both species examined by this study char-acteristically perform ßapping ßights in the under-story, consistent with their status as members of theachilles group phylogenetic clade (see Penz andDeVries 2002, DeVries et al. 2010). Of the variables
analyzed, the number of herbaceous species with ripefruit was the best predictor of butterßy occurrence(Tables 1 and 2). The period of ripening of zoochoricfruit in woody plants (November to February) wasnarrow relative to the same ripening period in her-baceous plants (November to May; Batalha and Man-tovani 2000). This narrow period could explain theabsence of relationships between the fruit phenologyof the woody plants and the occurrence of both but-terßy species. However, this observation does not im-ply that this feature does not affect the temporal dy-namics of these butterßy populations.
In view of the ßapping ßight behavior ofM. helenorand M. menelaus, it is possible that herbaceous plantswith ripe zoochoric fruits are a pro"table food re-source contributing to the energy intake of these but-terßies. Given the body sizes of the species (Ribeiroand Freitas 2011) and the energy intake required tocomplete the life cycle (Garcõa-Barros 2006), tempo-ral asynchrony with the food resources would be moreharmful to Morphinae than to their smaller counter-parts. Thus, our data support the hypothesis that thesespecies have been evolutionarily selected to occursynchronously with the availability of food resourcesin the understory, measured in this study in terms ofa period during which most herbaceous plant speciesfurnish ripe fruits.
The more seasonal a site is, the more synchronousthe phenology of species in the community is (Wolda1988). Our study reveals that in a strongly seasonalarea in the core of the Cerrado domain, the temporalvariations of food resources play an important role inthe temporal distribution of two species of fruit-feed-ing butterßies. The beginning of the wet season in theCerrado is marked by a higher development of repro-ductive and vegetative structures in plants. The ßushof zoochorous fruits, especially (but not exclusively)from herbaceous plants, is an important determinantof abundance for ßapping-ßight butterßies in the un-
Table 2. Results of hierarchical partitioning of variance, usingthe three GLM models testing the independent (I) and joint (J)effects of weather: relative humidity (Humid) and phenological:herbaceous and woody plants with mature zoochorous fruits (zoo-herb and zoowood, respectively) variables on the M. helenor, M.menelaus, and total butterfly abundance
Responsevariables
PredictorsTest parameters
I J Z.score!I Sign.
M. helenor abund. Zooherb 31.06 16.71 2.7 *Zoowood 4.94 9.58 (0.22 nsHumid. 13.17 16.74 0.72 ns
M. menelaus abund. Zooherb 33.36 16.92 2.36 *Zoowood 4.06 7.11 (0.41 nsHumid 24.73 17.08 1.98 *
Total abundance Zooherb 59.81 33.53 2.45 *Zoowood 8.67 16.72 (0.25 nsHumid 32.31 33.78 1.15 ns
The I values were then transformed to Z-scores (Z.score!I) to testthe signi"cance of each predictor variable. The sign (*) indicatessigni"cance values under P ) 0.05 levels and (ns) nonsigni"cant.
10 15 20 25
10
20
30
40
50
60
zoochorical plant richness
M. h
elen
or a
bund
ance
A
10 15 20 25
0
10
20
30
40
50
60
zoochorical plant richness
M. m
enel
aus
abun
danc
e
B
Fig. 6. Relationship between number of herbaceous species with ripe fruits and relative abundance of (A) M. helenor(Log(y) ! 0.18 & 0.14x; P ) 0.001) and (B) M. menelaus (Log(y) ! (2.84 & 0.24x; P ) 0.001) sampled from 2005 through2006 in PNB.
280 ENVIRONMENTAL ENTOMOLOGY Vol. 43, no. 2

derstory of Cerrado. These fruits are the main sourceof food when these species are searching for newleaves on plants that will nourish their offspring. Thisstudy reveals new possibilities in the research on tem-poral dynamics of communities of Cerrado butterßies,focusing more on the feeding niches of species than onindirect factors, such as climate.
Acknowledgments
The National Council for Scienti"c and TechnologicalDevelopment (CNPq) granted scholarships to I.K.M. andI.R.D. Coordination of Improvement of Higher Education(CAPES) granted scholarships to G.F.Jr and A.R.N. Supple-mentary "nancial resources were provided by PRONEX/CNPq/FAPDF. Finally, we would like to express our deepgratitude to Juliano Carregaro for his valuable suggestionsand contributions to the previous versions of this manuscript.
References Cited
Ackery, P. R. 1988. Hostplants and classi"cation: a review ofnymphalid butterßies. Biol. J. Linn. Soc. 33: 95!203.
Agassiz, L., and E.C.C. Agassiz. 1868. A journey in Brazil byProfessor and Mrs, p. 540. Louis Agassiz. 2nd ed. Ticknorand Fields, Boston, MA.
Batalha,M. A., andW.Mantovani. 2000. Reproductive phe-nological patterns of cerrado plant species at the Pe-de-Gigante Reserve (Santa Rita do Passa Quatro, SP, Brazil):a comparison between the herbaceous and woody ßoras.Rev. Bras. Biol. 60: 129!145.
Batalha, M. A., and F. R. Martins. 2004. Reproductive phe-nology of the cerrado plant community in Emas NationalPark (central Brazil). Aust. J. Bot. 52: 149!161.
Batalha, M. A., S. Aragaki, and W. Mantovani. 1997. Varia-coes fenologicas das especies do cerrado em Emas - Pi-rassununga, SP. Acta Botanica Brasilica 11: 61!78.
Berthier, S., E. Charron, and J. Boulenguez. 2006. Morpho-logical structure and optical properties of the wings ofMorphidae. Insect Sci. 13: 145!158.
Boinski, S., and N. L. Fowler. 1989. Seasonal patterns in atropicallowland forest. Biotropica 21: 223!233.
Brown Jr., K. S., and A.V.L. Freitas. 2000. Atlantic forestbutterßies: indicators for landscape conservation. Biotro-pica 32: 934!956.
Cassilde,C., P.Blandin, and J. F. Silvain. 2012. Phylogeny ofthe genus Morpho Fabricius 1807: insights from two mi-tochondrial genes (Lepidoptera: Nymphalidae). Ann.Soc. Entomol. Fr. 48: 173!188.
Chevan, A., and M. Sutherland. 1991. Hierarchical parti-tioning. Am. Statist. 45: 90!96.
Constantino, L. M. 1997. Natural history, immature stagesand hostplants of Morpho amathonte from western Co-lombia (Lepidoptera: Nymphalidae: Morphinae).Trop.Lepidoptera 8: 75!80.
Constantino, L.M., andG.Corredor. 2004. The biology andmorphology of the early stages of Morpho macrophthal-mus and Morpho peleides telamon (Nymphalidae: Mor-phinae) from Western Colombia. Boletõn Cientõ"co. Mus.Hist. Nat. Univ. Caldas. 8: 201!209.
Darwin, C. 1845. Journal of researches into the natural his-tory and geology of the countries visited during the voy-age of H.M.S. Beagle round the world, under the Com-mand of Capt, p. 500. In R. N. Fitz Roy 2nd ed. JohnMurray, London, United Kingdom.
DeVries, P. J., and T. R. Walla. 1999. Species diversity inspatial and temporal dimensions of fruit-feeding butter-
ßies from two Ecuadorian rainforests. Biol. J. Linn. Soc.68: 333!353.
DeVries, P. J., D. Murray, and R. Lande. 1997. Species di-versity in vertical, horizontal, and temporal dimensions ofa fruit-feeding butterßy community in an Ecuadorianrainforest. Biol. J. Linn. Soc. 62: 343!364.
DeVries, P. J., C. M. Penz, and R. I. Hill. 2010. Verticaldistribution, ßight behaviour and evolution of wing mor-phology in Morpho butterßies. J. Anim. Ecol. 79: 1077!1085.
Favretto, M. 2012. Borboletas e mariposas (Insecta: Lepi-doptera) do municõpio de Joacaba, Estado de Santa Ca-tarina, Brasil. EntomoBrasilis 5: 167!169.
Felfili, J. M. 1995. Diversity, structure, and dynamics of agallery forest in central Brazil. Vegetatio 117: 1!15.
Garcıa-Barros, E. 2006. Within and between species scalingin the weight, water, carbon and nitrogen contents of eggsand neonate larvae of twelve satyrine butterßies (Lepi-doptera: Nymphalidae). Eur. J. Entomol. 103: 559!568.
Grøtan, V., R. Lande, S. Engen, B. E. Sæther, and P. J.DeVries. 2012. Seasonal cycles of species diversity andsimilarity in a tropical butterßy community. J. Anim. Ecol.81: 714!723.
Guarino, G.S.E., and T.M.B. Walter. 2005. Fitossociologiadedois trechos inundaveisdeMatasdeGalerianoDistritoFederal, Brasil. Acta Botanica Brasilica 19: 431!442.
Guerra-Serrudo, J. F., and J. Ledezma-Arias. 2008. Biologiay morfologia deMorpho menelaus godartii (Lepidoptera:Nymphalidae: Morphinae) en el Parque Nacional Co-tapata (Bolivia). Ecologõa en Bolivia 43: 16!28.
Hamer, K. C., J. K. Hill, S. Benedick, N.Mustaffa, V. K. Chey,and M. Maryati. 2006. Diversity and ecology of carrion-and fruit-feeding butterßies in Bornean rain forest. Trop.Ecol. 22: 25!33.
Hunter, A. F., and M. J. Lechowicz. 1992. Foliage qualitychanges during canopy development of some northernhardwood trees. Oecologia 89: 316!323.
Janzen, D. H. 1987. How moths pass the dry season in aCosta Rican dry forest. Int. J. Trop. Insect Sci. 8: 489!500.
Lamas G. 2004. Tribe Morphini, pp. 192!201. In: G. Lamas(ed.), Checklist: Part 4A. Hesperoidea, Papilionoidea.Nymphalidae. Morphinae. Atlas of Neotropical Lepi-doptera.vol. 5. A.J.B. Heppner Ed., Association for Neotro-pical Lepidoptera, Scienti"c Publisher, Gainesville, FL.
Lenza, E., and C. A. Klink. 2006. Comportamento fenologicode especies lenhosas em um cerrado sentido restrito deBrasõlia, DF. Revista Brasileira de Botanica 29: 627!638.
Logan,M. 2010. BiostatisticaldesignanalysisusingR: aprac-tical guide. Wiley-Blackwell, Oxford, United Kingdom.
Mac Nally, R. 2000. Regression and model building in con-servation biology, biogeography and ecology: the distinc-tion between and reconciliation of ÔpredictiveÕ and Ôex-planatoryÕ models. Biodivers. Converv. 9: 655!671.
Mac Nally, R. 2002. Multiple regression and inference inecology and conservation biology: further comments onidentifying important predictor variables. Biodivers. Con-verv. 8: 1397!1401.
Mantovani,W., and F. R.Martins. 1988. Variacoes fenologi-cas das especies do cerrado da Reserva Biologica de MojiGuacu, estado de Sao Paulo. Revista Brasileira deBotanica 11: 101!112.
Miranda, I. S. 1995. Fenologia do estrato arboreo de umacomunidade de Alter do Chao, PA, Revista Brasileira deBotanica 18: 235!240.
Marquis, R. J., H. C. Morais, and I. R. Diniz. 2001. Patternsand correlates of interspeci"c variation in foliar insectherbivory and pathogen attack in Brazilian cerrado. J.Trop. Ecol. 17: 127!148.
April 2014 FREIRE ET AL.: TEMPORAL INFLUENCES OF TWO Morpho BUTTERFLIES 281

Marquis, R. J., H. C. Morais, and I. R. Diniz. 2002. Interac-tions among cerrado plants and their herbivores: uniqueor typical?pp. 306!328. InOliveiraP. S.,Marquis,R. J.TheCerrados of Brazil: Ecology and Natural History of aNeotropical Savanna. Columbia University Press, NewYork, NY.
Molleman, F., H.W. Krenn, M. E. Van Alphen, P. M. Brake-field, P. J. DeVries, and B. J. Zwann. 2005. Food intakeof fruit-feeding butterßies: evidence for adaptive varia-tion in proboscis morphology. Biol. J. Linn. Soc. 86: 333!343.
Morais,H.C., I.R.Diniz, andD.M.S. Silva. 1999. Caterpillarseasonality in a central Brazilian cerrado. Rev. Biol. Trop.47: 1025!1033.
Munhoz,C.B.R., and J.M.Felfili. 2005. Fenologia do estratoherbaceo-subarbustivo de uma comunidade de camposujo na Fazenda Agua Limpa no Distrito Federal, Brasil.Acta Botanical Brasilica 19: 979!988.
Oliveira-Filho, A. T., J. A. Ratter, and G. J. Shepherd. 1990.Floristic composition and community structure of a cen-tral Brazilian gallery forest. Flora 184: 103!117.
Penz, C. M., and P. J. DeVries. 2002. Phylogenetic analysisof Morpho butterßies (Nymphalidae, Morphinae ): im-plications for classi"cation and natural history. Am. Mus.Novit. 3374: 1!33.
Penz, C. M., P. J. DeVries, and N. Wahlberg. 2012. Diver-si"cation of Morpho butterßies (Lepidoptera, Nympha-lidae): a re-evaluation of morphological characters andnew insight from DNA sequence data. Syst. Entomol. 37:670!685.
Pinheiro, C.E.G., and J.V.C. Ortiz. 1992. Communities offruit-feeding butterßies along a vegetation gradient inCentral Brazil. J. Biogeogr. 19: 505!511.
Pinheiro, F., I. R. Diniz, D. Coelho, and M.P.S. Bandeira.2002. Seasonal pattern of insect abundance in the Bra-zilian cerrado. Austral Ecol. 27: 132!136.
RDevelopment Core Team. 2013. R: a language and environ-ment for statistical computing. R Foundation for StatisticalComputing, Vienna, Austria. (www.R-project.org).
Ribeiro, D. B., and A.V.L. Freitas. 2011. Large-sized insectsshow stronger seasonality than small-sized ones: a casestudy of fruit-feeding butterßies. Biol. J. Linn. Soc. 104:820!827.
Ribeiro, D. B., P. I. Prado, K. S. Brown, and A.V.L. Freitas.2010. Temporaldiversitypatterns andphenology in fruit-feeding butterßies in the Atlantic Forest. Biotropica 42:710!716.
Ribeiro, D. B., R. Batista, P. I. Prado, K. S. Brown, and A.V.L.Freitas. 2012. The importance of small scales to the fruit-feeding butterßy assemblages in a fragmented landscape.Biodivers. Conserv. 21: 811!827.
Roveratti, J. 2008. Flora vascular do cerrado sensu stricto doParque Nacional de Brasõlia, Distrito Federal, Brasil echave de identi"cacao das especies, p. 93. Master Degreedissertation, Universidade de Brasõlia. Brasõlia.
Ritter, C. D., R. Lemes, A.B.B. Morais, and C. S. Dambros.2011. Butterßies (Lepidoptera: Hesperioidea and Papil-ionoidea) frommixedombrophilous forest fragments,RioGrande do Sul, Brazil. Biota Neotrop. 11: 361!368.
Silva, N.A.P., M. R. Frizzas, and C.M.D. Oliveira. 2011a.Seasonality in insect abundance in the “Cerrado” of GoiasState, Brazil. Rev. Bras. Entomol. 55: 79!87.
Silva, I. A., D. M. Silva, G. H. Carvalho, and M. A. Batalha.2011b. Reproductive phenology of Brazilian savannasand riparian forests: environmental and phylogenetic is-sues. Ann. For. Sci. 68: 1207!1215.
Silva-Junior,M.C., andT.R. Sarmento. 2009. Comunidadeslenhosas no cerrado sentido restrito em duas posicoestopogra"cas na estacao ecologica do Jardim Botanico deBrasõlia, DF, Brasil. Rodriguesia 60: 277!294.
Sourakov, A., A. Duehl, and A. Sourakov. 2012. Foragingbehavior of the blue Morpho and other tropical butter-ßies: the chemical and electrophysiological basis of ol-factory preferences and the role of color. Psyche 2012:1!10.
Spix, J. B., and C.F.P.Martius. 1824. Travels in Brazil, in theyears 1817!1820, p. 290. Longman, Hurst, Rees, Orme,Brown, and Green, London, United Kingdom.
Thomas, C. D. 1991. Habitat use and geographic ranges ofbutterßies from the wet lowlands of Costa Rica. Biol.Conserv. 55: 269!281.
Uehara-Prado, M., K. S. Brown, A. Victor, and A. L. Freitas.2007. Species richness, composition and abundance offruit-feeding butterßies in the Brazilian Atlantic Forest:comparison between a fragmented and a continuouslandscape. Global Ecol. Biogeogr. 16: 43!54.
Urich, F. C., and T. Emmel. 1991. Life histories of Neotro-pical butterßies from Trinidad 3. Morpho peleides insu-laris (Nymphalidae: Morphinae). Trop. Lepid, (Gaines-ville) 2: 137!139.
Warming, E. 1892. Lagoa Santa, constibuicao para a geogra-"a "tobiologica, pp. 1!182. InE. Warming and M. G. Ferri(eds.), Lagoa Santa e a Vegetacao dos Cerrados Brasil-eiros. Belo Horizonte, Itatiaia.
Weatherspoon, M. R., Y. Cai, M. Crne, M. Srinivasarao, andK.H. Sandhage. 2008. 3D Rutile titania-based structureswith Morpho butterßy wing scale morphologies. Angew.Chem. Int. Ed. 47: 7921!7923.
Wolda, H. 1988. Insect seasonality: why? Annu. Rev. Ecol.Syst. 19: 1!18.
Young, A. M. 1975. Feeding behavior of Morpho butterßies(Lepidoptera: Nymphalidae: Morphinae) in a seasonaltropical environment. Rev. Biol. Trop. 23: 101!123.
Young, A. M., and A. Muyshondt. 1972. Geographical andecological expansion in tropical butterßies of the genusMorpho in evolutionary time. Rev. Biol. Trop. 20: 231!264.
Zar, J. H. 2010. Biostatistical analysis, 5th ed, Upper SaddleRiver, New Jersey.
Received 19 December 2012; accepted 7 January 2014.
282 ENVIRONMENTAL ENTOMOLOGY Vol. 43, no. 2
Copyright © 2022 FDOKUMEN




















