
Fossil Nelumbonaceae from the La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous),...
10
Fossil Nelumbonaceae from the La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous), Chubut, Patagonia, Argentina Maria A. Gandolfo a, * , Ruben N. Cuneo b,1 a L.H. Bailey Hortorium, Department of Plant Biology, Cornell University, Ithaca, NY 14853-5908, USA b MEF-Museo Paleontolo ´gico Egidio Feruglio, Trelew, Chubut, Argentina Received 29 April 2004; received in revised form 5 August 2004; accepted 23 September 2004 Abstract We describe here fossil leaves and fruits assignable to the family Nelumbonaceae Dumortier 1828. Fossils were collected at the Can ˜ado ´n del Irupe ´ locality, La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous), Chubut Province, Patagonia, Argentina. The fossil leaves are simple, with symmetrical and peltate lamina, orbicular in shape, and an entire margin. The venation is truly actinodromous with at least 15 primary veins, the secondary veins are poorly developed and intercalated with the primary veins, the third category vein is opposite, and the areoles are well-developed and 4- to 5-sided. These features are characteristic of the extant genus Nelumbo Adamson 1763, and the presence of these characters in the fossils allows their placement within this genus. Reproductive structures, similar to the fruit-receptacle of Nelumbo with fruits in situ, were found associated but not in organic connection with the leaves. These fossils constitute the only and oldest record for the family in the Southern Hemisphere. This record indicates that Nelumbo was more widespread in the past than today. D 2004 Elsevier B.V. All rights reserved. Keywords: Nelumbonaceae; fossils; Upper Cretaceous; La Colonia Formation; Patagonia; Argentina 1. Introduction What little is known about Maastrichtian floras of Southern South America is based primarily on five floras, four from Chile (Rı ´o de las Chinas, Quiriquina, Pichasca, and Cerro Guido) and one from Argentina (Estancia La Sara), which have been characterized mostly by palynological data (Troncoso and Romero, 1998). Although special attention has recently been given to the Upper Cretaceous La Colonia Formation due to its diverse and rich vertebrate fauna found in its continental facies (including fishes, turtles, snakes, plesiosaurs, birds, mammals, and dinosaurs; Albino, 2000; Bonaparte, 1985; Gasparini and De La Fuente, 2000; Pascual et al., 2000; Rougier et al., 2001), its 0034-6667/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.revpalbo.2004.09.007 * Corresponding author. Fax: +1 607 255 5407. E-mail addresses: [email protected] (M.A. Gandolfo)8 [email protected] (R.N. Cuneo). 1 Fax: +54 2965 432100. Review of Palaeobotany and Palynology 133 (2005) 169 – 178 www.elsevier.com/locate/revpalbo
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Fossil Nelumbonaceae from the La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous),...

www.elsevier.com/locate/revpalbo
Review of Palaeobotany and Paly
Fossil Nelumbonaceae from the La Colonia Formation
(Campanian–Maastrichtian, Upper Cretaceous),
Chubut, Patagonia, Argentina
Maria A. Gandolfoa,*, Ruben N. Cuneob,1
aL.H. Bailey Hortorium, Department of Plant Biology, Cornell University, Ithaca, NY 14853-5908, USAbMEF-Museo Paleontologico Egidio Feruglio, Trelew, Chubut, Argentina
Received 29 April 2004; received in revised form 5 August 2004; accepted 23 September 2004
Abstract
We describe here fossil leaves and fruits assignable to the family Nelumbonaceae Dumortier 1828. Fossils were collected at
the Canadon del Irupe locality, La Colonia Formation (Campanian–Maastrichtian, Upper Cretaceous), Chubut Province,
Patagonia, Argentina. The fossil leaves are simple, with symmetrical and peltate lamina, orbicular in shape, and an entire
margin. The venation is truly actinodromous with at least 15 primary veins, the secondary veins are poorly developed and
intercalated with the primary veins, the third category vein is opposite, and the areoles are well-developed and 4- to 5-sided.
These features are characteristic of the extant genus Nelumbo Adamson 1763, and the presence of these characters in the fossils
allows their placement within this genus. Reproductive structures, similar to the fruit-receptacle of Nelumbo with fruits in situ,
were found associated but not in organic connection with the leaves. These fossils constitute the only and oldest record for the
family in the Southern Hemisphere. This record indicates that Nelumbo was more widespread in the past than today.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Nelumbonaceae; fossils; Upper Cretaceous; La Colonia Formation; Patagonia; Argentina
1. Introduction
What little is known about Maastrichtian floras of
Southern South America is based primarily on five
floras, four from Chile (Rıo de las Chinas, Quiriquina,
0034-6667/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.revpalbo.2004.09.007
* Corresponding author. Fax: +1 607 255 5407.
E-mail addresses: [email protected] (M.A. Gandolfo)8
[email protected] (R.N. Cuneo).1 Fax: +54 2965 432100.
Pichasca, and Cerro Guido) and one from Argentina
(Estancia La Sara), which have been characterized
mostly by palynological data (Troncoso and Romero,
1998). Although special attention has recently been
given to the Upper Cretaceous La Colonia Formation
due to its diverse and rich vertebrate fauna found in its
continental facies (including fishes, turtles, snakes,
plesiosaurs, birds, mammals, and dinosaurs; Albino,
2000; Bonaparte, 1985; Gasparini and De La Fuente,
2000; Pascual et al., 2000; Rougier et al., 2001), its
nology 133 (2005) 169–178

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178170
paleoflora is scarcely known. Nevertheless, new
paleobotanical collections have revealed the presence
of an unexpectedly diverse plant assemblage.
Previous to this report, only two records of fossil
plants from the La Colonia Formation were known;
both records are of aquatic ferns. Archangelsky et al.
(1999) described megaspores and microspore massulae
of the fossil species Paleoazolla patagonica, while
Cuneo and Archangelsky (2000) reported megafossils
and microfossils with affinities to the extant genus
Regnellidium within the family Marsileaceae. Associ-
ated with these aquatic ferns, there are other pterido-
phytes, gymnosperms, and several angiosperms. This
contribution is focused on the taxonomy and descrip-
tion of the aquatic angiosperm family Nelumbonaceae
from the La Colonia Formation paleoflora.
2. Geology and age
The La Colonia Formation outcrops in an area
known as the Somuncura Massif; it is exposed between
Telsen and Sierra Rosada, Chubut province, Patagonia,
Argentina (Fig. 1). This formation was recognized by
Pesce (1979) as a homogeneous series of upward fining
Fig. 1. Locality map of outcrops of the La Colonia Formation
sediments, and it is divided into three portions (lower,
middle, and upper). Argillaceous sediments are char-
acteristic of the lower and middle portions of the
sequence. They are considered continental and thought
to probably represent lacustrine or lagoonal deposits.
The upper portion of the sequence changes to marginal
and shallow marine environment (Ardolino and Del-
pino, 1987; Ardolino and Franchi, 1996).
Ardolino and Franchi (1996) assigned a Senonian
(pre-Campanian) age to the lower and middle
sections. Later, Page et al. (1999) considered that an
unconformity in the base of the unit is inter-Senonian,
and, consequently, they assumed that the lower and
middle sections are at least Campanian in age. The
age of the upper section, based on ostracod and
foraminifera data collected in several localities is
Campanian–Maastrichtian, Upper Cretaceous (Ardo-
lino and Franchi, 1996).
3. Materials and methods
The fossil specimens studied here are fragments
and complete leaves and fruit impressions. They were
recovered from clays deposited in a littoral lagoon
(Maastrichtian, Upper Cretaceous) at Chubut province.

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178 171
environment. Although they seem to be quite fragile,
they are well preserved, suggesting low degree of
transportation. These fossil remains were found in
Fig. 2. Stratigraphic column of the Canadon d
association with other angiosperm leaves (at least four
morphotypes), another aquatic angiosperm (Nym-
phaeaceae seeds, Gandolfo et al., 2002), aquatic ferns
el Irupe locality, La Colonia Formation.

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178172
(Paleoazolla and Regnellidium), other pteridophytes,
and gymnosperms.
Fossil remains were collected at the Canadon del
Irupe locality (Fig. 2), La Colonia Formation, Chubut
province, Argentina and are housed at the Paleobotan-
ical collection of the Museo Paleontologico Egidio
Feruglio under the initials MPEF-Pb-Mz. Leaves,
receptacles, and fruits of extant species of Nelumbo
used for comparisons were observed from the herba-
rium collection of the L.H. Bailey Hortorium (BH).
Fossil leaves were described following the manual
of leaf architecture by the Leaf Architecture Working
Group (1999), and leaves and fruits were studied using
a stereomicroscope Zeiss Stemi SV-11. Photographs
were obtained with a Nikon 995 digital camera.
4. Systematic descriptions
4.1. Leaves
Genus Nelumbo Adamson 1763.
Nelumbo puertae Gandolfo and Cuneo, sp. nov.
(Plate IA–F).
Diagnosis: Simple, centrally peltate microphyll,
lamina orbiculate (L/W: 1), margin entire; first vein
category actinodromous, at least 12 primary veins
diverging regularly from the center of the lamina,
bifurcating at least once, and forming festooned
brochidodromous archs; poorly developed secondary
veins intercalated with the primary veins; third and
fourth category veins opposite percurrent; areoles
well-developed, 4- to 5-sided; marginal ultimate
venation looped, freely ending ultimate veins
unbranched to 1-branched, highest vein order fourth.
Holotype: MPEF-Pb-Mz 864-A and B; Plate IA.
Paratypes: MPEF-Pb-Mz 865–901; Plate IB, C and F.
Type locality: Canadon del Irupe; Mirasol Chico
Creek, Los Altares (4369-IV), Chubut province,
Patagonia, Argentina.
Horizon: uppermost part of the La Colonia Formation
at Canadon del Irupe locality.
Studied material: MPEF-Pb-Mz 864–901.
Derivatio nominis: puertae, in honor of Pablo Puerta,
Argentinean paleontological explorer.
Remarks: The leaves are simple microphylls charac-
terized by their symmetrical peltate lamina. The lamina
is rounded to orbicular with an entire margin, is at least
4 to 11 cm in diameter, and probably of membrana-
ceous texture (Plate IA–C). The petiole is inserted
centrally at the lamina, and, from its point of insertion,
the primary veins emerge radially (Plate IA,B,D,F ).
The first category venation is truly actinodromous, with
12–18 primary veins, emerging from the center of the
leaf (insertion point of the petiole) at acute angles (25–
308) in relation to other primary veins (Plate IA,B,D).
The course is straight, and the veins bifurcate once
maintaining the course; they then abruptly curve to join
another primary vein or a bifurcation of a primary vein,
forming brochidodromous arches (Plate IG). There are
veins of smaller caliber intercalated with the primary
veins that are interpreted as poorly developed secon-
dary veins. These veins have straight to slightly sinuous
course, and they are fused to third order veins forming a
bherring boneQ pattern. The third category vein is
opposite percurrent, with the veins emerging uniformly
from the primary veins at right to obtuse angles (90–
1008), their course is straight. The fourth category veinis the highest order and has an opposite percurrent
model (Plate ID,E). The areoles are well developed,
and they are 4- to 5- sided. Inside the areoles, the freely
ending ultimate veins that are unbranched to 1-
branched are noticeable. In addition to the festooned
brochidodromous arches formed by the primary veins,
there are other arches formed by veins of less caliber;
they originate a looped ultimate marginal venation
(Plate IC–E).
4.2. Reproductive structures
Associated Nelumbo-like structures.
(Plate IIA–C).
Structures inferred as reproductive organs were
found associated with the leaves. The remains consist
of numerous impressions similar to those described by
Samylina (1968) as ban inversely conical, accrescent
receptacle with fruits inside, like those of NelumboQand by Upchurch et al. (1994) as probably ba floral
axis.QThe Patagonian fossils are probably conical fruit-
like receptacles, at least 2.5 cm long and 1.1–1.5 cm in
diameter (Plate IIA–C). From the distal flat portion of
the receptacle, there are between 5–14 protuberances
that we have interpreted as fruits. They are surrounded
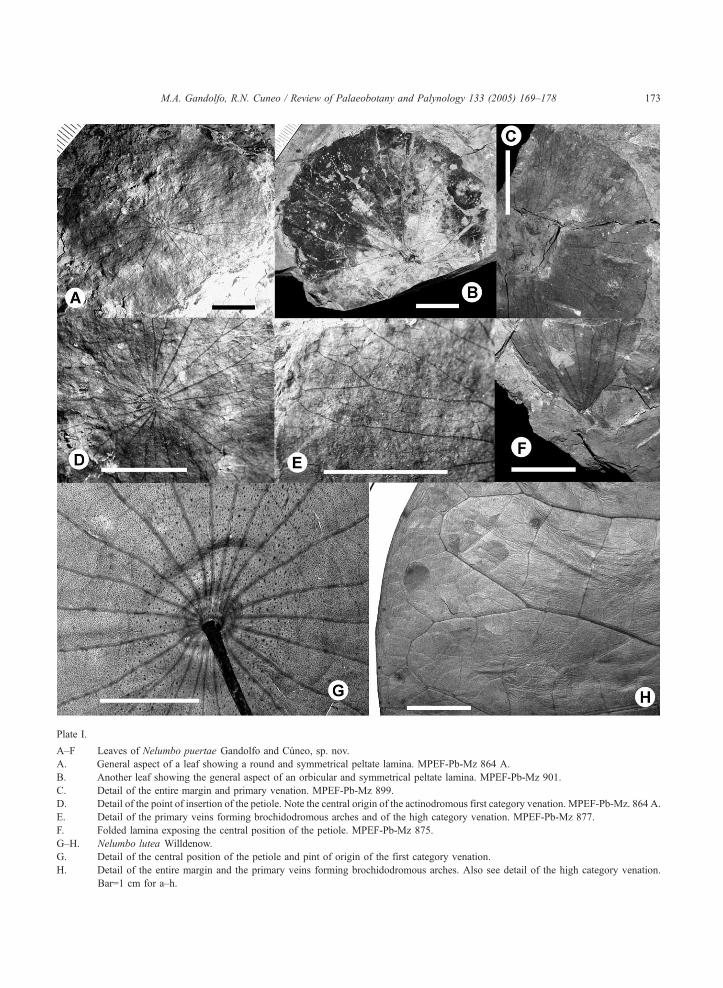
Plate I.
A–F Leaves of Nelumbo puertae Gandolfo and Cuneo, sp. nov.
A. General aspect of a leaf showing a round and symmetrical peltate lamina. MPEF-Pb-Mz 864 A.
B. Another leaf showing the general aspect of an orbicular and symmetrical peltate lamina. MPEF-Pb-Mz 901.
C. Detail of the entire margin and primary venation. MPEF-Pb-Mz 899.
D. Detail of the point of insertion of the petiole. Note the central origin of the actinodromous first category venation. MPEF-Pb-Mz. 864 A.
E. Detail of the primary veins forming brochidodromous arches and of the high category venation. MPEF-Pb-Mz 877.
F. Folded lamina exposing the central position of the petiole. MPEF-Pb-Mz 875.
G–H. Nelumbo lutea Willdenow.
G. Detail of the central position of the petiole and pint of origin of the first category venation.
H. Detail of the entire margin and the primary veins forming brochidodromous arches. Also see detail of the high category venation.
Bar=1 cm for a–h.
M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178 173

Plate II.
A–C Nelumbo-like fossil receptacle and fruits.
A. Receptacle laterally compressed. MPEF-Pb-Mz 914.
B. Top view of a receptacle with fruits in situ, note the wall of the receptacle. MPEF-Pb-Mz 903.
C. Detail of the fruits, note the apertures (respiratory pores). MPEF-Pb-Mz 920.
D–E. Nelumbo lutea Willdenow.
D. Top view of a receptacle with fruits in situ.
E. Detail of the fruits showing the respiratory pores. Bar=1 cm for figs. A–B and D–E, bar=0.5 cm for C.
M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178174

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178 175
by the thick wall of the receptacle. The wall of the
receptacle produces a rim surrounding the fruits. If our
interpretation is correct, each fruit is entrenched into a
single cavity and therefore separated from each other.
The fruits are rounded to oblong in shape and have a
small aperture at the top (Plate IIC). We assume that
these apertures are representing small respiratory pores.
Type locality: Canadon del Irupe; Mirasol Chico
Creek, Los Altares (4369-IV), Chubut province,
Argentina.
Horizon: uppermost part of the La Colonia Formation
at Canadon del Irupe locality.
Studied material: MPEF-Pb-Mz 902–920.
5. Discussion
From the morphological data gathered from the
Patagonian fossil leaves, it is clear that they belong to
the monotypic family Nelumbonaceae, allowing the
assignment of these leaves to the living genus
Nelumbo Adanson 1763. Nelumbo puertae leaves
are almost indistinguishable from extant ones.
Nelumbo, within the monotypic family Nelumbona-
ceae Dumortier, comprises two extant species,
Nelumbo nucifera Gaertner and N. lutea Willdenow.
Both species are aquatic perennial, have peltate
orbicular leaves, and produce nut-like indehiscent
fruits that are free within an accrescent receptacle
(Cronquist, 1981; Williamson and Schneider, 1993;
Takhtajan, 1996; Wiersema, 1997). Hickey and Wolfe
(1975) characterized the leaves of the family Nelum-
bonaceae as simple, truly peltate with entire margin,
and truly actinodromous with numerous primaries.
Upchurch et al. (1994) presented a list of leaf features
for modern Nelumbo. These features are the orbicular
lamina, the entire margin, the central petiole, the
medial primary vein giving rise to secondary veins
immediately beneath the leaf apex, the intercostal
primary loops uniform in size and shape, the tertiary
veins regular and transversely oriented, and, finally,
hexagonal areoles (Plate IG–H). The new fossil
species, N. puertae, shares five of the seven characters
enunciated by Upchurch et al. (1994). Nelumbo
puertae differs from extant species of Nelumbo in
the presence of poorly developed secondary category
venation, and the shape of the areoles. In addition, the
lamina of the fossil species is smaller than the extant
species in which sizes vary between 10–100 cm in
diameter (Williamson and Schneider, 1993; Wier-
sema, 1997). Furthermore, the fossil species has fewer
primary veins than the extant ones (19–23 veins,
Knowlton, 1930; personal observation, Plate IH).
Although the fossil record for the Nelumbonaceae is
extensive, there is no fossil record from the Southern
Hemisphere; all fossil species described as belonging
to the family come from the Northern Hemisphere.
Several fossil species of Nelumbo were described from
the Northern Hemisphere for sediments of Cretaceous
to Miocene in age. Kuprianova and Tarasevich (1983)
provided a summary of the fossil record for Nelumbo
pollen grains. The oldest leaves described as belonging
toNelumbo come from the Albian of Portugal (Saporta,
1894), later redescribed by Teixeira (1945). All the
Portuguese fossil species range from 20–30 cm in
diameter, have more than 20 primary veins, and the
tertiary vein category seems to be random reticulate
(see Teixeira, 1945, plate 1–3). However, their correct
taxonomic position is doubtful. These fossils are only
portion of leaves. They were exhumed from the same
sediments and locality where other leaves are attributed
as members of the family Nymphaeaceae, hence it is
possible that they are indeed members of the latter and
not Nelumbonaceae. A new reinvestigation of these
fossils is necessary to really assess their taxonomic
placement, which is beyond the scope of this paper.
Moreover, Collinson et al. (1993) and Drinnan et al.
(1994) considered as valid for Europe only the reports
of Nelumbo of Tertiary age; also Drinnan et al. (1994)
considered that possible leaves and reproductive
structures of Nelumbonaceae are present at the late
Albian of eastern North America.
In North America, there are at least 30 fossil
species of Upper Cretaceous and Tertiary age that
have been assigned to Nelumbo, among them N.
crossii Knowlton 1930, N. dawsoni (Hollick) Berry
1935, N. tenuifolia (Knowlton) Knowlton 1922 and
N. weymouthi R. W. Brown 1933. All these fossil
species were erected during the first half of the last
century and they are based mostly on single speci-
mens (Knowlton, 1930). They were described as
having entire margins and actinodromous venation,
and primary veins varying from 11 to 23; however,
many of the descriptions and illustrations lack enough
information to allow detailed comparisons with the
new species N. puertae.

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178176
Matsuo (1954) described the species Nelumbo
orientalis on the basis of leaf impressions from the
Upper Cretaceous of Japan, and he considered this
record as the oldest for the family known from East
Asia. Nonetheless, later on Matsuo (1962) transferred
this species to the species Nelumbium orientalis, a
fossil species also based on leaves remains, without
any explanation.
The possibility that the fossils described here could
belong to the fossil genus Nelumbites Berry 1911a
was also considered. The fossil genus Nelumbites was
erected by Berry (1911) to place fossil leaves with
characters similar to those of living Nelumbo.
Upchurch et al. (1994) emended its diagnosis. Based
on the revised diagnosis, we disregard the placement
of the Patagonian fossils within the genus Nelumbites
because they differ in lamina shape, position of the
petiole, tertiary category pattern, the ultimate venat-
ion, and lack of a midvein.
The fossil genus Protonelumbo Knowlton was
rejected as well as suitable for the fossils described
here. Knowlton (1930), who erected this fossil genus,
expressed that Protonelumbo leaves are similar to
those of Nelumbo but they differed in their margins.
Protonelumbo shows obtuse teeth or lobes, one for
each rib, with each lobe separated by deep rounded
sinuses. N. puertae has entire margin.
As mentioned earlier the family comprises two
modern species, N. lutea, a native of North and
Central America, and N. nucifera, which is wide-
spread in warmer parts of Asia and Australia. Both
species grow in ponds, lakes, slow streams and in
waters up to 2 m deep (Williamson and Schneider,
1993). The orbicular shape and the centered petiole of
N. puertae suggest that it had aquatic leaves as well.
Consequently, it is possible that N. puertae lived in a
very similar environment. This interpretation is also
supported by cooccurrence of the two aquatic ferns
(Regnellidium and Paleoazolla) and the sedimento-
logical data that defined the sediments as lacustrine/
lagoonal (Ardolino and Franchi, 1996).
With respect of the fruit-like receptacles, the
characters preserved are consistent to those of living
Nelumbo and with those described by Upchurch et al.
(1994) of the Quantico locality of the Lower Creta-
ceous Potomac Group of Virginia. Nelumbo produces
conical receptacles where the fruits (or disseminules
for Collinson, 1980) are free from each other, and
each of the fruits has a small respiratory pore
(Cronquist, 1981; Takhtajan, 1996; Plate IID,E).
Unfortunately, they are not in organic connection
with the leaves, and it would be incorrect to include
the receptacles within N. puertae. Until their relation-
ships with N. puertae can be determined, we prefer to
place them within the genus Nelumbo without a
species assignation.
Nelumbonaceae is considered as a bbasalQ eudicotfamily. Although it is not our intention to discuss at
length its taxonomic position, it is fair to mention
that the placement of Nelumbonaceae within the
order Nymphaeales was originally disputed by
Takhtajan (1970) who studied the morphology of
its pollen grains. Unexpectedly, based on molecular
data, the family Nelumbonaceae appears to be a
member of the Proteales clade formed by Protea-
ceae, Platanaceae and Nelumbonaceae, three fami-
lies considered in the past to be only distantly
related (Soltis et al., 2000, Qiu et al., 2000, Nandi
et al., 1998). Within the Proteales clade, Nelumbo-
naceae is the sister group of the pair Proteaceae+-
Platanaceae.
The fossils described here are of particular
significance. The discovery of leaves and fruits of
Nelumbo in Patagonia provide a new dimension to
our knowledge of the fossil record for the family
Nelumbonaceae and its present and past distribution.
As mentioned previously, today the family is
restricted to tropical and subtropical areas of North
and Central America and Asia and Australia.
However, this report supports the idea that, in the
past the family was more widespread. The fossils
described in this contribution constitute the only
fossil record for the family Nelumbonaceae in the
Southern Hemisphere.
Acknowledgements
This work was supported by the Franklin Research
Award of the American Philosophical Society awarded
to M. A. Gandolfo. In particular, we are in debt with
Ana Archangelsky and Gerardo Cladera that provided
information and the stratigraphic column of the local-
ity, and Pablo Puerta, Cynthia Gonzalez, Jose Carbal-
lido, Mariano Caffa, Maximo Dellocca of the Museo
Paleontologico E. Feruglio for assistance during

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178 177
fieldwork. Harufumi Nishida helped with the Japanese
literature consulted. We thank Kathleen B. Pigg and an
anonymous reviewer for their useful comments on the
original manuscript. We also want to express our
gratitude to the Curator of the L. H. Bailey Hortorium
Herbarium (BH), Cornell University for allowing the
use of the collection. Donna and Tim Schender
provided leaves and fruits of extant Nelumbo.
References
Albino, A.M., 2000. New record of snakes from the Cretaceous of
Patagonia (Argentina). Geodiversitas 22, 247–253.
Archangelsky, A., Phipps, C.J., Taylor, T.N., Taylor, E.L., 1999.
Paleoazolla, a new heterosporous fern from the Upper Creta-
ceous of Argentina. Am. J. Bot. 86, 1200–1206.
Ardolino, A., Delpino, D., 1987. Senoniano (continental-marino).
Comarca Norpatagonica, Provincia del Chubut, Argentina,
Actas vol. 3. Decimo Congreso Geologico Argentino, Argen-
tina, Tucuman, pp. 193–196.
Ardolino, A., Franchi, M., 1996. Hoja geologica 4366-I Telsen,
provincia del Chubut. Subsecretarıa de Minerıa de la Nacion,
Direccion General del Servicio Geologico, Boletın, vol. 215.
Buenos Aires, Argentina.
Berry, E.W., 1911. Systematic paleontology, Lower Cretaceous:
fossil plants. In: Clark, W.B. (Ed.), Lower Cretaceous. Maryland
Geol. Surv., Baltimore, USA, pp. 214–508.
Berry, E.W., 1935. A preliminary contribution to the floras of the
Whitemud and Ravenscrag formations. Canada Dept. Mines
Geol. Survey Mem. 182, 1–107.
Bonaparte, J.F., 1985. A horned Cretaceous carnosaur from
Patagonia. Natl. Geogr. Res. 1, 149–151.
Brown, R.W., 1933. Fossil plants from the Aspen Shale of
southwestern Wyoming. Proc. U. S. Natl. Mus. 82, 1–10.
Collinson, M.E., 1980. Recent and Tertiary seeds of the Nym-
phaeaceae sensu lato with the revision of Brasenia ovula
(Brong.) Reid and Chandler. Ann. Bot. 46, 603–632.
Collinson, M.E., Boulter, M.C., Holmes, P.L., 1993. Magnolio-
phyta (bAngiospermaeQ). In: Benton, M.J. (Ed.), The Fossil
Record, vol. 2. Chapman and Hall, London, United Kingdom,
pp. 809–841.
Cronquist, A., 1981. An Integrated System of Classification of
Flowering Plants. Columbia University Press, New York.
Cuneo, R., Archangelsky, A., 2000. Aquatic ferns from the
Cretaceous of Patagonia. Sixth Conf. Int. Org. Palaeobot.
(IOPC-VI). Qinhuangdao of Hebei, China, pp. 22 – 23.
(Abstract).
Drinnan, A.N., Crane, P.R., Hoot, S.B., 1994. Patterns of floral
evolution in early diversification of non-magnoliid dicotyledons
(eudicots). Plant Syst. Evol. 8, 93–122. (Suppl.).
Gandolfo, M.A., Cuneo, R.N., Archangelsky, A., 2002. Aquatic
angiosperms from the La Colonia Formation (Upper Creta-
ceous), Chubut, Patagonia, Argentina. Third Europ. Meet.
Palaeont. Stratig. Latin Am., pp. 54. Abstract.
Gasparini, Z., De La Fuente, M., 2000. Tortugas y plesiosaurios de
la Formacion La Colonia (Cretacico Superior) de Patagonia,
Argentina. Rev. Esp. Paleontol. 15, 23–35.
Hickey, L.J., Wolfe, J.A., 1975. The bases of angiosperm
phylogeny: vegetative morphology. Ann. Mo. Bot. Gard. 62,
538–589.
Knowlton, F.H., 1922. The Laramie flora of the Denver Basin with a
review of the Laramie problem. U.S. Geol. Surv. Prof. Pap. 130,
1–171.
Knowlton, F.H., 1930. The flora of the Denver and associated
formations of Colorado. U.S. Geol. Surv. Prof. Pap. 155, 1–142.
Kuprianova, L.A., Tarasevich, V.F., 1983. Pollen morphology of
recent and fossil species of the genus Nelumbo (Nelumbona-
ceae). Bot. Z. 68, 137–146.
Leaf Architecture Working Group, 1999. Manual of Leaf Archi-
tecture: Morphological Description and Categorization of
Dicotyledonous and Net-Veined Monocotyledonous Angio-
sperms. Smithsonian Institution, Washington, DC.
Matsuo, H., 1954. Discovery of Nelumbo from the Asuwa flora
(Upper Cretaceous) in Fukui prefecture in the inner side of central
Japan. Trans. Proc. Paleont. Soc. Japan, New Ser. 14, 155–158.
Matsuo, H., 1962. A study on the Asuwa Flora (Late Cretaceous
age) in the Hokuriku District, Central Japan. Sci. Rep.
Kanazawa Univ. 8, 228–231.
Nandi, O.I., Chase, M.W., Endress, P.K., 1998. A combined
cladistic analysis of angiosperms using rbcL and non-molecular
data sets. Ann. Mo. Bot. Gard. 85, 137–212.
Page, R., Ardolino, A., de Barrio, R., Franchi, M., Lizuain, A.,
Page, S., Silva Nieto, D., 1999. Estratigrafia del Jurasico y
Cretacico del Macizo de Somuncura, provincia de Rıo Negro y
Chubut. In: Caminos, R. (Ed.), Geologia Argentina. Segemar,
Anales, vol. 29, pp. 460–488.
Pascual, R., Goin, F.J., Gonzalez, P., Ardolino, A., Puerta, P.F.,
2000. A highly derived docodont from the Patagonian Late
Cretaceous: implications for Gondwanan mammals. Geodiver-
sitas 22, 395–414.
Pesce, A., 1979. Estratigrafıa del Arroyo Perdido en su tramo medio
e inferior, Provincia de Chubut. Septimo Cong. Geo. Arg.,
Neuquen, Actas, vol. 1, pp. 315–333.
Qiu, Y.-L., Lee, J., Bernasconia-Quadroni, F., Soltis, D.E, Soltis, P.S.,
Zanis, M., Zimmer, E.A., Chen, Z., Savolainen, V., Chase, M.W.,
2000. Phylogeny of basal angiosperms: analyses of five genes
from three genomes. Int. J. Plant Sci. 161 (Suppl. 6), S3–S27.
Rougier, G.M., Novacek, J., Ortiz-Jaureguizar, E., Pol, D., Puerta,
P.F., Muzzopappa, P., 2001. Mamıferos de la Formacion La
Colonia (Cretacico Superior): nuevas evidencias en favor del
aislamiento biogeografico de Patagonia durante el Cretacico
Tardıo-Terciario Temprano. Decimoseptimas Jorn. Arg. Paleont.
Vert., pp. 18R–19R. (Abstract).
Samylina, V.A., 1968. Early Cretaceous angiosperms of the Soviet
Union based on leaf and fruit remains. J. Linn. Soc. Bot. 61,
207–218.
de Marquis Saporta, G., 1894. Nouveaux details concernant les
Nympheinees. Nympheinees infracretacees. Comp. Rend. Sean.
L’Acad. Sci. 119, 837–853.
Soltis, D.E., Soltis, P.S., Chase, M.W., Mort, M.E., Albach, D.C.,
Zanis, M., Savolainen, V., Hahn, W.H., Hoot, S.B., Fay, M.F.,

M.A. Gandolfo, R.N. Cuneo / Review of Palaeobotany and Palynology 133 (2005) 169–178178
Axtell, M., Swensen, S.M., Prince, L.M, Kress, W.J., Nixon,
K.C., Farris, J.S., 2000. Angiosperm phylogeny inferred from
18S rDNA, rbcL, and atpB sequences. Bot. J. Linn. Soc. 133,
381–461.
Takhtajan, A., 1970. Origin and Dispersal of Flowering Plants.
Nauka, Leningrad.
Takhtajan, A., 1996. Diversity and Classification of Flowering
Plants. Columbia University Press, New York.
Teixeira, C., 1945. Nympheacees Fossiles du Portugal. Services
Geologiques du Portugal.
Troncoso, A., Romero, EJ., 1998. Evolucion de las comunidades
florısticas en el extremo sur de Sudamerica durante el cenofıtico.
Fortunato, R., Bacigalupo, N. Proc. Sixth Cong. Latinoam. Bot.,
Monog. Syst. Bot., vol. 68, pp. 149–172.
Upchurch, G.R, Crane, P.R., Drinnan, A.N., 1994. The megaflora
from the Quantico locality (Upper Albian), Lower Cretaceous
Potomac Group of Virginia. Virginia. Mus. Natl. Hist. Mem. 4,
1–57.
Wiersema, J.H., 1997. Nelumbonaceae Dumortier. Flora of North
America Editorial Committee, Flora of North America, North of
Mexico, Magnoliophyta: Magnoliidea and Hamamelidea, vol. 3.
Oxford University Press, Oxford, pp. 64–65.
Williamson, P.S., Schneider, E.L., 1993. Nelumbonaceae. In:
Kubitzki, K., Rohwer, J.G., Bittrich, V. (Eds.), The Families
and Genera of Vascular Plants. Springer-Verlag, New York,
pp. 470–473.




















