
Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt -taxonomy, biostratigraphy,...
32
Revue de Paléobiologie, Genève (juin 2008) 27 (1): 159-189 Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt - taxonomy, biostratigraphy, paleoecology and paleobiogeography Abdel-Mohsen M. MORSI 1 , Mahmoud FARIS 2 , Abd-Elfattah ZALAT 2 & Rehab F.M. SALEM 2 Abstract Detailed study of ostracodes from the Maastrichtian-Early Eocene sections at Gebel Nukhul and Wadi Feiran in west-central Sinai yielded 38 ostracode species and subspecies belonging to 26 genera. Biostratigraphically, they are divisible into five kinds : taxa restricted to the Maastrichtian, taxa crossing the K/P boundary, taxa restricted to the Paleocene, taxa crossing the P/E boundary, and taxa only found in the Early Eocene. The recorded faunas have a wide geographic distribution throughout North Africa and the Middle East. They are generally assigned to outer shelf - upper bathyal settings of normal marine environments and belong to the type of species termed as the “South Tethyan Type” that wandered along southern shores of the Tethys during this time. The presence of taxa which are also found in West Africa emphasizes faunal exchange between the Southern Tethys and the West African basins during the Maastrichtian - Early Eocene. Key words Maastrichtian, Paleocene, Eocene, Ostracoda, southern Tethys, Egypt. Geology Department, Faculty of Science, Ain Shams University, 566 Cairo, Egypt. Email : [email protected] 2 Geology Department, Faculty of Science,Tanta University, Tanta, Egypt I. INTRODUCTION Late Cretaceous-Early Eocene successions have been a target of a large number of studies worldwide as they rep- resent a critical interval in the earth history during which major changes in ocean circulation and global climates took place. These changes triggered major biotic pertur- bations and evolutionary innovations amongst marine microbenthos (e.g. TJALSMA &LOHMANN, 1983; THOM- AS, 1998), marine microplanktons (e.g. KELLY et al., 1996; AUBRY, 1998, CROUCH et al., 2001), and even ter- restrial mammals (e.g. GINGERICH, 2000). The Late Cre- taceous-Early Eocene rocks cover extensive areas of Egypt. In the area of west-central Sinai, they are well- exposed and are represented by a wide variety of sedi- ments belonging to different depths of marine environ- ments. In the present paper, we mainly aim to study the ostracode faunas of the Paleocene-Early Eocene interval in west-central Sinai for taxonomy, biostratigraphy, pa- leoecology and paleobiogeography. The Late Maastrich- tian interval has also been included to mark off the ostra- code faunal changes at the K/P transition. Two sections at Gebel Nukhul and Wadi Feiran, between latitudes 28º 40 and 29º 10 and longitudes 33º 10 and 33º 40, have been measured, sampled and examined for their ostracode con- tent (Fig. 1). Associating calcareous nannofossil assem- blages have been utilized in biostratigraphic zonation, to serve as a control and support for biostratigraphic evalu- ation of the present ostracode record. The information about the Maastrichtiaan-Early Eocene ostracodes of Egypt has increased remarkably in the re- cent years through several publications. Until the year 1990, only a limited number of studies dealing with Maastrichtian-Early Eocene ostracodes from Egypt was available (BASSIOUNI et al., 1977; BOUKHARY et al., 1982; EL SWEIFY, 1984; KHALIFA et al., 1984). Since 1990, when BASSIOUNI &LUGER published their volumi- nous paper on ostracodes from southern Egypt, a large number of studies has been published and the knowledge about the Maastrichtian-Early Eocene ostracodes of Egypt has substantially increased through many studies carried out by ISMAIL (1992, 1996), ELEWA &ISHIZAKI (1994), MORSI (1999, 2000), BASSIOUNI &MORSI (2000), SHAHIN (2000, 2005), SHAHIN &EL NADY (2001), AB- DEL SHAFY et al. (2002), ELEWA (2002), SPEIJER &MOR- SI (2002), MORSI &SPEIJER (2003), ISMAIL &IED (2004, 2005) and ELEWA &MORSI (2004). Besides, the publica- tions in the other biogeographically related North African and Middle East areas (ESKER, 1968; BASSIOUNI, 1969b, c, 1970; DONZE et al., 1982; DAMOTTE &FLEURY, 1987; HONIGSTEIN &ROSENFELD, 1995; EL-WAER, 1992; WHATLEY &ARIAS, 1993; ANDREU, 1996) also represent an indispensable source of information. Works in the ba- sins of West Africa such as APOSTOLESCU (1961), REY- MENT (1963, 1981), REYMENT &REYMENT (1959, 1980), OKOSUN (1987), CARBONNEL &JOHNSON (1989), CAR- BONNEL et al. (1990), CARBONNEL &OYEDE (1991), REYMENT &ARANKI (1991), DIGBEHI et al. (1994); CAR- BONNEL &MONCIARDINI (1995) and COLIN et al. (1998) ISSN 0253-6730
Transcript of Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt -taxonomy, biostratigraphy,...

Revue de Paléobiologie, Genève (juin 2008) 27 (1): 159-189
Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt - taxonomy, biostratigraphy, paleoecology and paleobiogeography
Abdel-Mohsen M. MORSI1, Mahmoud FARIS2, Abd-Elfattah ZALAT2 & Rehab F.M. SALEM2
AbstractDetailed study of ostracodes from the Maastrichtian-Early Eocene sections at Gebel Nukhul and Wadi Feiran in west-central Sinai yielded 38 ostracode species and subspecies belonging to 26 genera. Biostratigraphically, they are divisible into five kinds : taxa restricted to the Maastrichtian, taxa crossing the K/P boundary, taxa restricted to the Paleocene, taxa crossing the P/E boundary, and taxa only found in the Early Eocene. The recorded faunas have a wide geographic distribution throughout North Africa and the Middle East. They are generally assigned to outer shelf - upper bathyal settings of normal marine environments and belong to the type of species termed as the “South Tethyan Type” that wandered along southern shores of the Tethys during this time. The presence of taxa which are also found in West Africa emphasizes faunal exchange between the Southern Tethys and the West African basins during the Maastrichtian - Early Eocene.
Key words Maastrichtian, Paleocene, Eocene, Ostracoda, southern Tethys, Egypt.
� Geology Department, Faculty of Science, Ain Shams University, ��566 Cairo, Egypt. Email : [email protected] Geology Department, Faculty of Science,Tanta University, Tanta, Egypt
I. INTRODUCTION
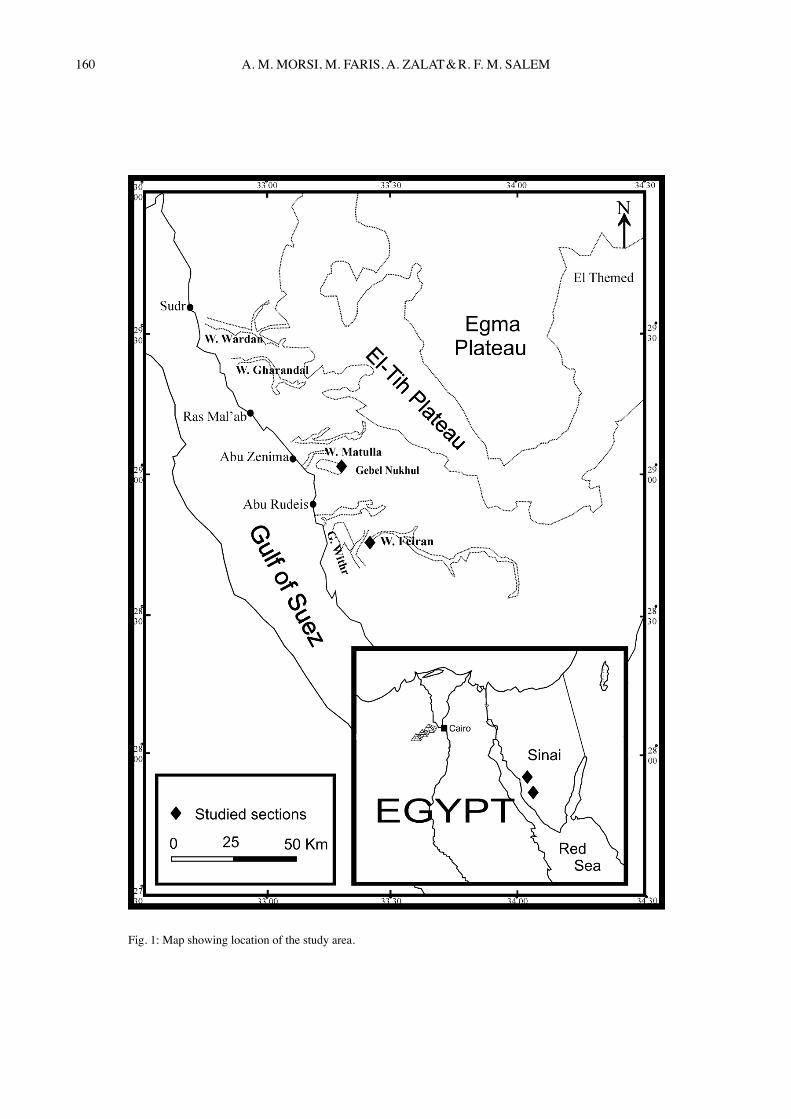
Late Cretaceous-Early Eocene successions have been a target of a large number of studies worldwide as they rep-resent a critical interval in the earth history during which major changes in ocean circulation and global climates took place. These changes triggered major biotic pertur-bations and evolutionary innovations amongst marine microbenthos (e.g. Tjalsma & lohmann, 1983; Thom-as, 1998), marine microplanktons (e.g. Kelly et al., 1996; aubry, 1998, CrouCh et al., 2001), and even ter-restrial mammals (e.g. GinGeriCh, 2000). The Late Cre-taceous-Early Eocene rocks cover extensive areas of Egypt. In the area of west-central Sinai, they are well-exposed and are represented by a wide variety of sedi-ments belonging to different depths of marine environ-ments. In the present paper, we mainly aim to study the ostracode faunas of the Paleocene-Early Eocene interval in west-central Sinai for taxonomy, biostratigraphy, pa-leoecology and paleobiogeography. The Late Maastrich-tian interval has also been included to mark off the ostra-code faunal changes at the K/P transition. Two sections at Gebel Nukhul and Wadi Feiran, between latitudes 28º 40́ and 29º 10́ and longitudes 33º 10́ and 33º 40́, have been measured, sampled and examined for their ostracode con-tent (Fig. 1). Associating calcareous nannofossil assem-blages have been utilized in biostratigraphic zonation, to serve as a control and support for biostratigraphic evalu-ation of the present ostracode record.
The information about the Maastrichtiaan-Early Eocene ostracodes of Egypt has increased remarkably in the re-cent years through several publications. Until the year 1990, only a limited number of studies dealing with Maastrichtian-Early Eocene ostracodes from Egypt was available (bassiouni et al., 1977; bouKhary et al., 1982; el sweify, 1984; Khalifa et al., 1984). Since 1990, when bassiouni & luGer published their volumi-nous paper on ostracodes from southern Egypt, a large number of studies has been published and the knowledge about the Maastrichtian-Early Eocene ostracodes of Egypt has substantially increased through many studies carried out by ismail (1992, 1996), elewa & ishizaKi (1994), morsi (1999, 2000), bassiouni & morsi (2000), shahin (2000, 2005), shahin & el nady (2001), ab-del shafy et al. (2002), elewa (2002), speijer & mor-si (2002), morsi & speijer (2003), ismail & ied (2004, 2005) and elewa & morsi (2004). Besides, the publica-tions in the other biogeographically related North African and Middle East areas (esKer, 1968; bassiouni, 1969b, c, 1970; donze et al., 1982; damoTTe & fleury, 1987; honiGsTein & rosenfeld, 1995; el-waer, 1992; whaTley & arias, 1993; andreu, 1996) also represent an indispensable source of information. Works in the ba-sins of West Africa such as aposTolesCu (1961), rey-menT (1963, 1981), reymenT & reymenT (1959, 1980), oKosun (1987), Carbonnel & johnson (1989), Car-bonnel et al. (1990), Carbonnel & oyede (1991), reymenT & aranKi (1991), diGbehi et al. (1994); Car-bonnel & monCiardini (1995) and Colin et al. (1998)
ISSN 0253-6730
160 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Fig. 1: Map showing location of the study area.

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 161
have been also considered since many ostracode elements in these areas are in common with the Southern Tethys areas.
II. LITHOSTRATIGRAPHY
During the Maastrichtian-Early Eocene, the study area was a part of an extensive epicontinental basin covering nearly all Egypt. With respect to their lithofacies and re-gional distributions, successions of this interval are di-vided into five lithostratigraphic units with a wide geo-graphic extension. These rock units are named, from older to younger, the Sudr, Dakhla, Tarawan, Esna and Thebes Formations (Figs 2, 3).
Sudr Formation (Ghorab, 1961)In the type locality at Wadi Sudr (west-central Sinai), this formation is composed of snow-white chalk, chalky lime-stone and argillaceous limestone (100-130 m thick). There, it is divisible from base to top into the Markha and Abu Zeneima Members (Ghorab, 1961). In the present study area, which is situated farther to the south, the Sudr Formation is distributed in the low lands between young-er highs. No complete section of this formation has been measured in the present study, but only ~28.5 m at Gebel Nukhul and ~44.5 m at Wadi Feiran. The measured part is composed of chalk, chalky limestones, argillaceous lime-stones and marly limestones belonging to the upper mem-ber of this formation (the Abu Zeneima Member). Based on calcareous nannoplankton, this part is assigned to the Late Maastrichtian. At Gebel Nukhul, it yields the Micula murus and Nephrolithus frequens Subzones (CC25c and CC26a). At Wadi Feiran, the Reinhardtites levis Zone (CC24), Arkhangelskiella cymbiformis, Lithraphidites quadratus, Micula murus Subzones (CC25a, b, c), Neph-rolithus frequens Subzone (CC26a) and lower part of the Micula prinsii Subzone (CC26b) are recognized. In both sections, the Sudr Formation is overlain by the Dakhla Formation with seeming conformity indicated by the presence of the Late Maastrichtian calcareous nannofos-sil Micula murus, Nephrolithus frequens and Micula prinsii Subzones. Although this conformable relationship was similarly documented by several authors in west-central Sinai (e.g. issawi et al., 1981; Cherif et al., 1989), some others noted a gap at the Sudr/Dakhla con-tact in neighbouring localities in this area (e.g. faris et al., 2005).
Dakhla Formation (Said, 1961)The type section for this unit lies along the scarp north of Mut at the Dakhla Oasis, Western Desert of Egypt. There, it was described to comprise yellowish to grayish shale, marl and clay with calcareous sandy and silty interbeds. It overlies the phosphate beds of the Duwi Formation and underlies the chalk and chalky limestone of the Tarawan Formation. At Gebel Nukhul, the Dakhla Formation con-sists of chalky argillaceous limestone and gray shale in
the lower part and calcareous gray shale in the upper part. At Wadi Feiran, it consists of multicoloured shale (grey, brown, black, greenish yellow) in its lower part and grades into grey marl in its upper part. The thickness of this formation varies from 32.4 m at Gebel Nukhul to 32 m at Wadi Feiran. The base of the Dakhla Formation comprises the Micula prinsii Subzone (CC26b) at the two studied sections, while its upper limit occurs within the lower part of the Discoaster mohleri Zone (NP7/8). Therefore, the Dakhla Formation is assigned to the latest Maastrichtian-Selandian age. Whereas the stratigraphic relationship with the overlying Tarawan Formation at Wadi Fieran is conformable, it is characterized by a mi-nor hiatus at Gebel Nukhul resulting in the absence of the Heliolithus kleinpellii Zone (NP6).
Tarawan Formation (awad & Ghobrial, 1965)At Gebel Tarawan in the Kharga Oasis area, the type lo-cality for this formation, it consists of chalk and chalky limestone. It rests on top of the Dakhla Formation and underlies the Esna Formation. In the studied sections, the Tarawan Formation is made up of yellow marl with chert bands changes into chalky limestone towards the top. The thickness of this formation at Gebel Nukhul and Wadi Feiran sections attains up to 2 m, and 1.2 m, respectively. The light colour and partly resistant appearance of the limestone make this unit clearly visible against the over-lying and underlying dark-coloured soft shale, and hence can be used as a good marker bed in the field. The Tar-awan Formation is assigned to the Late Paleocene (Thane-tian) as it occupies the calcareous nannofossil Zone Dis-coaster mohleri (NP7/8). In Wadi Feiran section, it lies conformably between two dark shale units (Dakhla and Esna) with no remarkable hiatus. At Gebel Nukhul, a mi-nor hiatus has been recorded at the Dakhla/Tarawan con-tact, indicated by the presence of erosion surface and absence of the Heliolithus kleinpellii Zone (NP6).
Esna Formation (beadnell, 1905)This formation consists of laminated, green and grey-green shale and clay in its type section at Gabal Oweina in the southern Nile Valley area. In the studied sections, it is made up of green-grey, brownish or blackish-grey shale and marl, either fissile or massive, with intercalated limestone bands and some gypsum veinlets. The total thicknesses of this unit are ~23.5 m and ~19.3 m in Geb-el Nukhul and Wadi Feiran sections, respectively. It con-formably overlies the Tarawan Formation and underlies the Thebes Formation. Based on the recognized calcare-ous nannofossil Discoaster multiradiatus, Tribrachiatus contortus and Discoaster bindosus Zones (NP9, NP10 and NP11), the Esna Formation is assigned as Late Paleo-cene (Thanetian) - Early Eocene (Ypresian).
Thebes Formation (Said, 1961)The Thebes Formation was first described at Gebel Gur-nah, opposite Luxor, behind the famous temple of El-Deir El-Bahari, southern Nile Valley. This formation is mainly

162 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Fig. 2: Detailed stratigraphic section of Gebel Nukhul.

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 163
Fig. 3: Detailed stratigraphic section at Wadi Feiran.

164 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
composed of marl and limestone with flint bands. Only the lowermost parts of the Thebes Formation at Gebel Nukhul (21.5 m thick.) and Wadi Feiran (40.8 m thick.) were measured. It conformably overlies the Esna Forma-tion in Wadi Feiran section. On the other hand, at Gebel Nukhul section, an unconformable relationship is ob-served between these two formations. Based on the pres-ence of the Discoaster bindosus and Tribrachiatus ortho-stylus Zones (NP11 and NP12), the Thebes Formation is assigned to the Early Eocene (Ypresian).
III. MATERIAL AND METHODOLOGY
The studied ostracode material is yielded from 96 rock samples collected from two sections measured at Gebel Nukhul and Wadi Feiran in west-central Sinai. From each sample, 100-200 grams were impregnated with a soda solution and washed over a 0.063 mm sieve. The investi-gated ostracodes were picked from the >250 µm residue size-fraction; the smaller size - fractions contained only juveniles. The number of individuals mentioned for each species is the sum of the adult carapaces and the largest number of either right or left adult valves. The study ma-terial is stored at the Geology Department, Faculty of Sci-ence, Tanta University (Tanta, Egypt). Representative specimens for all identified species have been photo-graphed by the first author using the Cam Scan SEM of Bremen University, Germany. Reference numbers are only given to the illustrated specimens, which are pre-served amongst the first author’s collection at the Geol-ogy Department, Faculty of Science, Ain Shams Univer-sity (Cairo, Egypt).
IV. SYSTEMATIC PALEONTOLOGY
The taxonomy is based on harTmann & puri (1974) and later established genera are treated according to their au-thors. The ostracode fauna gained from the studied Maas-trichtian to Lower Eocene succession are taxonomically classified into 38 species and subspecies belonging to 26 genera and 10 families. Species are arranged in alpha-betical order within each genus, those in open nomencla-ture at the end.
Subclass Ostracoda latreille, 1806Order Podocopida Müller, 1894Suborder Platycopa SarS, 1866Family Cytherellidae SarS, 1866
Genus Cytherella JoneS, 1849Type species: Cytherina ovata roemer, 1840
Cytherella cf. lagenalis Marliere, 1959Pl. I, figs 1-3
cf. 1959. Cytherella lagenalis marliere, p. 8, pl. 1, fig. 3.1999. Cytherella cf. lagenalis marliere. - morsi, p. 33, pl.
1, figs 1-3.2000. Cytherella cf. lagenalis marliere. - morsi, p. 50, pl.
1, figs 7-8.2004. Cytherella cf. lagenalis marliere. - ismail & ied, p.
99, pl. 1, figs 3-5.
Material: 11 specimens.Dimensions: L: 0.86-0.98 mm; H: 0.53-0.58 mm; H: 0.35-0.38 mm. Occurrence: Maastrichtian to Paleocene of east Sinai, Egypt (morsi, 1999, 2000; ismail & ied, 2004). In the present study, it occurs in the Late Maastrichtian to Early Eocene (Micula murus Subzone to Discoaster binodosus Zone).
Cytherella sinaensis MorSi, 1999Pl. I, figs 4-5
1999. Cytherella sinaensis morsi, p. 33, pl. 1, figs 4-6.2000. Cytherella sinaensis morsi. - morsi, p. 50, pl. 1, figs
9-10.
Material: 82 specimens.Dimensions: L: 0.81-0.94 mm; H: 0.49-0.57 mm; W: 0.29 mm.Occurrence: Maastrichtian and Paleocene of east-cen-tral Sinai, Egypt (morsi, 1999, 2000). Here, it is found in the Late Maastrichtian-Early Eocene (Reinhardtites levis - Tribrachiatus contortus Zones).
Genus Cytherelloidea alexander, 1929Type species: Cytherella williamsoniana jones, 1849
Cytherelloidea attiyaensis MorSi, 1999Pl. I, figs 6-7
1996. Cytherelloidea monmouthensis jenninGs. - ismail, p. 42, pl. 1, fig. 5.
1999. Cytherelloidea attiyaensis morsi, p. 37, pl. 1, figs 7-9.2005. Cytherelloidea melleguensis damoTTe & said.- sha-
hin, p. 754, pl. 1, figs 6-8. Material: 10 specimens.Dimensions: L: 0.57-0.61 mm; H: 0.36-0.40 mm.Discussion: The specimens illustrated by shahin (2005) as Cytherelloidea melleguensis damoTTe & said from the Paleocene of Egypt fit well into the present species as they possess the characteristic dorsal triangular rib with an incomplete lower arm. In C. melleguensis da-moTTe & said (in donze et al., 1982) the dorsal rib is shorter and a crescentric median rib, which is lacking in C. attiyaensis, is developed.Occurrence: Early Paleocene of Sinai, Egypt (ismail, 1996; morsi, 1999; shahin, 2005). In the present study, it is similarly recorded in the Early Paleocene (Crucipla-colithus tenuis - Ellipsolithus macellus Zones).

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 165
Cytherelloidea melleguensis daMotte & Said, 1982Pl. I, figs 8-10
1978. Cytherelloidea sp. said, p. 215, pl. 24, fig. 6.1982. Cytherelloidea melleguensis damoTTe & said, in donze
et al., p. 280, pl. 1, figs 2-5.1987. Cytherelloidea melleguensis damoTTe & said. - da-
moTTe & fleury, p. 93, pl. 1, fig. 6.1998. Cytherelloidea melleguensis damoTTe & said. - said-
benzarTi, pl. 1, fig. 4.2000. Cytherelloidea melleguensis damoTTe & said. - morsi,
p. 54, pl. 1, figs 11-12.non 2005. Cytherelloidea melleguensis damoTTe & said. -
shahin, p. 754, pl. 1, figs 6-8.
Material: 5 specimens.Dimensions: L: 0.68 mm; H: 0.43 mm; W: 0.27 mm.Occurrence: Late Campanian-Maastrichtian of Tunisia (said, 1978; donze et al., 1982; said-benzarTi, 1998), east Algeria (damoTTe & fleury, 1987) and east-cen-tral Sinai, Egypt (morsi, 2000). In the present area, it is also recorded in the Late Maastrichtian (Micula murus - Micula prinsii Subzones).
Suborder Podocopa SarS, 1866Superfamily Bairdiacea SarS, 1866
Family Bairdiidae SarS, 1866Genus Bairdia Mccoy, 1844
Type species: Bairdia curtus mCCoy, 1844.
Bairdia ilaroensis reyMent & reyMent, 1959Pl. I, figs 11, 13-14
1959. Bairdia ilaroensis reymenT & reymenT, p. 61, pl. 1, figs 1-7, text-figs 1a-b, 3a-m, 5a-h.
1981. Bairdia ilaroensis reymenT & reymenT. - rey-menT, p. 56, pl. 9, fig. 6-7.
1983. Bairdoppilata ilaroensis (reymenT & reymenT). - fosTer et al., p. 109, pl. 1, figs 5, 7-11.
1987. Bairdia ilaroensis reymenT & reymenT. - oKo-sun, p. 26, pl. 20, figs 7, 9-10.
1990. Bairdia ilaroensis reymenT & reymenT. - bassiou-ni & luGer, p. 780, pl. 1, fig. 15.
1990. Bairdoppilata ilaroensis (reymenT & reymenT). - Carbonnel et al., p. 674, pl. 1, fig. 19.
non 1991. Bairdoppilata ilaroensis (reymenT & reymenT). - Carbonnel & oyede, p. 69, pl. 1, fig.1.
non 1992. Bairdia ilaroensis reymenT & reymenT. - ismail, p. 45, pl. 1, fig. 5.
1992. Bairdia ilaroensis reymenT & reymenT. - el-waer, p. 47, pl. 4, figs 4-9.
1994. Bairdia gr. ilaroensis reymenT & reymenT. - Keen et al., pl. 16, fig. 5.
?1996. Bairdia ilaroensis reymenT & reymenT. - bassiouni & luGer, p. 8, pl. 1, figs 10-13.
1998. Bairdoppilata ilaroensis (reymenT & reymenT). - Colin et al., p. 291, pl. 1, figs 4-5.
2003. Bairdia ilaroensis reymenT & reymenT. - mor-si & speijer, p. 70, pl. 1, fig. 5.
Material : 9 specimens.
Dimensions : L : 0.88 -0.98 mm; H : 0.57-0.68 mm; W : 0.45-0.47 mm. Occurrence : The present species is widely distributed in the Maastrichtian to Early Eocene of West and North Af-rica. In Egypt, it is also known from the Maastrichtian to Early Eocene (bassiouni & luGer, 1990; morsi & spei-jer, 2003). In the present study, it is found in the Early Eocene (Discoaster multiradiatus - Tribrachiatus contor-tus Zones).
Bairdia aff. septentrionalis bonneMa, 1941Pl. I, fig. 12
aff. 1941. Bairdia septentrionalis bonnema, p. 108, pl. 2, figs 55-64; pl. 3, figs 1-8.
1995. Bairdia sp.1 honiGsTein & rosenfeld, p. 52, pl. 1, fig. 3.
1996. Bairdoppilata sp. ismail, p. 43, pl. 1, fig. 11.1999. Bairdia aff. septentrionalis bonnema. - morsi, p.
37, pl. 1, fig. 10.2000. Bairdia aff. septentrionalis bonnema. - morsi, p.
51, pl. 1, fig. 13.
Material : 61 specimens.Dimensions : L : 1.07-1.72 mm; H : 0.7-1.12 mm.Remarks : The present specimens closely resemble Bair-dia septentrionalis bonnema, 1941, from the Cretaceous of northeastern Holland, but differ in having a broader anterior margin.Occurrence : Paleocene of southern Israel (honiG-sTein & rosenfeld, 1995) and Maastrichtian-Paleocene of Egypt (ismail, 1996; morsi, 1999, 2000). In the pres-ent study, it occurs in the Late Maastrichtian-Paleocene (Reinhardtites levis - Discoaster mohleri Zones).
Bairdia sp.Pl. I, figs 15-16
Material : 4 specimens.Dimensions : L : 0.95-1.0 mm; H : 0.57-0.63 mm.Occurrence : Early Eocene (Tribrachiatus contortus Zone).
Superfamily Cypridacea baird, 1845Family Candonidae KaufMann, 1900Subfamily Paracypridinae SarS, 1923
Genus Paracypris SarS, 1866Type species : Paracypris polita sars, 1866.
Paracypris aff. jonesi bonneMa, 1941Pl. I, fig. 17
aff. 1941. Paracypris jonesi bonnema, p. 115, pl. 3, figs 24- 28.
1966. Paracypris n.sp. salahi, p. 7, pl. 1, figs 29-30, 34.1968. Paracypris jonesi bonnema. - esKer, p. 282, pl. 1,
fig. 13.1990. Paracypris jonesi bonnema. - bassiouni & luGer,

166 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
p. 83, pl. 2, figs 9, 11. 2000. Paracypris jonesi bonnema. - bassiouni & morsi,
p. 37, pl. 3, figs 1-4. 2005. Paracypris jonesi bonnema. - ismail & ied, p. 127,
pl. 2, fig. 1.
Material : One specimen.Dimensions : L : 1.10 mm; H : 0.40 mm.Remarks : Although the present specimen is strikingly similar to Paracypris jonesi which was originally de-scribed from the Maastrichtian of the Netherlands (bonnema, 1941), it has been assigned as Paracypris aff. jonesi since the nominate species originally comes from the Maastrichtian of northern Europe which be-longed to a different bioprovince during the Paleocene. Occurrence : Paleocene of Libya (salahi, 1966), Tuni-sia (esKer, 1968) and Paleocene to Early Eocene of Egypt (bassiouni & luGer, 1990; bassiouni & morsi, 2000; ismail & ied, 2005). In the present material, it is found in the Early Eocene (Tribrachiatus contortus Zone).
Family Pontocyprididae Müller, 1894Genus Abyssocypris Van den bold, 1974
Type species : Abyssocypris tipica Van den bold, 1974.
Abyssocypris? adunca (eSKer, 1968)Pl. I, fig. 18
1968. “Bythocypris” adunca esKer, p. 321, pl. 2, figs 10- 12; pl. 4, fig. 4.
1982. Abyssocypris? adunca (esKer). - donze et al., p. 281, pl. 2, figs 3-4.
1987. Abyssocypris? adunca (esKer). - damoTTe & fleury, p. 93, pl. 1, fig. 10.
1990. “Bythocypris” adunca esKer. - bassiouni & luGer, p. 781, pl. 2, fig. 4.
1998. Abyssocypris? adunca (esKer). - said-benzarTi, pl. 2, fig. 13.
2000. “Bythocypris” adunca esKer. - bassiouni & morsi, p. 33, pl. 2, fig. 3.
Material : 6 specimens.Dimensions : L : 0.60 mm; H : 0.30 mm.Occurrence : Late Campanian and Paleocene of Tunisia (esKer, 1968; donze et al., 1982; said-benzarTi, 1998), Paleocene of Algeria (damoTTe & fleury, 1987) and Late Paleocene - Early Eocene of Egypt (bassiou-ni & luGer, 1990; bassiouni & morsi, 2000). In the present material, it occurs in the Late Maastrichtian-Early Paleocene (Lithraphidites quadratus Subzone - Cruci-placolithus tenuis Zone).
Genus Pontocyprella lyubiMoVa, 1955Type species : Bairdia harrisiana jones, 1849.
Pontocyprella recurva eSKer, 1968Pl. II, fig. 1
1968. Pontocyprella recurva esKer, p. 323, pl. 1, figs 6-7.1978. Pontocyprella? cf. recurva esKer. - said, p. 220, pl.
24, fig. 15.1982. Pontocyprella recurva esKer. - bouKhary et al.,
pl. 2, fig. 10.1982. Pontocyprella recurva esKer. - donze et al., p.
281, pl. 2, figs 1-2.pars1987. Pontocyprella recurva esKer. - damoTTe & fleu-
ry, p. 93, pl. 1, fig. 12. 1990. Pontocyprella recurva esKer. - bassiouni & lu-
Ger, p. 785, pl. 3, fig. 12.1992. Pontocyprella recurva esKer. - el-waer, p.73, pl.
57, fig. 1-3. non 1996. Pontocyprella recurva esKer. – andreu, p. 105, pl.
1, figs 15-16.1998. Pontocyprella recurva esKer. - said-benzarTi, pl.
1, fig. 13.1999. Pontocyprella recurva esKer. - morsi, p. 38, pl. 1,
fig. 12.2000. Pontocyprella recurva esKer. - morsi, p. 51, pl. 1,
fig. 14.2000. Pontocyprella recurva esKer. - bassiouni & mor-
si, p. 37, pl. 3, figs 7-8.2004. Pontocyprella recurva esKer. - ismail & ied, p.
101, pl. 1, fig. 8.2005. Pontocyprella recurva esKer. - ismail & ied, p.
127, pl. 2, fig. 2.
Material : 11 specimens.Dimensions : L : 0.95-1.11 mm; H : 0.47-0.6 mm.Discussion : The specimens identified as Pontocyprella recurva Esker, 1968 from the Coniacian-Early Campan-ian of Morocco (andreu, 1996) deviate from the origi-nal illustration of esKer for this species from the Danian of Tunisia (esKer, 1968, pl. 1, fig 6) in being more elon-gate and having its posterodorsal margin gentler sloping.Occurrence : Maastrichtian-Paleocene of Tunisia (said, 1978; donze et al., 1982; said-benzarTi, 1998), Maas-trichtian-“Middle” Paleocene of Algeria (damoTTe & fleury, 1987), Maastrichtian of Libya (el-waer, 1992) and Maastrichtian-Early Eocene of Egypt (bouKhary et al. 1982, bassiouni & luGer, 1990; morsi, 1999; bassiouni & morsi, 2000; morsi, 2000; ismail & ied, 2004, 2005). In the present study, it is re-corded in the Late Maastrichtian-Early Paleocene (Neph-rolithus frequens Subzone Ellipsolithus macellus Zone).
Genus Argilloecia SarS, 1866Type species : Argilloecia cylindrica sars, 1866
Argilloecia sp. baSSiouni & luGer, 1990Pl. II, figs 2-4
1990. Argilloecia sp. bassiouni & luGer, p. 785, p1. 3, figs 2-4.
Material : 9 specimens.Dimensions : L : 0.49-0.53 mm; H : 0.22-0.24 mm; W : 0.2 mm.Occurrence : “Middle” to Late Paleocene of southern

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 167
Egypt (bassiouni & luGer, 1990). In the present work, it is found in the Late Maastrichtian-Early Eocene (Lithraphidites quadratus Subzone - Tribrachiatus contortus Zone).
Superfamily Cytheracea baird, 1850Family Cytheridae baird, 1850
Subfamily Cytherinae baird, 1850Genus Apateloschizocythere bate, 1972
Type species : Apaterloschizocythere geniculata BaTe, 1972.
Apateloschizocythere fimbrata baSSiouni & luGer, 1990
Pl. II, fig. 5
1990. Apateloschizocythere fimbrata bassiouni & luGer, p. 816, pl. 12, figs 16-21.
2004. Amphicytherura? sp. ismail & ied, p. 105, pl. 2, figs 1-2.
Material : A single specimen.Dimensions : L : 0.48 mm; H : 0.28 mm. Occurrence : Maastrichtian of Egypt (bassiou-ni & luGer, 1990; ismail & ied, 2004). In the present material, it is similarly recorded in the Late Maastrichtian (Reinhardtites levis Zone).
Genus Paraschizocythere el Sweify, 1984Type species : Paraschizocythere nodoreticulata el sweify, 1984.
Paraschizocythere hirsutonodosa el Sweify, 1984Pl. II, fig. 6
1982. Schizocythere? sp. donze et al., p. 282, p1. 2, figs 11-12; p1. 3, fig. 1.
1984. Paraschizocythere hirsutonodosa el sweify, p. 39, pl. 1, figs 8-10.
1990. Paraschizocythere hirsutonodosa el sweify. - bassiou-ni & luGer, p. 818, pl. 13, figs 7-11.
Material : A single specimen.Dimensions : L : 0.51 mm; H : 0.31 mm. Occurrence : Late Middle to early Late Maastrichtian of Tunisia (donze et al., 1982) and Middle Maastrichtian of Egypt (el sweify, 1984; bassiouni & luGer, 1990). In the present material it is recorded in the latest Maas-trichtian (Micula prinsii Subzone).
Family Bythocytheridae SarS, 1926Genus Bythoceratina hornibrooK, 1952
Type species : Bythoceratina mestayerae hornibrooK, 1952.
Bythoceratina hamzai baSSiouni & MorSi, 2000Pl. II, fig. 7
2000. Bythoceratina hamzai bassiouni & morsi, p. 40, pl. 4, figs 3-4.
Material : A single specimen.Dimensions : L : 0.38-0.4 mm; H : 0.69-0.73 mm.Discussion : Bythoceratina sp. bassiouni & luGer, 1990 from the Late Paleocene of southern Egypt is a strikingly similar species. It deviates, however, in having the anterior inflation overhanging dorsal margin and the vertical sulcus more anteriorly shifted.Occurrence : Early Eocene of the Farafra area in the Western Desert of Egypt (bassiouni & morsi, 2000). Here, it is also found in the Early Eocene (Tribrachiatus contortus Zone).
Family Krithidae MandelStaM, 1958Subfamily Krithinae MandelStaM, 1958
Genus Krithe brady, croSSKey & robertSon, 1874Type species : Ilyobates praetexta sars, 1866.
Krithe echolsae eSKer, 1968Pl. II, figs 8-10
1968. Krithe echolsae esKer, p. 33, pl. 3, figs 1-4.1978. Krithe echolsae esKer. - said, p. 232, pl. 26, figs 1-4.1982. Krithe echolsae esKer. - bouKhary et al., pl. 2, figs 8-9.1982. Krithe echolsae esKer. - donze et al., p. 283, fig. 4.1987. Krithe echolsae esKer. - honiGsTein et al., p. 42, pl. 1,
figs 5-6.1990. Krithe echolsae esKer. - bassiouni & luGer, p. 795, pl.
6, figs 10-11. 1995. Krithe echolsae esKer. - honiGsTein & rosenfeld, p.
53, pl. 1, figs 4-5.1999. Krithe echolsae esKer. - morsi, p. 38, pl. 1, figs 13-15.2000. Krithe echolsae esKer. - morsi, p. 56, pl. 1, figs 16-17.2000. Krithe echolsae esKer. - bassiouni & morsi, p. 43, pl.
5, figs 2-3.2004. Krithe echolsae esKer. - ismail & ied, p. 102, pl. 1, figs
9-10.
Material : 18 specimens.Dimensions : L : 0.69-0.82 mm; H : 0.34-0.43 mm; H : 0.33 mm.Occurrence : Late Campanian-Paleocene of Tunisia (es-Ker, 1968; said, 1978; donze et al., 1982; said-ben-zarTi, 1998), and Maastrichtian - Paleocene of Israel (honiGsTein et al., 1987; honiGsTein & rosenfeld, 1995) and Egypt (bouKhary et al., 1982; bassiou-ni & luGer, 1990; morsi, 1999, 2000; bassiou-ni & morsi, 2000; ismail & ied, 2004, 2005; shahin, 2005). In the present work, it is found in the Late Maas-trichtian-Early Eocene (Reinhardtites levis - Tribrachia-tus contortus Zones).
Krithe cf. solomoni honiGStein, 1984Pl. II, figs 11-12
cf. 1984. Krithe solomoni honiGsTein, p. 11, pl. 6, figs 9-12.1990. Krithe cf. solmoni honiGsTein. - bassiouni &
luGer, p. 795, pl. 6, figs 3, 6, 9, 12.
Material : 4 specimens.

168 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Dimensions : L : 0.27-0.30 mm; H : 0.57 mm. Remarks : The present material is closely similar to the nominate species; Krithe solomoni honiGsTein, howev-er, deviates in having its dorsal margin more sharply dip-ping downward at posterior end forming a more blunt concavity at upper part of posterior margin.Occurrence : “Middle” and Late Paleocene of southern Egypt (bassiouni & luGer, 1990). Here, it is recorded in the Late Maastrichtian (Micula murus Subzone) and Early Paleocene (Chiasmolithus danicus Zone).
Genus Parakrithe Van den bold, 1958Type species : Cytheridea (Dolocytheridea) vermunti Van den bold 1958.
Parakrithe crolifa baSSiouni & luGer, 1990Pl. II, figs 13-14
1979. Krithe sp. Cronin & Khalifa, p. 410, pl. 1, figs 26-17.pars 1981. Bythocypris kalambainaensis reymenT, p. 56, pl. 1,
fig. 4.1990. Parakrithe crolifa bassiouni & luGer, p. 796, pl.
6, figs 13-22.1995. Parakrithe? kalambainaensis reymenT. - honiG-
sTein & rosenfeld, p. 53, pl. 1, fig. 7. 1999. Parakrithe crolifa bassiouni & luGer. - morsi, p.
38, pl. 1, fig. 16.2000. Parakrithe crolifa bassiouni & luGer. - bassiou-
ni & morsi, p. 44, pl. 5, fig. 4.2002. Parakrithe crolifa bassiouni & luGer. - honiGs-
Tein et al., p. 372, pl. 1, fig. 12.2004. Parakrithe crolifa bassiouni & luGer. - ismail & ied,
p. 103, pl. 1, figs 11-12.2005. Parakrithe crolifa bassiouni & luGer. - shahin, p.
759, pl. 2, fig. 17.2005. Parakrithe crolifa bassiouni & luGer. - ismail & ied,
p. 131, pl. 2, fig. 16.
Material : 45 specimens.Dimensions : L : 0.52-0.63 mm; H : 0.25-0.31 mm.Occurrence : Late Paleocene of Nigeria (reymenT, 1981), Paleocene-Eocene of Israel (honiGsTein & rosen-feld, 1995; honiGsTein et al., 2002) and Maastrichtian - Middle Eocene of Egypt (Khalifa & Cronin, 1979, bassiouni & luGer 1990; morsi, 1999; bassiou-ni & morsi, 2000; ismail & ied, 2004, 2005). In the present study, it is recorded in the Late Maastrichtian-Early Eocene (Reinhardtites levis - Discoaster binodosus Zones).
Family Hemicytheridae Puri, 1953Subfamily Thaerocytherinae hazel, 1967Genus Martinicythere baSSiouni, 1969a
Type species : Martinicythere samalutensis bassiouni, 1969a.
Martinicythere bassiounii bassiounii honiGStein & roSenfeld, 1995
Pl. II, figs 15-17
1969a.Martinicythere sp. 3 bassiouni, pl. 19, fig. 6.1982. Martinicythere cf. vesiculosa (aposTolesCu). - donze et
al., p. 287, pl. 5, figs 9-10; pl. 14, figs 4-5.1987. Martinicythere cf. vesiculosa (aposTolesCu). - da-
moTTe & fleury, p. 95, pl. 2, figs 20-21.1995. Cristaeleberis reticulata bassiouni. - honiG-
sTein & rosenfeld, p. 56, pl. 2, figs 7-9.1995. Martinicythere bassiounii honiGsTein & rosenfeld, p. 60,
pl. 3, figs 7-8.1999. Martinicythere bassiounii honiGsTein & rosenfeld. -
morsi, p. 39, pl. 2, figs 1-4.2000. Martinicythere bassiounii honiGsTein & rosenfeld. -
morsi, p. 57, pl. 2, figs 8-10.2003. Martinicythere bassiounii bassiounii honiGsTein & rosen-
feld. - morsi & speijer, p. 73, pl. 3, figs 24-25.
Material : 31 specimens.Dimensions : L : 0.52-0.61 mm; H : 0.30-0.38 mm; W : 0.29 mm.Occurrence : Paleocene of Syria (bassiouni, 1969a) and Israel (honiGsTein & rosenfeld, 1995), Campan-ian to Early Paleocene of Tunisia (donze et al., 1982) and east Algeria (damoTTe & fleury, 1987) and Maas-trichtian to Late Paleocene of Egypt (morsi, 1999, 2000; morsi & speijer, 2003). In the present material, it is found in the Late Maastrichtian-Early Paleocene (Micula murus Subzone - Ellipsolithus macellus Zone).
Family Trachyleberididae SylVeSter-bradley, 1948
Subfamily Trachyleberidinae SylVeSter-bradley, 1948
Genus Acanthocythereis howe, 1963Type species : Acanthocythereis araneosa Howe, 1963.
Acanthocythereis? denticulata eSKer, 1968Pl. II, figs 18-19
1968. Acanthocythereis denticulata esKer, p. 328, pl. 2, figs 6-7; pl. 4, fig. 1.
1982. Acanthocythereis? denticulata (esKer). - donze et al., p. 293, pl. 11, figs 1-4.
1987. Acanthocythereis? denticulata (esKer). - da-moTTe & fleury, p. 97, pl. 3, fig. 11.
1990. Megommatocythere denticulata (esKer). - bassiou-ni & luGer, p. 825, pl. 7, fig. 7.
?1991. Megommatocythere cf. denticulata (esKer). - hon-iGsTein et al., p. 105, pl. 2, figs 11-12.
?1995. Megommatocythere cf. denticulata (esKer). - hon-iGsTein & rosenfeld, p. 53, pl. 1, fig. 8.
1999. Megommatocythere denticulata (esKer). - morsi, p. 41, pl. 2, fig. 15.
2000. Megommatocythere denticulata (esKer). - bassiou-ni & morsi, p. 58, pl. 8, fig. 17.
?2002. Megommatocythere cf. denticulata (esKer). - hon-iGsTein et al., p. 378, pl. 3, fig. 13.
?pars 2004. Megommatocythere denticulata (esKer). - is-mail & ied, p. 108, pl. 3, fig. 3.
?2005. Megommatocythere denticulata (esKer). - shahin, p. 361, pl. 3, fig. 13.
2005. Megommatocythere denticulata (esKer). - is-mail & ied, p. 138, pl. 3, fig. 14.

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 169
Material : 46 specimens.Dimensions : L : 0.99-1.06 mm; H : 0.53-0.56 mm; W : 0.49-0.53mm. Remarks : Very closely similar, but shorter, specimens were illustrated from the Paleocene-Middle Eocene of Israel (honiGsTein et al., 1991; honiGsTein & rosen-feld 1995; honiGsTein et al. 2002) and Maastrichtian-Early Eocene of northeastern Sinai, Egypt (ismail & ied, 2004; shahin, 2005). It is probable that these specimens represent an intraspecific variation of the present spe-cies. Occurrence : Late Maastrichtian to Early Paleocene of Tunisia (esKer, 1968; donze et al., 1982), Early - “Mid-dle” Paleocene of Algeria (damoTTe & fleury, 1987) and Maastrichtian-Early Eocene of Egypt (bassiou-ni & luGer, 1990; morsi, 1999; bassiouni & morsi 2000; ismail & ied, 2005). In the present study, it is re-corded in the Late Maastrichtian-Early Eocene (Nephro-lithus frequens Subzone - Discoaster binodosus Zone).
Acanthocythereis meslei donze & oertli, 1982
Two subspecies are known for this species : Acanthocy-thereis meslei meslei donze & oerTli, 1982 and Acan-thocythereis meslei paleocenica bassiouni & luGer, 1990. In the present work, only the nominate subspecies is recorded.
Acanthocythereis meslei meslei donze & oertli, 1982
Pl. II, figs 20-21
1982. Acanthocythereis? meslei donze & oerTli, in donze et al., p. 292, pl. 10, figs 1-10.
1987. Acanthocythereis? meslei donze & oerTli. - da-moTTe & fleury, p. 96, pl. 3, fig. 10.
?1996. Acanthocythereis meslei donze & oerTli. - andreu, p. 112, pl. 2, figs 10-12.
1999. Acanthocythereis meslei donze & oerTli. - morsi, p. 40, pl. 2, fig. 10.
Material : 4 specimens.Dimensions : L : 0.95-1.01 mm; H : 0.48-0.56 mm.Occurrence : The specimens assigned by andreu (1996) from the Coniacian-Early Campanian of Morocco as Acanthocythereis meslei are closely similar. However, they deviate in having a subtriangular posterior margin with a more pointed end, unlike in the original figures in donze et al. (1982, pl. 10, figs 1-10) where posterior end is broader and more rounded. Occurrence : Late Campanian and Maastrichtian of Tu-nisia (donze et al., 1982), Maastrichtian and earliest Pa-leocene of east Algeria (damoTTe & fleury, 1987) and Early Paleocene of Egypt (morsi, 1999). Here, it is re-corded in the Late Maastrichtian (Lithraphidites quadra-
tus - Nephrolithus frequens Subzones).
Acanthocythereis? posterotriangulata (MorSi, 1999)Pl. II, figs 22-23
1990. Oertliella? sp. bassiouni & luGer, p. 831, pl. 18, figs 6-10.
1995. Oertliella? cf. frescoensis (aposTolesCu). - honiG-sTein & rosenfeld, p. 58, pl. 3, fig. 8.
1999. Oertliella posterotriangulata morsi, p. 41, pl. 3, figs 1-5.2000. Oertliella? sp. bassiouni & luGer. - bassiouni & mor-
si, p. 35, pl. 9, figs 12-15.2003. Oertliella posterotriangulata morsi. - morsi & speijer,
p. 75, pl. 4, figs 36-37.
Material : 6 specimens.Dimensions : L : 0.60 (female), 0.69 mm (male); H : 0.33 mm (female), 0.38 mm (male).Occurrence : Paleocene to Early Eocene of Egypt (bassiouni & luGer, 1990; morsi, 1999; bassiou-ni & morsi, 2000; morsi & speijer, 2003) and Paleo-cene of southern Israel (honiGsTein & rosenfeld, 1995). In the present material, it is recorded from the Early Paleocene to Early Eocene (Cruciplacolithus tenuis - Tribrachiatus contortus Zones).
Genus Actinocythereis Puri, 1953Type species : Cythere exanthemata ulriCh & bassler, 1904.
Actinocythereis? coronata (eSKer, 1968)Pl. III, figs 1-3
1968. Cythereis coronata esKer, p. 323, pl. 1, figs 1-3; pl. 4, fig. 5.
1970. Mauritsina arabica bassiouni, p. 21, pl. 2, figs 8-9.1978. Actinocythereis? arabica (bassiouni). - said, p. 239, pl.
25, figs 20-21.1982. Mauritsina coronata (esKer). - bouKhary et al., pl. 1,
figs 5-8.1982. Actinocythereis? coronata (esKer). - donze et al., p.
291, pl. 9, figs 7-10; pl. 14, fig. 8.1987. Actinocythereis? coronata (esKer). - damoTTe & fleu-
ry, p. 96, pl. 3, figs 8-9.1990. Mauritsina coronata (esKer). - bassiouni & luGer, p.
812, pl .11, figs 13-15.1992. Mauritsina coronata (esKer). - ismail, p. 46, pl. 1, fig. 9.1995. Mauritsina coronata (esKer). - honiGsTein & rosen-
feld, p. 58, pl. 3, fig. 10.1996. Mauritsina coronata (esKer). - ismail, p. 44, pl. 2, figs
6-7.1998. Actinocythereis? coronata (esKer). - said-benzarTi, pl.
2, figs 21-22.1999. Mauritsina coronata (esKer). - morsi, p. 39, pl. 2, fig. 5.2000. Mauritsina coronata (esKer). - morsi, p. 57, pl. 2, fig. 11.2000. Mauritsina coronata (esKer). - bassiouni & morsi, p.
29, pl. 8, figs 1-2.2002. Mauritsina coronata (esKer). - honiGsTein et al., p.
376, pl. 3, fig. 4.2003. Mauritsina coronata (esKer). - morsi & speijer, p. 74,
pl. 3, fig. 29.

170 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
2004. Mauritsina coronata (esKer). - ismail & ied, p. 104, pl. 2, fig. 3.
2005. Mauritsina coronata (esKer). - ismail & ied, p. 134, pl. 3, fig. 5.
Material : 13 specimens.Dimensions : L : 0.80 mm (female), 0.92 mm (male); H : 0.50 mm (female), 0.56 mm (male).Occurrence : Late Campanian to Late Paleocene of Tu-nisia (esKer, 1968; said, 1978; donze et al., 1982; said-benzarTi, 1998), Maastrichtian to “Middle” Pa-leocene of east Algeria (damoTTe & fleury, 1987), Maastrichtian of Libya (Keen et al., 1994), Paleocene and Early Eocene of Israel (honiGsTein & rosenfeld, 1995; honiGsTein et al., 2002), Paleocene and Early Eo-cene of Jordan (bassiouni, 1970) and Maastrichtian to Early Eocene of Egypt (bouKhary et al. 1982; bassiou-ni & luGer, 1990; ismail, 1992, 1996; morsi 1999, 2000; bassiouni & morsi, 2000; morsi & speijer, 2003; ismail & ied, 2004, 2005). In the present material, it is found in the Late Maastrichtian-Early Eocene (Neph-rolithus frequens Subzone - Discoaster binodosus Zone).
Genus Aphrikanecythere daMotte & oertli, 1982Type species : Aphrikanecythere phumatoides da-moTTe & oerTli, 1982.
Aphrikanecythere phumatoides daMotte & oertli, 1982
Pl. III, figs 4-5
1978. Atlanticythere? sp. said, p. 242, pl. 27, figs 4-6.1982. Aphrikanecythere phumatoides Damotte & Oertli, in
donze et al., p. 288, pl. 6, figs 1-9.1987. Aphrikanecythere phumatoides Damotte & Oertli. - da-
moTTe & fleury, p. 95, pl. 2, figs 22-23.pars 2004. Trachyleberis spininodosa (el sweify). - ismail & ied,
p. 112, pl. 4, fig. 8; non fig. 9.
Material : 3 specimens.Dimensions : L : 0.73-0.74 mm; H : O.40 mm.Occurrence : Middle-Late Maastrichtian of Tunisia (said, 1978; donze et al, 1982) and Maastrichtian of east Algeria (damoTTe & fleury, 1987) and Egypt (is-mail & ied, 2004). Here, this species is found in the Late Maastrichtian (Arkhangelskiella cymbiformis Subzone).
Genus Cristaeleberis baSSiouni, 1970Type species : Cristaeleberis reticulata bassiouni, 1970.
Cristaeleberis reticulata baSSiouni, 1970Pl. III, fig. 6
1970. Cristaeleberis reticulata bassiouni, p. 26, pl. 3, figs 5-7.
1982. Cristaeleberis reticulata bassiouni. - bouKhary et
al., pl. 2, figs 5-7.?1984. Cristaeleberis reticulata bassiouni. - honiGsTein, p.
34, pl. 10, figs 1-4; pl. 15, figs 19-20.1984. Cristaeleberis reticulata bassiouni. - Khalifa et al.,
pl. 2, fig. 1a-b.1999. Cristaeleberis reticulata bassiouni. - morsi, p. 41, pl.
2, fig. 11.
Material : A single female specimen.Dimensions : L : 0.55 mm; H : 0.31 mm.Occurrence : “Middle” Paleocene of Jordan (bassiouni, 1970) and Paleocene of Egypt (bouKhary et al., 1982; Khalifa et al., 1984; morsi, 1999). In the present mate-rial, it is recorded in the Early Paleocene (Cruciplacoli-thus tenuis Zone).
Genus Cythereis JoneS, 1849Type species : Cytherina ciliata REUSS, 1846.
Remark : The genus Cythereis is widely known as a Cre-taceous genus that was extinct at the end of this period. Although the species recorded herein were originally de-scribed from the Late Cretaceous, their record later in the Paleocene makes the generic assignment questionable. The lack of separate valves among the present material did not allow for investigation of internal features for ge-neric inspection.
Cythereis? mesa honiGStein, 1984
Cythereis? mesa mesa honiGStein, 1985Pl. III, fig. 7
1984. Cythereis mesa ventroleviata honiGsTein, p. 22, pl. 13, figs 4-6.
1985. Cythereis mesa mesa. honiGsTein, p. 240.1993. Cythereis mesa mesa honiGsTein. - honiGsTein et al.,
pl. 1, figs 1-2, 6-8.1995. Cythereis mesa mesa honiGsTein. - honiGsTein & rosen-
feld, p. 60, pl. 3, fig. 3.1999. Cythereis mesa mesa honiGsTein. - morsi, p. 41, pl. 2,
fig. 12.2000. Cythereis mesa mesa honiGsTein. - morsi, p. 60, pl. 2,
fig. 14.2004. Cythereis mesa mesa honiGsTein. - ismail & ied, p.
107, pl. 2, figs 4-5.2005. Mauritsina teiskotensis (aposTolesCu). - shahin, p.
761, pl. 3, fig. 12.
Material : A single specimen.Dimensions : L : 1.00 mm; H : 0.56 mm. Occurrence : Late Santonian, Campanian and Paleocene of Israel (honiGsTein, 1984; honiGsTein et al., 1993; honiGsTein & rosenfeld, 1995) and Maastrichtian-Pa-leocene of Egypt (morsi, 1999, 2000; ismail & ied, 2004; shahin, 2005). In the present area, it occurs in the Early Paleocene (Cruciplacolithus tenuis Zone).
Cythereis? mesa ventroreticulata honiGStein, 1984Pl. III, fig. 8

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 171
1984. Cythereis mesa ventroreticulata honiGsTein, p. 22, pl. 13, figs 7-11.
1993. Cythereis mesa ventroreticulata honiGsTein. - honiG-sTein et al., pl. 1, figs 3, 5, 9, 10.
1995. Cythereis mesa ventroreticulata honiGsTein. - honiG-sTein & rosenfeld, p. 60, pl. 3, fig. 4.
1999. Cythereis mesa ventroreticulata honiGsTein. - morsi, p. 41, pl. 2, figs 13-14.
2000. Cythereis mesa ventroreticulata honiGsTein. - morsi, p. 60, pl. 2, fig. 15.
2004. Cythereis mesa ventroreticulata honiGsTein. - is-mail & ied, p. 107, pl. 2, figs 6-7.
2005. Cythereis mesa ventroreticulata honiGsTein. - shahin, p. 760, pl. 3, fig. 4.
Material : 10 specimens.Dimensions : L : 0.95 mm; H : 0.50 mm.Remarks : Cythereis? mesa ventroreticulata is distin-guished from Cythereis? mesa mesa by its reticulate lat-eral surface, whereas in C. mesa mesa the lateral surface is smooth. Occurrence : Late Santonian, Campanian and Paleocene of Israel (honiGsTein, 1984; honiGsTein et al., 1993; honiGsTein & rosenfeld, 1995) and Maastrichtian-Pa-leocene of Egypt (morsi, 1999, 2000; ismail & ied, 2004; shahin, 2005). Here, it is found in the Late Maas-trichtian-Early Paleocene (Micula prinsii Subzone - El-lipsolithus macellus Zone)
Genus Doricythereis Gründel, 1976Type species : Mauritsina jordanica bassiouni, 1970.
Doricythereis martinii (baSSiouni, 1970)Pl. III, figs 9-10, 14
1970. Mauritsina martinii bassiouni, p. 21, pl. 1, figs 1-2.1977. Mauritsina martinii bassiouni. - bassiouni et al., p. 5,
pl. 1, fig. 4a-b.1982. Mauritsina martinii bassiouni. - bouKhary et al., pl.
1, fig. 2. 1995. Mauritsina jordanica nodoreticulata bassiouni. - hon-
iGsTein & rosenfeld, p. 56, pl. 3, figs 1-2.1999. Mauritsina martinii bassiouni. - morsi, p. 40, pl. 2,
figs 7-9.2000. Megommatocythere sp. bassiouni & morsi, p. 59, pl. 8,
fig. 18.2002. Mauritsina jordanica nodoreticulata bassiouni. - hon-
iGsTein et al., p. 375, pl. 3, fig. 1.2004. Mauritsina jordanica nodoreticulata bassiouni. - is-
mail & ied, p. 105, pl. 2, figs 1-2.2005. Mauritsina jordanica nodoreticulata bassiouni. - is-
mail & ied, p. 134, pl. 3, figs 6-7.
Material : 266 specimens.Dimensions : L : 1.12-1.13 mm (females), 1.40-1.41 mm (males); H : 0.62 mm (females), 0.68 mm (males); W : 0.67 mm (female).Occurrence : Early Eocene of Jordan (bassiouni, 1970) and Paleocene to Early Eocene of Egypt (bassiouni et al., 1977; bouKhary et al., 1982; morsi 1999; is-
mail & ied, 2004, 2005) and Israel (honiGsTein & rosen-feld, 1995; honiGsTein et al., 2002). In the present work, it is recorded from the Paleocene-Early Eocene (Ellipsolithus macellus - Discoaster binodosus zones).
Genus Kefiella donze & Said, 1982Type species : Kefiella maresi donze & said, 1982.
Kefiella sp.Pl. III, figs 11-13
2004. Acanthocythereis meslei donze & oerTli. - ismail & ied, p. 107, pl. 2, fig. 8.
Material : 16 specimens.Dimensions : L : 0.78-0.79 mm (females), 0.93 mm (male); H : 0.48-0.49 mm (females), 0.53 mm (male); W : 0.44 mm (female), 0.39 mm (male). Description : Carapace medium-sized, subrectangular in lateral view. Anterior margin broadly rounded, posterior margin narrower with rounded lower part and a straight upper part. Dorsal margin straight, weakly inclined pos-teriorly; in left valve, it joins posterior margin at an ob-tuse cardinal angle and anteriorly builds a hinge ear at connection with anterior margin. Ventral margin almost straight, gently rising posteriorly. Maximum length me-dian, maximum height at eye tubercle, at about one fifth of length. Sculpture composed of reticulation, small spines and differently developed ribs. Reticulation mod-erately coarse, variable in shape, fining at anterior part of lateral surface, followed anteriorly by a row of coarse reticules along well-developed anterior marginal rib; in front of this marginal rib a similar row of reticules bor-ders anterior margin. Posterior margin occupied by a marginal rim; both end margins decorated with small spines. Ventral rib well-developed, occupied by seven small tubercles. Median rib thin, convex, begins behind subcentral tubercle, ends almost below posterodorsal cor-ner. Dorsal rib weak, carries a row of about six small spines. Left valve slightly larger than right valve; greatest overlap at anterodorsal and posterodorsal corners. Eye-spot small. Subcentral tubercle pronounced. Outline in dorsal view suboval with compressed ends; maximum width shortly behind middle. Hinge of right valve con-sists of a small anterior tooth, a smooth median groove and a strong smooth posterior tooth situated at the poste-rior dorsal angle. Marginal zone of moderate width. Inner margin and line of concrescence coincide. Selvage devel-oped. Internal features not observed. Sexual dimorphism not observed.Discussion : Kefiella maresi donze & said, 1982 (in donze et al., 1982), which was first described from the Maastrichtian of Tunisia, resembles the present species. However, it differs in displaying shorter longitudinal ven-tral and dorsal ribs, stronger muscle node and thicker re-ticulation muri. The reticules aligned at the anterior mar-gin in K. maresi are wider than high, whereas in the pres-

172 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
ent species they are higher than wide. Furthermore, K. maresi shows longitudinal striations ventrally which are not shown in Kefiella sp.Occurrence : This species was previously illustrated in the Paleocene rocks of northeast Sinai (ismail & ied, 2004). Here, it is recorded from the Early Paleocene to Early Eocene (Ellipsolithus macellus - Tribrachiatus contortus Zones).
Genus Ordoniya al-SheiKhly, 1985Type species : Hazelina ordoniya bassiouni, 1970.
Ordoniya ordoniya (baSSiouni, 1970)Pl. III, figs 15-16
1970. Hazelina ordoniya bassiouni, p. 31, pl. 4, figs 4-8.1982. Hazelina ordoniya bassiouni. - donze et al., p. 284, pl.
3, figs 2-3.1984. Hazelina ordoniya bassiouni. - Khalifa et al., pl. 1, fig. 7.1985. Ordoniya (Ordoniya) ordoniya (bassiouni). - al-sheiKh-
ly, p. 246, text-fig. 3; pl. 1, figs 1-11; pl. 2, figs 1, 3-4.1990. Ordoniya ordoniya (bassiouni). - bassiouni & luGer,
p. 832, pl. 16, figs 7-8, 10.1992. Hazelina ordoniya bassiouni. - ismail, p. 50, pl. 2, fig. 6. 1996. Ordoniya ordoniya (bassiouni). - andreu, p. 116, pl. 3,
figs 14-15.1999. Ordoniya ordoniya (bassiouni). - morsi, p. 41, pl. 3,
figs 1-5.2000. Ordoniya ordoniya (bassiouni). - morsi, p. 62, pl. 3,
fig. 7.2000. Ordoniya ordoniya (bassiouni). - bassiouni & morsi,
p. 35, pl. 9, figs 16-17.2003. Ordoniya ordoniya (bassiouni). - morsi & speijer, p.
75, pl. 4, fig. 38.2004. Ordoniya ordoniya (bassiouni). - ismail & ied, p. 109,
pl. 3, figs 9-10.2005. Ordoniya ordoniya (bassiouni). - shahin, p. 762, pl. 3,
fig. 21.
Material : 10 specimens.Dimensions : L : 0.65-0.68 mm; H : 0.33-0.39 mm. Occurrence : Early to “Middle” Paleocene of Jordan (bassiouni, 1970), Late Maastrichtian to Early Paleo-cene of Tunisia (donze et al., 1982), Paleocene of Mo-rocco (andreu, 1996) and Maastrichtian to Early Eo-cene of Egypt (Khalifa et al., 1984; bassiouni & luGer, 1990; ismail 1992; morsi, 1999, 2000; bassiou-ni & morsi, 2000; morsi & speijer, 2003; ismail & ied, 2004; shahin, 2005). In the present sections, it occurs in the Late Maastrichtian-Early Paleocene (Reinhardtites levis - Ellipsolithus macellus Zones).
Ordoniya bulaqensis baSSiouni & luGer, 1990Pl. III, figs 17-18
1990. Ordoniya bulaqensis bassiouni & luGer, p. 832, pl. 16, figs 5-9, 11-15.
1995. Ordoniya bulaqensis bassiouni & luGer. - honiG-
sTein & rosenfeld, p. 54, pl. 2, figs 1-2. 1996. Ordoniya bulaqensis bassiouni & luGer. - ismail, p.
45, pl. 3, figs 11-12.1999. Ordoniya bulaqensis bassiouni & luGer. - morsi, p.
42, pl. 3, figs 8-11.2000. Ordoniya bulaqensis bassiouni & luGer. - bassiou-
ni & morsi, p. 61, pl. 9, fig. 18. 2002. Ordoniya bulaqensis bassiouni & luGer. - ho-
niGsTein et al., p. 378, pl. 4, fig. 2.2003. Ordoniya bulaqensis bassiouni & luGer. - mor-
si & speijer, p.75, pl. 4, fig. 39.2005. Ordoniya ordoniya (bassiouni). - ismail & ied, p. 138,
pl. 3, figs 15-16.2005. Ordoniya bulaqensis bassiouni & luGer. - shahin, p.
762, pl. 3, fig. 17.
Material : 3 specimens.Dimensions : L : 0.61-0.71 mm; H : 0.33-0.40 mm.Occurrence : Paleocene to Early-“Middle” Eocene of Is-rael (honiGsTein & rosenfeld, 1995; honiGsTein et al., 2002) and Paleocene to Early Eocene of Egypt (bassiouni & luGer, 1990; ismail, 1996; morsi 1999; bassiouni & morsi, 2000; morsi & speijer, 2003; is-mail & ied, 2005; shahin, 2005). In the present study, it is found in the Early Paleocene (Markalius inversus - Chiasmolithus danicus Zones).
Ordoniya hasaensis (baSSiouni, 1970)Pl. III, figs 19-20
1970. Hazelina hasaensis bassiouni, p. 33, pl. 5, figs 5-6.1977. Hazelina hasaensis bassiouni. - bassiouni et al., 1977,
p. 3, pl. 1, fig. 12 a-c.1991. Ordoniya hasaensis (bassiouni). - honiGsTein et al., p.
104, pl. 2, fig. 5.1999. Ordoniya hasaensis (bassiouni). - morsi, p. 42, pl. 3,
fig. 13.2002. Ordoniya hasaensis (bassiouni). - honiGsTein et al., p.
378, pl. 4, fig. 3.2004. Ordoniya hasaensis (bassiouni). - ismail & ied, p. 109,
pl. 3, figs 5-6. 2005. Ordoniya hasaensis (bassiouni). - shahin, p. 762, pl. 3,
fig. 19.
Material : 8 specimens.Dimensions : L : 0.74 -0.85 mm; H : 0.40-0.53 mm.Occurrence : Late Paleocene of Jordan (bassiouni, 1970), Paleocene-Middle Eocene of Egypt (bassiouni et al., 1977; morsi 1999; ismail & ied, 2004; shahin, 2005) and Early-Middle Eocene of Israel (honiGsTein et al., 1991, 2002). In the present material, it is recorded in the Early Eocene (Discoaster binodosus Zone).
Ordoniya maanensis (baSSiouni, 1970)Pl. IV, figs 1-2
1970. Hazelina maanensis bassiouni, p. 33, pl. 5, figs 1-2.1977. Hazelina maanensis bassiouni. - bassiouni et al., p. 3,
pl. 1, fig. 13.

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 173
1995. Ordoniya maanensis (bassiouni). - honiGsTein & ro-senfeld, p. 56, pl. 2, fig. 4.
2000. Ordoniya burmaensis (bassiouni). - bassiouni & mor-si, p. 61, pl. 9, figs 19-20.
2000. Ordoniya maanensis (bassiouni). - bassiouni & mor-si, p. 61, pl. 10, figs 1-3.
2002. Ordoniya maanensis (bassiouni). - honiGsTein et al., p. 378, pl. 4, fig. 1.
2005. Ordoniya maanensis (bassiouni). - shahin, p. 762, pl. 3, fig. 20.
Material : 15 specimens.Dimensions : L : 0.79-0.83 mm; H : 0.43-0.45 mm. Occurrence : Early Eocene of Jordan (bassiouni, 1970), Paleocene and Early Eocene of Israel (honiG-sTein & rosenfeld, 1995; honiGsTein, et al., 2002) and Egypt (bassiouni et al., 1977; bassiouni & morsi, 2000; shahin, 2005) and. In the present area, it is re-corded from the Early Paleocene to Early Eocene (Cruci-placolithus tenuis - Tribrachiatus contortus zones).
Genus Paracosta Siddiqui, 1971Type species : Costa (Paracosta) decilvis siddiqui, 1971.
Paracosta parakefensis baSSiouni & luGer, 1990Pl. IV, figs 3-6
1990. Paracosta parakefensis bassiouni & luGer, p. 834, pl. 19, figs 10, 13-23.
1999. Paracosta parakefensis bassiouni & luGer. - morsi, p. 43, pl. 3, figs 14-15.
2000. Paracosta parakefensis bassiouni & luGer. - bassiou-ni & morsi, p. 36, pl. 10, figs 4-5.
2003. Paracosta parakefensis bassiouni & luGer. - mor-si & speijer, p. 76, pl. 4 figs 43-46, pl. 5, figs 47-49.
2004. Paracosta parakefensis bassiouni & luGer. - ismail & ied, p. 111, pl. 4, figs 4-5.
2005. Paracosta parakefensis bassiouni & luGer. - ismail & ied, p. 139, pl. 4, figs 1-4.
Material : 24 specimens.Dimensions : L : 0.70-081 mm (females), 0.89 mm (male); H : 0.41-0.43 mm (females), 0.46 mm (male); W : 0.34 mm (male).Occurrence : Paleocene-Early Eocene of Egypt (bassiouni & luGer, 1990; morsi, 1999; bassiou-ni & morsi, 2000; morsi & speijer, 2003; ismail & ied, 2004, 2005). In the present area, it occurs in the Early Paleocene-Early Eocene (Chiasmolithus danicus - Dis-coaster binodosus Zones).
Genus Phacorhabdotus howe & laurencich, 1958Type species : Phacorhabdotus texanus howe & lau-renCiCh, 1958.
Phacorhabdotus inaequicostatus colin & donze, 1982
Pl. IV, figs 7-8
1982. Phacorhabdotus inaequicostatus Colin & donze, in donze et al., p. 296, pl. 13, fig. 11.
1995. Phacorhabdotus inaequicostatus Colin & donze. - ho-niGsTein & rosenfeld, pl. 1, figs 9-11.
1999. Phacorhabdotus inaequicostatus Colin & donze. - morsi, p. 54, pl. 1, figs 9-11.
2004. Phacorhabdotus inaequicostatus Colin & donze. - is-mail & ied, p. 114, pl. 4, figs 1-2.
2005. Phacorhabdotus inaequicostata Colin & donze. - sha-hin, p. 764, pl. 4, figs 13-14.
Material : 6 specimens.Dimensions : L : 0.77 mm; H : 0.55 mm.Occurrence : Latest Campanian and Maastrichtian of Tunisia (donze et al., 1982), Paleocene of southern Is-rael (honiGsTein & rosenfeld, 1995) and Egypt (mor-si, 1999; ismail & ied, 2004; shahin, 2005). In the pres-ent study, it is recorded in the Early Paleocene (Crucipla-colithus tenuis - Fasciculithus tymbaniformis Zones).
Genus Reticulina baSSiouni, 1969bType species : Carinocythereis (Reticulina) heluanensis bassiouni, 1969b
Reticulina proteros baSSiouni, 1969bPl. IV, figs 9-12
1969b. Carinocythereis (Reticulina) scitula proteros bassiou-ni, p. 11, pl.1, fig. 8; pl. 2, figs 6-7.
1978. Reticulina scitula proteros bassiouni. - said, p. 261, pl. 29, figs 5-8.
1982. Reticulina proteros bassiouni. - donze et al., p. 287, pl. 5, figs 7-8.
1984. Reticulina proteros bassiouni. - Khalifa et al., pl. 2, fig. 8 a-b.
1987. Reticulina proteros bassiouni. - damoTTe & fleury, p. 95, pl. 2, figs 17-19.
1990. Reticulina proteros bassiouni. - bassiouni & luGer, p. 836, pl. 20, figs 16-21.
1991. Reticulina proteros bassiouni. - honiGsTein et al., p. 104, pl. 2, fig. 7.
1995. Reticulina proteros bassiouni. - honiGsTein & rosen-feld, p. 60, pl. 3, figs 5-6.
1998. Reticulina proteros bassiouni. - said-benzarTi, pl. 3, fig. 18.
1999. Reticulina proteros bassiouni. - morsi, p. 43, pl. 3, fig. 17.
2000. Reticulina proteros bassiouni. - bassiouni & morsi, p. 36, pl. 10, figs 6-8.
2000. Reticulina proteros bassiouni. - shahin, p. 303, pl. 7, fig. 2.
2002. Reticulina proteros bassiouni. - honiGsTein et al., p. 376, pl. 5, fig. 51.
2003. Reticulina proteros bassiouni. - morsi & speijer, p. 76, pl. 3, figs 7-8.
2005. Reticulina proteros bassiouni. - ismail & ied, p. 140, pl. 4, figs 5-6.
Material : 49 specimens.Dimensions : L : 0.87-0.88 (females), 0.95 mm (male);

174 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
H : 0.53 mm (female), 0.46 mm (male); H : 0.38 mm (male).Occurrence : Paleocene of east Algeria (damoTTe & fleury, 1987), Paleocene to Early Eocene of Jordan (bassiouni, 1969b) and Tunisia (said, 1978; donze et al., 1982; said-benzarTi, 1998), Paleocene and Middle Eocene of Israel (honiGsTein et al., 1991; honiG-sTein & rosenfeld, 1995; honiGsTein et al., 2002) and late Late Paleocene and Early Eocene of Egypt (bassiou-ni & luGer, 1990; morsi, 1999; shahin, 2000; morsi & speijer, 2003; ismail & ied, 2005; shahin, 2005). In the present area, it is recorded in the Early Pa-leocene-Early Eocene (Ellipsolithus macellus - Discoast-er binodosus Zones).
Subfamily Buntoniinae aPoStoleScu, 1961Genus Soudanella aPoStoleScu, 1961
Type species : Soudanella laciniosa laciniosa aposTo-lesCu, 1961.
Soudanella laciniosa triangulata aPoStoleScu, 1961Pl. IV, figs 13-14
1961. Soudanella laciniosa triangulata aposTolesCu, p. 810, p1. 7, figs 130-135.
1963. Soudanella cf. S. laciniosa triangulata aposToles-Cu. - barsoTTi, p. 1525, p1. 3, fig. 20.
?1966. Buntonia (Buntonia) virgulata aposTolesCu. - salahi, p. 10, p1. 2, fig. 10.
1968. Soudanella laciniosa triangulata aposTolesCu. - GréKoff, p. 17, p1. 3, fig. 44.
1970. Soudanella laciniosa triangulata aposTolesCu. - bassiouni, p1. 3, fig. 11 a-c.
non 1973. Soudanella laciniosa triangulata aposTolesCu. - neufVille, pl. 6. 11, fig. 1a-c.
1977. Soudanella laciniosa triangulata aposTolesCu. - bassiouni et al., p. 2, p1.3, figs 2-5.
1978. Soudanella cf. laciniosa triangulata aposTolesCu. - said, p. 227, pl. 25, figs 10-11.
1982. Soudanella laciniosa triangulata aposTolesCu. - donze et al., p. 295, p1. 12, figs 2-3.
1987. Soudanella laciniosa triangulata aposTolesCu. - oKosun, p. 56, p1. 6, figs 15, 17-19.
1990. Soudanella laciniosa triangulata aposTolesCu. - bassiouni & luGer, p. 847, p1. 24, fig. 18.
1992 Soudanella laciniosa triangulata aposTolesCu. - ismail, p. 51, p1. 2, fig. 10.
1995. Soudanella laciniosa triangulata aposTolesCu. - aref, p. 128, p1. 1, fig. 12.
1998. Soudanella laciniosa triangulata aposTolesCu. - said-benzarTi, pl. 2, fig. 3.
2000. Soudanella laciniosa triangulata aposTolesCu. - bassiouni & morsi, p. 67, pl. 12, fig. 6.
2004. Soudanella laciniosa triangulata aposTolesCu. - ismail & ied, p. 115, pl. 5, figs 14-15.
2005. Soudanella laciniosa triangulata aposTolesCu. - shahin, p. 765, pl. 4, fig. 15.
2005. Soudanella laciniosa triangulata aposTolesCu. - ismail & ied, p. 145, pl. 5, fig. 11.
Material : 6 specimens.Dimensions : L : 0.76-0.85 mm; H : 0.39-0.44 mm. Occurrence : This subspecies is widely known in from the Early Paleocene of Senegal, Late Paleocene of Nige-ria and Libya, and Late Paleocene to Early Eocene of Algeria, Tunisia, Jordan and Egypt. In the present work, it is recorded in the Early Eocene (Discoaster binodosus - Tribrachiatus orthostylus Zones).
Family Xestoleberididae SarS, 1928Genus Xestoleberis SarS, 1866
Type species : Cythere aurantica baird, 1838.
Xestoleberis tunisiensis eSKer, 1968Pl. IV, fig. 17
1968. Xestoleberis tunisiensis esKer, p. 332, pl. 2, figs 13-14; pl. 4, fig. 2.
1978. Xestoleberis tunisiensis esKer. - said, p. 263, pl. 29, figs 10-12.
1982. Xestoleberis tunisiensis esKer. - donze et al., p. 282, pl. 2, fig. 9.
1987. Xestoleberis tunisiensis esKer. - damoTTe & fleury, p. 94, pl. 1, figs 17-18.
1990. Xestoleberis tunisiensis esKer. - bassiouni & luGer, p. 847, pl. 25, figs 1-7.
1992. Xestoleberis tunisiensis esKer. - ismail, p. 51, pl. 2, fig. 8. 1996. Xestoleberis tunisiensis esKer. - bassiouni & luGer, p.
73, pl. 24, figs 14, 17.1998. Xestoleberis tunisiensis esKer. - said-benzarTi, pl. 3,
fig. 21.2000. Xestoleberis tunisiensis esKer. - bassiouni & morsi, p.
68, pl. 12, figs 11-14.2003. Xestoleberis tunisiensis esKer. - morsi & speijer, p. 78,
pl. 5, fig. 57.
Material : 4 specimens.Dimensions : L : 0.55 mm; H : 0.38 mm.Occurrence : Late Campanian to Paleocene of Tunisia (said, 1978; donze et al., 1982; said-benzarTi, 1998), Maastrichtian of Algeria (damoTTe & fleury, 1987), Maastrichtian to Early Eocene of Egypt (bassiou-ni & luGer, 1990; ismail, 1992; bassiouni & morsi, 2000; morsi & speijer, 2003; ismail & ied, 2005) and Eocene of Somalia (bassiouni & luGer, 1996). In the present work, it is recorded in the Early Eocene (Tribra-chiatus contortus - Discoaster binodosus Zones).
Xestoleberis sp.Pl. IV, figs 15-16, 18
Material : 6 specimens.Dimensions : L : 0.47 – 0.49 mm; H : 0.28-0.31 mm; W : 0.32 mm.Remarks : This species differs from Xestoleberis tu-nisiensis in having a more elongate lateral outline and a symmetrically convex dorsal margin making the position of greatest height at mid-length, whereas in X. tunisiensis

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 175
the dorsal margin is asymmetric and the maximum is be-hind the middle. In dorsal view, maximum width is cen-tral in the present species, but behind the middle in X. tunisiensis.Occurrence : Late Maastrichtian (Arkhangelskiella cym-biformis - Micula prinsii Subzones).
V. BIOSTRATIGRAPHY
Among the large number of publications available on the Late Cretaceous-Early Eocene ostracodes from south Tethyan areas, biostratigraphic information handling the Maastrichtian - Early Eocene south Tethyan ostracodes has increased substantially in the recent years through several works in Egypt (bouKhary et al., 1982; bassiou-ni & luGer; 1990; ismail, 1992, 1996; morsi, 1999; bassiouni & morsi, 2000; shahin, 2000; spei-jer & morsi, 2002; morsi & speijer, 2003; is-mail & ied, 2004, 2005; shahin, 2005) as well as in the other Middle East areas (e.g. esKer, 1968; bassiouni, 1969b, 1969c, 1970; donze et al., 1982; da-moTTe & fleury, 1987; honiGsTein & rosenfeld, 1995; said-benzarTi, 1998; honiGsTein et al., 1991, 2002). Studies carried out in the basins of West Africa such as aposTolesCu (1961), reymenT (1963, 1981), reymenT & reymenT (1959, 1980), Carbonnel et al. (1990), Colin et al. (1998) have been also considered as many ostracode taxa found there are also recorded in Egypt.
Stratigraphic distributionIn the present studied sections, several horizons yielded a well-diversified ostracode faunas. The distribution charts of the ostracode species recorded in the studied Maas-trichtian to Early Eocene sections (Tables 1, 2) reveal a stratigraphic differentiation with time. As observed from the assembled range chart (Table 3), which depends on species records in the studied sections, dated by calcare-ous nannoplankton, as well as the other records in and outside Egypt, the recorded taxa can be categorized into the following kinds:1- Taxa restricted to the Maastrichtian : This kind in-cludes the taxa which are found here only in the Maas-trichtian part of the sections and have never been record-ed in higher intervals anywhere else. These taxa are Cyth-erelloidea melleguensis, Apateloschizocythere fimbrata, Paraschizocythere hirsutonodosa and Aphrikanecythere phumatoides. They can be used as markers for the Maas-trichtian interval.2- Taxa crossing the K/P boundary : It incorporates the taxa which are found here in the Maastrichtian and cross into the Paleocene. This kind of taxa includes Cytherella cf. lagenalis, Bairdia ilaroensis, Bairdia aff. septentriona-lis, Abyssocypris? adunca, Pontocyprella recurva, Argil-loecia sp. bassiouni & luGer, Krithe echolsae, Krithe cf. solomoni, Parakrithe crolifa, Martinicythere bassiou-nii, Actinocythereis? coronata, Acanthocythereis meslei
meslei, Cythereis? mesa mesa, Cythereis? mesa ventrore-ticulata, Acanthocythereis? denticulata, Ordoniya ordo-nyia, Phacorhabdotus inaequicostatus and Xestoleberis tunisiensis.3- Paleocene taxa : This group includes the taxa, which are confined to the Paleocene. They are represented only by two taxa, namely Cytherelloidea attiyaensis and Cris-taeleberis reticulata. They can be used as markers for the Paleocene. Cytherelloidea attiyaensis is so far only known in the Early Paleocene (morsi, 1999) and record-ed here in the same interval (NP2-NP4 Zones). Cristaele-beris reticulata was previously recorded in the Paleocene of Egypt (bouKhary et al., 1982; Khalifa et al., 1984; morsi, 1999) and Jordan (bassiouni, 1970). In the pres-ent sections, it is recorded only from the Early Paleocene (NP2 Zone).4- Taxa crossing the P/E boundary : This group includes the taxa which first appear in the Paleocene and also extend higher into the Early Eocene. The taxa listed are Paracy-pris aff. jonesi, Doricythereis martinii, Acanthocythereis? posterotriangulata, Ordoniya bulaqensis, Ordoniya ha-saensis, Ordoniya maanensis, Paracosta parakefensis, Re-ticulina proteros and Soudanella laciniosa triangulata. Of these, Ordoniya hasaensis and Soudanella laciniosa trian-gulata are found in the present sections only in the Early Eocene.5- Early Eocene Taxa : This kind comprises the species, which first appear in the Early Eocene. They include Bythoceratina hamzai, which was recorded previously from the same interval in the Farafra area (Western Des-ert of Egypt), and Bairdia sp., which is so far known only in the present study area and its complete stratigraphic range is still unknown.
DiscussionFrom the fore-mentioned ostracode faunal distribution, the delineation of the K/P boundary in the present stud-ied sections by means of ostracodes is distinct. It is char-acterized by the disappearance of the typical Maastrich-tian taxa represented by Cytherelloidea melleguensis, Apateloschizocythere fimbrata, Paraschizocythere hir-sutonodosa and Aphrikanecythere phumatoides and in-troduction of new taxa first appearing in the Paleocene, represented by Cytherelloidea attiyaensis, Paracypris jonesi, Doricythereis martinii, Cristaeleberis reticulata, Acanthocythereis? posterotriangulata, Ordoniya bu-laqensis, Ordoniya hasaensis, Ordoniya maanensis, Paracosta parakefensis, Reticulina proteros and Sou-danella laciniosa triangulata. On the other side, the rec-ognition of the different stages of the Paleocene based on the recorded taxa is generally difficult since the taxa first appearing in the Paleocene either range throughout the entire Paleocene, or even range higher into the Eocene. The only exception concerns Cytherelloidea attiyaensis that is so far known only in the Early Paleocene and its occurrence may be used on this basis to indicate this in-terval. It is also worth noting that although Ordoniya ha-

176 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Form
ati
on
Nan
nofo
ssil
zon
es/s
ub
zon
es
Sam
ple
No.
Arg
illo
ecia
sp.
Bas
siouni
& L
uger
Cyt
her
ello
idea
mel
leguen
sis
Kri
the
cf.
solo
moni
Cyt
her
ella
cf.
lagen
ali
s
Bair
dia
aff.
sep
tentr
ionali
s
Cyt
her
ella
sin
aen
sis
Para
krit
he
croli
fa
Ord
oniy
a o
rdoniy
a
Mart
inic
yther
e bass
iounii
bass
iounii
Act
inocy
ther
eis?
coro
nata
Aca
nth
ocy
ther
eis?
den
ticu
lata
Para
schiz
ocy
ther
e hir
suto
nodosa
Kri
the
echols
ae
Cyt
her
eis?
mes
a m
esa
Ponto
cypre
lla r
ecurv
a
Cyt
her
eis?
mes
a v
ento
reti
cula
ta
Ord
oniy
a b
ula
qen
sis
Cri
stael
eber
is r
etic
ula
ta
Aca
nth
ocy
ther
eis?
post
erotr
iangula
ta
Phaco
rhabdotu
s in
aeq
uic
ost
ata
Cyt
her
ello
idea
att
iyaen
sis
Kef
iell
a s
p.
Dori
cyth
erei
s m
art
inii
Ord
oniy
a m
aanen
sis
Para
cost
a p
ara
kefe
nsi
s
Ret
iculi
na p
rote
ros
Bair
dia
ila
roen
sis
Bair
dia
sp.
Xes
tole
ber
is t
unis
iensi
s
Thebes N-71 R
N-68 R
N-67 R R
N-66 R R R R
N-65 R R R R R
N-64 R R R R R C R R R R
N-63 R R R R
N-62 R R R R R R R R
N-61 R R R
N-60 R R R R R
N-59 R
N-57 R R R R
Tar. N-55 R
N-53 R
N-51 R
N-50 R R
N-49 R R R
N-48 R R R R R R
N-46 R R R R
N-45 R R R R R R R
N-44 R R R R
N-43 R R R
N-42 R R R
N-41 R R R R R
N-40 R R R R R
N-39 R R R R R R R R R
N-38 R R R R
N-37 R R R C R
N-36 R R R
N-35 R
N-34 R R
N-32 R R
N-31 R R
N-30 R R R R R
N-29 R R R R R R R R
N-27 R R R R R R
N-25 R
N-23 R R R
N-21 R R R R
N-17 R R R R
N-16 R R
N-15 R R
N-14 R R
N-10 R
N-5 R
N-3 R
N-1 R
Table 1, Morsi et al.
NP3
NP2
NP1
NP
4
Age
Ear
ly E
oce
ne
Ypre
sian
NP11
Esn
a
NP
10
NP
9
Pal
eoce
ne
Than
.
NP7/8
M. m
uru
sN
. fr
equen
sM
. pri
nsi
i
Lat
e C
reta
ceous
Lat
e M
aast
rich
tian
Sudr
Dak
hla
NP5
Dan
ian
Sel
andia
n
a
b
a
b
Abundance
A (abundant): >25 specimens
C (common): 6-25 specimens
R (rare): 1-5 specimens
Than. Thantian
Tar. Tarawan Formation
b
a
Table 1: Stratigraphic distribution of ostracode taxa with number of specimens at each level in Gebel Nukhul section.

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 177
Fo
rm
ati
on
Na
nn
ofo
ssil
zo
nes/
sub
zo
nes
Sa
mp
le N
o.
Ord
oniy
a o
rdoniy
a
Bair
dia
aff
. se
pte
ntr
ionali
s
Kri
the e
chols
ae
Para
kri
the c
roli
fa
Apate
losc
hiz
ocyth
ere
fim
bra
ta
Cyth
ere
lla s
inaensi
s
Aphri
kanecyth
ere
phum
ato
ides
Xest
ole
beri
ssp
.
Acanth
ocyth
ere
is m
esl
ei
mesl
ei
Abyss
ocypri
s? a
dunca
Arg
illo
ecia
sp.
Bass
iouni
& L
uger
Mart
inic
yth
ere
bass
iounii
bass
iounii
Ponto
cypre
lla r
ecurv
a
Acti
nocyth
ere
is? c
oro
nata
Acanth
ocyth
ere
is? d
enti
cula
ta
Phacorh
abdotu
s in
aequic
ost
atu
s
Cyth
ere
lloid
ea a
ttiy
aensi
s
Acanth
ocyth
ere
is? p
ost
ero
tria
ngula
ta
Cyth
ere
is? m
esa
vento
reti
cula
ta
Ord
oniy
a m
aanensi
s
Kri
the
cf.
solo
moni
Para
cost
a p
ara
kefe
nsi
s
Reti
culi
na p
rote
ros
Kefi
ell
a s
p.
Dori
cyth
ere
is m
art
inii
Bair
dia
ila
roensi
s
Cyth
ere
lla
cf.
lagenali
s
Para
cypri
saff
.jo
nesi
NP12 WF-123
WF-121 R R
WF-119 R
WF-118 R
WF-117 C
WF-116 R
WF-115 R R
WF-114 R
WF-111 R R R
WF-109 R R R R R R
WF-108 R R R R R R R R R R
WF-107 R R R C R R
WF-106 C R R
WF-105 R R R
WF-104 R R R R
WF-102 R
WF-101 R
WF-100 R
Tar. WF-97 R R R
WF-96 R
NP6 WF-95 R R
WF-93 R R
WF-92 R
WF-91 R R R R
WF-90 R R C R R R
WF-89 R R R R
WF-87 R
WF-86 R R
NP3 WF-82 R R R R
WF-81 R R C R R R R R R R
WF-80 R C C R R R R R
WF-79 R C R R
WF-78 R
WF-76 R R R R
WF-75 R R R R R R R R
WF-74 R R R
WF-70 R R R
WF-68 R R R R R
WF-65 R R
WF-61 R R R R
WF-55 R R R
WF-48 R R R
WF-42 R R R R
WF-35 R
WF-30 R
WF-17 R R R R R R
WF-11 R R R R R
WF-05 R R R
WF-01 R R R R
Table 2, Morsi et al.
Sela
nd
ian
Ag
e
Earl
y E
ocen
e
Yp
resi
an
Pale
ocen
e
Th
an
eti
an
Thebes
Esn
a
NP
9
NP11
NP
10
NP7/8
Dak
hla
NP5
NP
4
NP2
NP1
Late
Cre
taceo
us
Late
Maast
rich
tian
Su
dr
CC24
CC
25
N. fr
equens
M. pri
nsi
i
Dan
ian
c
b
a
b
a
c
b
a
b
Abundance
A (abundant): >25 specimens
C (common): 6-25 specimens
R (rare): 1-5 specimens
Tar.: Tarawan Formation
a
Table 2: Stratigraphic distribution of ostracode taxa with number of specimens at each level in Wadi Feiran section.

178 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Ost
raco
de sp
ecie
sW
. Afr
ica
Mor
occo
Alg
eria
Tuni
siaLi
bya
Egyp
tIs
rael
Jord
an
12
34
56
78
Cyt
here
lla c
f. la
gena
lis
Maa
stric
ht.-E
. Eoc
ene
Cyt
here
lla si
naen
sis
M
aast
richt
.-E. E
ocen
e
C
ythe
rello
idea
atti
yaen
sis
Ea
rly P
aleo
cene
Cyt
here
lloid
ea m
elle
guen
sis
L.
Cam
pani
an-M
aast
richt
ian
M
aast
richt
ian
Baird
ia il
aroe
nsis
Maa
st.-P
aleo
cene
Maa
stric
ht.-E
. Eoc
ene
Baird
ia a
ff. se
pten
trio
nalis
M
aast
richt
.-Pal
eoce
nePa
leoc
ene
Ab
ysso
cypr
is?
adun
ca
Pa
leoc
ene
L. C
amp/
.-Pal
eoce
ne
Maa
stric
ht.-E
. Eoc
ene
Para
cypr
is a
ff. jo
nesi
Pa
leoc
ene
Pale
ocen
ePa
leoc
ene-
E. E
ocen
e
Po
ntoc
ypre
lla re
curv
a
M
aast
richt
. Pal
eoce
neM
aast
richt
. Pal
eoce
neM
aast
richt
ian
Maa
stric
ht.-E
. Eoc
ene
Argi
lloec
ia sp
. Bas
siou
ni &
Lug
er
Maa
stric
ht.-E
. Eoc
ene
Kri
the
echo
lsae
C
ampa
nian
-Pal
eoce
ne
Maa
stric
ht.-E
. Eoc
ene
Maa
stric
ht.-P
aleo
cene
K
rith
e cf
. sol
omon
i
Maa
stric
ht -
E. P
aleo
c.
Pa
rakr
ithe
crol
ifaLa
te P
aleo
cene
Maa
stric
ht.-M
. Eoc
ene
Pale
ocen
e-E.
Eoc
ene
M
artin
icyt
here
bas
siou
nii b
assi
ouni
i
C
ampa
nian
-E. P
aleo
c.C
ampa
nian
-E. P
aleo
cene
M
aast
richt
. L. P
aleo
cene
Pale
ocen
e
Actin
ocyt
here
is?
coro
nata
Maa
stric
ht.-M
. Pal
eoc.
L.C
ampa
nian
-L. P
aleo
cene
Maa
stric
htia
nM
aast
richt
.-E. E
ocen
ePa
leoc
ene-
E. E
ocen
ePa
leoc
.-E. E
oc.
Dor
icyt
here
is m
artin
ii
L. P
aleo
cene
-E. E
ocen
ePa
leoc
ene-
E. E
ocen
eEa
rly E
ocen
eAp
ater
losc
hizo
cyth
ere
fimbr
ata
M
aast
richt
ian
Para
schi
zocy
ther
e hi
rsut
onod
osa
M
.-L. M
aast
richt
ian
M
aast
richt
ian
Acan
thoc
ythe
reis
mes
lei m
esle
i
L.C
amp.
-Maa
stric
ht.
Maa
stric
ht. -
E. P
aleo
c.M
aast
richt
.E. P
aleo
cene
Aphr
ikan
ecyt
here
phu
mat
oide
s
M
aast
richt
ian
M. L
. Maa
stric
htia
n
Maa
stric
htia
n
C
rist
aele
beri
s ret
icul
ata
Pa
leoc
ene
M
. Pal
eoce
neC
ythe
reis
? m
esa
mes
a
Maa
stric
ht.-P
aleo
cene
Sant
onia
n-Pa
leoc
ene
C
ythe
reis
? m
esa
vent
rore
ticul
ata
M
aast
richt
.-E.P
aleo
cene
Sant
onia
n-Pa
leoc
ene
Ac
anth
ocyt
here
is?
dent
icul
ata
E.-M
. Pal
eoce
neL.
Maa
satri
cht.-
E.Pa
leoc
.
Maa
stric
ht.-E
. Eoc
ene
Acan
thoc
ythe
reis
? po
ster
otri
angu
lata
Pa
leoc
ene-
E. E
ocen
eM
. Pal
eoce
ne-E
.Eoc
ene
O
rdon
iya
ordo
niya
Pa
leoc
ene
L.
Maa
st.-E
.Pal
eoce
ne
Maa
stric
ht.-E
. Eoc
ene
E. P
aleo
cene
E.-M
.Pal
eoce
neO
rdon
iya
bula
qens
is
Pale
ocen
e-E.
Eoc
ene
M.-
L. P
aleo
cene
O
rdon
iya
hasa
ensi
s
Pale
ocen
e-M
. Eoc
ene
E.-M
. Eoc
ene
L.Pa
leoc
ene
Ord
oniy
a m
aane
nsis
Pa
leoc
ene-
E. E
ocen
ePa
leoc
ene-
E. E
ocen
eE.
Eoc
ene
Para
cost
a pa
rake
fens
is
Pale
ocen
e-E.
Eoc
ene
Retic
ulin
a pr
oter
os
Pa
leoc
ene
M. P
aleo
cene
-E. E
ocen
e
L. P
aleo
cene
-E. E
ocen
ePa
leoc
ene-
M.E
ocen
eE.
Pal
eoc.
-E. E
oc.
Phac
orha
bdot
us in
aequ
icos
tatu
s
L.C
amp.
-Maa
stric
ht.
Pa
leoc
ene
Pale
ocen
e
Soud
anel
la la
cini
osa
tria
ngul
ata
Early
Pal
eoce
ne
L.
Pal
eoce
ne-E
. Eoc
ene
Late
Pal
eoce
neL.
Pal
eoce
ne-E
. Eoc
ene
L.
Pal
eoc.
-E. E
oc.
Xest
oleb
eris
tuni
sien
sis
Maa
stric
htia
nL.
Cam
pani
an-P
aleo
cene
M
aast
richt
.-E. E
ocen
e
Tabl
e 3:
Geo
grap
hic
and
stra
tigra
phic
dis
tribu
tions
of t
he re
cord
ed o
stra
code
taxa
insi
de a
nd o
utsi
de E
gypt
; 1. r
eym
enT &
rey
men
T (1
959)
, rey
men
T (1
981)
, fo
sTer
et
al. (
1983
), o
Ko
sun
(198
7), C
ar
bo
nn
el e
t al.
(199
0), C
oli
n e
t al.
(199
8), 2
. an
dr
eu (1
996)
, 3. d
am
oTT
e & f
leu
ry (1
987)
, 4. e
sKer
(196
8),
said
(197
8),
do
nze
et a
l. (1
982)
, sa
id-b
enza
rTi,
(199
8), 5
. ba
rso
TTi (
1963
), sa
lah
i (19
66),
el-w
aer
(199
2), K
een
et a
l. (1
994)
, wh
aTle
y &
ar
ias (
1993
), 6.
ba
ssio
un
i et
al.
(197
7), C
ro
nin
& K
ha
lifa
(197
9), b
ou
Kh
ary
et a
l. (1
982)
, ba
ssio
un
i & l
uG
er (1
990)
, is
ma
il (1
992)
, ism
ail
(199
6), b
ass
iou
ni &
mo
rsi
(200
0),
mo
rsi
(19
99,
2000
), sh
ah
in (
2005
), a
bd
el s
ha
fy e
t al
. (2
002)
, m
or
si &
spe
ijer
(20
03),
ism
ail
& ie
d (
2004
, 20
05),
7. h
on
iGsT
ein
(19
84),
ho
niG
sTei
n &
ro
sen
feld
(199
5), h
on
iGsT
ein
et a
l. (1
991,
200
2), 8
. ba
ssio
un
i (19
69b,
c, 1
970)
.

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 179
saensis, Ordoniya maanensis, Reticulina proteros and Soudanella laciniosa triangulata generally range from the Paleocene to the Early Eocene, they have never been recorded in Egypt older than the Late Paleocene. The P/E boundary in the present sections is only roughly delin-eated being accompanied by the disappearance of Cris-taeleberis reticulata and Phacorhabdotus inaequicostata and first occurrence of Bairdia sp. and Bythoceratina sp.
VI. INTEGRATED PALEOECOLOGY AND PALEOBIOGEOGRAPHY
Ostracodes are mainly benthic crustaceans, which inhabit almost all types of aquatic environments, from fresh to ma-rine to hypersaline waters provided that oxygen and suffi-cient food are supplied. In such environments, these small organisms are plentiful, nevertheless species diversity, like most of the other invertebrates, are highest in marine con-ditions. They are characterized by their long fossil history, abundance, diversity, and control on their spatial distribu-tion by ecological parameters and lack of pelagic larvae. Therefore, they can be successfully used in paleoenviron-mental as well as paleogeographic interpretations. Among the several works carried out on the Maastrichtian-Early Eocene ostracodes of Egypt and the paleobiogeographi-cally related areas, those of barsoTTi (1963), esKer (1968), bassiouni (1969b, c, 1970), reymenT & rey-menT, (1981), donze et al. (1982), honiGsTein (1984), peypouqueT et al. (1986), damoTTe & fleury (1987), bassiouni & luGer (1990), Carbonnel et al. (1990), (ismail (1992), Keen et al. (1994), damoTTe (1995), andreu (1996), Colin et al. (1998), elewa et al. (1999), morsi (1999, 2000), bassiouni & morsi (2000), sha-hin & el nady (2001), abdel shafy et al. (2002), spei-jer & morsi (2002), morsi & speijer (2003), is-mail & ied (2004), elewa & morsi (2004) and shahin (2005) made paleoecological and paleobiogeographic contributions.
In the present study, diverse assemblages of ostracode faunas have been released from the Sudr, Dakhla and Esna Formations; rare faunas have been extracted from the Tarawan and Thebes Formations. They all cover the time interval extending from the Maastrichtian to the Early Eocene. The ostracode fauna recorded are domi-nated by taxa known for the marine environment. Most of them show a wide geographic distribution along the southern realm of the Tethys, from east Algeria in the west to Israel and Jordan in the east (Table 3). They in-clude species belonging to the genera Pontocyprella, Bythoceratina, Krithe, Aphrikanecythere, Ordoniya and Reticulina, which are known from the deeper parts of the shelf (outer shelf to upper bathyal). Species representing these genera are Pontocyprella recurva, Bythoceratina hamzai, Krithe echolsae, Ordoniya ordoniya, O. bulaqen-sis, O. maanensis and Reticulina proteros. They are found together with Actinocythereis? coronata, Acanthocy-
thereis? denticulata, Acanthocythereis? posterotrianglata and Paracosta parakefensis, which were interpreted by bassiouni & luGer (1990), based on foraminiferal evi-dence, to implicate an outer shelf setting. Of these spe-cies, Krithe echolsae, Reticulina proteros and probably Ordoniya bulaqensis can also be found even deeper in the upper bathyal domain, together with Acanthocythereies meslei which is regarded as a species characteristic for this latter depth range (donze et al., 1982). Among the recorded assemblages, Aphrikanecythere phumatoides and Phacorhabdotus inaequicostatus also belong to this group of taxa and have been recognized by the latter au-thors in the domain of the upper bathyal and the outer shelf. Cythereis? mesa seems to be a more localized spe-cies; it is so far only known from southern Israel and Si-nai Peninsula. Only three of the taxa recorded, namely Bairdia ilaroensis, Parakrithe crolifa and Soudanella la-ciniosa triangulata, are also found in the basins of West Africa, (Table 3). These taxa seem to have inhabited the shallow inner shelf as well as the deep outer shelf depths (bassiouni & luGer, 1990). Their northward migration to the southern entities of the Tethys was likely via the shallow Trans-Saharan seaway (barsoTTi, 1963; rey-menT & reymenT, 1980; bassiouni & luGer, 1990, Carbonnel et al., 1990; Keen et al., 1994; morsi, 1999; bassiouni & morsi, 2000; morsi & speijer, 2003), around the West African Coast (bassiouni & luGer, 1990), or both (Fig. 4). The recorded ostracode fauna other than those discussed above belong to the genera Cytherella, Cytherelloidea, Bairdia, (?)Abyssocypris, Martinicythere and Cristaeleberis. Cytherella and Bair-dia are generally known to occupy all depths of marine waters. The occurrence of Cytherella may indicate low oxygen levels (whaTley, 1991). Although Cytherelloi-dea is generally known for shallow, warm waters, species belonging to this genus have been recognized by bassiouni & luGer (1990) and morsi (1999) for the outer infraneritic depths as indicated by accompanying foraminifera. Abyssocypris is more predominant in the deeper marine conditions. Most of the known species of the genus Cristaleberis are assigned to the middle-outer shelf (bassiouni & luGer, 1990) or outer shelf-upper bathyal (donze et al., 1982).
From the fore-mentioned discussion, it is recognized that the ostracode faunas extracted from the present studied sections are largely dominated by the type of species wandering along southern realm of the Tethys, the type referred to as the “South Tethyan Type” by bassiou-ni & luGer (1990), damoTTe (1995), morsi (1999) and morsi & speijer (2003). Less dominant, species so far known to have a localized distribution also co-occur. From the analysis of the known and inferred ecology of the present ostracode taxa, both generic and specific, it is possible to assign them generally to the deeper realms of the marine environment (outer shelf - upper bathyal).

180 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
currence of Cytherelloidea attiyaensis that may be used to indicate the Early Paleocene interval. The Paleocene/Eocene boundary is accompanied by the disappearance of Cristaeleberis reticulata and Phacorhabdotus inae-quicostatus and first occurrence of Bairdia sp. and Bytho-ceratina hamzai. The observations about the paleobio-geographic distribution of the ostracodes throughout the studied interval show that the ostracode faunas extracted from the study sections are largely dominated by the type of species wandering along the southern realm of the Te-thys, termed by bassiouni & luGer (1990) as the “South Tethyan Type”. They can be generally assigned to the deeper realms of the marine environment (outer shelf - upper bathyal).
ACKNOWLEDGEMENTSThe authors would like to thank Prof. M.A. bouKhary (Ain Shams University, Cairo) and Prof. M.A. hewaidy (Al-Azhar University, Cairo) for critical review and valu-able comments. Much gratitude is due to Prof. j. Kuss (University of Bremen, Germany) for affording the facil-ity of SEM-photography of the ostracode material. A spe-cial appreciation is due to Dr. H. mai (University of Bre-men, Germany) for technical assistance.
REFERENCESabdel ashafy, e., n. ibrahim & i.m. ied (2002) - Ostracode
biostratigraphy and paleobiogeography of the Upper Cretaceous in the northern part of the Gulf of Suez, Egypt. Egyptian Journal of Paleontology, 2 : 157-198.
VII. CONCLUSION
The studied Maastrichtian-Early Eocene successions in the area of west-central Sinai are differentiated into five lithostratigraphic units, namely from older to younger the Sudr Formation (Middle to Late Maastrichtian), Dakhla Formation (Late Maastrichtian to early Late Paleocene), Tarawan Formation (Late Paleocene), Esna Formation (Late Paleocene to Early Eocene), and Thebes Formation (Early Eocene). From these, thirty eight ostracode spe-cies and subspecies have been recorded from the sections of Gebel Nukhul and Wadi Feiran. They have been taxo-nomically described. Biostratigraphically, they can be subdivided into taxa restricted to the Maastrichtian, taxa crossing the K/P boundary, taxa marking the Paleocene, taxa crossing the P/E boundary and Early Eocene marker taxa. The K/P boundary in the studied sections is charac-terized by the disappearance of the typical Maastrichtian taxa represented by Apateloschizocythere fimbrata, Aphrikanecythere phumatoides, Cytherelloidea melleg-uensis and Paraschizocythere hirsutonodosa, and intro-duction of new taxa that first appear in the Paleocene such as Cytherelloidea attiyaensis, Doricythereis martinii, Or-doniya bulaqensis, Ordoniya hasaensis, Reticulina prote-ros and Soudanella laciniosa triangulata. The subdivi-sion of the Paleocene into Danian, Selandian, and Thane-tian based on the recorded ostracodes is generally difficult herein since the taxa first appearing in the Paleocene ei-ther occur throughout the entire Paleocene, or even high-er into the Eocene. The only exception concerns the oc-
Fig. 4: Paleogeographic map of North Africa and the Middle East showing possible migration routes during the Late Paleocene - Early Eocene (modified after Keen et al. 1994); Trans-Saharan passage (solid arrow), West African passage (dashed arrow).

Maastrichtian-Early Eocene ostracodes from west-central Sinai, Egypt 181
al-sheiKhly, s.a.j. (1985) - The new genus Ordoniya (Ostra-coda) from the Paleogene of the Middle East. Procures of the 2nd Jordanian Geology Conference, Amman, 238-273.
andreu, B. (1996) - Ostracodes du Crétacé supérieur-Paléo-cène des rides Sud-Rifaines, Maroc septentrional. Revue de Micropaléontologie, 39 (2) : 93-123.
aposTolesCu, V. (1961) - Contribution à l’étude paléontologi-que (Ostracodes) et stratigraphique des bassins cretacés et tertiaires de 1’Afrique Occidentale. Revue de l’Insti-tut Français du Pétrole, 16 (7/8) : 779- 867.
aref, m. (1995) - Early Eocene Ostracoda from the Thebes Formation along the Red Sea coastal area, Egypt. Egyp-tian Journal of Geology, 39 (1) : 113-131.
aubry, m.-p. (1998) - Early Paleogene calcareous nannoplank-ton evolution : A tale of climatic amelioration. In : au-bry, m.-p., s.G. luCas & w.a. berGGren (eds). Late Paleocene-early Eocene climatic and biotic events in the marine and terrestrial records, Columbia University Press:, New York, 158-203.
awad, G.h. & m.G. Ghobrial (1965) - Zonal stratigraphy of the Kharga Oasis. Ministry of Industry, General Egyp-tian organization for geological research and Mining, Geological Survey, 34 : 1-77.
barsoTTi, G. (1963) - Paleocenic ostracods of Libya (Sirte Ba-sin) and their wide African distribution. Revue de l’Ins-titut Français du Pétrole, 18 (10/11) : 202-217.
bassiouni, m.a.a. (1969a) - Ostracoden aus dem Eozän von Ägypten. 2. Die Unterfamilien Hemicytherinae, Thae-rocytherinae und Campylocytherinae. Geologisches Jahrbuch, 88 : 203-234.
bassiouni, m.a.a. (1969b) - Einige Costa- und Carinocytbe-reis (Reticulina)- Arten aus dem Paleozän und Eozän von Jordanien (Ostracoda). Neues Jahrbuch für Geolo-gische und Paläontologische Abhandlungen, 134 (1) : 1-16.
bassiouni, m.a.a. (1969c) - Einige Buntonia- und Soudanella - Arten (Ostracoda, Crustacea) aus dem Eozän von Jor-danien. Paläontologische Zeitschrift, 43 : 205-214.
bassiouni, m.a.a. (1970) - Ostracoden (Mauritsininae und Trachyleberidinae) und ihre Bedeutung für die Biostra-tigraphie des Maastrichts und Alttertiärs von Jordanien. Beihefte zum Geologischen Jahrbuch, 106 : 5-52.
bassiouni, m.a.a., m.a. bouKhary & h.s. anan (1977) - Ostracodes from Gebel Gurnah, Nile Valley, Egypt. Proceedings of the Egyptian Academy of Science, 30 : 1-9.
bassiouni, m.a.a. & p. luGer (1990) - Maastrichtian to Early Eocene Ostracoda from southern Egypt. Paleontology, Paleoecology, Paleobiogeography and Biostratigraphy. Berliner geowissenschaftliche Abhandlungen (A), 120 : 755-928.
bassiouni, m.a.a. & p. luGer (1996) - Middle Eocene Ostra-coda from northern Somalia. Courier Forschungsinsti-tut Senckenberg, 129 : 1-139.
bassiouni, m.a.a. & a.m. morsi (2000) - Paleocene-lower Eocene ostracodes from El Quss Abu Said Plateau (Fa-rafra Oasis), Western Desert, Egypt. Palaeontographica (A), 257 : 27-84.
beadnell, h.j.l. (1905) - The Relation of the Eocene and Cre-taceous systems in the Esna-Aswan reach of the Nile Valley. Quarterly Journal of the Geological Society of London, 61 : 667-678.
bonnema, J.H. (1941) - Ostracoden aus der Kreide des Unter-grundes der nordostlichen Niederlande. Natuurhisto-risch Maandblad, 29 : 9-12; 30 : 1-6, 35 p.
bouKhary, m.a., C. GuerneT & h. mansour (1982) - Os-tracodes du Tertiaire inférieur de l’Egypte. Cahiers de Micropaléontologie 1 : 13-20.
Carbonnel, G., K. alzouma & m. diKouma (1990) - Les os-tracodes paléocènes du Niger : taxinomie-un témoigna-ge de l’existence éventuelle de la Mer Transsaharienne? Geobios, 23 : 671-697.
Carbonnel, G. & A. Johnson (1989) - Les ostracodes paléogè-nes du Togo : taxonomie, biostratigraphie, apports dans l’organisation et l’évolution du bassin. Geobios 22 (4) : 409-443.
Carbonnel, G. & C. monCiardini (1995) - Ostracodes paléo-gènes au débouché de la mer transsaharienne (Mali, Ni-ger). Documents du Laboratoire de Géologie de l’Uni-versité de Lyon, 135 : 1-123.
Carbonnel, G. & m. oyede (1991) - Les ostracodes du site d’Onigbolo (Paléocène, Benin). Leur place dans le bas-sin Togo-Nigeria. Annales de Paléontologie, 77 (2) : 65-106.
Cherif, o. h., i. a. al-rifaiy, f. i. al-afifi & o. h. orabi (1989) - Foraminiferal biostratigraphy and paleoecolo-gy of some Cenomanian-Turonian exposures in west-central Sinai, Egypt. Revue de Micropaléontologie, Pa-ris, 31, 243-262.
Colin, j.-p., y. Tambareau & a. KrasheninniKoV (1998) - Maastrichtian and Paleocene ostracode assemblages of Mali (Western Africa). Dela-Opera SAZU 4 razr., 34 (2) : 273-345.
Cronin, T.m. & h. Khalifa (1979) - Middle and Late Eocene Ostracoda from Gebel El Mereir, Nile Valley, Egypt. Micropaleontology, 25 : 397-411.
CrouCh, e.m., C. heilmann-Clausen, h. brinlhuis, h.e.G. morGans, K.m. roGers, h. eGGer & b. sChmiTz (2001) - Global dinoflagellate event associated with the late Paleocene thermal maximum. Geology, 29(4) : 315-318.
damoTTe, r. (1995) - The biostratigraphy and paleobiogeogra-phy of the Upper Cretaceous-basal Tertiary ostracods from North Africa, Mali and Congo. Cretaceous Re-search, 16 : 35-366.
damoTTe, r. & j.j. fleury (1987) - Ostracodes Maastrichtiens et Paléocènes du Djebel Dyr, près de Tebessa, (Algérie orientale). Géologie Méditerranéenne, 14 (2) : 87-107.
diGbehi, b.z., G. Carbonnel & p. dufaure (1994) - Contri-butions de l’analyse microfaunistique (ostracodes et fo-raminifères) à la reconstitution paléogéographique du bassin ‘onshore’ de côte d’Ivoire à la limite Paléocène/Eocène. Revue de Micropaléontologie, 77 (2) : 65-106.
donze, p., j.-p. Colin, r. damoTTe, h. oerTli, j.-p. peypou-queT & r. said (1982) - Les Ostracodes du Campanien Terminal à l’Éocène inférieur de la coupe de Kef, Tuni-sie Nord-Occidentale. Bulletin des Centres de Recher-che Exploration-Production Elf-Aquitaine, 6 : 273-355.
elewa, a.m. (2002) - Paleobiogeography of Maastrichtian to early Eocene ostracoda of North and West Africa and the Middle East. Micropaleontology, 48 (4) : 391-398.
elewa, a.m.T., m.a. bassiouni & p. luGer (1999) - Multi-variate data analysis as a tool for reconstructing pal-aeoenvironments : The Maastrichtian to early Eocene Ostracoda of southern Egypt.- Bulletin of Faculty of Sci-

182 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Abbreviations:L : length, H : height, W : width, RVC : right view carapace, LVC : left view carapace, DVC : dorsal view carapace.
Plate I
Fig. 1-3 : Cytherella cf. lagenalis marliere, 1958Gebel Nukhul section, 1, sample N-27, RH1, L 0.98 mm, LVC; 2, sample N-62, RH2, L 0.90 mm, RVC; Wadi Feiran section, 3, sample WF-108, RH3, W 0.38 mm, DVC.
Fig. 4-5 : Cytherella sinaensis morsi, 1999Wadi Feiran section, 4, sample WF-81, RH4, W 0.29 mm, DVC; Gebel Nukhul section, 5, sample N-62, RH5, L 0.88 mm, LVC.
Fig. 6-7 : Cytherelloidea attiyaensis morsi, 1999 Wadi Feiran section, sample WF-81, 7, RH6, L 0.61 mm, LVC; 8, RH7, L 0.57 mm, RVC.
Fig. 8-10 : Cytherelloidea melleguensis damoTTe & said, 1982 Gebel Nukhul section, sample N-29, 8, RH8, L 0.68 mm, LVC; 9, RH9, width 0.27 mm, DVC; 10, RH10, length 0.68 mm, RVC.
Fig. 11, 13-14 : Bairdia ilaroensis reymenT & reymenT Gebel Nukhul section, sample N-64, 11, RH11,W 0.45 mm, DVC; 13, RH13, L 0.95 mm, RVC; 14, RH14, L 0.98 mm, LVC.
Fig. 12 : Bairdia aff. septentrionalis bonnema, 1941Wadi Feiran section, sample WF-42, RH12, L 1.10 mm, RVC.
Fig. 15-16 : Bairdia sp.Gebel Nukhul section; sample N-64, Gebel Nukhul section; 15, RH15, L 1.00 mm, LVC; 16, RH16, L 0.95 mm, RVC.
Fig. 17 : Paracypris aff. jonesi bonnema, 1941 Wadi Feiran section, sample WF-108, RH17, L 1.10 mm, RVC.
Fig. 18 : Abyssocypris? adunca (esKer, 1968)Wadi Feiran section, sample WF-17, RH18, L 0.60 mm, LVC.
ence, Minia University, 12(2) : 1-20.ElEwa, a.M. & K. IshIzaKI (1994) - Ostracodes from Eocene
rocks of the El Sheikh Fadl - Ras Gharib stretch, the Eastern Desert, Egypt- Biostratigraphy and paleoenvi-ronments. Earth Science, 48 (2) : 143-157.
elewa, a.m.T. & a.m. morsi (2004) - Palaeobiotope analysis and palaeoenvironmental reconstruction of the Paleo-cene-Early Eocene ostrcodes from east-central Sinai, Egypt. In : beaudoin, a.b. & head, m.j. (eds). The Palynology and Micropaleontology of Boundaries, Geological Society of London, Special Publications, 230 : 293-308.
el-sweify, a.a.r. (1984) - Senonian and Paleocene Ostracoda (Cytherellidae, Cytheridae, Hemicytheridae and Tra-chyleberididae) from Dakhla Oasis (Edmonstone), Egypt. Unpublished M.Sc. Thesis, Ain Shams Univer-sity, Cairo, 118 p.
el-waer, a. (1992) - Tertiary and Upper Cretaceous ostracoda from NW offshore, Libya. Their Taxonomy, biostratig-raphy and correlation with adjacent areas. Petroleum Research Center Special Publications, Tripoli, 445 p.
esKer, G.l. (1968) - Danian ostracodes from Tunisia. Micro-paleontology, 14 : 319-333.
faris, m., s.n. ayyad, h.a. el nahass & K.a. al wosabi (2005) - Integrated planktonic foraminifera and calcare-ous nannofossil biostratigraphy of the Upper Creta-ceous-lower Eocene formations, west central Sinai,
Egypt. Proceedings of the 4th international Conference on the Geology of Africa, Assiut, Egypt, 2 : 769-791.
fosTer, C.a., f.m. swain & s.w. peTTers (1983) - late Pa-leocene ostracoda from Nigeria. Revista Española de Micropaleontologia, 15 : 103-166.
Ghorab, M.A. (1961) : Abnormal stratigraphic features in Ras Ghareb oil field. 3rd Arab Petroleum Congress, Alexan-dria, 2, 10 pp.
GinGeriCh, p. d. (2000) - Paleocene/Eocene boundary and con-tinental vertebrate faunas of Europe and North America. In : sChmiTz, b., b. sundquisT & f. p. andersson (Hrsg.). Early Paleogene warm climates and biosphere dynamics. GFF, Stockholm, 122 (1) : 57-59.
GréKoff, n. (1968) - Sur la valeur stratigraphique el les rela-tions paléogéographiques de quelques ostracodes du Crétacé, du Paléocène et de l’Éocène inferieur d’Algérie Orientale. Proceedings of the 3rd African Micropaleon-tological Colloquium, Cairo : 227-248.
harTmann, G. & h.s. puri (1974) - Summary of neontological and paleontological classification of Ostracoda. Mitteilun-gen Hamburg Zoologisches Museum Institut, 70 : 7-73.
honiGsTein, a. (1984) - Senonian ostracodes from Israel. Bul-letin of the Geological Survey of Israel, 78 : 1-48.
honiGsTein, a. (1985) - Cythereis mesa mesa (Ostracoda), a correct name for Cythereis mesa ventroleviata Honig-stein, 1984. Israel Journal of Earth-Sciences, 34 (4) : 240 p.

Plate I

184 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Plate II
Fig. 1 : Pontocyprella recurva esKer, 1958Wadi Feiran section, sample WF-89, RH19, L 1.00 mm, RVC.
Fig. 2-4 : Argilloecia sp. bassiouni & luGer, 1990Wadi Feiran section, 2, sample WF-42, RH20, L 0.49 mm, LVC; 3, sample WF-55, RH21, L 0.51 mm, RVC; 4, sample WF-17, RH22, W 0.20 mm, DVC.
Fig. 5 : Apateloschizocythere fimbrata bassiouni & luGer, 1990 Wadi Feiran section, sample WF-05, RH23, L 0.48 mm, RVC.
Fig. 6 : Paraschizocythere hirsutonodosa el sweify, 1984Gebel Nukhul section, sample N-66, RH24, L 0.51 mm, RVC.
Fig. 7 : Bythoceratina hamzai bassiouni & morsi, 2000Wadi Feiran section, sample WF-109, RH25, L 0.73 mm, RVC.
Fig. 8-10 : Krithe echolsae esKer, 1968Gebel Nukhul section, sample N-29, 8, RH26, W 0.33 mm, DVC; 9, RH27, L 0.73 mm, RVC; Wadi Feiran section, 10, sample WF-90, RH28, L 0.69 mm, LVC.
Fig. 11-12 : Krithe cf. solomoni honiGsTein, 1984Wadi Feiran section, sample WF-11, 11, RH29, L 0.57 mm, RVC; 12, RH30, L 0.57 mm, LVC.
Fig. 13-14 : Parakrithe crolifa bassiouni & luGer, 1990 Gebel Nukhul section, 13, sample N-62, RH31, L 0.52 mm, RVC; 14, sample N-64, RH32, L 0.63 mm, LVC.
Fig. 15-17 : Martinicythere bassiounii bassiounii honiGsTein & rosenfeld, 2002 Wadi Feiran section, sample WF-81, 15, RH33, W 0.29 mm, DVC; 16, RH34, L 0.58 mm, RVC; 17,
RH35, L 0.61 mm, LVC.Fig. 18-19 : Acanthocythereis? denticulata esKer, 1968
Gebel Nukhul section, sample N-62, 18, RH36, L 1.03 mm, RVC; 19, sample N-64, RH37, W 0.53 mm, DVC.
Fig. 20-21 : Acanthocythereis meslei meslei donze & oerTli, 1982Wadi Feiran section, 20, sample WF-55, RH38, L 0.95 mm, RVC; 21, sample WF-48, RH39, L 1.00 mm, LVC.
Fig. 22-23 : Acanthocythereis? posterotriangulata (morsi, 1999)Wadi Feiran section, sample WF-80, 22, RH40, male, L 0.69 mm, RVC; 23, RH41, female, L 0.60 mm, RVC.
honiGsTein, a., a. almoGi-labin & a. rosenfeld (1987) - Combined ostracod and planktonic foraminiferal biozo-nation of the Late Coniacian-Early Maastrichtian in Is-rael. Journal of Micropalaeontology, 6 (2) : 31-60.
honiGsTein, a., f. hirsCh, a. rosenfeld & a. flexer (1993) - Cythereis mesa. Methusalem or Lazarus? Zetteliana, Munich, 20 : 343-347.
honiGsTein, a. & a. rosenfeld (1995) - Paleocene ostracods from southern Israel. Revue de Micropaléontologie, 38 (1) : 49-62.
honiGsTein, a., a. rosenfeld & C. benjamini (1991) - Ostra-cods and foraminifera from the Early-Middle Eocene of Qeren Sartaba, Jordan Valley. Journal of Micropalae-ontology, 10 : 95-107.
honiGsTein, a., a. rosenfeld & C. benjamini (2002) - Eo-cene ostracode faunas from the Negev, southern Israel : Taxonomy, stratigraphy and paleobiogeography. Jour-nal of Micropalaeontology, 48 (4) : 365-389.
ismail, a.a. (1992) - Late Campanian to Early Eocene Ostra-coda from Esh El Mallaha area, Eastern Desert, Egypt. Revue de Micropaléontologie, 35 (1) : 39-52.
ismail, a.a. (1996) - Biostratigraphy and Paleoecology of
Maastrichtian - early Eocene ostracods of west - central Sinai, Egypt. Revue de Paléobiologie, 15 (l) : 37-54.
ismail, a.s.a. & i.b. ied (2004) - Paleontology, paleoecology, paleobiogeography of the Maastrichtian - Early Paleo-gene Ostracoda, northeast Sinai, Egypt. Egyptian Jour-nal of Paleontology, 4 : 95-125.
ismail, a.s.a. & i.b. ied (2005) - Maastrichtian - Lower Eo-cene ostracodes from Safaga area, Eastern Desert, Egypt. Egyptian Journal of Paleontology, 5 : 119-159.
issawi, b., m. el-hinnawi, l. el-KhawaGa, s. labib & n. anany (1981) - Contribution to the geology of Wadi Feiran area, Sinai, Egypt. Geological Survey of Egypt, internal report, Cairo, 43 pp.
Keen, m.C., s.s.j. al-sheiKhly, a. elsoGher & a.m. Gam-mudi (1994) - Tertiary ostracods of North Africa and the Middle East. In : simmons, M. D. (ed.). Micropaleon-tology and Hydrocarbon Exploration in the Middle East. Chapman & Hall, London : 371-388.
Kelly, d. C., T.j. bralower, j.C. zaChos, i. premoli sil-Va & e. Thomas (1996) - Rapid diversification of plank-tonic foraminifera in the tropical Pacific (ODP Site 865) during the late Paleocene thermal maximum. Geology,

Plate II

186 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Plate III
Fig. 1-3 : Actinocythereis? coronata (esKer, 1968)Gebel Nukhul section, 1, sample N-23, RH42, female, L 0.80 mm, RVC; 2, sample N-29, RH43, male, L 0.87 mm, RVC; Wadi Feiran section, 3, sample WF-119, RH44, male, L 0.92 mm, LVC
Fig. 4-5 : Aphrikanecythere phumatoides damoTTe & oerTli, 1982Wadi Feiran section, sample WF-11, 4, RH45, L 0.73 mm, LVC; 5, RH46, L 0.74 mm, LVC.
Fig. 6 : Cristaeleberis reticulata bassiouni, 1970Gebel Nukhul section, sample N-38, RH47, L 0.55 mm, RVC.
Fig. 7 : Cythereis? mesa mesa honiGsTein, 1984Gebel Nukhul section, sample N-39, RH48, L 1.00 mm, RVC.
Fig. 8 : Cythereis? mesa ventroreticulata honiGsTein, 1984Gebel Nukhul section, sample N-39, RH49, L 0.95 mm, LVC.
Fig. 9-10, 14 : Doricythereis martinii (bassiouni, 1970) Gebel Nukhul section, sample N-64, 9, RH50, female, L 1.12 mm, RVC; 10, RH51, female, W 0.67 mm,
DVC; 11, RH52, male, L 1.24 mm, RVC.Fig. 11-13 : Kefiella sp.
Wadi Feiran section, sample WF-108, 11, RH53, female, L 0.78 mm, RVC; 12, RH54, male, W 0.39 mm, DVC; 13, RH55, male, L 0.83 mm, RVC.
Fig. 15-16 : Ordoniya ordoniya (bassiouni, 1970)Wadi Feiran section, sample WF-75, females, 15, RH56, L 0.66 mm, LVC; 16, RH57, L 0.66 mm, RVC.
Fig. 17-18 : Ordoniya bulaqensis bassiouni & luGer, 1990Wadi Feiran section, sample WF-81, 17, RH58, L 0.61 mm, RVC; 18, RH59, L 0.70 mm, RVC.
Fig. 19-20 : Ordoniya hasaensis (bassiouni, 1970)Wadi Feiran section, sample WF-119, 19, RH60, male, L 0.81 mm, RVC Gebel Nukhul section, sample N-63, 20, RH61, female, L 0.85 mm, LVC.
24 (5) : 423-426.Khalifa, h., a.r. ei-younsy & m.a. el bouKhary (1984)
- The Cretaceous/ Paleocene boundary as defined by os-tracodes at the north western approach of Kharga Oasis, Western Desert, Egypt. Bulletin of the Faculty of Sci-ence, Assiut University, 1 : 159-173.
Khalifa. h. & T.m. Cronin (1979) - Ostracodes de l’Éocène Moyen de El Sheikh Fadl, Est de Beni Mazar, Hauts Egypte. Revue de Micropaléontologie, 22 : 172-185.
Marliére, R. (1958) - Ostracodes du Montien de Mons et ré-sultats de leur étude. Mémoires de la Sociéte Belge de Géologie, Paléontologie et Hydrologie, 8 (5) : 1-53.
morsi, a.m. (1999) - Paleocene to Early Eocene ostracodes from the area of east-central Sinai, Egypt. Revue de Paléobiologie, 18 (1) : 31-55.
morsi, a.m. (2000) - Senonian ostracodes from east-central Sinai, Egypt; Biostratigraphic and paleobiogeographic implications. Revue de Micropaléontologie, 43 (1-2) : 47-70.
morsi, a.m. & r.p. speijer (2003) - High-resolution Ostra-code Records of the Paleocene/ Eocene Transition in the South Eastern Desert of Egypt-Taxonomy, Biostratigra-phy, Paleoecology and Paleobiogeography. Senckenber-giana lethaea, 83 (1/2) : 61-93.
neufVille, E.M.H. (1973) - Upper Cretaceous - Paleogene os-tracoda from the South Atlantic. Special Publications of the Paleontological Institut, University of Uppsala, 1 : 205 p.
oKosun, e.a. (1987) - Ostracod biostratigraphy of the eastern Dahomey basin, Niger Delta and the Benue trough of
Nigeria. Bulletin of the Geological Survey of Nigeria, 41 : 1-151.
peypouqueT, j.p., f. GrousseT & p. mourGuiarT (1986) - Pa-leooceanography of the Mesogean Sea based on ostra-cods of the northern Tunisian continental shelf between the Late Cretaceous and Early Paleogene. Geologische Rundschau, 75 : 159-174.
reymenT, r.a. (1963) - Studies on Nigerian Upper Cretaceous and Lower Tertiary Ostracoda : II. Danian, Paleocene and Eocene Ostracoda. Stockholm Contribution in Ge-ology, 10 : 1-286.
reymenT, r.a. (1981) - The Ostracoda of the Kalambaina For-mation (Paleocene), northwestern Nigeria. Bulletin of the Paleontological Institut of the University of Uppsa-la, 9 : 51-65.
reymenT, r.a. & j.f. aranKi (1991) - On the Tertiary genus Soudanella Apostolescu (1961) (Ostracoda, Crustacea). Journal of Micropalaeontology, 10 (1) : 23-28.
reymenT, r.a. & e.r. reymenT (1959) - Bairdia ilaroensis sp. nov. aus dem Paleozän Nigeriens und die Gültigkeit der Gattung Bairdopillata (Ostr. Crust.). Stockholm Contribution in Geology, 3 : 59-70.
reymenT, r.a. & e.r. reymenT (1980) - The Paleocene Trans-Saharan transgression and its ostracod fauna. In : salem, m. j. & busreVil, m. i. (eds.). The Geology of Libya. Academic Press, London, 1 : 245-254.
said, r. (1961) - Tectonic framework of Egypt and its influence on distribution of foraminifera. Bulletin of the American Association of Petroleum Geologists, 45 : 198-218.
said, r. (1978) - Etude stratigraphique et micropaléontologi-

Plate III

188 A. M. MORSI, M. FARIS, A. ZALAT & R. F. M. SALEM
Plate IV
Fig. 1-2 : Ordoniya maanensis (bassiouni, 1970)Gebel Nukhul section, 1, sample N-62, RH62, female, L 0.79 mm, RVC; 2, sample N-60, RH63, male, L 0.83 mm, RVC.
Fig. 3-6 : Paracosta parakefensis bassiouni & luGer, 1990Wadi Feiran section, sample WF-106, 3, RH64, female, L 0.77 mm, LVC; 4, RH65, female, L 0.70 mm, RVC; 5, RH66, male, W 0.34 mm, DVC; 6, RH67, male, L 0.89 mm, RVC.
Fig. 7-8 : Phacorhabdotus inaequicostatus Colin & donze, 1982Gebel Nukhul section, sample N-39, 7, RH68, juvenile, L 0.54 mm, RVC; 9, sample N-48, RH69, L 0.77 mm, LVC.
Fig. 9-12 : Reticulina proteros bassiouni, 1969bGebel Nukhul section, sample N-63, 9, RH70, female, L 0.88 mm, LVC; sample N-64, 10, 11, males, 10, RH71, W 0.38 mm, DVC, 11, RH72, L 0.93 mm, RVC; sample N-61, 12, RH73, female, L 0.91 mm, RVC.
Fig. 13-14 : Soudanella laciniosa triangulata aposTolesCu, 1961Wadi Feiran section, sample WF-118, 13, RH74, L 0.85 mm, RVC; 14, RH75, L 0.76 mm, RVC.
Fig. 17 : Xestoleberis tunisiensis esKer, 1968Gebel Nukhul section, sample N-66, RH76, L 0.55 mm, RVC.
Fig. 15-16, 18 : Xestoleberis sp.Wadi Feiran section, sample WF-55, 15, RH77, W 0.32 mm, DVC; sample WF-75, 17, RH78, L 0.49 mm, RVC; sample WF-17, 18, RH79, L 0.48 mm, LVC.
que du passage Crétacé-Tertiaire du Synclinal d’Elles (Région Siliana-Sers) Tunisie centrale. Thèse de Docto-rat 3ème Cycle, Université Paris, 5 : 275 p.
said-benzarTi, R. (1998) - Les ostracodes du Campanien Su-périeur à l’Yprésien de la coupe d’Elles (Tunisie du Centre Nord) - Biostratigraphie, paléoécologie et paéo-biogéographie. Bulletin des Centres de Recherche Ex-ploration-Production, Mémoire 20 : 197-211.
salahi, d. (1966) - Ostracodes du Crétacé supérieur et du Ter-tiaire en provenance d’un sondage de la région de Zeiten (Libye). Revue de l’Institut Français du Pétrole, 21 (1) : 3-43.
shahin, a. (2000) - Tertiary ostracods of Gebel Withr, south-western Sinai, Egypt : palaeontology, biostratigraphy and palaeobiography. Journal of African Earth Scienc-es, 31 (2) : 285-316.
shahin, a. (2005) - Maastrichtian to Middle Eocene ostracodes from Sinai, Egypt : Systematics, biostratigraphy and pa-leobiogeography. Revue de Paléobiologie, 24 (2) : 749-779.
shahin, a. & h. el nady (2001) - Late Cretaceous - Early Ter-tiary ostracodes from northeastern Sinai, Egypt : Bio-stratigraphy and Palaeobiogeography. Egyptian Journal Paleontolology, 1 : 149-191
speijer, r.p. & a.m. morsi (2002) - Ostracode turnover and sea-level changes associated with the Paleocene-Eocene thermal maximum. Geology, 30 (1) : 23-26.
Thomas, E. (1998) - Biogeography of the late Paleocene ben-thic foraminiferal extinction. In : M. - p. aubry, s. G. luCas & w. a. berGGren (eds). Late Paleocene-Early Eocene Climatic and Biotic Events in the Marine and Terrestrial Records. Columbia University Press, New York, 214-243.
Tjalsma, r. C. & G.p. lohmann (1983) - paleocene-Eocene bathyal and abyssal benthic foraminifera from the At-lantic Ocean. Micropaleontology, Special Publication, 4 : 90 pp.
whaTley, r.C. (1991) - The platycopid signal : a means of detecting kenoxic events using Ostracoda. Journal of Micropalaeontology, 10 : 181-183.
whaTley, r.C. & C.f. arias (1993) - Paleogene ostracoda from the Tripoli Basin, Libya. Revista Española de Mi-cropaleontologia, 15 (2) : 125-154.
Accepté juin 2008

Plate IV





















