
Paleoecology and Biostratigraphy of Marginal Marine Gulf Coast Eocene Vertebrate Localities
Upload
unisouthafrCategory
view
0download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Shellfish gathering, marine paleoecology and modern human behavior:perspectives from cave PP13B, Pinnacle Point, South Africaq
Antonieta Jerardino a,*, Curtis W. Marean b
aCatalan Institution for Research and Advanced Studies (ICREA)/Department of Prehistory, Ancient History and Archaeology, University of Barcelona, c/Montalegre, 6-8,E-08001 Barcelona, Spainb Institute of Human Origins, School of Human Evolution and Social Change, PO Box 872402, Arizona State University, Tempe, AZ 85287-2402, USA
a r t i c l e i n f o
Article history:Received 31 August 2008Accepted 22 December 2008
Keywords:Middle Stone AgeMossel BayShell middenMollusks
a b s t r a c t
Systematic collection of shellfish has been increasingly recognized as an important component of humanadaptation to aquatic environments and as part of the archaeological evidence found in association withthe appearance of early Homo sapiens. Over the last forty years, South Africa has played a prominent rolein recording the earliest evidence of shellfish in and substantial expansion of the early human diets asshown by several Middle Stone Age (MSA) coastal sites along the west and south coasts. In this paper, wereport on the abundance of marine invertebrate species from PP13B cave and interpret these abundancesin terms of paleoenvironmental changes, the likely shellfish procurement behaviors involved in bothrocky and sandy shore contexts, and the significance of the collection of marine shells for purposes otherthan food collection. Possible cognitive implications of shellfish gathering as a reflection of modernbehavior are also suggested.
� 2010 Elsevier Ltd. All rights reserved.
Introduction
Shellfish collecting has become increasingly recognized as animportant component of human adaptation to aquatic environ-ments and for its dramatic role on other human developments,such as higher population densities, greater sedentism, and long-distance migrations by people (Erlandson, 2001). Pinnacle PointSite (PP) 13B has provided the earliest known evidence for shellfishcollection at w164 ka and shows that a coastal adaptation waspresent in South Africa long before the postulated out of Africamigration w70e60 ka (Marean et al., 2007). In that paper, weprovided taxonomic abundance of the shellfish assemblages fromonly one part of the site (the Northeastern excavations into theLightly Cemented Middle Stone Age [LC-MSA]), and most of thatsample dates to Marine Isotope Stage 6 (MIS 6). In a prior publi-cation (Marean et al., 2004), we presented taxonomic abundancefrom the test excavations conducted in 2000 at sites PP9, PP13A,and PP13B from occupations that were at that time undated. Herewe extend that analysis to all the occupations at PP13B, coveringa span of time nowwell constrained betweenw170 and 90 ka, from
excavations undertaken between 2000 and 2006. Our sample isnow large enough and sufficiently well dated for us to documentshifts in the way Middle Stone Age (MSA) people changed theirshellfish collection habits and link those to broader changes in theirterrestrial and marine environment.
From early on, South Africa has played a prominent role in therecord for the earliest expansions of thehumandiet into shellfish. Thefirst evidence for systematic shellfish collection byMSA people alongthe south coast of SouthAfricawas initiallymade at Klasies River (KR)during excavations between 1967 and 1968 as reflected by the largequantities, density, and presence of marine shell throughout thestratigraphic sequence of this site (Speed, 1969; Voigt, 1973). Thisdiscovery was confirmed soon afterwards by observations from thewest coast at Sea Harvest and Hoedjiespunt (Volman, 1978; Klein,1979), and together these sites documented shellfish collection atleast by the early stages of MIS 5 (w123e74 ka). The analysis of MSAshellfish remains has evolved steadily since the early methodicalwork by Liz Voigt on the KR shellfish assemblage (Voigt, 1973, 1982).Right at the onset of these initial observations, it was recognized thatpeoplehadutilized shellfish as a source of food formany thousandsofyears, although the time depth of this subsistence practice increasedwith improved dating methods (Deacon and Geleijnse, 1988; Jacobset al., 2006; Marean et al., 2007).
Changes in sea surface temperatures, foraging range within tidalzones, availability of substrate type where shellfish thrived, dietarypreferences, andshellfishcollectingactivitieswere inferredasa result
q This article is part of ‘The Middle Stone Age at Pinnacle Point Site 13B, a CoastalCave near Mossel Bay (Western Cape Province, South Africa)’ Special Issue.* Corresponding author.
E-mail addresses: [email protected], [email protected] (A. Jerardino).
Contents lists available at ScienceDirect
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
0047-2484/$ e see front matter � 2010 Elsevier Ltd. All rights reserved.doi:10.1016/j.jhevol.2010.07.003
Journal of Human Evolution 59 (2010) 412e424

Author's personal copy
of quantitative analyses involving several mollusk species (Voigt,1973, 1982). Klein (1979) first noticed that Patella granatina (laterdescribed as Cymbula granatina) from the MSA site of Sea Harvestwere significantly larger on average than those from Later Stone Age(LSA) contexts along the same coastline, suggesting to Klein that SeaHarvest occupants engaged in less intensive shellfish collecting aspart of an overall less effective utilization of coastal resources byMSApeople. Thisobservationwasmentioned inpassingbyVolman(1978),and soonafterVoigt (1982)alsonoticed that somespeciesofmollusks(Cymbula oculus then described as Patella oculus, and Turbo sarmati-cus) fromMSA layers at KR were also larger than those of LSA origin,citing a combination of animal size range availability and humanpreference behind this difference. Based on additional samplesrecovered fromfurtherexcavationsatKR, Thackeray (1988)expandedon certain aspects of Voigt’s study by emphasizing species frequen-cies and shell density as proxies for environmental changes, such asshifts in the geomorphology of the coastline (rocky shores versussandy beaches) and fluctuations in cave/shore distance as a result ofsea level change. Thackeray (1988) also noticed that Turbo sarmaticusopercula from MSA levels were considerably larger than those fromLSA horizons, but did not elaborate any further on this observation.Klein (1989: 332) linked the larger MSA shells explicitly todemography by arguing that LSA people were more numerous thanMSA groups leading to the intense collection of shellfish anddepression of shellfish body size (see below).
With an increasing interest in MSA studies in the early 1990s,other coastalMSAsiteswere excavated systematically. Henshilwoodet al. (2001) at Blombos Cave followed the lead of prior investiga-tions, but also added other angles of interpretation. Based on westcoast LSA observations (Buchanan, 1988), higher frequencies ofPerna perna (brown mussels) were explained in terms of differentcollecting strategies, where these rocky shore mussels were pref-erentially collected for transport over longer distances as thesespecies remain fresh longer. Thus, higher abundances of brownmussels were taken as indicative of longer distances to the shoreand, hence, lower sea levels. Henshilwood et al. (2001) also inter-preted shifting ratios of shell versus opercula frequencies of Turbosarmaticusasevidence forprocessingof shellfishat collectionpoints,where heavy shells were removed and separated from the fleshbefore transporting the catch back to the cave. They alsomade a casefor the sizesof limpet shells and Turbo sarmaticusopercula asproxiesfor lower human population densities during MSA times.
From there on, the argument for lower MSA population densi-ties dominates the literature on MSA shellfish analysis (Halkettet al., 2003; Parkington, 2003; Klein et al., 2004; Steele and Klein,2005; Avery et al., 2008). Both, Klein et al. (2004) and Parkington(2003) also pointed out that MSA shell middens showed a lowernumber of marine invertebrate species when compared to their LSAcounterparts, suggesting that MSA people exploited marineresources more selectively or less intensively, as already indicatedby Klein (1979), stressing that this difference is perhaps alsoa reflection of a diminished capacity among MSA groups to exploitmarine resources to the extent that LSA people did. These wereperhaps some of the first references to cognitive capacities of MSApeople in the context of MSAmarine shell studies in South Africa. Afew years earlier, though, McBrearty and Brooks (2000) andErlandson (2001) reflected on shellfishing in the context ofarchaeological evidence formodern behavior in earlyHomo sapiens.Although limited shellfishing was practiced by other homininspecies in East Africa, Europe, and Asia that were broadlycontemporaries of MSA people in southern Africa (Erlandson, 2001;Stringer et al., 2008a; but see also Klein and Steele, 2008; Stringeret al., 2008b), more systematic shellfish collecting by MSA H.sapiens leading to the accumulation of large and relatively denseshell middens appears to have been part of a more broad based
economic strategy that supported population growth and demo-graphic expansion of this species into the rest of the world. Also,and in our opinion, the cognitive implications of shellfish gatheringas a possible reflection of modern behavior, if any, have not yet beenunpacked and understood beyond its timing in human evolution(McBrearty and Brooks, 2000).
With few exceptions (Stiner, 1993, 1994), discussions onmodernor nonmodern human behavior in the context of hominin subsis-tence activities have largely been dominated by the analysis of largevertebrate fauna, mostly from terrestrial environments (Mareanand Assefa, 1999). Despite the fact that marine shells dominatemuch of the sediment volume at several MSA sites in South Africa,marine shells have remained largely relegated to providing whatwe consider background observations relative to the largemammalfauna and stone artifacts (Erlandson, 2001), especially in thecontext of deriving inferences of MSA capabilities and modernbehavior. Thus, it is now time for shellfish gathering and theanalysis of marine invertebrates in theMSA to be launched to a newlevel of analytical and conceptual sophistication.
Shellfish gathering can be an important ingredient in overalladaptation, and, thus, we need to integrate it more fully into ourdiscussions of MSA adaptations. Only then will we be able toevaluate its relevance for understanding the development of“modern human behavior.” To begin this, we did the following.First, in the field we plotted in 3D to a total station all observedshellfish remains, thus treating the shellfish in the same way as thelithic artifacts and mammalian fauna. Ultimately this will allowdetailed vertical and horizontal spatial control and analyses; theusefulness of that approach has already been shown in Mareanet al. (2007). We then examined all shell fragments from theplotted and screened material (see below), compiled lists of speciesabundance for PP13B (Fig.1), and interpreted these changes relativeto paleoenvironmental and sea level changes. Attention is also paidto the task of procuring shellfish resources along the south coast ofSouth Africa where the diverse communities are found in bothrocky and sandy habitats. We also examine the collection of marineshells for purposes other than food collection. Metrical andtaphonomical observations on shell sizes will be incorporated ina further publication once the entire assemblage is analyzed. Whenthis analysis is completed, PP13B shellfish remains are likely to bethe most heavily analyzed in a southern African MSA site.
Analytical methods
Marine shell remains were recovered mostly as plotted finds(plotted in 3D to a total station) and asmaterial retained in a stackedset of 1.5mm, 3mm, and 10mmsieves (seeMarean et al., 2010). Thisanalysis does not include the material from the 1.5 mm sieves, asonly very tiny fragments were found in that sieve size. Marine shellremains were identified wherever possible to a generic or specificlevel (Kilburn and Rippey, 1982; Ridgway et al., 1998; Branch et al.,2007), whereupon minimum number of individuals (MNI) andweightswere established. For bivalves, the highest number of eitherleft or right hinges was used to determine MNIs. For limpets, andmost whelks and winkles (Oxystele spp., Burnupena spp., Nucellasquamosa), the number of apices was quantified (Jerardino, 1997);however, numbers of operculawere used for establishing counts forTurbo sarmaticus as these preserve better than the shell of thisspecies. In a few instances, MNI ¼ 1 was established when nocountable shell parts were encountered but taxonomically identi-fiable fragments were found. Each shell fragment, whether count-able or not, was weighed separately with a precision scale.
Maximum dimensions of plotted shell (whole or fragmented)were measured with a caliper in order to control for differentialfragmentation across space and between stratigraphic horizons.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424 413
Author's personal copy
Preservational status of marine shell was further established viamore qualitative evaluation of the degree of chalkiness or burningand through a subjective assessment of shell fragmentationexpressed as a percentage of the original whole shell. Taphonomicinferences derived from these observations are not presented here,but will be included in a separate publicationwith updated data. Allthe above analytical data were entered into a spreadsheet. About65% of the excavatedmarine shell from PP13B has been analyzed sofar, and we targeted our analysis presented here to fully sample therange of stratigraphic aggregates. Based on qualitative assessments,however, we do not believe that the species abundances presentedhere will change substantially with the addition of more material.
The excavations at PP13B were conducted within small strati-graphic lenses and features (StratUnits) that were then groupedinto larger stratigraphic aggregates (what many archaeologists maycall layers, see Marean et al., 2010). In our analysis some of theseaggregates are grouped into larger analytical units based on similarartifactual and geological composition and time. Table 1 providesa listing of the full set of stratigraphic aggregates and our assign-ment to analytical units.
Results
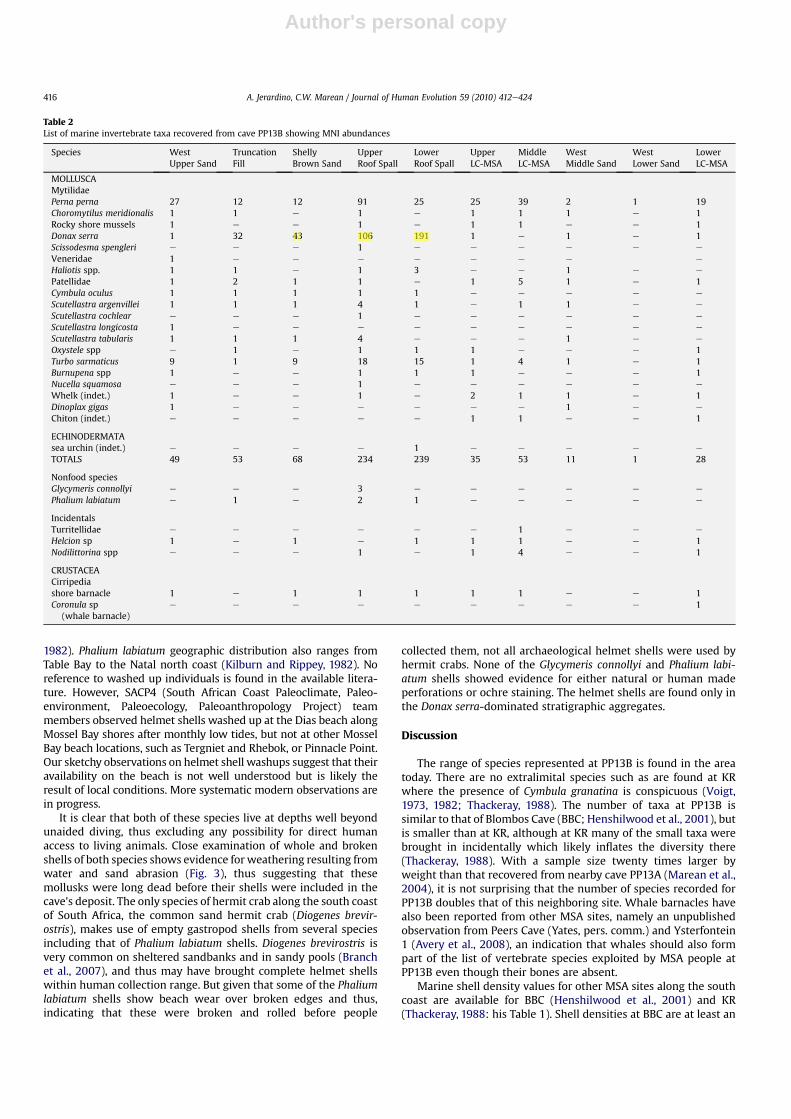
A total of at least 28 categories of marine invertebrates wereidentified (Table 2 and 3): 14 mollusks to species level (Perna perna,Choromytilus meridionalis, Donax serra, Scissodesma spengleri,
Cymbula oculus, Scutellastra argenvillei, Scutellastra cochlear, Scu-tellastra longicosta, Scutellastra tabularis, Turbo sarmaticus, Nucellasquamosa, Dinoplax gigas, Glycymeris connollyi, and Phalium labi-atum), another six mollusks to genus level (Haliotis spp., Oxystelespp., Helcion sp., Nodilittorina spp., Burnupena spp., and Coronulasp.), four other mollusks to Family level (Mytilidae, Patellidae,Turritellidae, and Veneridae), and one to Subphylum Crustacea andanother to Subphylum Echinodermata (sea urchin), as well as twoothers to a broad category of mollusks (winkles, whelks, andchitons) to which, respectively, many Families and an entire Classbelong. Chitons are represented by a few plates of the large-bodiedDinoplax gigas. Nevertheless, the majority of chiton remains atPP13B belong to a much smaller, as yet unidentified species.Specimens identified as rocky shore mussels were the fragmentarydiagnostic remains of badly preserved Perna perna or Choromytilusmeridionalis. Shell remains classified as Patellidae are limpetremains that could belong to either the Scutellastra or Cymbulagenera.
Nonfood species are also listed in Tables 2 and 3 and theirrelevance in the context of MSA behavior is discussed below. Othernondietary taxa such as a few tiny shells were incorporated into theshell assemblage, probably due to the fact that some are epibionts,which are species that cling to others such as shore barnacles, andothers often are observed attached to the byssus or threads of rockyshore mussels. These later taxa were, therefore, classified as“incidentals.”
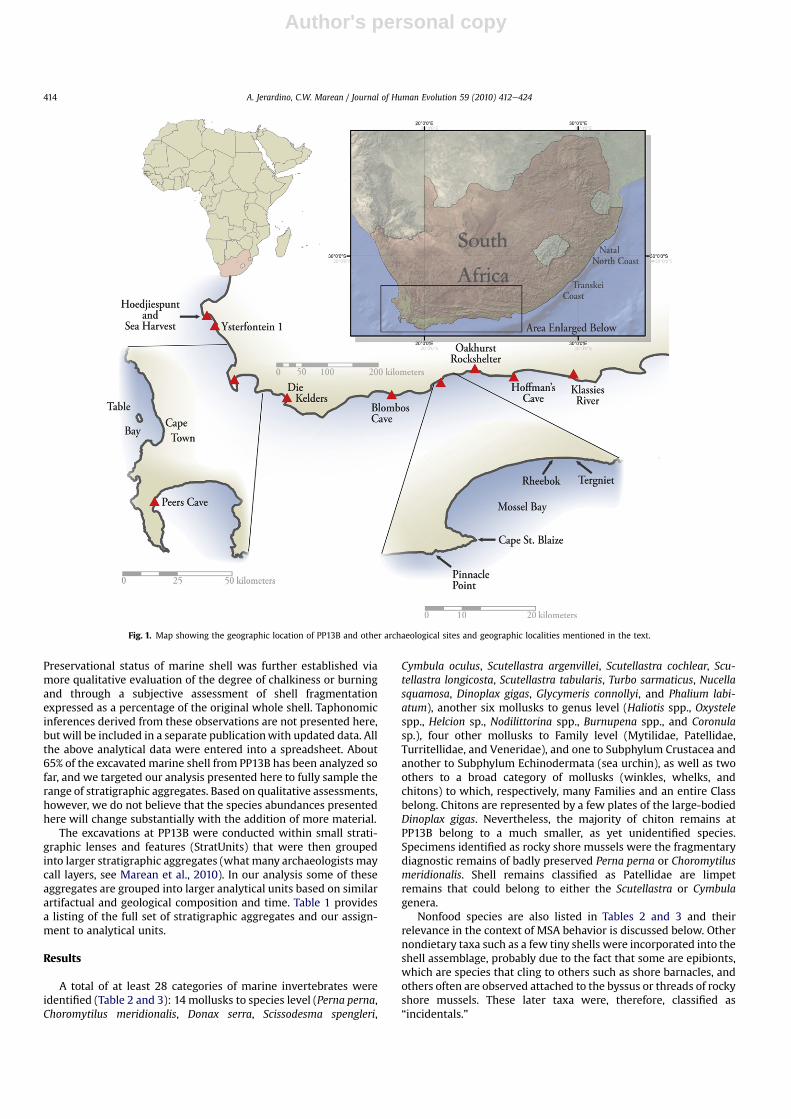
Fig. 1. Map showing the geographic location of PP13B and other archaeological sites and geographic localities mentioned in the text.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424414

Author's personal copy
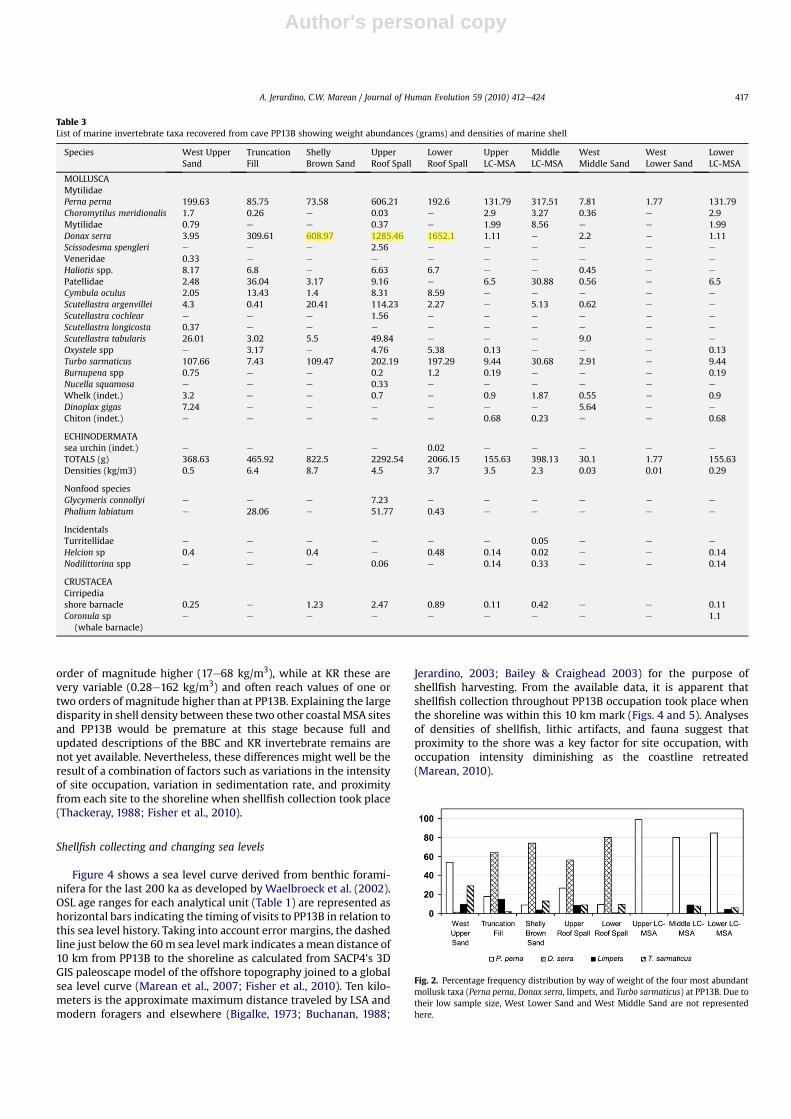
Overall, the PP13B assemblage is dominated by Perna perna,Donax serra, Turbo sarmaticus, and limpet species (mainly Scu-tellastra argenvillei and Scutellastra tabularis; Tables 2 and 3). Pernaperna dominates in all analytical units from the Northeast andWestexcavation areas, whereas Donax serra dominates in those exca-vated from the Eastern area. The sample size of shell remains is,however, markedly uneven through the PP13B sequence, withWestLower Sands andWest Middle Sands consisting of only a handful ofshells (Table 2 and 3). For this reason, many of the comparisons ofspecies representation shown and discussed here exclude the WestMiddle and Lower Sand units. However, we do note that the relativerarity of shellfish in these analytical units is interesting in and ofitself, as it likely represents either diagenesis of the shells or timeswhen people were inhabiting the cave and collecting fewer shell-fish than at other times. The West Upper Sands departs from thispattern and has a higher density than the underlying layers, but stillmuch lower density than either the Eastern or Northeastern areas.The overall counts of West Upper Sands are, nonetheless, suffi-ciently large as to be useful for description and analysis.
During the MIS 6 and MIS 5e occupations of PP13B between 164and 115 ka, MSA people harvestedmostly Perna pernawith 99e84%byweight of shell, followed bymodest numbers of Turbo sarmaticus(6e7%), limpets (4e7.7%), and very fewDonax serra (0e0.7%; Fig. 2).Shell densities appear to increase over time, showing valuesbetween 0.3 and 3.5 kg/m3 (Table 3). Interestingly, the whalebarnacle fragment recovered from the LC-MSA Lower with anassociated optically stimulated lighting (OSL) weightedmean age of164 ka is tentatively identified as Coronula diadema, and is indica-tive of scavenging of beached whale meat and blubber with Coro-nula barnacles attached to the skin of the animal (Jerardino andParkington, 1993). The Coronula barnacle is almost specificallyattached to the humpback whale (Megaptera novaeangliae) andfound very rarely on southern right whales (Eubalaena australis),probably as a result of Coronula barnacle larvae settling on southernright whales when sharing common migration routes (Best, 1991).Both of these whale species approach the southern African coast-line in large numbers and more closely than other whale species,and are also known to beach along this coastline occasionally. Themigration season today lasts from May through November.
A radical shift in the shellfish frequencies and shell densitiesoccurs in the Eastern area, which dates to the earlier phases of MIS5 (Fig. 2). Although the same range of species were harvestedbetween about 115 and 90 ka, Donax serra dominates the Easternarea with values between 56% and 79% by shell weight. Theremaining species were collected with variable intensity, withPerna perna showing values between 9e26%, Turbo sarmaticuscontributing with 1.5e13%, and limpets making up only 1e2.5% ofthe collected shellfish by weight. However, in the middle of thissequence in the Upper Roof Spall (URS) there is a clear increase inPerna perna and consequent decrease in Donax serra of about 20%.Shell densities continue to increase throughout most of theanalytical units excavated from the Eastern area, reaching highestlevels in the Shelly Brown Sand (SBS) with nearly 5 kg/m3. TheWestUpper Sands, which may be the last occupation before the site wasclosed by a dune w90 ka, documents a return to a Perna pernadominated assemblage (54%), followed by Turbo sarmaticus (29%),small numbers of limpets (8.7%), and only traces of Donax serra(1.1%). Shell density in West Upper Sand decreases sharply to0.5 kg/m3 (Table 3) relative to the Eastern area.
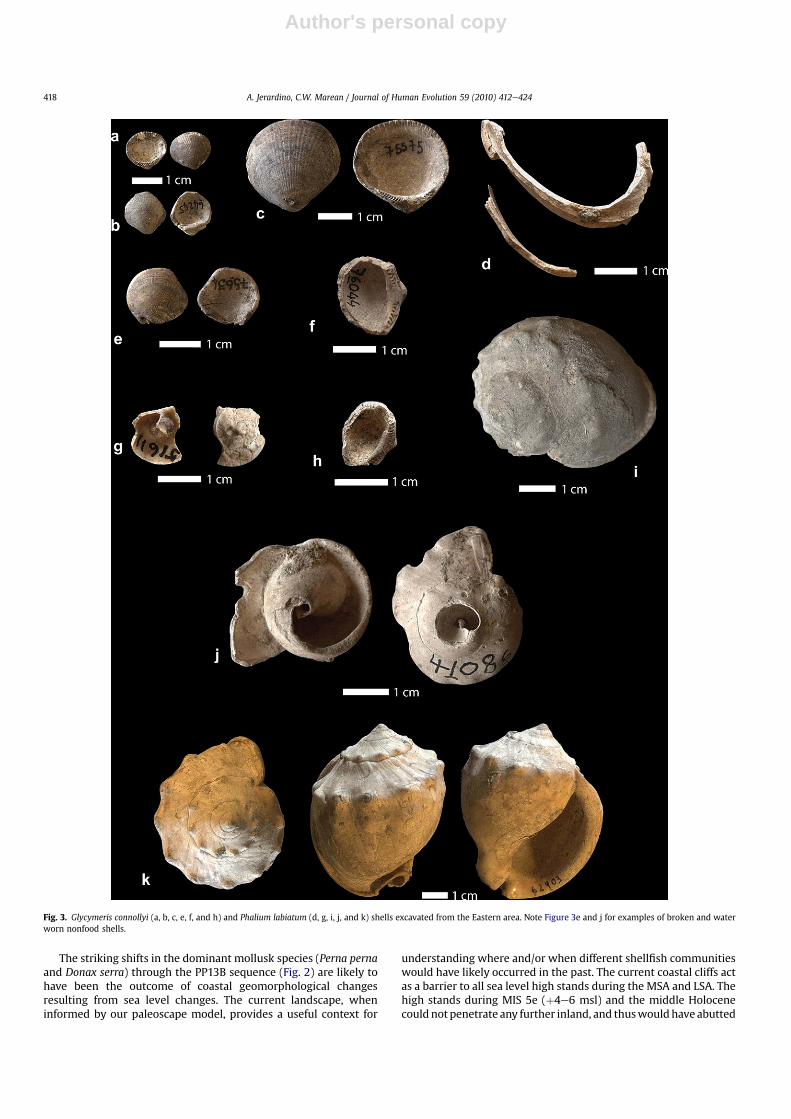
In addition to small mollusk species of no subsistence value, twoother species with relatively larger shells were also collected byMSA people and for a purpose other than subsistence (Tables 2 and3). Five valves of Glycymeris connollyi (known as ‘dog cockle’ or‘bitter sweet’), along with one whole shell and a broken shell withapex of Phalium labiatum (helmet shell), were found in the URS.Another whole Phalium labiatum was found in the Truncation Fill,and another broken shell with apex was recovered from the LRS. Afurther complete helmet shell was found associated with an ochre‘pencil’ in recent excavations between the Western and Easternareas, but is not reported here.
The current geographic distribution of Glycymeris connollyi isfrom Table Bay to the Natal north coast, with shell sizes reachinga maximum length of 32 mm. Glycymeris connollyi lives in sand atocean depths of 20e100 m, with single valves commonly washed upon the shore, usually worn and bleached (Kilburn and Rippey, 1982).Phalium labiatum is a slow moving predator snail living subtidallyand inshore among rocks or on sand at depths of 100e150 m deep,with maximum recorded shell size of 78 mm (Kilburn and Rippey,
Table 1The relation between the analytical units used here and the stratigraphic aggregates for the excavations (see Marean et al., 2010)a
Analytical units Stratigraphic aggregate Adjusted age range (ka) MIS assignment
West-Upper Sand (A) LB Sand 1 91e94 5cDB Sand 2 91e102 5cLB Sand 2 91e102 5cDB Sand 3 91e102 5c
East area shell Midden (B) Truncation Fill 35e39 3Shelly Brown Sand 91e98 5cUpper Roof Spall 91e98 5cLower Roof Spall 106e114 5d
LC-MSA Upper (C) LC-MSA Upper 115e133 May have a 5c and5e component
LC-MSA Middle (D) LC-MSA Middle 120e130 5e
West-Middle Sand (E) LBG Sand 94e134 Likely includes 2 occupationswith hiatus or truncation between
DB Sand 4a 117e166 Likely early MIS 6LBG Sand 2 117e166 Likely early MIS 6
West-Lower Sand (F) DB Sand 4b 152e166 Likely early MIS 6LBG Sand 3 152e349 Likely early MIS 6DB Sand 4c 152e349 Likely early MIS 6LBG Sand 4 152e349 Likely early MIS 6
LC-MSA Lower (G) LC-MSA Lower 153e174 Early MIS 6
a Age ranges are described by Marean et al. (2010) and are conservative estimates based on OSL ages (Jacobs, 2010), UeTh ages (Marean et al., 2010), and stratigraphicdetails. Alphabetical letters associated with analytical units correspond to those appearing in Figure 4.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424 415
Author's personal copy
1982). Phalium labiatum geographic distribution also ranges fromTable Bay to the Natal north coast (Kilburn and Rippey, 1982). Noreference to washed up individuals is found in the available litera-ture. However, SACP4 (South African Coast Paleoclimate, Paleo-environment, Paleoecology, Paleoanthropology Project) teammembers observed helmet shells washed up at the Dias beach alongMossel Bay shores after monthly low tides, but not at other MosselBay beach locations, such as Tergniet and Rhebok, or Pinnacle Point.Our sketchy observations on helmet shell washups suggest that theiravailability on the beach is not well understood but is likely theresult of local conditions. More systematic modern observations arein progress.
It is clear that both of these species live at depths well beyondunaided diving, thus excluding any possibility for direct humanaccess to living animals. Close examination of whole and brokenshells of both species shows evidence for weathering resulting fromwater and sand abrasion (Fig. 3), thus suggesting that thesemollusks were long dead before their shells were included in thecave’s deposit. The only species of hermit crab along the south coastof South Africa, the common sand hermit crab (Diogenes brevir-ostris), makes use of empty gastropod shells from several speciesincluding that of Phalium labiatum shells. Diogenes brevirostris isvery common on sheltered sandbanks and in sandy pools (Branchet al., 2007), and thus may have brought complete helmet shellswithin human collection range. But given that some of the Phaliumlabiatum shells show beach wear over broken edges and thus,indicating that these were broken and rolled before people
collected them, not all archaeological helmet shells were used byhermit crabs. None of the Glycymeris connollyi and Phalium labi-atum shells showed evidence for either natural or human madeperforations or ochre staining. The helmet shells are found only inthe Donax serra-dominated stratigraphic aggregates.
Discussion
The range of species represented at PP13B is found in the areatoday. There are no extralimital species such as are found at KRwhere the presence of Cymbula granatina is conspicuous (Voigt,1973, 1982; Thackeray, 1988). The number of taxa at PP13B issimilar to that of Blombos Cave (BBC; Henshilwood et al., 2001), butis smaller than at KR, although at KR many of the small taxa werebrought in incidentally which likely inflates the diversity there(Thackeray, 1988). With a sample size twenty times larger byweight than that recovered from nearby cave PP13A (Marean et al.,2004), it is not surprising that the number of species recorded forPP13B doubles that of this neighboring site. Whale barnacles havealso been reported from other MSA sites, namely an unpublishedobservation from Peers Cave (Yates, pers. comm.) and Ysterfontein1 (Avery et al., 2008), an indication that whales should also formpart of the list of vertebrate species exploited by MSA people atPP13B even though their bones are absent.
Marine shell density values for other MSA sites along the southcoast are available for BBC (Henshilwood et al., 2001) and KR(Thackeray, 1988: his Table 1). Shell densities at BBC are at least an
Table 2List of marine invertebrate taxa recovered from cave PP13B showing MNI abundances
Species WestUpper Sand
TruncationFill
ShellyBrown Sand
UpperRoof Spall
LowerRoof Spall
UpperLC-MSA
MiddleLC-MSA
WestMiddle Sand
WestLower Sand
LowerLC-MSA
MOLLUSCAMytilidaePerna perna 27 12 12 91 25 25 39 2 1 19Choromytilus meridionalis 1 1 e 1 e 1 1 1 e 1Rocky shore mussels 1 e e 1 e 1 1 e e 1Donax serra 1 32 43 106 191 1 e 1 e 1Scissodesma spengleri e e e 1 e e e e e e
Veneridae 1 e e e e e e e e
Haliotis spp. 1 1 e 1 3 e e 1 e e
Patellidae 1 2 1 1 e 1 5 1 e 1Cymbula oculus 1 1 1 1 1 e e e e e
Scutellastra argenvillei 1 1 1 4 1 e 1 1 e e
Scutellastra cochlear e e e 1 e e e e e e
Scutellastra longicosta 1 e e e e e e e e e
Scutellastra tabularis 1 1 1 4 e e e 1 e e
Oxystele spp e 1 e 1 1 1 e e e 1Turbo sarmaticus 9 1 9 18 15 1 4 1 e 1Burnupena spp 1 e e 1 1 1 e e e 1Nucella squamosa e e e 1 e e e e e e
Whelk (indet.) 1 e e 1 e 2 1 1 e 1Dinoplax gigas 1 e e e e e e 1 e e
Chiton (indet.) e e e e e 1 1 e e 1
ECHINODERMATAsea urchin (indet.) e e e e 1 e e e e e
TOTALS 49 53 68 234 239 35 53 11 1 28
Nonfood speciesGlycymeris connollyi e e e 3 e e e e e e
Phalium labiatum e 1 e 2 1 e e e e e
IncidentalsTurritellidae e e e e e e 1 e e e
Helcion sp 1 e 1 e 1 1 1 e e 1Nodilittorina spp e e e 1 e 1 4 e e 1
CRUSTACEACirripediashore barnacle 1 e 1 1 1 1 1 e e 1Coronula sp
(whale barnacle)e e e e e e e e e 1
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424416
Author's personal copy
order of magnitude higher (17e68 kg/m3), while at KR these arevery variable (0.28e162 kg/m3) and often reach values of one ortwo orders of magnitude higher than at PP13B. Explaining the largedisparity in shell density between these two other coastal MSA sitesand PP13B would be premature at this stage because full andupdated descriptions of the BBC and KR invertebrate remains arenot yet available. Nevertheless, these differences might well be theresult of a combination of factors such as variations in the intensityof site occupation, variation in sedimentation rate, and proximityfrom each site to the shoreline when shellfish collection took place(Thackeray, 1988; Fisher et al., 2010).
Shellfish collecting and changing sea levels
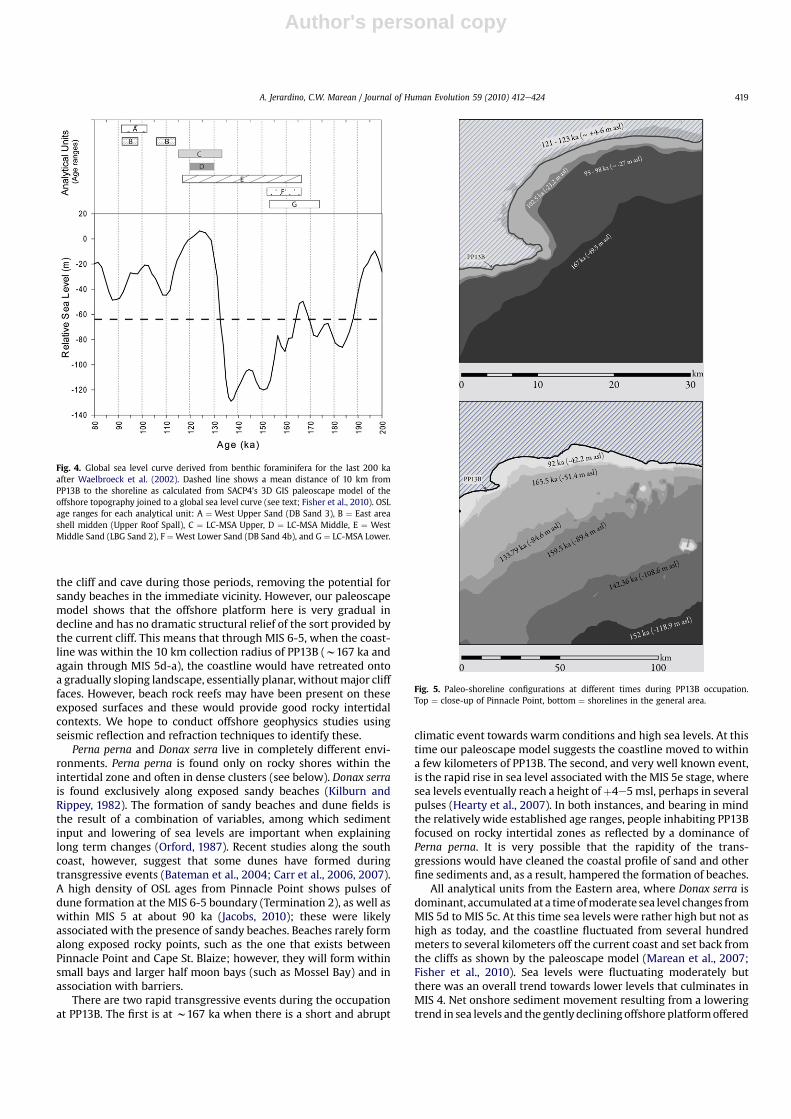
Figure 4 shows a sea level curve derived from benthic forami-nifera for the last 200 ka as developed by Waelbroeck et al. (2002).OSL age ranges for each analytical unit (Table 1) are represented ashorizontal bars indicating the timing of visits to PP13B in relation tothis sea level history. Taking into account error margins, the dashedline just below the 60m sea level mark indicates a mean distance of10 km from PP13B to the shoreline as calculated from SACP4’s 3DGIS paleoscape model of the offshore topography joined to a globalsea level curve (Marean et al., 2007; Fisher et al., 2010). Ten kilo-meters is the approximate maximum distance traveled by LSA andmodern foragers and elsewhere (Bigalke, 1973; Buchanan, 1988;
Jerardino, 2003; Bailey & Craighead 2003) for the purpose ofshellfish harvesting. From the available data, it is apparent thatshellfish collection throughout PP13B occupation took place whenthe shoreline was within this 10 km mark (Figs. 4 and 5). Analysesof densities of shellfish, lithic artifacts, and fauna suggest thatproximity to the shore was a key factor for site occupation, withoccupation intensity diminishing as the coastline retreated(Marean, 2010).
Table 3List of marine invertebrate taxa recovered from cave PP13B showing weight abundances (grams) and densities of marine shell
Species West UpperSand
TruncationFill
ShellyBrown Sand
UpperRoof Spall
LowerRoof Spall
UpperLC-MSA
MiddleLC-MSA
WestMiddle Sand
WestLower Sand
LowerLC-MSA
MOLLUSCAMytilidaePerna perna 199.63 85.75 73.58 606.21 192.6 131.79 317.51 7.81 1.77 131.79Choromytilus meridionalis 1.7 0.26 e 0.03 e 2.9 3.27 0.36 e 2.9Mytilidae 0.79 e e 0.37 e 1.99 8.56 e e 1.99Donax serra 3.95 309.61 608.97 1285.46 1652.1 1.11 e 2.2 e 1.11Scissodesma spengleri e e e 2.56 e e e e e e
Veneridae 0.33 e e e e e e e e e
Haliotis spp. 8.17 6.8 e 6.63 6.7 e e 0.45 e e
Patellidae 2.48 36.04 3.17 9.16 e 6.5 30.88 0.56 e 6.5Cymbula oculus 2.05 13.43 1.4 8.31 8.59 e e e e e
Scutellastra argenvillei 4.3 0.41 20.41 114.23 2.27 e 5.13 0.62 e e
Scutellastra cochlear e e e 1.56 e e e e e e
Scutellastra longicosta 0.37 e e e e e e e e e
Scutellastra tabularis 26.01 3.02 5.5 49.84 e e e 9.0 e e
Oxystele spp e 3.17 e 4.76 5.38 0.13 e e e 0.13Turbo sarmaticus 107.66 7.43 109.47 202.19 197.29 9.44 30.68 2.91 e 9.44Burnupena spp 0.75 e e 0.2 1.2 0.19 e e e 0.19Nucella squamosa e e e 0.33 e e e e e e
Whelk (indet.) 3.2 e e 0.7 e 0.9 1.87 0.55 e 0.9Dinoplax gigas 7.24 e e e e e e 5.64 e e
Chiton (indet.) e e e e e 0.68 0.23 e e 0.68
ECHINODERMATAsea urchin (indet.) e e e e 0.02 e e e e e
TOTALS (g) 368.63 465.92 822.5 2292.54 2066.15 155.63 398.13 30.1 1.77 155.63Densities (kg/m3) 0.5 6.4 8.7 4.5 3.7 3.5 2.3 0.03 0.01 0.29
Nonfood speciesGlycymeris connollyi e e e 7.23 e e e e e e
Phalium labiatum e 28.06 e 51.77 0.43 e e e e e
IncidentalsTurritellidae e e e e e e 0.05 e e e
Helcion sp 0.4 e 0.4 e 0.48 0.14 0.02 e e 0.14Nodilittorina spp e e e 0.06 e 0.14 0.33 e e 0.14
CRUSTACEACirripediashore barnacle 0.25 e 1.23 2.47 0.89 0.11 0.42 e e 0.11Coronula sp
(whale barnacle)e e e e e e e e e 1.1
Fig. 2. Percentage frequency distribution by way of weight of the four most abundantmollusk taxa (Perna perna, Donax serra, limpets, and Turbo sarmaticus) at PP13B. Due totheir low sample size, West Lower Sand and West Middle Sand are not representedhere.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424 417
Author's personal copy
The striking shifts in the dominant mollusk species (Perna pernaand Donax serra) through the PP13B sequence (Fig. 2) are likely tohave been the outcome of coastal geomorphological changesresulting from sea level changes. The current landscape, wheninformed by our paleoscape model, provides a useful context for
understanding where and/or when different shellfish communitieswould have likely occurred in the past. The current coastal cliffs actas a barrier to all sea level high stands during the MSA and LSA. Thehigh stands during MIS 5e (þ4e6 msl) and the middle Holocenecouldnot penetrate any further inland, and thuswouldhave abutted
Fig. 3. Glycymeris connollyi (a, b, c, e, f, and h) and Phalium labiatum (d, g, i, j, and k) shells excavated from the Eastern area. Note Figure 3e and j for examples of broken and waterworn nonfood shells.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424418
Author's personal copy
the cliff and cave during those periods, removing the potential forsandy beaches in the immediate vicinity. However, our paleoscapemodel shows that the offshore platform here is very gradual indecline and has no dramatic structural relief of the sort provided bythe current cliff. This means that through MIS 6-5, when the coast-line was within the 10 km collection radius of PP13B (w167 ka andagain through MIS 5d-a), the coastline would have retreated ontoa gradually sloping landscape, essentially planar, withoutmajor clifffaces. However, beach rock reefs may have been present on theseexposed surfaces and these would provide good rocky intertidalcontexts. We hope to conduct offshore geophysics studies usingseismic reflection and refraction techniques to identify these.
Perna perna and Donax serra live in completely different envi-ronments. Perna perna is found only on rocky shores within theintertidal zone and often in dense clusters (see below). Donax serrais found exclusively along exposed sandy beaches (Kilburn andRippey, 1982). The formation of sandy beaches and dune fields isthe result of a combination of variables, among which sedimentinput and lowering of sea levels are important when explaininglong term changes (Orford, 1987). Recent studies along the southcoast, however, suggest that some dunes have formed duringtransgressive events (Bateman et al., 2004; Carr et al., 2006, 2007).A high density of OSL ages from Pinnacle Point shows pulses ofdune formation at the MIS 6-5 boundary (Termination 2), as well aswithin MIS 5 at about 90 ka (Jacobs, 2010); these were likelyassociated with the presence of sandy beaches. Beaches rarely formalong exposed rocky points, such as the one that exists betweenPinnacle Point and Cape St. Blaize; however, they will form withinsmall bays and larger half moon bays (such as Mossel Bay) and inassociation with barriers.
There are two rapid transgressive events during the occupationat PP13B. The first is at w167 ka when there is a short and abrupt
climatic event towards warm conditions and high sea levels. At thistime our paleoscape model suggests the coastline moved to withina few kilometers of PP13B. The second, and very well known event,is the rapid rise in sea level associated with the MIS 5e stage, wheresea levels eventually reach a height ofþ4e5msl, perhaps in severalpulses (Hearty et al., 2007). In both instances, and bearing in mindthe relatively wide established age ranges, people inhabiting PP13Bfocused on rocky intertidal zones as reflected by a dominance ofPerna perna. It is very possible that the rapidity of the trans-gressions would have cleaned the coastal profile of sand and otherfine sediments and, as a result, hampered the formation of beaches.
All analytical units from the Eastern area, where Donax serra isdominant, accumulated at a timeofmoderate sea level changes fromMIS 5d to MIS 5c. At this time sea levels were rather high but not ashigh as today, and the coastline fluctuated from several hundredmeters to several kilometers off the current coast and set back fromthe cliffs as shown by the paleoscape model (Marean et al., 2007;Fisher et al., 2010). Sea levels were fluctuating moderately butthere was an overall trend towards lower levels that culminates inMIS 4. Net onshore sediment movement resulting from a loweringtrend in sea levels and the gently decliningoffshoreplatformoffered
Fig. 4. Global sea level curve derived from benthic foraminifera for the last 200 kaafter Waelbroeck et al. (2002). Dashed line shows a mean distance of 10 km fromPP13B to the shoreline as calculated from SACP4’s 3D GIS paleoscape model of theoffshore topography joined to a global sea level curve (see text; Fisher et al., 2010). OSLage ranges for each analytical unit: A ¼ West Upper Sand (DB Sand 3), B ¼ East areashell midden (Upper Roof Spall), C ¼ LC-MSA Upper, D ¼ LC-MSA Middle, E ¼ WestMiddle Sand (LBG Sand 2), F ¼ West Lower Sand (DB Sand 4b), and G ¼ LC-MSA Lower.
Fig. 5. Paleo-shoreline configurations at different times during PP13B occupation.Top ¼ close-up of Pinnacle Point, bottom ¼ shorelines in the general area.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424 419

Author's personal copy
the right conditions for the formation of sandy beaches withincollection reach of PP13B. This scenario is likely to explain thedominance of Donax serra in these occupations and the relativepaucity of Perna perna and other rocky intertidal fauna.
Our conclusion that sandy beaches were probably well devel-oped during MIS 5d and MIS 5c as a result of lowering sea levelsseems to be supported by observations from KR dating to after MIS5c. Donax serra, while rare overall at KR, dominates the shellfishassemblage with a short and strong spike in abundance just beforemember RF that precedes the Howiesons Poort (HP) horizons. KMmarine shell concentrations are recorded at their lowest ever in HPdeposits (Thackeray, 1988: his Fig. 1). According to Deacon andGeleijnse (1988: 9), member RF is a distinctive 0.5 m thick sandhorizon with a few thin partings of occupational material and roofblocks, and also rich in “.chance inclusions like small mammalremains.”, all of which was interpreted as indicative of slowdeposition and low occupational frequency. Thackeray (1988)interpreted the lowest concentrations of marine shell in theuppermost HP deposits as an indication of greater distance to theshoreline as a result of lowering of sea levels, a contention sup-ported by SACP4’s 3D GIS paleoscape model (Fisher et al., 2010). Animproved chronology for the KR sequence would certainly help testthis interpretation. The relatively intense collection of Donax serra,as reflected by abundant quantities of this species at PP13B Easternarea (a small portion of which has been sampled), was possibleprobably because of rapid lowering of sea levels initiated around110,000 years ago that led to the formation of long and exposedsandy beaches. Of course, the ability by MSA people to exploit thistype of shoreline is a precondition, a matter that is discussed below.
Exploiting shellfish among rocks and beach on the south coast
Donax serra (white mussel) burrows in sandy substrate and isfound invariable concentrations alongexposed sandybeaches in thewest and south coasts of South Africa (McLachlan and Hanekom,1979; Kilburn and Rippey, 1982). Along the south coast, thisspecies occupies themid-intertidal zone, while along thewest coastit is found deeper within the lower intertidal and shallow subtidalzone (McLachlan et al., 1979; McLachlan and Hanekom,1979; Donnand Els, 1990). Easier access to this resource along the south coastwas also the case in the past as reflected by their higher frequenciesin nearby LSA shell middens when compared to their equivalentalong thewest coast (Döckel,1998; Binneman, 2004/2005; Jerardinoet al., 2009). Foraging for Donax serra involves walking down thebeach slope aboutmean tide level and feeling gentlywith the feet, asthis species burrows down quickly when disturbed, which is anability that Donax species have evolved in order to maintain theirposition on highly dynamic beaches (Donn and Els, 1990). Smallerindividuals (30e45 mm in shell length) are found at sand depths of0e7 cm while larger individuals (45e60 mm shell length) arelocated between 7 and 15 cm into the sand (McLachlan andHanekom, 1979). Once a patch of Donax serra is located, hands areused to dig quickly and grab as many shellfish as possible beforemanyof themcanburrow furtherdown into the sand. This collectionmethod is currently followed by local fishermen in search for baitand snacking (Kilburn and Rippey, 1982).
Perna perna is the dominant rocky shore mussel on the southand east coasts, often forming dense beds on semihorizontal andsteep rock surfaces and also pools where water movement is brisk,from the mid-intertidal to a fewmeters below (Kilburn and Rippey,1982; Van Erkom Schurink and Griffiths, 1990; Branch et al., 2007).The rocks on which they grow can include rough and relativelysteep Table Mountain Sandstone bedrock, such as is present todayat Pinnacle Point, or more gently and horizontally bedded ‘beachrock’ and eroded aeolianite, such as near Tergniet and Reebok
within Mossel Bay. Further to the east, Perna perna grows also ontillite, shale, and dolorite (Hockey et al., 1988). Aware of tidalmovements that follow the lunar cycle, modern shellfish foragers ofthe South African east coast (Transkei) harvest intensively duringspring low tides with some opportunistic collecting at other timesas seen also along the southern Pacific coast of South America(Bigalke, 1973; Siegfried et al., 1985; Durán et al., 1987). Harvest ofPerna perna and other rocky shore mollusks is done on the mostaccessible rocky platforms by women of all ages and youngsters,with the collection starting in the upper reaches of shellfish bedsand then moving down to the more exposed rocks washed by thesurf. Care is taken from dangerous waves by briefly movingupslope, with catches being piled up temporarily on dry rockshigher up the tidal gradient (Bigalke,1973; Bigalke and Voigt,1973).
The collection of Donax serra requires walking down the beachprofile at about the level of the hip, or higher for relatively shortpeople as are most individuals from indigenous southern Sangroups, and dealing with wave action. The aid of a container and/oran accompanying helper to receive the catch would assist withlarger catches of Donax serra. Rocky intertidal species (Perna pernaand limpet colonies), on the other hand, are largely visible, fixed torocks to a large degree, and their collection entails relativelyshallow entry into the water during low tides. Containers are notalways necessary and the aid of a helper is seldom needed whencollecting mollusks from rocks. Hence, the procurement of Donaxserra involves more than simple foraging on an encounter basis.Rather, it seems to require a relatively more complex set of vari-ables such as anticipation, planning, and possibly also additionalhelp in the form of dilly bags and/or cooperationwith other people.It could be argued that shellfish collection along rocky shores mightwell have required dealing with the same complex set of variablesmentioned above, particularly the use of moon cycles for predictingtidal movements. Although this might have been the case, rockyshore mollusks can also be collected when their hard substrate isnot fully exposed by low tides (Bigalke, 1973; Siegfried et al., 1985;Durán et al., 1987). The harvest of any substantial numbers of Donaxserra seems to leave collectors with little choice but to anticipate,plan, provide bags for containing the catch, and possibly also tocooperate with others. In the context of hunting, these skills areusually indicative of an aspect of behavioral modernity (McBreartyand Brooks, 2000). Further and more conclusive data grounded intaphonomic studies and strong understanding of the main vari-ables involved in shellfish collection through actualistic studiesshould shed more light on this issue (Bird and Bliege-Bird, 1997;Bird et al., 2002).
Collection of nonfood species
Glycymeris connollyi and Phalium labiatum specimens describedherewere likely collected from the beach and brought to the site bypeople for nonutilitarian purposes. The geology of PP13B is devoidof fossil shell material and, although sea gulls are known to preyupon sandy shore invertebrates such as Donax serra, the spoils offeeding activities are invariably strewn immediately above the highwater mark (Siegfried, 1977). It is also evident that sea gulls wouldnot have been attracted to empty and/or broken shells, thusmakingthem an unlikely agent of dispersal beyond the usual radius.Although the coastline would have been as close to PP13B as it istoday, or about 4,000 m away at most during MIS 5, major stormsare also unlikely to have been responsible for the occurrence ofGlycymeris connollyi and Phalium labiatum specimens in this site.Large waves would have either washed some of the archaeologicalmaterial out of the cave and/or added other shells from otherspecies with evidence of sea weathering. Our field and laboratoryobservations do not support either of these scenarios. Likewise, it is
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424420

Author's personal copy
difficult to explain why relatively large sea weathered shells fromother species are also not present in the Donax serra-dominatedassemblages if Phalium labiatum and Glycymeris connollyi madetheir way into the sites by chance while harvesting Donax serra insandy beach environments. It is true that Phalium labiatum andGlycymeris connollyi were recovered from stratigraphic layerswhere larger amounts of shell material was excavated. However,possible sample size effects need not concern us here, as evensmaller numbers of Phalium labiatum shells than the ones found atPP13B have been recovered from other MSA and LSA sites wheremuch larger volumes of deposit were excavated (see below).
Why would MSA people collect water worn shells, both wholeand broken? Before attempting to answer this question, it shouldfirst be noted that such an occurrence is not unique to PP13B cave aswater worn helmet shells without human modification have alsobeen found at other South African MSA and LSA sites along thesouth coast, such as MSA deposits at KR (Voigt, 1982; Thackeray,1988), LSA deposits at Oakhurst rock shelter (A.J., pers. obs.), andLSA deposits at Hoffman’s Cave (Kyriacou, pers. comm.). Also, beachworn and perforated helmet shells have been reported for DieKelders Cave LSA layers (Schweitzer, 1979; A.J., pers. obs.). Helmetshells from the same genus but different species have beenencountered at sites in the eastern Mediterranean Basin, includingUpper Paleolithic burials and Neolithic worship sites (Reese, 1989,2005; Bar-Yosef Mayer, 2005).
Nonperforated Glycymeris connollyi shells have also been exca-vated from LSA levels of Hoffman’s Cave (Kyriacou, pers. comm.) andDie Kelders (Schweitzer, 1979) and specimens from different Glycy-meris species have also been recovered from Upper Paleolithic andlater sites in Cyprus, Italy, and the Levant including Sefunim Cave inMountCarmel andRasel-Kelb (D.Bar-Yosef, pers. comm.;Reese,1982,1998, 1999; Stiner, 1994). LSA hunter-gatherers along the south andwest coasts of South Africa used Glycymeris connollyi and/or Glycy-meris quecketti shells as pendants (Schweitzer, 1979; Schweitzer andWilson, 1982; Inskeep, 1987; Parkington and Poggenpoel, 1987;
Robey, 1987; Sealy et al., 2002; Smith, 2006), just as Middle andUpper Paleolithic groups seem to have done in the Mediterraneanbasin, including the site of Qafzeh (Reese, 1999; Taborin, 2003; Bar-Yosef Mayer, 2005; Bar-Yosef Mayer et al., 2009). At Qafzeh, dated to92 ka, Bar-Yosef Mayer et al. (2009) report Glycymeris shells used aspendants in associationwith early modern humans.
The beauty of these shells is evident to the modern eye, withsmoothness and near perfect bilateral symmetry in the case of Gly-cymeris connollyi and alluring gloss, round body and harmonicallyshaped spiral forming a lowapex in the case of Phalium labiatum. Thisaesthetic appreciation probably applied to LSA people as well sincethey specifically collected Glycymeris connollyi, and in one instancealso Phalium labiatum, from a range of other shells available to themformaking pendants. The fact that MSA people collected these shellsalso for apparent nondietary purposes seems to suggest the sameaesthetic appreciation of these shells by MSA people around MIS 5d.Moreover, the fact that broken and/or water worn shells were alsocollected may imply that MSA people were satisfied with picking up“second best choice” after whole fresh specimens, something thatmodern humans did later on in history with helmet shells and otherspecies (Reese, 1982, 1989, 1999; Alvarez-Fernández, 2005: 364).
We do not argue that Glycymeris connollyi and Phalium labiatumshells from PP13B cave were “pre-forms” for pendant or beadmaking. Instead, we suggest that these shells were selected fortheir particular aesthetic qualities just as LSA hunter-gatherersselected particular shells, and not any other shell, as keepsakes andoften also for making pendants. Voigt (1982: 165) arrived ata similar conclusion when discussing the presence of a few largewater worn helmet shells and that of a large subtidal whelk ina similar condition at KR. MSA people might have conferred somespecial value to these particular shells because it seems to havewarranted the collection of even broken and/or weathered speci-mens, a choice that is not repeated with other shells from differentspecies. Moreover, the spatial distribution of Glycymeris and helmetshells in PP13B cave is not random but rather clustered, arguing
Fig. 6. Spatial distribution of Glycymeris and Phalium labiatum shells in the context of cave PP13B.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424 421

Author's personal copy
against their inclusion into the deposits by storm waves, othernonhuman agents, or by chance when collecting Donax serramussels (Figs. 6 and 7). We contend that this could potentially beregarded as a w110 ka old “material proxy” for material engage-ment, representation, and symbolic thought that lie at the base ofaesthetic appreciations (Gell, 1998). In the context of relatedobservations, this finding is significantly older than the use of shellbeads byMSA people in southern Africa (Henshilwood et al., 2004),older and broadly contemporary with some of the earliest use ofshells as beads in the Levant and North Africa (Vanhaeren et al.,2006; Bouzouggar et al., 2007; Bar-Yosef Mayer et al., 2009), andsubsequent to some of the earliest use of pigment at PP13B caveabout 164 ka (Marean et al., 2007). It is thus expected that nonfoodshells will soon be identified and studied more regularly at otherUpper andMiddle Paleolithic sites now that their relevance is beinghighlighted.
Conclusions
Awide range of mollusk species was gathered and brought backto PP13B, but this collection focused on three to four taxa at anygiven time, namely Perna perna, Donax serra, Scutellastra andCymbula limpets, and Turbo sarmaticus. Both rocky shore and sandybeaches were exploited within a foraging radius to the nearestshore of nomore than 10 km from the cave and with no extralimitalspecies represented in the catches.
Species frequencies appear to be in good accord with predictedcoastal geomorphological changes as a response to sea levelchange. Highest and dominating frequencies of the sandy shoredweller, Donax serra, are recorded for MIS 5d to MIS 5c, at a time ofrapid sea level regression and when OSL dating of dunes in thePinnacle Point area show pulses of dune formation and their likelyassociation with well developed sandy beaches. On the other hand,rocky shore species, such as Perna perna, followed by Turbo sar-maticus and limpets, dominate the shellfish assemblage of PP13B ata time of transgressive events. The outcome of rapid rising sealevels would have likely involved the cleaning away of sand andfine sediments from the coastal profile and exposure of extensiverocky shores.
Fromavailable ethnohistorical observationsgathered in theSouthAfrican east coast and elsewhere, it appears that shellfish harvestingby modern humans involves consideration of a number of variables,such as, but not limited to, tidal movements, accessibility to shellfishbeds, wave action, and minimal extractive technology. These vari-ables apply to both rocky shore and sandy beach habitats, withperhaps added complexity when harvesting in the latter. Anticipa-tion, planning and cooperationwith a helper, and/or use of dilly bagsseem to be a precondition for the harvest of quantities (beyonda handful) ofDonax serra as reflected in the large and relatively densevolumes of marine shell present in the Eastern excavation area andother MSA sites. Consideration of this type of shellfish collection hasa bearing on discussions about behavioral modernity among MSAshellfish foragers. However, further shellfish foraging studies(ethnohistory and actualistic studies) should complement ourquantifiedobservations presentedhere andallowus tomove beyondthe entertainment of informed speculation. The analysis of theremaining 35% of the excavated marine shell assemblage (almostentirely from the East excavation area) from PP13Bwill also improvethe empirical foundation of these observations.
Helmet and Glycymeris shells, most of which were probablyempty when collected, were also brought back to PP13B fornonutilitarian purposes around 110 ka and during the accumulationof Donax serra-dominated deposits from the East excavation area.Their spatial distribution on the cave floor is not random but rela-tively clustered. None of these nondietary shells at PP13B areperforated and the occurrence of helmet shells has been recorded atother MSA and LSA coastal sites in South Africa and the easternMediterranean Basin. Glycymeris shells, with and without perfora-tionoruse-wearmarks aspendants, haveonlybeen recorded for LSAsites in South Africa, but these are also present in early modernhuman Middle Paleolithic sites and Holocene sites in the Levant.Broken helmet shells that had retainedmuch of the apex and part ofthe whorl and damaged Glycymeris worn around the flaked edgeappear to have been equally valued and regarded as worth takingback to the site. We thus suggest that these ‘beauty shells’ werecollected by MSA people for their particular aesthetic qualities andretained as keepsakes. The recognition of these traits seems toforeshadow the use of these types of shells as pendants later on inprehistory. A thorough inventory of helmet and Glycymeris shellsand possibly other types of shells in similar weathering conditionfrom South African MSA and LSA contexts, recording of theiroccurrence on modern beaches, and their detailed analysis would
Fig. 7. Comparison of the spatial distribution of Glycymeris and Phalium labiatum shells(D) in excavated quadrants of the Eastern area of PP13B cave with that of majorfindings in Upper Roof Spall and Shelly Brown Sand only. A ¼ vertebrate fauna andlithic artifacts, B ¼ other marine shell, C ¼ possible hearth areas as reflected bymagnetic susceptibility measurements (�108 m3/kg; see Herries and Fisher, 2010).Note that Upper Roof Spall and Shelly Brown Sand thin out significantly towards thesouth.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424422

Author's personal copy
certainly help understand their significance and relevance, if any, forearly modern human behavior.
Acknowledgements
The support of Mary Leslie and the South African HeritageResources Agency towards this project is gratefully acknowledged.Many thanks to D. Bar-Yosef, D. Reese, Ana Pinto Llona, and SarahWurz for making bibliographic references available, and to T. Steelealso for bibliographic references and discussions on the subject ofthis paper. Access to Die Kelders Cave 1 helmet shells stored atIZIKO: South African Museum is also gratefully acknowledged.Erich Fisher compiled Figures 1, 5, 6, and 7, Hope Williams gener-ated Figures 3 and 4, and Maria Grazia Galimberti helped withFigure 2. This research was funded by the National Science Foun-dation (USA; grants # BCS-9912465, BCS-0130713, and BCS-0524087 to Marean), funding from the Huxleys, the Hyde FamilyFoundation, the Institute for Human Origins, and Arizona StateUniversity.
References
Alvarez-Fernández, E., 2005. La explotación de los moluscos marinos durante elPalaeolítico superior y el Mesolítico en la región cantábrica y en el Valle delEbro: pasado y presente de la investigación. Munibe 57, 359e368.
Avery, G., Halkett, D., Orton, J., Steele, T., Klein, R.G., 2008. The Ysterfontein 1 MiddleStone Age rockshelter and the evolution of coastal foraging. S. Afr. Archaeol. Soc.Goodwin Ser. 10, 66e89.
Bailey, G., Craighead, A.S., 2003. Late Pleistocene and Holocene coastal palae-oeconomies: a reconsideration of the molluscan evidence from Northern Spain.Geoarchaeology 18, 175e204.
Bar-Yosef Mayer, D.E., 2005. The exploitation of shells as beads in the Paleolithic andNeolithic of the Levant. Paleorient 31, 176e185.
Bar-Yosef Mayer, D.E., Vandermeersch, B., Bar-Yosef, O., 2009. Shells and ochre inMiddle Palaeolithic Qafzeh cave, Israel: indications for modern behavior.J. Hum. Evol. 56, 307e314.
Bateman, M.D., Holmes, P.J., Carr, A.S., Horton, B.P., Jaiswal, M.K., 2004. Aeolianiteand barrier dune construction spanning the last two glacialeinterglacial cyclesfrom the southern Cape coast, South Africa. Quatern. Sci. Rev. 23, 1681e1698.
Best, P., 1991. The presence of coronuline barnacles on a southern right whaleEubalaena australis. S. Afr. J. Marine Sci. 11, 585e587.
Bigalke, E.H., 1973. The exploitation of shellfish by coastal tribesmen of the Transkei.Annls. Provincial Cape Mus. (Nat. Hist.) 9, 159e175.
Bigalke, E.H., Voigt, E.A., 1973. The inter-disciplinary aspect of a study of shellfishexploitation by indigenous coastal communities. S. Afr. Mus. Assoc. Bull. 10,256e261.
Binneman, J., 2004/2005. Archaeological research along the south-eastern Capecoast Part I: open-air shell middens. S. Afr. Field Archaeol 13 and 14, 49e77.
Bird, D.W., Bliege-Bird, R.L., 1997. Contemporary shellfish gathering strategiesamong the Meriam of the Torres Strait Islands, Australia: testing predictions ofa central place foraging model. J. Archaeol. Sci. 24, 39e63.
Bird, D.W., Richardson, J.L., Veth, P.M., Barham, A.J., 2002. Explaining shellfishvariability in middens on the Meriam Islands, Torres Strait, Australia.J. Archaeol. Sci. 29, 457e469.
Bouzouggar, B., Barton, N., Vanhaeren, M., D’Errico, F., Collcutt, S., Higham, T.,Hodge, E., Parfitt, S., Rhodes, E., Schwenninger, J., Stringer, C., Turner, E.,Ward, S., Moutmir, A., Stambouli, A., 2007. 82,000-year-old shell beads fromNorth Africa and implications for the origins of modern human behavior. Proc.Natl. Acad. Sci. USA 104, 9964e9969.
Branch, G.M., Griffiths, C.L., Branch, M.L., Beckley, L.E., 2007. Two Oceans: a Guide tothe Marine Life of Southern Africa. Struik, Cape Town.
Buchanan, W., 1988. Shellfish in Prehistoric Diet: Elands Bay, SW Cape Coast, SouthAfrica. Cambridge Monographs in African Archaeology 31, BAR InternationalSeries 455, Oxford.
Carr, A.S., Bateman, M.D., Holmes, P.J., 2007. Developing a 150 ka luminescencechronology for the barrier dunes of the southern Cape, South Africa. Quatern.Geochronol 2, 110e116.
Carr, A.S., Thomas, D.S.G., Bateman, M.D., 2006. Climatic and sealevel controls onLate Quaternary eolian activity on the Agulhas Plain, South Africa. Quatern. Res.65, 252e263.
Deacon, H.J., Geleijnse, V.B., 1988. The stratigraphy and sedimentology of the mainsite sequence, Klasies River, South Africa. S. Afr. Archaeol. Bull. 43, 5e14.
Döckel, W., 1998. Reinvestigation of the Matjes River Rock Shelter. Unpublished MAdissertation. University of Stellenbosch, Stellenbosch.
Donn, T., Els, S.F., 1990. Burrowing times of Donax serra from the south and westcoasts of South Africa. The Veliger 33, 355e358.
Durán, L.R., Castilla, J.C., Oliva, D., 1987. Intensity of human predation on rockyshores at Las Cruces in central Chile. Environ. Conserv. 14, 143e149.
Erlandson, J., 2001. The Archaeology of aquatic adaptations: paradigms for a newmillennium. J. Archaeol. Res. 9, 287e350.
Fisher, E., Bar-Mathews, M., Jacobs, Z., Jerardino, A., Marean, C.W., 2010. Middle andLate Pleistocene paleoscape modeling along the southern coast of South Africa.Quatern. Sci. Rev. 29, 1382e1398.
Gell, A., 1998. Art and Agency: an Anthropological Theory. Oxford University Press,Oxford.
Halkett, D., Hart, T., Yates, R., Volman, T.P., Parkington, J.E., Orton, J., Klein, R.G., Cruz-Uribe, K., Avery, G., 2003. First excavation of intact Middle Stone Age layers atYsterfontein, Western Cape Province, South Africa: implications for MiddleStone Age ecology. J. Archaeol. Sci. 30, 955e971.
Hearty, P.J., Hollin, J.T., Neumann, A.C., OÆLeary, M.J., McCulloch, M., 2007. Globalsea-level fluctuations during the Last Interglaciation (MIS 5e). Quatern. Sci. Rev.26, 2090e2112.
Herries, A.I.R., Fisher, E.C., 2010. Multi-dimensional modelling of magnetic miner-alogy as a proxy for fire use and spatial patterning: evidence from the MiddleStone Age sea cave site of Pinnacle Point 13B (Western Cape, South Africa).J. Hum. Evol. 59 (3e4), 306e320.
Henshilwood, C.S., d’Errico, F., Vanhaeren, M., Van Niekerk, K., Jacobs, Z., 2004.Middle Stone Age shell beads from South Africa. Science 304, 404.
Henshilwood, C.S., Sealy, J.C., Yates, R., Cruz-Uribe, K., Goldberg, P., Grine, F.E.,Klein, R.G., Poggenpoel, C., Van Niekerk, K., Watts, I., 2001. Blombos Cave,southern Cape, South Africa: preliminary report on the 1992e1999 excavationsof the Middle Stone Age levels. J. Archaeol. Sci. 28, 421e448.
Hockey, P.A.R., Bosman, A.L., Siegfried, W.R., 1988. Patterns and correlates of shell-fish exploitation by coastal people in Transkei: an enigma of protein production.J. Appl. Ecol. 25, 353e363.
Inskeep, R.R. (Ed.), 1987. Nelson Bay Cave. Cape Province, South Africa: The Holo-cene Levels. British Archaeological Reports International Series, 357(i)þ(ii)(Oxford).
Jacobs, Z., 2010. An OSL chronology for the sedimentary deposits from PinnaclePoint Cave 13B - a punctuated presence. J. Hum. Evol. 59 (3e4), 289e305.
Jacobs, Z., Duller, G.A.T., Wintle, A.G., Henshilwood, C.S., 2006. Extending thechronology of deposits at Blombos Cave, South Africa, back to 140 ka usingoptical dating of single and multiple grains of quartz. J. Hum. Evol. 51, 255e273.
Jerardino, A., 1997. Changes in shellfish species composition and mean shell sizefrom a Late-Holocene record of the west coast of southern Africa. J. Archaeol.Sci. 24, 1031e1044.
Jerardino, A., 2003. Precolonial settlement and subsistence along sandy beachessouth of Elands Bay, west coast, South Africa. S. Afr. Archaeol. Bull. 58, 53e62.
Jerardino, A., Horwitz, L., Mazel, A., Navarro, R., 2009. Just before Van Riebeeck:glimpses into terminal LSA lifestyle at Connies Limpet Bar, West Coast of SouthAfrica. S. Afr. Archaeol. Bull. 64, 75e86.
Jerardino, A., Parkington, J., 1993. New evidence for whales on archaeological sitesin the south-western Cape. S. Afr. J. Sci. 89, 6e7.
Kilburn, R., Rippey, E., 1982. Sea Shells of Southern Africa. Macmillan, Johannesburg.Klein, R.G., 1979. Stone age exploitation of animals in southern Africa. Am. Sci. 67,
151e160.Klein, R.G., 1989. The Human Career: Human Biological and Cultural Origins, first ed.
The University of Chicago Press, Chicago.Klein, R.G., Avery, G., Cruz-Uribe, K., Halkett, D., Parkington, J.E., Steele, T.,
Volman, T.P., Yates, R., 2004. The Ysterfontein 1 Middle Stone Age site, SouthAfrica, and early human exploitation of coastal resources. Proc. Natl. Acad. Sci.USA 101, 5708e5715.
Klein, R.G., Steele, T., 2008. Gibraltar data are too sparse to inform on Neanderthalexploitation of coastal resources. Proc. Natl. Acad. Sci. USA 105 (51), E115.
Marean, C.W., 2010. Pinnacle Point Cave 13B (Western Cape Province, South Africa)in context: The Cape Floral Kingdom, Shellfish, and Modern Human Origins.J. Hum. Evol. 59 (3e4), 425e443.
Marean, C.W., Assefa, Z.,1999. Zooarchaeological evidence for the faunal exploitationbehavior of Neandertals and Early Modern Humans. Evol. Anthropol. 8, 22e37.
Marean, C.W., Bar-Matthews, M., Bernatchez, J., Fisher, E., Goldberg, P., Herries, A.,Jacobs, Z., Jerardino, A., Karkanas, P., Mercier, N., Minichillo, M., Nilssen, P.J.,Thompson, E., Tribolo, C., Valladas, H., Williams, H., 2007. Early human use ofmarine resources and pigment in South Africa during the Middle Pleistocene.Nature 449, 905e908.
Marean, C.W., Bar-Matthews, M., Fisher, E., Goldberg, P., Herries, A.I.R., Karkanas, P.,Nilssen, P.J., Thompson, E., 2010. The stratigraphy of the Middle Stone Agesediments at Pinnacle Point Cave 13B (Mossel Bay, Western Cape Province,South Africa). J. Hum. Evol. 59 (3e4), 234e255.
Marean, C.W., Nilssen, P.J., Brown, K., Jerardino, A., Stynder, D., 2004. Paleoanthro-pological investigations of Middle Stone Age sites at Pinnacle Point, Mossel Bay(South Africa): archaeology and hominid remains from the 2000 field season.J. Paleoanthropol. 2, 14e83.
McBrearty, S., Brooks, A., 2000. The revolution that wasn’t: a new interpretation ofthe origin of modern human behaviour. J. Hum. Evol. 39, 453e563.
McLachlan, A., Hanekom, N., 1979. Aspects of the biology, ecology and seasonalfluctuations in biochemical composition of Donax serra in the East Cape. S. Afr. J.Zool. 14, 183e193.
McLachlan, A., Wooldridge, T., van der Horst, G., 1979. Tidal movements of themacrofauna on an exposed sandy beach in South Africa. J. Zool. Lond. 188,433e442.
Orford, J., 1987. Coastal processes: the coastal response to sea-level variation. In:Devoy, R.J.N. (Ed.), Sea Surface Studies: a Global View. Croom Helm, pp.415e463.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424 423

Author's personal copy
Parkington, J., 2003. Middens and moderns: shellfishing and the Middle Stone Ageof the Western Cape, South Africa. S. Afr. J. Sci. 99, 243e247.
Parkington, J., Poggenpoel, C., 1987. Diepkloof rock shelter. In: Parkington, J., Hall, M.(Eds.), Papers in the Prehistory of the Western Cape, South Africa. BritishArchaeological Reports International Series 332(ii), pp. 269e293. Oxford.
Reese, D.S., 1982. Marine and fresh-water mollusks from the Epipaleolithic site ofHayonim Terrace, Western Galilee, northern Israel, and other east Mediterra-nean sites. Paleorient 8, 83e90.
Reese, D.S., 1989. On Cassid lips and helmet shells. Bull. Am. Schools Orient. Res.275, 33e39.
Reese, D., 1998. Middle Palaeolithic shells from Ras el-Kelb. In: Copeland, L.,Moloney, N. (Eds.), The Mousterian Site of Ras El-Kelb, Lebanon, Oxford, p. 67.
Reese, D.S., 1999. Marine invertebrates. In: Simmons, A.H. (Ed.), Faunal Extinction inan Island Society: Pygmy Hippopotamus Hunters of Cyprus. Kluwer Academic/Plenum Publishers, New York, pp. 188e191.
Reese, D.S., 2005. The Çatalhöyük shells. In: Hodder, I. (Ed.), Inhabiting Çatalhöyük:Reports from the 1995e1999 Seasons. McDonald Institute Monographs, Cam-bridge, pp. 123e127.
Ridgway, S.A., Reid, D.G., Taylor, J.D., Branch, G.M., Hodgson, A.N., 1998. A cladisticphylogeny of the family Patellidae (Mollusca: Gastropoda). Phil. Trans. Roy. Soc.Lond. B 353, 1645e1671.
Robey, T.S., 1987. The stratigraphic and cultural sequence at Tortoise Cave, Verlor-envlei. In: Parkington, J., Hall, M. (Eds.), Papers in the Prehistory of the WesternCape, South Africa. British Archaeological Reports International Series 332(ii),pp. 294e325. Oxford.
Schweitzer, F.R., 1979. Excavations at Die Kelders, Cape Province, South Africa.Annls. S. Afr. Mus. 78, 101e233.
Schweitzer, F.R., Wilson, M.L., 1982. Byneskranskop 1: a Late Quaternary living sitein the southern Cape Province, South Africa. Annls. S. Afr. Mus. 88, 1e203.
Sealy, J., Maggs, T., Jerardino, A., Kaplan, J., 2002. Excavations at Melkbosstrand: vari-abilityamongherder sitesonTableBay, SouthAfrica.S.Afr.Archaeol. Bull. 59,17e28.
Siegfried,W.R.,1977.Mussel-droppingbehaviourofKelpGulls. S.Afr. J. Sci. 73,337e341.Siegfried, W.R., Hockey, P.A.R., Crowe, A.A., 1985. Exploitation and conservation of
brown mussel stocks by coastal people of Transkei. Environ. Conserv. 12,303e307.
Smith, A.B., 2006. Excavations at Kasteelberg, and the Origins of the Khoekhoen inthe Western Cape, South Africa. British Archaeological Reports S1537 Oxford.
Speed, E., 1969. Prehistoric shell collectors. S. Afr. Archaeol. Bull. 24, 193e196.
Steele, T., Klein, R.G., 2005. Mollusk and tortoise size as proxies for stone agepopulation density in South Africa: implications for the evolution of humancultural capacity. Munibe 57, 5e21.
Stiner, M.C., 1993. Small animal exploitation and its relation to hunting, scavenging,and gathering in the Italian Mousterian. In: Peterkin, G.L., Bricker, H.M.,Mellars, P. (Eds.), Hunting and Animal Exploitation in the Later Palaeolithic andMesolithic of Eurasia. Papers of the American Anthropological Association, vol.4, pp. 101e119. Washington, D.C.
Stiner, M., 1994. Honor Among Thives: a Zooarchaeological Study of NeandertalEcology. Princeton University Press, Princeton.
Stringer, C.B., Finlayson, J.C., Barton, R.N.E., Cáceres, I., Fernández-Jalvo, Y., 2008b.Reply to Klein and Steele: Neanderthals and their South African contempo-raries. Proc. Natl. Acad. Sci. USA 105 (51), E116.
Stringer, C.B., Finlayson, J.C., Barton, R.N.E., Fernández-Jalvo, Y., Cáceres, I.,Sabin, R.C., Rhodes, E.J., Currant, A.P., Rodríguez-Vidal, J., Giles-Pacheco, F.,Riquelme-Cantal, J.A., 2008a. Neanderthal exploitation of marine mammals.Proc. Natl. Acad. Sci. USA 105 (38), 14319e14324.
Taborin, Y., 2003. La mer et les premiers hommes modernes. In: Vandermeersch, B.(Ed.), Échanges et Diffusion Dans la Préhistroire Méditerranéenne. Editions duComité des Travaux Historiques et Scientifiques, Paris, pp. 113e122.
Thackeray, F., 1988. Molluscan fauna from Klasies River, South Africa. S. Afr.Archaeol. Bull. 43, 27e32.
Van Erkom Schurink, C., Griffiths, C., 1990. Marine mussels of southern Africadtheirdistribution patterns, standing stocks, exploitation and culture. J. Shellfish Res.9, 75e85.
Vanhaeren, M., d’Errico, F., Stringer, C., James, S.L., Todd, J.A., Mienis, H.K., 2006.Middle Palaeolithic shell beads in Israel and Algeria. Science 312, 1785e1788.
Voigt, E.A., 1973. Stone age molluscan utilization at Klasies River Mouth Caves. S.Afr. J. Sci. 69, 306e309.
Voigt, E.A., 1982. The molluscan fauna. In: Singer, R., Wymer, J. (Eds.), The MiddleStone Age at Klasies River Mouth in South Africa. University of Chicago Press,Chicago, pp. 155e186.
Volman, T.P., 1978. Early archaeological evidence for shellfish collecting. Science201, 911e913.
Waelbroeck, C., Labeyrie, L., Michel, E., Duplessy, J.C., McManus, J.F., Lambeck, K.,Balbon, E., Labracherie, M., 2002. Sea-level and deep water temperaturechanges derived from benthic foraminifera isotopic records. Quatern. Sci. Rev.21, 295e305.
A. Jerardino, C.W. Marean / Journal of Human Evolution 59 (2010) 412e424424
Copyright © 2022 FDOKUMEN




















