
FLUOXETINE ALTERS MU OPIOID RECEPTOR EXPRESSION IN OBESE ZUCKER RAT EXTRAHYPOTHALAMIC REGIONS
10
289 Intern. J. Neuroscience, 116:289–298, 2006 Copyright 2006 Taylor & Francis Group, LLC ISSN: 0020-7454 / 1543-5245 online DOI: 10.1080/00207450500403231 FLUOXETINE ALTERS MU OPIOID RECEPTOR EXPRESSION IN OBESE ZUCKER RAT EXTRAHYPOTHALAMIC REGIONS ITZIAR CHURRUCA MARÍA P. PORTILLO Department of Nutrition and Food Science Faculty of Pharmacy University of the Basque Country Vitoria (Spain) JOSÉ MARÍA ZUMALABE Department of Personality, Evaluation and Psychological Treatments Faculty of Pharmacy University of the Basque Country Vitoria (Spain) MARÍA T. MACARULLA Department of Nutrition and Food Science Faculty of Pharmacy University of the Basque Country Vitoria (Spain) Received 10 February 2005. This work has been supported by a Basque Government grant (GV PI-1999-51) and a 1/UPV/ EHU 00081.327-E-15320/2003 grant. Itziar Churruca is a doctoral fellowship from the Basque Gov- ernment. The authors thank Prof. J. C Pérez Cobo for his bibliographical revision, and J. M. Rodríguez Robledo for his technical assistance. Address correspondence to Itziar Churruca, Department of Nutrition and Food Science, School of Pharmacy, University of the Basque Country, Paseo de la Universidad 7, 01006 Vitoria. Spain. E-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of FLUOXETINE ALTERS MU OPIOID RECEPTOR EXPRESSION IN OBESE ZUCKER RAT EXTRAHYPOTHALAMIC REGIONS

289
Intern. J. Neuroscience, 116:289–298, 2006Copyright 2006 Taylor & Francis Group, LLCISSN: 0020-7454 / 1543-5245 onlineDOI: 10.1080/00207450500403231
FLUOXETINE ALTERS MU OPIOID RECEPTOREXPRESSION IN OBESE ZUCKER RATEXTRAHYPOTHALAMIC REGIONS
ITZIAR CHURRUCAMARÍA P. PORTILLO
Department of Nutrition and Food ScienceFaculty of PharmacyUniversity of the Basque CountryVitoria (Spain)
JOSÉ MARÍA ZUMALABE
Department of Personality, Evaluationand Psychological TreatmentsFaculty of PharmacyUniversity of the Basque CountryVitoria (Spain)
MARÍA T. MACARULLA
Department of Nutrition and Food ScienceFaculty of PharmacyUniversity of the Basque CountryVitoria (Spain)
Received 10 February 2005.This work has been supported by a Basque Government grant (GV PI-1999-51) and a 1/UPV/
EHU 00081.327-E-15320/2003 grant. Itziar Churruca is a doctoral fellowship from the Basque Gov-ernment. The authors thank Prof. J. C Pérez Cobo for his bibliographical revision, and J. M. RodríguezRobledo for his technical assistance.
Address correspondence to Itziar Churruca, Department of Nutrition and Food Science, Schoolof Pharmacy, University of the Basque Country, Paseo de la Universidad 7, 01006 Vitoria. Spain.E-mail: [email protected]

290 I. CHURRUCA ET AL.
LAURA SÁENZ DEL BURGOJON ZARATEENRIQUE ECHEVARRÍA
Department of PhysiologyFaculty of Pharmacy, University of the Basque CountryVitoria (Spain)
The aim of this article was to describe the effects of chronic fluoxetine on muopioid receptor expression in obese Zucker rat extrahypothalamic regions. Maleobese Zucker (fa/fa) rats were administered with fluoxetine (10 mg/kg; i.p.)daily for two weeks. Brain regional immunostaining for mu opioid receptorwas carried out. An increase in the numbers of neural cells immunostained formu opioid receptor in caudatus-putamen, dentate gyrus, lateral septum, amygdala,and frontal, parietal, and piriform cortices was observed. Increased mu opioidreceptor expression in the central amygdaloid nuclei suggests a decreased opioidergictone at this level that could be involved in fluoxetine anorectic action.
Keywords amygdale, anorexia, fluoxetine, mu opioid receptor, obese Zuckerrat
INTRODUCTION
Mu opioid receptors are widely distributed throughout several regions of thecentral nervous system, such as the basal ganglia, limbic structures and fron-tal, parietal, and temporal cortices (Ding et al., 1996; Kaneko et al., 1995;Hiller et al., 1994). Thus, opioid peptides can mediate pleasure, reward, andemotions in rodents through its action on opioid receptors (Belluzi & Stein,1977).
Opioid neural circuits play a physiological role in the control of feeding(Yoemans & Gray, 2002). In fact, it has been described that administrationof mu opioid receptor agonists in the hypothalamic paraventricular nucleusincreases food intake in rats (Koch et al., 1995). Moreover, in several centralnervous system regions, such as brainstem, nucleus accumbens, ventral pallidum,and amygdala, opioids have been suggested to be involved in food-inducedreward (Berridge, 1996). Thus, enhancement of hedonic palatability of foodhas been proposed to be on the basis of opioid agonists-induced hyperphagia,when these drugs are administered centrally (Pecina & Berridge, 1995).
An interaction of antidepressant drugs with the opioidergic system hasbeen suggested (Valverde et al., 1994). Thus, alterations in mu opioid receptor

FLUOXETINE AND OPIOIDS 291
expression (Gandarias et al., 1998) and subsequent antinociceptive effects(De Felipe et al., 1986; Eschalier e tal., 1981) in rats have been reported afterchronic imipramine treatment (Humel et al., 1994; Tura & Tura, 1990), andit has been described that imipramine changes enkephalin levels in rat stria-tum (De Felipe et al., 1985) and enhances enkephalin release from striatalnerve endings (Kurumaji et al., 1988). However, in the case of selectiveserotonin reuptake inhibitors, it has been reported that fluoxetine treatmentgenerates an increase in the density of neurons expressing mu opioid recep-tors in several rat prosencephalic regions, suggesting a pharmacological ac-tion on opioid neural circuits (Gandarias et al., 1999).
Whole brain decreased concentrations of mu opioid receptors have beenreported in obese Zucker rats, suggesting the involvement of opioid neuralcircuits in hypersensitivity to painful stimuli and resistance to morphine anal-gesic effect, showed by this genetic obesity model (Roane et al., 1988). Moreover,it has been suggested that endogenous opioids could contribute to hyperph-agia and obesity in obese Zucker rats (Marin-Bivens & Olster, 1999). In fact,both lean and obese Zucker rats display weight loss and reduced intake fol-lowing chronic central administration of opioid receptor antagonists (Cole etal., 1997).
Chronic fluoxetine administration increases the numbers of neural cellspositively immunostained for mu opioid receptor in several hypothalamicregions in obese Zucker rats, suggesting the involvement of hypothalamicopioid system in fluoxetine-induced anorexia (Churruca et al., 2004), little isknown concerning the effects of this drug on opioid extrahypothalamic neu-ral circuits in genetically obese animal models. This being the case, thisstudy focused on describing the effects of chronic fluoxetine administrationon regional prosencephalic mu opioid receptor immunostaining, in obese Zuckerrats.
MATERIALS AND METHODS
Male obese Zucker (fa/fa) rats were obtained from CRIFFA (Barcelona, Spain)and maintained in a temperature (22 ± 2 °C) and humidity (50%)-controlledroom with 12 h-12 h light-dark cycle (lights on at 08:00 h) and given freeaccess to standard rat chow diet (pelleted) and water. Rats were carefullyhandled and all experiments were carried out in accordance with the law,avoiding animal suffering. Both treated and control animals were observed inorder to detect fluoxetine-induced behavioural changes. Rats assigned to fluoxetinetreatment (fluoxetine hydrochloride, Lilly, Madrid, Spain) were subjected to

292 I. CHURRUCA ET AL.
daily intraperitoneal injections (10 mg/kg) for two weeks. The control groupwas given 0.9% NaCl solution in the same volume and duration as the fluoxetine-treated group. Twenty-four h after the last treatment, experimental animalswere anaesthetized intraperitoneally with 15% choral hydrate (1.2 mg/kg)(Sigma, St. Louis, MO) and perfused transcardially under deep anaesthesiawith saline plus 50 mM phosphate buffer, pH 7.4, followed by 4% paraform-aldehyde (Sigma, St. Louis, MO). The brains were removed, cut into smallerpieces, and then immersed in the same fixative medium overnight. Theywere stored for 2 days in 0.1 M phosphate buffer containing 30% sucrose at4ºC. Fifty micrometer sections were cut using a cryostatic microtome (Cryocut3.000, Leica España, Barcelona, Spain) with a stereotaxic atlas guide andimmunostained for mu opioid receptor with polyclonal antisera raised in rab-bits. The antigens were detected by the avidin-peroxidase technique, using 3,3'-diaminobenzidine as chromogen (Sigma, St. Louis, MO). Following re-duction of endogenous peroxidases with 1% hydrogen peroxide (Sigma, St.Louis, MO) and blocking of nonspecific background staining with 5% nor-mal goat serum (NGS) (Sigma, St. Louis, MO), the sections were incubatedwith the following immunoreagents: 1-primary antisera: rabbit anti mu opidreceptor (Chemicon International Inc., Temecula, CA), a commercially ob-tained polyclonal antibodiy raised in rabbits (dilution 1:1,000); 2-goat antirabbit immunoglobulin (goat anti rabbit biotinilated), Chemicon InternationalInc., Temecula, CA, dilution 1:200; 3-avidin-peroxidase complex: strept ABCcomplex HRP (Dako A/S, Glostrup, Denmark), dilution 1:300; 4-chromogen:3, 3´-diaminobenzidine (Sigma, St. Louis, MO), 0.3 mg/mL in 0.2 M TrisHCl buffer containing 0.03% hydrogen peroxide. Each step was followed byan appropriate wash per triplicate in phosphate buffer saline and 0.3% TritonX-100 (Sigma, St. Louis, MO) was used. Sections were carefully extended,dehydrated and mounted (DPX mountant for histology, Fluka Chemie AG,Buchs, Switzerland), and examined with an Olympus BX50F optic micro-scope (Olympus Optical Co. Ltd., Japan). The selected prosencephalic re-gions were the caudatus-putamen (CP), the dentate gyrus (DG), the lateralseptum (LS), the amygdala (AM), and the frontal cortex (F), parietal cortex(P), and piriform cortex (Pir). Counts of positively immunostained neural cellbodies were made, in order to calculate cell density (expressed as stainedcells/mm2) in every selected extrahypothalamic region. Data were expressedas mean ± SEM. Statistical analysis was performed using SPSS 8.0 (SPSSInc. Michigan, USA). Mann-Whitney test was used for comparisons betweenboth groups. p values < .05 were considered statistically significant.
FLUOXETINE AND OPIOIDS 293
RESULTS
Utilization of a specific polyclonal antibody against mu opioid receptors showedthe presence of round cells intensely immunostained in the frontal, parietal,and piriform cortices of the rat brain. Stained cell bodies were also present inseveral regions, such as the amygdala, the dentate gyrus, the lateral septum,and the caudatus-putamen. However, the authors did not appreciate the pres-ence of stained fibers in these regions.
Fluoxetine was well tolerated in all cases and a lack of fluoxetine-in-duced behavioral changes was observed in treated animals, with respect tocontrols.
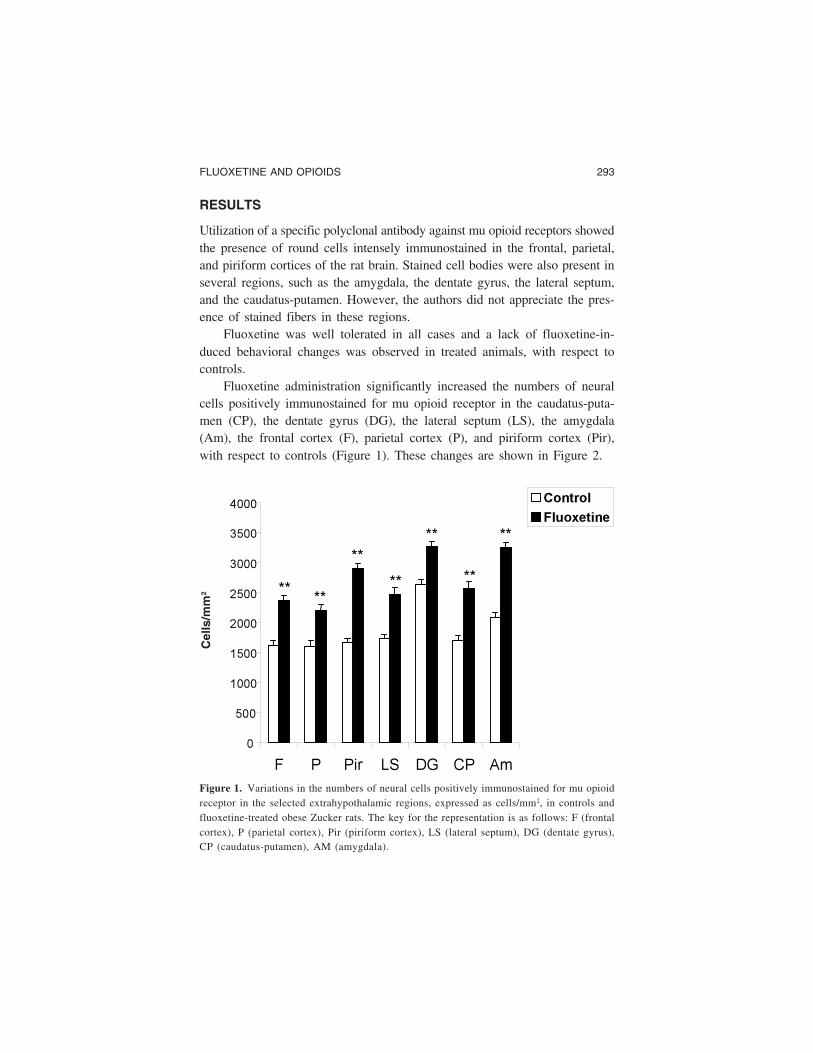
Fluoxetine administration significantly increased the numbers of neuralcells positively immunostained for mu opioid receptor in the caudatus-puta-men (CP), the dentate gyrus (DG), the lateral septum (LS), the amygdala(Am), the frontal cortex (F), parietal cortex (P), and piriform cortex (Pir),with respect to controls (Figure 1). These changes are shown in Figure 2.
0
500
1000
1500
2000
2500
3000
3500
4000
F P Pir LS DG CP Am
Control
Fluoxetine
**
**
**
**
**
**
**
Cel
ls/m
m2
Figure 1. Variations in the numbers of neural cells positively immunostained for mu opioidreceptor in the selected extrahypothalamic regions, expressed as cells/mm2, in controls andfluoxetine-treated obese Zucker rats. The key for the representation is as follows: F (frontalcortex), P (parietal cortex), Pir (piriform cortex), LS (lateral septum), DG (dentate gyrus),CP (caudatus-putamen), AM (amygdala).

294 I. CHURRUCA ET AL.
Figure 2. Light micrographs showing mu opioid receptor positively immunostained neuralcells in the frontal cortex, in controls (A) and fluoxetine-treated (B) obese Zucker rats.10×. (See Color Plate VII at end of issue.)
(A)
(B)

FLUOXETINE AND OPIOIDS 295
DISCUSSION
Orexigenic effects of opioid neural circuits, which are involved in the physi-ological regulation of ingestive behavior (Glass et al., 1999), as well as thepreferential increase in the intake of highly palatable food induced by muopioid agonists in rodents, are well known. In fact, opioid neural circuits incentral amygdaloid nuclei are involved in the regulation of feeding related toalimentary preferences and food-induced rewarding (Berridge, 1996; Glass etal., 1999). It must be pointed out that mu opioid receptor agonist local ad-ministration in central amygdaloid nuclei increases feeding, due in part toactivation of neurons in the shell of the nucleus accumbens (Levine et al.,2004). Thus, opioid receptor modulation of a metabolically sensitive ion channelin rat amygdala neurons has been reported (Chen et al., 2001).
Mu opioid agonist intracerebral injection in central amygdaloid nucleiboth increases food intake and mu opioid receptor internalization at this levelin rats, due to activation of mu opioid receptor expressing neural cells in thisextrahypothalamic prosencephalic region (Levine et al., 2004). It has beenalso described that diet-induced obesity in rats increases mu opioid receptorbinding in several amygdaloid regions, suggesting a decreased release ofendogenous opioid peptides, probably due to increased leptin and insulinserum levels (Smith et al., 2002).
Fluoxetine, a selective serotonin reuptake inhibitor (Wong et al., 1995)and a widely used antidepressant drug (Mackay et al., 1997), can alter bodyweight (Goldstein et al., 1997) and increases the density of neurons express-ing mu opioid receptors in several rat prosencephalic regions (Gandarias etal., 1999), suggesting a pharmacological action on prosencephalic opioid neuralcircuits.
Obese Zucker rats are hyperleptinemic and show an hyperphagic eatingpattern, which is similar to that displayed by obese binge eaters (Jarosz &Metzger, 2002). Weight loss and reduced food intake after central adminis-tration of opioid receptor antagonists has been reported in this well-charac-terized obesity model (Cole et al., 1997; Mitch et al., 1993), suggesting theinvolvement of opioidergic neural circuits in hyperphagia (19). Moreover, ithas been demonstrated that obese female Zucker rats show decreased con-centrations of mu opioid receptors in whole brain, that could partially explaintheir hypersensitivity to painful stimuli and their resistance to the analgesiceffects of morphine (Roane et al., 1988).
In obese Zucker rats, a significant increase in mu opioid receptor expres-sion in several hypothalamic regions after chronic fluoxetine treatment has

296 I. CHURRUCA ET AL.
been reported (Churruca et al., 2004), suggesting a role for opioids in fluoxetine-induced anorexia in this genetically obese and hyperphagic model.
Accordingly, in the results reported here, chronic fluoxetine treatment inobese Zucker rats generated a statistically significant increase in the densityof neurons expressing mu opioid receptors in several regions, such as piri-form, parietal and frontal cortices, amygdala, caudatus-putamen, lateral septumand dentate gyrus, with respect to controls, suggesting a pharmacologicalaction of fluoxetine on mu opioid receptor function in obese Zucker rat extra-hypothalamic regions.
Fluoxetine decreases food intake and increases mu. opioid receptor ex-pression in the central amygdaloid nuclei of obese Zucker rats, suggesting adecreased opioidergic tone at this level that could be involved in fluoxetineanorectic action.
REFERENCES
Belluzi, J. D., & Stein, L. L. (1977). Enkephalin may mediate euphoria and drive-reduction reward. Nature, 266, 556–558.
Berridge, K. C. (1996). Food reward: brain substrates of wanting and liking. Neuro-science Biobehavioral Review, 20(1), 1–25.
Chen, X., Marrero, H. G., & Freedman, J. E. (2001). Opioid receptor modulation ofa metabolically sensitive ion channel in rat amygdala neurons. Journal of Neuro-science, 21(23), 9092–90100.
Churruca, I., Casis, L., Portillo, M. P., Macarulla, M. T., Zárate, J., Pascual, J., &Echevarria, E. (2004). Fluoxetine alters mu opioid receptor expression in obeseZucker rat hypothalamus. Neurosci Res Comm. 35(1) (in press). Published Online:17 Aug 2004.
Cole, J. L., Berman, N., & Bodnar, R. J. (1997). Evaluation of chronic opioid recep-tor antagonist effects upon weight and intake measures in lean and obese Zuckerrats. Peptides, 18(8), 1201–1207.
De Felipe, M. C., De Ceballos, M. L., Gil, C., & Fuentes, J. A. (1985). Chronicantidepressant treatment increases enkephalin levels in nucleus accumbens andstriatum of the rat. European Journal of Pharmacology, 112, 119–122.
De Felipe, M. C., De Ceballos, M. L., Gil, C., & Fuentes, J. A. (1986). Hypoalgesiainduced by antidepressant in mice: A case for opioids and serotonin. EuropeanJournal of Pharmacology, 125, 193–199.
Ding, Y. Q., Kaneko, T., Nomura, S., & Mizuno, N. (1996). Immunohistochemicallocalization of mu-opioid receptors in the central nervous system of the rat. Journalof Comp Neuroloyg, 367(3), 375–402.
Eschalier, A., Montasturc, J. L., Devoize, J. L., Rigal, F., Gaillard-Plaza, G., & Pechadre,

FLUOXETINE AND OPIOIDS 297
J. C. (1981). Influence of naloxone and metisergide on the analgesic effect ofclomipramine in rats. European Journal of Pharmacology, 76, 1–7.
Gandarias, J. M., Echevarría, E., Acebes, I., Silió, M., & Casis, L. (1998). Effects ofimipramine administration on mu-opioid receptor immunostaining in the rat fore-brain. Arzneim-Forsch/Drug Research, 48 (II), 717–719.
Gandarias, J. M., Echevarría, E., Acebes, I., Abecia, L. C., Casis, O., & Casis, L.(1999). Effects of fluoxetine administration on mu-opioid receptor immunostainingin the rat forebrain. Brain Research, 817, 236–240.
Glass, M. J., Billington, C. J., & Levine, A. S. (1999). Opioids and food intake:Distributed functional neural pathways? Neuropeptides, 33(5), 360–368.
Goldstein, D. J., Hamilton, S. H., Masica, D. N., & Beasley, C. M. (1997). Fluoxetinein medically stable, depressed geriatric patients: Effect on weight. Journal ofClinical Psychopharmacology, 17, 365–369.
Hiller, J. M., Zhang, Y., Bing, T., Gioannini, T. L., Stone, E. A., & Simon, E. J.(1994). Immunohistochemical localization of mu-opioid receptors in rat brainusing antibodies generated against a peptide sequence present in a purified mu-opioid binding protein. Neuroscience, 62, 829–841.
Humel, T., Humel, C., Friedel, Y., Pauli, E., & Kobal, G. (1994). A comparison ofthe antinociceptive effects of imipramine, tramadol and anpirtoline. British Journalof Clinical Pharmacology, 37, 325–333.
Jarosz, P. A., & Metzger, B. L. (2002). The effect of opioid antagonism on foodintake behavior and body weight in a biobehavioral model of obese binge eat-ing. Biological Research Nurs, 3(4), 198–209.
Kaneko, T., Minami, M., Satoh, M., & Mizuno, N. (1995). Immunocytochemical lo-calization of mu-opioid receptor in the rat caudate-putamen. Neuroscience Letters,184(3), 149–152.
Koch, J. E., Glass, M. J., Cooper, M. L., & Bodnar, R. J. (1995). Alterations indeprivation, glucoprivic and sucrose intake following general, mu and kappa opioidantagonists in the hypothalamic paraventricular nucleus of rats. Neuroscience,66(4), 951–957.
Kurumaji, A., Mitsushio, H., & Takashima, M. (1988). Chronic dietary treatment withantidepressants decrease brain met-enkephalin-like immunoreactivity in the rat.Psychopharmacology, 94, 188–192.
Levine, A. S., Olszewski, P. K., Mullett, M. A., Pomonis, J. D., Grace, M. K., Kotz,C. M., & Billington, C. J. (2004). Intra-amygdalar injection of DAMGO: Effectson c-Fos levels in brain sites associated with feeding behavior. Brain Research,1015(1–2), 9–14.
Mackay, F. J., Dunn, N R., Wilton, L V., Pierce, G L., Freemantle, S. N., & Mann,R. D. (1997). A comparison of fluvoxamine, fluoxetine, sertraline and paroxetineexamined by observational cohort studies. Pharmacoepidemiol Drug Safe, 6, 235–246.

298 I. CHURRUCA ET AL.
Marin-Bivens, C. L., & Olster, D. H. (1999). Opioid receptor blockade promotes weightloss and improves the display of sexual behaviors in obese Zucker female rats.Pharmacol Biochem Behav, 63(3), 515–520.
Mitch, C. H., Leander, J. D., Mendelsohn, L. G., Shaw, W. N., Wong, D. T., Cantrell,B. E., Johnson, B. G., Reel, J. K., Snoddy, J. D., & Takemori, A. E. (1993). 3,4-Dimethyl-4-(3-hydroxyphenyl)piperidines: Opioid antagonists with potent anorectantactivity. Journal of Med Chem, 36(20), 2842–2850.
Pecina, S., & Berridge, K. C. (1995). Central enhancement of taste pleasure by intra-ventricular morphine. Neurobiology, 3, 269–280.
Roane, D. S., Iadarola, M. J., & Porter, J. R. (1988). Decreased [3H]-naloxone bind-ing and elevated dynorphin-A(1-8) content in Zucker rat brain. Physiol Behav43(3), 371–374.
Smith, S. L., Harrold, J. A., & Williams, G. (2002). Diet-induced obesity increasesmu opioid receptor binding in specific regions of the rat brain. Brain Research,953(1–2), 215–222.
Tura, B., & Tura, S. (1990). The analgesic effect of tryciclic antidepressants. BrainResearch, 518, 19–22.
Valverde, O., Micó, J. A., Maldonado, R., Mellado, M., & Gibert-Rahola, J. (1994).Participation of opioid and monoaminergic mechanisms on the antinociceptiveeffect induced by tricyclic antidepressants in two behavioural pain test in mice.Prog Neuropsychopharmacol Biol Psychiatry, 18, 1073–1092.
Wong, D. T., Bymaster, F P., & Englemam, E. A. (1995). Prozac (fluoxetine, Lilly110140), the first selective serotonin uptake inhibitor and an antidepressant drug:twenty years since its first publication. Life Science, 57, 411–441.
Yeomans, M. R., & Gray, R. W. (2002). Opioid peptides and the control of humaningestive behaviour. Neuroscience Biobehavioral Reviews, 26, 713–728.




















