
Fluorescence recovery after photobleaching reveals that LPS rapidly transfers from CD14 to hsp70 and...
12
Erratum Fluorescence recovery after photobleaching reveals that LPS rapidly transfers from CD14 to hsp70 and hsp90 on the cell membrane Triantafilou, K., Triantafilou, M., Ladha, S., Mackie, A., Fernandez, N., Dedrick, R. L. and Cherry, R. (2001). J. Cell. Sci. 114, 2535-2545 The order of authors of the paper is incorrect in the printed version. The correct list of authors is shown below and in the the online versions Kathy Triantafilou, Martha Triantafilou, Shab Ladha, Alan Mackie, Russell L. Dedrick, Nelson Fernandez and Richard Cherry.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Fluorescence recovery after photobleaching reveals that LPS rapidly transfers from CD14 to hsp70 and...

Erratum
Fluorescence recovery after photobleaching reveals that LPS rapidly transfers from CD14 to hsp70 andhsp90 on the cell membraneTriantafilou, K., Triantafilou, M., Ladha, S., Mackie, A., Fernandez, N., Dedrick, R. L. and Cherry, R. (2001). J. Cell. Sci. 114,2535-2545
The order of authors of the paper is incorrect in the printed version. The correct list of authors is shown below and in thethe online versions
Kathy Triantafilou, Martha Triantafilou, Shab Ladha, Alan Mackie, Russell L. Dedrick, Nelson Fernandez and RichardCherry.

INTRODUCTION
The recognition of bacterial lipopolysaccharide (LPS) orendotoxin by the innate immune system provokes a stronginflammatory response, triggering the production and releaseof potent inflammatory mediators. Although Gram-positiveand Gram-negative bacteria differ in the composition of theirouter cell wall, the host reactions to them are similar. In asevere bacterial infection caused by either form of bacterium,inflammatory mediators are produced and released in excessand can lead to haemodynamic shock, accompanied bysymptoms such as fever, tachycardia, tachypnoea and possiblyending in multiple organ failure and sometimes death (Bone,1991).
The outer cell wall of Gram-negative bacteria, which iscomposed of LPS, appears to be the cause of the immuneactivation. Our current understanding of the innate immunerecognition of Gram-negative bacteria is based on the seminaldiscovery of LPS-binding protein (LBP) (Tobias et al., 1986)and the elucidation of its structure and function (Wright et al.,1989; Schumann et al., 1990), followed by the identification ofCD14 as an LPS receptor (Wright et al., 1990). CD14 existsas a membrane bound protein (mCD14) and also as a soluble
form (sCD14) (Durieux et al., 1994) that binds LPS.Complexes of soluble CD14 (sCD14) and LPS bind to a yetunknown receptor and activate cells of non-myeloid lineage(Frey et al., 1992; Labeta et al., 1993; Pugin et al., 1993; Leeet al., 1993; Demters et al., 1994). It has been suggested thatthe same signalling receptor might be involved in bothmCD14-LPS and sCD14-LPS complexes. CD14 has also beenimplicated in mediating inflammatory responses to bacterialpeptidoglycan (PGN) and lipoteichoic acid (LTA) (Weidemannet al., 1994; Gupta et al., 1996), which are the majorcomponents of the cell wall of Gram-positive bacteria.
However, because CD14 is a glycosylphosphatidylinositol(GPI)-linked protein and does not transverse the cellmembrane, it has been suggested that it cannot deliver a signalfor activation against LPS. It has been suggested that CD14mediates LPS responses by interacting with other signaltransducing molecules (Ingalls and Golenbock, 1995; Ingallset al., 1997; Pugin et al., 1994; Gegner et al., 1995). The theoryof the existence of multiple LPS receptors has been furtherstrengthened by several lines of evidence, which havedemonstrated that CD14 blocking monoclonal antibodies(mAbs) only partially inhibit LPS binding (Lynn et al., 1993;Shapira et al., 1994; Blondin et al., 1997; Troelstra et al., 1997;
2535
Although CD14 has been implicated in the immunerecognition of bacterial lipopolysaccharide (LPS) fromGram-negative bacteria and also peptidoglycan (PGN) andlipoteichoic acid (LTA) from the outer cell wall of Gram-positive bacteria, accumulating evidence has suggestedthe possible existence of other functional receptor(s). Inthis study, we have used fluorescence recovery afterphotobleaching (FRAP) in order to get the first dynamicpicture of the innate recognition of bacteria. We have foundthat the diffusion coefficient of CD14 remains unaffectedafter LPS ligation and that the diffusion coefficients ofFITC-LPS and FITC-LTA bound to cells differ from thatof CD14. Furthermore, FITC-LPS/LTA rapidly becomeimmobile when bound to cells, suggesting that FITC-LPS/LTA must briefly associate with CD14 in the initial
attachment process and rapidly move on to an immobilereceptor or to a complex of receptors. Further FRAPexperiments revealed that heat shock protein 70 (hsp70)and hsp90 are immobile in cell membranes, and antibodiesagainst them were found to block the transfer of LPS to theimmobile receptor and to inhibit interleukin 6 productionupon LPS stimulation. These experiments indicated thatLPS transfers from CD14 to hsp70 and hsp90, which maybe part of an LPS/LTA multimeric receptor complex. Thus,hsps are implicated as mediators of the innate activation bybacteria.
Key words: Endotoxin, Lipopolysaccharide, FRAP, Lateral diffusion,CD14
SUMMARY
Fluorescence recovery after photobleaching revealsthat LPS rapidly transfers from CD14 to hsp70 andhsp90 on the cell membraneKathy Triantafilou1,‡,§, Martha Triantafilou1, Shab Ladha2, Alan Mackie2, Russell L. Dedrick3, NelsonFernandez1 and Richard Cherry1
1University of Essex, Department of Biological Sciences, Central Campus, Wivenhoe Park, Colchester, CO4 3SQ, UK2Department of Food Biophysics, Institute of Food Research, Norwich Research Park, Colney Lane, Norwich, NR4 7UA, UK3Molecular Immunology, XOMA (US) LLC, 2910 Seventh Street, Berkeley, CA 94710, USA‡Present address: School of Biological Sciences, University of Portsmouth, King Henry Building, King Henry I Street, Portsmouth, PO1 2DY, UK§Author for correspondence (e-mail: [email protected])
Accepted 6 April 2001Journal of Cell Science 114, 2535-2545 (2001) © The Company of Biologists Ltd
RESEARCH ARTICLE

2536
Triantafilou et al., 2000c). Furthermore, several reports havesuggested the existence of LPS transducers, such asCD11c/CD18 (Ingalls and Golenbock, 1995), the humanhomologues of Toll (which are known as Toll-like receptor 2(TLR2) (Kirschning et al., 1998) and TLR4 (Poltorac et al.,1998)), moesin (Tohme et al., 1999) and, most recently, heatshock proteins (Byrd et al., 1999), suggesting that the innateimmune recognition of bacteria is highly complex.
Even though the exact mechanism whereby the innateimmune system recognizes bacteria remains to be elucidated,the acquired immune response is less of a mystery. Monks etal. and Grakoui et al. have provided the first dynamic pictureof the multimolecular choreography of receptors that takesplace as the T cell engages an antigen-presenting cell (APC)(Monks et al., 1998; Grakoui et al., 1999). This revealed themolecular interactions and microdomains involved in APC-Tcell engagement. It was shown that the APC-T cell contactzone, the immunological synapse, consisted of multiplereceptors, which moved to different domains throughout thesynapse while delivering different signals.
The innate immune recognition, being the more archaic formof host defence, lacks the complex interaction betweentwo different cells but still possesses the multimolecularinvolvement of a variety of proteins and receptors. In this study,we set out to investigate whether the receptors involved inthe innate bacterial recognition are also part of a compleximmunological choreography involving multiple receptors andmicrodomains. It is possible that the central molecule involvedin recognition of bacteria, CD14, being a GPI-linked protein,exists in detergent-insoluble glycolipid-enriched domains(DIGs) or GPI-microdomains, in which signalling moleculesmight accumulate following CD14 ligation by LPS.
In this study, fluorescence recovery after photobleaching(FRAP) was used in order to analyse the mobility of CD14 onthe cell membrane before and after LPS stimulation. We reportfor the first time that the diffusion coefficient of CD14 remainsvirtually unchanged before and after LPS stimulation.Furthermore, we find that FITC-LPS and FITC-LTA, whenbound to cells, have similar diffusion coefficients (which differfrom that of CD14) in a fraction of cells and are completelyimmobile in the remainder. The results suggest that FITC-LPS/LTA must initially bind to CD14 but are subsequentlyrapidly transferred to a transducing molecule or complex ofmolecules that is immobile. Here, we present evidence thatsuggests that these molecules are heat shock proteins (hsps) 70and 90. We find that these proteins are completely immobileon the plasma membrane and antibodies against them, whenadded prior to LPS stimulation, block the progressiveimmobilization of LPS and inhibit synthesis of interleukin 6(IL-6) and subsequent cell activation. We thus propose thathsp70 and hsp90 are implicated in cell activation, probably aspart of a multimeric transducing complex.
MATERIALS AND METHODS
MaterialsRough LPS from Salmonella minessota Re595 was purchased fromList Labs (Campbell, CA). All fine chemicals and human pooledserum were purchased from Sigma (St Louis, MO). Fluoresceinisothiocyanate (FITC) and OG were purchased from Molecular
Probes Europe (Leiden, The Netherlands). Hybridoma cells secreting26ic (anti-CD14) was obtained from the American Type CultureCollection (ATCC, Rockville, MD). Monoclonal antibody, MY4,directed against the functional domain of CD14 was obtained fromCoulter (San Francisco, CA). Recombinant human LBP (rLBP)(Theofan et al., 1994) and rsCD14 were provided by XOMA(Berkeley, CA) LLC. Hsp70 rabbit polyclonal serum was obtainedfrom Dako (Cambridge, UK). Hsp90-specific rabbit polyclonal serumwas obtained from Santa Cruz Biotechnology (Santa Cruz, CA). IL-6 enzyme-linked immunosorbent assay (ELISA) kit was obtainedfrom Diaclone (Besancon, France).
CellsChinese hamster ovary (CHO) cells transfected with hCD14 cDNA inthe expression vector pRc/RSV (CHO-CD14) were kindly provided byS. Viriyakosol and T. Kirkland (University of California) (Viriyakosoland Kirkland, 1995). CHO cells were maintained in DMEM/F12(Dulbecco’s Modified Eagle’s Medium (DMEM)/Ham’s F12 1:1 mix)from Gibco BRL with 2 mM L-glutamine, 7.5% foetal calf serum(FCS), 500 µg ml−1 gentamycin sulfate (G418, Sigma). Cells weregrown in 80 cm3 tissue culture flasks (Nunc). Trypsin/EDTA (0.05%trypsin/0.53 mM EDTA) was used for passaging the cells.
The MonoMac 6 cell line was obtained from the Institute ofImmunology, University of Munich, Germany. MonoMac 6 cells werecultured in 5% CO2 at 37°C in Iscove modified Dulbecco medium(IMDM; Gibco BRL) containing 10% FCS.
The human umbilical vein endothelial cell line (ECV-304) wasobtained from the ECACC (Cambridge, UK). The cells weremaintained in Medium 199 supplemented with Glutamax® (GibcoBRL) and 10% FCS. Cells were grown in 80 cm3 flasks (Nunc).Cultures of high viability were obtained by trypsinizing the cells in0.25% trypsin/EDTA, followed by seeding the cells at a density of1×105 cells per 25 cm3 flask.
For FRAP experiments, the cells were seeded in Lab-tek slides(Nunc) at a density of 10,000 cells per well and cultured for a further72 hours prior to the experiments.
Preparation of PE-Fab probeR-phycoerythrin (PE)-Fab probe was prepared as previously described(Smith et al., 1998; Triantafilou et al., 2000b). Briefly IgG waspurified and digested with papain. Aliquots of purified Fab fragmentswere labelled with PE, pyridyl disulphide derivative (MolecularProbes, Eugene, OR). Briefly, 1.25 mg of Fab was concentrated to5 mg ml−1 in a Centristart-1 microconcentrator (Sartorius). Thiol-reactive maleimide residues were introduced into Fab by incubatingwith 5 mM succinimidyl trans-4-(N-maleimidylmethyl) cyclohexane-1-carboxylate (SMCC) in dimethyl sulphoxide for 2 hours at roomtemperature. Excess SMCC was removed by extensive dialysis against100 mM phosphate buffer, 150 mM NaCl pH 7.5.
In parallel, 2.0 mg of R-phycoerythrin, pyridyl disulphide derivative(average 2.2 pyridyl residues) was incubated with 50 mMdithiothreitol (DTT) for 15 minutes at room temperature. Excess DTTwas eliminated by dialysis against 100 mM phosphate buffer, 150 mMNaCl pH 7.5. The PE was then incubated with the Fab for 20 hoursat 4°C in the dark. After this time, in order to prevent Fab disulphidereduction or aggregation, a 20 M excess of N-ethyl maleimide wasadded.
Purification of PE-labelled FabPreparative size exclusion chromatography was performed on a Bio-Rad 5000T HRLC controlled by a 486 PC. 300 µl labelled Fab wereloaded onto a Bio-Select SEC 250-5 column, eluted with 50 mMphosphate buffer, 150 mM NaCl pH 6.8 at 0.1 ml minute−1 and 200µl fractions collected. Integration was performed usingValueChrom integration analysis software (Bio-Rad). PE-Fabconcentration was determined using a molar extinction coefficient of1.96×10−6 M−1cm−1 for PE.
JOURNAL OF CELL SCIENCE 114 (13)

2537Lateral diffusion of LPS receptors
Preparation of OG-Fab probesAntibodies against hsp70 and hsp90 were digested with papainovernight and purified in order to yield Fab fragments. Aliquots ofpurified Fab, 400 µg (1 mg ml−1) were mixed with 20% by volumeof sodium bicarbonate buffer (1 M, pH 8.0) and 40 µl of freshlyprepared OG 488 succinimide dissolved in DMSO (10 mg ml−1). Themixture was placed in the dark for 1 hour at room temperature. Thereaction was terminated by adding ~10% by volume of hydroxylamine(1.5 M, pH 8.5) and incubated for a further hour in the dark. Theconjugate was subsequently separated from the unlabelled Fab anddye by passing the mixture through a PD-10 column that hadpreviously been equilibrated with PBS.
Preparation of FITC-LPS/LTALPS or LTA were labelled with FITC according to a previouslydescribed protocol (Troelstra et al., 1997). Briefly, LPS/LTA (4 mg)was made monomeric by treatment with 2 ml of 0.5% triethylamine(Sigma) and sonication for 15 minutes on ice. After the sonication,200 µl of 100 mM EDTA (BDH,) were added. The pH was thenadjusted to 5 by adding 10 µl of 1 N HCl. 800 µl of 0.25 M boratebuffer (pH 10.5) containing 20 mg FITC (Molecular Probes) wereadded to the LPS solution. The mixture was sonicated again for 1minute and, after the addition of 1 ml 1.6% sodium deoxycholate(Sigma), was incubated for 18 hours at 37°C while rotating.Aggregates were pelleted by centrifuging the mixture at 10,000 g andthe supernatant was concentrated in a dialysis bag with polyethyleneglycol 6000 (BDH). Afterwards, the conjugate was dialysed againstPBS for 1 hour and passed through a PD-10 column (Pierce) in orderto separate the FITC-LPS/LTA from the free FITC molecules. Thefractions containing FITC-LPS/LTA were pooled, concentrated anddialysed against PBS. The concentration of FITC in the finalpreparation was determined spectrophotometrically at 492 nm, withan extinction coefficient for FITC (E492) of 8.5×104 M−1 cm−1. Todetermine the amount of LPS, the level of 2-keto-3-deoxyoctulosonicacid (KDO) was measured by the thiobarbiturate assay as describedpreviously (Brade and Galanos, 1983). The FITC-LPS probe wasassayed for its KDO content in order to estimate the LPSconcentration and the absorbance at 492 nm was used to determinethe FITC content. KDO and FITC data were used to calculate thelabelling ratio. This revealed that a high degree of efficiency had beenachieved, yielding a 1:1 labelling ratio for LPS.
FITC-LPS/LTA flow cytometric binding assayThe binding of FITC-LPS/LTA to CHO cells transfected with wild-type CD14 was measured by flow cytometry. CHO cells, 3×105 persample were incubated with increasing concentrations of FITC-LPSor FITC-LTA (1–250 ng) in the presence and absence of 5% humanpooled serum (HPS). Samples were incubated for 30 minutes at 22°C,gently mixed. For blocking experiments, cells were first incubatedwith 1 µg MY4 (Coulter) mAb directed against a functional domainof CD14. Subsequently, the cells were washed three times in PBSbefore cytometric analysis. The mean fluorescence of 10,000 cells wasmeasured in a flow cytometer (FACalibur®, Becton Dickinson). Thecells were not gated.
Direct immunofluorescence for flow cytometry500,000 cells were harvested and washed twice with PBS, followedby incubation with PBS, 0.02% bovine serum albumin (BSA), 0.02%NaN3 for 30 minutes. The probe was added for 1 hour at roomtemperature. To remove the unbound probe, the cells were washedtwice with PBS, 0.02% BSA, 0.02% NaN3. Finally the cells werewashed twice with PBS, 0.02% donor calf serum (DCS), 0.02% NaN3.The final cell pellet was resuspended in 500 µl of PBS prior to flowcytometric analysis.
Cell labelling for FRAP experimentsCHO-CD14, MonoMac 6 and ECV-304 cells were allowed to adhere
on eight-well microchamber slides for 72 hours at 37°C. Afterwards,cells were washed twice in PBS prior to labelling with PE-Fab or OG-Fab for 30 minutes at 22°C or FITC-LPS/LTA for 30 minutes at 22°Cin serum-free medium supplemented with 5% HPS or 1 µg ml−1 ofLBP and 1 µg ml−1 of rsCD14. To avoid aggregation, FITC-LPS/LTAprobes were sonicated prior to cell labelling. After washing with PBSand serum-free medium, the cells were imaged at the desiredtemperature using a temperature-controlled microscope stage.
FRAP measurementsFRAP measurements were performed as previously described (Ladhaet al., 1994; Ladha et al., 1996). Briefly, slides containing labelledcells were placed onto a temperature controlled microscope stage(Physitemp, Clifton, NJ; Model TS-4) and allowed to equilibrate tothe desired temperature for FRAP measurements. After equilibration,the beam of an argon ion laser (Innova, Palo Alto, CA; 100-10) wasfocused onto the desired area on the cell. The laser beam was ofGaussian cross-sectional intensity, with a half-width at 1/e2 height ofthe laser beam at its point of focus equal to 2.15 µm spot radius. FRAPmeasurements were recorded and analysed as previously described(Ladha et al., 1996).
IL-6 inductionMonoMac 6 cells (5×105) were mixed with 50 µl of serial dilutionsof LPS in the presence of 1% HPS in serum-free medium. After 2.5hours, the supernatant was collected and analysed for IL-6 using anELISA (Diaclone, France).
RESULTS
Generation and characterization of PE-Fab probe forFRAPIn order to study the mobility of CD14 on the cell surface beforeand after LPS stimulation, a fully characterized probe of smallsize and high fluorescent yield was prepared. Phycobiliprotein-Fab conjugates were prepared from PE and Fab fragments ofthe mAb 26ic. This antibody, which is specific for CD14, waschosen because it binds to a non-functional domain of CD14that is distinct from the LPS binding domain (Wright et al.,1990). Therefore the binding of the LPS to CD14 would notinterfere with the binding of the probe.
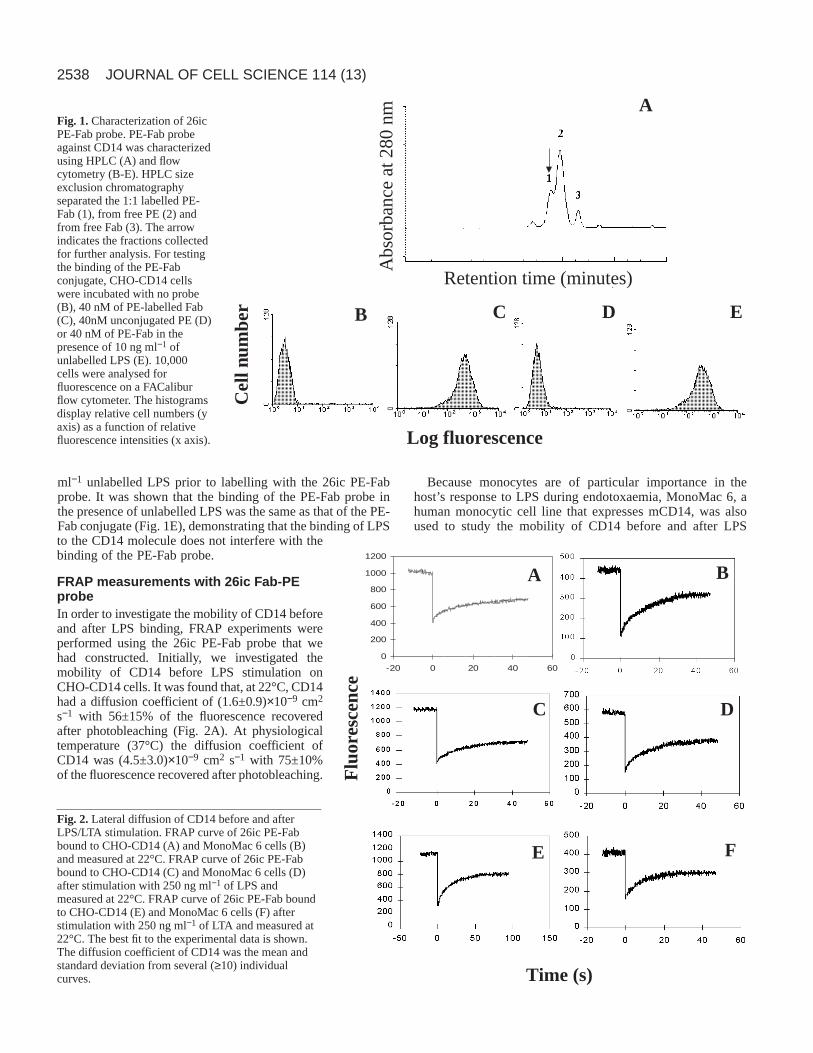
Fab fragments of the mAb 26ic were conjugated in a 1:1molar ratio to PE. The PE-Fab conjugate, which had amolecular mass higher than either of the two starting materials,was purified by HPLC size exclusion chromatography. UsingHPLC the PE-Fab conjugate was eluted first, followed by thefree PE and Fab. Fig. 1A shows a typical HPLC profileobtained with a PE-Fab probe, in which three major peaks areshown corresponding to PE-Fab (1), free PE (2) and unlabelledFab (3). The arrow indicates the 1:1 PE-Fab fractions collectedfor flow cytometric and FRAP experiments.
The 1:1 PE-Fab was then tested for specificity and bindingto the CD14 receptor on CHO-CD14 cells (Fig. 1B-E). Flowcytometric analysis revealed a high peak fluorescence intensitywhen 4 pmol of the 1:1 PE-Fab was used (Fig. 1C), whencompared with the fluorescence intensity of unlabelled cells(Fig. 1B). Nonspecific binding of the PE was checked byincubating cells with 4 pmol of unconjugated PE, and peakfluorescence intensity was reduced by 94% (Fig. 1D). Usingflow cytometry, the binding of the PE-Fab conjugate was alsocompared with that of the PE-Fab conjugate in the presence ofunlabelled LPS. CHO-CD14 cells were incubated with 10 ng
2538
ml−1 unlabelled LPS prior to labelling with the 26ic PE-Fabprobe. It was shown that the binding of the PE-Fab probe inthe presence of unlabelled LPS was the same as that of the PE-Fab conjugate (Fig. 1E), demonstrating that the binding of LPSto the CD14 molecule does not interfere with thebinding of the PE-Fab probe.
FRAP measurements with 26ic Fab-PEprobeIn order to investigate the mobility of CD14 beforeand after LPS binding, FRAP experiments wereperformed using the 26ic PE-Fab probe that wehad constructed. Initially, we investigated themobility of CD14 before LPS stimulation onCHO-CD14 cells. It was found that, at 22°C, CD14had a diffusion coefficient of (1.6±0.9)×10−9 cm2
s−1 with 56±15% of the fluorescence recoveredafter photobleaching (Fig. 2A). At physiologicaltemperature (37°C) the diffusion coefficient ofCD14 was (4.5±3.0)×10−9 cm2 s−1 with 75±10%of the fluorescence recovered after photobleaching.
Because monocytes are of particular importance in thehost’s response to LPS during endotoxaemia, MonoMac 6, ahuman monocytic cell line that expresses mCD14, was alsoused to study the mobility of CD14 before and after LPS
JOURNAL OF CELL SCIENCE 114 (13)
B C D E
Log fluorescence
Cel
l num
ber
A
Retention time (minutes)
Abs
orba
nce
at 2
80 n
m
Fig. 1.Characterization of 26icPE-Fab probe. PE-Fab probeagainst CD14 was characterizedusing HPLC (A) and flowcytometry (B-E). HPLC sizeexclusion chromatographyseparated the 1:1 labelled PE-Fab (1), from free PE (2) andfrom free Fab (3). The arrowindicates the fractions collectedfor further analysis. For testingthe binding of the PE-Fabconjugate, CHO-CD14 cellswere incubated with no probe(B), 40 nM of PE-labelled Fab(C), 40nM unconjugated PE (D)or 40 nM of PE-Fab in thepresence of 10 ng ml−1 ofunlabelled LPS (E). 10,000cells were analysed forfluorescence on a FACaliburflow cytometer. The histogramsdisplay relative cell numbers (yaxis) as a function of relativefluorescence intensities (x axis).
0
200
400
600
800
1000
1200
-20 0 20 40 60
A B
C D
E F
Time (s)
Flu
ores
cenc
e
Fig. 2.Lateral diffusion of CD14 before and afterLPS/LTA stimulation. FRAP curve of 26ic PE-Fabbound to CHO-CD14 (A) and MonoMac 6 cells (B)and measured at 22°C. FRAP curve of 26ic PE-Fabbound to CHO-CD14 (C) and MonoMac 6 cells (D)after stimulation with 250 ng ml−1 of LPS andmeasured at 22°C. FRAP curve of 26ic PE-Fab boundto CHO-CD14 (E) and MonoMac 6 cells (F) afterstimulation with 250 ng ml−1 of LTA and measured at22°C. The best fit to the experimental data is shown.The diffusion coefficient of CD14 was the mean andstandard deviation from several (≥10) individualcurves.

2539Lateral diffusion of LPS receptors
stimulation. It was found that, at 22°C, CD14 had a diffusioncoefficient of (1.35±0.6)×10−9 cm2 s−1 (Fig. 2B), almostidentical to that obtained with the CHO transfectants, althoughthe fluorescence recovery was somewhat higher.
After CHO-CD14 cells were stimulated with 5 ng ml−1 ofunlabelled LPS in 5% HPS, the diffusion coefficient of CD14remained virtually unchanged, (1.2±0.7)×10−9 cm2 s−1 with51±13% of the fluorescence recovered after photobleaching(Table 1). Even when higher concentrations of LPS were usedto stimulate the cells (250 ng ml−1), the diffusion coefficient ofCD14 remained essentially the same, (1.5±0.7)×10−9 cm2 s−1
with 50±8% fluorescence recovery (Fig. 2C). Similar resultswere obtained when MonoMac 6 cells were stimulated with250 ng ml−1 of LPS. The diffusion coefficient of CD14 onMonoMac 6 cells after stimulation with LPS was(1.5±0.6)×10−9 cm2 s−1 with 70±20% of the fluorescencerecovered after photobleaching (Fig. 2D), very similar to thediffusion coefficient obtained prior to LPS stimulation.Furthermore, FRAP experiments performed on CHO-CD14and MonoMac 6 cells with the 26ic Fab-PE probe in thepresence and absence of unlabelled LPS revealed almostidentical results (data not shown), suggesting that the bindingof LPS did not affect the lateral diffusion of CD14 in the lipidbilayer.
It has been suggested that CD14 might also function as acellular receptor for peptidoglycan or LTA from Gram-positivebacteria, and so we investigated whether stimulation with LTAwould affect the mobility of CD14. Before and after LTAstimulation (250 ng ml−1), the CD14 lateral diffusioncoefficient remained essentially the same. On CHO-CD14cells, CD14 exhibited diffusion coefficients of (1.9±0.8)×10−9,with 80±16% of the fluorescence recovered after photo-bleaching (Fig. 2E). After LTA stimulation on MonoMac 6cells, CD14 exhibited diffusion coefficients of (1.6±0.5)×10−9,with 75±12% of the fluorescence recovered after photo-
bleaching (Fig. 2F). Table 1 summarizes the results obtainedfrom FRAP experiments using the 26ic Fab-PE probe.
Generation and characterization of FITC-LPS probefor FRAPIn order to test whether LPS remains bound to CD14, wedecided to label the ligand itself and to use it as a probe forFRAP measurements. Using a protocol described previously(Troelstra et al., 1997), we prepared an FITC-conjugated LPSof high fluorescent yield that enabled us to detect lowconcentrations of LPS. This conjugate had previously beenshown to be biological active, by testing its ability to induceTNF-α production (Triantafilou et al., 2000a).
The FITC-LPS probe was then tested for specificity andbinding to CHO-CD14 cells (Fig. 3). Flow cytometric analysisrevealed a high peak fluorescence intensity when 10 ng ml−1
of FITC-LPS were used (Fig. 3B) compared with thefluorescence intensity of unlabelled cells (Fig. 3A).Nonspecific binding of the FITC-LPS probe was checked byincubating cells with a 100-fold excess of unlabelled LPS, inwhich peak fluorescence intensity was almost identical to thatof the unlabelled cells (Fig. 3C).
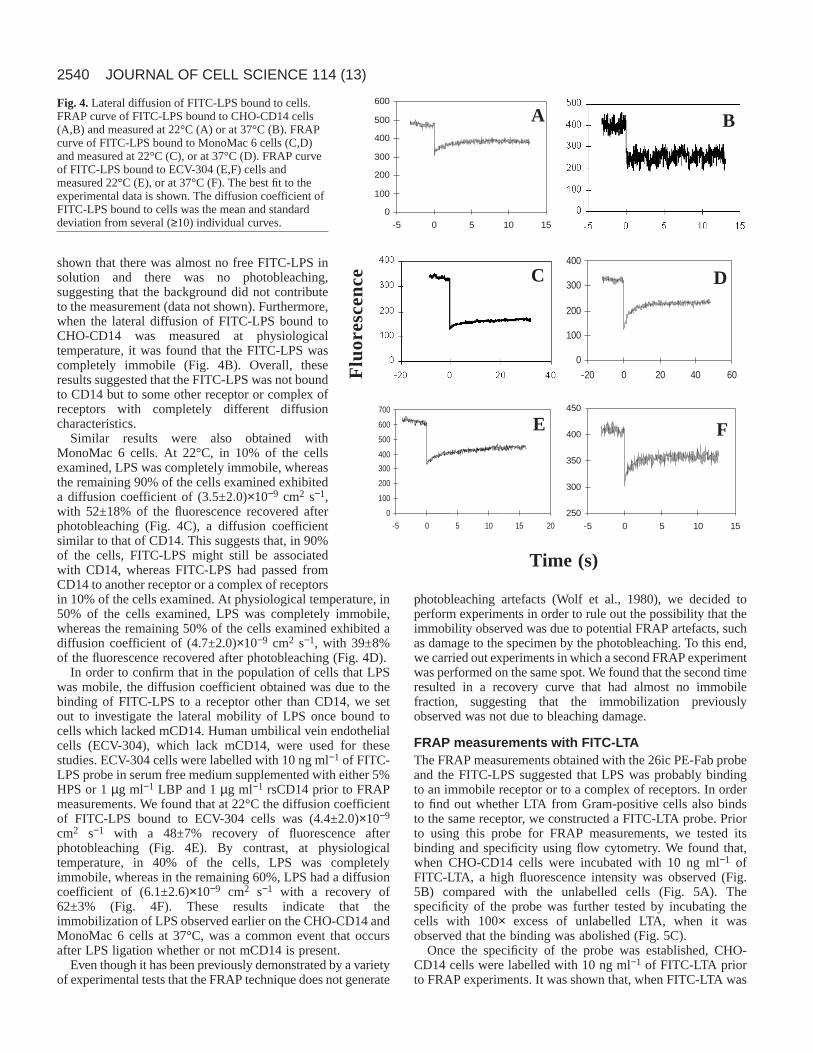
FRAP measurements with FITC-LPSIn order to examine the mobility of LPS by FRAP, CHO-CD14cells were labelled with 10 ng ml−1 of FITC-LPS in serum-freemedium supplemented with either 5% HPS or 1 µg ml−1 ofrLBP and 1 µg ml−1 rsCD14. Surprisingly, it was found that,in 60% of the cells examined, LPS was completely immobileand, in 40% of the cells examined, LPS had a diffusioncoefficient of (1.1±0.3)×10−8 cm2 s−1 with 53±10% recovery,much different to that observed for CD14 (Fig. 4A). In orderto verify that the fast diffusion coefficient observed in 40% ofthe cells was not free FITC-LPS in solution, FRAPmeasurements of the background were performed. It was
Table 1. Lateral diffusion of CD14 on CHO-CD14 and MonoMac 6 cellsCells Probe used °C Stimulated Diffusion coefficient (cm2/s)* Percentage recovery*
CHO-CD14 PE-Fab 22 − (1.58±0.9)×10−9 56±15CHO-CD14 PE-Fab 37 − (4.45±3.0)×10−9 75±10CHO-CD14 PE-Fab 22 5 ng/ml LPS (1.21±0.7)×10−9 51±13CHO-CD14 PE-Fab 22 250 ng/ml LPS (1.52±0.7)×10−9 49±8CHO-CD14 PE-Fab 22 250 ng/ml LTA (1.94±0.8)×10−9 80±16
MonoMac 6 PE-Fab 22 − (1.35±0.6)×10−9 78±18MonoMac 6 PE-Fab 22 250 ng/ml LPS (1.49±0.6)×10−9 70±20MonoMac 6 PE-Fab 22 250 ng/ml LTA (1.61±0.5)×10−9 75±12
*The values represent the mean ± standard deviation from several determinations (≥10).
A B C
Log fluorescence
Cel
l nu
mbe
r
Fig. 3.Characterization of FITC-LPS probe.CHO-CD14 cells were incubated with no FITC-LPS probe (A), 10 ng ml−1 of FITC-LPS probe(B), or with 10 ng ml−1 of FITC-LPS in thepresence of 100× excess of unlabelled LPS (C).10,000 cells were analysed for fluorescence on aFACalibur flow cytometer. The histogramsdisplay relative cell numbers (y axis) as afunction of relative fluorescence intensities (xaxis).
2540
shown that there was almost no free FITC-LPS insolution and there was no photobleaching,suggesting that the background did not contributeto the measurement (data not shown). Furthermore,when the lateral diffusion of FITC-LPS bound toCHO-CD14 was measured at physiologicaltemperature, it was found that the FITC-LPS wascompletely immobile (Fig. 4B). Overall, theseresults suggested that the FITC-LPS was not boundto CD14 but to some other receptor or complex ofreceptors with completely different diffusioncharacteristics.
Similar results were also obtained withMonoMac 6 cells. At 22°C, in 10% of the cellsexamined, LPS was completely immobile, whereasthe remaining 90% of the cells examined exhibiteda diffusion coefficient of (3.5±2.0)×10−9 cm2 s−1,with 52±18% of the fluorescence recovered afterphotobleaching (Fig. 4C), a diffusion coefficientsimilar to that of CD14. This suggests that, in 90%of the cells, FITC-LPS might still be associatedwith CD14, whereas FITC-LPS had passed fromCD14 to another receptor or a complex of receptorsin 10% of the cells examined. At physiological temperature, in50% of the cells examined, LPS was completely immobile,whereas the remaining 50% of the cells examined exhibited adiffusion coefficient of (4.7±2.0)×10−9 cm2 s−1, with 39±8%of the fluorescence recovered after photobleaching (Fig. 4D).
In order to confirm that in the population of cells that LPSwas mobile, the diffusion coefficient obtained was due to thebinding of FITC-LPS to a receptor other than CD14, we setout to investigate the lateral mobility of LPS once bound tocells which lacked mCD14. Human umbilical vein endothelialcells (ECV-304), which lack mCD14, were used for thesestudies. ECV-304 cells were labelled with 10 ng ml−1 of FITC-LPS probe in serum free medium supplemented with either 5%HPS or 1 µg ml−1 LBP and 1 µg ml−1 rsCD14 prior to FRAPmeasurements. We found that at 22°C the diffusion coefficientof FITC-LPS bound to ECV-304 cells was (4.4±2.0)×10−9
cm2 s−1 with a 48±7% recovery of fluorescence afterphotobleaching (Fig. 4E). By contrast, at physiologicaltemperature, in 40% of the cells, LPS was completelyimmobile, whereas in the remaining 60%, LPS had a diffusioncoefficient of (6.1±2.6)×10−9 cm2 s−1 with a recovery of62±3% (Fig. 4F). These results indicate that theimmobilization of LPS observed earlier on the CHO-CD14 andMonoMac 6 cells at 37°C, was a common event that occursafter LPS ligation whether or not mCD14 is present.
Even though it has been previously demonstrated by a varietyof experimental tests that the FRAP technique does not generate
photobleaching artefacts (Wolf et al., 1980), we decided toperform experiments in order to rule out the possibility that theimmobility observed was due to potential FRAP artefacts, suchas damage to the specimen by the photobleaching. To this end,we carried out experiments in which a second FRAP experimentwas performed on the same spot. We found that the second timeresulted in a recovery curve that had almost no immobilefraction, suggesting that the immobilization previouslyobserved was not due to bleaching damage.
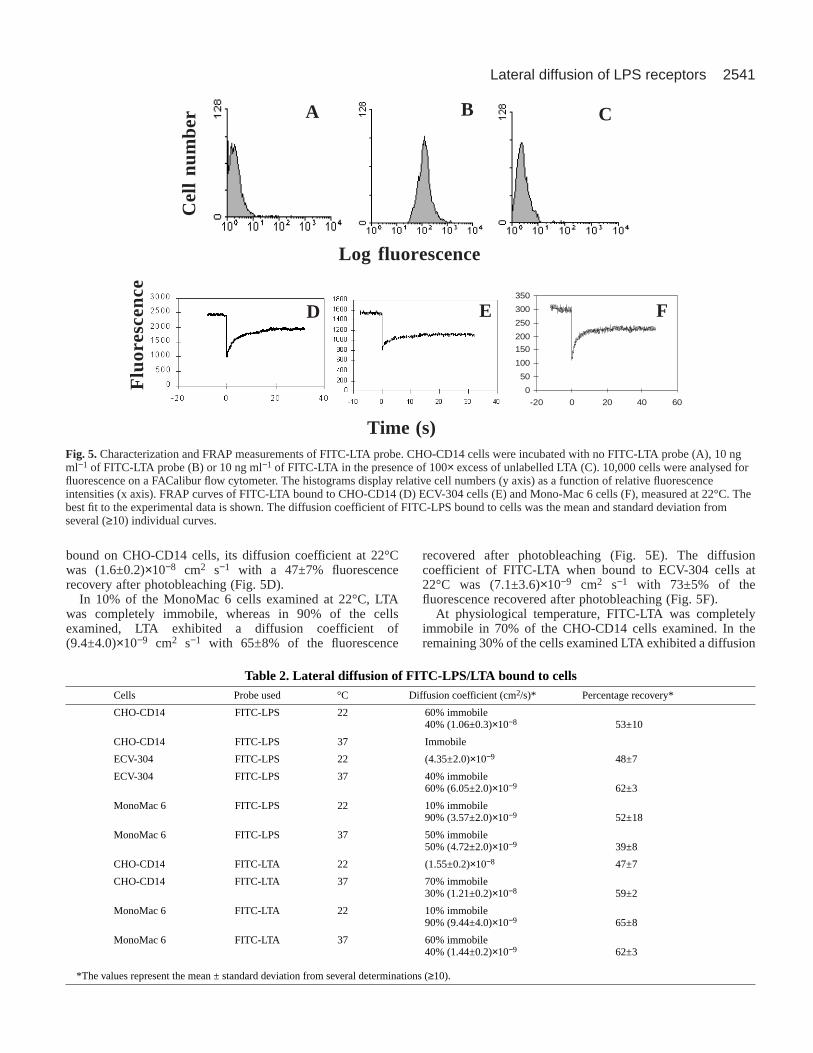
FRAP measurements with FITC-LTAThe FRAP measurements obtained with the 26ic PE-Fab probeand the FITC-LPS suggested that LPS was probably bindingto an immobile receptor or to a complex of receptors. In orderto find out whether LTA from Gram-positive cells also bindsto the same receptor, we constructed a FITC-LTA probe. Priorto using this probe for FRAP measurements, we tested itsbinding and specificity using flow cytometry. We found that,when CHO-CD14 cells were incubated with 10 ng ml−1 ofFITC-LTA, a high fluorescence intensity was observed (Fig.5B) compared with the unlabelled cells (Fig. 5A). Thespecificity of the probe was further tested by incubating thecells with 100× excess of unlabelled LTA, when it wasobserved that the binding was abolished (Fig. 5C).
Once the specificity of the probe was established, CHO-CD14 cells were labelled with 10 ng ml−1 of FITC-LTA priorto FRAP experiments. It was shown that, when FITC-LTA was
JOURNAL OF CELL SCIENCE 114 (13)
0
100
200
300
400
500
600
-5 0 5 10 15
A B
0
100
200
300
400
500
600
700
-5 0 5 10 15 20
E
250
300
350
400
450
-5 0 5 10 15
F
C
0
100
200
300
400
-20 0 20 40 60
D
Flu
ore
sce
nce
Time (s)
Fig. 4.Lateral diffusion of FITC-LPS bound to cells.FRAP curve of FITC-LPS bound to CHO-CD14 cells(A,B) and measured at 22°C (A) or at 37°C (B). FRAPcurve of FITC-LPS bound to MonoMac 6 cells (C,D)and measured at 22°C (C), or at 37°C (D). FRAP curveof FITC-LPS bound to ECV-304 (E,F) cells andmeasured 22°C (E), or at 37°C (F). The best fit to theexperimental data is shown. The diffusion coefficient ofFITC-LPS bound to cells was the mean and standarddeviation from several (≥10) individual curves.
2541Lateral diffusion of LPS receptors
bound on CHO-CD14 cells, its diffusion coefficient at 22°Cwas (1.6±0.2)×10−8 cm2 s−1 with a 47±7% fluorescencerecovery after photobleaching (Fig. 5D).
In 10% of the MonoMac 6 cells examined at 22°C, LTAwas completely immobile, whereas in 90% of the cellsexamined, LTA exhibited a diffusion coefficient of(9.4±4.0)×10−9 cm2 s−1 with 65±8% of the fluorescence
recovered after photobleaching (Fig. 5E). The diffusioncoefficient of FITC-LTA when bound to ECV-304 cells at22°C was (7.1±3.6)×10−9 cm2 s−1 with 73±5% of thefluorescence recovered after photobleaching (Fig. 5F).
At physiological temperature, FITC-LTA was completelyimmobile in 70% of the CHO-CD14 cells examined. In theremaining 30% of the cells examined LTA exhibited a diffusion
D E
A B C
Log fluorescence
Cel
l nu
mbe
r
0
50
100
150
200
250
300
350
-20 0 20 40 60
F
Time (s)
Flu
ore
sce
nce
Fig. 5.Characterization and FRAP measurements of FITC-LTA probe. CHO-CD14 cells were incubated with no FITC-LTA probe (A), 10 ngml−1 of FITC-LTA probe (B) or 10 ng ml−1 of FITC-LTA in the presence of 100× excess of unlabelled LTA (C). 10,000 cells were analysed forfluorescence on a FACalibur flow cytometer. The histograms display relative cell numbers (y axis) as a function of relative fluorescenceintensities (x axis). FRAP curves of FITC-LTA bound to CHO-CD14 (D) ECV-304 cells (E) and Mono-Mac 6 cells (F), measured at 22°C. Thebest fit to the experimental data is shown. The diffusion coefficient of FITC-LPS bound to cells was the mean and standard deviation fromseveral (≥10) individual curves.
Table 2. Lateral diffusion of FITC-LPS/LTA bound to cellsCells Probe used °C Diffusion coefficient (cm2/s)* Percentage recovery*
CHO-CD14 FITC-LPS 22 60% immobile40% (1.06±0.3)×10−8 53±10
CHO-CD14 FITC-LPS 37 Immobile
ECV-304 FITC-LPS 22 (4.35±2.0)×10−9 48±7
ECV-304 FITC-LPS 37 40% immobile60% (6.05±2.0)×10−9 62±3
MonoMac 6 FITC-LPS 22 10% immobile90% (3.57±2.0)×10−9 52±18
MonoMac 6 FITC-LPS 37 50% immobile50% (4.72±2.0)×10−9 39±8
CHO-CD14 FITC-LTA 22 (1.55±0.2)×10−8 47±7
CHO-CD14 FITC-LTA 37 70% immobile30% (1.21±0.2)×10−8 59±2
MonoMac 6 FITC-LTA 22 10% immobile90% (9.44±4.0)×10−9 65±8
MonoMac 6 FITC-LTA 37 60% immobile40% (1.44±0.2)×10−9 62±3
*The values represent the mean ± standard deviation from several determinations (≥10).

2542
coefficient of (1.2±0.2)×10−9 cm2 s−1 with 59±2% of thefluorescence recovered after photobleaching. When MonoMac6 cells were used, LTA was completely immobile in 60% ofthe cells examined, whereas LTA displayed a diffusioncoefficient of (1.4±0.2)×10−9 cm2 s−1 in 40%, with 62±3% ofthe fluorescence recovered after photobleaching. These resultsare similar to those obtained with FITC-LPS on CHO-CD14and MonoMac 6 cells, suggesting that both LPS and LTAtransfer to the same receptor.
FRAP measurements of the background in both cell types(i.e. away from the cells) showed much weaker fluorescenceintensities and no photobleaching, verifying that the diffusioncoefficients obtained were not free FITC-LTA probe (data notshown). FRAP data obtained using FITC-LPS/LTA probes aresummarized in Table 2.
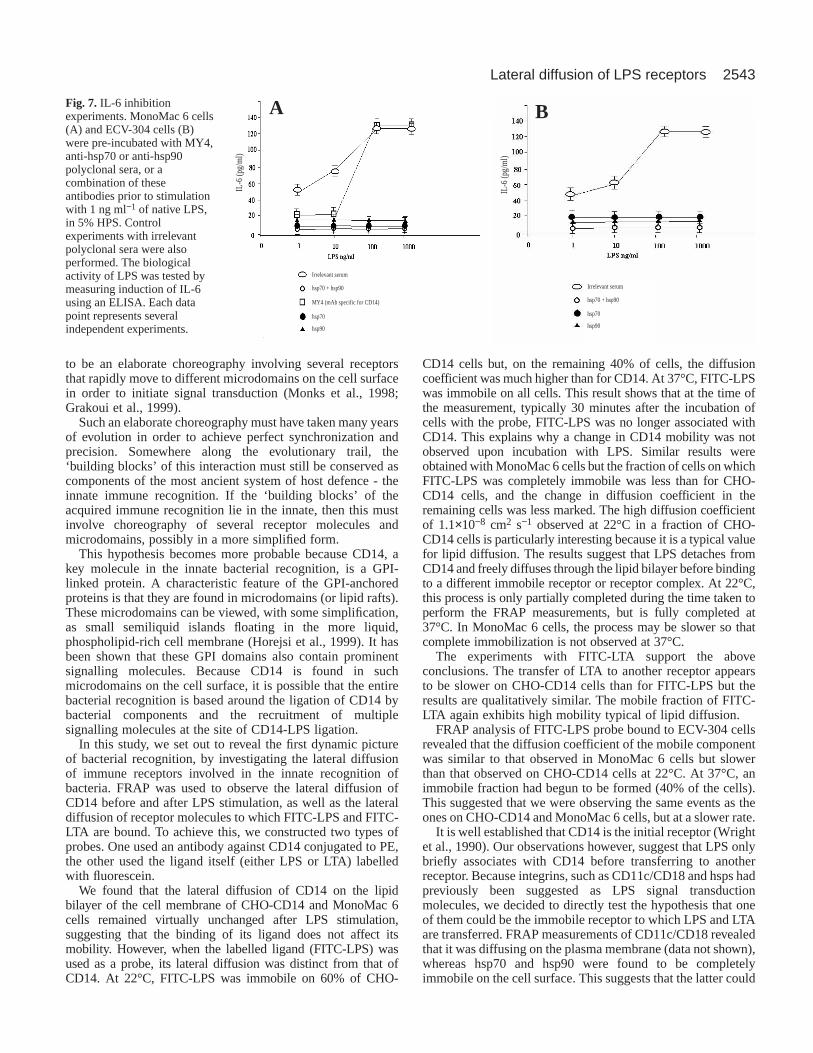
FRAP measurements of heat shock proteinsOur data indicated that FITC-LPS/LTA were probably bindingto either an immobile receptor or a complex of receptors, andso we set out to reveal the identity of this receptor. Probesagainst integrin CD11c/CD18, hsp70 and hsp90 were generatedby conjugating antibodies against these molecules to OG 488.All probes were characterized and tested for specificity by flowcytometry prior to using them for FRAP experiments (data notshown). Integrins such as CD11c/CD18, which had previouslybeen suggested as LPS ‘signal transducers’, were found to bediffusing on the plasma membrane before and after LPSstimulation (data not shown). By contrast, hsp70 (Fig. 6A) andhsp90 (Fig. 6B), which had recently been implicated inmediating macrophage activation by taxol (Byrd et al., 1999),a plant-derived antitumour agent that elicits cell signalsindistinguishable to LPS, were found to have either very lowmobility of were found to be immobile on MonoMac 6 cells,thus suggesting that these proteins were probably the immobilereceptor(s) that we had observed. The lateral diffusions of hsp70and hsp90 were also measured on ECV-304 cells, which lackmCD14, yielding similar results (Fig. 6C,D). In order to
determine whether hsp70 and hsp90 were the immobilereceptors that LPS was transferred to, we investigated the effectof anti-hsp antibodies on the immobilization of LPS usingFRAP. It was revealed that in the presence of anti-hsp70 andanti-hsp90 antibodies, LPS remained mobile at 37°C, with adiffusion coefficient of (3.3±0.1)×10−9 cm2 s−1 and recovery of78±4%, suggesting that hsp70 and hsp90 were the immobilereceptors that LPS was transferring to.
IL-6 induction experimentsOur FRAP data suggested that the heat shock proteins were theimmobile receptor that we were observing, and so antibodyinhibition experiments of IL-6 induction were performed inorder to determine the roles of hsp70 and hsp90 in thebiological response to LPS. IL-6 secretion was measured afterLPS stimulation. MonoMac 6 cells were pre-incubated withMY4 (CD14 specific blocking mAb), anti-hsp70, anti-hsp90 oran irrelevant polyclonal serum and an isotype control. It wasfound that antibodies against hsp70 and hsp90 inhibited theresponse by 85%, whereas a combination of hsp70 and hsp90antibodies completely inhibited IL-6 secretion (Fig. 7A),strongly suggesting that hsp70 and hsp90 are involved in LPSsignal transduction. Similar results were obtained with ECV-304 cells, which lack mCD14 (Fig. 7B).
DISCUSSION
The recognition of foreign antigens is the first and most crucialstep in both innate and adaptive immunity. Even though theinnate immune recognition is not well understood, themolecular interactions involved in the acquired antigenrecognition have been revealed. It is generally accepted that theacquired immune response involves the engagement of twotypes of cells, APCs and T cells. Recently it was revealed thatthe interaction between the two is more complex thanpreviously imagined. The APC-T cell interaction was shown
JOURNAL OF CELL SCIENCE 114 (13)
0
200
400
600
800
1000
-20 0 20 40 60
A B
C D
Time (s)
Flu
ore
scen
ce
Fig. 6.FRAP measurements of heatshock proteins. FRAP curve ofhsp70 OG-Fab bound to MonoMac6 (A) and ECV-304 cells (B) andmeasured at 22°C. FRAP curve ofhsp90 OG-Fab bound to MonoMac6 (C) and ECV-304 cells (D) andmeasured at 22°C. The best fit to theexperimental data is shown. Thediffusion coefficients of hsp70 andhsp90 were the mean and standarddeviation from several (≥10)individual curves.
2543Lateral diffusion of LPS receptors
to be an elaborate choreography involving several receptorsthat rapidly move to different microdomains on the cell surfacein order to initiate signal transduction (Monks et al., 1998;Grakoui et al., 1999).
Such an elaborate choreography must have taken many yearsof evolution in order to achieve perfect synchronization andprecision. Somewhere along the evolutionary trail, the‘building blocks’ of this interaction must still be conserved ascomponents of the most ancient system of host defence - theinnate immune recognition. If the ‘building blocks’ of theacquired immune recognition lie in the innate, then this mustinvolve choreography of several receptor molecules andmicrodomains, possibly in a more simplified form.
This hypothesis becomes more probable because CD14, akey molecule in the innate bacterial recognition, is a GPI-linked protein. A characteristic feature of the GPI-anchoredproteins is that they are found in microdomains (or lipid rafts).These microdomains can be viewed, with some simplification,as small semiliquid islands floating in the more liquid,phospholipid-rich cell membrane (Horejsi et al., 1999). It hasbeen shown that these GPI domains also contain prominentsignalling molecules. Because CD14 is found in suchmicrodomains on the cell surface, it is possible that the entirebacterial recognition is based around the ligation of CD14 bybacterial components and the recruitment of multiplesignalling molecules at the site of CD14-LPS ligation.
In this study, we set out to reveal the first dynamic pictureof bacterial recognition, by investigating the lateral diffusionof immune receptors involved in the innate recognition ofbacteria. FRAP was used to observe the lateral diffusion ofCD14 before and after LPS stimulation, as well as the lateraldiffusion of receptor molecules to which FITC-LPS and FITC-LTA are bound. To achieve this, we constructed two types ofprobes. One used an antibody against CD14 conjugated to PE,the other used the ligand itself (either LPS or LTA) labelledwith fluorescein.
We found that the lateral diffusion of CD14 on the lipidbilayer of the cell membrane of CHO-CD14 and MonoMac 6cells remained virtually unchanged after LPS stimulation,suggesting that the binding of its ligand does not affect itsmobility. However, when the labelled ligand (FITC-LPS) wasused as a probe, its lateral diffusion was distinct from that ofCD14. At 22°C, FITC-LPS was immobile on 60% of CHO-
CD14 cells but, on the remaining 40% of cells, the diffusioncoefficient was much higher than for CD14. At 37°C, FITC-LPSwas immobile on all cells. This result shows that at the time ofthe measurement, typically 30 minutes after the incubation ofcells with the probe, FITC-LPS was no longer associated withCD14. This explains why a change in CD14 mobility was notobserved upon incubation with LPS. Similar results wereobtained with MonoMac 6 cells but the fraction of cells on whichFITC-LPS was completely immobile was less than for CHO-CD14 cells, and the change in diffusion coefficient in theremaining cells was less marked. The high diffusion coefficientof 1.1×10−8 cm2 s−1 observed at 22°C in a fraction of CHO-CD14 cells is particularly interesting because it is a typical valuefor lipid diffusion. The results suggest that LPS detaches fromCD14 and freely diffuses through the lipid bilayer before bindingto a different immobile receptor or receptor complex. At 22°C,this process is only partially completed during the time taken toperform the FRAP measurements, but is fully completed at37°C. In MonoMac 6 cells, the process may be slower so thatcomplete immobilization is not observed at 37°C.
The experiments with FITC-LTA support the aboveconclusions. The transfer of LTA to another receptor appearsto be slower on CHO-CD14 cells than for FITC-LPS but theresults are qualitatively similar. The mobile fraction of FITC-LTA again exhibits high mobility typical of lipid diffusion.
FRAP analysis of FITC-LPS probe bound to ECV-304 cellsrevealed that the diffusion coefficient of the mobile componentwas similar to that observed in MonoMac 6 cells but slowerthan that observed on CHO-CD14 cells at 22°C. At 37°C, animmobile fraction had begun to be formed (40% of the cells).This suggested that we were observing the same events as theones on CHO-CD14 and MonoMac 6 cells, but at a slower rate.
It is well established that CD14 is the initial receptor (Wrightet al., 1990). Our observations however, suggest that LPS onlybriefly associates with CD14 before transferring to anotherreceptor. Because integrins, such as CD11c/CD18 and hsps hadpreviously been suggested as LPS signal transductionmolecules, we decided to directly test the hypothesis that oneof them could be the immobile receptor to which LPS and LTAare transferred. FRAP measurements of CD11c/CD18 revealedthat it was diffusing on the plasma membrane (data not shown),whereas hsp70 and hsp90 were found to be completelyimmobile on the cell surface. This suggests that the latter could
A
Irrelevant serum
hsp70 + hsp90
MY4 (mAb specific for CD14)
hsp70
hsp90
Irrelevant serum
hsp70 + hsp90
hsp70
hsp90
IL-6
(pg/
ml)
IL-6
(pg/
ml)
A BFig. 7. IL-6 inhibitionexperiments. MonoMac 6 cells(A) and ECV-304 cells (B)were pre-incubated with MY4,anti-hsp70 or anti-hsp90polyclonal sera, or acombination of theseantibodies prior to stimulationwith 1 ng ml−1 of native LPS,in 5% HPS. Controlexperiments with irrelevantpolyclonal sera were alsoperformed. The biologicalactivity of LPS was tested bymeasuring induction of IL-6using an ELISA. Each datapoint represents severalindependent experiments.

2544
well be the immobile receptors to which LPS/LTA istransferred. Therefore, we decided to investigate the effect ofanti-hsp70 and anti-hsp90 antibodies on the immobilization ofLPS. We found that, once hsp70 and hsp90 were blocked,FITC-LPS remained mobile, suggesting that these were thereceptors that LPS was transferring to. Furthermore, wedecided to test whether hsp70 and hsp90 were involved insignal transduction by performing antibody inhibitionexperiments of IL-6 induction. It was found that incubationwith polyclonal sera against these hsps, prior to LPSstimulation, completely inhibited IL-6 secretion, suggestingthat hsp70 and hsp90 are part of the LPS multimeric receptorand are involved in LPS signal transduction.
Previous investigations have shown that a variety ofmolecules including CD11c/CD18 (Ingalls and Golenbock,1995), the Toll-like family of receptors (Kirschning et al.,1998), moesin (Tohme et al., 1999) and heat shock proteins(Byrd et al., 1999) are involved in LPS signal transduction. Theevents that occur between binding of LPS to CD14 and signaltransduction have, however, not been established. Ourexperiments show that LPS transfers from CD14 to immobilereceptors that are very probably hsp70 and hsp90. The lack ofmobility suggest that these heat shock proteins could be partof a large multimeric complex that includes other moleculesthat have previously been implicated in signal transduction butdoes not include CD14.
In conclusion our data suggest that the heat shock family ofproteins might be part of an LPS multimeric receptor but thatthey are not physically associated with the CD14 receptor.Therefore, we propose a model based on our findings that,following CD14 ligation by bacterial components, a complexof receptors is probably formed within the lipid rafts involvingseveral signalling molecules, including hsp70 and hsp90 (Fig.8). CD14 releases LPS, catalysing the transfer of LPS to asignal transducing complex that involves hsp70 and 90, leadingto cell activation. In the absence of CD14, this receptorcomplex is still formed.
This work was supported by the BBSRC. We would like to thankKeith Wilson for helpful discussions. We would also like to thankTheodore Kirkland and Suganya Viriyakosol for kindly providingtheir CHO-CD14 cells.
REFERENCES
Blondin, C., Ledur, A., Cholley, B., Caroff, M. and Haeffner Cavaillon, N.
(1997). Lipopolysaccharide complexed with soluble CD14 binds to normalhuman monocytes. Eur. J. Immunol. 27, 3303-3309.
Bone, R. C. (1991). The pathogenesis of sepsis. Ann. Intern. Med.115, 457-450.
Brade, H. and Galanos, C. (1983). A method to detect 2-keto-3-deoxyoctanatand related compounds on pherograms and chromatograms. Anal. Biochem.132, 158-159.
Byrd, C. A., Bornmann, W., Erdjument-Bromage, H., Tempst, P.,Pavletich, N., Rosen, N., Nathan, C. F. and Ding, A. (1999). Heat shock90 mediates macrophage activation by Taxol and bacteriallipopolysaccharide. Proc. Natl. Acad. Sci. USA96, 5645-5650.
Demters, P. A., Zhou, D. and Powell, D. E. (1994). Response of neutrophilsto LPS. Different signalling pathways for CD18 mediated adhesion and Fc-mediated phagocytosis. J. Immunol. 153, 2137-2144.
Durieux, J. J., Vita, N., Popescu, O., Guette, F., Calzadawack, J., Munker,R., Schmidt, R. E., Lupker, J., Ferrara, P., Zieglerheitbrock, H. L. et al.(1994). The 2 soluble forms of the lipopolysaccharide receptor, CD14 -characterization and release by normal human monocytes. Eur. J. Immunol.24, 2006-2012.
Frey, E. A., Miller, D. S., Jahr, T. G., Sundan, A., Bazil, V., Espevik, T.,Finlay, B. B. and Wright, S. D. (1992). Soluble CD14 participates in theresponse of cells to lipopolysaccharide. J. Exp. Med.176, 1668-1671
Gegner, J. A., Ulevitch, R. J. and Tobias, P. S. (1995). Lipopolysaccharide(LPS) Signal-transduction and clearance - dual roles for LPS binding-protein and membrane CD14. J. Biol. Chem. 270, 5319-5325.
Grakoui, A., Bromley, S. K., Sumen, C., Davis, M. M., Shaw, A. S., Allen,P. M. and Dustin, M. L. (1999). The immunological synapse: a molecularmachine controlling T cell activation. Science285, 221-227.
Gupta, D., Kirkland, T. N., Viriyakosol, S. and Dziarski, R. (1996). CD14is a cell-activating receptor for bacterial peptidoglycan. J. Biol. Chem. 271,23310-23316.
Horejsi, V., Drbal, K., Cebecauer, M., Cerny, J., Brdicka, T., Angelisova,P. and Stockinger, H. (1999). GPI-microdomains: a role in signalling viaimmunoreceptors. Immunol. Today20, 356-361.
Ingalls, R. R. and Golenbock, D. T. (1995). CD11c/CD18, a transmembranesignaling receptor for lipopolysaccharide. J. Exp. Med. 181, 1472-1479.
Ingalls, R. R., Arnaout, M. A. and Golenbock, D. (1997). Outside-insignalling by lipopolysaccharide through a tailless integrin. J. Immunol. 159,433-440.
Kirschning, C. J., Wesche, H., Ayers, T. M. and Rothe, M. (1998). HumanToll-like receptor 2 confers responsiveness to bacterial lipopolyssacharide.J. Exp. Med. 188, 2091-2097.
Labeta, M. O., Durieux, J. J., Fernandez, N., Hermann, R. and Ferrara,P. (1993). Release from a human monocyte-like cell line of two differentsoluble forms of the lipopolysaccharide receptro, CD14. Eur. J. Immunol.23, 2144-2151.
Ladha, S., Mackie, A. and Clark, D. (1994). Cheek cell membrane fluiditymeasured by fluorescence recovery after photobleaching and steady stateanisotropy. J. Membr. Biol. 142, 223-229.
Ladha, S., Mackie, A., Harvey, L., Clark, D., Lea, E., Brullemans, M. andDuclohier, H. (1996). Lateral diffusion in planar lipid bilayers. A FRAPinvestigation of its modulation by lipid composition, cholesterol oralamethicin content and divalent cations. Biophys. J. 71, 1364-1373.
Lee, J. D., Kravchenko, V., Kirkland, T. N., Han, J., Mackman, N.,Moriarty, A., Leturcq, D., Tobias, P. S. and Ulevitch, R. J. (1993).Glycosyl-phosphatidylinositol-anchored or integral membrane forms ofCD14 mediate identical cellular-responses to endotoxin. Proc. Natl. Acad.Sci. USA90, 9928-9934
Lynn, W. A., Liu, Y. and Golenbock, D. T. (1993). Neither CD14 nor serumis absolutely necessary for activation of mononuclear phagocytes bybacterial lipopolysaccharide. Infect. Immun. 61, 4456-4461
Monks, C. R. F., Freiberg, B. A., Kupfer, H., Sciaky, N. and Kupfer, A.(1998). Three-dimensional segregation of supramolecular activation clustersin T cells. Nature395, 82-86.
Poltorac, A., He, X. L., Smirnova, I., Liu, M. Y., VanHuffel, C., Du, X.,Birdwell, D., Alejos, E., Silva, M., Galanos, C. et al. (1998). DefectiveLPS signaling in C3H/Hej and C57BL/10ScCr mice: mutations in TLR4gene. Science282, 2085-2088.
Pugin, J., Schurermaly, C. C., Leturcq, D., Moriarty, A., Ulevitch, R. J.and Tobias, P. S. (1993). Lipopolysaccharide activation of humanendothelial and epithelial cells is mediated by lipopolysaccharide-binding protein and soluble CD14. Proc. Natl Acad. Sci. USA90, 2740-2748
Pugin, J., Heumann, D., Tomasz, A., Kravchenko, V. V., Akamatsu, Y.,
JOURNAL OF CELL SCIENCE 114 (13)
CD14 Signal transductioncomplex containinghsp70 and hsp90
LPS
LPS
LPS
Fig. 8.Schematic representation of LPS transfer from CD14. LPStransfers from CD14 and is released in the lipid bilayer of themembrane before it reaches the signal transduction complex thatcontains hsp70 and hsp90.

2545Lateral diffusion of LPS receptors
Nishijima, M., Glauser, M. P., Tobias, P. S. and Ulevitch, R. J. (1994).CD14 is a pattern-recognition receptor. Immunity1, 509-516.
Schumann, R. R., Leong, S. R., Flaggs, G. W., Gray, P. W., Wright, S. D.,Mathison, J., Tobias, P. and Ulevitch, R. (1990). Structure and functionof lipopolysaccharide binding protein. Science249, 1429-1433.
Shapira, L., Takashiba, S., Amar, S. and Van Dyke, T. E. (1994).Porphyronomas gingivalis lipopolysaccharide stimulation of humanmonocytes: dependence on serum and CD14 receptor. Oral Microbiol.Immunol.9, 112-117.
Smith, P. R., Wilson, K. M., Morrison, I. E. G., Cherry, R. J. andFernandez, N. (1998). Imaging of individual cell surface MHC moleculesusing fluorescent particles. In MHC Biochemistry and Genetics. (eds N.Fenandez and G. Butcher), pp. 133-151. Oxford University Press.
Theofan, G., Horwitz, A. H., Williams, R. E., Liu, P., Chan, I., Birr, C.,Carroll, S. F., Meszaros, K., Parent, J. B., Kasler, H. et al. (1994). Anamino-terminal fragment of human lipopolysaccharide-binding proteinretains lipid A binding but not CD14-stimulatory activity. J. Immunol. 152,3623-3629.
Tobias, P. S., Soldau, K. and Ulevitch, R. J. (1986). Isolation of alipopolysaccharide-binding acute phase reactant from rabbit serum. J. Exp.Med. 164, 785-793.
Tohme, Z. N., Amar, S. and Van Dyke, T. E. (1999). Moesin functions as alipopolysaccharide receptor on human monocytes. Infect. Immun. 67, 3215-3220.
Triantafilou, K., Triantafilou, M. and Fernandez, N. (2000a).Lipopolysaccharide labelled with Alexa 488 Hydrazide as a novel probe forLPS binding studies. Cytometry41, 316-320
Triantafilou, K., Triantafilou, M. and Wilson, K. M. (2000b).Phycobiliprotein-Fab conjugates as probes for single particle fluorescenceimaging. Cytometry41, 226-234
Triantafilou, M., Triantafilou, K. and Fernandez, N. (2000c). Rough andsmooth forms of fluorescein-labelled bacterial endotoxin exhibit CD14/LBPdependent and independent binding that is influenced by endotoxinconcentartion. Eur. J. Biochem. 267, 2218-2226.
Troelstra, A., Antal-Szalmas, P., De Graaf-Miltenburg, L. A., Weersink,A. J., Verhoef, J., Van kessel, K. P. M. and Van Strijp, J. A. G. (1997).Saturable CD14-dependent binding of fluorescein-labelledlipopolysaccharide to human monocytes. Infect. Immun. 65, 2272-2277.
Viriyakosol, S. and Kirkland, T. N. (1995). A region of human CD14required for lipopolysaccharide-binding. J. Biol. Chem. 270, 362-368.
Weidemann, B., Brade, H., Rietschel, E. T., Dziarski, R., Bazil, V.,Kusumoto, S., Flad, H. D. and Ulmer, A. J. (1994). Soluble peptidoglycan-induced monokine production can be blocked by anti-CD14 monoclonal-antibodies and by lipid-A partial structures. Infect. Immun. 62, 4709-4715.
Wolf, D. E., Edidin, M. and Dragsten, P. (1980). The effect of bleaching lighton measurements of lateral diffusion in cell membranes by the fluorescencephotobleaching recovery method. Proc. Natl. Acad. Sci. USA 77, 2043-2045.
Wright, S. D., Tobias, P. S., Ulevitch, R. J. and Ramos, R. A. (1989).Lipopolysaccharide (LPS) binding protein opsonises LPS-bearing particlesfor recognition by a novel receptor on macrophages. J. Exp. Med. 170, 1231-1241
Wright, S. D., Ramos, R. A., Tobias, P. S., Ulevitch, R. J. and Mathison,J. C. (1990). CD14, a receptor for complexes of lipopolysaccharide (LPS)and LPS binding-protein. Science249, 1431-1433.




















