
Final Total - HUG
54
Influenza surveillance in Switzerland Sentinella study Winter Season 2004 – 2005 National Centre of Influenza Central Laboratory of Virology University Hospitals of Geneva Geneva, Switzerland FEDERAL OFFICE OF PUBLIC HEALTH BERNE - SWITZERLAND
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Final Total - HUG

1/54
Influenza surveillance in Switzerland Sentinella study
Winter Season 2004 – 2005
National Centre of Influenza
Central Laboratory of Virology University Hospitals of Geneva
Geneva, Switzerland
FEDERAL OFFICE OF PUBLIC HEALTH BERNE - SWITZERLAND

2/54
National Centre of Influenza
Central Laboratory of Virology, University Hospital of Geneva 24, rue Micheli-du-Crest, 1211 GENEVA 14 – SWITZERLAND
Dr Yves THOMAS & : +41/22 372 40 81 Fax: +41/22 372 40 88 , : [email protected]
Dr Laurent KAISER & : +41/22 372 40 96 , : [email protected]
Dr Werner WUNDERLI & : +41/22 372 40 86

3/54
Table of content 1. Acknowledgements .....................................................................................................................4 2. Summary - Résumé - Zusammenfassung...................................................................................5
2.1. Summary...............................................................................................................................5 2.2. Résumé..................................................................................................................................6 2.3. Zusammenfassung................................................................................................................7
3.1. Introduction...............................................................................................................................8 4. Method of detection for respiratory viruses...............................................................................8
4.1. Clinical identification of influenza cases............................................................................8 4.2. Detection of respiratory viruses...........................................................................................9 4.3. Characterization of influenza viruses................................................................................11 4.4. Identification of influenza A (H5N1) virus ......................................................................12
5. Results ........................................................................................................................................15 5.1. Viral detection ....................................................................................................................15 5.2. Characterization of the influenza viruses..........................................................................18
5.2.1. Influenza A (H3N2) ....................................................................................................18 5.2.2. Influenza A (H1N1) ....................................................................................................22 5.2.3. Influenza B ..................................................................................................................24
5.3. Patients with influenza infection .......................................................................................25 5.3.1. Frequency of viruses detected in a particular age group ..........................................25 5.3.2. Clinical data of patients infected with influenza.......................................................26
5.4. Evolution of the epidemic in Europe.................................................................................28 5.5. Influenza A/H2N2 ..............................................................................................................30 5.6. Avian influenza...................................................................................................................31
5.6.1. Avian influenza epidemics in South East Asia, 2004-2005 .....................................31 5.6.2. Detection of avian influenza viruses in Switzerland.................................................32
6. Recommended composition of the 2004/05 influenza vaccine ..............................................36 7. Discussion ..................................................................................................................................37 8. Bibliography...............................................................................................................................41 Annexe 1.........................................................................................................................................43 Annexe 2 : Detection of respiratory viruses during the 2004/05 season....................................44 Annexe 3 : Antigenic analysis of influenza A (H3N2)-like strains............................................45 Annexe 4 : Antigenic analysis of influenza A (H1N1)-like strains............................................51 Annexe 5 : Antigenic analysis of influenza B-like strains ..........................................................52 Annexe 6 : Alignement de l’hemagglutinine de souches récentes influenza A (H5N1) ..........54

4/54
1. Acknowledgements
We would like to thank all members of the Sentinella network, in particular the
practitioners who actively participated in the identification of patients, provided data,
and took samples. This report is the result of a close collaboration with the Swiss
Federal Office of Public Health and we should like to especially thank Dr Reto
Hagmann for his continued support and active participation.
Our activity is equally dependent upon a collaboration with other international
centres, notably the European Influenza Surveillance Scheme (EISS) network and
the World Health Organization (WHO), and we should like to thank Dr Alan Hay, Dr
Lin Yi Pu and Mrs Vicki Gregory, members of the WHO reference centre in London
(MRC, UK). Certain developments have been carried out in close collaboration with
Dr Olav Hungnes, Oslo, Norway. Finally, we would like to thank all members of the
Central Laboratory of Virology, University Hospitals of Geneva, in particular, Sabine
Nobs-Grunenwald for her valuable work and without whom this report would not have
been possible.

5/54
2. Summary - Résumé - Zusammenfassung
2.1. Summary
The 2004 -2005 influenza epidemic was of moderate intensity and characterized by a
rather late onset in the season. Our surveillance activity began in September 2004
and terminated at the end of April 2005 after a period of 31 weeks. The first sporadic
cases were identified in December and the detection peak was reached at week 5 in
2005 with an epidemic peak which stretched over 7 to 8 weeks until mid-March 2005.
Eight hundred and thirty-five samples were analysed, all in patients presenting with
an influenza syndrome. Of these, 345 (41%) were positive for a respiratory virus by
cell culture. The overwhelming majority (90%) was an influenza virus in the form of
type A for 268 (87%) cases, and type B in 41 (13%). Among these influenza viruses,
73% were of subtype A (H3N2) and 11%, A (H1N1). Characterization of influenza A
viruses by hemagglutination inhibition assay showed that these viruses were
antigenically related to the influenza reference strain A/Shantou/1219/2004 (H3N2),
itself very closely related to the A/California/7/2004 (H3N2) strain. These strains are
recently-appeared variants which can be distinguished antigenically from the 2004-
2005 vaccine strain A/Fujian/411/2002 (H3N2) vaccine strain. The influenza A virus
(H1N1) which predominated this year was antigenically close to the 2004-2005
vaccine strain A/New Caledonia/20/99 (H1N1). The influenza B strains detected
during this season belonged to two groups. Forty-four percent were antigenically
close to the influenza B strain included in the 2002-2003 season vaccine, and 46%
were antigenically close to the strain included in the 2004-2005 vaccine. Finally, three
influenza B strains showed a decrease of antigenic recognition with the overall
antisera used and were unable to be definitively identified at present.

6/54
2.2. Résumé
L'épidémie grippale 2004 – 2005 a été modérée en intensité et d'une apparition
assez tardive dans la saison. Notre surveillance a débuté en septembre 2004 pour
s'achever fin avril 2005 après une période de 31 semaines. Les premiers cas
sporadiques ont été identifiés dès le mois de décembre et le pic de détection a eu
lieu dans la semaine 5 de 2005, avec un pic épidémique qui s'est étendu sur environ
7 à 8 semaines jusqu'à mi-mars 2005. 835 échantillons ont été analysés, tous chez
des patients présentant un syndrome grippal. 345 de ces échantillons (41%) se sont
révélés positifs pour un virus respiratoire par culture. L'immense majorité, soit 90%,
était un virus grippal distribué sous forme de virus influenza de type A pour 268
(87%) et influenza de type B pour 41 (13%). Parmi les virus influenza, 73% des virus
étaient de sous-type A (H3N2) et 11% A (H1N1). La caractérisation des virus
influenza A (H3N2) par inhibition de l'hémagglutination a montré que ces virus étaient
antigéniquement proches de la souche de référence influenza A/Shantou/1219/2004
(H3N2) elle-même très proche de la souche influenza A/California/7/2004 (H3N2).
Ces souches sont des variants récemment apparus qui se distinguent
antigéniquement de la souche vaccinale 2004-2005 influenza A/Fujian/411/2002
(H3N2). Les virus influenza A (H1N1) mis en évidence cette année étaient
antigéniquement proches de la souche vaccinale 2004-2005 influenza A/New
Caledonia/20/99 (H1N1). Les souches influenza B détectées au cours de cette
saison appartenaient à deux groupes. 44% étaient antigéniquement proches de la
souche influenza B incluse dans le vaccin de la saison 2002-2003 et 46% étaient
antigéniquement proches de la souche incluse dans le vaccin 2004-2005 et
finalement 3 souches influenza B ont montré une diminution de reconnaissance
antigénique avec l’ensemble des antisera utilisés et n'ont pas pu être définitivement
caractérisés à ce jour.

7/54
2.3. Zusammenfassung
Die Grippe Epidemie 2004/2005 war von einer mässigen Intensität und begann
ziemlich spät. Die Überwachung der Grippe wurde im September 2004 gestartet und
wurde Ende April 2005 nach 31 Wochen beendet. Die ersten sporadischen Fälle von
Grippe wurden im Dezember nachgewiesen und das Maximum der Epidemie lag in
der 5. Woche (2005). Die Epidemie dauerte etwa 7 – 8 Wochen und war Mitte März
beendet.
835 Proben von Patienten mit Symptomen einer Grippe wurden untersucht. Davon
enthielten 345 Proben (41%) ein mittels Kultur nachweisbares respiratorisches Virus.
Die grosse Mehrheit der positiven Proben, d.h. 90% enthielten ein Grippevirus wovon
268 (87%) Influenza A und 41 (13%) Influenza B enthielten. Von den Influenza A
Viren gehörten 73% zum Subtyp A (H3N2) und 11% zum Subtyp A (H1N1).
Die Charakterisierung der Influenza A (H3N2) mit dem Hämagglutinationshemmtest
ergab, dass die Mehrheit der Viren verwandt waren mit dem Referenz Stamm
Influenza A/Shantou/1219/2004. Dieser Stamm selbst ist nahe verwandt mit
Influenza A/Kalifornien/7/2004 (H3N2). Diese zwei Varianten sind erst kürzlich
aufgetreten und unterscheiden sich von dem im Impfstoff (2004/2005) enthaltenen
Influenza A/Fujian/411/2002 (H3N2).
Die diese Jahr nachgewiesenen Influenza A (H1N1) waren mit dem im Impfstoff
enthaltenen Virus Influenza A/Neu/Kaledonien/20/99 (H1N1) verwandt.
Die Influenza B Viren welche diese Saison nachgewiesen werden konnten, gehörten
zwei verschiedenen Gruppen an. 44% waren verwandt mit dem Influenza B Virus
welcher im Impfstoff der Saison 2002/2003 enthalten war und 46% waren verwandt
mit dem Stamm welcher im Impfstoff der Saison 2004/2005 enthalten war. 3
Influenza B Viren zeigten eine reduzierte Affinität mit den geprüften Antiseren. Diese
Stämme konnten bis jetzt nicht definitiv charakterisiert werden.

8/54
3.1. Introduction
The Sentinella surveillance network relies upon more than 200 general practitioners
and paediatricians distributed throughout Switzerland to identify patients in their
practice with influenza-like illness as defined by strict clinical criteria. This definition is
based on the presence of fever associated with at least one additional respiratory
symptom. Respiratory sampling, either a pharyngeal or a nasopharyngeal swab, is
carried out weekly in a sub-group of patients and sent to the National Influenza
Centre for the identification of an eventual influenza virus by cell culture. The main
target of the virologic detection system used is influenza virus, but it is also capable
of detecting other respiratory viruses. Once identified, the influenza virus undergoes
a series of tests to determine its sub-type and antigenic characteristics, and to
determine if the circulating strains are covered by the vaccine strains.
Apart from the human circulating influenza A and B strains, the 2004-2005 season is
once again marked by the risk of an avian flu pandemic. To date, no case has been
identified outside south-east Asia, but the surveillance laboratories must be equipped
with the appropriate molecular diagnostic tools in readiness to diagnose the
emergence of a new avian influenza virus in persons returning from regions where
this virus is circulating.
4. Method of detection for respiratory viruses
4.1. Clinical identification of influenza cases
Clinical identification is based on the network of practitioners in the community who
document during their daily consultations the total number of flu syndromes observed
compared with the total number of consultations carried out. The case definition used
for a flu syndrome is the presence of fever greater than 38° with or without an
impression of sickness, myalgia, or a change of general state. In addition to fever, an
acute respiratory symptom such as cough or rhinorrhea must be present. Among the
cases which meet the clinical criteria definitions, a sub-group of practitioners select

9/54
each week up to two cases in which a nasopharyngeal or pharyngeal swab is taken
for despatch to the reference laboratory. These samples are placed in viral transport
medium and mailed to the centre the same day. The geographical distribution of the
subgroup of practitioners who take swabs is shown in Figure 1.
4.2. Detection of respiratory viruses
Detection of respiratory viruses is carried out according to the procedure shown in
Figure 2. Three different cell lines are inoculated (MDCK, LLC-MK2 et A549) at
different temperatures (37° and 33°C). After three days of incubation, screening of
cultures by monoclonal antibodies capable of detecting respiratory viruses influenza
A and B, parainfluenza 1, 2 and 3, adenovirus and respiratory syncytial virus is
conducted. A positive result leads to further, more specific screening to identify the
virus family concerned.
Figure 1 : Geographical distribution of the participants in the Sentinella network who do the sampling. Each participant is represented by a white dot. The region is distinguished by the number and the colour. Our country is divided in 6 regions comprising different number of cantons.
I
II
III
IV
V
VI

10/54
Patient with symptoms
Nasal and throat swab (practitioner)
Samples shipped to the laboratory
Inoculation on cell cultures
(33 and 37°C)
Day 7 Screening with
monoclonal antibodies (immunofluorescence)
Negative Positive
Characterisation with Monoclonal antibodies
influenza A, B Parainfluenza-virus 1, 2, 3, virus respiratory syncytial-virus,
adenovirus Hemagglutination Test (HA)
HA titre < 1/8 HA titre ≥ 1/8
Re-inoculation of the sample on Determination of the type (A,B) and cell culture, to increase the HA titre subtype (A/H1N1, A/H3N2); Comparison of the antigenic similarities with the vaccine strains by the hemagglutination inhibition test; Distinction of Influenza A (H1N1 and Figure 2 : Procedure applied for the detection of respiratory viruses by cell culture
H1N2) by real-time PCR

11/54
4.3. Characterization of influenza viruses
When an influenza virus is positively identified by cell culture, it is characterized by a
hemagglutination inhibition assay technique with specific antisera from immunised
ferrets. The results from this typing will be interpreted according to an antigenic table
adapted and established at the beginning of each influenza season with the aim to
use the antisera of strains which can be reasonably assumed to be circulating in the
human population during the season under study. The choice of these antisera is
equally essential in order to be able to differentiate the sub-types circulating and their
antigenic relation with the strains included in the vaccine. The titers obtained with
each clinical strain are identified and compared with standard antisera and allow a
precise identification of its antigenic behaviour. The criteria established for the 2004-
2005 season are described in table 1. As described in the legend to the table, the
homologous titers allow a comparison with the titers obtained with the circulating
influenza strains and to define the antigenic relation of these strains. This technique
permits an identification of the antigenic variations present on the hemagglutinin (HA)
which is an essential target of the immune and vaccine response. For example, in the
table illustrating the influenza A (H3N2) virus, it can be observed that the influenza
strains A/Wyoming/3/2003, A/Wellington/1/2004 and A/California/7/2004 show an
inhibition of titers comparable with antisera targeted against the influenza strain
A/Wyoming/3/2003. However, this technique does not permit to observe antigenic
variations related to the neuraminidase (NA) and it is thus impossible to differentiate
viruses with the same HA but with a different NA (e.g., H1N1 and H1N2 viruses). This
is particularly important as these viruses have circulated in Switzerland during the
last few years. To overcome this problem, we have developed a RT-PCR capable of
identifying an influenza virus type H1 containing a NA of type N1 or N2. This tool is
now applied systematically when a type H1 virus is identified.
Real-time RT-PCR techniques available in our laboratory allow to identify influenza
viruses A and B and to discriminate viruses of types N1 and N2 (annexe 1, van
Elden, 2001). Although more sensitive than cell culture, these techniques are not
used systematically for the routine detection of influenza viruses, but only to confirm
cell culture or in specific cases. To date, even molecular techniques used for

12/54
sequencing do not allow to discriminate precisely antigenic behaviour and thus the
phenotype of similar viruses on a phylogenetic level. Hence, the inhibition of
hemagglutination assay remains the reference technique for the identification of
antigenic variations.
4.4. Identification of influenza A (H5N1) virus
For almost 2 years, the H5N1 virus has been circulating mostly in the avian batteries
of south-east Asia and is continuing nowadays (WHO, 17.06.2005). Several tens of
human cases have been documented with a massive mortality leading to rapid death
by diffuse pulmonary attack. At present, there exists a probable case of inter-human
transmission (Ungchusak, 2005) but this phenomenon remains limited and, to date,
the virus does not appear to have acquired the essential characteristics to support an
inter-human chain of transmission to efficiently spread in man. However, the threat of
this risk has to be taken seriously in consideration of the exponential aspect of the
epidemic in the avian world, the increasing number of human cases observed, and
the increasing number of countries reporting cases coupled with the difficulty to
rapidly identify patients in regions where medical and laboratory infrastructures are
limited.
For this reason, it is essential that each reference laboratory may be able to propose
diagnostic tools to allow the rapid identification of a suspected case. RT-PCR is the
preferred technique of choice for an initial screening of a suspected case. It allows to
avoid the culture of a virus which is potentially extremely dangerous and,
theoretically, also permits a more rapid diagnosis. Cell culture as well as molecular
diagnostic techniques are confronted with several unknown factors concerning this
virus. The phenotypic and antigenic variability of the influenza A (H5N1) virus can
make its identification difficult even with established techniques, and it is therefore
essential that the international surveillance networks for influenza virus share their
most recent data. For detection by molecular tools, RT-PCR techniques require
knowledge of the sequence of the targeted genome. Hence, the genomic sequencing
of viruses detected in the various countries where cases are observed must

13/54
Table 1 : Hemagglutination inhibition (IHA) of the titer of reference influenza strains incubated with each of the reference antisera. The IHA titer obtained after incubation of a given strain with its corresponding antiserum is mentioned in bold. This value is called the homologous titer (HT). The titer obtained with a strain isolated from a sentinella sample (Sen) is then compared with the HT titer. If the ratio Sen/HT is low, the strain is considered as antigenically related to the reference strain. If the ratio is high, the strain is considered as antigenically different from the reference strain. Les 2 lignées principales des virus influenza B sont indiquées en Bleu. Yam : Yamagata-like ; Vict : Victoria-like.
Influenza A (H3N2)
Antisera
Strains A/Panama/2007/99 A/Wyoming/3/03 A/Shantou/1219/04 A/Wellington/1/04 A/California/7/04
A/Panama/2007/99 5120 640 80 640 < 160
A/Wyoming/3/2003 640 5120 640 1280 2560
A/Shantou/1219/2004 320 640 2560 1280 320
A/Wellington/1/2004 320 5120 2560 2560 1280
A/California/7/2004 < 160 5120 2560 320 2560
Influenza A (H1N1)
Antisera
Strains A/N.Caledonia/20/99 A/Beijing/262/95 A/Madag./57794/00 A/Egypt/96/02
A/N.Caledonia/20/99 20480 640 10240 2560
A/Beijing/262/95 5120 10240 2560 1280
A/Madagascar/57794/2000 5120 320 2560 2560
A/Egypt/96/2002 (H1N2) 5120 640 5120 5120
Influenza B Antisera
Strains B/Jiangsu/ 10/2003
B/Sichuan/
379/99
B/Harbin/ 7/94
B/Beijing /184/93
B/HK/ 335/01
B/Shandong/ 7/97
B/Brisbane/32/02
B/Jiangsu/10/2003 10240 1280 640 320 160 < 160 < 160 B/Sichuan/379/99 < 160 256 1280 320 640 < 160 < 160
B/Harbin/7/94 < 160 1280 5120 640 2560 320 < 160
B/Beijing/184/93 < 160 1280 5120 2560 1280 160 < 160
B/HK/335/2001 < 160 < 160 640 < 160 2560 10240 10240
B/Shandong/7/97 < 160 < 160 < 160 < 160 320 10240 2560
B/Brisbane/32/2002 < 160 < 160 160 < 160 320 2560 2560
Yam
Vict

14/54
imperatively be made available to all laboratories participating in global surveillance
as rapidly as possible. This is not the case at present, despite the existence of an
international influenza surveillance network under the auspices of the WHO. A further
difficulty is to maintain a register of non-infectious viral RNA types or plasmids from
the most recent cases. When faced with such an epidemic, with a virus capable of a
rapid and perpetual mutation, or capable of reassortment, it is essential to adapt
diagnostic tools as fast as possible to be able to obtain pertinent data in time. The
technique developed and perfected in our laboratory relies on a RT-PCR targeting of
the HA of the H5N1 virus. We have three types of tests at our disposal which permit
to detect avian influenza viruses having circulated recently either in the avian world or
in human cases. The controls used are DNA from RT-PCR derived from the genomic
RNA of human or avian influenza viruses and plasmids encoding for the HA of
viruses from human cases infected in Viet Nam during 2004.
To facilitate the availability of a control to allow the development of these diagnostic
tests, a bank of molecular material such as positive controls is in the process of being
set up under the auspices of the EISS. In this way, these controls can be distributed
to European laboratories participating in flu surveillance. Within this context, we have
produced plasmids based on influenza A (H5N1) virus sequences. Beyond the
technical aspects, the identification strategy of suspected cases of influenza A
(H5N1) virus is based not only on the identification of initial clinical cases, but also on
the capacity of laboratories to propose an alternative diagnosis. Therefore, we
propose to carry out a series of molecular analyses for each sporadic suspected case
which allows to identify both all influenza viruses circulating and all human respiratory
viruses, thus providing clinicians and health authorities with an alternative diagnosis
in the event of a clinical suspicion.
At last, we have at our disposal a specific antiserum targeted against the human
influenza strain A/Hong Kong/156/97 (H5N1). This antiserum has been provided by
the WHO to all national reference laboratories participating in the influenza
surveillance network. This reactive can be used on a sample which would have
escaped all recognition by standard antisera and on which would cause a problem of
suspected infection by an influenza A (H5N1) virus. The antigenic table of this
antiserum is given in Table 2.

15/54
Table 2 : Hemagglutination inhibition titer of influenza A (H5N1) inactivated virus with standard
influenza antisera
Standard Antisera (WHO) A/Hong Kong
156/97 (H5N1) A/Moscow
10/99
(H3N2)
A/Panama 2007/99 (H3N2)
A/Hong-Kong 1550/02 (H3N2)
A/N-Caledonia 20/99
(H1N1)
A/Egypt 96/02
(H1N2)
Inactivated
A (H5N1) strain (WHO 1997)
81920
< 80
< 80
< 80
< 80
< 80
5. Results
5.1. Viral detection
Surveillance began on 18 September 2004 and ended on 22 April 2005 after 31
weeks. Eight hundred and thirty-five samples were obtained from 56 Sentinella
practitioners, and a few samples were sent in by hospital laboratories. Of these, 309
influenza viruses were detected, representing a mean positive rate of 37% for the
whole season (Figure 3a). From this, we can conclude that almost 4 of 10 suspicions
of flu infection were confirmed. At the peak of the season, the proportion of flu
infection considerably increased and reached 57%. Among the 309 influenza viruses
detected, the majority was influenza A (H3N2) virus (73%), followed by influenza B
(13%) and influenza A (H1N1) virus (11%). Eight influenza A viruses could not be
sub-typed (3%). Thirty-six respiratory viruses from a family other than influenza virus
were also detected (10%; Figure 3b). Among these other viruses, respiratory
syncytial virus was isolated 14 times, adenovirus, 12 times, parainfluenza 1, 4 times,
parainfluenza 2, 2 times, and parainfluenza, 4 times (Figure 3b). Details of these data
are shown in the green table of the annexe 2.
Among the influenza viruses, the majority detected was of type A, 268 of 309 (87%).
Two hundred and twenty-five (73%) were of sub-type A (H3N2) and 35 (11%) of sub-
type A (H1N1). Of those remaining, 41 (13%) influenza type B were detected (Figure
3c).

16/54
a)41% positive
59% Negative
b)
3% ADV
1% PIV11 PIV2
90% Influenza
4% RSV1% PIV3
c)
73% inf. A/H3N2
13% inf. B
11% inf. A/H1N1
3% inf. A
Figure 3 : Proportion of nasopharyngeal samples positive for influenza or other respiratory viruses during the 2004/05 season (n = 835). a) Percentage of positive and negative samples received during the season. b) Percentage and type of different respiratory viruses detected. c) Percentage of type and subtype of influenza viruses detected (Inf. = influenza viruses)
345 positive
309 influenza viruses

17/54
During the first weeks of surveillance at the end of September 2004, no influenza
virus was detected. The first cases were detected at the end of November and
followed by sporadic but regular detections until week 48. From the last weeks of
December, there was a regular increase in positive cases in parallel to an increase in
medical consultations for flu syndromes. The detection rate peak was observed
during week 5 with 45 influenza viruses detected, which corresponded to 64% of all
positive samples collected (Figure 4). The medical consultation rate for flu syndrome
reached its maximum one week later with 61.7 flu consultations for 1000 medical
consultations. This was followed by a decrease both in the detection rate and the
number of medical consultations and passed under the epidemic index threshold
during week 11, being mid-March 2005 (Figure 4). The eight-week phase considered
as epidemic was rather short with medical consultations remaining above the level of
15 out of 1000. In comparison, the mean duration of flu epidemics observed during
the past 12 years has been nine weeks.
The kinetics of the circulating viruses were recorded for all six regions of Switzerland.
Medical contacts for influenza-like illness (MC-ILI) exceeded baseline values in all
regions during weeks 2 and 3 and reached the maximum value during weeks 6 and 7
(Figure 5). The different influenza viruses were detected in all regions. However, no
influenza A (H1N1) virus was detected in region 6, and no influenza B virus was
detected in region 2. The interpretation of this observation remains difficult as the
number of samples from these regions is too small. Thus, the flu epidemic in
Switzerland has been homogenous, as observed in all European countries.

18/54
0
10
20
30
40
50
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16
Weeks
Num
ber
of v
irus
es (n
)
0
10
20
30
40
50
60
70
MC
-ILI (
‰)
Other respiratory viruses
Inf. A (H1N1)
Inf.A
Inf. A (H3N2)
Inf. B
MC-ILI 2004/05 (‰)
Threshold (15 ‰)
Figure 4 : Type of viruses detected per week. inf. A (H1N1) : influenza A (H1N1) virus; inf. A (H3N2) : influenza A (H3N2) virus; Inf. A: influenza A viruses which were not subtyped, inf. B : influenza B virus. MC-ILI: medical contacts for influenza-like illness; threshold : percentage of medical contacts for influenza-like illness indicating the presence of an epidemic (15 ‰).
5.2. Characterization of the influenza viruses
5.2.1. Influenza A (H3N2)
Of 225 influenza A (H3N2) strains detected, only four strains showed an enhanced
recognition with the antiserum A/Wyoming/3/2003, the 2004-2005 vaccine strain
(annexe 3). In contrast, the great majority of strains demonstrated good recognition
with the antisera influenza A/California/7/2004 et A/Shantou/1219/2004 and are
therefore antigenically quite distinct from the vaccine strain (189/225, 84%). Finally,
21 influenza strains showed a decrease of the titer in inhibition of the
hemagglutination with the antisera overall. The influenza strains A/California/7/2004 and A/Shantou/1219/2004 are very close
antigenically and reactions are detected with their specific antisera (see Table 1).

19/54
02468
1012141618202224262830
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16
Positive samples (n)
0
10
20
30
40
50
60
70
80
90
100‰ M edical consult.
IBIA/H3N2IA/H1N1IA undet.Baseline%MC 2004/05
02468
1012141618202224262830
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16
Positive samples (n)
0
10
20
30
40
50
60
70
80
90
100‰ Medical consult.
IBIA/H3N2IA/H1N1IA undet.Baseline%MC 2004/05
Region 1 (GE, VD, VS, NE) Region 2 (BE, FR, JU)
02468
1012141618202224262830
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16
Positive samples (n)
0
10
20
30
40
50
60
70
80
90
100‰ Medical consult.
IBIA/H3N2IA/H1N1IA undet.Baseline%MC 2004/05
02468
1012141618202224262830
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16
Positive samples (n)
0
10
20
30
40
50
60
70
80
90
100‰ M edical consult.
IBIA/H3N2IA/H1N1IA undet.Baseline%MC 2004/05
Region 3 (BS, BL, AG, SO) Region 4 (LU, ZG, NW, OW, UR, SZ, GL)
02468
1012141618202224262830
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14
Positive samples (n)
0
10
20
30
40
50
60
70
80
90
100‰ Medical consult.
IBIA/H3N2IA/H1N1IA undet.Baseline%MC 2004/05
02468
1012141618202224262830
39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16
Positive samples (n)
0
10
20
30
40
50
60
70
80
90
100‰ M edical consult.
IBIA/H3N2IA/H1N1IA undet.Baseline%MC 2004/05
Region 5 (ZH, SH, TG, SG, AI, AR) Region 6 (TI, GR) Figure 5 : Number of influenza viruses detected per week and per region. A/H3N2 : influenza A (H3N2) viruses; IA/H1N1 : influenza A (H1N1) viruses; IA undetermined : influenza A virus that have not been subtyped; IB: influenza B viruses. ‰ MC: medical consultation for ILI

20/54
The genetic analysis confirms this small difference : there are less than 10
nucleotides of difference between the HA sequences of these two strains (Figure 6).
It is therefore reasonable to consider them as similar.
Among the 189 influenza A (H3N2) strains close to the influenza strains
A/Shantou/1219/2004 and A/California/7/2004, three populations can be
distinguished. The first is formed of strains which demonstrate significant cross-
reactions with three antisera: A/Shantou/1219/2004, A/Wellington/1/2004 and
A/Wyoming/3/2003 (86/189). The second population shows cross-reactions with the
antisera A/Shantou/1219/2004 and A/Wellington/1/2004, but a significant decrease of
recognition with the antiserum A/Wyoming/3/2003 (34/189). Finally, the third
population shows an affinity with the influenza antiserum A/Shantou/1219/2004 only
and a significant decrease of recognition with the antisera A/Wellington/1/2004 and
A/Wyoming/3/2003 (69/189). Hence, the two latter populations show a clear
decrease of recognition with the specific antiserum of the vaccine strain. This would
suggest that the influenza A (H3N2) strains have the possibility to escape or to be
less sensitive to the immune protection conferred by the 2004-2005.
The influenza antiserum A/California/7/2004 became available during the season and
allowed to distinguish between the influenza A (H3N2) strains. Sixty-one influenza A
(H3N2) strains were able to be analysed with this antiserum. Thirty-eight of 61 strains
were close to A/California/7/2004, and 19 were close to A/Shantou/1219/2004. Four
strains showed decreased titres with all the antisera used (annexe 3). This illustrates
that although similar, these two strains correspond to variants which are antigenically
slightly different.

21/54
Figure 6 : Phylogenetic comparison of nucleotide sequences encoding for H3 hemagglutinins
In conclusion, the influenza strains A/Shantou/1219/2004 and A/California/7/2004
represented those which predominantly circulated in Switzerland during the 2004-
2005 season. These recently-appeared variants differ antigenically from the strain
contained in the 2004-2005 vaccine, A/Wyoming/3/2003 (H3N2). Moreover, the
influenza strain A/California/7/2004 (H3N2) has been chosen to be included in the
2005-2006 influenza vaccine.

22/54
5.2.2. Influenza A (H1N1)
Thirty-five influenza A (H1N1) viruses were detected in Switzerland during this
session. Overall, these strains were antigenically close to the vaccine strain A/New
Caledonia/20/99 (H1N1) and to the most recent influenza strain
A/Madagascar/57794/2000 (H1N1) (annexe 4). Antigenic (Table 1) and genetic
(Figure 7) analyses show that these two strains are very close. The HA sequences of
these two strains are closely related (Figure 7 red arrows) ; in fact less than 10
nucleotides separate the HA of recently-detected strains worldwide (in blue at the top
of the Figure 7) with the influenza strain A/New Caledonia/20/99 (H1N1). For this
reason, the antigenic composition of influence A (H1N1) strains to be included in the
2005-2006 vaccine has not been modified.
No influenza A (H1N1) strain was close to the former vaccine strain A/Beijing/262/95
(H1N1) which circulated in Switzerland in 1995-1996. Moreover, no influenza A
(H1N2) strain has been detected in Switzerland this year. One patient was co-infected by an influenza A and an influenza B strain. The sample
contained both an influenza A (H1N1) and an influenza B strain. Unfortunately, the
presence of two strains in the same sample did not permit the use of the reaction to
hemagglutination inhibition to determine the relationship. The clinical sample was
sent to London to be studied in more depth. A RT-PCR reaction confirmed the
presence of the two viruses, the influence A (H1N1) virus being predominant. The
latter could be typed as being antigenically close to the influenza vaccine strain
A/New Caledonia/20/99 (H1N1).

23/54
Figure 7: Phylogenetic comparison of nucleotide sequences encoding H1 hemagglutinins of
human influenza viruses. Red arrows indicate the representative strains of influenza A (H1N1) that recently circulated.

24/54
5.2.3. Influenza B Two distinct lineages of influenza B strains have been regularly detected over the
past years in Switzerland, as in Europe : the lineage “Yamagata-like” and the lineage
“Victoria-like” (Figure 8). To be able to type circulating influenza B strains, antisera
specific for these two groups have been used (table 1).
Figure 8 : Phylogenetic comparison of nucleotide sequences encoding hemagglutinins of influenza B virus. Names in red indicate representative strains.
This year, the two lineages have been detected in equal proportions in Switzerland.
First, 44% (n=18) were antigenically close to the influenza strain B/Shandong/7/97,
itself very close to the influenza strain B/Hong Kong/330/2001 (Figure 8). This type of
strain has already circulated in Switzerland during the 2002-2003 season and has
even been included in the vaccine composition for that season. 46% (n=19) influenza
B viruses were antigenically close to influenza strains B/Jiangsu/10/2003 and
B/Sichuan/379/99, the first strain being included in the 2004-2005 vaccine (annexe
5). Among these, seven were closer to the most recent influenza strain
B/Jiangsu/10/2003 than the influenza B/Sichuan/379/99 one. Three influenza B
strains showed decreased titers with the reference antisera B/Shandong/7/97 and
Victoria-like
Yamagata-like

25/54
B/Sichuan/379/99 and are presently undergoing molecular analysis and a more
precise typing. Finally, an influenza B strain could not be typed as the sample came
from a patient co-infected by two influenza strains: an influenza B and an influenza A
(H1N1) strain.
5.3. Patients with influenza infection
5.3.1. Frequency of viruses detected in a particular age group The number of influenza viruses detected during the season has been stratified by
age group (Figure 9). As observed during past seasons, the majority of samples were
from the age groups “active” in the population, i.e., between 10 and 49 years old. The
younger age groups (less than 20 years old) do not appear to have been especially
affected this year in comparison to the 2003-2004 season where patients from this
group represented more than 40% of all those infected. In contrast, during this
season, this age group represented only 27% of all patients. In the age group of over
70 years old, the number of samples as well as the number of viruses detected is
higher than that observed in the groups of over 50 and 60 years old. This was also
observed during the 1997-1998 season. The season saw the appearance of an
antigenic variant of the influenza A (H3N2) virus : influenza A/Sydney/5/97. Thus, the
circulation of an antigenic variation close to the influenza strains
A/Shantou/1219/2004 and A/California/7/2004 (H3N2) appears to have had a more
severe effect than usual on those over 70 years old. This observation is confirmed by
the fact that the predominant virus in the age groups over 60 years old is an influenza
A (H3N2) virus. The influenza B virus infected indifferently all age groups. However,
influenza A (H1N1) virus infected mainly those less than 50 years old (Figure 9).
26/54
0
5
10
15
20
25
30
35
40
45
50
<= 1 2-4 5-9 10-19 20-29 30-39 40-49 50-59 60-69 70 <= ?
Age of patients
influ
enza
vir
uses
(n)
(H1N1)
(H3N2)
B
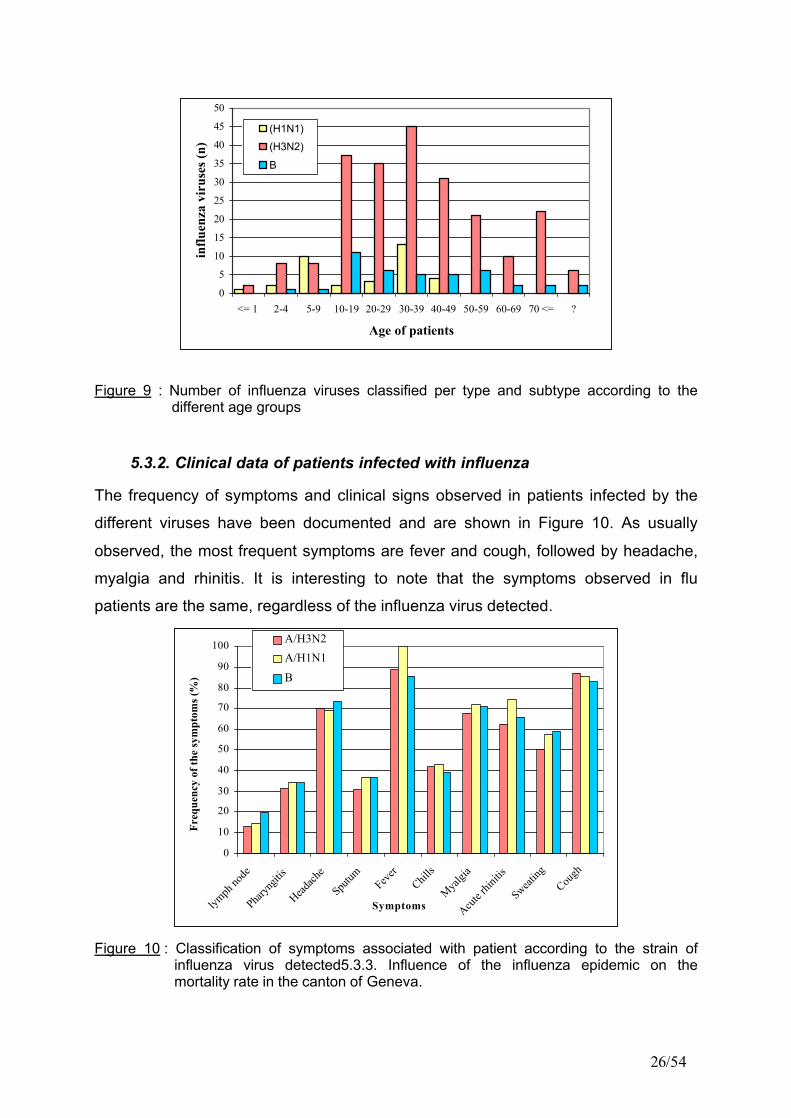
Figure 9 : Number of influenza viruses classified per type and subtype according to the different age groups
5.3.2. Clinical data of patients infected with influenza
The frequency of symptoms and clinical signs observed in patients infected by the
different viruses have been documented and are shown in Figure 10. As usually
observed, the most frequent symptoms are fever and cough, followed by headache,
myalgia and rhinitis. It is interesting to note that the symptoms observed in flu
patients are the same, regardless of the influenza virus detected.
0
10
20
30
40
50
60
70
80
90
100
lymph
node
Pharyn
gitis
Headach
e
Sputum Fev
erChil
ls
Myalgi
a
Acute
rhinit
is
Sweating
Cough
Symptoms
Freq
uenc
y of
the
sym
ptom
s (%
)
A/H3N2A/H1N1
B
Figure 10 : Classification of symptoms associated with patient according to the strain of influenza virus detected5.3.3. Influence of the influenza epidemic on the mortality rate in the canton of Geneva.

27/54
The elderly (over 60 years old) represent the main population at high risk of serious
complications following an influenza infection. To evaluate the severity of the 2004-
2005 influenza epidemic, we have analysed in particular the weekly mortality rate in
this age group in the canton of Geneva and compared these Figures with the weekly
rate of MC-ILI and the number of viruses detected in the canton (Figure 11). In
addition, the 2005 mortality rate has been compared with rates registered during the
seasons 1991 to 2004 and the mean of these weekly mortality rates has been
calculated plus two standard deviations. Results of these comparisons are shown in
Figure 11.
As observed at national level, the influenza epidemic in Geneva reached a peak
during week 6 : the rate of influenza viruses detected reached a maximum of 5/7
viruses detected during that week and the MC-ILIC value culminated at 11%. A
detailed analysis of the mortality rates for the following weeks showed no significant
increase when compared with the mean calculated for 1991-2004. Therefore, no
peak in mortality was observed consecutive to the influenza epidemic during the
2004-2005 season. This was not the case for the 2003-2004 season where eight
additional deaths in the canton of Geneva alone were observed one to two weeks
after the peak of MC-ILI (Thomas et al, 2004).

28/54
0
1
2
3
4
5
6
7
8
9
1 0
1 1
1 2
1 3
1 4
1 5
38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 202 0 0 4 W e e k 2 0 0 5
MC
-IL
I (%
) / V
irus
es n
umbe
r (n
)
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
1 1 0
1 2 0D
eath rate (n)
Medical consultations GE (%)
Influenza viruses detected (n)
Death rate >=60 years (n)
Threshold
Death rate Mean+2sd >= 60 years 1991-2004
Figure 11 : Influence of the influenza epidemic on the mortality in people older than 60 years in the canton of Geneva during the 2004/05 season.
Medical consultations GE (%): medical contacts for influenza-like symptoms in the canton of Geneva; Influenza viruses : influenza A and B viruses detected by the Sentinel network, in the canton of Geneva. Death rate : number of weekly deaths obtained from the records of the Register Office of the Canton of Geneva published by the Public Health Authorities of Geneva; Mean +2sd: mean number of the weekly deceased people older than sixty years-old + two standard deviation registered between 1991 and 2004
5.4. Evolution of the epidemic in Europe The influenza epidemic started at different times according to the various countries.
Some western European countries were the first to report outbreaks, such as Ireland
where the epidemic started relatively early (week 52, Figure 12). A few weeks later,
the epidemic culminated in the United Kingdom, Spain and Portugal (weeks 1 and 2,
Figure 12). Other countries such as France, Switzerland and Italy followed with a
maximum activity registered during week 5 (beginning of February). Finally, countries
in the more eastern part of Europe observed an epidemic which peaked during
weeks 6 and 7 (Slovenia, The Netherlands and The Czech Republic), week 8
(Norway) and week 10 (Germany).

29/54
Figure 12 : Detection of influenza viruses in European countries according to the week of sampling. Inf.A : influenza A viruses; Inf. A H1 (influenza A (H1N1) or influenza A (H1N2)); Inf B : influenza B virus; Inf A untyped : influenza A virus not subtyped; Inf A H3 : influenza A (H3N2). Data obtained from the European Influenza Surveillance Scheme, http://www.eiss.org.
Ireland
Spain
France
Slovenia
Norway

30/54
As in Switzerland, it was observed that several different types of influenza viruses
were circulating simultaneously: influenza A (H3N2 and H1N1) viruses and influenza
B viruses. Influenza A (H3N2) viruses circulated mainly in all European countries
apart from Slovenia where the influenza B was predominant. They were antigenically
close to the influenza A/California/7/2004 and A/Wellington/1/2004 strains. Influenza
A (H1N1) viruses were also detected in several European countries but in small
quantity. All were antigenically close to the influenza vaccine strain A/New
Caledonia/20/99 (H1N1). Influenza B viruses circulated quite significantly in almost all
European countries (Figure 12). The two main lineages of the influenza B virus were
detected in Europe during the 2004-2005 season. However, the proportions varied
according to the country. Viruses antigenically close to the influenza
B/Jiangsu/10/2003 strain and belonging to the Yamagata-like lineage (Figure 8)
mainly circulated in western and northern Europe (Portugal, France, Ireland, United
Kingdom, Norway...). In contrast, viruses antigenically close to the influenza
B/Shandong/7/97 strain belonging to the Victoria-like lineage (Figure 8) were
observed mainly in countries in central and southern Europe (Germany, Switzerland,
Italy, Poland and Romania).
5.5. Influenza A/H2N2
In February 2005, the College of American Pathologists (CAP) sent quality controls to
laboratories. For this purpose, currently circulating influenza A (H3N2 and H1N1) and
B viruses are normally used. In October 2004, a panel of controls containing
influenza A (H2N2) virus was inadvertently distributed and subsequently identified by
a reference laboratory participating to the proficiency testing. This virus was similar to
influenza A (H2N2) viruses that circulated in humans in 1957-58 and caused the so-
called Asian influenza pandemic. This influenza A (H2N2) was easily transmitted
from human to human. It continued to circulate in humans and caused annual
epidemics until 1968 when it vanished after the emergence of influenza A (H3N2)
viruses that caused the next pandemic. Therefore, persons born after 1968 are
expected to have no or only limited immunity to influenza A (H2N2). This A (H2N2)
virus is not contained in current trivalent influenza vaccines. The control panel

31/54
containing this A (H2N2) virus was sent by the CAP to more than 3750 laboratories in
eighteen countries (http://www.who.int/csr/disease/influenza/h2n2_2005_04_12/en/).
Since 8 April, WHO has been working directly with the US Department of Health and
Human Services and the Centers for Disease Control and Prevention (CDC) as well
as the relevant Canadian health agencies to assure rapid destruction of the samples
containing the influenza A (H2N2) virus. In addition, WHO through its regional and
country offices contacted the Ministries of Health of the remaining 16 involved
countries and territories to support this international effort. On 21 April 2005, 15/16
countries confirmed the destruction of the samples. No human infection of laboratory
workers has been detected until now.
5.6. Avian influenza
5.6.1. Avian influenza epidemics in South East Asia, 2004-2005
In January 2004, WHO reported officially a large number of avian influenza
epidemics in the eight countries of South East Asia. More than 100 million animals in
breeding farms either died following these epidemics or were culled as a preventive
measure to check the evolution of the epidemic. On 10 March 2004, 35 human cases
had been reported in Viet Nam and Thailand alone. Of these, 24 died following an
infection by the virus (Tran et al, 2004, de Jong et al, 2005). On 19 July 2004, after a
silence of several months, new avian influenza epidemics were observed in several
batteries of Viet Nam, followed by Thailand. Human cases have been reported in
Thailand, Viet Nam and Cambodia, close to the Vietnamese border. This re-
appearance of the virus occurred in two additional phases (Table 3). The first ranged
from mid-July to the beginning of October 2004 with nine human cases and eight
deaths in Viet Nam and Thailand. The second and last phase began in mid-
December and still persists. As at 17 June 2005, this last epidemic phase had
caused 63 cases of human infection in Viet Nam and Cambodia; to date, 22 persons
have died (Table 3). The isolated viruses, both in sick animals and human victims,
were the influenza A (H5N1) viruses, genetically close to the viruses already detected
in previous years in the same region. (Li et al., 2004).

32/54
Table 3 : Cumulative number of confirmed human cases of avian influenza A (H5N1) reported to WHO (WHO, cumulative A/H5N1 cases)
5.6.2. Detection of avian influenza viruses in Switzerland
At the Swiss National Centre for Influenza, the detection of avian influenza virus is
conducted by RT-PCR. For the detection of influenza A (H5N1) virus, we have at our
disposal specific reactives of the sub-type A/H5. These reactives must be able to
recognise the viruses despite their evolution over time. For example, the influenza A
(H5N1) viruses that circulated in 1987 in human is genetically slightly different than
the one that circulated in 2003 (Horimoto et al., 2004, Li et al, 2004). To overcome
this phenomenon, three different combinations of primers have been chosen and
developed in Geneva. The hemagglutinin H5 of the virus has been targeted. Two
combinations are constituted of primers for a detection by Real-Time RT-PCR
(orange and green combinations, Figure 13), and one combination is constituted of a
primer for a classic PCR (blue combination, Figure 13).

33/54
Figure 13 : Schéma des régions de l’hemagglutinine de virus influenza A/H5 reconnues par les différents jeux de primers (les sondes centrales ne sont pas représentées). Les combinaisons oranges et vertes sont utilisées pour une PCR en real-time et les sondes bleues sont utilisées en PCR classique. These primer combinations are regularly tested with positive controls, if available. For
security reasons, no live influenza A (H5N1) virus strain is distributed to countries
outside the country of detection. For this reason, only the RNA and the DNA of the
viruses are distributed as positive controls. Our primer sequences are also regularly
verified by alignment with the genomic sequences of the influenza A (H5N1) virus
recently detected and available in data banks such as Genbank (National Institutes of
Health) and ISDB (Los Alamos National Laboratory). Indeed, it is essential to
guarantee the efficiency of our detection reactions despite the constant modification
of the influenza genome over time due to a well-known phenomenon of the influenza
viruses : the antigenic shift. The sequences of two influenza A (H5N1) viruses of
avian origin detected recently (January 2005) in Viet Nam have been provided by the
National Influenza Centre of the National Institute of Hygiene and Epidemiology
(NIHE) in Hanoi. First sample was collected in the South of Vietnam (annexe 6,
sequence 2) and second one in the North (annexe 6, sequence 3). The alignment of
these two sequences with other influenza A (H5N1) strains detected in humans since
1997 demonstrates a satisfactory conservation of the zones recognised by our
primers. In contrast, a site of three nucleotides is missing from the sequence of the
influenza virus sampled in the north of Viet Nam. This zone is also the target of one
of our primers, the HARev. Thus, the primer combination including the HARev may
not be able to detect these types of strains. This observation well illustrates the limits
H5 Clivage
site

34/54
of molecular technique detection. However, the two other primer combinations
demonstrate practically no difference with the new sequences. Theoretically, our set
of primers should be able to detect new strains of influence A (H5N1). However, a
more recent positive control is necessary to experiment and confirm this observation.
Unfortunately, at the present time, we have been unable to obtain such controls from
countries affected by the avian influenza epidemics.
We have also available heterogeneous positive controls to validate our reactives for a
large variety of influenza (H5N1) strains. This concerns DNA comprising the
sequence encoding pour hemagglutinin of two strains of avian origin having
circulated in 1984 and 1997 and a strain detected in a human case in Viet Nam in
2004. The results of the Real-Time RT-PCR reactions carried out on the human
strain control (Figure 14a) and on the avian strains (Figure 14b) demonstrate our
primer detection efficiency.
It is important to have available adequate, standard positive controls in sufficient
quantity to allow laboratories to develop a molecular detection test for the influenza A
(H5N1) virus. One of the tasks of the European group, EISS, is to help the
laboratories of the international surveillance network to achieve this objective. As an
active member of the EISS, Switzerland participates actively to the establishment of a
data bank of molecular material to enable access to positive controls for diagnostic
tests. Within this framework, the Central Laboratory of Virology at the University
Hospitals of Geneva, in collaboration with the Norwegian Institute of Public Health in
Oslo, has developed a positive control for PCR. This type of control can help
advance a large number of European laboratories to establish or develop a detection
technique either by classic PCR or a Real-Time RT-PCR. It can also be used to
create the standardisation of detection techniques between laboratories in the
community network and EISS (CNRL) reference laboratories (CNRL).

35/54
Combinaison 1 Combinaison 2 (HA5For, HA5Rev, HA5Pro) (H5VietFor, H5VietRev, H5VietPro) A/Duck/Potsdam/2243/84 H5N6 A/Duck/Singapore-Q/F119/97
Figure 14 : Detection of three different positive controls with two Real-Time RT-PCR primer combinations. Positive controls are constituted of plasmid containing a part of the hemagglutinin sequence of A/Vietnam/1203/2004 (H5N1) and DNA containing the encoding sequence for hemagglutinin for the A/Duck/Postdam/2243/84 (H5N6) and A/Duck/Singapore-Q/F119/97 (H5N3) viruses.
The construction consists of an amplification of the most variable part of the avian
virus genome, the entire encoded sequence of the hemagglutinin. This sequence has
been amplified from the influenza A/Duck/Singapore-Q/F119/97 (H5N3) virus. The
hemagglutinin shows a good conservation with the hemagglutinins of the avian
influenza A (H5N1) virus detected in humans. The results of the amplification
obtained by PCR were then introduced into a plasmid which allowed to largely
reproduce this construction into bacteria. A diagram of this construction is shown in
Figure 15. The next steps will be to also obtain the hemagglutinin of viruses detected
in human cases.
A/Vietnam/1203/04 (H5N1) dilutions 10-3, 10-4 et 10-5

36/54
Figure 15 : Schéma du vecteur porteur de la séquence de l’hemagglutinine H5 du virus
aviaire A/Duck/Singapore/F119-3/97 (H5N3). En noir est représenté la séquence
codante pour l’hemagglutinine HA5
6. Recommended composition of the 2004/05 influenza vaccine
The 25th of February 2005, WHO recommended that vaccines to be used in the
2005/06 influenza season and in the northern hemisphere contain the following
(WHO, 25.02.2005) :
- an influenza A/New Caledonia/20/99 (H1N1)-like virus
- an influenza A/California/7/2004(H3N2)-like virus a
- an influenza B/Shanghai/361/2002-like virus b
a A/New York/55/2004 is available as a vaccine virus bThe currently used vaccine viruses are B/Shanghai/361/2002, B/Jiangsu/10/2003 and B/Jilin/20/2003
HA5 1750 bps
M13
T7

37/54
7. Discussion
The intensity of the influenza epidemic observed this season was medium to high.
Different influenza viruses circulated in Switzerland namely influenza A (H3N2),
influenza A (H1N1) and influenza B. During the winter an influenza A (H3N2) virus
strain appeared which was antigenically distinct from the strain that circulated in
2003-2004 and which was included in the 2004-05 vaccine. Viruses which were
detected during the 2003/2004 season were antigenically close to influenza
A/Fujian/411/2002 and A/Wyoming/3/2003. This winter however, the influenza A
(H3N2) strains which circulated most frequently in Europe and in the United States
were antigenically related to influenza A/California/7/2004 and A/Shantou/1219/2004.
This means that an antigenic drift occurred which clearly could be detected with
inhibition of the hemagglutination assays. In Geneva, some infections with this virus
were observed in immuno-compromised patients vaccinated with the 2004-2005
influenza vaccine suggesting that the protection was diminished. The phylogenetic
analysis (Figure 6) shows that influenza A/California/7/2004 and influenza
A/Shantou/1219/2004 strains are distinguishable from the vaccine strains influenza
A/Fujian/411/2002 and A/Wyoming/3/2003.
The frequency with which the different influenza A (H3N2) circulated in Switzerland
since 1995 are represented in Figure 16. During the last ten years, a particular
influenza A (H3N2) virus circulated between 2 and 3 seasons on average before a
new variant showed up. Normally the new variant and the previous strain circulated
on parallel during a season before the older one disappeared. This was the case for
example for influenza A/Sydney75/97 (H3N2) (Figure 16) which appeared in 1997. It
became the dominating strain after influenza A/Wuhan/354/95 and
A/Johannesburg/33/94. In the same manner, influenza A/Fujian/411/2002 started to
circulate at the end of the 2002/03 season and became dominant during the following
season 2003-04.
The decision on the composition of the vaccine for the northern hemisphere takes
place in February each year. If a variant shows up later on during the season, the
vaccine might protect less against the new strain circulating during the following
season. The virus can then be transmitted more easily to vaccinated people. This

38/54
might have severe consequences. Such a situation was observed during the 1997-98
season where influenza A/Sydney/5/97 (H3N2) appeared. At that time influenza
A/Wuhan/354/95 was included in the 1997-98 influenza vaccine and as a
consequence gave only partial protection against A/Sydney/5/97 (H3N2). On parallel
high rates of medical consultations for influenza-like illness were observed. As a
consequence a higher mortality rate of the more than 60 years old people was
observed in the canton of Geneva (Thomas et al., 1998).
This year, the variant influenza A/California/7/2004 (Figure 16) was predominant from
the very beginning of the season. The vaccine contained the antigens from the
influenza A virus which circulated the previous years namely influenza
A/Fujian/411/2002 (Thomas et al, 2004). Although the epidemic was less severe than
the epidemic observed in 1997-98, it still was moderate to strong.
95/9
6
96/9
7
97/9
8
98/9
9
99/0
0
00/0
1
01/0
2
02/0
3
03/0
4
04/0
5
0102030405060708090
100
Influenza virus rate (%)
Influenza A (H3N2) virus
A/Thessaloniki/1/95 A/Johannesburg/33/94A/Wuhan/359/95A/Sydney/5/97A/Moscow/10/99A/Fujian/411/02A/California/7/2004
Figure 16 : Influenza A (H3N2) strains which circulated in Switzerland since 1995. Each colour represents a particular influenza strain. The values represent the rate of each strain in comparison to all influenza A strains detected during the season.
Influenza B virus were also observed this year. Two lineages of influenza B,
circulating already in previous years in Switzerland, were observed at the same
frequency this season. The two variants belonged to the Yamagata and to the
Victoria lineages. As one can see in Figure 17, influenza B strains belonging to the
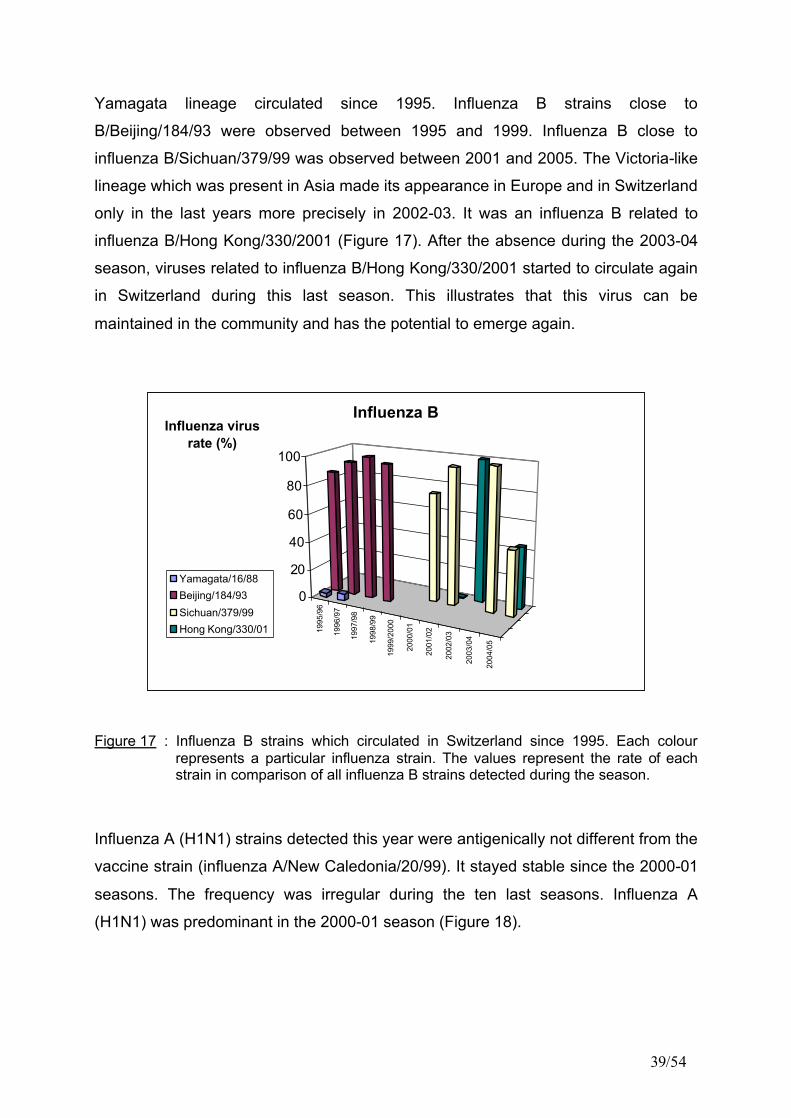
39/54
Yamagata lineage circulated since 1995. Influenza B strains close to
B/Beijing/184/93 were observed between 1995 and 1999. Influenza B close to
influenza B/Sichuan/379/99 was observed between 2001 and 2005. The Victoria-like
lineage which was present in Asia made its appearance in Europe and in Switzerland
only in the last years more precisely in 2002-03. It was an influenza B related to
influenza B/Hong Kong/330/2001 (Figure 17). After the absence during the 2003-04
season, viruses related to influenza B/Hong Kong/330/2001 started to circulate again
in Switzerland during this last season. This illustrates that this virus can be
maintained in the community and has the potential to emerge again.
1995
/96
1996
/97
1997
/98
1998
/99
1999
/200
0
2000
/01
2001
/02
2002
/03
2003
/04
2004
/05
0
20
40
60
80
100
Influenza virus rate (%)
Influenza B
Yamagata/16/88Beijing/184/93Sichuan/379/99Hong Kong/330/01
Figure 17 : Influenza B strains which circulated in Switzerland since 1995. Each colour represents a particular influenza strain. The values represent the rate of each strain in comparison of all influenza B strains detected during the season.
Influenza A (H1N1) strains detected this year were antigenically not different from the
vaccine strain (influenza A/New Caledonia/20/99). It stayed stable since the 2000-01
seasons. The frequency was irregular during the ten last seasons. Influenza A
(H1N1) was predominant in the 2000-01 season (Figure 18).

40/54
1995
/96
1997
/98
1999
/200
0
2001
/02
2003
/04
0
20
40
60
80
100Influenza virus
rate (%)
Influenza A (H1N1) viruses
Singapoore/6/86Texas/36/91Bayern/7/95Wuhan/371/95New Caledonia/20/99Egypt/96/2002Madagascar/57794/2000
Figure 18 : Influenza A (H1N1) strains which circulated in Switzerland since 1995. Each colour represents a particular influenza strain. The values represent the rate of each strain in comparison to all influenza A strains detected during the season.
Recent outbreaks of avian influenza among chickens show that, the influenza A
(H5N1) virus is endemic in Asia today: The reservoir is in aquatic birds which gives
the virus the possibility to infect regularly birds with terrestrial breeding. The increase
of the number of epidemics in poultry flocks increases the risk of the passage of the
avian influenza A (H5N1) virus to man. Such events are now more frequently
described. The virus caused numerous epidemics which was not the case until
recently. Eight countries in South Asia East were affected in spring 2004. Nowadays,
avian influenza A (H5N1) virus is still causing epidemics in poultry farms in Vietnam
and in Indonesia. In total 107 human victims were proven of whom 54 died (update
from WHO: June 17th, 2005). Until now infection of humans by the avian influenza A
(H5N1) virus was only observed after exposure to infected birds. No transmission
between humans could be proven. However, the risk exists that the virus adapts to
man and starts circulating. For this reason the National Centre of Influenza must be
able to detect such viruses in man. Different combinations of primers were developed
and tested for the detection of this avian influenza A (H5N1) virus. The virus is highly
variable a fact which requires a continuous verification and adaptation of the
reagents.

41/54
8. Bibliography
de Jong MD, Bach VC, Phan TQ, Vo MH, Tran TT, Nguyen BH, Beld M, Le TP, Truong HK, Nguyen VV, Tran TH, Do QH, Farrar J. Fatal avian influenza A (H5N1) in a child presenting with diarrhea followed by coma. N Engl J Med. 2005 Feb 17;352(7):686-91.
Horimoto T, Fukuda N, Iwatsuki-Horimoto K, Guan Y, Lim W, Peiris M, Sugii S, Odagiri T, Tashiro M, Kawaoka Y., Antigenic differences between H5N1 human influenza viruses isolated in 1997 and 2003. J Vet Med Sci. 2004 Mar; 66(3):303-5.
Li KS, Guan Y, Wang J, Smith GJ, Xu KM, Duan L, Rahardjo AP, Puthavathana P, Buranathai C, Nguyen TD, Estoepangestie AT, Chaisingh A, Auewarakul P, Long HT, Hanh NT, Webby RJ, Poon LL, Chen H, Shortridge KF, Yuen KY, Webster RG, Peiris JS. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature. 2004 Jul 8;430(6996):209-13.
Thomas Y., L. Kaiser, W. Wunderli. Influenza Surveillance in Switzerland, Sentinella study. Winter 2003-04. 2004.
Thomas Y., W. Wunderli. Influenza Surveillance in Switzerland, Sentinella study. Winter 1997-98, 1998.
Tran TH, Nguyen TL, Nguyen TD, Luong TS, Pham PM, Nguyen VC, Pham TS, Vo CD, Le TQ, Ngo TT, Dao BK, Le PP, Nguyen TT, Hoang TL, Cao VT, Le TG, Nguyen DT, Le HN, Nguyen KT, Le HS, Le VT, Christiane D, Tran TT, Menno de J, Schultsz C, Cheng P, Lim W, Horby P, Farrar J; World Health Organization International Avian Influenza Investigative Team. 2004, Avian influenza A (H5N1) in 10 patients in Vietnam. N Engl J Med. 2004 Mar 18;350(12):1179-88. Epub 2004 Feb 25.
Ungchusak K, Auewarakul P, Dowell SF, Kitphati R, Auwanit W, Puthavathana P, Uiprasertkul M, Boonnak K, Pittayawonganon C, Cox NJ, Zaki SR, Thawatsupha P, Chittaganpitch M, Khontong R, Simmerman JM, Chunsutthiwat S. 2005, Probable person-to-person transmission of avian influenza A (H5N1). N Engl J Med. 2005 Jan 27;352(4):333-40
Van Elden LJ, Nijhuis M, Schipper P, Schuurman R, van Loon AM, Simultaneous detection of influenza viruses A and B using real-time quantitative PCR; J Clin Microbiol 2001 Jan;39(1):196-200
WHO, 25.02.2005. Weekly epidemiological record, Recommended composition of influenza virus vaccine composition for use in the 2005-06 influenza season, WER 80, n° 8, p71-75.
WHO, 15.04.2005. Update: International response to the inadvertent distribution of H2N2 influenza virus: Destruction of virus panels proceeding, http://www.who.int/csr/disease/ influenza/h2n2_2005_04_15/en/index.html

42/54
WHO, 17.06.2005. Report of cumulative number of confirmed human cases of avian influenza A (H5N1) reported to WHO : http://www.who.int/csr/disease/avian_influenza/ country/cases_table_2005_06_17/en/index.html

43/54
Annexe 1
Primers and probes used in real-time PCR for the detection and characterisation of influenza viruses
Target virus Primers/ Probes
Sequence Target gene
Forward primer 5’- GGA CTG CAG CGT AGA CGC TT -3’ Reverse primer 5’- CAT CCT GTT GTA TAT GAG GCC CAT -3’ Influenza A
Probe 5’- CTC AGT TAT TCT GCT GGT GCA CTT GCC A -3’ Matrix
Forward primer 5’- AAA TAC GGT GGA TTA AAT AAA AGC AA -3’ Reverse primer 5’- CCA GCA ATA GCT CCG AAG AAA -3’ Influenza B
Probe 5’- CAC CCA TAT TGG GCA ATT TCC TAT GGC -3’ Hemagglutinin
Forward primer 5’- ATG GTA ATG GTG TTT GGA TAG GAA G -3’ Reverse primer 5’- AAT GCT GCT CCC ACT AGT CCA G -3’ Neuraminidase 1
(N1) Probe 5’- TGA TTT GGG ATC CTA ATG GAT GGA CAG -3’
Neuraminidase
Forward primer 5’- AAG CAT GGC TGC ATG TTT GTG -3’ Reverse primer 5’- ACC AGG ATA TCG AGG ATA ACA GGA -3’ Neuraminidase 2
(N2) Probe 5’- TGC TGA GCA CTT CCT GAC AAT GGG CT -3’
Neuraminidase
HA5For 5’- CCCAAATATGTGAAATCAAACAGATT-3’ HA5Rev 5’- CAAATAGTCCTCTCTTTTTTCTTCTTCTC-3’ HA5Pro 5’- TGCGACTGGACTCAGAAATACCCCTCA-3’
Hemagglutinin
H5VietFor 5’-GGA TGG CAG GGA ATG GTA GA-3’ H5VietRev 5’-TCTATTGCCTTTTGAGTGGATTCTT-3’
H5VietPro 5’-TGGGTACCACCATAGCAACGAGCAGG-3’ Hemagglutinin
WHO H5-1 5’-GCCATTCCACAACATACACCC-3’
Influenza A/H5
WHO H5-2 5’-TAAATTCTCTATCCTCCTTTCCAA-3’ Hemagglutinin
Forward primer 5’- GGC AAC AGG AAT GAA GAA TGT TCC-3’ Reverse primer 5’- AAT CAG ACC TTC CCA TCC ATT TTC-3’ Influenza A/H7
Probe 5’ AGG CCT ATT TGG TGC TAT AGC GGG TTT CAT -3’
Hemagglutinin

44/54
Annexe 2 : Detection of respiratory viruses during the 2004/05 season. MC-ILI: proportion of medical consultations for influenza-like illness; influ A: influenza A; not typed: influenza A which could not be sub-typed. Influ. B: influenza B; other resp. viruses: other respiratory viruses.
Week MC-ILI Samp. Influ. A Other resp. Total virus Total virus
03/04 Dates
(‰) rec. undet. H3N2 H1N1
Influ.B
viruses (n) (%)
39 18-sept-04 24-sept-04 1.1 10 1Para3 1 10
40 25-sept-04 01-oct-04 2 6 0 0
41 02-oct-03 08-oct-04 2 4 1Para1, 1ADV 2 50
42 09-oct-04 15-oct-04 1.2 11 0 0
43 16-oct-04 22-oct-04 1.2 7 0 0
44 23-oct-04 29-oct-04 1.9 6 0 0
45 30-oct-04 05-nov-04 2.1 14 0 0
46 06-nov-04 12-nov-04 2.5 22 0 0
47 13-nov-04 19-nov-04 2.8 33 1Para2 1 3
48 20-nov-04 26-nov-04 3.6 24 1 1VRS 2 8
49 27-nov-04 03-déc-04 2.4 20 1 2ADV 3 15
50 04-déc-04 10-déc-04 2.8 13 1 1Para1, 1Para3 3 23
51 11-déc-04 17-déc-04 3.2 24 1 1 4
52 18-déc-04 24-déc-04 4.3 35 2 1 5 2ADV, 1VRS 11 31
53 25-déc-04 31-déc-04 8.6 22 2 2 2 1ADV 7 32
1 01-janv-05 07-janv-05 9.5 35 12 2 2ADV, 3VRS,1Para1 20 57
2 08-janv-05 14-janv-05 9.4 33 5 3 8 24
3 15-janv-05 21-janv-05 16 42 1 19 1 2ADV, 1Para3,1VRS 25 60
4 22-janv-05 28-janv-05 25.9 61 1 22 6 2 1VRS 32 52
5 29-janv-05 05-févr-05 45.8 70 40 2 3 1Para1,1Para2 47 67
6 05-févr-05 11-févr-05 61.7 61 2 30 1 2 1VRS 36 59
7 12-févr-05 18-févr-05 59.5 71 2 33 5 1 1VRS 42 59
8 19-févr-05 25-févr-05 40.4 60 1 25 2 2 2VRS 32 53
9 26-févr-05 05-mars-05 25.8 51 1 15 4 1 1ADV 22 43
10 05-mars-05 11-mars-05 17.7 37 11 2 7 20 54
11 12-mars-05 18-mars-05 13.1 31 5 1 6 2VRS 14 45
12 19-mars-05 25-mars-05 7.2 16 1 2 5 1Para3 9 56
13 26-mars-05 01-avril-05 3.9 9 1 2 1ADV, 1VRS 5 56
14 02-avril-05 08-avril-05 2.1 5 1 1 20
15 09-avril-05 15-avril-05 0.8 1 1 1 100
16 16-avril-05 22-avril-05 2.1 1 0 0
Total 835 8 225 35
268 41
309 36 345 41 %

45/54
Annexe 3 : Antigenic analysis of influenza A (H3N2)-like strains
Antisera
A (H3N2) strains A/Shantou A/California A/Wellington A/Wyoming
A/Shantou/1219/04 (H3N2) 256 32 128 64
A/California/4/04 (H3N2) 256 256 32 512
A/Wellington/1/04 (H3N2) 256 128 256 512
A/Wyoming/3/03 (H3N2) 64 256 128 512
Date Seq N° Typisation A/Shan A/Cal A/Well A/Wyo
08-févr-05 7336 InfA H3N2 California/7/04 512 512 256 512
18-févr-05 7793 InfA H3N2 California/7/04 512 512 128 256
07-févr-05 7267 InfA H3N2 California/7/04 512 256 128 256
09-févr-05 7401 InfA H3N2 California/7/04 512 256 64 128
22-mars-05 9034 InfA H3N2 California/7/04 256 256 128 256
14-févr-05 7552 InfA H3N2 California/7/04 256 256 64 256
04-mars-05 8386 InfA H3N2 California/7/04 256 256 64 256
09-mars-05 8562 InfA H3N2 California/7/04 256 256 64 256
14-mars-05 8701 InfA H3N2 California/7/04 256 256 64 256
16-mars-05 8817 InfA H3N2 California/7/04 256 256 64 256
18-mars-05 8989 InfA H3N2 California/7/04 256 256 64 128
08-févr-05 7320 InfA H3N2 California/7/04 256 256 32 128
03-mars-05 8331 InfA H3N2 California/7/04 128 256 64 256
03-mars-05 8306 InfA H3N2 California/7/04 128 256 64 128
15-févr-05 7615 InfA H3N2 California/7/04 128 256 32 128
03-févr-05 7037 InfA H3N2 California/7/04 256 128 64 128
07-mars-05 8444 InfA H3N2 California/7/04 128 128 64 128
14-mars-05 8713 InfA H3N2 California/7/04 128 128 64 128
14-avr-05 9799 InfA H3N2 California/7/04 128 128 64 128
17-mars-05 8895 InfA H3N2 California/7/04 128 128 32 128
09-mars-05 8563 InfA H3N2 California/7/04 128 128 32 64
03-mars-05 8334 InfA H3N2 California/7/04 128 128 32 128
03-mars-05 8298 InfA H3N2 California/7/04 128 128 32 64
02-févr-05 6974 InfA H3N2 California/7/04 64 128 64 64
27-janv-05 6811 InfA H3N2 California/7/04 64 128 32 32
03-mars-05 8325 InfA H3N2 California/7/04 32 128 32 64
01-mars-05 8233 InfA H3N2 California/7/04 64 64 64 128
28-févr-05 8156 InfA H3N2 California/7/04 64 64 32 64
08-mars-05 8496 InfA H3N2 California/7/04 64 64 32 64
27-janv-05 6801 InfA H3N2 California/7/04 32 64 64 64
28-févr-05 8165 InfA H3N2 California/7/04 32 64 32 64
08-févr-05 7344 InfA H3N2 California/7/04 32 64 32 32
01-mars-05 8223 InfA H3N2 California/7/04 32 64 32 32

46/54
Date Seq N° Typisation A/Shan A/Cal A/Well A/Wyo
14-mars-05 8714 InfA H3N2 California/7/04 32 64 32 32
14-mars-05 8718 InfA H3N2 California/7/04 32 64 32 32
22-févr-05 7926 InfA H3N2 California/7/04 32 64 16 32
04-mars-05 8379 InfA H3N2 California/7/04 16 64 32 32
25-févr-05 8097 InfA H3N2 California/7/04 16 64 32 16
07-févr-05 7270 InfA H3N2 Shantou/1219/04 512 128 128 256
06-janv-05 5970 InfA H3N2 Shantou/1219/04 512 256 512
01-mars-05 8222 InfA H3N2 Shantou/1219/04 256 128 64 128
14-mars-05 8696 InfA H3N2 Shantou/1219/04 256 128 32 64
17-févr-05 7732 InfA H3N2 Shantou/1219/04 256 128 32 256
14-mars-05 8702 InfA H3N2 Shantou/1219/04 256 64 64 128
21-févr-05 7858 InfA H3N2 Shantou/1219/04 256 64 64 64
09-mars-05 8556 InfA H3N2 Shantou/1219/04 256 64 32 64
22-mars-05 9031 InfA H3N2 Shantou/1219/04 256 32 128 128
14-mars-05 8707 InfA H3N2 Shantou/1219/04 256 32 64 64
24-févr-05 8047 InfA H3N2 Shantou/1219/04 256 32 64 64
16-févr-05 7662 InfA H3N2 Shantou/1219/04 256 16 64 64
17-févr-05 7738 InfA H3N2 Shantou/1219/04 256 256 256
18-janv-05 6398 InfA H3N2 Shantou/1219/04 256 256 512
21-févr-05 7861 InfA H3N2 Shantou/1219/04 256 128 256
21-févr-05 7855 InfA H3N2 Shantou/1219/04 256 128 128
21-févr-05 7859 InfA H3N2 Shantou/1219/04 256 128 128
18-févr-05 7799 InfA H3N2 Shantou/1219/04 256 128 128
17-févr-05 7753 InfA H3N2 Shantou/1219/04 256 128 128
16-févr-05 7660 InfA H3N2 Shantou/1219/04 256 128 256
16-févr-05 7664 InfA H3N2 Shantou/1219/04 256 128 128
10-févr-05 7461 InfA H3N2 Shantou/1219/04 256 128 256
03-févr-05 7043 InfA H3N2 Shantou/1219/04 256 128 256
03-févr-05 7045 InfA H3N2 Shantou/1219/04 256 128 256
02-févr-05 6987 InfA H3N2 Shantou/1219/04 256 128 256
01-févr-05 6944 InfA H3N2 Shantou/1219/04 256 128 256
20-janv-05 6520 InfA H3N2 Shantou/1219/04 256 128 128
20-janv-05 6521 InfA H3N2 Shantou/1219/04 256 128 128
17-janv-05 6350 InfA H3N2 Shantou/1219/04 256 128 256
07-janv-05 6002 InfA H3N2 Shantou/1219/04 256 128 256
06-janv-05 5979 InfA H3N2 Shantou/1219/04 256 128 128
26-nov-04 4689 InfA H3N2 Shantou/1219/04 256 128 128
15-févr-05 7608 InfA H3N2 Shantou/1219/04 256 64 128
15-févr-05 7623 InfA H3N2 Shantou/1219/04 256 64 128
09-févr-05 7379 InfA H3N2 Shantou/1219/04 256 64 128
27-janv-05 6804 InfA H3N2 Shantou/1219/04 256 64 64
20-janv-05 6512 InfA H3N2 Shantou/1219/04 256 64 128
19-janv-05 6441 InfA H3N2 Shantou/1219/04 256 64 128
08-mars-05 8498 InfA H3N2 Shantou/1219/04 128 128 128 128
09-mars-05 8558 InfA H3N2 Shantou/1219/04 128 64 64 128
23-févr-05 7977 InfA H3N2 Shantou/1219/04 128 64 32 64
22-févr-05 7936 InfA H3N2 Shantou/1219/04 128 64 32 64

47/54
Date Seq N° Typisation A/Shan A/Cal A/Well A/Wyo
22-févr-05 7931 InfA H3N2 Shantou/1219/04 128 32 128 64
17-mars-05 8909 InfA H3N2 Shantou/1219/04 128 32 32 32
25-févr-05 8092 InfA H3N2 Shantou/1219/04 128 128 256
23-févr-05 7959 InfA H3N2 Shantou/1219/04 128 128 256
23-févr-05 7966 InfA H3N2 Shantou/1219/04 128 128 128
18-févr-05 7795 InfA H3N2 Shantou/1219/04 128 128 256
10-févr-05 7465 InfA H3N2 Shantou/1219/04 128 128 128
25-janv-05 6678 InfA H3N2 Shantou/1219/04 128 128 128
14-janv-05 6300 InfA H3N2 Shantou/1219/04 128 128 256
07-janv-05 5997 InfA H3N2 Shantou/1219/04 128 128 128
07-janv-05 6000 InfA H3N2 Shantou/1219/04 128 128 128
25-févr-05 8094 InfA H3N2 Shantou/1219/04 128 64 64
24-févr-05 8038 InfA H3N2 Shantou/1219/04 128 64 128
22-févr-05 7927 InfA H3N2 Shantou/1219/04 128 64 64
22-févr-05 7933 InfA H3N2 Shantou/1219/04 128 64 64
21-févr-05 7856 InfA H3N2 Shantou/1219/04 128 64 64
18-févr-05 7798 InfA H3N2 Shantou/1219/04 128 64 64
18-févr-05 7803 InfA H3N2 Shantou/1219/04 128 64 64
16-févr-05 7661 InfA H3N2 Shantou/1219/04 128 64 256
15-févr-05 7629 InfA H3N2 Shantou/1219/04 128 64 128
15-févr-05 7633 InfA H3N2 Shantou/1219/04 128 64 128
15-févr-05 7635 InfA H3N2 Shantou/1219/04 128 64 128
15-févr-05 7614 InfA H3N2 Shantou/1219/04 128 64 64
09-févr-05 7405 InfA H3N2 Shantou/1219/04 128 64 128
09-févr-05 7409 InfA H3N2 Shantou/1219/04 128 64 128
09-févr-05 7377 InfA H3N2 Shantou/1219/04 128 64 64
09-févr-05 7389 InfA H3N2 Shantou/1219/04 128 64 64
08-févr-05 7356 InfA H3N2 Shantou/1219/04 128 64 128
08-févr-05 7319 InfA H3N2 Shantou/1219/04 128 64 64
08-févr-05 7331 InfA H3N2 Shantou/1219/04 128 64 64
07-févr-05 7271 InfA H3N2 Shantou/1219/04 128 64 128
02-févr-05 6978 InfA H3N2 Shantou/1219/04 128 64 128
01-févr-05 6941 InfA H3N2 Shantou/1219/04 128 64 128
01-févr-05 6945 InfA H3N2 Shantou/1219/04 128 64 128
01-févr-05 6936 InfA H3N2 Shantou/1219/04 128 64 64
27-janv-05 6802 InfA H3N2 Shantou/1219/04 128 64 64
26-janv-05 6715 InfA H3N2 Shantou/1219/04 128 64 128
25-janv-05 6689 InfA H3N2 Shantou/1219/04 128 64 128
25-janv-05 6677 InfA H3N2 Shantou/1219/04 128 64 64
24-janv-05 6621 InfA H3N2 Shantou/1219/04 128 64 256
24-janv-05 6618 InfA H3N2 Shantou/1219/04 128 64 128
21-janv-05 6550 InfA H3N2 Shantou/1219/04 128 64 128
20-janv-05 6516 InfA H3N2 Shantou/1219/04 128 64 64
13-janv-05 6265 InfA H3N2 Shantou/1219/04 128 64 128
10-janv-05 6065 InfA H3N2 Shantou/1219/04 128 64 128
07-janv-05 6003 InfA H3N2 Shantou/1219/04 128 64 64
30-nov-04 4817 InfA H3N2 Shantou/1219/04 128 64 64
25-févr-05 8098 InfA H3N2 Shantou/1219/04 128 32 128

48/54
Date Seq N° Typisation A/Shan A/Cal A/Well A/Wyo
22-févr-05 7924 InfA H3N2 Shantou/1219/04 128 32 64
22-févr-05 7925 InfA H3N2 Shantou/1219/04 128 32 64
22-févr-05 7938 InfA H3N2 Shantou/1219/04 128 32 64
18-févr-05 7797 InfA H3N2 Shantou/1219/04 128 32 64
15-févr-05 7631 InfA H3N2 Shantou/1219/04 128 32 64
08-févr-05 7322 InfA H3N2 Shantou/1219/04 128 32 128
08-févr-05 7353 InfA H3N2 Shantou/1219/04 128 32 64
08-févr-05 7359 InfA H3N2 Shantou/1219/04 128 32 64
07-févr-05 7273 InfA H3N2 Shantou/1219/04 128 32 64
04-févr-05 7084 InfA H3N2 Shantou/1219/04 128 32 32
03-févr-05 7048 InfA H3N2 Shantou/1219/04 128 32 64
02-févr-05 6984 InfA H3N2 Shantou/1219/04 128 32 64
19-janv-05 6439 InfA H3N2 Shantou/1219/04 128 32 32
23-déc-04 5676 InfA H3N2 Shantou/1219/04 128 32 64
04-janv-05 5868 InfA H3N2 Shantou/1219/04 128 16 64
14-mars-05 8720 InfA H3N2 Shantou/1219/04 64 32 16 32
01-mars-05 8231 InfA H3N2 Shantou/1219/04 64 16 32 32
11-févr-05 7487 InfA H3N2 Shantou/1219/04 64 128 64
25-févr-05 8096 InfA H3N2 Shantou/1219/04 64 64 64
23-févr-05 7969 InfA H3N2 Shantou/1219/04 64 64 128
23-févr-05 7965 InfA H3N2 Shantou/1219/04 64 64 64
08-févr-05 7339 InfA H3N2 Shantou/1219/04 64 64 64
07-févr-05 7269 InfA H3N2 Shantou/1219/04 64 64 128
03-févr-05 7039 InfA H3N2 Shantou/1219/04 64 64 64
02-févr-05 6972 InfA H3N2 Shantou/1219/04 64 64 64
02-févr-05 6977 InfA H3N2 Shantou/1219/04 64 64 64
02-févr-05 6986 InfA H3N2 Shantou/1219/04 64 64 64
27-janv-05 6807 InfA H3N2 Shantou/1219/04 64 64 64
25-janv-05 6676 InfA H3N2 Shantou/1219/04 64 64 64
24-janv-05 6622 InfA H3N2 Shantou/1219/04 64 64 128
10-janv-05 6067 InfA H3N2 Shantou/1219/04 64 64 128
29-déc-04 5732 InfA H3N2 Shantou/1219/04 64 64 128
22-déc-04 5625 InfA H3N2 Shantou/1219/04 64 64 128
04-mars-05 8378 InfA H3N2 Shantou/1219/04 64 32 64
23-févr-05 7970 InfA H3N2 Shantou/1219/04 64 32 64
17-févr-05 7736 InfA H3N2 Shantou/1219/04 64 32 64
17-févr-05 7748 InfA H3N2 Shantou/1219/04 64 32 64
17-févr-05 7755 InfA H3N2 Shantou/1219/04 64 32 64
17-févr-05 7737 InfA H3N2 Shantou/1219/04 64 32 64
17-févr-05 7739 InfA H3N2 Shantou/1219/04 64 32 32
15-févr-05 7621 InfA H3N2 Shantou/1219/04 64 32 64
15-févr-05 7611 InfA H3N2 Shantou/1219/04 64 32 64
15-févr-05 7636 InfA H3N2 Shantou/1219/04 64 32 64
11-févr-05 7484 InfA H3N2 Shantou/1219/04 64 32 64
09-févr-05 7383 InfA H3N2 Shantou/1219/04 64 32 64
09-févr-05 7407 InfA H3N2 Shantou/1219/04 64 32 64
08-févr-05 7340 InfA H3N2 Shantou/1219/04 64 32 64
08-févr-05 7327 InfA H3N2 Shantou/1219/04 64 32 32

49/54
Date Seq N° Typisation A/Shan A/Cal A/Well A/Wyo
08-févr-05 7330 InfA H3N2 Shantou/1219/04 64 32 32
04-févr-05 7073 InfA H3N2 Shantou/1219/04 64 32 64
03-févr-05 7040 InfA H3N2 Shantou/1219/04 64 32 64
02-févr-05 6970 InfA H3N2 Shantou/1219/04 64 32 64
02-févr-05 6983 InfA H3N2 Shantou/1219/04 64 32 64
02-févr-05 6976 InfA H3N2 Shantou/1219/04 64 32 32
01-févr-05 6938 InfA H3N2 Shantou/1219/04 64 32 64
31-janv-05 6885 InfA H3N2 Shantou/1219/04 64 32 64
31-janv-05 6892 InfA H3N2 Shantou/1219/04 64 32 64
27-janv-05 6805 InfA H3N2 Shantou/1219/04 64 32 64
26-janv-05 6725 InfA H3N2 Shantou/1219/04 64 32 64
25-janv-05 6684 InfA H3N2 Shantou/1219/04 64 32 64
25-janv-05 6691 InfA H3N2 Shantou/1219/04 64 32 64
25-janv-05 6675 InfA H3N2 Shantou/1219/04 64 32 32
24-janv-05 6629 InfA H3N2 Shantou/1219/04 64 32 32
19-janv-05 6443 InfA H3N2 Shantou/1219/04 64 32 32
18-janv-05 6402 InfA H3N2 Shantou/1219/04 64 32 64
13-janv-05 6264 InfA H3N2 Shantou/1219/04 64 32 64
11-janv-05 6136 InfA H3N2 Shantou/1219/04 64 32 64
11-janv-05 6128 InfA H3N2 Shantou/1219/04 64 32 32
11-févr-05 7489 InfA H3N2 Shantou/1219/04 64 16 32
08-févr-05 7341 InfA H3N2 Shantou/1219/04 64 16 32
12-janv-05 6178 InfA H3N2 Shantou/1219/04 32 32 64
02-févr-05 6988 Inf A H3N2 Wyoming/3/03 64 128 256
03-janv-05 5779 Inf A H3N2 Wyoming/3/03 32 64 128
15-févr-05 7619 Inf A H3N2 Wyoming/3/03 32 64 128
04-févr-05 7091 Inf A H3N2 Wyoming/3/03 32 32 128
22-févr-05 7932 Influenza A H3N2 : 16 32 32 32
23-févr-05 7978 Influenza A H3N2 : 16 32 32 32
08-févr-05 7342 Influenza A H3N2 : 16 32 32 16
28-févr-05 8166 Influenza A H3N2 : 16 32 16 16
24-janv-05 6624 Influenza A H3N2 : 32 32 64
27-janv-05 6808 Influenza A H3N2 : 32 32 64
27-janv-05 6813 Influenza A H3N2 : 32 32 64
01-févr-05 6933 Influenza A H3N2 : 32 32 64
02-févr-05 6975 Influenza A H3N2 : 32 32 64
08-févr-05 7351 Influenza A H3N2 : 32 32 64
08-févr-05 7326 Influenza A H3N2 : 32 32 64
09-févr-05 7376 Influenza A H3N2 : 32 32 64
15-févr-05 7625 Influenza A H3N2 : 32 32 64
17-févr-05 7746 Influenza A H3N2 : 32 32 64
01-févr-05 6937 Influenza A H3N2 : 32 32 32
02-févr-05 6979 Influenza A H3N2 : 32 32 32
08-févr-05 7337 Influenza A H3N2 : 32 32 32
09-févr-05 7394 Influenza A H3N2 : 32 32 32
19-janv-05 6440 Influenza A H3N2 : 16 32 32
04-févr-05 7095 Influenza A H3N2 : 16 32 32

50/54
Date Seq N° Typisation A/Shan A/Cal A/Well A/Wyo
21-janv-05 6573 Influenza A H3N2 : 8 32 64 Samples sent to MRC London for further analysis

51/54
Annexe 4 : Antigenic analysis of influenza A (H1N1)-like strains
Antisera
A (H1N1) strains A/New Cal. A/Beij. A/Madaga.
A/New Caledonia/20/99 2048 64 1024 A/Beijing/262/95 512 1024 256 A/Madagascar/57794 512 32 256
IHA titre Date Seq. N° Typisation
A/NC A/Beij A/Madag 11.03.2005 8719 A/Madagascar/57794/00 1024 256 256 26.03.2005 9163 A/Madagascar/57794/00 512 32 256 30.03.2005 9286 A/Madagascar/57794/00 512 64 256 06.04.2005 9497 A/Madagascar/57794/00 512 32 256 08.03.2005 8493 A/Madagascar/57794/00 512 64 256 28.02.2005 8703 A/Madagascar/57794/00 512 64 128 15.03.2005 8816 A/Madagascar/57794/00 512 32 128 22.03.2005 9066 A/Madagascar/57794/00 512 32 128 24.01.2005 6714 A/Madagascar/57794/00 256 128 512 27.12.2004 5741 A/Madagascar/57794/00 256 64 256 14.01.2005 6348 A/Madagascar/57794/00 256 64 256 27.01.2005 6889 A/Madagascar/57794/00 256 32 256 27.01.2005 7357 A/Madagascar/57794/00 256 32 256 22.12.2004 5673 A/Madagascar/57794/00 256 32 128 10.01.2005 6260 A/Madagascar/57794/00 256 32 128 04.02.2005 7332 A/Madagascar/57794/00 256 32 128 10.02.2005 7554 A/Madagascar/57794/00 256 32 128 28.02.2005 8230 A/Madagascar/57794/00 256 64 128 03.03.2005 8384 A/Madagascar/57794/00 256 16 128 25.02.2005 8219 A/Madagascar/57794/00 256 32 64 14.02.2005 7628 A/Madagascar/57794/00 256 15.02.2005 7754 A/Madagascar/57794/00 256 32 21.02.2005 7937 A/Madagascar/57794/00 256 32 14.12.2004 5365 A/Madagascar/57794/00 128 32 256 24.01.2005 6720 A/Madagascar/57794/00 128 64 256 15.02.2005 7675 A/Madagascar/57794/00 128 32 256 27.12.2004 5742 A/Madagascar/57794/00 128 32 128 10.01.2005 6188 A/Madagascar/57794/00 128 32 128 02.02.2005 7072 A/Madagascar/57794/00 128 32 128 27.01.2005 7352 A/Madagascar/57794/00 128 64 128 18.01.2005 6442 A/Madagascar/57794/00 128 16 64 24.01.2005 6734 A/Madagascar/57794/00 128 32 64 14.02.2005 7632 A/Madagascar/57794/00 128 16.02.2005 7757 A/Madagascar/57794/00 128 64 07.03.2005 8619 A/Madagascar/57794/00

52/54
Annexe 5 : Antigenic analysis of influenza B-like strains
Antisera
B strains B/Jiang. B/Harb. B/Sich. B/HK. B/Beij. B/Shan.
B/Jiangsu/10/2003 1024 64 128 16 32 < 16
B/Harbin/7/94 < 16 512 128 256 64 32 B/Sichuan/379/99 < 16 128 256 64 32 < 16 B/Hong Kong/335/2001 < 16 64 < 16 256 < 16 1024 B/Beijing/184/93 < 16 512 128 128 256 16 B/Shandong/7/97 < 16 < 16 < 16 32 < 16 1024
Antisera Date Seq N° Typisation B/Jiang B/Har B/Sich B/HK B/Bris B/Beij B/Shan
5-janv-05 5972 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 1024
2-févr-05 7046 InfB Shandong/7/97 <16 <16 <16 <16 64 <16 512
6-janv-05 6001 InfB Shandong/7/97 <16 <16 <16 <16 64 <16 512
28-févr-05 8226 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
13-févr-05 7637 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
29-janv-05 6886 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
24-janv-05 6690 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
23-déc-04 5710 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
23-déc-04 5709 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
21-déc-04 5675 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
20-déc-04 5580 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 512
30-mars-05 9287 InfB Shandong/7/97 <16 <16 <16 <16 16 <16 512
25-mars-05 9277 InfB Shandong/7/97 <16 <16 <16 <16 16 <16 512
18-mars-05 9039 InfB Shandong/7/97 <16 <16 <16 <16 16 <16 512
18-mars-05 8983 InfB Shandong/7/97 <16 <16 <16 <16 16 <16 512
27-déc-04 5733 InfB Shandong/7/97 <16 <16 <16 <16 32 <16 256
10-déc-04 5303 InfB Shandong/7/97 <16 <16 <16 8 32 <16 256
8-mars-05 8560 InfB Shandong/7/97 <16 <16 <16 <16 16 <16 256
22-mars-05 9164 InfB Jiangsu/10/02 256 128 64 32 <16 64 <16
24-févr-05 8161 InfB Jiangsu/10/03 256 64 64 32 <16 16 <16
21-févr-05 7974 InfB Jiangsu/10/03 256 64 64 32 <16 16 <16
2-févr-05 7093 InfB Jiangsu/10/03 256 64 64 32 <16 16 <16
1-févr-05 6981 InfB Jiangsu/10/03 256 64 64 64 <16 16 <16
27-déc-04 5738 InfB Jiangsu/10/03 256 64 64 32 <16 32 <16
8-févr-05 7412 InfB Jiangsu/10/03 256 32 32 32 <16 16 <16
10-mars-05 8716 B/Sichuan/379/99 128 128 64 64 <16 32 <16
21-mars-05 9275 B/Sichuan/379/99 128 64 64 <16 <16 <16 <16
18-mars-05 9274 B/Sichuan/379/99 128 64 64 <16 <16 <16 <16
22-mars-05 9070 B/Sichuan/379/99 128 64 64 32 <16 32 <16
21-mars-05 9029 B/Sichuan/379/99 128 64 64 32 <16 16 <16

53/54
Antisera Date Seq N° Typisation B/Jiang B/Har B/Sich B/HK B/Bris B/Beij B/Shan
16-mars-05 8907 B/Sichuan/379/99 128 64 64 32 <16 32 <16
16-mars-05 8906 B/Sichuan/379/99 128 64 64 32 <16 32 <16
16-mars-05 8903 B/Sichuan/379/99 128 64 64 32 <16 16 <16
31-mars-05 9365 B/Sichuan/379/99 128 64 64 32 <16 16 <16
9-mars-05 8623 B/Sichuan/379/99 128 16 64 32 <16 16 <16
9-mars-05 8624 B/Harbin/7/94 128 64 32 32 <16 16 <16
11-mars-05 9273 B/Sichuan/379/109 64 256 256 256 <16 32 64
20-déc-04 5579 Influenza B 128 32 32 8 <16 8 <16
10-mars-05 8710 Influenza B 64 32 32 16 <16 8 <16
10-févr-05 7490 Influenza B 64 32 32 32 <16 16 <16
7-mars-05 8619 Co-infection A/H1 and B

54/54
Annexe 6 : Alignement de l’hemagglutinine de souches récentes influenza A (H5N1) Les séquences de souches influenza A (H5N1) d’origine aviaire détectés chez l’homme ont été
alignées, dont 2 souches provenant de cas humains du Vietnam détectés au cours de l’année 2005 :
séquence 2 (Vietnam Sud) et séquence 3 (Vietnam Nord). Les primers et les sondes centrales
utilisées pour la détection des virus influenza A (H5N1) par PCR Sont mentionnés. En bleu sont
représentés les primers de PCR classique, en vert et rouge les primers de PCR en temps réel. La
région mentionnée en orange montre une délétion dans la séquence du virus du Nord du Vietnam).
Les 2 séquences de virus humains influenza A (H5N1) du Vietnam ont été aimablement fournies par
le laboratoire National de l’Influenza, du Vietnam du NIHE (National Institute of Hygiene and
Epidemiology) de Hanoi.
Who H5-1 HA5For HA5 Pro 910 1 2 3 4 5 6 7 8 9
10
12 13 14 15 16 17 18
11
H5Viet For H5VietPro HARev 1030 1 2 3 4 5 6 7 8 9
10
12 13 14 15 16 17 18
11
H5VietRev 1150 1 2 3 4 5 6 7 8 9
10
12 13 14 15 16 17 18
11
Who H5-2 1275 1 2 3 4 5 6 7 8 9
10
12 13 14 15 16 17 18
11




















