
Fc-mOX40L Fusion Protein Produces Complete Remission and Enhanced Survival in 2 Murine Tumor Models
11
Fc-mOX40L Fusion Protein Produces Complete Remission and Enhanced Survival in 2 Murine Tumor Models Rebecca E. Sadun,* Wen-En Hsu,* Nan Zhang,* Yu-Chih Nien,w Scott A. Bergfeld,* Helen Sabzevari,z M. E. Christine Lutsiak,z Leslie Khawli,y Peisheng Hu,* and Alan L. Epstein* Summary: OX40L is a member of the tumor necrosis factor superfamily that provides a costimulatory signal to CD4 + and CD8 + T cells while inhibiting the effects of suppressive CD4 + CD25 + regulatory T cells. Because of this dual activity, OX40L may provide significant antitumor immunity in tumor- bearing mice. To study its clinical potential, a fusion protein consisting of mOX40L linked to the C-terminus of the Fc fragment of immunoglobulin was genetically engineered. After demonstrating its potency in vitro, several assays were performed to evaluate its antitumor effect in comparison to the OX40 agonist antibody OX86. Dosing studies in Colon 26-bearing and renal cell carcinoma (RENCA)-bearing mice showed that although OX86 produced modest tumor regression, Fc-mOX40L produced complete remission in both tumor models. Survival studies confirmed these results and showed that Fc-mOX40L treatment produced lasting responses throughout the 5-month observation period. Flow cytometric analysis of treated and untreated tumors and tumor-draining lymph nodes identified a qualitative difference in the activity of Fc-mOX40L compared with OX86 treatment as evidenced by differences in lymphoid and macrophage populations. These studies reflect the profound therapeutic potential of Fc-mOX40L, which substantially exceeds the agonist antibody OX86 in ability to produce complete tumor remissions and promote long-term survival in solid tumor models. Key Words: Fc-mOX40L, OX40L, CD134L, antibody immu- notherapy, TNFSF, fusion proteins (J Immunother 2008;31:235–245) O X40 (TNFRSF4, CD134) is a membrane-associated glycoprotein member of the tumor necrosis factor receptor superfamily (TNFRSF). This receptor is tran- siently expressed on the surface of T cells after T-cell receptor ligation. 1–3 Its natural ligand OX40L (TNFSF4, CD134L) belongs to the tumor necrosis factor ligand superfamily (TNFSF) 4,5 and is found primarily on antigen presenting cells, including activated B cells, dendritic cells (DCs), and macrophages. In addition, OX40L is expressed on activated endothelial cells 6–11 and has been reported to be up-regulated on CD4 + T cells during antigen simulation. 6,12 Early studies of primary T-cell responses revealed OX40L to be a potent costimulatory molecule for sustaining the CD4 + T-cell response. 13–15 In vivo, OX40 signaling augments tumor-specific priming by stimulating and expanding the natural repertoire of the host’s tumor-specific CD4 + T cells. 8,10,16,17 Additionally, OX40 signaling is believed to increase CD4 + T-cell differentiation from effector to memory cells. 18 In addition to OX40L’s effects on CD4 + T cells, recent studies have shown that OX40 signaling also directly costimulates CD8 + T cells. 19,20 OX40L stimula- tion results in the clonal expansion and increased effector function of CD8 + T cells, with additive and synergistic effects when delivered in conjunction with B7.1 or 4-1BBL. 21 Anti-OX40L monoclonal antibody (mAb) suppressed interferon (IFN)-g expression and proliferation of CD8 + T cells in coculture with DCs, suggesting that DC OX40L is largely responsible for CD8 + expression of IFN-g. 22 In addition, in vivo administration of the agonist anti-OX40 antibody, OX86, was found to increase survival and potency of antigen-specific CD8 + memory T cells. 23 Most recently it was discovered that IL-18, which increases antigen-specific CD4 + and CD8 + T cells, acts through the OX40 pathway, by increasing DC expression of OX40L and by increasing antigen- specific T-cell expression of OX40 when delivered alone or in conjunction with peptide. 24 Importantly, in addition to expression on CD4 + and CD8 + T cells, OX40 has also been found to be expressed on both naive and activated CD4 + CD25 + regulatory T cells (Tregs). 25,26 OX40 signaling may provide an inhibitory signal that abrogates Treg-mediated suppression when delivered to antigen-engaged naive T cells. 27 The agonist antibody OX86 was found to inhibit the suppressive effects of Tregs while restoring effector T-cell proliferation and cytokine production. 28 Using a graft-versus-host disease model, it was confirmed that OX40 is nearly as potent as glucocorticoid-induced tumor Copyright r 2008 by Lippincott Williams & Wilkins From the Departments of *Pathology; wMicrobiology, Keck School of Medicine, University of Southern California, Los Angeles; yGenentech Inc, One DNA Way, South San Francisco, CA; and zLaboratory of Tumor Immunology and Biology, National Cancer Institute, National Institutes of Health, Bethesda, MD. Supported in part by grants from Cancer Therapeutics Laboratories Inc (Los Angeles, CA) and the Philip Morris External Research Program (Linthicum Heights, MD). Financial Disclosure: The authors have declared there are no financial conflicts of interest in regard to this work. Reprints: Prof Alan L. Epstein, Department of Pathology, Keck School of Medicine, University of Southern California, 2011 Zonal Avenue, HMR 205, Los Angeles, CA 90033 (e-mail: [email protected]). BASIC STUDY J Immunother Volume 31, Number 3, April 2008 235
Transcript of Fc-mOX40L Fusion Protein Produces Complete Remission and Enhanced Survival in 2 Murine Tumor Models

Fc-mOX40L Fusion Protein Produces Complete Remissionand Enhanced Survival in 2 Murine Tumor Models
Rebecca E. Sadun,* Wen-En Hsu,* Nan Zhang,* Yu-Chih Nien,w Scott A. Bergfeld,*Helen Sabzevari,zM. E. Christine Lutsiak,z Leslie Khawli,y Peisheng Hu,* and Alan L. Epstein*
Summary: OX40L is a member of the tumor necrosis factor
superfamily that provides a costimulatory signal to CD4+ and
CD8+ T cells while inhibiting the effects of suppressive
CD4+CD25+ regulatory T cells. Because of this dual activity,
OX40L may provide significant antitumor immunity in tumor-
bearing mice. To study its clinical potential, a fusion protein
consisting of mOX40L linked to the C-terminus of the Fc
fragment of immunoglobulin was genetically engineered. After
demonstrating its potency in vitro, several assays were performed
to evaluate its antitumor effect in comparison to the OX40
agonist antibody OX86. Dosing studies in Colon 26-bearing and
renal cell carcinoma (RENCA)-bearing mice showed that
although OX86 produced modest tumor regression, Fc-mOX40L
produced complete remission in both tumor models. Survival
studies confirmed these results and showed that Fc-mOX40L
treatment produced lasting responses throughout the 5-month
observation period. Flow cytometric analysis of treated and
untreated tumors and tumor-draining lymph nodes identified a
qualitative difference in the activity of Fc-mOX40L compared
with OX86 treatment as evidenced by differences in lymphoid and
macrophage populations. These studies reflect the profound
therapeutic potential of Fc-mOX40L, which substantially exceeds
the agonist antibody OX86 in ability to produce complete tumor
remissions and promote long-term survival in solid tumor models.
Key Words: Fc-mOX40L, OX40L, CD134L, antibody immu-
notherapy, TNFSF, fusion proteins
(J Immunother 2008;31:235–245)
OX40 (TNFRSF4, CD134) is a membrane-associatedglycoprotein member of the tumor necrosis factor
receptor superfamily (TNFRSF). This receptor is tran-
siently expressed on the surface of T cells after T-cellreceptor ligation.1–3 Its natural ligand OX40L (TNFSF4,CD134L) belongs to the tumor necrosis factor ligandsuperfamily (TNFSF)4,5 and is found primarily onantigen presenting cells, including activated B cells,dendritic cells (DCs), and macrophages. In addition,OX40L is expressed on activated endothelial cells6–11 andhas been reported to be up-regulated on CD4+ T cellsduring antigen simulation.6,12
Early studies of primary T-cell responses revealedOX40L to be a potent costimulatory molecule forsustaining the CD4+ T-cell response.13–15 In vivo,OX40 signaling augments tumor-specific priming bystimulating and expanding the natural repertoire of thehost’s tumor-specific CD4+ T cells.8,10,16,17 Additionally,OX40 signaling is believed to increase CD4+ T-celldifferentiation from effector to memory cells.18
In addition to OX40L’s effects on CD4+ T cells,recent studies have shown that OX40 signaling alsodirectly costimulates CD8+ T cells.19,20 OX40L stimula-tion results in the clonal expansion and increased effectorfunction of CD8+ T cells, with additive and synergisticeffects when delivered in conjunction with B7.1 or4-1BBL.21 Anti-OX40L monoclonal antibody (mAb)suppressed interferon (IFN)-g expression and proliferationof CD8+ T cells in coculture with DCs, suggesting thatDC OX40L is largely responsible for CD8+ expression ofIFN-g.22 In addition, in vivo administration of theagonist anti-OX40 antibody, OX86, was found toincrease survival and potency of antigen-specific CD8+
memory T cells.23 Most recently it was discovered thatIL-18, which increases antigen-specific CD4+ and CD8+
T cells, acts through the OX40 pathway, by increasingDC expression of OX40L and by increasing antigen-specific T-cell expression of OX40 when delivered aloneor in conjunction with peptide.24
Importantly, in addition to expression on CD4+
and CD8+ T cells, OX40 has also been found to beexpressed on both naive and activated CD4+CD25+
regulatory T cells (Tregs).25,26 OX40 signaling mayprovide an inhibitory signal that abrogates Treg-mediatedsuppression when delivered to antigen-engaged naiveT cells.27 The agonist antibody OX86 was found to inhibitthe suppressive effects of Tregs while restoring effectorT-cell proliferation and cytokine production.28 Using agraft-versus-host disease model, it was confirmed thatOX40 is nearly as potent as glucocorticoid-induced tumorCopyright r 2008 by Lippincott Williams & Wilkins
From the Departments of *Pathology; wMicrobiology, Keck Schoolof Medicine, University of Southern California, Los Angeles;yGenentech Inc, One DNA Way, South San Francisco, CA; andzLaboratory of Tumor Immunology and Biology, National CancerInstitute, National Institutes of Health, Bethesda, MD.
Supported in part by grants from Cancer Therapeutics Laboratories Inc(Los Angeles, CA) and the Philip Morris External Research Program(Linthicum Heights, MD).
Financial Disclosure: The authors have declared there are no financialconflicts of interest in regard to this work.
Reprints: Prof Alan L. Epstein, Department of Pathology, Keck Schoolof Medicine, University of Southern California, 2011 Zonal Avenue,HMR 205, Los Angeles, CA 90033 (e-mail: [email protected]).
BASIC STUDY
J Immunother � Volume 31, Number 3, April 2008 235

necrosis factor receptor ligand (GITRL) at abrogatingTregs in mice.28
Because of the ability of the OX40-OX40L pathwayto stimulate effector and memory CD4+ and CD8+ Tcells and to suppress CD4+CD25+ Tregs, it waspostulated that OX40 signaling might provide significantantitumor activity. Indeed, it was discovered thatengagement of OX40 during tumor priming enhancesa tumor-specific T-cell response that can increase thepercentage of mice that become tumor-free.29 Tumor-specific CD8+ T-cell responses were found to besustained for several weeks and have been sufficient totreat established tumors, even under immunocompromis-ing conditions.30 Further studies have confirmed thatOX86 has a synergistic effect in combination withgranulocyte macrophage colony-stimulating factor31 orthe 4-1BB32 agonist antibody 2A in the treatment ofColon 26 tumors. In addition, 2A antibody treatmentsenhance the 1-year survival of mice bearing Colon 26hepatic metastases.32 OX40L has also been shown to beeffective in tumor models as disparate as the Lewis lungcarcinoma33 and lymphomas.34
To-date, most in vivo studies of OX40’s antitumoreffects have been performed using the OX86 agonistantibody. It has been postulated, however, that thenatural tumor necrosis factor (TNF) ligand may producefewer side effects,35 advocating in favor of the use ofOX40L for the treatment of cancer. Several laboratorieshave endeavored to create OX40L-containing moleculeswith adequate bioavailability,36 but few investigationshave been published on their in vivo activity. Thesestudies have been complicated by the fact that the OX40Lis composed of a homo-trimer which is required forbinding to the OX40 receptor.5 Given the evidence thatconjugation of TNF ligands to the Fc portion ofantibodies can extend half-life while providing addedeffector functions including antibody-dependent cellcytotoxicity,37 we produced an Fc-mOX40L fusionprotein consisting of 2 homo-trimers per complex. Weherein provide evidence of both in vitro and in vivoactivity of this molecule for the treatment of murinetumors, with results suggesting that Fc-mOX40L hassignificantly greater antitumor activity than the well-studied agonist antibody OX86.
MATERIALS AND METHODS
Antibodies and Cell LinesAnti-CD3 (145-11C clone), phycoerythrin (PE)-anti
CD4 (RM4-5 clone), PE-anti-CD8a (53-6.7 clone), PE-anti-CD25 (PC61 clone), PE-anti-CD11c (HL3 clone),PE-anti-CD49b (DX5 clone) mAb, and HRP-streptavidinwere purchased from BD Pharmingen (San Diego, CA).Hybridomas, including rat antimouse CD4 (GK1.5), anti-CD8b (H35), and anti-CD25 (PC61) mAbs were pur-chased from American Type Culture Collection (ATCC,Manassas, VA). Anti-OX40 (OX86) mAb-producingcell line was obtained from the European Cell CultureCollection (Wiltshire, UK). For immunohistochemical
staining, both primary rabbit antimouse granzyme B andsecondary biotinylated goat antirabbit IgG polyclonalantibodies were purchased from Abcam Inc (Cambridge,MA).
The NS0 murine myeloma cell line was obtainedform Lonza Biologics (Slough, UK). The Colon 26colorectal adenocarcinoma and RENCA renal cellcarcinoma cell lines were obtained from ATCC.
Reagents and MiceThe Glutamine Synthetase Gene Amplification
System with expression plasmids pEE6/hCMV-B andpEE12 was purchased from Lonza Biologics (Slough,UK). The murine OX40L cDNA was a generous gift fromDr Sugamura from the Department of Microbiology andImmunology, Tohoku University, Japan. Restrictionendonucleases, T4 DNA ligase, Vent polymerase, andother molecular biology reagents were obtained fromeither New England Biolabs (Beverly, MA) or BoehringerMannheim (Indianapolis, IN). Characterized and dia-lyzed fetal calf sera were purchased from Hyclone Corp(Logan, UT), and RPMI 1640 medium, HybridomaSelective Medium without L-glutamine, minimal essentialmedium nonessential amino acids solution (100� ), andphosphate-buffered saline (PBS) were purchased fromGIBCO LifeTechnologies (San Diego, CA). The murineIL-2 enzyme-linked immunosorbent assay (ELISA) kitwas purchased from BD Biosciences (San Jose, CA).
Six-week-old female BALB/c mice were obtainedfrom Harlan Sprague-Dawley (Indianapolis, IN). Allexperiments were performed in accordance with Institu-tional Animal Care and Use Committee protocols andinstitutional guidelines for the proper humane care anduse of animals in research.
Construction of Fc-mOX40LFc-mOX40L was cloned using previously described
methods.35 The Fc-mOX40L cDNA extracellular domainwas amplified using primers 50-AAG GAA AAA AGCGGC CGC CAA CTC TCT TCC TCT CCG GCA-30 and50-GGC GAA TTC TCA CAG TGG TAC TTG GTTCAC AGT-30. The polymerase chain reaction fragmentthen was inserted into the pEE12 vector via Not I andEcoR I sites in the C-terminus of the Fc sequence. Thevector with the Fc-mOX40L fusion gene was transfectedby electroporation into NS0 cells.
Expression and Purification of Fc-mOX40LThe Fc-mOX40L was expressed in NS0 murine
myeloma cells for long-term stable expression as per themanufacturer’s protocol (Lonza Biologics). The highestproducing clone, determined by indirect ELISA screeningfor murine Fc in a 24-hour assay, was scaled-up forincubation in an aerated 3 or 8-L stir flask bioreactorusing 2.5% dialyzed fetal calf serum. The secreted fusionprotein was then purified from the clarified cell culturesupernatant by tandem protein-A affinity and ion-exchange chromatography, as described previously.38
Sadun et al J Immunother � Volume 31, Number 3, April 2008
236 r 2008 Lippincott Williams & Wilkins

The fusion protein was analyzed by ELISA to verify thepresence and proper folding of the OX40L extracellulardomain and was further analyzed by high-pressure liquidchromatography (HPLC) and sodium dodecyl sulfate-polyacrylamide gel electrophoresis to verify properassembly and purity.
In Vitro Activity AssayThe bioactivity of the OX40L moiety was deter-
mined by ELISA measurement of IL-2 production fromsplenocytes aseptically removed from 6-week-old femaleBALB/c mice. After the red blood cells were lysed usingthe BD Pharm Lyse Lysing buffer (BD Pharmingen),single cell suspensions were washed twice in PBS andincubated in a 24-well plate (1.5� 106 cells/well) pre-coated with 5 mg/mL anti-CD3 (145-11C clone) in thepresence of 2 mg/mL Fc-mOX40L or OX86. After 48hours, interleukin (IL)-2 production was determined bysandwich ELISA (BD Biosciences) for the above culturesupernatants according to the manufacture’s protocol. Allassays were performed in triplicate.
Murine Treg and CD8+ T-cell ProliferationAssays
To demonstrate the effect of the Fc-mOX40L fusionprotein on T-cell subsets, a panel of Fc fusion proteinsproduced in our laboratory was tested in mouse Treg andCD8+ cell proliferation assays. For these assays, spleno-cytes from normal C57BL/6 mice were subjected to cellpurification using either a Miltenyi Biotec CD4+CD25+
Regulatory T Cell Isolation Kit (Treg) or CD8a microbe-ads (CD8+ cells). All wells contained irradiated antigen-presenting cells at a concentration of 1� 105/well andanti-CD3 (1 mg/mL). The Fc fusion proteins were testedin triplicate using 2 concentrations (1 and 10 mg/mL) inwhich CD8+ cells alone (5� 104/well), Treg cells alone(5� 104 cells/well), and a 1:1 combination of both cellpopulations (5� 104 of each/well) were tested. In thisway, background data of proliferation was obtained assome of the Fc fusion proteins induced proliferation ofreporter and Treg cells. 3H-thymidine was added for thelast 18 hours of culture. For analysis, the triplicate wellswere averaged and the percent suppression was calculatedby the equation:
CD8þ alone� ðCD8þ=Treg combination� Treg aloneÞ � 100
CD8þ alone:
Immunotherapeutic StudiesSix-week-old female BALB/c mice were injected
subcutaneously in the left flank with a 0.2-mL inoculumcontaining approximately 5� 106 Colon 26 or RENCAcells. When tumors reached 0.5 cm in diameter 5 to 7 daysafter tumor implantation, groups of mice (n=5) wereintravenously (IV) treated with a 0.1-mL inoculumcontaining incrementally increasing equimolar concentra-tions of Fc-mOX40L or OX86 (1 to 150 mg/dose). Allgroups of mice were treated daily � 5 and tumor growthwas monitored every other day by caliper measurement in
3 dimensions enabling tumor volumes to be calculated bythe formula length�width� height. The results wereexpressed as the mean±SD. Significance levels (P values)were determined using the Wilcoxon rank-sum test.
Evaluation of ToxicityToxicity was evaluated by monitoring the activity
levels of treated mice and by serum analysis for thepresence of autoimmune antibodies. For the later study,indirect immunofluorescence microscopy was performedon 2% paraformaldehyde (Polysciences, Warrington, PA)fixed HeLa cells grown on printed glass slides using serafrom treated mice 3 weeks after initiation of therapy.Positive controls were run in parallel using sera fromautoimmune patients. Sera were diluted in PBS and usedneat, 1:2 and 1:4. F(ab)02-specific fluorescein isothiocya-nate–conjugated goat antimouse or human antibodies(Cappel Laboratories, Cochranville, PA) were then usedas secondary reagents. The slides were then counter-stained with Evans Blue, mounted with 1:1 PBS:glycerol,and observed under a Leitz fluorescence microscope.
Survival StudyGroup of BALB/c mice (n=5) were injected with
Colon 26 or RENCA cells as described above. Five toseven days after tumor implantation, mice were treatedwith 1 nmol/dose Fc-mOX40L (100-mg dose level) orOX86 (200-mg dose level) daily � 5, and survival ofthe mice was recorded for 120 days. Significance levels(P values) were determined using the Wilcoxon rank-sumtest.
Flow Cytometry of Tumor-infiltratingLymphocytes
Mice from the control group and Fc-mOX40L-treated and OX86-treated groups injected with 500 pmol/dose (50-mg/dose and 75-mg/dose, respectively) were killedby sodium pentobarbital overdose 9 days after initiationof treatment when tumor reduction was at its greatest.Tumors and tumor-draining lymph nodes were removedand the lymphoid populations were isolated as describedpreviously.39 PE-labeled anti-CD4, anti-CD8, anti-CD11c,anti-CD25, anti-CD62L, anti-CCR7, and anti-CD49bantibodies were used to stain the tumor-infiltratinglymphocytes for flow cytometric analysis. After gatingon lymphocytes, the percentages of all lymphocytepopulations were calculated.
Morphologic StudiesTumors from treated and control Colon 26-bearing
mice were removed on day 15 posttumor implantation.Tumors were fixed in 10% neutral buffered formalin(VWR Scientific, West Chester, PA) and paraffin-embedded sections from Colon 26 tumor-bearing micewere stained with hematoxylin and eosin for morphologicanalysis.
J Immunother � Volume 31, Number 3, April 2008 Fc-mOX40L Immunotherapy
r 2008 Lippincott Williams & Wilkins 237
Leukocyte Subset Depletion andCombination Studies
To deplete CD4+, CD8+, CD25+, or natural killer(NK) cells, mice were injected IP on days 0, 7, and 14posttumor implantation with 0.5mg of purified anti-CD4antibody (GK1.5), anti-CD8 antibody (H35), anti-CD25antibody (PC61), or anti-NK antibody (rabbit antimouse/rat asialo GM1 polyclonal antibody, Cedarlane Labora-tories) in a 1-mL inoculum in PBS. Depletion of specificT-cell subsets was confirmed by flow cytometry analysisof lymph nodes of inoculated mice using antibody clonesthat differ from those used for depletion (data notshown). Tumor volumes were calculated 3� /wk bycaliper measurement in 3 dimensions for all groups ofmice.
RESULTS
Construction and Expression ofFc-mOX40L Fusion Protein
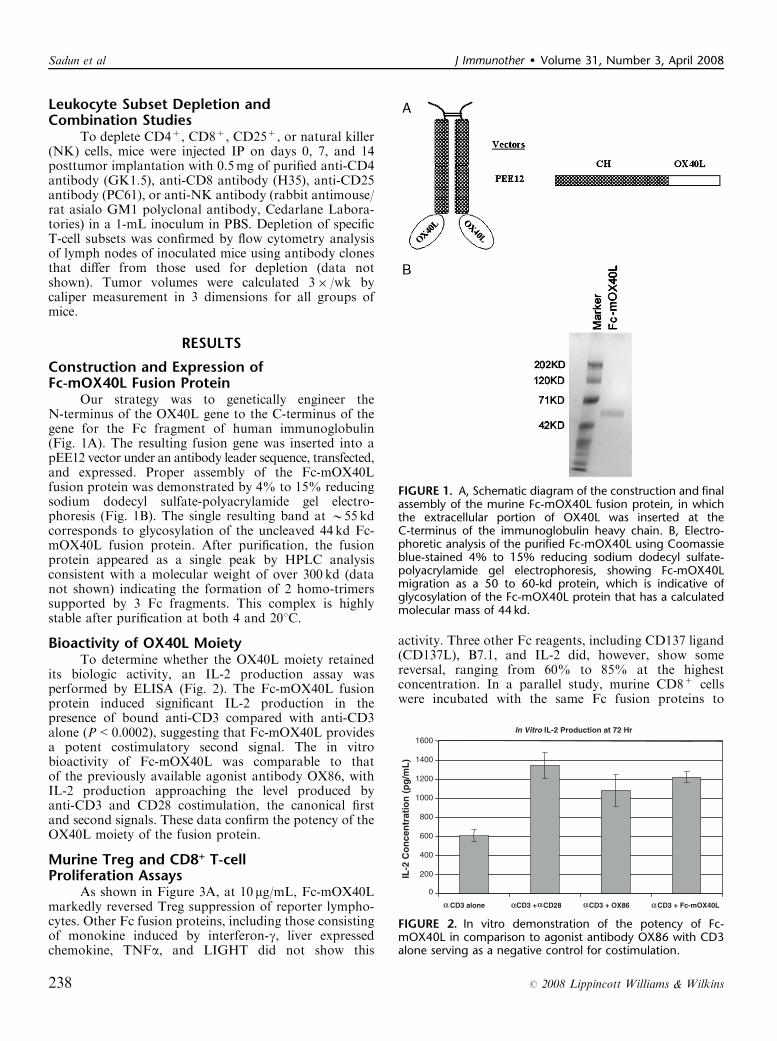
Our strategy was to genetically engineer theN-terminus of the OX40L gene to the C-terminus of thegene for the Fc fragment of human immunoglobulin(Fig. 1A). The resulting fusion gene was inserted into apEE12 vector under an antibody leader sequence, transfected,and expressed. Proper assembly of the Fc-mOX40Lfusion protein was demonstrated by 4% to 15% reducingsodium dodecyl sulfate-polyacrylamide gel electro-phoresis (Fig. 1B). The single resulting band at B55 kdcorresponds to glycosylation of the uncleaved 44 kd Fc-mOX40L fusion protein. After purification, the fusionprotein appeared as a single peak by HPLC analysisconsistent with a molecular weight of over 300 kd (datanot shown) indicating the formation of 2 homo-trimerssupported by 3 Fc fragments. This complex is highlystable after purification at both 4 and 201C.
Bioactivity of OX40L MoietyTo determine whether the OX40L moiety retained
its biologic activity, an IL-2 production assay wasperformed by ELISA (Fig. 2). The Fc-mOX40L fusionprotein induced significant IL-2 production in thepresence of bound anti-CD3 compared with anti-CD3alone (P<0.0002), suggesting that Fc-mOX40L providesa potent costimulatory second signal. The in vitrobioactivity of Fc-mOX40L was comparable to thatof the previously available agonist antibody OX86, withIL-2 production approaching the level produced byanti-CD3 and CD28 costimulation, the canonical firstand second signals. These data confirm the potency of theOX40L moiety of the fusion protein.
Murine Treg and CD8+ T-cellProliferation Assays
As shown in Figure 3A, at 10 mg/mL, Fc-mOX40Lmarkedly reversed Treg suppression of reporter lympho-cytes. Other Fc fusion proteins, including those consistingof monokine induced by interferon-g, liver expressedchemokine, TNFa, and LIGHT did not show this
activity. Three other Fc reagents, including CD137 ligand(CD137L), B7.1, and IL-2 did, however, show somereversal, ranging from 60% to 85% at the highestconcentration. In a parallel study, murine CD8+ cellswere incubated with the same Fc fusion proteins to
FIGURE 1. A, Schematic diagram of the construction and finalassembly of the murine Fc-mOX40L fusion protein, in whichthe extracellular portion of OX40L was inserted at theC-terminus of the immunoglobulin heavy chain. B, Electro-phoretic analysis of the purified Fc-mOX40L using Coomassieblue-stained 4% to 15% reducing sodium dodecyl sulfate-polyacrylamide gel electrophoresis, showing Fc-mOX40Lmigration as a 50 to 60-kd protein, which is indicative ofglycosylation of the Fc-mOX40L protein that has a calculatedmolecular mass of 44 kd.
In Vitro IL-2 Production at 72 Hr
0
200
400
600
800
1000
1200
1400
1600
α α α ααCD3 alone CD3 + CD28 CD3 + OX86 CD3 + Fc-mOX40L
IL-2
Co
nce
ntr
atio
n (
pg
/mL
)
FIGURE 2. In vitro demonstration of the potency of Fc-mOX40L in comparison to agonist antibody OX86 with CD3alone serving as a negative control for costimulation.
Sadun et al J Immunother � Volume 31, Number 3, April 2008
238 r 2008 Lippincott Williams & Wilkins

determine their effects on this critical effector cellpopulation. The results of these studies (Fig. 3B) indicatethat IL-2–Fc had the greatest proliferative response onCD8+ cells as expected, but that Fc-mOX40L also hadmoderate proliferative effects on this subpopulation ofT cells. As shown in Figure 3C, several of the fusionproteins directly caused the proliferation of Treg cells. Asthe suppressive Treg cells do not proliferate to anti-CD3stimulation in vitro, this is an indication that these cellsare losing their suppressive phenotype. This backgroundproliferation was subtracted in the percentage suppres-sion calculations. From these data, it seems that one of
the key actions of Fc-mOX40L may be the reversal ofTreg suppression on tumor-infiltrating T cells.
Immunotherapeutic Dosing StudiesA dosing study was performed on Colon 26-bearing
and RENCA-bearing BALB/c mice using IV equimolardoses of Fc-mOX40L (1 to 100 mg/dose) or OX86 (1.5 to150 mg/dose), with IV injection of PBS serving as anegative control (Fig. 4). At 1 mg/dose, Fc-mOX40Ldid not produce measurable tumor reduction in eithertreatment model, whereas the molar equivalent of OX86,1.5 mg/dose, produced minimal tumor regression in theColon 26 model (Fig. 4A). As the concentration of thetreatments increased, so did the rapidity with which Fc-mOX40L produced complete remission in the Colon 26model, whereas the partial remission effects of OX86plateaued and failed to show continued dose-dependencein Colon 26-bearing mice (Fig. 4A). In the RENCAmodel, 15 mg/dose to 150 mg/dose produced identicalresults, in which OX86 failed to bring about measurablereduction in tumor volume, whereas Fc-mOX40L rapidlyproduced complete remission at molar equivalent doses(Fig. 4B).
Toxicity StudiesThe appearance of treated mice and their activity
levels did not suggest the presence of autoimmune sideeffects after therapy. This was confirmed by indirectimmunofluorescence studies of mouse sera that, unlikepositive controls, showed the absence of antibodies tonormal cellular antigens.
Survival StudiesSurvival studies were performed using equimolar
concentrations of Fc-mOX40L (50 mg/dose) or of OX86(75 mg/dose). All mice treated with Fc-mOX40L survivedfor the full observation period of 113 days, whereastreatment with OX86 succeeded only in extending themedian survival from 55 to 70 days in Colon 26-bearingmice (Fig. 4C) and from 45 days to 65 days in RENCA-bearing mice (Fig. 4D). In contrast to Fc-mOX40L-treated mice, no control mice or OX86-treated micesurvived beyond 80 days in either tumor model(P<0.05).
Leukocyte Subset Depletion andCombination Studies
To assess the role of specific leukocyte populationsin Fc-mOX40L–mediated immunotherapy, individual cellpopulations were depleted by monoclonal antibodyadministration. Depletion was confirmed by flow cyto-metry analysis of splenic populations (data not shown).As seen in Figure 5A, anti-CD4 and anti-CD25 depletionproduced, by themselves, significant reduction in tumorsize, whereas anti-NK depletion also brought aboutmeasurable reduction in tumor size. When Fc-mOX40L–treated mice were depleted by these procedures (Fig. 5B),anti-CD4 and anti-NK treatment enhanced the antitumoreffects of Fc-mOX40L treatment, whereas anti-CD8abrogated the effect of Fc-mOX40L treatment, indicating
FIGURE 3. Effect of different Fc fusion proteins on (A)percentage suppression of CD8+ cells by Treg cells, (B)proliferation of CD8+ T cells, and (C) proliferation of Tregcells. Note that the Fc-mOX40L induced the proliferationof CD8+ cells but markedly inhibited Treg suppression ofproliferating CD8+ T cells.
J Immunother � Volume 31, Number 3, April 2008 Fc-mOX40L Immunotherapy
r 2008 Lippincott Williams & Wilkins 239

that Fc-mOX40L is dependent upon CD8+ T cells for itstherapeutic effects.
Characterization of Tumor-infiltratingLymphocytes
Using equimolar doses of Fc-mOX40L and OX86(50 and 75 mg/dose, respectively), the relative compositionof tumor-infiltrating lymphocytes was determined by flowcytometry at day 23 (Table 1). In Colon 26-bearing mice,both Fc-mOX40L and OX86 induced a significant tumorinfiltration of CD11c+ cells, whereas only in the
Fc-mOX40L–treated mice, there was a significant in-crease in tumor infiltration of CD8+ cells (6.73% of gatedlymphocytes in comparison to 0.7% in the controland 0.4% in OX86-treated mice). Both Fc-mOX40Land OX86 treatment produced a decrease in the tumor-infiltrating CD25+ and CD49+ cells in the Colon 26model, and in the tumor-draining lymph nodes of Colon26-bearing mice, there was a decrease in CD11c+ andCD49+ cells.
In RENCA-bearing mice, almost none of the samepatterns were observed. In particular, there was no
10 pmole/dose
0
0.2
0.4
0.6
0.8
1
5 7 9 12 14 16 19 21 23
Days after tumor implantation
tum
or
volu
me
(cm
3)
Control
OX86
Fc-mOX40L
100 pmole/dose
0
0.2
0.4
0.6
0.8
1
5 7 9 12 14 16 19 21 23
Days after tumor implantation
tum
or
volu
me
(cm
3)
Control
OX86
500 pmole/dose
0
0.2
0.4
0.6
0.8
1
5 7 9 12 14 16 19 21 23
tum
or
volu
me
(cm
3) Control
OX86
500 pmole/dose
0
0.1
0.2
0.3
0.4
5 7 9 12 14 16 19 21 23
Days after tumor implantation
tum
or
volu
me
(cm
3)
Control
OX86
A
B
C
D RENCA Kaplan Meier Survival Plot
0.00
0.25
0.50
0.75
1.00
0 30 60 90 120
Days Following Tumor Implantation
Est
imat
ed P
roba
bilit
y o
f S
urvi
val
p<0.001
Fc-mOX40L
OX86
PBS Control
Days after tumor implantation
Colon 26 Kaplan Meier Survival Plot
0.00
0.25
0.50
0.75
1.00
0 30 60 90 120
Days Following Tumor Implantation
Est
imat
ed P
roba
bilit
y o
f S
urvi
val
p<0.001
Fc-mOX40L
OX86
PBS Control
Fc-mOX40L
Fc-mOX40L
Fc-mOX40L
FIGURE 4. Dose response of Fc-mOX40L in (A) Colon 26-bearing and (B) RENCA-bearing BALB/c mice. Kaplan-Meier survivalanalysis of Fc-mOX40L in (C) Colon-26-bearing and (D) RENCA-bearing BALB/c mice.
Sadun et al J Immunother � Volume 31, Number 3, April 2008
240 r 2008 Lippincott Williams & Wilkins

Fc-mOX40L treatment-induced increase in tumor-infil-trating CD8+ cells, the presumed effector cell forTNFSF-induced antitumor activity. Also notable, RE-NCA tumor-draining lymph nodes had much greaterpercentages of positive staining cells across the board,and whereas even untreated RENCA tumor-draininglymph nodes had more lymphocytes than Colon 26-
bearing mice, Fc-mOX40L treatment was able to increasefurther the percentage of CD8+ and CD11b+ cells,though CD25+ cells were also increased in the tumor-draining lymph nodes of RENCA-bearing mice.
Morphologic StudiesHematoxylin and eosin slides were prepared from
paraffin-embedded tumor tissues removed from Colon26-bearing and RENCA-bearing mice 6 days after thecompletion of treatment (day 15) with daily � 5 injectionsof equimolar Fc-mOX40L (100 mg/dose) and the OX86agonist antibody (150 mg/dose) (Fig. 6). Tumors fromPBS-treated control mice showed a necrotic core typicalof such large tumors. Tumors from mice treated witheither Fc-mOX40L or OX86, although much smallerbecause of successful treatment, had prominent necrosis.In Colon 26-bearing mice, Fc-mOX40L inducedcoagulative necrosis throughout the tumor, with highmagnification revealing infiltration of mononuclearcells. OX86 treatment, however, produced necrosisconcomitant with heavy neutrophil infiltration and aliquefactive pattern of tumor lysis. In RENCA-bearingmice, the same pattern of necrosis was observed as withColon 26-bearing mice, though the tumors were largerand the necrosis less pronounced (data not shown).Morphologic analysis of tumors treated with either Fc-mOX40L or OX86 did not show evidence of vasculardamage such as vascular thrombosis or red cell extra-vasation.
DISCUSSIONOver the past several years, many costimulatory
ligands belonging to the TNFSF and B7 families havebeen investigated, both by our laboratory and others.Although many of these costimulatory molecules haveshown a degree of in vivo antitumor activity, fewmolecules have been as promising as OX40L. Signalingthrough OX40 stimulates both primary and effectorresponses, and activates both CD4+ and CD8+ effectorcells. Remarkably, activation using OX86 has beenshown to be tumor-specific and effector cell specific,actually inhibiting the response of suppressor Treg cells.Although OX86 has been shown by others to reducetumor growth or synergize with other immunotherapies,in our hands, Fc-mOX40L is far more potent than OX86and is capable of producing complete and lastingremissions.
The genetically engineered Fc-mOX40L expressedby NS0 cells was shown to remain intact (Fig. 1B) and toretain its bioactivity (Fig. 2). HPLC analysis revealed thatthe molecule had a molecular weight of approximately300 kd indicating that 3 Fc fusion proteins were joinedtogether to form 2 trimers of OX40L. As previouslyshown, OX40L consists of 3 peptides accounting for thegeneration of the 2 trimer configuration.36 In IL-2induction studies, Fc-mOX40L seems to have greaterbioactivity than OX86, stimulating B15% more T-cellproduction of IL-2. This degree of costimulation fallswithin 10% of the maximal IL-2 production induced by
0
100
200
300
400
500
600
700
800
5 7 9 12 14 16 19 21 23
days
tum
or
volu
me
(mm
3)
no treatment
anti-CD4 (days 0, 7, 14)
anti-CD8 (days 0, 7, 14)
anti-CD25 (day 0)
anti-NK (days 0, 7, 14)
TREATMENT
0
100
200
300
400
500
600
700
800
5 7 9 12 14 16 19 21 23
days
tum
or
volu
me
(mm
3)
no treatment
Fc-mOX40L (30ug/dose +anti-CD4 (days 0, 7, 14)
Fc-mOX40L 30ug/dose +anti-CD8 (days 0, 7, 14)
Fc-mOX40L (30ug/dose) + anti-CD25 (day 0)
Fc-mOX40L(30ug/dose) +anti-NK (days 0, 7, 14)
Fc-mOX40L (30ug/dose)
TREATMENT
A
B
FIGURE 5. Effect of T cell subpopulation depletion on Colon26 tumor growth either (A) alone or (B) in combination withFc-mOX40L treatment. Note that only depletion of CD8+
T cells reversed the therapeutic effects of Fc-mOX40L.
TABLE 1. Immunophenotype of Tumor and TDLNs in Fc-mOX40L–treated, OX86-treated, and Control-treated Mice
RENCA Tissue Control (%) Fc-OX40L (%) OX86 (%)
CD4+ 0 0.44 0.55CD8+ 0.24 0.87 0.50CD11b+ 10.24 6.46 5.55CD25+ 0.60 0.56 0.11CD49+ 4.25 2.47 3.79
Colon 26 tumorCD4+ 3.81 32.13 8.19CD8+ 0.70 6.73 0.41CD11b+ 2.77 8.98 6.68CD25+ 1.42 0.42 0.51CD49+ 23.32 1.49 9.99CD62L+/CCR7+ 1.71 13.88 4.40CD4+/CD25� 2.21 26.02 4.90
RENCA TDLNCD4+ 23.28 23.94 15.83CD8+ 4.28 10.15 5.30CD11b+ 5.81 20.97 8.20CD25+ 4.23 17.06 4.75CD49+ 14.76 18.93 7.73
Colon 26 TDLNCD4+ 2.34 2.94 5.23CD8+ 1.59 2.38 3.04CD11b+ 1.68 10.02 5.85CD25+ 1.64 2.00 2.66CD49+ 3.67 6.26 9.27
J Immunother � Volume 31, Number 3, April 2008 Fc-mOX40L Immunotherapy
r 2008 Lippincott Williams & Wilkins 241

CD3/CD28 stimulation. Indeed, Fc-mOX40L-inducedIL-2 production exceeded that produced by othercostimulatory fusion proteins produced in our laboratory,including B7.1-Fc,40 Fc-mCD137L,35 and Fc-mGITRL.41
The fact that Fc-mOX40L’s potency exceeds that ofB7.1-Fc, which captures the costimulatory activity of theB7-CD28 signaling pathway, suggests that Fc-mOX40L
has an unparalleled capacity to activate CD8+ effectorT cells. As important, Treg activity assays showed thatFc-mOX40L, like Fc-mGITRL41 inhibits Treg suppres-sion yet enhances CD8+ T-cell proliferation (Fig. 3). In arecent study by Vu et al,42 a newly created Foxp3gfpknockin model showed that OX40 was dispensable for thegenesis and suppressor functions of naturally occurringCD4+Foxp3+ Tregs, but stimulating OX40 on theFoxp3+ Treg cells negated their ability to suppressCD8+ effector cell proliferation, IFN-g production,and cell-mediated allograft rejection. Moreover, OX40costimulation profoundly inhibited Foxp3 gene expres-sion. Based on their data, these investigators haveidentified OX40 as a key negative regulator of Foxp3+
Treg cells.In vivo, Fc-mOX40L was also found to be
extremely potent, consistent with early studies conductedby Ali et al43 using mOX40L-Ig fusion protein prepara-tions. In Figure 4A, it can be seen that Fc-mOX40L isinactive at 1 mg/dose, but at 10 mg/dose and higherproduces complete remission in both Colon 26-bearingand RENCA-bearing mice. At 50 mg/dose, Fc-mOX40Lprevents appreciable tumor growth in both Colon 26 andRENCA models (Figs. 4A, B). Fc-mOX40L was testedat up to 100 mg/dose (not shown), wherein it producedcomplete remission much as seen with 50 mg/dose. Bycomparison, OX86 agonist antibody produced a partialremission, plateauing at 80% reduction in tumor volume,even at the 150 mg/dose.
The therapeutic window of Fc-mOX40L is ideal foran immunotherapeutic drug, because 10 mg/dose pro-duced complete remission in all treated mice, but micetreated with doses as great as 100 mg/dose did not showany signs of toxicity. Toxicologic studies in rhesusmonkeys with an anti-OX40 agonist antibody did notreveal side effects, and at necropsy, monkeys were foundto have enlarged splenic and gut-associated lymph nodesfor up to 28 days posttreatment, a finding consistent forT-cell activators.44 Immunohistochemistry confirmedspleen and lymph node lymphocyte hyperplasia andshowed no signs of destruction owing to inflammation,suggesting OX40-OX40L pathway stimulation to be arelatively safe immunotherapeutic response.44 Sera
FIGURE 6 Morphologic analysis of tumors from (A) control,(B) Fc-mOX40L-treated, and (C) OX86-treated Colon 26-bearing mice at day 15 posttumor implantation (6 d aftercompletion of immunotherapy). In panel A, note thepresence of small areas of patchy necrosis (white arrow)amidst nests of viable cells (black arrows) in phosphate-buffered saline-treated control tumors. By contrast, panel Bshows Fc-mOX40L–treated tumor with massive coagulativenecrosis (yellow arrow) along with significant mononuclearinfiltration (teal arrow). Panel C shows OX86-treated tumorswith large, central areas of liquefactive necrosis rich inneutrophils (purple arrow) along with small areas of viabletumor (green arrow). (hematoxylin and eosin, �100, insets�400).
Sadun et al J Immunother � Volume 31, Number 3, April 2008
242 r 2008 Lippincott Williams & Wilkins

obtained from Fc-mOX40L-treated mice did not showany positivity for normal cellular antigens by indirectimmunofluorescence microscopy indicating that auto-immune complications were not induced. Because Fc-mOX40L produces complete remission at doses far lowerthan agonist antibody but has similar tolerability at veryhigh doses, Fc-mOX40L is an even safer antitumortherapy.
In survival studies, Fc-mOX40L again proved to besuperior to the well-known OX86 antibody. As seen inFigure 4A, the partial remission produced by 75 mg/doseOX86 in Colon 26-bearing mice was not a lastingresponse, with all mice dying before the middle of thethird month and a mean increase in life expectancy ofonly 2 weeks. Fc-mOX40L at 50 mg/dose, however,produced lasting remissions and mice survived throughthe completion of the study, roughly 4 months later. InFigure 4B, the same trend was evident in RENCA-bearing mice, with OX86 increasing mean survival byroughly 3 weeks, but OX86-treated mice dying early inthe third month. Fc-mOX40L mice, however, were able tosustain lasting remission in the very aggressive RENCAmodel, with all surviving through the end of the study. Asshown in Figure 5, depletion studies in treated micebearing Colon 26 tumors showed that CD8+ T cells werecritical for Fc-mOX40L immunotherapy, but unlike otherimmunotherapeutic ligands such as the chemokine liverexpressed chemokine,45,46 NK cells did not seem to berequired for tumor regression.
In Figure 6, treatment of Colon 26-bearing mice canbe seen to induce tumor necrosis in excess of that whichwould be expected for the size of the tumor. Interestingly,although both Fc-mOX40L and OX86 induced tumorlysis, they did so with a different pattern of necrosis. Inparticular, Fc-mOX40L produced massive central necro-sis and OX86 produced a peripheral ring of necrosis.Although treatment-induced necrosis was less pro-nounced in the RENCA model, it also followed the samepattern (data not shown). The central necrosis induced byFc-mOX40L is likely an indicator of better prognosisbecause many treatments have been found to fail becauseof their inability to overcome the high oncotic pressure oftumors that makes infiltration and deep permeationchallenging.47–49 The trimer Fc complex of the Fc-mOX40L may account for its higher activity thanOX86, which acts as an agonist on the OX40 receptor.Consistent with this notion, Morris et al36 also found thatthe natural ligand is more potent than agonist antibodiesto the OX40 receptor for the immunotherapy of solidtumors.
Finally, flow cytometric analyses of tumor-infiltrat-ing lymphocytes presented in Table 1 showed that,although the overall number of CD8+ cells in Fc-mOX40L–treated mice was only marginally greater thanthat seen in OX86-treated mice, the number ofCD4+CD25� and CD62L+CCR7+ cells was substan-tially higher in Fc-mOX40L–treated mice than in OX86-treated mice (5.3-fold and 8.1-fold increase, respectively).These results suggest that the activation status of
cytotoxic T cells may be more important than the overallnumber of infiltrating lymphocytes in tumor.CD62L+CCR7+ coexpression has been shown tocharacterize a population of T memory cells known ascentral memory cells (Tcm), which studies suggest confersuperior tumor immunity.50 Furthermore, CD4+CD25�
T helper cells have been shown to influence thedevelopment of Tcm memory cells.51 It thus seemsthat natural stimulation of the OX40 receptor byFc-mOX40L succeeds in producing a more physiologicresponse by promoting established OX40 pathways,including the stimulation of T helper cells and theactivation of cytotoxic CD8+ T effector and centralmemory cells.
By all measures, Fc-mOX40L succeeds in activatingan antitumor response, as would be expected of many ofthe ligand members of the TNF superfamily. Thoughother receptor members of the TNFR superfamily havebeen studied for their therapeutic potential with sub-stantial but not total success, OX40 seems to have severaladvantages over its superfamily sisters. In one study,investigators identified a dichotomy between the costi-mulatory action of OX40 and 4-1BB, finding that OX40enhances early effector function and late accumulationsurvival, whereas the early effects of 4-1BB may act in anegative, regulatory fashion and serve to limit primaryCD8+ responses.52
In summary, we provide the first description of anFc-mOX40L fusion protein that demonstrates in vitroimmune stimulation and in vivo antitumor activity. Inour hands, this Fc-mOX40L has proven more potent thanthe OX86 agonist antibody, both in vitro and in vivo.Stimulation of the OX40-OX40L pathway has provento be as potent as B7.1 stimulation,21 but has the addedbenefits of producing an antigen-specific immune re-sponse23,24 and Treg inhibition. Additionally, stimulationof the OX40-OX40L pathway has been shown to produceminimal to no toxicity, even when significant immuneactivation is induced.43 In light of the significant andlasting tumor remission and increased survival broughtabout by Fc-mOX40L in both Colon 26-bearing andRENCA-bearing mice, this highly potent fusion proteinseems to be a strong candidate for immunotherapy ofsolid tumors, meeting and exceeding the high expectationsfor OX86.
ACKNOWLEDGMENTSThe authors thank James Pang for his assistance with
the animal experiments, Mandy Han for her assistancewith cloning procedures, and Janet Kim for her assistancewith the immunohistochemistry. The authors also acknowl-edge the support of Dr Jeffrey Schlom, National CancerInstitute, Bethesda, MD.
REFERENCES1. Mallett S, Fossum S, Barclay AN. Characterization of the MRC
OX40 antigen of activated CD4 positive T lymphocytes—a moleculerelated to nerve growth factor receptor. Embo J. 1990;9:1063–1068.
J Immunother � Volume 31, Number 3, April 2008 Fc-mOX40L Immunotherapy
r 2008 Lippincott Williams & Wilkins 243

2. Calderhead DM, Buhlmann JE, van den Eertwegh AJ, et al. Cloningof mouse OX40: a T cell activation marker that may mediate T-Bcell interactions. J Immunol. 1993;151:5261–5271.
3. Paterson DJ, Jefferies WA, Green JR, et al. Antigens of activated ratT lymphocytes including a molecule of 50,000 Mr detected only onCD4 positive T blasts. Mol Immunol. 1987;24:1281–1290.
4. Miura S, Ohtani K, Numata N, et al. Molecular cloning andcharacterization of a novel glycoprotein, gp34, that is specificallyinduced by the human T-cell leukemia virus type I transactivatorp40tax. Mol Cell Biol. 1991;11:1313–1325.
5. Godfrey WR, Fagnoni FF, Harara MA, et al. Identification of ahuman OX-40 ligand, a costimulator of CD4+ T cells withhomology to tumor necrosis factor. J Exp Med. 1994;180:757–762.
6. Stuber E, Neurath M, Calderhead D, et al. Cross-linking of OX40ligand, a member of the TNF/NGF cytokine family, inducesproliferation and differentiation in murine splenic B cells. Immunity.1995;2:507–521.
7. Ohshima Y, Tanaka Y, Tozawa H, et al. Expression and functionof OX40 ligand on human dendritic cells. J Immunol. 1997;159:3838–3848.
8. Weinberg AD, Wegmann KW, Funatake C, et al. Blocking OX-40/OX-40 ligand interaction in vitro and in vivo leads to decreasedT cell function and amelioration of experimental allergic encepha-lomyelitis. J Immunol. 1999;162:1818–1826.
9. Imura A, Hori T, Imada K, et al. The human OX40/gp34 systemdirectly mediates adhesion of activated T cells to vascularendothelial cells. J Exp Med. 1996;183:2185–2195.
10. Pippig SD, Pena-Rossi C, Long J, et al. Robust B cell immunitybut impaired T cell proliferation in the absence of CD134 (OX40).J Immunol. 1999;163:6520–6529.
11. Takasawa N, Ishii N, Higashimura N, et al. Expression of gp34(OX40 ligand) and OX40 on human T cell clones. Jpn J Cancer Res.2001;92:377–382.
12. Soroosh P, Ine S, Sugamura K, et al. OX40-OX40 ligand interactionthrough T cell-T cell contact contributes to CD4 T cell longevity.J Immunol. 2006;176:5975–5987.
13. Gramaglia I, Jember A, Pippig SD, et al. The OX40 costimulatoryreceptor determines the development of CD4 memory by regulatingprimary clonal expansion. J Immunol. 2000;165:3043–3050.
14. Maxwell JR, Weinberg A, Prell RA, et al. Danger andOX40 receptor signaling synergize to enhance memory T cellsurvival by inhibiting peripheral deletion. J Immunol. 2000;164:107–112.
15. Gramaglia I, Weinberg AD, Lemon M, et al. Ox-40 ligand: a potentcostimulatory molecule for sustaining primary CD4 T cell responses.J Immunol. 1998;161:6510–6517.
16. Rogers PR, Song J, Gramaglia I, et al. OX40 promotes Bcl-xL andBcl-2 expression and is essential for long-term survival of CD4T cells. Immunity. 2001;15:445–455.
17. Murata K, Ishii N, Takano H, et al. Impairment of antigen-presenting cell function in mice lacking expression of OX40 ligand.J Exp Med. 2000;191:365–374.
18. Weinberg AD, Vella AT, Croft M. OX-40: life beyond the effectorT cell stage. Semin Immunol. 1998;10:471–480.
19. Wang HC, Klein JR. Multiple levels of activation of murineCD8(+) intraepithelial lymphocytes defined by OX40 (CD134)expression: effects on cell-mediated cytotoxicity, IFN-gamma, andIL-10 regulation. J Immunol. 2001;167:6717–6723.
20. Bansal-Pakala P, Halteman BS, Cheng MH, et al. Costimulationof CD8 T cell responses by OX40. J Immunol. 2004;172:4821–4825.
21. Serghides L, Bukczynski J, Wen T, et al. Evaluation of OX40ligand as a costimulator of human antiviral memory CD8 T cellresponses: comparison with B7.1 and 4-1BBL. J Immunol. 2005;175:6368–6377.
22. Fujita T, Ukyo N, Hori T, et al. Functional characterization ofOX40 expressed on human CD8+ T cells. Immunol Lett. 2006;106:27–33.
23. Ruby CE, Redmond WL, Haley D, et al. Anti-OX40 sti-mulation in vivo enhances CD4+ memory T cell survival and
significantly increases recall responses. Eur J Immunol. 2007;37:157–166.
24. Maxwell JR, Yadav R, Rossi RJ, et al. IL-18 bridges innate andadaptive immunity through IFN-gamma and the CD134 pathway.J Immunol. 2006;177:234–245.
25. Gavin MA, Clarke SR, Negrou E, et al. Homeostasis and anergy ofCD4(+)CD25(+) suppressor T cells in vivo. Nat Immunol. 2002;3:33–41.
26. McHugh RS, Whitters MJ, Piccirillo CA, et al. CD4(+)CD25(+)immunoregulatory T cells: gene expression analysis reveals afunctional role for the glucocorticoid-induced TNF receptor.Immunity. 2002;16:311–323.
27. Takeda I, Ine S, Killeen N, et al. Distinct roles for the OX40-OX40ligand interaction in regulatory and nonregulatory T cells.J Immunol. 2004;172:3580–3589.
28. Valzasina B, Guiducci C, Dislich H, et al. Triggering of OX40(CD134) on CD4(+)CD25+ T cells blocks their inhibitory activity:a novel regulatory role for OX40 and its comparison with GITR.Blood. 2005;105:2845–2851.
29. Weinberg AD, Rivera MM, Prell R, et al. Engagement of the OX-40receptor in vivo enhances antitumor immunity. J Immunol. 2000;164:2160–2169.
30. Lee SJ, Myers L, Muralimohan G, et al. 4-1BB and OX40dual costimulation synergistically stimulate primary specificCD8 T cells for robust effector function. J Immunol. 2004;173:3002–3012.
31. Gri G, Gallo E, Di Carlo E, et al. OX40 ligand-transduced tumorcell vaccine synergizes with GM-CSF and requires CD40-Apcsignaling to boost the host T cell antitumor response. J Immunol.2003;170:99–106.
32. Pan PY, Zang Y, Weber K, et al. OX40 ligation enhances pri-mary and memory cytotoxic T lymphocyte responses in animmunotherapy for hepatic colon metastases. Mol Ther. 2002;6:528–536.
33. Andarini S, Kikuchi T, Nukiwa M, et al. Adenovirus vector-mediated in vivo gene transfer of OX40 ligand to tumor cellsenhances antitumor immunity of tumor-bearing hosts. Cancer Res.2004;64:3281–3287.
34. Kaneko H, Hori T, Yanagita S, et al. Introduction of OX40 ligandinto lymphoma cells elicits anti-lymphoma immunity in vivo. ExpHematol. 2005;33:336–343.
35. Zhang N, Sadun RE, Arias RS, et al. Targeted and untargetedCD137L fusion proteins for the immunotherapy of experimentalsolid tumors. Clin Cancer Res. 2007;13:2758–2767.
36. Morris NP, Peters C, Montler R, et al. Development andcharacterization of recombinant human Fc:OX40L fusion proteinlinked via a coiled-coil trimerization domain. Mol Immunol.2007;44:3112–3121.
37. Taylor L, Bachler M, Duncan I, et al. In vitro and in vivoactivities of OX40 (CD134)-IgG fusion protein isoforms withdifferent levels of immune-effector functions. J Leukoc Biol. 2002;72:522–529.
38. Li J, Hu P, Khawli LA, et al. chTNT-3/hu IL-12 fusion protein forthe immunotherapy of experimental solid tumors. Hybrid Hybri-domics. 2004;23:1–10.
39. Sanderson K, Scotland R, Lee P, et al. Autoimmunity in a phase Itrial of a fully human anti-cytotoxic T-lymphocyte antigen-4monoclonal antibody with multiple melanoma peptides andMontanide ISA 51 for patients with resected stages III and IVmelanoma. J Clin Oncol. 2005;23:741–750.
40. Liu A, Hu P, Khawli LA, et al. Combination B7-Fc fusion proteintreatment and Treg cell depletion therapy. Clin Cancer Res.2005;11:8492–8502.
41. Hu P, Arias RS, Sadun RE, et al. Construction and pre-clinicalcharacterization of Fc-mGITRL for the immunotherapy of cancer.Clin Cancer Res. 2008;14:579–588.
42. Vu MD, Xiao X, Gao W, et al. OX40 costimulation turns offFoxp3+ Tregs. Blood. 2007;110:2501–2510.
43. Ali SA, Ahmad M, Lynam J, et al. Anti-tumour therapeuticefficacy of OX40L in murine tumour model. Vaccine. 2004;22:3585–3594.
Sadun et al J Immunother � Volume 31, Number 3, April 2008
244 r 2008 Lippincott Williams & Wilkins

44. Weinberg AD, Thalhofer C, Morris N, et al. Anti-OX40 (CD134)administration to nonhuman primates: immunostimulatory effectsand toxicokinetic study. J Immunother. 2006;29:575–585.
45. Li J, Hu P, Khawli LA, et al. LEC/chTNT-3 fusion protein for theimmunotherapy of experimental solid tumors. J Immunother.2003;26:320–331.
46. Li J, Hu P, Khawli LA, et al. Complete regression of experi-mental solid tumors by combination LEC/chTNT-3 immuno-therapy and CD25(+) T-cell depletion. Cancer Res. 2003;63:8384–8392.
47. Karakousis CP, Rao U, Holtermann OA, et al. Tourniquet infusionchemotherapy in extremities with malignant lesions. Surg GynecolObstet. 1979;149:481–490.
48. Di Paolo A, Bocci G. Drug distribution in tumors: mechanisms, rolein drug resistance, and methods for modification. Curr Oncol Rep.2007;9:109–114.
49. Heldin CH, Rubin K, Pietras K, et al. High interstitial fluid pressure—an obstacle in cancer therapy. Nat Rev Cancer. 2004;4:806–813.
50. Wherry EJ, Teichgraber V, Becker TC, et al. Lineage relationshipand protective immunity of memory CD8+ T cell subsets. NatImmunol. 2003;4:225–234.
51. Northrop JK, Shen H. CD8 T-cell memory: only the good ones last.Curr Opin Immnol. 2004;16:451–455.
52. Lee SW, Park Y, Song A, et al. Functional dichotomy betweenOX40 and 4-1BB in modulating effector CD8 T cell responses.J Immunol. 2006;177:4464–4472.
J Immunother � Volume 31, Number 3, April 2008 Fc-mOX40L Immunotherapy
r 2008 Lippincott Williams & Wilkins 245




















