Fc-mediated transport of nanoparticles across airway epithelial cell layers
8
Fc-mediated transport of nanoparticles across airway epithelial cell layers Driton Vllasaliu a, ⁎, Cameron Alexander a , Martin Garnett a , Mike Eaton b , Snow Stolnik a a Division of Drug Delivery and Tissue Engineering, School of Pharmacy, University of Nottingham, Nottingham, NG7 2RD, UK b UCB Pharma, 208 Bath Road, Slough, Berkshire, SL1 3WE, UK abstract article info Article history: Received 5 September 2011 Accepted 10 December 2011 Available online xxxx Keywords: IgG FcRn pathway Mucosal delivery Nanoparticle transport Surface modification In a study directed towards non-invasive delivery of therapeutic biomacromolecules, we examined whether surface modification of sub-200 nm model nanoparticles with the Fc portion of IgG promotes their cell uptake and transport across the airway epithelial cells. The study initially confirms the expression of the relevant receptor, namely neonatal Fc receptor (FcRn), by Calu-3 cell layers simulating the airway epithelium and demonstrates FcRn-mediated cell association, internalization and transcellular transport of molecular IgG. Surface decoration of nanoparticles with the Fc portion of IgG enhanced both cell uptake and translocation of the particulate system across the cell layers, in a manner strongly suggesting FcRn involvement in these processes. The study further demonstrates the potential of Fc-modified nanoparticles to ‘shuttle’ a model therapeutic antibody fragment across the epithelial cell layers. Fc-modified nanoparticles are transported in the μg/h/cm 2 range, presenting a substantial increase in transport capacity in comparison to molecular IgG (ng/h/cm 2 range), therefore warranting consideration of the FcRn transcytotic pathway for further inves- tigation as a means to achieve transmucosal delivery of nanoparticulate systems that could act as carriers of a range of biotherapeutics. © 2011 Elsevier B.V. All rights reserved. 1. Background Recent advances in biotechnology have produced a considerable increase in the number of proteins that might act as biological thera- peutics. However, this trend has not been paralleled by advances in novel delivery routes for this class of therapeutics. Non-invasive ad- ministration of biotherapeutics across the mucosal surfaces would be desirable, however, absorption of macromolecules through the mucosae is severely limited by the physical and biochemical barriers associated with this route. Improvement in mucosal absorption of some protein therapeutics is potentially achievable by inducing tran- sient opening of the epithelial tight junctions, but this strategy appears to be limited to delivery of smaller peptides and proteins [1–5], whilst delivery of larger proteins, such as antibodies (M W approximately 150,000 g mol -1 ), by using tight junction opening strategies is inefficient [6]. Transcellular transport of large therapeutic molecules across the epithelium, rather than relying on the paracellular route, would potentially overcome this problem. Unlike the non- specificity of the paracellular route, the transcellular pathway may be a receptor-mediated process and hence potentially selective to the molecule of interest. Immunoglobulin G (IgG) is actively transferred from mother to offspring, conferring short-term passive immunity [7,8], and has been shown to traverse a number of mucosal surfaces in man where it is thought to play an important role in the function of IgG in immune surveillance and host defense at mucosal surfaces [9]. IgG is transported by the neonatal Fc receptor (FcRn) [10,11]. From a drug delivery perspective, the IgG/FcRn transport pathway could be exploited to deliver biotherapeutics as this system is functional in several organs and tissues, including the lung [12] and intestine [13]. The primary aim of our study was to determine whether trans- port of nanoparticles across polarized epithelial cells could be promoted by coating their surfaces with the Fc portion of IgG. Although the IgG/FcRn pathway has previously been investigated for mucosal delivery of protein therapeutics in molecular form, to the best of our knowledge this system has not been considered for transepithelial transport of nanoparticles. Our studies were con- ducted using a human bronchial epithelial cell line, Calu-3, as an in vitro model of the human airway epithelium. Initial work assessed whether this cell line expresses FcRn. Cellular uptake of molecular IgG (in solution) and its transport across the cell layers was initially determined and the role of FcRn in these processes established. Subsequent transport experiments and related confo- cal microscopy studies assessed cell uptake and translocation of Fc-modified nanoparticles across the epithelial cell layers, whilst at the same time determining the role of FcRn in the assessed cell trafficking processes. Journal of Controlled Release xxx (2012) xxx–xxx ⁎ Corresponding author at: Division of Drug Delivery and Tissue Engineering, School of Pharmacy, University of Nottingham, Nottingham, NG7 2RD, UK. Tel.: +44 1158467046; fax: + 44 115 9515102. E-mail address: [email protected] (D. Vllasaliu). COREL-06125; No of Pages 8 0168-3659/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2011.12.009 Contents lists available at SciVerse ScienceDirect Journal of Controlled Release journal homepage: www.elsevier.com/locate/jconrel Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of nanoparticles across airway epithelial cell layers, J. Control. Release (2012), doi:10.1016/j.jconrel.2011.12.009 NANOMEDICINE
-
Upload
nottingham -
Category
Documents
-
view
0 -
download
0
Transcript of Fc-mediated transport of nanoparticles across airway epithelial cell layers

Journal of Controlled Release xxx (2012) xxx–xxx
COREL-06125; No of Pages 8
Contents lists available at SciVerse ScienceDirect
Journal of Controlled Release
j ourna l homepage: www.e lsev ie r .com/ locate / jconre l
NANOMEDICIN
E
Fc-mediated transport of nanoparticles across airway epithelial cell layers
Driton Vllasaliu a,⁎, Cameron Alexander a, Martin Garnett a, Mike Eaton b, Snow Stolnik a
a Division of Drug Delivery and Tissue Engineering, School of Pharmacy, University of Nottingham, Nottingham, NG7 2RD, UKb UCB Pharma, 208 Bath Road, Slough, Berkshire, SL1 3WE, UK
⁎ Corresponding author at: Division of Drug Deliveryof Pharmacy, University of Nottingham, Nottingham1158467046; fax: +44 115 9515102.
E-mail address: [email protected] (D
0168-3659/$ – see front matter © 2011 Elsevier B.V. Alldoi:10.1016/j.jconrel.2011.12.009
Please cite this article as: D. Vllasaliu, et a(2012), doi:10.1016/j.jconrel.2011.12.009
a b s t r a c t
a r t i c l e i n f oArticle history:Received 5 September 2011Accepted 10 December 2011Available online xxxx
Keywords:IgGFcRn pathwayMucosal deliveryNanoparticle transportSurface modification
In a study directed towards non-invasive delivery of therapeutic biomacromolecules, we examined whethersurface modification of sub-200 nmmodel nanoparticles with the Fc portion of IgG promotes their cell uptakeand transport across the airway epithelial cells. The study initially confirms the expression of the relevantreceptor, namely neonatal Fc receptor (FcRn), by Calu-3 cell layers simulating the airway epithelium anddemonstrates FcRn-mediated cell association, internalization and transcellular transport of molecular IgG.Surface decoration of nanoparticles with the Fc portion of IgG enhanced both cell uptake and translocationof the particulate system across the cell layers, in a manner strongly suggesting FcRn involvement in theseprocesses. The study further demonstrates the potential of Fc-modified nanoparticles to ‘shuttle’ a modeltherapeutic antibody fragment across the epithelial cell layers. Fc-modified nanoparticles are transportedin the μg/h/cm2 range, presenting a substantial increase in transport capacity in comparison to molecularIgG (ng/h/cm2 range), therefore warranting consideration of the FcRn transcytotic pathway for further inves-tigation as a means to achieve transmucosal delivery of nanoparticulate systems that could act as carriers of arange of biotherapeutics.
© 2011 Elsevier B.V. All rights reserved.
1. Background
Recent advances in biotechnology have produced a considerableincrease in the number of proteins that might act as biological thera-peutics. However, this trend has not been paralleled by advances innovel delivery routes for this class of therapeutics. Non-invasive ad-ministration of biotherapeutics across the mucosal surfaces wouldbe desirable, however, absorption of macromolecules through themucosae is severely limited by the physical and biochemical barriersassociated with this route. Improvement in mucosal absorption ofsome protein therapeutics is potentially achievable by inducing tran-sient opening of the epithelial tight junctions, but this strategyappears to be limited to delivery of smaller peptides and proteins[1–5], whilst delivery of larger proteins, such as antibodies (MW
approximately 150,000 g mol−1), by using tight junction openingstrategies is inefficient [6]. Transcellular transport of large therapeuticmolecules across the epithelium, rather than relying on the paracellularroute, would potentially overcome this problem. Unlike the non-specificity of the paracellular route, the transcellular pathway may bea receptor-mediated process and hence potentially selective to themolecule of interest.
and Tissue Engineering, School, NG7 2RD, UK. Tel.: +44
. Vllasaliu).
rights reserved.
l., Fc-mediated transport of n
Immunoglobulin G (IgG) is actively transferred from mother tooffspring, conferring short-term passive immunity [7,8], and hasbeen shown to traverse a number of mucosal surfaces in manwhere it is thought to play an important role in the function of IgGin immune surveillance and host defense at mucosal surfaces [9].IgG is transported by the neonatal Fc receptor (FcRn) [10,11]. Froma drug delivery perspective, the IgG/FcRn transport pathway couldbe exploited to deliver biotherapeutics as this system is functionalin several organs and tissues, including the lung [12] and intestine[13].
The primary aim of our study was to determine whether trans-port of nanoparticles across polarized epithelial cells could bepromoted by coating their surfaces with the Fc portion of IgG.Although the IgG/FcRn pathway has previously been investigatedfor mucosal delivery of protein therapeutics in molecular form, tothe best of our knowledge this system has not been considered fortransepithelial transport of nanoparticles. Our studies were con-ducted using a human bronchial epithelial cell line, Calu-3, as anin vitro model of the human airway epithelium. Initial workassessed whether this cell line expresses FcRn. Cellular uptake ofmolecular IgG (in solution) and its transport across the cell layerswas initially determined and the role of FcRn in these processesestablished. Subsequent transport experiments and related confo-cal microscopy studies assessed cell uptake and translocation ofFc-modified nanoparticles across the epithelial cell layers, whilstat the same time determining the role of FcRn in the assessed celltrafficking processes.
anoparticles across airway epithelial cell layers, J. Control. Release

2 D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
NANOMEDICIN
E
2. Methods
2.1. Cell culture
Calu-3 cells (American Type Culture Collection, USA) were seededon Transwell® filters (12 mm diameter, 0.4 μmpore size; Corning LifeSciences, Holland) at 105 cells/cm2. Cells were cultured using Eagle'sMinimum Essential Medium (EMEM, LGC standards, UK) supplemen-ted with penicillin (100 U/mL), streptomycin (0.1 mg/mL), ampho-tericin (0.25 μg/mL) and Fetal Bovine Serum (FBS, 10% v/v, Sigma-Aldrich, UK). Cells were cultured using air-interfaced culture (AIC)conditions, created on day two post-seeding, with subsequent culturemedium replacement every other day. Cell confluence was confirmedby transepithelial electrical resistance measurements (EVOM, WorldPrecision Elements, USA). Filter-cultured cell layers were used13–14 days after seeding in all experiments described below.
2.2. Expression of FcRn by Calu-3 cells
Cell layers were fixed in paraformaldehyde, washed with PBS andpermeabilized with Triton X-100 (0.1% v/v in PBS; 10 min incuba-tion). Cells were then incubated with bovine serum albumin (BSA)in PBS (1% w/v) for 1 h, followed by a goat, anti-human FcRn antibody(polyclonal, Santa Cruz, USA) incubation at 10 mg/mL in 1% w/v BSA/PBS for 30–60 min. Following extensive washing, cells were treatedwith donkey, anti-goat TRITC-IgG (Santa Cruz, USA) diluted in 1%w/v BSA/PBS (as per supplier's instructions) for 30–60 min. Cellswere then washed, filters excised and mounted on glass slides using4′,6-diamidino-2-phenylindole (DAPI)-containing mounting medium(Slowfade Gold®, Invitrogen, UK) for confocal imaging (Leica TCS SP2systemmounted on a Leica DMIRE2 invertedmicroscope). Control ex-periments were conducted where the cells were treated with either anirrelevant primary antibody (polyclonal goat, anti-human Endoglin, anendothelial cell marker) followed by a secondary donkey, anti-goatTRITC-IgG or the secondary antibody only.
2.3. Cell association and cell uptake of molecular IgG
Although bovine IgG from the FBS-supplemented culture medium isnot expected to bind to human FcRn [14], cell layers were serum-starved by incubating with Hank's Balanced Salt Solution (HBSS) for 1 h,whilst replacing HBSS twice to remove any residual bovine IgG. FITC-IgG (5 μg/mL, Sigma-Aldrich, UK) was applied to the apical side of celllayers at pH6.0 or 7.4 inHBSS bufferedwith 2-(N-Morpholino)ethanesul-fonic acid (MES) or 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid(HEPES), respectively, and incubated for 3 h. To assess the cell associationof FITC-IgG, samples (100 μl) were removed from the apical side andFITC-IgG quantified by fluorescence at 485 nm/535 nm (MFX microtiterplate fluorometer, Dynex Technologies, USA). To account for pH sensitiv-ity of FITC, where FITC-IgGwas applied at pH 6.0, the calibration curves ofFITC-IgG were constructed at pH 6.0.
For competition experiments, FITC-IgG (5 μg/mL) and unlabelledIgG (20 μg/mL, Sigma-Aldrich, UK) in HBSS (pH 6.0) were applied tothe cell layers in combination for 3 h. Apical solutions were sampled(100 μl) for fluorescence quantitation of FITC-IgG (as above).
To analyse the cell internalization of molecular IgG, followingincubation of cell layers with FITC-IgG solution (in the conditionsspecified above), apical solutions were aspirated and cells washedextensively. Cells were then permeabilized and detached with TritonX-100 (0.1% v/v), harvested and centrifuged. FITC-IgG in the superna-tants was quantified by fluorescence (as above). The calibration curveused for quantitation was in this instance constructed by progressive-ly diluting FITC-IgG in 0.1% v/v Triton X-100 in HBSS. Temperaturedependency of IgG uptake was also determined by measuring theuptake of FITC-IgG by Calu-3 cells at 4 °C and 37 °C. In this instance2.5 μg of IgG was applied apically and cell uptake measured following
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
extensive washing and cell permeabilization following a 3-hour incu-bation with FITC-IgG at either 4 °C or 37 °C.
2.4. Transport of molecular IgG across the Calu-3 layers: saturation andreceptor competition
Cells layers were serum-starved (as above) and IgG applied to theapical side at 50 ng/mL, 500 ng/mL and 5000 ng/mL (in HBSS, pH 6.0).Its apical-to-basolateral translocation was determined by measuringIgG presence in the basolateral medium (HBSS, pH 7.4) every30 min over 3 h. IgG was then quantified by ELISA using NUNC-Immuno 96 MicroWell™ (NUNC, Denmark) plates, 1% BSA w/v inPBS as the blocking buffer, horseradish peroxidase (HRP)-conjugatedanti-human IgG (The Binding Site, UK), diluted 1:1000 in blockingbuffer as the detection antibody and 3,3,5,5-tetramethylbenzidine(TMB, Autogen Bioclear Ltd, UK) as the substrate, with the reactionstopped using 2.5 M sulphuric acid (20 μl) and absorbance (at450 nm) determined using a Dynex microplate reader.
In the receptor competition experiment a solution of (human) IgG(1 μg/mL) and rabbit IgG (rIgG, 50 μg/mL) (in HBSS, pH 6.0) wereadded to the apical side of the cell layers; IgG was quantified (byELISA, in the manner described above) in the basolateral mediumperiodically (every 30 min over 3 h).
2.5. Surface modification of nanoparticles with Fc
Yellow–green nanoparticles (FluoSpheres®, 20 nm; Invitrogen,USA), yellow–orange (Fluoresbrite®, 50 nm) andnon-fluorescent nano-particles (Polybead®, 50 nm; Polysciences Europe GmbH, Germany)were used in the experiments. The nanoparticle suspensionswere dilut-ed in purified, sterile-filteredwater and incubatedwith the Fc portion ofIgG (referred to as ‘Fc’) or fluorescein isothiocyanate (FITC)-labelled Fc(‘FITC-Fc’) (Jackson Immunoresearch Europe Ltd) for 3 h at room tem-perature, with gentle stirring. 20 mg Fc, or FITC-Fc, in PBS buffer wasused per 1 mL of (undiluted) nanoparticle stock suspension. After incu-bation of nanoparticles with Fc (or FITC-Fc), the resulting suspensionwas further diluted inHBSS (MES-buffered; pH6.0) to achieve an overalldilution of 1 in 100 for all nanoparticle stock suspensions. Fc-modifiednanoparticles were applied to cells at these conditions in all cases. Forconfocal imaging of nanoparticle cell uptake Fc-modified yellow–
green nanoparticles or FITC-Fc-modified yellow–orange nanoparticleswere used, whilst particle size characterization and quantitation ofnanoparticle uptake and transport experiments were conducted usingyellow–orange (Fc-modified or unmodified) and unlabelled (Fc-adsorbed) nanoparticles.
2.6. Nanoparticle size characterization
The mean diameter and size distribution of nanoparticles (unmo-dified and Fc-coated) was determined by Dynamic Light Scattering(DLS) using a Malvern (UK) system. Fc-modified nanoparticles weresuspended in HBSS (pH 6.0) for their size analysis. The result repre-sents the mean of ten measurements at 25 °C.
2.7. Cell uptake of Fc-modified nanoparticles: imaging by confocalmicroscopy
Serum-starved cell layers were incubated with FITC-Fc-modifiedyellow–orange nanoparticles or Fc-modified yellow–green nanoparti-cles in HBSS (pH 6.0) for 2 h. Cells were then washed, fixed and pro-cessed for confocal imaging. Where zonula occludens-1 (ZO-1)staining was performed, cells were washed, permeabilized, washedagain and incubated with 1% BSA/PBS (for 1 h). BSA/PBS solutionwas replaced with mouse, anti-human ZO-1 antibody (Zymed, USA)at 10 mg/mL (in 1% BSA/PBS) for 1 h. Following washing, cells wereincubated with Cy5-labelled goat, anti-mouse antibody (Invitrogen,
anoparticles across airway epithelial cell layers, J. Control. Release

3D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
NANOMEDICIN
E
USA) at 2 mg/mL (in 1% BSA/PBS) for 1 h. Samples were then processfor imaging as described above.
Lysosomal staining. Lysotracker® Red DND-99 (Invitrogen, USA)was added to the suspension of Fc-modified nanoparticles (in HBSS,pH 6.0) at a final concentration of 100 nM. This suspension was ap-plied to confluent and serum-starved cell layers for 2 h. Followingthe incubation period, the apical solution was removed and cellswashed extensively (with PBS). Cells were then fixed with parafor-maldehyde, stained for nuclei with DAPI and processed for imagedby confocal microscopy in the manner described previously. As a con-trol, Lysotracker™ staining was also performed in cells treated withunmodified nanoparticles.
2.8. Cell uptake of Fc-modified nanoparticles: quantitative analysis
2.8.1. Cell uptake of Fc-modified nanoparticles, unmodified nanoparticlesand effect of ‘free’, soluble IgG competitor
Fc-modified, or unmodified nanoparticles (yellow–orange) wereapplied (in HBSS, pH 6.0) to serum-starved cell layers and incubatedfor 3 h. In a competition experiment, Fc-modified nanoparticles wereapplied to the cells in the presence of soluble, ‘free’ IgG (5 μg/mL,added separately 5 min prior to the application of nanoparticles).Following incubation, apical solutions were removed and cellspermeabilized, detached from the filters, harvested and centrifuged(as above). Nanoparticles were quantified in the supernatant by fluo-rescence (Excitation 529, Emission 546), using calibration curves andthe extent of cell uptake was expressed as % relative to the appliedamount.
2.8.2. Effect of unlabelled Fc-modified nanoparticles on cell uptake oflabelled Fc-modified nanoparticles (‘particulate competitor experiment’)
In a ‘particulate competitor experiment’ Fc-modified yellow–
orange nanoparticles were applied to the cell layers in combinationwith unlabelled Fc-nanoparticles (in MES-buffered HBSS, pH 6.0);the latter were used at 5-fold higher concentration. Following a 3-hour incubation, the apical solution was aspirated, cells washed, per-meabilized, detached from filters, pelleted by centrifugation and thesupernatant used for nanoparticle quantitation by fluorescence (asdescribed above).
2.8.3. Effect of temperature on cell uptake of Fc-modified nanoparticlesFc-bearing nanoparticles (yellow–orange) were applied to serum-
starved Calu-3 cell layers (in HBSS, pH 6.0). The cells were previouslyequilibrated in HBSS (pH 6.0) at either 4 °C or 37 °C. Cells wereincubated with Fc-nanoparticles at these conditions for 3 h. Apicalmedium was then removed, cells extensively washed and cellspermeabilized, detached from the Transwell® filters, harvested andcentrifuged. Nanoparticles were quantified in the supernatant byfluorescence as above.
2.9. Transport of Fc-modified nanoparticles across Calu-3 cell layers
Transport of Fc-adsorbed and unmodified (yellow–orange) nano-particles across the cell layers was determined by application ofnanoparticles on the apical side and sampling the basolateral solutionregularly for 3 h (sampled volumes were replaced with fresh HBSS).Nanoparticles were then quantified by fluorescence (against standardcalibration curves) and transport was expressed as % relative to theapplied nanoparticle amount.
In the ‘particulate competitor experiment’ Fc-modified (yellow–
orange) nanoparticles, were applied in combination with Fc-modified but unlabelled nanoparticles (used at 5-fold higher concen-tration), and analysis performed as described above.
In assessing how the co-adsorption of Fc and an antibody frag-ment on the surface of nanoparticles affects their transport behaviouracross the epithelial cell layers, equal amounts of Fc and an
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
investigational therapeutic (anti-cancer) antibody fragment (F(ab)2;MW 95815), code-named ‘CTM01’ (UCB, UK) were incubated with(yellow–orange) nanoparticles for 3 h. Fc-CTM01-modified nanopar-ticles were then applied to the apical surface of serum-starved celllayers and their apical-to-basolateral transport determined by sam-pling the basolateral solution at 30 min intervals and fluorescencemeasurement, as described above.
The effect of temperature on Fc-nanoparticle transport was alsodetermined by conducting a transport experiment (in a mannerdescribed previously, for 3 h) whilst incubating the cells with Fc-adsorbed nanoparticles at 4 °C or 37 °C.
2.10. Statistical analysis
Comparison between two groups was analysed for statistical sig-nificance using Student's t-test. Data are presented as mean±SD. Adifference of pb0.05 was considered statistically significant.
3. Results
3.1. Expression of FcRn in Calu-3 cells
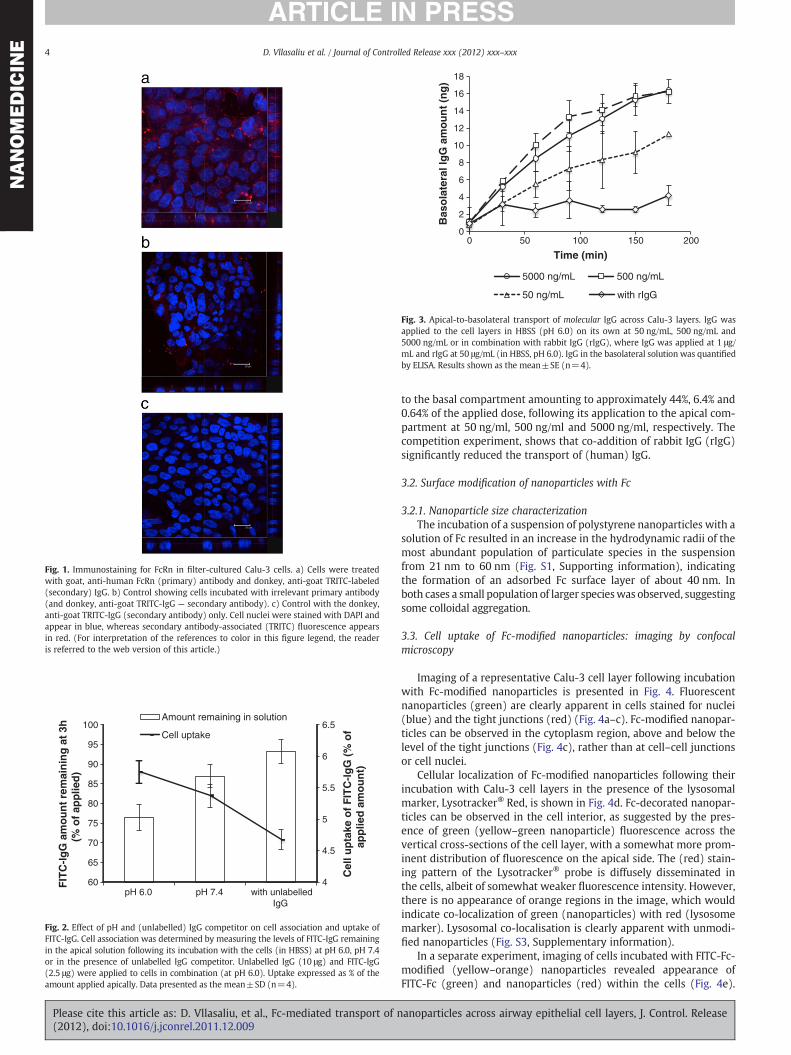
Fig. 1 shows confocal micrographs of representative air interfacegrown Calu-3 cell layers, immunostained for FcRn. Cell layers weretreated with the primary, anti-FcRn antibody, followed by a ‘staining’with TRITC-labelled (red) secondary antibody. Red (TRITC) fluores-cence, indicative of FcRn presence, can be observed in the cell cytosolregion and across the vertical axis of the cell layer (Fig. 1a), suggest-ing that the receptor is distributed within the cells and on both apicaland basolateral sides of nuclei. To ensure that the positive stainingobserved in Fig. 1a is due to the presence of FcRn two control exper-iments were conducted. Cells were subjected to treatment with an ir-relevant primary antibody, followed by the same secondary antibody(Fig. 1b) or the cells were incubated with the secondary antibodyonly (Fig. 1c). Whilst treatment with an irrelevant primary antibodyresulted in observation of some secondary antibody-associated fluo-rescence, the level of staining is notably different to cells exposed toanti-FcRn antibody (Fig. 1a). No red fluorescence was detected inthe second control experiment where the cells were treated withthe secondary (TRITC-labelled) antibody only (Fig. 1c).
Following demonstration that Calu-3 cells express FcRn, furtherexperiments were conducted to determine its functionality in cell up-take and transport of molecular IgG. Fig. 2 depicts the levels of bothcell association and uptake of molecular FITC-IgG in different condi-tions: following incubation with the cells at pH 6.0 or 7.4 and in thepresence of unlabelled IgG acting as a “competitor” (at pH 6.0). Signif-icantly lower levels of FITC-IgG remained in the apical solution at theend of the incubation interval at pH 6.0, indicating a higher extent ofcell association, compared to pH 7.4 and in the presence of unlabelledIgG competitor (p=0.04 and p=0.015, respectively). Although thecellular uptake of FITC-IgG appears to be lower at pH 7.4 comparedto pH 6.0, this difference was not statistically significant. Co-application of unlabelled IgG ‘competitor’ however significantly sup-presses cellular uptake of FITC-IgG (p=0.007).
Fig. 3 illustrates apical-to-basolateral transport of molecular IgGacross the cell layers following its application at increasing concentra-tions and in the presence of a competing ligand for FcRn (Fig. 3). Thecurve profiles representing cumulative amounts of IgG in the basolat-eral solution reveal a significant increase in transport when theapplied dose of IgG was increased from 50 to 500 ng/mL, whilst thebasolateral IgG levels measured following the apical addition at5000 ng/mL were similar to those obtained with 500 ng/mL, indicat-ing no further increase in transport with increasing apical IgG concen-tration. The data also shows that proportionally less FITC-IgG wastransported with increasing concentration of the applied material tothe apical compartment, with the proportion of FITC-IgG translocated
anoparticles across airway epithelial cell layers, J. Control. Release
0
2
4
6
8
10
12
14
16
18
0 50 100 150 200
Bas
ola
tera
l Ig
G a
mo
un
t (n
g)
Time (min)
5000 ng/mL 500 ng/mL
50 ng/mL with rIgG
Fig. 3. Apical-to-basolateral transport of molecular IgG across Calu-3 layers. IgG wasapplied to the cell layers in HBSS (pH 6.0) on its own at 50 ng/mL, 500 ng/mL and5000 ng/mL or in combination with rabbit IgG (rIgG), where IgG was applied at 1 μg/mL and rIgG at 50 μg/mL (in HBSS, pH 6.0). IgG in the basolateral solution was quantifiedby ELISA. Results shown as the mean±SE (n=4).
Fig. 1. Immunostaining for FcRn in filter-cultured Calu-3 cells. a) Cells were treatedwith goat, anti-human FcRn (primary) antibody and donkey, anti-goat TRITC-labeled(secondary) IgG. b) Control showing cells incubated with irrelevant primary antibody(and donkey, anti-goat TRITC-IgG — secondary antibody). c) Control with the donkey,anti-goat TRITC-IgG (secondary antibody) only. Cell nuclei were stained with DAPI andappear in blue, whereas secondary antibody-associated (TRITC) fluorescence appearsin red. (For interpretation of the references to color in this figure legend, the readeris referred to the web version of this article.)
4
4.5
5
5.5
6
6.5
60
65
70
75
80
85
90
95
100
pH 6.0 pH 7.4 with unlabelledIgG
Cel
l up
take
of
FIT
C-I
gG
(%
of
app
lied
am
ou
nt)
FIT
C-I
gG
am
ou
nt
rem
ain
ing
at
3h(%
of
app
lied
)
Amount remaining in solution
Cell uptake
Fig. 2. Effect of pH and (unlabelled) IgG competitor on cell association and uptake ofFITC-IgG. Cell association was determined by measuring the levels of FITC-IgG remainingin the apical solution following its incubation with the cells (in HBSS) at pH 6.0, pH 7.4or in the presence of unlabelled IgG competitor. Unlabelled IgG (10 μg) and FITC-IgG(2.5 μg) were applied to cells in combination (at pH 6.0). Uptake expressed as % of theamount applied apically. Data presented as the mean±SD (n=4).
4 D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
NANOMEDICIN
E
to the basal compartment amounting to approximately 44%, 6.4% and0.64% of the applied dose, following its application to the apical com-partment at 50 ng/ml, 500 ng/ml and 5000 ng/ml, respectively. Thecompetition experiment, shows that co-addition of rabbit IgG (rIgG)significantly reduced the transport of (human) IgG.
3.2. Surface modification of nanoparticles with Fc
3.2.1. Nanoparticle size characterizationThe incubation of a suspension of polystyrene nanoparticles with a
solution of Fc resulted in an increase in the hydrodynamic radii of themost abundant population of particulate species in the suspensionfrom 21 nm to 60 nm (Fig. S1, Supporting information), indicatingthe formation of an adsorbed Fc surface layer of about 40 nm. Inboth cases a small population of larger specieswas observed, suggestingsome colloidal aggregation.
3.3. Cell uptake of Fc-modified nanoparticles: imaging by confocalmicroscopy
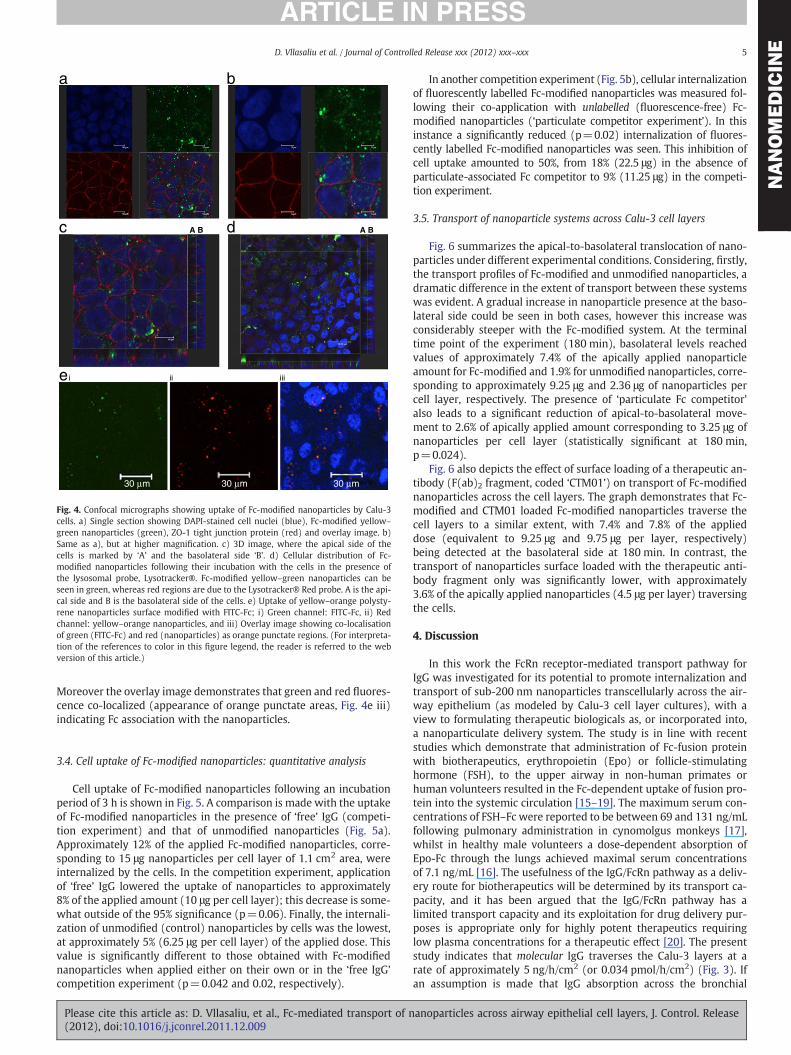
Imaging of a representative Calu-3 cell layer following incubationwith Fc-modified nanoparticles is presented in Fig. 4. Fluorescentnanoparticles (green) are clearly apparent in cells stained for nuclei(blue) and the tight junctions (red) (Fig. 4a–c). Fc-modified nanopar-ticles can be observed in the cytoplasm region, above and below thelevel of the tight junctions (Fig. 4c), rather than at cell–cell junctionsor cell nuclei.
Cellular localization of Fc-modified nanoparticles following theirincubation with Calu-3 cell layers in the presence of the lysosomalmarker, Lysotracker® Red, is shown in Fig. 4d. Fc-decorated nanopar-ticles can be observed in the cell interior, as suggested by the pres-ence of green (yellow–green nanoparticle) fluorescence across thevertical cross-sections of the cell layer, with a somewhat more prom-inent distribution of fluorescence on the apical side. The (red) stain-ing pattern of the Lysotracker® probe is diffusely disseminated inthe cells, albeit of somewhat weaker fluorescence intensity. However,there is no appearance of orange regions in the image, which wouldindicate co-localization of green (nanoparticles) with red (lysosomemarker). Lysosomal co-localisation is clearly apparent with unmodi-fied nanoparticles (Fig. S3, Supplementary information).
In a separate experiment, imaging of cells incubated with FITC-Fc-modified (yellow–orange) nanoparticles revealed appearance ofFITC-Fc (green) and nanoparticles (red) within the cells (Fig. 4e).
anoparticles across airway epithelial cell layers, J. Control. Release
a b
c A B d A B
ei ii iii
30 µm 30 µm 30 µm
Fig. 4. Confocal micrographs showing uptake of Fc-modified nanoparticles by Calu-3cells. a) Single section showing DAPI-stained cell nuclei (blue), Fc-modified yellow–
green nanoparticles (green), ZO-1 tight junction protein (red) and overlay image. b)Same as a), but at higher magnification. c) 3D image, where the apical side of thecells is marked by ‘A’ and the basolateral side ‘B’. d) Cellular distribution of Fc-modified nanoparticles following their incubation with the cells in the presence ofthe lysosomal probe, Lysotracker®. Fc-modified yellow–green nanoparticles can beseen in green, whereas red regions are due to the Lysotracker® Red probe. A is the api-cal side and B is the basolateral side of the cells. e) Uptake of yellow–orange polysty-rene nanoparticles surface modified with FITC-Fc; i) Green channel: FITC-Fc, ii) Redchannel: yellow–orange nanoparticles, and iii) Overlay image showing co-localisationof green (FITC-Fc) and red (nanoparticles) as orange punctate regions. (For interpreta-tion of the references to color in this figure legend, the reader is referred to the webversion of this article.)
5D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
NANOMEDICIN
E
Moreover the overlay image demonstrates that green and red fluores-cence co-localized (appearance of orange punctate areas, Fig. 4e iii)indicating Fc association with the nanoparticles.
3.4. Cell uptake of Fc-modified nanoparticles: quantitative analysis
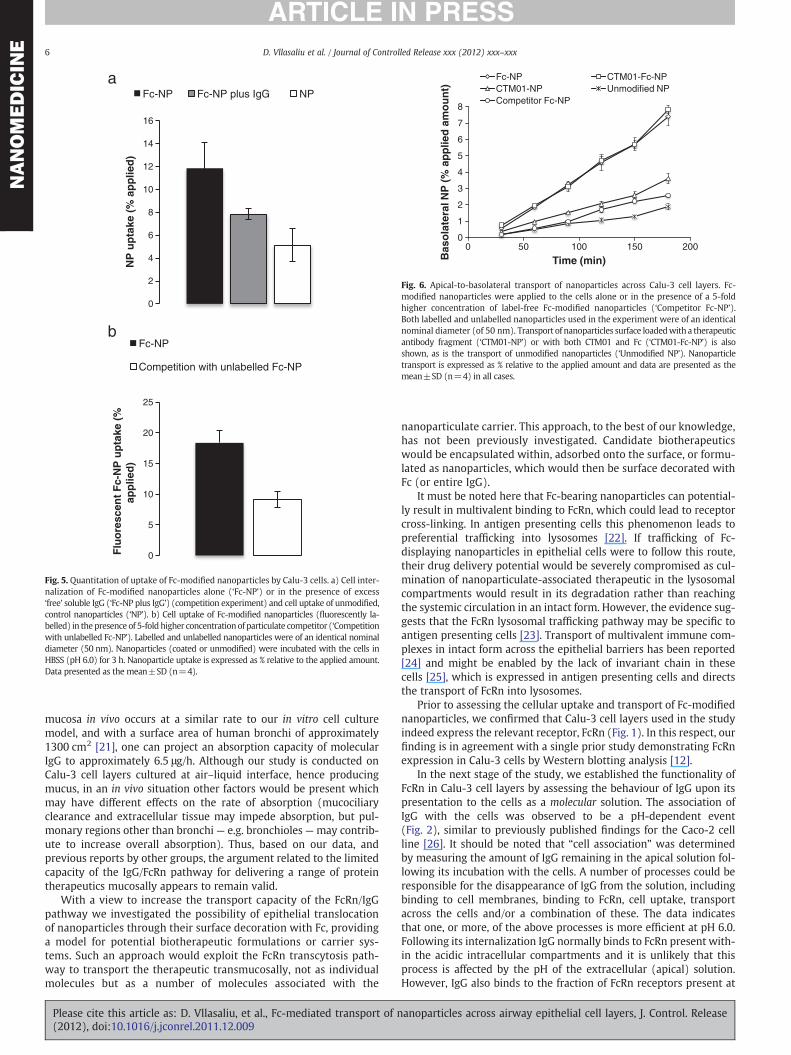
Cell uptake of Fc-modified nanoparticles following an incubationperiod of 3 h is shown in Fig. 5. A comparison is made with the uptakeof Fc-modified nanoparticles in the presence of ‘free’ IgG (competi-tion experiment) and that of unmodified nanoparticles (Fig. 5a).Approximately 12% of the applied Fc-modified nanoparticles, corre-sponding to 15 μg nanoparticles per cell layer of 1.1 cm2 area, wereinternalized by the cells. In the competition experiment, applicationof ‘free’ IgG lowered the uptake of nanoparticles to approximately8% of the applied amount (10 μg per cell layer); this decrease is some-what outside of the 95% significance (p=0.06). Finally, the internali-zation of unmodified (control) nanoparticles by cells was the lowest,at approximately 5% (6.25 μg per cell layer) of the applied dose. Thisvalue is significantly different to those obtained with Fc-modifiednanoparticles when applied either on their own or in the ‘free IgG’competition experiment (p=0.042 and 0.02, respectively).
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
In another competition experiment (Fig. 5b), cellular internalizationof fluorescently labelled Fc-modified nanoparticles was measured fol-lowing their co-application with unlabelled (fluorescence-free) Fc-modified nanoparticles (‘particulate competitor experiment’). In thisinstance a significantly reduced (p=0.02) internalization of fluores-cently labelled Fc-modified nanoparticles was seen. This inhibition ofcell uptake amounted to 50%, from 18% (22.5 μg) in the absence ofparticulate-associated Fc competitor to 9% (11.25 μg) in the competi-tion experiment.
3.5. Transport of nanoparticle systems across Calu-3 cell layers
Fig. 6 summarizes the apical-to-basolateral translocation of nano-particles under different experimental conditions. Considering, firstly,the transport profiles of Fc-modified and unmodified nanoparticles, adramatic difference in the extent of transport between these systemswas evident. A gradual increase in nanoparticle presence at the baso-lateral side could be seen in both cases, however this increase wasconsiderably steeper with the Fc-modified system. At the terminaltime point of the experiment (180 min), basolateral levels reachedvalues of approximately 7.4% of the apically applied nanoparticleamount for Fc-modified and 1.9% for unmodified nanoparticles, corre-sponding to approximately 9.25 μg and 2.36 μg of nanoparticles percell layer, respectively. The presence of ‘particulate Fc competitor’also leads to a significant reduction of apical-to-basolateral move-ment to 2.6% of apically applied amount corresponding to 3.25 μg ofnanoparticles per cell layer (statistically significant at 180 min,p=0.024).
Fig. 6 also depicts the effect of surface loading of a therapeutic an-tibody (F(ab)2 fragment, coded ‘CTM01’) on transport of Fc-modifiednanoparticles across the cell layers. The graph demonstrates that Fc-modified and CTM01 loaded Fc-modified nanoparticles traverse thecell layers to a similar extent, with 7.4% and 7.8% of the applieddose (equivalent to 9.25 μg and 9.75 μg per layer, respectively)being detected at the basolateral side at 180 min. In contrast, thetransport of nanoparticles surface loaded with the therapeutic anti-body fragment only was significantly lower, with approximately3.6% of the apically applied nanoparticles (4.5 μg per layer) traversingthe cells.
4. Discussion
In this work the FcRn receptor-mediated transport pathway forIgG was investigated for its potential to promote internalization andtransport of sub-200 nm nanoparticles transcellularly across the air-way epithelium (as modeled by Calu-3 cell layer cultures), with aview to formulating therapeutic biologicals as, or incorporated into,a nanoparticulate delivery system. The study is in line with recentstudies which demonstrate that administration of Fc-fusion proteinwith biotherapeutics, erythropoietin (Epo) or follicle-stimulatinghormone (FSH), to the upper airway in non-human primates orhuman volunteers resulted in the Fc-dependent uptake of fusion pro-tein into the systemic circulation [15–19]. The maximum serum con-centrations of FSH–Fc were reported to be between 69 and 131 ng/mLfollowing pulmonary administration in cynomolgus monkeys [17],whilst in healthy male volunteers a dose-dependent absorption ofEpo-Fc through the lungs achieved maximal serum concentrationsof 7.1 ng/mL [16]. The usefulness of the IgG/FcRn pathway as a deliv-ery route for biotherapeutics will be determined by its transport ca-pacity, and it has been argued that the IgG/FcRn pathway has alimited transport capacity and its exploitation for drug delivery pur-poses is appropriate only for highly potent therapeutics requiringlow plasma concentrations for a therapeutic effect [20]. The presentstudy indicates that molecular IgG traverses the Calu-3 layers at arate of approximately 5 ng/h/cm2 (or 0.034 pmol/h/cm2) (Fig. 3). Ifan assumption is made that IgG absorption across the bronchial
anoparticles across airway epithelial cell layers, J. Control. Release
a
b
0
2
4
6
8
10
12
14
16N
P u
pta
ke (
% a
pp
lied
)
Fc-NP Fc-NP plus IgG NP
0
5
10
15
20
25
Flu
ore
scen
t F
c-N
P u
pta
ke (
%ap
plie
d)
Fc-NP
Competition with unlabelled Fc-NP
Fig. 5. Quantitation of uptake of Fc-modified nanoparticles by Calu-3 cells. a) Cell inter-nalization of Fc-modified nanoparticles alone (‘Fc-NP’) or in the presence of excess‘free’ soluble IgG (‘Fc-NP plus IgG’) (competition experiment) and cell uptake of unmodified,control nanoparticles (‘NP’). b) Cell uptake of Fc-modified nanoparticles (fluorescently la-belled) in thepresence of 5-fold higher concentrationof particulate competitor (‘Competitionwith unlabelled Fc-NP’). Labelled and unlabelled nanoparticles were of an identical nominaldiameter (50 nm). Nanoparticles (coated or unmodified) were incubated with the cells inHBSS (pH 6.0) for 3 h. Nanoparticle uptake is expressed as % relative to the applied amount.Data presented as the mean±SD (n=4).
0
1
2
3
4
5
6
7
8
0 50 100 150 200
Bas
ola
tera
l NP
(%
ap
plie
d a
mo
un
t)
Time (min)
Fc-NP CTM01-Fc-NPCTM01-NP Unmodified NPCompetitor Fc-NP
Fig. 6. Apical-to-basolateral transport of nanoparticles across Calu-3 cell layers. Fc-modified nanoparticles were applied to the cells alone or in the presence of a 5-foldhigher concentration of label-free Fc-modified nanoparticles (‘Competitor Fc-NP’).Both labelled and unlabelled nanoparticles used in the experiment were of an identicalnominal diameter (of 50 nm). Transport of nanoparticles surface loadedwith a therapeuticantibody fragment (‘CTM01-NP’) or with both CTM01 and Fc (‘CTM01-Fc-NP’) is alsoshown, as is the transport of unmodified nanoparticles (‘Unmodified NP’). Nanoparticletransport is expressed as % relative to the applied amount and data are presented as themean±SD (n=4) in all cases.
6 D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
NANOMEDICIN
E
mucosa in vivo occurs at a similar rate to our in vitro cell culturemodel, and with a surface area of human bronchi of approximately1300 cm2 [21], one can project an absorption capacity of molecularIgG to approximately 6.5 μg/h. Although our study is conducted onCalu-3 cell layers cultured at air–liquid interface, hence producingmucus, in an in vivo situation other factors would be present whichmay have different effects on the rate of absorption (mucociliaryclearance and extracellular tissue may impede absorption, but pul-monary regions other than bronchi— e.g. bronchioles—may contrib-ute to increase overall absorption). Thus, based on our data, andprevious reports by other groups, the argument related to the limitedcapacity of the IgG/FcRn pathway for delivering a range of proteintherapeutics mucosally appears to remain valid.
With a view to increase the transport capacity of the FcRn/IgGpathway we investigated the possibility of epithelial translocationof nanoparticles through their surface decoration with Fc, providinga model for potential biotherapeutic formulations or carrier sys-tems. Such an approach would exploit the FcRn transcytosis path-way to transport the therapeutic transmucosally, not as individualmolecules but as a number of molecules associated with the
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
nanoparticulate carrier. This approach, to the best of our knowledge,has not been previously investigated. Candidate biotherapeuticswould be encapsulated within, adsorbed onto the surface, or formu-lated as nanoparticles, which would then be surface decorated withFc (or entire IgG).
It must be noted here that Fc-bearing nanoparticles can potential-ly result in multivalent binding to FcRn, which could lead to receptorcross-linking. In antigen presenting cells this phenomenon leads topreferential trafficking into lysosomes [22]. If trafficking of Fc-displaying nanoparticles in epithelial cells were to follow this route,their drug delivery potential would be severely compromised as cul-mination of nanoparticulate-associated therapeutic in the lysosomalcompartments would result in its degradation rather than reachingthe systemic circulation in an intact form. However, the evidence sug-gests that the FcRn lysosomal trafficking pathway may be specific toantigen presenting cells [23]. Transport of multivalent immune com-plexes in intact form across the epithelial barriers has been reported[24] and might be enabled by the lack of invariant chain in thesecells [25], which is expressed in antigen presenting cells and directsthe transport of FcRn into lysosomes.
Prior to assessing the cellular uptake and transport of Fc-modifiednanoparticles, we confirmed that Calu-3 cell layers used in the studyindeed express the relevant receptor, FcRn (Fig. 1). In this respect, ourfinding is in agreement with a single prior study demonstrating FcRnexpression in Calu-3 cells by Western blotting analysis [12].
In the next stage of the study, we established the functionality ofFcRn in Calu-3 cell layers by assessing the behaviour of IgG upon itspresentation to the cells as a molecular solution. The association ofIgG with the cells was observed to be a pH-dependent event(Fig. 2), similar to previously published findings for the Caco-2 cellline [26]. It should be noted that “cell association” was determinedby measuring the amount of IgG remaining in the apical solution fol-lowing its incubation with the cells. A number of processes could beresponsible for the disappearance of IgG from the solution, includingbinding to cell membranes, binding to FcRn, cell uptake, transportacross the cells and/or a combination of these. The data indicatesthat one, or more, of the above processes is more efficient at pH 6.0.Following its internalization IgG normally binds to FcRn present with-in the acidic intracellular compartments and it is unlikely that thisprocess is affected by the pH of the extracellular (apical) solution.However, IgG also binds to the fraction of FcRn receptors present at
anoparticles across airway epithelial cell layers, J. Control. Release

7D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
NANOMEDICIN
E
the plasma membrane [27] and this process is likely to be facilitatedby the acidic extracellular solution (pH 6.0) [28], which may accountfor our observation. Detection of a larger level of fluorescently la-belled IgG in the extracellular apical solution following its co-application with the competitor molecule (label-‘free’ IgG) comparedto its addition per se suggests that its disappearance from the solutionis a receptor-dependent process. Cell internalisation of IgG was foundto be temperature dependent (Supplementary information, Fig. S2).Although cell uptake of IgG does not appear statistically different atpH 6.0 and 7.4, it is significantly reduced by the addition of competi-tor, unlabelled IgG (Fig. 2), indicating a specific, FcRn-mediated inter-nalization route.
Involvement of a receptor-mediated process was also confirmedin the course of apical-to-basolateral transport of molecular IgG. Thestudy assessing the translocation of IgG following its application at in-creasing concentrations reveal process saturation (Fig. 3). To assessthat the transport of applied IgG across the Calu-3 layers is indeedmediated by FcRn, an excess of rabbit IgG (rIgG) was used as a com-petitive ligand, based on the previously documented ability of rIgGto bind to human FcRn [14]. A clear reduction in apical-to-basolateral transport of IgG (Fig. 3) in such experimental conditionscorroborates with the transport saturation data in suggesting areceptor-mediated effect and further suggests that the transport ofIgG is mediated by FcRn.
Calculating the approximate mass balance from the uptake andtransport data, it is notable that the mass balance of IgG at pH 6.0 islower than that at pH 7.4 and in the presence of competing IgG (byapproximately 5–10%). The possible reason for this is the more effi-cient binding of IgG to the fraction of FcRn present at the plasmamembrane at pH 6.0, relative to pH 7.4 and in the presence of com-peting IgG. It is likely that this membrane bound IgG (which wasnot specifically determined in the study) was removed during thewashing step with PBS (pH >7.0) and hence lost prior to determiningcell internalization of IgG.
Our model therapeutic carrier system utilises polystyrene nano-particles, surface decorated with the Fc portion of IgG. IgG was previ-ously demonstrated to non-covalently adsorb to polystyrene latexalmost irreversibly [29,30] with surface immobilized moleculesadopting random orientation [31,32], including those that potentiallypermit its binding to the receptor. In our study Fc, as the portion ofthe molecule that triggers the process of IgG translocation acrossthe epithelial surfaces [28] in vivo, was adsorbed onto polystyrenenanoparticles. This process created a layer with on approximatethickness of 40 nm, indicating multilayer formation. We postulatedthat, similarly IgG, Fc would adopt random orientations, resulting insome of the molecules being available for binding to FcRn to initiateinternalization into and ultimately transport of nanoparticles acrossthe epithelial cells.
Confocal microscopy studies revealed the presence of Fc-modifiednanoparticles within the cell interior and below the level of the tightjunctions (Fig. 4c), whereby fluorescence from (FITC-) Fc co-localizedwith that of the nanoparticles (Fig. 4d). This indicates that Fcremained associated with the nanoparticles within the cells, withoutevident separation of the targeting ligand and nanoparticles. Further-more, staining of the cells with the Lysotracker® probe to identifyactive lysosomal compartments revealed that, in Fc-nanoparticletreated cells, lysosomal compartments appear diffusely distributedthroughout the cell cytosol, rather than being clustered in the peri-nuclear regions, where the latter would suggest presence of activelysosomes [33,34]. The Fc-modified nanoparticles could also be clearlyidentified to be located separately from the Lysotracker® probe, whichis a different pattern to that observed with unmodified nanoparticles(where co-localization with Lysotracker® was clearly apparent,Supplementary information, Fig. S3), indicating that the endocytosisof Fc-modified nanoparticles generally does not terminate inside thelysosomes.
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
Experiments quantifying the extent of nanoparticle uptakehighlighted that surface modification of nanoparticles with Fc signif-icantly promoted their endocytosis (Fig. 5a), relative to unmodified(ligand-free) nanoparticles.
Further work attempted to establish whether Fc-modified nano-particles actually exploit the FcRn pathway to cross the epithelialcell layer. To this end, Fc-nanoparticle uptake was found to betemperature dependent (Supplementary information, Fig. S4a).Competition experiments using either ‘free’ soluble or nanoparticleimmobilized ligand (IgG and Fc, respectively), the latter termed‘particulate competitor’, were conducted. The data (Fig. 5) indicatesthat, although the application of soluble IgG competitor appears todecrease the uptake of Fc-modified nanoparticles, the produced effectwas not sufficiently statistically significant. Interestingly, in the‘particulate competitor’ experiment, the presentation of the compet-ing ligand in the same particle-immobilized and potentially multiva-lent manner as the tested nanoparticle system produced a markedlyreduced cellular uptake of Fc-modified nanoparticles, indicating thatthe manner in which the ligand is presented at the cell surface maybe an important parameter and that cell internalization of Fc-modified nanoparticles is a saturable and FcRn-mediated process.
In establishing the usefulness of the IgG/FcRn pathway for the pur-pose of delivering therapeutics transmucosally for a systemic effect,studying the actual transport of our Fc-nanosystem across the epithe-lial cells was essential. The transport of Fc-modified nanoparticles oc-curred to a significantly greater extent at 37 °C compared to 4 °C(Supplementary information, Fig. S4b), signifying the involvementof an energy-requiring process. Comparing the transport of Fc-modified and unmodified nanoparticles across the epithelial celllayers (Fig. 6), a significantly higher transport for Fc-modified nano-particles was evident, relative to their unmodified counterparts. Thisis an important observation and it clearly demonstrates that surfacedecoration with Fc promotes nanoparticle transport (in the apical-to-basolateral direction) across the Calu-3 cell layers. In the competi-tion experiment using nanoparticle immobilized ligand (‘particulatecompetitor’), a significantly reduced transport of Fc-modified nano-particles across the cell layers was evident, suggesting that theirtransport is indeed mediated by FcRn.
The nanoparticle transport experiments also tested the transportof nanoparticles surface loaded with a model therapeutic antibodyfragment (‘CTM01’). The surface loading of the antibody fragmentwas conducted as co-adsorption with Fc or adsorption without Fc.Such an experiment was designed to assess whether surface presenceof a relatively large biotherapeutic, other than Fc, would affect trans-cellular transport of nanoparticles, in addition to testing the potentialof Fc-modified nanoparticulate carrier system to transport biothera-peutics transepithelially. The data (Fig. 6) demonstrates similar trans-port levels of Fc-modified and Fc-CTM01-modified nanoparticles,whilst nanoparticles surface loaded with only CTM01 traversed thecell layers considerably less efficiently. The experimental set up wasnot designed to quantify the model therapeutic antibody fragment.However, our data would suggest that it is possible to use Fc-displaying nanosystems to ‘shuttle’ therapeutics across the epitheli-um provided that the protein remained associated with the nanopar-ticle carrier. This is likely to be the case considering the strong andalmost irreversible capacity of immunoglobulins to adsorb to latex[29,30].
It is important to note here that, compared to molecular IgG, Fc-modified nanoparticles exhibited a substantially increased capacityto transport biologics across the epithelial cell layers. Whilst molecu-lar IgG traversed the cell layers at a rate in the ng/h/cm2 range (ap-proximately 5.4 ng/h/cm2), the amount of the transported materialwas in the μg/h/cm2 range (approximately 2.5 μg/h/cm2) for Fc-modified nanoparticles. Interestingly, the transport of unmodifiednanoparticles is also notably larger (approximately 630 ng/h/cm2)than that of IgG in solution, although the ability of ligand-free
anoparticles across airway epithelial cell layers, J. Control. Release

8 D. Vllasaliu et al. / Journal of Controlled Release xxx (2012) xxx–xxx
NANOMEDICIN
E
nanoparticles to traverse the epithelial cell layers in the apical-to-basolateral direction has been demonstrated before [35–37]. Never-theless our study demonstrates that the capacity of the IgG/FcRntranscytotic pathway to carry therapeutics from the mucosal lumeninto the systemic circulation may be considerably enhanced by utilis-ing Fc-decorated nanocarriers, or therapeutics formulated as nano-particulates, offering noteworthy potential for future transmucosaldelivery of nanomedicines.
5. Conclusion
This work investigated whether transport of nanoparticles (whichcould be formulated from, or serve as carriers for, biomolecules)across polarized epithelial cells might be facilitated by surface modi-fication with the Fc portion of IgG. The adsorption of Fc on the surfaceof sub-200 nm nanoparticles promoted both cell uptake and trans-port of nanoparticles across the epithelial layers of airway Calu-3 cul-tures. Although detailed intracellular trafficking pathways wouldneed to be investigated to unambiguously claim that apical-to-basolateral transport of Fc-modified nanoparticles occurred by thesame transcytotic process responsible for IgG transport, our cell up-take and transport studies of Fc-bearing nanoparticles in the presenceof appropriate competitor systems strongly indicate the involvementof FcRn in cell uptake and transport behaviour of Fc-displaying nano-particles. The transport of Fc-modified nanoparticles across the epi-thelial cell layers was remarkably more efficient compared tomolecular IgG. Overall our work shows that the FcRn pathway couldbe exploited to deliver appropriately formulated nanomedicinestransmucosally, whilst at the same time improving its capacity to de-livery bioactives to potentially sensible, therapeutically adequatelevels.
Declaration of interest
This work was funded by a BBSRC Industrial CASE grant with UCBPharma.
Appendix A. Supplementary data
Supplementary data to this article can be found online at doi:10.1016/j.jconrel.2011.12.009.
References
[1] H.L. Luessen, B.J. de Leeuw,M.W. Langemeyer,A.B. de Boer, J.C.Verhoef, H.E. Junginger,Mucoadhesive polymers in peroral peptide drug delivery. VI. Carbomer and chitosanimprove the intestinal absorption of the peptide drug buserelin in vivo, Pharm. Res. 13(1996) 1668–1672.
[2] P. Sinswat, P. Tengamnuay, Enhancing effect of chitosan on nasal absorption ofsalmon calcitonin in rats: comparison with hydroxypropyl- and dimethyl-beta-cyclodextrins, Int. J. Pharm. 257 (2003) 15–22.
[3] Y.J. Zhang, C.H. Ma, W.L. Lu, X. Zhang, X.L. Wang, J.N. Sun, Q. Zhang, Permeation-enhancing effects of chitosan formulations on recombinant hirudin-2 by nasaldelivery in vitro and in vivo, Acta Pharmacol. Sin. 26 (2005) 1402–1408.
[4] M. Thanou, B.I. Florea, M.W. Langemeyer, J.C. Verhoef, H.E. Junginger, N-trimethylatedchitosan chloride (TMC) improves the intestinal permeation of the peptide drugbuserelin in vitro (Caco-2 cells) and in vivo (rats), Pharm. Res. 17 (2000) 27–31.
[5] B.I. Florea, M. Thanou, H.E. Junginger, G. Borchard, Enhancement of bronchialoctreotide absorption by chitosan and N-trimethyl chitosan shows linear invitro/in vivo correlation, J. Control. Release 110 (2006) 353–361.
[6] M. Miyamoto, H. Natsume, I. Satoh, K. Ohtake, M. Yamaguchi, D. Kobayashi, K.Sugibayashi, Y. Morimoto, Effect of poly-L-arginine on the nasal absorption ofFITC-dextran of different molecular weights and recombinant human granulocytecolony-stimulating factor (rhG-CSF) in rats, Int. J. Pharm. 226 (2001) 127–138.
[7] L.G. Morphis, D. Gitlin, Maturation of the maternofoetal transport system forhuman gamma-globulin in the mouse, Nature 228 (1970) 573.
[8] F.W. Brambell, The transmissionof immunity frommother to young and the catabolismof immunoglobulins, Lancet 2 (1966) 1087–1093.
[9] B.L. Dickinson, K. Badizadegan, Z. Wu, J.C. Ahouse, X. Zhu, N.E. Simister, R.S. Blumberg,W.I. Lencer, Bidirectional FcRn-dependent IgG transport in a polarizedhuman intestinalepithelial cell line, J. Clin. Invest. 104 (1999) 903–911.
Please cite this article as: D. Vllasaliu, et al., Fc-mediated transport of n(2012), doi:10.1016/j.jconrel.2011.12.009
[10] N.E. Simister, A.R. Rees, Isolation and characterization of an Fc receptor from neonatalrat small intestine, Eur. J. Immunol. 15 (1985) 733–738.
[11] N.E. Simister, K.E.Mostov, An Fc receptor structurally related toMHC class I antigens,Nature 337 (1989) 184–187.
[12] G.M. Spiekermann, P.W. Finn, E.S. Ward, J. Dumont, B.L. Dickinson, R.S. Blumberg,W.I. Lencer, Receptor-mediated immunoglobulin G transport acrossmucosal barriersin adult life: functional expression of FcRn in the mammalian lung, J. Exp. Med. 196(2002) 303–310.
[13] E.J. Israel, S. Taylor, Z. Wu, E. Mizoguchi, R.S. Blumberg, A. Bhan, N.E. Simister,Expression of the neonatal Fc receptor, FcRn, on human intestinal epithelialcells, Immunology 92 (1997) 69–74.
[14] R.J. Ober, C.G. Radu, V. Ghetie, E.S. Ward, Differences in promiscuity for antibody-FcRn interactions across species: implications for therapeutic antibodies, Int.Immunol. 13 (2001) 1551–1559.
[15] A.J. Bitonti, J.A. Dumont, S.C. Low, R.T. Peters, K.E. Kropp, V.J. Palombella, J.M. Stattel,Y. Lu, C.A. Tan, J.J. Song, A.M. Garcia, N.E. Simister, G.M. Spiekermann,W.I. Lencer, R.S.Blumberg, Pulmonary delivery of an erythropoietin Fc fusion protein in non-humanprimates through an immunoglobulin transport pathway, Proc. Natl. Acad. Sci. U. S.A. 101 (2004) 9763–9768.
[16] J.A. Dumont, A.J. Bitonti, D. Clark, S. Evans, M. Pickford, S.P. Newman, Delivery ofan erythropoietin-Fc fusion protein by inhalation in humans through an immu-noglobulin transport pathway, J. Aerosol Med. 18 (2005) 294–303.
[17] S.C. Low, S.L. Nunes, A.J. Bitonti, J.A. Dumont, Oral and pulmonary delivery of FSH-Fc fusion proteins via neonatal Fc receptor-mediated transcytosis, Hum. Reprod.20 (2005) 1805–1813.
[18] A.J. Bitonti, J.A. Dumont, Pulmonary administration of therapeutic proteins usingan immunoglobulin transport pathway, Adv. Drug. Deliv. Rev. 58 (2006)1106–1118.
[19] J.A. Dumont, S.C. Low, R.T. Peters, A.J. Bitonti, Monomeric Fc fusions: impact onpharmacokinetic and biological activity of protein therapeutics, BioDrugs 20(2006) 151–160.
[20] M. Sakagami, Y. Omidi, L. Campbell, L.E. Kandalaft, C.J.Morris, J. Barar,M. Gumbleton,Expression and transport functionality of FcRn within rat alveolar epithelium: astudy in primary cell culture and in the isolated perfused lung, Pharm. Res. 23(2006) 270–279.
[21] R.R. Mercer, Morphometry for alpha particle hits of critical targets in the lungs,Final Technical Report, in, USDOE Office of Energy Research, Washington, 1998.
[22] S.W. Qiao, K. Kobayashi, F.E. Johansen, L.M. Sollid, J.T. Andersen, E. Milford, D.C.Roopenian, W.I. Lencer, R.S. Blumberg, Dependence of antibody-mediated pre-sentation of antigen on FcRn, Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 9337–9342.
[23] L. Ye, X. Liu, S.N. Rout, Z. Li, Y. Yan, L. Lu, T. Kamala, N.K. Nanda, W. Song, S.K.Samal, X. Zhu, The MHC class II-associated invariant chain interacts with the neo-natal Fc gamma receptor and modulates its trafficking to endosomal/lysosomalcompartments, J. Immunol. 181 (2008) 2572–2585.
[24] M. Yoshida, S.M. Claypool, J.S. Wagner, E. Mizoguchi, A. Mizoguchi, D.C. Roopenian,W.I. Lencer, R.S. Blumberg, Human neonatal Fc receptor mediates transport of IgGinto luminal secretions for delivery of antigens to mucosal dendritic cells, Immunity20 (2004) 769–783.
[25] E.S.Ward, R.J. Ober, Multitasking by exploitation of intracellular transport functions:themany faces of FcRn, in: F.W. Alt (Ed.), Advances in Immunology, Academic Press,2009, pp. 77–117.
[26] K. Sato, J. Nagai, N. Mitsui, Y. Ryoko, M. Takano, Effects of endocytosis inhibitorson internalization of human IgG by Caco-2 human intestinal epithelial cells, LifeSci. 85 (2009) 800–807.
[27] T.A. Waldmann, E.A. Jones, The role of cell-surface receptors in the transport andcatabolism of immunoglobulins, CIBA Found. Symp. 9 (1972) 5–23.
[28] D.C. Roopenian, S. Akilesh, FcRn: the neonatal Fc receptor comes of age, Nat. Rev.Immunol. 7 (2007) 715–725.
[29] J. Serra, J. Puig, A.Martin, F. Galisteo, M. Galvez, R. Hidalgoalvarez, On the adsorptionof igg onto polystyrene particles — electrophoretic mobility and critical coagulationconcentration, Colloid Polym. Sci. 270 (1992) 574–583.
[30] F. Galisteo-Gonzalez, A. Martin-Rodriguez, R. Hidalgo-Alvarez, Adsorption ofmonoclonal IgG on polystyrene microspheres, Colloid Polym. Sci. 272 (1994)352–358.
[31] V.V. Shmanai, T.A. Nikolayeva, L.G. Vinokurova, A.A. Litoshka, Oriented antibodyimmobilization to polystyrene macrocarriers for immunoassay modified withhydrazide derivatives of poly(meth)acrylic acid, BMC Biotechnol. 1 (2001) 4.
[32] E. Imbert-Laurenceau, M.C. Berger, G. Pavon-Djavid, A. Jouan, W. Migonney, Surfacemodification of polystyrene particles for specific antibody adsorption, Polymer 46(2005) 1277–1285.
[33] A.M. Arsham, T.P. Neufeld, A genetic screen in Drosophila reveals novel cytopro-tective functions of the autophagy–lysosome pathway, PLoS One 4 (2009) e6068.
[34] P. Boya, R.A. Gonzalez-Polo, D. Poncet, K. Andreau, H.L. Vieira, T. Roumier, J.L.Perfettini, G. Kroemer, Mitochondrial membrane permeabilization is a criticalstep of lysosome-initiated apoptosis induced by hydroxychloroquine, Oncogene22 (2003) 3927–3936.
[35] O. Harush-Frenkel, E. Rozentur, S. Benita, Y. Altschuler, Surface charge of nano-particles determines their endocytic and transcytotic pathway in polarizedMDCK cells, Biomacromolecules 9 (2008) 435–443.
[36] F. Fazlollahi, S. Angelow, N.R. Yacobi, R. Marchelletta, A.S. Yu, S.F. Hamm-Alvarez,Z. Borok, K.J. Kim, E.D. Crandall, Polystyrene nanoparticle trafficking acrossMDCK-II, Nanomedicine 7 (2011) 588–594.
[37] A. des Rieux, E.G. Ragnarsson, E. Gullberg, V. Preat, Y.J. Schneider, P. Artursson,Transport of nanoparticles across an in vitro model of the human intestinal follicleassociated epithelium, Eur. J. Pharm. Sci. 25 (2005) 455–465.
anoparticles across airway epithelial cell layers, J. Control. Release