
Faunal assemblages of the mussel Mytilus galloprovincialis in and around Alsancak Harbour (Izmir...
17
Faunal assemblages of the mussel Mytilus galloprovincialis in and around Alsancak Harbour (Izmir Bay, eastern Mediterranean) with special emphasis on alien species Melih Ertan Çinar a, ⁎ , Tuncer Katağan a , Ferah Koçak b , Bilal Öztürk a , Zeki Ergen a , Ahmet Kocatas a , Mesut Önen a , Fevzi Kirkim a , Kerem Bakir a , Güley Kurt a , Ertan Dağli a , Sermin Açik a , Alper Doğan a , Tahir Özcan a a Ege University, Faculty of Fisheries, Department of Hydrobiology, 35100, Bornova, Izmir, Turkey b Dokuz Eylül University, Institute of Marine Sciences and Technology, Inciralti, Turkey Received 11 December 2006; received in revised form 10 May 2007; accepted 10 May 2007 Available online 23 May 2007 Abstract The faunal assemblages of the mussel Mytilus galloprovincialis were examined seasonally at four stations located in and around Alsancak Harbour between January and September 2004. A total of 155 species belonging to 14 systematic groups were found in the assemblages, of which Polychaeta had the highest number of species and individuals. The assemblages were mainly composed of species tolerant to pollution. The number of individuals (maximum 209,000 ind m - 2 ) and biomass (maximum 24,563 g m - 2 ) of the assemblage were generally higher at the stations located in the harbours, whereas the station not located in the harbours had the highest number of species and diversity index values (maximum 4.19). The mussel faunal assemblages differed mainly according to seasons and locations (ANOSIM test), but seasonal samples collected at each station constituted separate groups in the MDS plot, which are moderately correlated with a combination of environmental variables such as temperature, the concentrations of total nitrogen and phosphate phosphorus (ρ r = 0.46, BIOENV test). The environmental conditions at each station, the structure of substratum to which the mussel was attached and the biotic interactions among the associated faunal components seemed to be main factors influencing the faunal assemblages of the mussel in the area. The combined effect of season and location significantly changed the community parameters such as the number of species and individuals, and the values of diversity, evenness and biomass (two-way ANOVA test). The factors significantly influencing these community parameters were the algae biomass on mussels, total nitrogen and dissolved oxygen concentrations. The density of the mussel was negatively correlated with the diversity and evenness values of samples. The mussel community included ten alien species: Pseudonereis anomala, Polydora cornuta, Streblospio gynobranchiata, Hydroides dianthus, Hydroides elegans, Maera hamigera, Stenothoe gallensis, Paradella dianae, Anadara demiri and Celleporaria brunnea. These species comprised 31% of the total number of individuals found during this study. H. elegans was the most dominant species among aliens, reaching a maximum density of 110,700 ind m - 2 (spring, at station 3, 63% of the total population) in the area. The total abundance of aliens was positively correlated with that of natives, but the total biomass value of aliens was negatively correlated with that of natives. © 2007 Elsevier B.V. All rights reserved. Keywords: Zoobenthos; Pollution; Aliens; Community structure; Mytilus galloprovincialis; Izmir Bay; Aegean Sea; Turkey; Eastern Mediterranean Sea Available online at www.sciencedirect.com Journal of Marine Systems 71 (2008) 1 – 17 www.elsevier.com/locate/jmarsys ⁎ Corresponding author. E-mail address: [email protected] (M.E. Çinar). 0924-7963/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.jmarsys.2007.05.004
Transcript of Faunal assemblages of the mussel Mytilus galloprovincialis in and around Alsancak Harbour (Izmir...

Available online at www.sciencedirect.com
s 71 (2008) 1–17www.elsevier.com/locate/jmarsys
Journal of Marine System
Faunal assemblages of the mussel Mytilus galloprovincialis in andaround Alsancak Harbour (Izmir Bay, eastern Mediterranean) with
special emphasis on alien species
Melih Ertan Çinar a,⁎, Tuncer Katağan a, Ferah Koçak b, Bilal Öztürk a, Zeki Ergen a,Ahmet Kocatas a, Mesut Önen a, Fevzi Kirkim a, Kerem Bakir a, Güley Kurt a,
Ertan Dağli a, Sermin Açik a, Alper Doğan a, Tahir Özcan a
a Ege University, Faculty of Fisheries, Department of Hydrobiology, 35100, Bornova, Izmir, Turkeyb Dokuz Eylül University, Institute of Marine Sciences and Technology, Inciralti, Turkey
Received 11 December 2006; received in revised form 10 May 2007; accepted 10 May 2007Available online 23 May 2007
Abstract
The faunal assemblages of the mussel Mytilus galloprovincialis were examined seasonally at four stations located in and aroundAlsancak Harbour between January and September 2004. A total of 155 species belonging to 14 systematic groups were found inthe assemblages, of which Polychaeta had the highest number of species and individuals. The assemblages were mainly composedof species tolerant to pollution. The number of individuals (maximum 209,000 ind m−2) and biomass (maximum 24,563 g m−2) ofthe assemblage were generally higher at the stations located in the harbours, whereas the station not located in the harbours had thehighest number of species and diversity index values (maximum 4.19). The mussel faunal assemblages differed mainly accordingto seasons and locations (ANOSIM test), but seasonal samples collected at each station constituted separate groups in the MDSplot, which are moderately correlated with a combination of environmental variables such as temperature, the concentrations oftotal nitrogen and phosphate phosphorus (ρr=0.46, BIOENV test). The environmental conditions at each station, the structure ofsubstratum to which the mussel was attached and the biotic interactions among the associated faunal components seemed to bemain factors influencing the faunal assemblages of the mussel in the area. The combined effect of season and location significantlychanged the community parameters such as the number of species and individuals, and the values of diversity, evenness andbiomass (two-way ANOVA test). The factors significantly influencing these community parameters were the algae biomass onmussels, total nitrogen and dissolved oxygen concentrations. The density of the mussel was negatively correlated with the diversityand evenness values of samples. The mussel community included ten alien species: Pseudonereis anomala, Polydora cornuta,Streblospio gynobranchiata, Hydroides dianthus, Hydroides elegans, Maera hamigera, Stenothoe gallensis, Paradella dianae,Anadara demiri and Celleporaria brunnea. These species comprised 31% of the total number of individuals found during thisstudy. H. elegans was the most dominant species among aliens, reaching a maximum density of 110,700 ind m−2 (spring, at station3, 63% of the total population) in the area. The total abundance of aliens was positively correlated with that of natives, but the totalbiomass value of aliens was negatively correlated with that of natives.© 2007 Elsevier B.V. All rights reserved.
Keywords: Zoobenthos; Pollution; Aliens; Community structure;Mytilus galloprovincialis; Izmir Bay; Aegean Sea; Turkey; Eastern Mediterranean Sea
⁎ Corresponding author.E-mail address: [email protected] (M.E. Çinar).
0924-7963/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jmarsys.2007.05.004

2 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
1. Introduction
The mussel Mytilus galloprovincialis builds updense beds particularly on hard substrate in themediolittoral and upper infralittoral zones in theMediterranean (Bellan-Santini, 1969; Kocatas, 1978).Mussel beds are known to harbour a variety of animalsin interstitial spaces or on surfaces provided by musselsand algae growing on their shells (Kocatas, 1978;Tsuchiya and Bellan-Santini, 1989; Tsuchiya, 2002;Chintiroglou et al., 2004). Associated species comprisehighly mobile as well as more sedentary organisms.Mobile species roam in the complexmatrix of the musselssearching for prey or detritus retained in interstitial spaces.Sedentary species attach self-constructed tubes betweenthe mussels feeding primarily on allochthonous materialthat is continuously imported into the mussel bed (Thieland Ullrich, 2002). Beadman et al. (2004) reported thatmussels might alter the infaunal benthic community ofadjacent and interstitial sediment through provision of acomplex habitat, input of organically rich material andlarval removal through filter feeding.
Many investigations on fauna living in association withM. galloprovincialis have been carried out in the westernMediterranean (Bellan, 1969; Bellan-Santini, 1969; Bel-lan, 1980; Tsuchiya and Bellan-Santini, 1989), in theAegean Sea (Uysal, 1970; Kocatas, 1978; Lantzouni et al.,1998; Damianidis and Chintiroglou, 2000; Chintiroglou etal., 2004), in the Sea of Marmara (Topaloglu and Kihara,
Fig. 1. Map of the investigated area wi
1993) and in the Black Sea (Bacescu et al., 1965; Kiselevaand Slavina, 1966; Milovidova, 1966; Kiseleva, 1968;Băcescu, 1977; Çinar and Gönlügür-Demirci, 2005).These studies showed that a number of species areassociated with this mussel and comprised a peculiarcommunity.
As M. galloprovincialis can tolerate severe pollutionand occurs abundantly on artificial substrate such aspilings of harbours or buoys, biomonitoring of the musselbed assemblages can provide valuable information aboutthe impact of pollution on hard substrate assemblages(Bellan, 1980; Chintiroglou et al., 2004). Accumulationsof a large amount of sediments within the dense byssalthreads and shells of mussels in organically pollutedenvironments stimulate the settlement of soft bottomanimals (i.e. Capitella capitata) on mussel beds (Kocatas,1978; Çinar and Gönlügür-Demirci, 2005). In addition,biodeposit production by mussels may affect theenvironment both in the bed and the ambient waters(Tsuchiya, 2002).
The inner part of Izmir Bay, where Alsancak Harbouris located, has been subjected to various pollutiondischarges since the 1960 and is considered to be one ofthe most-polluted environments of the Mediterranean Seaover the last two decades. Due to dense pollution anddegradation, azoic conditions together with intenseodours have been previously reported from the area(Müezzinoğlu et al., 2000; Doğan et al., 2005). Recentstudies on soft bottom zoobenthic communities carried
th the location of sampling sites.

3M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
out by Çinar et al. (2006) and Ergen et al. (2006) in thearea showed that a number of species, including thosesensitive to pollution, have gradually re-colonized thearea after the establishment of “The Grand Canal Project”on February 2000, which aimed to treat waste waterspouring into the bay. However, the recent status ofzoobenthic assemblages on hard substrate in the area hasnot been investigated. However, Çinar and Ergen (2005)and Çinar (2005) found an alien species (Pseudonereisanomala) and a new species (Syllis ergeni) on the musselbed in the area, respectively.
The objectives of the present study are to investigatethe spatial and temporal patterns of zoobenthos associatedwith the mussel M. galloprovincialis after an episode ofsevere pollution and the establishment of a Waste WaterTreatment Plant in the area, to assess the environmentalfactors affecting the structure of benthic communities inthe area, and to determine the importance of alien specieswithin the communities.
2. Material and methods
2.1. Study site and sampling
Samples of M. galloprovincialis were collected sea-sonally during January 2004–September 2004 at 4 stationslocated in and around Alsancak Harbour (Fig. 1). At eachstation, three replicates were taken in 0.2 m depth byscraping off an area of 400 cm−2 using a spatula. At station1 located in AlsancakHarbour, mussel sampleswere takenfrom a piling of the harbour. At station 2 located inAlsancak Harbour, mussel samples were taken from aconcrete block dumped into the sea. At station 3 located inPasaport Harbour, mussel samples were collected on aniron platform used for approaching small boats. At station4 located near Çakalburnu Lagoon, mussel samples werecollected on a concrete block dumped into the sea.
The scraped material was put in jars containing 10%seawater–formalin solution. In the laboratory, thematerialwas sorted according to major taxonomic groups under a
Table 1Physico-chemical properties of water samples taken near stations in each sea
Winter Spring
1 2 3 4 1 2 3
Temperature (°C) 11 13 11 9 16.5 16 15.5Salinity (ppt) 35.1 36.3 37 36.3 33.9 37.4 36Oxygen (mg.l−1) 8.6 8 8.4 8.4 6.8 7.6 7.4pH 8.06 8.21 8.3 8.3 8.02 8.06 8.02Total N (μg at l−1) 7.62 11.4 5.09 3.88 3.24 4.75 2.95PO4−P (μg at l−1) 1.29 1.91 1.35 0.92 1.85 2.52 1.85Silicate (μg at l−1) 11.3 19.2 12.6 8.8 10.4 11.3 11.1
stereomicroscope and preserved in 70% ethanol. Thespecimens were then identified and counted, and total wetweight of each species was estimated using a balance of0.0001 sensitivity. The material was deposited at theMuseum of Faculty of Fisheries, Ege University (ESFM).
2.2. Laboratory and statistical analyses
Surface water samples were taken using a Nansenbottle at each station during the sampling period. Tem-perature and dissolved oxygen concentration were deter-mined in the field. Water samples for the analyses ofsalinity, pH, total nitrogen, phosphate phosphorus andsilicate were poured into bottles after pre-filtration,freezing and then transferred to the laboratory immedi-ately. Salinity, pH and nutrients were analyzed accordingto the Mohr–Knudsen method, pH meter and spectro-photometer method, respectively (Parsons et al., 1984).
Community parameters such as number of species,number of individuals, Shannon–Weaver's diversityindex (log2 base) (H′), Pielou's evenness index (J′) andthe total biomass value (wet weight) were calculated foreach station in each sampling period. The temporaltrends at each station were tested using both one-wayand two-way ANOVA. To better determine temporaldistribution patterns, the abundance data of all stationsin each sampling period were analyzed using cluster andmultidimensional scaling (MDS) techniques, based onthe Bray–Curtis similarity (group average technique),using the PRIMER package (Clarke and Warwick,2001). Prior to the cluster and multidimensional scalinganalyses, the raw data (mumber of individuals) weretransformed using transformation of yji=log(xji+1).Pairwise analysis of similarities (ANOSIM) was carriedout to test differences in the benthic assemblages in timeand space. SIMPER analysis was performed to identifythe percentage contribution of each species to the overallsimilarity (dissimilarity) within each of the groups iden-tified from the cluster analysis. Spearman rank and Pear-son correlation analyses were used to assess relationships
son
Summer Fall
4 1 2 3 4 1 2 3 4
16 26 26 26 25 23 23 23 2136.3 38.6 37.4 37.2 38.3 39.8 39.8 39.5 38.68 4.8 2 4 6 5.2 4.8 4.8 7.68.2 8 8.05 8.08 8.11 8.11 8.09 8.1 8.10.26 5.2 4.9 0.67 0.22 2.84 2.39 0.67 4.751.54 4.51 4.5 4.35 4.04 2.64 2.8 2.85 5.299.6 7.54 4.57 4.69 3.66 5.37 4.91 5.49 9.94

Table 2List of species collected during the study and their maximum density (individuals.m−2) of each species at each station [D: Dominance (%),F: Frequency (%), W: Winter, Sp: Spring, S: Summer, F: Fall] ⁎Species that represent new records for the Turkish fauna; ⁎⁎species with a dominancevalue lower than 0.10%
1 2 3 4 %D %F
PoriferaPorifera (sp.1) − − − 1/S ⁎⁎ 12.5Porifera (sp.2) − − 1 − ⁎⁎ 2.08
Hydrozoa⁎Hydrodendron mirabile (Hincks, 1866) − − − + − 2.08⁎Cordylophora caspia (Pallas, 1771) − + − − − 2.08Obelia dichotoma (Linnaeus, 1758) + + + − − 16.7⁎Bougainvillia muscus (Allman, 1863) + + + + − 31.3Ectopleura crocea (L. Agassiz 1862) + + + − − 18.8Lafoeina sp. + − − − − 2.08Ventromma halecioides (Alder, 1859) − − − + − 4.17Hydrozoa (sp.) − − + − − 2.08
ActinariaParanthus rugosus Andrès, 1880 − − 25/W − ⁎⁎ 2.08Aiptosiogeton comatus (Andres, 1881) 2400/S 200/Sp 200/Sp 100/Sp 0.17 33.3Actinaria (sp.) 25/F − − 100/S ⁎⁎ 4.17
TurbellariaTurbellaria sp.1 225/F 50/F 300/F 175/F ⁎⁎ 45.8Turbellaria sp.2 100/Sp 25/F − 75/F ⁎⁎ 18.8Turbellaria sp.3 25/F 150/F 75/F 550/F ⁎⁎ 18.8Turbellaria sp.4 150/F 200/F 2600/F 475/F 0.28 47.9Turbellaria sp.5 25/Sp − − 100/Sp ⁎⁎ 4.17
NemertiniHaplonemertea sp. − − 200/F − ⁎⁎ 4.17Heteronemertea sp. 150/Sp − 525/W 100/Sp ⁎⁎ 12.5Tubulanus cf. polymorphus Renier, 1804 150/Sp − − 300/S ⁎⁎ 10.4⁎Tubulanus cf. linearis (Mc Intosh, 1874) 50/F − − − ⁎⁎ 2.08Lineus cf. ruber (Müller, 1774) 14550/S − 775/F 7300/F 3.17 58.3Emplectonema gracile (Johnston, 1837) 375/F − 750/F 25/F 0.10 31.3⁎Prosorhochmus claparedii Keferstein, 1862 300/F − − 1450/S 0.12 18.8⁎Nemertopsis bivittata (Delle Chiaje, 1841) 225/F 25/F 425/F − ⁎⁎ 14.6
NematodaNematoda (sp.) 50/Sp − 375/F 50/Sp ⁎⁎ 20.8
OligochaetaThalassodrilides gurwitschi (Hrabe, 1971) − − − 50/Sp ⁎⁎ 4.17
PolychaetaHarmothoe sp. − − 25/S − ⁎⁎ 2.08Eumida sanguinea (Oersted, 1843) 25/W 375/F 50/Sp 25/Sp ⁎⁎ 18.8Ophiodromus pallidus (Claparède, 1864) 1300/Sp 1875/F 12525/Sp 1625/F 2.69 92Syllidia armata Quatrefages, 1865 75/S 50/Sp 275/Sp − ⁎⁎ 25Podarkeopsis galangaui Laubier, 1961 − − 100/F 25/F ⁎⁎ 10.4Myrianida benazzi Cognetti, 1953 − 25/Sp 25/Sp 50/Sp ⁎⁎ 10.4Myrianida brachycephalus (Marenzeller, 1874) − − 475/F 25/F ⁎⁎ 8.3Myrianida quindecimdentatus Langerhans, 1884 − − 475/F − ⁎⁎ 2.08⁎Myrianida neapolitanus Cognetti, 1953 25/W − 250/W 75/W ⁎⁎ 10.4Myrianida sp. − − − 25/Sp ⁎⁎ 2.08Exogone naidina Oersted, 1845 − − 25/Sp 50/Sp ⁎⁎ 12.5Salvatoria clavata (Claparède, 1863) − − − 1150/F ⁎⁎ 10.4⁎Salvatoria neapolitana (Goodrich, 1930) − − − 25/W ⁎⁎ 2.08
4 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17

Table 2 (continued )
1 2 3 4 %D %F
Syllis gracilis Grube, 1840 − − 100/S 200/F ⁎⁎ 14.6Syllis columbretensis Campoy, 1982 − − 25/W − ⁎⁎ 2.08Syllis ergeni Çinar, 2005 − 125/W 1175/F − 0.1 20.8⁎Syllis pectinans Haswell, 1920 950/S 1375/Sp 3800/F 1775/S 0.63 45.8Syllis vittata (Grube, 1840) 275/W 725/W 125/F 100/W 0.11 35.4Trypanosyllis zebra (Grube, 1840) − − − 1950/F 0.10 10.4Neanthes caudata (Delle Chiaje, 1828) 25/Sp 25/F 25/Sp − ⁎⁎ 8.33Neanthes succinea (Frey & Leuckart, 1847) 175/W 2050/Sp 175/F − 0.31 43.7Nereis falsa Quatrefages, 1865 450/F 3150/Sp 2575/Sp 125/Sp 0.89 83.3Nereis sp. − − − 50/W ⁎⁎ 2.08Ceratonereis costae (Grube, 1840) − − − 25/F ⁎⁎ 2.08Ceratonereis hircinicola (Eisig, 1870) − − − 25/W ⁎⁎ 2.08Platynereis sp. 25/W − 25/S − ⁎⁎ 4.17Platynereis dumerilii (Audouin & Milne-Edwards, 1833) − 25/F 75/W − ⁎⁎ 6.25Websterinereis glauca (Claparède, 1870) − − − 1175/F ⁎⁎ 2.08Pseudonereis anomala Gravier, 1899 175/F 125/F 225/F 100/F ⁎⁎ 39.6Nematonereis unicornis (Grube, 1840) − − − 25/Sp ⁎⁎ 2.08Schistomeringos rudolphi (Delle Chiaje, 1828) 6675/Sp 13650/Sp 18323/W 3100/F 6.5 94Ophryotrocha sp. 25/Sp 25/F 100/F − ⁎⁎ 14.6Protoaricia oerstedii (Claparède, 1864) − − − 450/S ⁎⁎ 8.3Nainereis laevigata (Grube, 1855) − − − 825/F ⁎⁎ 6.3Polydora coeca Webster, 1879 525/Sp 25/S 100/S − ⁎⁎ 18.8Polydora cornuta Bosc, 1802 75/Sp 1600/F 325/S 25/Sp 0.19 52.1Polydora sp. 25/F 125/Sp 600/S − ⁎⁎ 20.8Pseudopolydora antennata (Claparède, 1868) 50/Sp 25/F − 100/S ⁎⁎ 12.5Prionospio multibranchiata Berkeley & Berkeley, 1927 50/Sp 100/F 25/W 175/F ⁎⁎ 20.8Streblospio gynobranchiata Rice & Levin, 1998 50/S − 25/Sp − ⁎⁎ 4.17Capitella capitata (Fabricius, 1780) 25/Sp 25/Sp 175/Sp 75/F ⁎⁎ 27.1Capitella cf. capitata (Fabricius, 1780) − − 400/W 25/W ⁎⁎ 8.3Aphelochaeta sp./spp. − − − 1025/F ⁎⁎ 8.3Caulleriella cf. alata (Southern, 1914) − − − 500/F ⁎⁎ 4.2Caulleriella bioculata (Keferstein, 1862) − − − 125/Sp ⁎⁎ 2.1Caulleriella sp.1 − − − 25/W ⁎⁎ 4.2Caulleriella sp.2 − − − 200/S ⁎⁎ 10.4Cirriformia cf .tentaculata (Montagu, 1808) − 325/Sp 875/W 75/S 0.19 31.3Dodecaceria cf. sextentaculata (Delle Chiaje, 1822) − − 25/W − ⁎⁎ 2.08Timarete sp. − − − 6075/F 1.31 25Ctenodrilus serratus (Schmidt, 1857) 250/Sp 25/Sp 75/W − ⁎⁎ 16.7Polyophthalmus pictus (Dujardin, 1839) − 25/F − 25/W ⁎⁎ 6.25Ophelina modesta Støp-Bowitz, 1958 − − 25/W − ⁎⁎ 2.08Sabellaria alcocki Gravier, 1906 100/F − − 250/Sp ⁎⁎ 16.7Sabellaria alveolata (Linnaeus, 1767) − − − 125/F ⁎⁎ 6.25Polycirrus sp. − − − 25/F ⁎⁎ 2.08Terebella lapidaria Linnaeus, 1767 2050/F 2000/Sp 1200/Sp 850/F 0.78 92Bispira viola (Grube, 1863) − − 50/W − ⁎⁎ 2.08Demonax brachychona (Claparède, 1870) 25/S 75/F − − ⁎⁎ 4.17Pomatoceros lamarckii (Quatrefages, 1865) − − 25/S 75/S ⁎⁎ 4.17Hydroides dianthus (Verrill, 1873) 8675/Sp 1300/Sp 33050/Sp 1550/F 5.3 81Hydroides elegans (Haswell, 1883) 18475/F 54925/F 110700/Sp 6975/F 25.4 88Vermiliopsis infundibulum (Philippi, 1844) − − 25/W − ⁎⁎ 4.17Vermiliopsis striaticeps (Grube, 1862) − − − 575/F ⁎⁎ 4.17Simplaria pseudomilitaris (Thiriot-Quievreux, 1965) − − 675/S 1875/F 0.13 18.8Neodexiospira pseudocorrugata (Bush, 1904) − − − 1325/F 0.11 4.17
Phoronida⁎Phoronis hippocrepia Wright, 1856 25/S 25/Sp − − ⁎⁎ 4.17
(continued on next page)
5M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17

Table 2 (continued )
1 2 3 4 %D %F
BryozoaBowerbankia gracilis (Leidy, 1855) + + + + − 31.3Bowerbankia sp. + + + + − 8.33Buskia nitens Alder,1857 + − − − − 4.17Buskia socialis Hincks,1887 + + + − − 20.8Bugula neritina (Linnaeus,1758) − + + − − 20.8Bugula stolonifera Ryland,1960 + + + + − 27.1Electra monostachys (Busk, 1854) − + − − − 2.08Cryptosula pallasiana (Moll, 1803) + − + + − 16.7⁎Celleporaria brunnea (Hincks, 1884) − − − + − 8.33Schizoporella errata (Waters, 1848) − − − + − 2.08
PantopodaAmmothella appendiculata (Dohrn, 1881) 21150/F 50/F 8750/F 1100/F 1.92 50Anoplodactylus pygmaeus (Hodge, 1864) 9975/F 25/S 23200/Sp 75/S 2.18 41.7Anolodactylus sp. 125/F − − − ⁎⁎ 2.08Nymphon gracile Leach, 1814 375/F 250/F 125/F − ⁎⁎ 29.2Tanystylum conirostre (Dohrn, 1881) 600/F 250/S 9125/Sp 600/F 1.62 33.3
CrustaceaCirripediaBalanus amphitrite (Darwin, 1854) 2700/F 10250/F 2025/S 75/W 1.1 54.2Balanus eburneus Gould,1841 125/F 75/F − 150/S ⁎⁎ 16.7Balanus perforatus Bruguiére 1789 475/Sp 275/Sp 675/S 475/F 0.2 47.9Balanus improvisus Darwin, 1854 − 100/S 575/S − ⁎⁎ 12.5
AmphipodaCorophium acutum Chevreux, 1908 6825/F 3550/F 10500/F 125/F 2.32 56.3Elasmopus affinis (Della Valle, 1893) − − 575/F 5225/W 0.89 27.1Elasmopus rapax A.Costa, 1853 8075/F 2125/S 4050/S − 1.23 37.5Ericthonius brasiliensis (Dana, 1855) 3375/F 12828/S 3075/S 3350/S 1.68 39.6Hyale crassipes (Heller, 1866) 3425/S − − 3625/Sp 0.85 45.8Jassa marmorata Holmes, 1903 100/Sp − 2025/Sp 5750/Sp 0.38 25Maera hamigera Haswell, 1880 − 50/Sp − − ⁎⁎ 6.25Stenothoe monoculoides (Montagu, 1815) 25/S 75/S 25/S − ⁎⁎ 10.4Stenothoe gallensis Walker, 1904 − − 400/Sp − ⁎⁎ 4.17
IsopodaJanira maculosa Leach, 1814 − 125/Sp − − ⁎⁎ 2.08Cymodoce tuberculata Costa in Hope, 1851 50/Sp 50/S 875/F 450/S ⁎⁎ 22.9⁎Paradella dianae (Menzies, 1962) − − − 1075/W ⁎⁎ 6.25Sphaeroma serratum (Fabricius, 1787) 25/Sp 825/Sp 3075/W 150/W 0.63 47.9
TanaidaceaLeptochelia savignyi (Kroyer, 1842) − − − 600/F ⁎⁎ 12.5Tanais dulongii (Audouin, 1826) 50/F 3875/F 12450/S 4900/F 1.55 54.2
DecapodaPalaemon elegans Rathke, 1837 − 50/F − − ⁎⁎ 4.17Brachynotus sexdentatus (Risso, 1827) 100/S 625/F 275/S 25/F 0.1 41.7Pilumnus spinifer H. Milne-Edwards, 1834 50/F 50/F 275/S − ⁎⁎ 6.25Pisidia bluteli (Risso, 1816) − − − 25/S ⁎⁎ 2.08
MolluscaGastropodaFissurella nubecula (Linnaeus, 1758) − − − 1525/F ⁎⁎ 2.08Gibbula adansonii (Payraudeau, 1826) − − − 100/F ⁎⁎ 6.25Hexaplex trunculus (Linné, 1758) − − − 25/F ⁎⁎ 2.08Auriculinella bidentata (Montagu, 1808) − − 25/F − ⁎⁎ 2.08
BivalviaAnadara demiri (Piani, 1981) 25/F 25/W 50/Sp 375/F ⁎⁎ 16.7Mytilus galloprovincialis Lamarck, 1819 50550/S 23850/S 77175/F 73925/S 30.82 100Modiolus barbatus (Linnaeus, 1758) − − − 25/Sp ⁎⁎ 2.08
6 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17

Table 2 (continued )
1 2 3 4 %D %F
Modiolarca subpicta (Cantraine, 1835) − − 25/F − ⁎⁎ 4.17Musculus costulatus (Risso, 1826) − − 25/F 25/F ⁎⁎ 4.17Anomia ephippium Linnaeus, 1758 100/F 4900/F 1225/F 2875/F 0.82 37.5Ctena decussata (O.G. Costa, 1829) − − − 25/W ⁎⁎ 4.17Paphia rhomboides (Pennant, 1777) − − − 50/S ⁎⁎ 4.17Petricola lithophaga (Philippson, 1788) 25/F − 25/Sp − ⁎⁎ 4.17Sphenia binghami Turton, 1822 175/S 25/F 1400/Sp 75/S ⁎⁎ 33.3Ruditapes decussatus (Linnaeus, 1758) − − − 625/F ⁎⁎ 4.17
EchinodermataAmphipholis squamata (Delle Chiaje, 1829) − − − 4775/S 0.58 20.8
TunicataStyela canopus (Savigny, 1816) 475/F − 125/F − ⁎⁎ 18.8Ascidia mentula Müller, 1776 − 5575/F 700/F − 0.54 20.8Ascidia sp. − − 325/F 25/F ⁎⁎ 4.17⁎Molgula occidentalis Traustedt, 1882 100/Sp − 300/Sp − ⁎⁎ 10.4Botryllus schlosseri (Pallas, 1766) 50/S − 25/Sp 25/S ⁎⁎ 12.5Pyura dura Heller, 1877 150/S − − 25/S ⁎⁎ 4.17
7M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
between environmental variables and community parame-ters. Relationships betweenmultivariate community struc-ture and environmental variables were examined using theBIOENV procedure (Clarke and Warwick, 2001).
3. Results
3.1. Physico-chemical analysis
Physico-chemical features of stations in seasons areshown in Table 1. Salinity was usually high in fall (max.39.8 psu) and temperature (max. 26 °C) in summer. Inwinter, dissolved oxygen, pH, total nitrogen and silicateattained their maximum values. The highest phosphatephosphorus value (5.29 μg at l−1) was calculated atstation 4 in fall, while it was generally high at all stationsin summer.
3.2. Faunistical and ecological analysis
Faunistic analysis of the mussel samples revealed atotal of 155 benthic species belonging to Porifera (2species), Cnidaria (11 species, 174 individuals), Turbel-laria (5 species, 430 individuals), Nemertini (8 species,3486 individuals), Nematoda (1 species, 31 individuals),Oligochaeta (1 species, 3 individuals), Polychaeta (66species, 45,175 individuals), Phoronida (1 species, 2individuals), Bryozoa (10 species), Pantopoda (5 species,5716 individuals), Crustacea (23 species, 11,099 indivi-duals), Mollusca (15 species, 31491 individuals), Echi-nodermata (1 species, 569 individuals) and Tunicata (6species, 625 individuals) (Table 2). This study added 13
new records to the inventory of the marine faunainhabiting the coasts of Turkey and 5 new records(Myrianida neapolitanus, Salvatoria neapolitana, Syllispectinans, Phoronis hippocrepia and Paradella dianae)to that of the eastern Mediterranean (see Table 2).
Polychaeta accounted for 46% of the total indivi-duals, followed by Mollusca (32%) and Crustacea(11.2%). The maximum density of polychaetes wasfound at station 3 in spring (174,450 ind m−2 ), that ofCrustacea at station 3 in summer (20,650 ind m−2) andthat of Mollusca at station 3 in fall (78,625 ind m−2).
The most dominant species in the area were M.galloprovincialis (30.8% of total individuals), Hy-droides elegans (25.4%), Schistomeringos rudolphi(6.5%), H. dianthus (5.3%), Lineus cf. ruber (3.2%),Ophiodromus pallidus (2.7%), Corophium acutum(2.3%) and Anoplodactylus pygmaeus (2.2%), respec-tively, all comprising 78.4% of total specimens(Table 2). The density of M. galloprovincialis reachedits maximum (77,175 ind m−2) at station 3 in fall and itsminimum (50 ind m−2) at station 2 in fall.
The most frequent species on the mussel sampleswere M. galloprovincialis (occurred in 100% of thesamples), S. rudolphi (94%), O. pallidus (92%), Tere-bella lapidaria (92%), H. elegans (88%), Nereis falsa(83%) and H. dianthus (81%), respectively (Table 2).
Mollusca accounted for 98% (mostly M. gallopro-vincialis) of the total biomass (25,578 g) in musselsamples, followed by Crustacea (0.96%) and Polychaeta(0.44%). The maximum biomass value of Mollusca wasencountered at station 4 in fall (24,200 g m2), that ofCrustacea at station 3 in winter (1540 g m2) and that of

Table 3Results of two-way ANOVA of the community parameters
Sources ofvariation
df Number ofspecies
Number ofindividuals
Evenness index(J′)
DiversityIndex (H′)
Biomass
C=0.37ns C=0.35⁎⁎ C=0.31ns C=0.30ns C=0.31ns
MS P MS P MS P MS P MS P
Stations 3 351 ⁎ 26,828,746 ⁎ 0.006 ns 0.15 ns 327,876 ⁎
Post hoc test 2, 3 3 – – 2Seasons 3 128 ⁎ 2,544,706 ns 0.04 ⁎⁎ 1.26 ⁎ 147,718 ⁎⁎
Post hoc test Fall – Summer Fall Summer, FallStations×seasons 9 94 ⁎ 4,542,642 ⁎⁎ 0.03 ⁎ 0.93 ⁎ 136,431 ⁎
Residual 32 19 1,695,502 0.008 0.22 40,283
Post hoc test (Tukey's HSD) shows the stations and sampling periods which determine statistical differences. MS: Mean square; df: degrees offreedom; C: Cochrane's test. ⁎Pb0.01, ⁎⁎Pb0.05, ns = non significant.
Table 4Two-way crossed analysis testing for differences among seasons(averaged across sites) and among sites (averaged across seasons)
R value P
Global differencesAmong seasons 0.96 ⁎
Among stations 0.95 ⁎
Pairwise tests SeasonsFall versus winter 1 ⁎
Fall versus spring 0.97 ⁎
Fall versus summer 0.96 ⁎
Winter versus spring 0.94 ⁎
Winter versus summer 0.98 ⁎
Spring versus summer 0.87 ⁎
Stations1 versus 2 0.94 ⁎
1 versus 3 1 ⁎
1 versus 4 0.92 ⁎
2 versus 3 0.99 ⁎
2 versus 4 1 ⁎
3 versus 4 1 ⁎
⁎Pb0.01.
8 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
Polychaeta at station 3 in spring (345 g m2). Among thegroups living onM. galloprovincialis, Crustacea had thehighest biomass score (49%), followed by Polychaeta(23%) and Tunicata (14%). The species with the highestindividual biomass were M. galloprovincialis (98% oftotal biomass), Sphaeroma serratum (0.52%), Balanusamphitrite (0.24%) and Porifera sp. 1 (0.11%). Thebiomass of M. galloprovincialis reached its maximum(24,210 g m2) at station 3 in fall.
Temporal variations in number of species andindividuals, and biomass, diversity and evenness valuesat all stations are presented in Fig. 2. The number ofspecies was always high in fall (maximum 47species.400 cm−2 at station 4) and always low inwinter, and changed significantly according to seasonsat each station (Pb0.05), except for station 2. At stations1 and 4, there was a similar fluctuation in number ofindividuals, having a peak in summer, followed by a lessdecline in fall. The highest number of individuals(209,000 ind m−2) were encountered at station 3 in Fall,the lowest (7800 ind m−2) at stations 1 and 2 in winter.The number of individuals at each station changedaccording to seasons (Pb0.05), except for station 4. Theseasonality was not important on the total biomassvalues at stations 1 and 4 (PN0.05), but was important atstations 2 and 3. The biomass of theM. galloprovincialiscommunity ranged from 445 g m−2 (fall, at station 2) to24,563 g m−2 (Fall, at station 3). Temporal changes inboth evenness and diversity index values were notstatistically significant at stations 2 and 3 (PN0.05)(Fig. 2). The highest diversity and evenness values weredetermined at stations in different seasons; at stations 1(H′=3.3, J′=0.7) and 4 (H′=4.2, J′=0.76) in fall, atstation 2 (H′=2.89, J′=0.74) in spring and at station 3(H′=3.1, J′=0.65) in winter (Fig. 2).
Table 3 shows that the combined effect of season andlocation has a significant impact on all community
parameters, whereas evenness and diversity values werehomogenous between stations and number of indivi-duals between seasons. The number of species andbiomass values significantly changed according toseasons and locations. ANOSIM also revealed signifi-cant differences in the structure of assemblages in spaceand time (Table 4). Pairwise tests showed that thehighest difference were between fall and winter, andstation 4 and others (R value=1).
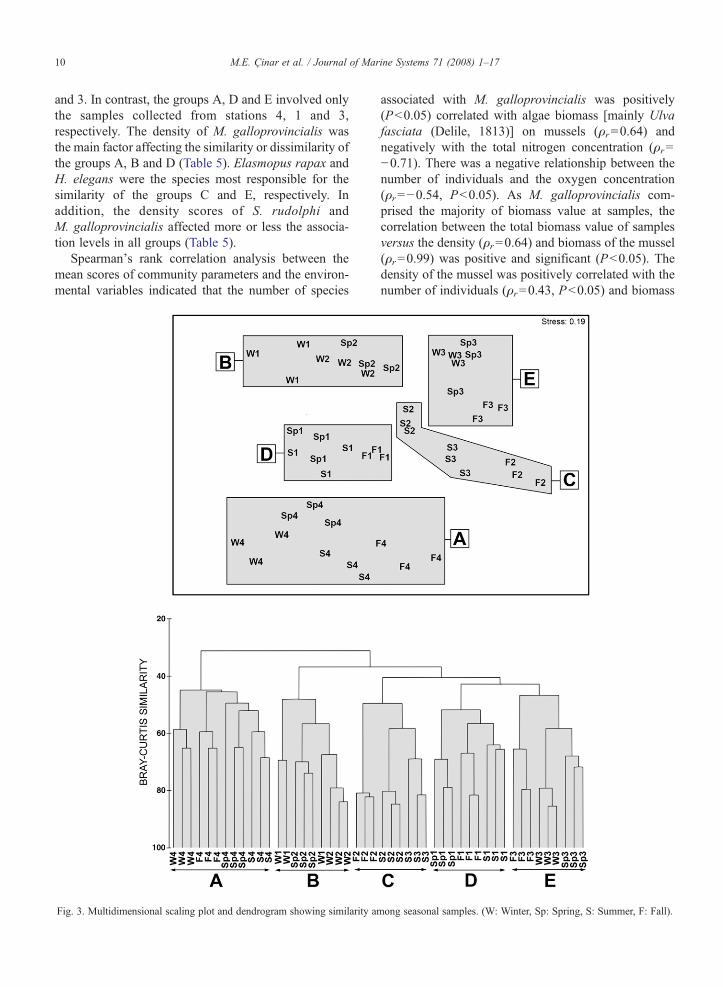
Based on Bray–Curtis similarity values higher than43%, five groups of stations (A–E) can be recognized(Fig. 3). The stress value for the two-dimensional MDSplot was 0.19, indicating a proper group separation.Samples collected from the same station tended to jointo each other, but the group B contained samples bothfrom stations 1 and 2, and the group C from stations 2

Fig. 2. Temporal fluctuations in the mean number of species, faunal densities (number of individuals per m−2), biomass (g wet weight per m−2),diversity index and evenness index at each station, with ±standard error. One-way ANOVA was used to find out if the means values of theseparameters are significant according to sampling time at each station (⁎Pb0.01, ⁎⁎Pb0.05, ns = not significant). (W: Winter, Sp: Spring, S: Summer,F: Fall).
9M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
10 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
and 3. In contrast, the groups A, D and E involved onlythe samples collected from stations 4, 1 and 3,respectively. The density of M. galloprovincialis wasthe main factor affecting the similarity or dissimilarity ofthe groups A, B and D (Table 5). Elasmopus rapax andH. elegans were the species most responsible for thesimilarity of the groups C and E, respectively. Inaddition, the density scores of S. rudolphi andM. galloprovincialis affected more or less the associa-tion levels in all groups (Table 5).
Spearman's rank correlation analysis between themean scores of community parameters and the environ-mental variables indicated that the number of species
Fig. 3. Multidimensional scaling plot and dendrogram showing similarity am
associated with M. galloprovincialis was positively(Pb0.05) correlated with algae biomass [mainly Ulvafasciata (Delile, 1813)] on mussels (ρr=0.64) andnegatively with the total nitrogen concentration (ρr=−0.71). There was a negative relationship between thenumber of individuals and the oxygen concentration(ρr=−0.54, Pb0.05). As M. galloprovincialis com-prised the majority of biomass value at samples, thecorrelation between the total biomass value of samplesversus the density (ρr=0.64) and biomass of the mussel(ρr=0.99) was positive and significant (Pb0.05). Thedensity of the mussel was positively correlated with thenumber of individuals (ρr=0.43, Pb0.05) and biomass
ong seasonal samples. (W: Winter, Sp: Spring, S: Summer, F: Fall).

Table 5Species much contributing to similarity within each assemblage of the habitats (as shown in Fig. 3), with their average similarity
Associations in Fig. 3 A B C D E
Average similarity 47 MD 58 MD 59 MD 57 MD 56 MD
Lineus cf. ruber 9 2450 – – – – 11 5275 – –Ophiodromus pallidus – – – – 6 700 6 575 11 5475Neanthes succinea – – 10 700 – – – – – –Schistomeringos rudolphii 6 975 10 3150 8 1025 8 2350 10 10,100Nereis falsa – – 11 975 – – – – 6 1000Timarete sp. 12 2700 – – – – – – – –Terebella lapidaria – – 8 675 – – 6 625 – –Hydroides dianthus – – – – – – 9 2875 9 10,800Hydroides elegans – – 8 250 6 13,775 – – 14 51,275Tanystylum conirostre – – – – – – – – 6 3925Balanus amphitrite – – – – 9 2550 – – – –Hyale crassipes 8 1050 – – – – 8 950 – –Ericthonius brasiliensis – – – – 9 3150 – – – –Elasmopus affinis 6 1700 – – – – – – – –Elasmopus rapax – – – – 10 1875 – – – –Corophium acutum – – – – 7 1475 – – – –Mytilus galloprovincialis 19 15,425 29 8025 9 13,925 14 23,275 9 18,775
Bold numbers indicate the highest score of species contribution in each assemblage.MD: Mean density (individuals m−2).
11M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
of the associated fauna (ρr=0.44), and the number ofspecies (ρr=0.40) and individuals of crustaceans(ρr=0.40). However, it was negatively correlated withthe diversity (ρr=−0.54) and evenness (ρr=−0.67)index values of samples. The mussel's biomass alsonegatively affected the diversity (ρr=−0.32) andevenness (ρr=−0.44) index values of samples, and thebiomass of polychaetes (ρr=−0.35).
The BIOENV analysis was applied in order to relatezoobenthos structural changes to environmental para-meters variability. The correlation between environ-mental and biological similarity matrix was rather low.Temperature, and the concentrations of total nitrogenand phosphate phosphorus constituted the combinationof parameters which gave the highest rank correlationvalue (ρr=0.46).
3.3. Alien species and their importance in thecommunity
The present collection included ten alien species;P. anomala, Polydora cornuta, Streblospio gynobran-chiata, Hydroides dianthus, H. elegans, Maera hami-gera, Stenothoe gallensis, P. dianae, Anadara demiri andCelleporaria brunnea. Among them, H. elegans [max.110,700 ind m−2 (spring, station 3) and 135 g m−2 (fall,station 2)] andH. dianthus [max. 33,050 ind m−2 (spring,station 3) and 82.5 g m−2 (spring, station 3)] formeddense populations in the area. The other alien specieswith the high density and biomass values in the area were
P. cornuta (max. 1600 ind m−2 and 0.4 g m−2 at station 2in fall), P. dianae (max. 1075 ind m−2 and 5.75 g m−2 atstation 4 in winter) and S. gallapagensis (max. 400 indm−2 and 0.83 g m−2 at station 3 in spring). The otherspecies were represented by low density and biomass,thoughP. anomala had a high biomass value (22.3 gm−2)at station 3 in fall. Aliens comprised the majority ofnumber of individuals at station 2 in fall (61%, 56,800 indm−2) and station 3 in spring (81%, 143,800 ind m−2).Samples taken in fall were characterized by having densepopulations of alien species. Among the stations, station 4had lower population density of alien species. Overall datashowed that aliens comprised 31% of number ofindividuals found during this study. There was a weakpositive correlation between the total density of alienspecies and that of native ones (Pearson correlationr=0.31, Pb0.05, n=48). However, a negative correlationwas found between the total biomass of alien speciesand that of native ones (Pearson correlation, r=−0.34,Pb0.05, n=48).
Spearman rank correlation analysis showed that therewas a negative correlation between the mussel densityand the densities of A. demiri (ρr=−0.57, Pb0.05), H.elegans (ρr=−0.48, Pb0.05) and H. dianthus (ρr=−0.54, Pb0.05). Only algae biomass on the mussel wassignificantly correlated with the density of P. anomala(ρr=0.44). The dense settlement of H. elegans alsoencouraged the settlement of H. dianthus (ρr=0.71). Nosignificant correlation was found between the densitiesof alien species and the environmental variables, except

12 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
for the correlation value (ρr=0.54) estimated betweenpH and the density of P. anomala.
4. Discussion
The present study shows that hard substratum in andaround Alsancak Harbour, which is located in thepolluted inner part of Izmir Bay, was densely coveredwith the mussel, M. galloprovincialis. The maximumdensity of M. galloprovincialis was found to be 77,175ind m−2, whereas Kocatas (1978) reported the maxi-mum density of the mussel as 52,850 ind m−2 inAlsancak Harbour and as 24,025 ind m−2 in the outsideof the harbour. The maximum density of the mussel wasreported to be 10,408 ind m−2 in Thermaikos Gulf(northern Aegean Sea) (Chintiroglou et al., 2004),37,300 ind m−2 on the coast of Marseille (France)(Tsuchiya and Bellan-Santini, 1989), 13,525 ind m−2 inthe Tyrrhenian Sea (Ardizzone et al., 1996) and 4250ind m−2 on the Atlantic coast of Morocco (Naciri,1997–1998). The maximum biomass (wet weight) ofM.galloprovincialis was found to be 24,200 g m2 inAlsancak Harbour. Bellan-Santini (1965) found also asimilar biomass of M. galloprovincialis (29,950 g m2)in the polluted harbour of Marseille. Hammond andGriffiths (2004) showed that M. galloprovincialisattained maximum percentage cover, average musselsize, bed depth and biomass at intermediate exposures,declining both towards the most sheltered and mostexposed sites. The present study supports this finding inthat the highest mean biomass of M. galloprovincialis(651 g m2) was found at station 4, which is not locatedin a harbour, in contrast to other stations.
A high mortality of M. galloprovincialis wasobserved at station 2 in fall; only 2–6 live specimenswere found in samples together with dead shells. Suchmortality of the mussel was previously determined fromthe area by Uysal (1978) and the reason for this massmortality was reported to be due to the low dissolvedoxygen concentration and high temperature. The cageexperiment undertaken by Uysal (1978) showed that allmussel specimens died over 26 °C temperature. Asstation 2 is located in the innermost part of Izmir Bay,where the circulation is weak, the high temperaturetogether with the low dissolved oxygen concentration insummer (see Table 1) may have resulted in the massdestruction of the mussel bed in the area. Seed (1976)summarized the reasons for the mortality of mussels assenility, physical factors (storms, salinity, excessive siltand temperature) and natural enemies [predators (e.g.crabs), competitors for food and space (e.g. barnacles),forms which attack the shell (e.g. Polydora) and
parasites (e.g. Pinnotheres)]. In this study, no boringspecies was found on M. galloprovincialis. Althoughthis study reported four polydorid species, they werespecies that constructed tubes within sediments accu-mulated among shells of the mussel. Stylochid flat-worms (Turbellaria) and decapod crustaceans are knownto be the main predators of mussels (Seed, 1976;O'Connor and Newman, 2003). We found densepopulations of five species of flatworms and the crabBrachynotus sexdentatus on the mussel beds aroundAlsancak Harbour. The mussel density in the area wasfound to be positively correlated the density offlatworms (r=0.50, Pb0.05) that may indicate thepredation of these species on the mussel.
A total of 155 zoobenthic organisms were found to beassociated with M. galloprovincialis in the area. Thefirst faunistic account on the mussel community in IzmirBay was made by Uysal (1970), who reported only 25animals on the mussel. The hard bottom community inAlsancak Harbour was previously studied by Kocatas(1978), who found 48 animals on the mussel attached towalls of quay in Alsancak Harbour. The present studycollected 79 animals at the same station. In terms ofspecies composition, there is only 42% similarity(Jaccard Similarity Index) between the present studyand the study by Kocatas (1978). Both studies indicatedthat the hard bottom communities in Alsancak Harbourwere mainly composed of species tolerant to pollutedenvironments. Although H. elegans and S. rudolphiiwere the most dominant species in both studies, thedensity of the isopod S. serratum reached up to 30,750ind m−2 on mussel samples collected from the harbourin January 1972 (Kocatas, 1978) but had a maximumdensity of 3075 ind m−2 on samples collected in January2004 (this study). Kocatas (1978) also investigatedbenthic assemblages of the mussel in Izmir Bay (outsidethe harbour) and encountered 111 animals, of whichE. rapax (17,175 ind m−2), S. serratum (10,300 ind m−2)and H. elegans (3500 ind m−2) were the most importantones. E. rapax was also reported to form densepopulations on the mussel community in ThermaikosBay (1258 ind m−2) (Chintiroglou et al., 2004) and inAlsancak Harbour (8075 ind m−2) (this study). Koçaket al. (1999) found 37 animals on fouling panelssubmerged in the polluted Levent Marina (nearAlsancak Harbour) and pointed out the high abundancesof H. elegans, Balanus eburneus, B. amphitrite,M. galloprovincialis and Ciona intestinalis. Chintir-oglou et al. (2004) postulated that the structure ofpolychaete fauna ofM. galloprovincialis assemblages inthe Mediterranean Sea was largely homogenous.Assemblages in ports are generally considered to be

13M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
assemblages of invertebrates in heavy polluted waters(Bellan-Santini, 1969; Koçak et al., 1999).
The effectiveness of the waste water treatment plantto reduce pollution in the area, which has becomeoperative in 2000, was studied and discussed by Çinaret al. (2006) and Ergen et al. (2006). Çinar et al. (2006)found a total of 232 species (including species sensitiveto pollution) on soft bottom in and around Alsancakharbour in 2004, where azoic conditions were previouslydetermined by Doğan et al. (2005) (based on the materialcollected in 1995 and 1996) and Ergen et al. (2006)(based on the material collected between 1997–2002). Inaddition to this de-faunation of the area, a decline wasencountered in the concentrations of nitrite, nitrate andammonium in surface water over time (see Çinar et al.,2006). As no datum is available concerning the effects ofsevere pollution in the area on the hard bottomcommunities after 1972, we cannot properly detect theimpact of the improved water quality on the hard bottomcommunities. However, the faunal components of M.galloprovincialis assemblages in and around AlsancakHarbour are more diversified (155 species) whencompared to those reported for other Mediterraneancoasts. The highest number of faunal species previouslyfound on this mussel was 111 in the Mediterranean Sea(Kocatas, 1978), 131 on the eastern Atlantic coast(Portugal) (Saldanha, 1974), 48 in the Bosphorus Strait(Topaloglu and Kihara, 1993) and 99 in the Black Sea(Kiseleva, 1968). This result clearly shows that there hasbeen an environmental improvement which makes thearea more favourable for the settlement of benticorganisms. The difference in the number of speciesassociated with M. galloprovincialis in different regionscould also be attributed to the taxonomic expertise of theresearchers, the total area and age structure of the musselbed sampled, height on the shore, structural complexityof the bed, etc. (Hammond and Griffiths, 2004).
Due to high individual biomass of M. galloprovin-cialis, the total biomass of species associated with thismussel comprised almost 3% of the total biomass. Thieland Ullrich (2002) also found that the associated faunausually constituted only about 2–8% of the totalbiomass within mussel beds.
Impacts of a mussel bed on the environments and thebiota associated with it can be summarized as follows(see Ragnarsson and Raffaelli, 1999); 1) the structuralcomplexity of the mussel matrix creates microhabitatswhich provide organisms with shelter and a refuge frompredation; 2) mussels deposit large amounts of faecesand pseudofaeces as well as enhancing biodeposition offine particulate matter as a result of changes in the nearbed hydrodynamic regime; and 3) dense beds of mussels
can filter out large quantities of fine particulate matter,including larvae. A recent study concerning soft bottomzoobenthos carried out in the same area (Çinar et al.,2006) gives us a comparative background about howmany species are associated with the sediment accumu-lated among mussel shells. Forty one species (mainlypolychaetes) were found both hard and soft substrates inthe area, 42 species attached to shells of the mussel andthe others (72 species) specific to the mussel beds(mobile fauna). The species attached to shells by theirtubes are filter feeders, the mobile forms are generallycarnivores or omnivores, and species associated withsediments among shells are deposit-feeders. Organicallypolluted environments of the area (inner part of IzmirBay) seem to provide a good nursery ground fordetritivors and filter feeders which in turn stimulatedensely settlements of predators such as nemerteans andpycnogonids. Nemerteans were also abundantly foundon the purple mussel beds Perumytilus purpuratus(Lamarck, 1819) along the coast of Chile (Thiel andUllrich, 2002). Both nemerteans and pycnogonids werefound at all stations in the area, but the pycnogonids(especially A. pygmaeus and Tanystylum conirostre)were abundantly encountered at station 3 where shells ofM. galloprovincialis were densely occupied by thehydroids Ectopleura crocea and Bougainvillia muscus.Pycnogonids were known to feed exclusively onhydroids (Genzano, 2002). Because of the densepredations of the pycnogonids on E. crocea andB. muscus at stations 2 and 3, the majority of specimensbelonging to these hydroids were found withouthydrotheca. The omnivores and carnivore polychaetessuch as hesionids, syllids, nereidids and dorvilleids(especially S. rudolphii) comprised up to 12% of totalnumber of individuals found during this study. Damia-nidis and Chintiroglou (2000) reported that the majorityof polychaetes on M. galloprovincialis were filterfeeders, mainly serpulids and sabellids (33.5% of totalindividuals versus 66% of total individuals of poly-chaetes in this study), and carnivores (29% versus 26%in this study). The highest number of individuals offilter-feeders found in this study was due to the highabundance of H. elegans (55% of total individuals ofpolychaetes), which was less important on the musselbeds in Thermaikos Gulf (1.23%) (Damianidis andChintiroglou, 2000). The difference in the abundance ofH. elegans between these studies is mainly due to thedifference of environmental features at the two loca-tions; polluted in Alsancak Harbour, relatively clean inThermaikos Gulf. H. elegans is known to occur abun-dantly in polluted harbours (Zibrowius, 1992; Koçaket al., 1999). Karalis et al. (2003) and Chintiroglou et al.

14 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
(2004) also reported that biotic interactions betweenorganisms played a significant role on the compositionof the hard bottom biotic assemblages in the AegeanSea.
Community structure and dynamics of animals onmussel beds or patches are considered to be influencedby a number of factors; difference in mussel species(Tsuchiya, 2002), the situation of the bed such as age orsize structures of mussels (Tsuchiya and Bellan-Santini,1989; Tsuchiya, 2002; Beadman et al., 2004), environ-mental properties and organic enrichment around thebed (Tokeshi and Romero, 2000; Saier, 2002; Adamiet al., 2004), the amount of fine particles accumulated inpatches and quality of ambient water (Tsuchiya, 2002),biotic interactions on the bed (Thiel and Ullrich, 2002;Chintiroglou et al., 2004), the type of the surface towhich the bed is attached (Adami et al., 2004; People,2006), and climatic change (Smith et al., 2006). TheMDS analysis showed that the community structure onM. galloprovincialis different mainly according to thelocations; samples from stations 1 (except for wintersamples), 3 (except for summer samples) and 4constituted different groups. The environmental condi-tions such as temperature, the concentrations of totalnitrogen and phosphate phosphorus seemed to beresponsible for this separation (BIOENV test). Howev-er, the substrate characteristics of stations appeared to bealso an important factor for the faunal difference amongstations; samples taken from concrete blocks at stations1, 2 and 4, from an iron platform at station 3. Station 4 islocated far from the source of pollution discharges in thearea and subjected to more water renewals, in contrast toother stations which are located in Alsancak andPasaport harbours. The diversity index values at station4 was generally high [range: 1.6 (summer)–4.19 (fall)]and a total of 37 species, which are sensitive topollution, such as Trypanosyllis zebra, Websterinereisglauca, Salvatoria clavata and Amphipholis squamata,were found only at this station. Çinar et al. (2006) alsoshowed that the soft bottom benthic community nearstation 4 was significantly different to those in and nearAlsancak Harbour. The iron platform at station 3attracted the dense settlements of the alien speciesH. elegans and H. dianthus. The result of post hoc testin Table 3 showed that station 3 was responsible for thedifference between stations in terms of number ofspecies and individuals. People (2006) showed that thetypes and amounts of artificial structures added to anenvironment can affect the types, distribution andabundance of organisms on mussels.
The density of the mussel was also found to becorrelated with the number of individuals (ρr=0.43) and
biomass of all associated fauna (ρr=0.44). However, thebiomass of the mussel was negatively correlated withthe faunal diversity. Chintiroglou et al. (2004) found norelationship between mussel density and the abundanceand diversity of the assemblage. However, Damianidisand Chintiroglou (2000) suggested that the faunaassociated with M. galloprovincialis appeared to berelated largely to mussel density.
The species composition and abundance on M. gallo-provincialis significantly varied among seasons and sta-tions (see Table 4). However, the diversity and evennessvalues of samples did not change significantly betweenstations and the number of individuals between seasons(Table 3). However, the combined effects of stations andseasons on community parameters were important. Adamiet al. (2004) also found that species diversity on the musselBrachidontes rodriguezii (d'Orbigny, 1846) in warm-temperate zone of Argentina showed significant temporalbut not spatial fluctuations. Hammond and Griffiths (2004)determined significant differences in faunal communitycompositions based both on infaunal abundance and bio-mass between sites. However, Saier (2002) found nosignificant differences in species number and abundancesbetween locations and between years on the mussel beds(Mytilus edulis L.) in the Wadden Sea.
The soft bottom in and around Alsancak Harbour wasreported to be densely invaded by alien species,comprising more than 70% of number of individuals ofzoobenthos in the area (Çinar et al., 2006). This study alsoshowed that the hard bottom in the area was also invadedby ten alien species, namely P. anomala, P. cornuta,S. gynobranchiata, H. dianthus, H. elegans,M. hamigera,S. gallensis, P. dianae, A. demiri and Celleporariabrunnea, accounting up to 80% of species populationsat station 3 in spring. These species, except for P. dianae,were previously reported from the Turkish coast (seeÇinar et al., 2005; Koçak, 2007). P. dianae was firstreported from the polluted Civitavecchia harbour (Tyrre-henian Sea, Italy) within the Mediterranean basin byForniz andMaggiore (1985). This species was previouslyknown from the temperate and tropical areas (seeHarrison and Holdich, 1982) and might have beenintroduced to the Mediterranean harbours by ship'shulls. Although Zenetos et al. (2005) classified thisspecies as casual, the high abundance of this species inAlsancak Harbour (maximum 1075 ind m−2) indicatedthat it has become well established in the MediterraneanSea. The alien serpulid species, H. dianthus and H.elegans, are considered as the primary organisms in thefouling event in the Mediterranean Sea (Koçak et al.,1999), which were derived from the northern Atlantic (H.dianthus) and the tropical/sub tropical region (H. elegans)

15M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
by shipping (Zibrowius, 1992). They established denselyamong shells of M. galloprovincialis in the area,suggesting that they have a potential to cause economictrouble to ship owners and others using sea-water intakepipes in the area. The presence of the alien amphipods,M.hamigera and S. gallensis, in the polluted harbours in thisstudy is interesting as these species were previouslyreported on algae in clean waters (Ruffo, 1982, 1993).
The abundance of the above-mentioned alien speciesdo not seem to interfere with the pattern of abundance ofother species (r=0.33). However, the biomass of alienspecies was negatively correlated with that of native ones(r=−34). A similar correlation (r=0.33) was foundbetween the abundance of alien species and that of nativeones inhabiting soft substratum in and near AlsancakHarbour (Çinar et al., 2006). Unlike this study, Çinaret al. (2006) also found a positive but weak correlationbetween the biomass of aliens versus natives. Rana-singhe et al. (2005) outlined a similar finding in asouthern California embayment in that abundance ofalien species was positively correlated with the abun-dance and richness of other species and that aliens did nothave a negative impact on the overall community level,possibly due to the biogenic structures built by aliens.The wealth of detritus material in the inner part of IzmirBay might hinder any possible competition betweenspecies and the still polluted conditions restrict thesettlement of pollution-sensitive species, which havegreater competitive abilities. The mass calcareousstructures built by the alien serpulid tube worms(H. elegans and H. dianthus) seem to provide a suitablemicroenvironments for sessile (mainly hydroids) andmobile (mainly pycnogonids and syllid polychaetes)species.
Mussels are known to be keystone consumers andbioturbators and their ability to filter and process largequantities of suspended material can profoundly influ-ence the dynamics of coastal and estuarine systems, withconsequent implications for local patterns of biodiversity(Seed, 1996). This study indicated that the musselM. galloprovincialis as a habitat supported a highlydiversified faunal components in this polluted part ofIzmir Bay. The biomonitoring of this community in thecourse of time would enable us predicting the dimen-sional impacts of pollution on the prevailing ecosystems.
Acknowledgments
We are much indebted to Dr. Dimitris Vafidis (Greece)for the identification of Actinarians, to Drs. Juan Junoy(Spain) and Jon L. Norenberg (USA) for the identificationof nemerteans, to Dr. Alfonso Ramos (Spain) for the
identification of Ascidian species, to Dr. Tomás Munilla(Spain) for the identification of pycnogonid species, toChristian Emig (France) for the identification ofP. hippocrepia and to personnel of R/V EGESÜF fortheir help in collecting the benthic material. This work hasbeen financially supported by Scientific Research Projectsof Ege University (Project no.: 03 SÜF 005).
References
Adami, M.L., Tablado, A., López Gappa, J., 2004. Spatial andtemporal variability in intertidal assemblages dominated by themussel Brachidontes rodriguezii (d'Orbigny, 1846). Hydrobiolo-gia 520, 49–59.
Ardizzone, G.D., Belluscio, A., Gravina, M.F., Somaschini, A., 1996.Colonization and disappearance of Mytilus galloprovincialis Lam.on an artificial habitat in the Mediterranean Sea. Estuarine, Coastaland Shelf Science 43, 665–676.
Băcescu, M., 1977. Les biocenoses benthiques de la Mer Noire.Biologie des eaux saumatres de la Mer Noire Premiere partie,pp. 128–134.
Bacescu,M.,Müller, G., Skolka, H., Petran, A., Elian, V., Gomoiu,M.T.,Bodeanu, N., Stănescu, S., 1965. Cercetări de ecologie Marină însectorul predeltaic în conditiile Anilor 1960–1961. EcologieMarină.Editura Academiei Republicii Populare Romăne, Bucureşti, pp.185–344.
Beadman, H.A., Kaiser, M.J., Galanidi, M., Shucksmith, R., Willows,R.I., 2004. Changes in species richness with stocking density ofmarine bivalves. Journal of Applied Ecology 41, 464–475.
Bellan, G., 1969. Polychètes des horizons supérieurs de l'étageinfralittoral rocheux dans la région Provençale. Tethys 1, 349–366.
Bellan, G., 1980. Relationship of pollution to rocky substratumpolychaetes on the French Mediterranean coast. Marine PollutionBulletin 11, 318–321.
Bellan-Santini, D., 1965. Étude quantitative du peuplement a Mytilusgalloprovincialis Lamarck en eau moyennement polluée. Rapportet Procès-verbaux des Réunion Commission Internationale pourl'exploration scientifique de la Mer Méditerranée 18, 85–89.
Bellan-Santini, D., 1969. Contribution à l'étude des peuplementsinfralittoraux sur substrat rocheux (Etude qualitative et quantitativede la frange supérieure). Recueil des Travaux de la Station Marined'Endoume 47, 5–294.
Chintiroglou, C.C., Damianidis, P., Antoniadou, C., Lantzouni, M.,Vafidis, D., 2004. Macrofauna biodiversity of mussel bedassemblages in Thermaikos Gulf (northern Aegean Sea). Helgo-land Marine Research 58, 62–70.
Çinar, M.E., 2005. Syllis ergeni: a new species of Syllidae (Annelida:Polychaeta) from Izmir Bay (Aegean Sea, eastern MediterraneanSea). Zootaxa 1036, 43–54.
Çinar, M.E., Ergen, Z., 2005. Lessepsian migrants expanding theirdistributional ranges; Pseudonereis anomala (Polychaeta: Nerei-didae) in Izmir Bay (Aegean Sea). Journal of the Marine BiologicalAssociations of the United Kingdom 85, 313–321.
Çinar, M.E., Gönlügür-Demirci, G., 2005. Polychaete assemblages onshallow-water benthic habitats along the Sinop Peninsula (BlackSea, Turkey). Cahiers de Biologie Marine 46, 253–263.
Çinar, M.E., Bilecenoglu, M., Öztürk, B., Katagan, T., Aysel, V., 2005.Alien species on the coasts of Turkey. Mediterranean MarineScience 6, 119–146.
Çinar, M.E., Katagan, T., Öztürk, B., Egemen, Ö., Ergen, Z., Kocatas,A., Önen, M., Kirkim, F., Bakir, K., Kurt, G., Dagli, E., Kaymakçi,

16 M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
A., Açik, S., Dogan, A., Özcan, T., 2006. Temporal changes ofsoft-bottom zoobenthic communities in anda round AlsancakHarbour (Izmir Bay, Aegean Sea), with special attention to theautecology of exotic species. Marine Ecology 27, 229–246.
Clarke, K.R., Warwick, R.M., 2001. Change in Marine Communities:An Approach to Statistical Analysis and Interpretation, 2ndedition. PRIMER-E, Plymouth.
Damianidis, P., Chintiroglou, C.-C., 2000. Structure and functions ofPolychaetofauna living in Mytilus galloprovincialis assemblagesin Thermaikos Gulf (north Aegean Sea). Oceanologica Acta 23,323–337.
Doğan, A., Çinar, M.E., Önen, M., Ergen, Z., Katağan, T., 2005.Seasonal analysis of soft bottom zoobenthic communities inpolluted and unpolluted areas of Izmir Bay (Aegean Sea).Senckenbergiana Maritima 35, 133–145.
Ergen, Z., Çinar, M.E., Dagli, E., Kurt, G., 2006. Seasonal dynamics ofsoft-bottom polychaetes in Izmir Bay (Aegean Sea, easternMediterranean). Scientia Marina 70, 197–205 (Suppl.).
Forniz, C., Maggiore, F., 1985. New records of Sphaeromatidae fromthe Mediterranean Sea (Crustacea, Isopoda). Oebalia 11-3,779–783.
Genzano, G.N., 2002. Associations between pycnogonids andhydroids from the Buenos Aires littoral zone, with observationson the semi-parasitic life cycle of Tanystylum orbiculare(Ammotheiidae). Scientia Marina 66, 83–92.
Hammond, W., Griffiths, C.L., 2004. Influence of wave exposure onSouth African mussel beds and their associated infaunalcommunities. Marine Biology 144, 547–552.
Harrison, K., Holdich, D.M., 1982. Revision of the genera Dyname-nella, Ischyromene, Dynamenopsis and Cymodocella (Crustacea:Isopoda), including a new genus and five new species ofEubranchiate Sphaeromatids from Queensland waters. Journal ofCrustacean Biology 2, 84–119.
Karalis, P., Antoniadou, C., Chintiroglou, C., 2003. Structure of theartificial hard substrate assemblages in ports in Thermaikos Gulf(North Aegean Sea). Oceanologica Acta 26, 215–224.
Kiseleva, M.I., 1968. Comparative characteristics of the biocenoses ofMytilus galloprovincialis on the Crimia and Caucas coasts. RevueRoumaine de Biologie Série de Zoologie 13, 403–409.
Kiseleva, M.I., Slavina, O.Y., 1966. Quantitative distribution ofmacrobenthos along the coast of Caucas. Naukovaya Dumka Kiev55–74.
Koçak, F., 2007. A new alien bryozoan Celleporaria brunnea (Hincks,1884) in the Aegean Sea (eastern Mediterranean). Scientia Marina71, 191–195.
Koçak, F., Ergen, Z., Çinar, M.E., 1999. Fouling organisms and theirdevelopments in a polluted and an unpolluted marina in theAegean Sea (Turkey). Ophelia 50, 1–20.
Kocatas, A., 1978. Contribution à l'étude des peuplements deshorizons supérieurs de substrat rocheux du golfe d'Izmir (Turquie).Scientific monographs of the Faculty of Science Ege University12, 1–93 (in Turkish).
Lantzouni, M., Voultsiadou, E., Chintiroglou, C., 1998. Preliminaryobservations on Amphipod assemblages associated with Mytilusgalloprovincialis Lamarck Beds from Thermaikos Gulf (AegeanSea). Rapport de la Commission Internationale pour l'Explorationscientifique de la Mer Méditerranée 35, 458–459.
Milovidova, H.O., 1966. Deep biocenoses in the Novorosisk Gulf.Naukovaya Dumka Kiev 75–89.
Müezzinoğlu, A., Sponza, D., Köken, I., Alparslan, N., Akyarli, A.,Öztüre, N., 2000. Hydrogen sulfide and odour control in IzmirBay. Water, Air and Soil Pollution 123, 245–257.
Naciri, M., 1997–1998. Dynamique d'une population de moules, Myti-lus galloprovincialis (Lmk.), vivant sur la côte atlantique marocaine.Bulletin de l'Institut Scientifique Rabat, vol. 21, pp. 43–50.
O'Connor, W.A., Newman, L.J., 2003. Predation of cultured mussels,Mytilus galloprovincialis, by stylochid flatworms, Imoginemcgrafti, from Twofold Bay, New South Wales, Australia. AsianFisheries Science 16, 133–140.
Parsons, T.R., Matia, Y., Lalli, C.M., 1984. A Manual of Chemical andBiological Methods for Seawater Analysis. Pergamon Press, NewYork. 173 pp.
People, J., 2006. Mussel beds on different types of structures supportdifferent macroinvertebrate assemblages. Austral Ecology 31,271–281.
Ragnarsson, S.A., Raffaelli, D., 1999. Effects of the mussel Mytilusedulis L. on the invertebrate fauna of sediments. Journal ofExperimental Marine Biology and Ecology 241, 31–43.
Ranasinghe, J.A., Mikel, T.K., Velarde, R.G., Weisberg, S.B.,Montagne, D.E., Cadien, D.B., Dalkey, A., 2005. The prevalenceof non-indigenous species in southern California embayments andtheir effects on benthic macroinvertebrate communities. BiologicalInvasions 7, 679–686.
Ruffo S. (ed.), 1982. The Amphipoda of the Mediterranean. Part 1.Gammaridea (Acanthonotozomatidae to Gammaridae), by Bellan-Santini D,KaramanG., Krapp-SchickelG., LedoyerM.,MyersA.A.,Ruffo S., Schieke U. Mémoires de l'Institut océanograohique,Monaco, 13: i–xiv + 1 map + 1–364.
Ruffo S. (ed.), 1993. The Amphipoda of the Mediterranean. Part 3.Gammaridea (Melphidippidae to Talitridae), Ingolfiellidea,Caprellidea, by Bellan-Santini D., Karaman G., Krapp-SchickelG., Ledoyer M., Ruffo S. Mémoires de l'Institut océanographique,Monaco, 13: 365–576. Mémoires de l'Institut océanographique,Monaco, 13: xxi–xxv + 577–814.
Saldanha, L., 1974. Estudo do povoamento dos horizontes superioresda rocha litoral da costa da Arrábida (Portugal). Publicaçăo doMuseu e Laboratório Zoológico e Antropólógico Faculdade deCiências de Lisboa 5, 1–382.
Saier, B., 2002. Subtidal and intertidal mussel beds (Mytilus edulis L.)in the Wadden Sea: diversity differences of associated epifauna.Helgoland Marine Research 56, 44–50.
Seed, R., 1976. Ecology. In: Bayne, B.L. (Ed.), Marine Mussels: TheirEcology and Physiology. Cambridge University Press, pp. 13–65.
Seed, R., 1996. Patterns of biodiversity in the macro-invertebrate faunaassociated with mussel patches on rocky shores. Journal of theMarine Biological Associations of the United Kingdom 76,203–210.
Smith, J.R., Fong, P., Ambrose, R.F., 2006. Dramatic declines inmussel bed community diversity: response to climatic change?Ecology 87, 1153–1161.
Thiel, M., Ullrich, N., 2002. Hard rock versus soft bottom: the faunaassociated with intertidal mussel beds on hard bottoms along thecoast of Chile, and considerations on the functional role of messelbeds. Helgoland Marine Research 56, 21–30.
Tokeshi, M., Romero, L., 2000. Spatial overlap and coexistence in amussel-associated polychaete assemblage on a South Americanrocky shore. Pubblicazioni della Stazione Zoologica di Napoli.Marine Ecology 21, 247–261.
Topaloglu, B., Kihara, K., 1993. Community of Mediterranean musselMytilus galloprovincialis Lamarck, 1819 in the Bosphorus Strait.Journal of Tokyo University of Fisheries 80, 113–120.
Tsuchiya, M., 2002. Faunal structures associated with patches ofmussels on East Asian coasts. Helgoland Marine Research 56,31–36.

17M.E. Çinar et al. / Journal of Marine Systems 71 (2008) 1–17
Tsuchiya, M., Bellan-Santini, D., 1989. Vertical distribution of shallowrocky shore organisms and community structure of mussel beds(Mytilus galloprovincialis) along the coast of Marseille, France.Mésogée 49, 91–110.
Uysal, H., 1970. Biological and ecological investigations on themussels (Mytilus galloprovincialis Lamarck) living in the coastlineof Turkey. Scientific Reports of the Faculty of Science EgeUniversity 79, 1–77 (in Turkish).
Uysal, H., 1978. The effects of some pollutants on Mytilusgalloprovincialis Lam. and Paracentrotus lividus Lam. in the
Bays of Izmir and Aliaga. IVes Journées Étude Pollutions Antalya313–317.
Zenetos, A., Çinar,M.E., Pancucci-Papadopoulou,M.A., Harmelin, J.G.,Furnari, G., Andaloro, F., Bellou, N., Streftaris, N., Zibrowius, H.,2005. Annotated list of marine alien species in the Mediterraneanwith records of the worst invasive species. Mediterranean MarineScience 6, 63–118.
Zibrowius, H., 1992. Ongoing modification of the Mediterraneanmarine fauna and flora by the establishment of exotic species.Mésogée 51, 83–107.




















