Extracellular Matrix-mediated Tissue Remodeling Following Axial Movement of Teeth
15
http://jhc.sagepub.com/ Journal of Histochemistry & Cytochemistry http://jhc.sagepub.com/content/55/2/127 The online version of this article can be found at: DOI: 10.1369/jhc.6A7018.2006 2007 55: 127 J Histochem Cytochem Yamane, Ellen Begole, Carla Evans and Thomas G.H. Diekwisch Xianghong Luan, Yoshihiro Ito, Sean Holliday, Cameron Walker, Jon Daniel, Therese M. Galang, Tadayoshi Fukui, Akira Extracellular Matrix-mediated Tissue Remodeling Following Axial Movement of Teeth Published by: http://www.sagepublications.com On behalf of: Official Journal of The Histochemical Society can be found at: Journal of Histochemistry & Cytochemistry Additional services and information for http://jhc.sagepub.com/cgi/alerts Email Alerts: http://jhc.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: What is This? - Feb 1, 2007 Version of Record >> by guest on December 10, 2013 jhc.sagepub.com Downloaded from by guest on December 10, 2013 jhc.sagepub.com Downloaded from
Transcript of Extracellular Matrix-mediated Tissue Remodeling Following Axial Movement of Teeth

http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/55/2/127The online version of this article can be found at:
DOI: 10.1369/jhc.6A7018.2006
2007 55: 127J Histochem CytochemYamane, Ellen Begole, Carla Evans and Thomas G.H. Diekwisch
Xianghong Luan, Yoshihiro Ito, Sean Holliday, Cameron Walker, Jon Daniel, Therese M. Galang, Tadayoshi Fukui, AkiraExtracellular Matrix-mediated Tissue Remodeling Following Axial Movement of Teeth
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Feb 1, 2007Version of Record >>
by guest on December 10, 2013jhc.sagepub.comDownloaded from by guest on December 10, 2013jhc.sagepub.comDownloaded from

ARTICLE
Extracellular Matrix-mediated Tissue Remodeling FollowingAxial Movement of Teeth
Xianghong Luan, Yoshihiro Ito, Sean Holliday, Cameron Walker, Jon Daniel, Therese M. Galang,Tadayoshi Fukui, Akira Yamane, Ellen Begole, Carla Evans, and Thomas G.H. Diekwisch
Department of Oral Biology (XL,YI,SH,CW,JD,TMG,TGHD) and Department of Orthodontics (XL,YI,SH,TMG,EB,CE,TGHD),University of Illinois at Chicago College of Dentistry, Chicago, Illinois, and Department of Pharmacology, Tsurumi UniversitySchool of Dental Medicine, Yokohama, Japan (TF,AY)
SUMMARY Tooth eruption is a multifactorial process involving movement of existing tis-sues and formation of new tissues coordinated by a complex set of genetic events. We haveused the model of the unopposed rodent molar to study morphological and genetic mecha-nisms involved in axial movement of teeth. Following extraction of opposing upper molars,lower molars supererupted by 0.13 mm. Labeled tissue sections revealed significant amountsof new bone and cementum apposition at the root apex of the unopposed side followingsupereruption for 12 days. Newly apposited cementum and alveolar bone layers wereapproximately 3-fold thicker in the experimental vs the control group, whereas periodontalligament width was maintained. Tartrate-resistant acid phosphatase staining indicated boneresorption at the mesial alveolar walls of unopposed molars and provided in tandem withnew bone formation at the distal alveolar walls an explanation for the distal drift of molars inthis model. Microarray analysis and semiquantitative RT-PCR demonstrated a significantincrease in collagen I, integrin b5, and SPARC gene expression as revealed by comparisonbetween the unopposed molar group and the control group. Immunohistochemical veri-fication revealed increased levels of integrin b5 and SPARC labeling in the periodontalligament of the unopposed molar. Together our findings suggest that posteruptive axialmovement of teeth was accomplished by significant formation of new root cementum andalveolar bone at the root apex in tandem with upregulation of collagen I, integrin b5, andSPARC gene expression. (J Histochem Cytochem 55:127–140, 2007)
KEY WORDS
tooth movement
tooth eruption
bone resorption
matrix remodeling
THE CONCEPT that tooth movement is facilitated by ex-tensive bone tissue reorganization was originally pro-posed by Sandstedt (1904) andOppenheim (1911–1912).Both authors described the various processes of boneformation and absorption on aspects of the movingtooth and established the bone extracellular matrix(ECM) as a dynamic tissue facilitating tooth movement.In the century that followed Sandstedt’s andOppenheim’sseminal papers, the process of tissue remodeling as a
consequence of orthodontic forces has been extensivelystudied (reviewed in Sandy et al. 1993; McCulloch et al.2000). A number of studies have established toothmovement as a complex process involving highly re-fined interactions between growth factors, transcrip-tion factors, and structural proteins (Shimono et al.2003; Sringkarnboriboon et al. 2003). In addition,remodeling of the collagenous ECM of the periodontalligament (PDL) and alveolar bone has been identified asa molecular key process facilitating tooth movement(Nakagawa et al. 1994; Karimbux andNishimura 1995).
Effects of the ECM on cell behavior are often medi-ated by integrin cell surface receptors and/or matricel-lular glycoproteins. Integrins are a family of cell surfacereceptors that attach cells to the matrix and affectcell cycle regulations, directing cells to live or die, toproliferate, or to exit the cell cycle and differentiate(Boudreau and Jones 1999; Giancotti and Ruoslahti
Correspondence to: Thomas G.H. Diekwisch, DMD, PhD (sc.),PhD (phil.), Professor and Head, Department of Oral Biology, AllanG. Brodie Sr. Endowed Chair for Orthodontic Research, Professorfor Anatomy/Cell Biology, Bioengineering, Orthodontics, Periodon-tics, Director, Allan G. Brodie Laboratory for Craniofacial Genetics,UIC College of Dentistry, 801 South Paulina, Chicago, IL 60612.E-mail: [email protected]
Received for publication May 14, 2006; accepted September 14,2006 [DOI: 10.1369/jhc.6A7018.2006].
TheJournal
ofHistoch
emistry&
Cytoch
emistry
C The Histochemical Society, Inc. 0022-1554/06/$3.30 127
Volume 55(2): 127–140, 2007
Journal of Histochemistry & Cytochemistry
http://www.jhc.org
by guest on December 10, 2013jhc.sagepub.comDownloaded from

1999). SPARC (osteonectin, BM-40) modulates cellularinteraction with the ECM by binding to structuralmatrix proteins such as collagen I or by interacting withhydroxyapatite (Bornstein 2000; Brekken and Sage2000; Remelli et al. 2002). Consequentially, signalsfrom matricellular mediators such as integrins orSPARC may regulate and control the manufacture andorganization of newly secreted ECMmolecules, notablycollagen I, by affecting cell behavior. It is thereforeconceivable that signals from periodontal pressure ortension receptors, because they are affected by occlud-ing teeth or mechanical stimulation and may triggermatricellular mediators such as integrins and SPARC,in turn would cause a change in ECM synthesis, espe-cially collagen I. This synthesis of new ECM might beone cause for tooth movement to occur.
We have used the model of the unopposed rodentmolar to study the effects of axial tooth movement onmatrix remodeling and to identify matricellular media-tors. As a first step, we have characterized the amountof mouse molar supereruption in this model using skel-etal preparations, ultrathin ground sections, fluorescentlabels, and histochemical bone markers. These studiesdemonstrated a significant degree of molar tooth super-eruption associated with new cementum and alveolarbone mineralized tissue deposition. In addition, wehave performed tartrate-resistant acid phosphatase(TRAP) staining to detect sites and levels of bone re-sorption in the periodontium and bony sockets ofaffected teeth. To reveal the molecular factors thatmight be involved in matrix remodeling during toothmovement, we have used an ECM gene-based micro-array assay to identify candidate genes. The first threecandidate genes that emerged from our preliminarystudies were collagen I, integrin b5, and SPARC. Wethen verified these findings using semiquantitative RT-PCR and immunohistochemistry. Together our findingsindicated that posteruptive axial movement of teeth in-volves significant formation of new apical root cemen-tum and alveolar bone in tandem with upregulation ofcollagen I, integrin b5, and SPARC gene expression.
Materials and Methods
Experimental Animals and Extraction Technique
Swiss-Webster mice, 35 days old (Charles River; Wilmington,MA) were chosen as experimental animals. Animal experi-ments were approved by and performed according to the
guidelines of the UIC Animal Care Committee. Mice wererandomly divided into control and experimental groups. Micewere anesthetized using ketamine (100 mg/kg) and xylazine(5 mg/kg). Following successful anesthesia, all three left-sidemaxillary molars were extracted (Figure 1). After extraction,pressure was applied to achieve hemostasis. Buprenorphinewas administered as an analgesic SC at a dose of 0.05 mg/kgbefore ending the procedure. Doses of analgesic were adminis-tered every 12 hr for up to 2 days postprocedure. Mandiblesfrom untreated animals were used as controls. For fluorescentlabeling, fluorochromes were applied at concentrations of50 mg/kg (tetracycline hydrochloride), 25 mg/kg (alizarinred), and 30 mg/kg (calcein blue).
Tissue Processing
A group of 15 mice received an IP injection of tetracyclinehydrochloride as a vital stain at the time of extraction. Sub-sequently, they received injections of alizarin red at 4 days andcalcein blue after 8 days. Mice were sacrificed on day 12 byusing CO2 gas and stored in 70% ethanol for overnight fixa-tion. Tissues were dehydrated in a graded series of ethanols,followed by acetone, and then infiltrated with resin (Tech-novit 2000; EXAKT Technologies, Oklahoma City, OK).Subsequently, samples were embedded and prepared for un-decalcified ground sections, fluorescent microscopy, and vonKossa’s and Villanueva’s Osteochrome stain.
For immunohistochemistry, tissues of 15mice were fixedwith10% buffered formalin for 1 week, decalcified in 5% EDTA for2 weeks, and dehydrated in a graded series of ethanols. Sub-sequently, tissues were embedded in paraffin wax, cut into 5-mm-thick sections, and prepared for immunohistochemistry.
Morphometry
To determine the total eruption rate over 12 days in anunopposed situation, three mandibles were skeletonized, andthe difference between molar heights in the unopposed andopposed situation was determined. Molar height in the un-opposed model was compared with molar height in a controlgroup in which opposing molars maintained occlusion. Re-sulting macrographs were oriented by aligning the base ofthe mandible parallel to the base of the image while a secondparallel was constructed through the occlusal plane of theopposed molar. Mandibles were photographed en face usingthe posterior prominences of both condyles and ramus to es-tablish a retrograde plane of orientation.
Alveolar bone and cementum apposition were measuredusing computerized techniques by measuring the distance be-tween the initially stained mineralized zone to the final marginof bone or cementum.Measurements were taken from sagittalsections through the center of the first and second molar.Values were compared between the experimental side and thecontrol side using the paired t-test statistical analysis.
’
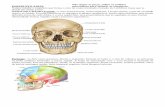
Figure 1 Unopposed mouse molar skull preparations illustrating tooth extraction procedures and unopposed side supereruption. (A) Caudalview onto a skeletonized upper jaw illustrating the position of the incisors (Inc) and maxillary molars (M1, M2, M3). All three left molars wereextracted to create an unopposed situation (M1x). (B) Base of the upper jaw following extraction of the left maxillary molars (X). Note thesockets of alveolar bone remaining immediately after extraction (X). (C) Close-up of an extracted first (M1) and second (M2) maxillary mousemolar. Note the sharp inclination of the mesial (mes) root of the first maxillary molar, providing an indication of the distal (dist) tooth driftin the rodent dentition. Bars: A,B 5 1 mm; C 5 500 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
128 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from

TheJournal
ofHistoch
emistry&
Cytoch
emistry
Axial Movement of Teeth 129
by guest on December 10, 2013jhc.sagepub.comDownloaded from

TheJournal
ofHistoch
emistry&
Cytoch
emistry
130 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from

Histochemistry and von Kossa’s Procedure
Plastic-embedded sections were stained with either Villanueva’sOsteochrome stain to detect newly mineralized tissue matrixor von Kossa’s stain as a mineralized tissue marker. For vonKossa’s procedure, tissue sections were treated with 5% silvernitrate solution as previously described (Diekwisch et al.1995). To detect osteoclasts, paraffin sections were stainedfor TRAP using the Sigma acid phosphatase detectionkit 386A (Sigma; St Louis, MO) according to the manufac-turer’s instructions.
Microarray Technology
For microarray analysis, three mandibles from the controland the experimental group (day 6) each were removed andapical regions prepared using surgical instruments. The dis-sected apical region of each molar contained apical root tipincluding cementum, PDL, and adjacent alveolar bone. Day 6was chosen based on preliminary studies indicating high levelsof gene expression on day 6.
mRNA was extracted from the tissues using Trizol LS Re-agent (Invitrogen; Carlsbad, CA) according to the manufac-turer’s instructions. Quality and quantity of the RNA wastested using spectrophotometry and agarose gel electropho-resis. Optimal quality of RNA from one mouse from eachgroup was used, hybridized, and converted to cDNA via re-verse transcription. cDNA was labeled by 32P isotope andexposed to a GEArray Q Series Mouse Extracellular Matrixand Adhesion Molecules Gene Array (SuperArray; Frederick,MD). The response of 96 genes associated with the ECM andadhesion molecules, plus positive and negative controls, wasevaluated using a software package provided by SuperArray.
Semiquantitative RT-PCR
Total RNA was extracted from apical tissues (apical regionincluding root tip, cementum, PDL, and alveolar bone ofall three molars) of three mice (per group) using the Trizol LSReagent (Invitrogen) and its protocol. Quality and quantity ofthe RNA was tested with spectrophotometry and agarose gelelectrophoresis in the same way samples were prepared formicroarray analysis. RNA was then reverse transcribed andthe product of cDNA was amplified using selected primersand the PCR technique. Primers were selected for type I col-lagen, SPARC,and integrinb5basedon results from themicro-array analysis. Primer sequences were as follows: collagen I—59CGT CGGAGCAGACGGGAG TTT 39 (forward) and 59CAG AGT TTG GAA CTT ACT GTC 39 (reverse), integrinb5—59 TTG CCA AGT TCC AAA GTG A 39 (forward) and59GCG TGA CCT TTT TAT TTC AT 39 (reverse), SPARC—59 CTG CGT GTG AAG AAG ATC CA 39 (forward) and 59
CAT GTG GGT TCT GAC TGG TG 39 (reverse), and b-actin—59GAG CAAGAGAGGTAT CCTGAC 39 (forward)and 59 TTC ATG GAT GCC ACA GGA TTC 39. RT-PCRproducts were then run through agarose gel electrophoresisalong with a b-actin internal control and a DNA ladder.
Immunohistochemistry
Tissue sections were prepared from experimental and controlmolars in sagittal orientation, and mid-sagittal sections wereused for further analysis. Slides were deparaffinized and tis-sues were rehydrated. Immunoreactions were performedusing a polyclonal primary antibody for integrin b5 (SantaCruz Biotechnology; Santa Cruz, CA) and a monoclonal anti-body for SPARC (Santa Cruz Biotechnology). Sections wereincubated with primary antibody at room temperature for2 hr. Dilution of the primary antibodies was previously deter-mined in preliminary trials. The integrin b5 primary antibodywas diluted using PBS. The SPARC primary antibody wasdiluted using a buffered solution of sodium chloride, HEPES,and calcium chloride. Sections were washed three times in PBSand incubated for 10 min with the appropriate secondaryantibody, anti-mouse antibody for SPARC, and anti-rabbitIgG for integrin b5. After three additional washes in PBS,sections were incubated 10 min with streptavidin–enzymeconjugate and then washed three additional times with PBS.Immunoreaction signals were detected using an AEC substrate–chromogen mixture (color substrate), counterstained withhematoxylin, and mounted with GVA mount. The followingcontrols were performed to test for antibody specificity:(1) tissue controls—specificity of the antibody was evaluatedin various tissues, (2) antibody controls by using a dilutionseries, (3) controls with preadsorbed antibody to exclude un-specific binding, (4) controls with preimmune serum to con-trol for binding to serum components, and (5) omission ofprimary antibody as a systematic control.
Results
Measurements of relative levels of molar supereruptionbased on skeletal preparations were performed to deter-mine skeletal dimensions of tooth movement. Whereasin control experiments the occlusal plane of the left andright molar was parallel to the baseline, there was asignificant amount of supereruption on the unopposedside of the experimental group (Figure 1). The amountof molar supereruption on the unopposed side in com-parison to the control side was 0.13 mm (60.06 mm)after 12 days.
’
Figure 2 Tissue apposition in unopposed molars as revealed by fluorescent labels and ultrathin ground sections. (A,B) Overview micrographsillustrating ultrathin cross-sections through a control molar (A) and through an unopposed molar (B). Both were labeled using fluorescentmarkers for tetracycline (green label, after 4 days), alizarin red (red label, after 8 days), and calcein (blue label, after 12 days). Compare the apexof the distal root of the first molar in both images (dr), and note the significant deposition of new root cementum and alveolar bone on theunopposed side (B). The tooth crown (cr) in A is identified for orientation purposes. (C,D) High-magnification images of unopposed molarsindicating the position of fluorescent labels: c1 (tetracycline label, cementum), c2 (alizarin red label, cementum), c3 (calcein blue label,cementum), ab1 (tetracycline label, alveolar bone), ab2 (alizarin red label, alveolar bone), and ab3 (calcein blue label, alveolar bone). In D thepositions of the periodontal ligament (lig) and alveolar bone (ab) are also indicated. Bars: A,B 5 500 mm; C 5 100 mm; D 5 50 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Axial Movement of Teeth 131
by guest on December 10, 2013jhc.sagepub.comDownloaded from

Changes in mineralized tissue dimensions were as-sessed using fluorescent bone markers, and changeswere measured on mid-root sagittal ultrathin groundsections using fluorescent microscopy. Our results re-vealed highly significant differences in the thickness ofalveolar bone and cementum apposition in the unop-posed (experimental) group vs the opposed (control)group in the unopposed mouse molar model (88.26 mmexperimental vs 28.64 mm control for new alveolarbone and 78.81 mm experimental vs 30.61 mm controlfor new cementum). In comparison, levels of mineral-ized tissue apposition were remarkably similar foralveolar bone and cementum (88.26 mm alveolar boneand 78.81 mm cementum for the experimental groupand 28.64 mm alveolar bone and 30.61 mm cementumfor the control group) (Figure 2; Table 1).
Based on the same sections that were used in thepreviously described labeling studies, mean PDL diame-ter was calculated from mid-root cut sagittal sectionsand compared between experimental and controlgroup. In opposition to the dramatic changes observedin mineralized tissue apposition, PDL width remainedunaffected from supereruption in unopposed situations,and total PDL width was z80 mm in both unopposedand opposed conditions (Figure 3; Table 1).
Using the TRAP-staining procedure, distinct TRAP-positive demarcation linings containing multiple osteo-clasts were recorded along the mesial alveolar wall ofthe unopposed molars (Figure 4). In contrast, controlmolars featured only a few single osteoclasts and noTRAP-positive alveolar linings (Figure 4). There wereno osteoclasts or indications of TRAP activity at theapex of unopposed molars (Figure 4). These findingssuggest the presence of bone resorption at the mesialalveolar walls.
To determine whether newly formed apical tissueswere mineralized, von Kossa’s technique for the detec-tion of inorganic phosphate was applied. There was asignificantly enhanced layer of mineralized cementumin the experimental group in comparison to the controlgroup (Figure 5). The entire cellular cementum regionwas labeled by von Kossa’s procedure, indicating thatthe newly formed apical cementum was mineralized(Figure 5). The mineralizing cementoblast contours at
the ligament/cementum interface resembled patterns ofnormal apical cementogenesis (Figure 5).
ECM microarray comparisons were performedusing mRNA extracted from root apices of super-
Table 1 Tooth apex mineralized tissue apposition and periodontal ligament (PDL) dimensions in unopposed and control molars after12 daysa
Experimental Control Mean difference SD Significance
Alveolar bone 88.26 6 19.20 28.64 6 5.65 59.63 17.86 .0.001***Cementum 78.81 6 20.55 30.61 6 7.45 48.20 22.23 .0.001***PDL width 83.06 6 12.08 81.57 6 12.41 1.49 20.62 0.783
aThere were highly significant differences in the thickness of alveolar bone and cementum apposition in the unopposed (experimental) group vs the opposed(control) group in the unopposed mouse molar model (88.26 mm experimental vs 28.64 mm control for new alveolar bone, and 78.81 mm experimental vs 30.61 mmcontrol for new cementum). In comparison, levels of mineral tissue apposition were remarkably similar for alveolar bone and cementum (88.26 mmalveolar bone and78.81 mm cementum for the experimental group and 28.64 mm alveolar bone and 30.61 mm cementum for the control group). Moreover, PDL width remainedunaffected from supereruption in unopposed situations, and total PDL width was about 80 mm in both unopposed and opposed conditions.***Highly significant.
Figure 3 Twelve-day apposition statistics for apical alveolar bone,cementum, and periodontal ligament (PDL) in unopposed andcontrol molars. (A) Thickness of newly apposited cementum andalveolar bone was z2.5- to 3-fold higher in the unopposed molargroup than in the opposed molar group. There was a total of 80–90 mm each of both alveolar bone and cementum apposited in theunopposed group, whereas the apposition values for both bone andcementum were significantly less in the opposed molar group, eachtotaling z30 mm. Interestingly, periodontal ligament widthremained unaffected from supereruption in unopposed situations,and total PDL width was z80 mm in both unopposed and opposedconditions (B). Exp., experimental; Cont., control; PDL, periodontalligament. Exact values are documented in Table 1.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
132 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from

Figure 4 Osteoclast activity as visualized via tartrate-resistant acid phosphatase (TRAP) staining indicates bone resorption at themesial alveolarwalls of unopposed mouse molars. (A,B) Comparable mid-sagittal sections through mouse jaws with the second molar in the center and a smallfragment of the third molar at the rightmargin of themicrograph.A is from a control and B is from amouse jaw that remained in an unopposedsituation for 9 days. (C–E) Higher-magnification images of the three selected root areas marked in B by an asterisk. Note the distinct TRAP-positive lining (TRAP) containing multiple osteoclasts (oc) along the mesial alveolar wall of the unopposed molars. In contrast, control molarsfeatured only a few single osteoclasts (oc). The following were marked for orientation purposes: alveolar bone (ab), control group (contr.), andexperimental group (exp.). Bars: A,B 5 500 mm; C–E 5 100 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Axial Movement of Teeth 133
by guest on December 10, 2013jhc.sagepub.comDownloaded from

TheJournal
ofHistoch
emistry&
Cytoch
emistry
134 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from

erupted and control molars and commercially availableECM gene-specific arrays. Our results based on micro-array analysis of 96 selected ECM genes documenteda 4.4-fold increase in collagen I gene expression, a1.9-fold increase in integrin b5 gene expression, and a2.3-fold increase in osteonectin (SPARC) gene expres-sion, whereas the remainder of the ECM genes inves-tigated in this assay were significantly below the 2-foldthreshold (Table 2).
Semiquantitative RT-PCR analysis and densitometrywere performed to verify microarray data on changes inECM gene expression in unopposed molar root tissues.Using semiquantitative RT-PCR, all three gene prod-ucts, collagen I, integrin b5, and SPARC (osteonectin),demonstrated significantly higher expression levelsin the unopposed molar group compared with theopposed molar control group, indicating an activeinvolvement of ECM genes in tissue remodeling thatoccurs during axial tooth movement (Figure 6). Therewas no difference in b-actin control gene expressionlevels between the experimental and the control group,and the negative control reaction (Neg) did not yieldany product in all four experiments (Figure 6). Den-sitometric evaluation of RT-PCR results yielded a morethan 2-fold increase in collagen I and SPARC gene ex-pression and a slight increase (1.3-fold) in integrin b5gene expression (Figure 6E).
To verify semiquantitative RT-PCR and microarrayresults on a protein level, immunohistochemical studieswere performed using same-thickness adjacent sectionsand identical reaction conditions in a single experiment.Studies were focused on integrin b5 and osteonectin,whereas the ubiquitous collagen I was not used forthese studies. Immunoreactions revealed specific label-ing reactions for both integrin b5 and osteonectin in thePDL, whereas surrounding dental tissues such as alveo-lar bone or cementum exhibited little or no reactionproducts (Figure 7 and Figure 8). Labeling intensity forintegrin b5 and osteonectin in the unopposed molargroup vs the control group was dramatically enhancedunder otherwise identical conditions on adjacent same-thickness sections (Figure 7). Careful study of the mi-crographs revealed localization of integrin b5 reactionproducts to the cell surfaces of PDL fibroblasts whereasosteonectin (SPARC) was distributed in ECMs includ-ing the adjacent bone and cementum matrix (Figure 8).Control immunoreactions were negative.
Discussion
In the present study we have used the model of theunopposed mouse molar as a model system to inves-tigate tissue dynamics and ECM remodeling followingaxial movement of teeth. Our data established that in the12-day unopposed mouse molar model, unopposedmolars erupted 0.13 mm above the level of the contra-lateral side. Historically, the model of the unopposedmouse molar was conceived in the early 1950s at theUniversity of Illinois by Viennese emigres to distinguishgenetic and acquired traits in the functional anatomyof dentition but has been used more recently to assesspatterns of posteruptive tooth movement (Schneider andMeyer 1965; Cohn 1966; Anneroth and Ericsson 1967;Levy and Maillard 1980; Compagnon and Woda 1991).
Our results demonstrated that layers of similarthickness of both mineralized tissues, alveolar boneand cementum, were deposited at the root apex follow-ing removal of the antagonistic tooth. Morphometricalevaluation of fluorescent labels on ultrathin ground sec-tions revealed that significant amounts of cementumand alveolar bone were gradually deposited at the apexof unopposed molars, indicating that posteruptive axialmovement was greatly facilitated by the active deposi-tion of mineralized tissues at the apex of the affectedtooth. Whereas some investigators reported lesseramounts of cementum deposition in comparison to al-veolar bone deposition (Schneider and Meyer 1965;Cohn 1966) or complete absence of newly formed ce-mentum (Levy and Maillard 1980), other studies(Anneroth and Ericsson 1967; Compagnon and Woda1991) report similar findings of significant amounts ofnew cementum deposition after loss of antagonists. Ourfindings indicated that the rate by which both mineral-ized tissues were deposited was approximately identical
’
Figure 5 Formation of new mineralized tissues at the apex of unopposed molars. To determine whether newly formed apical tissueswere mineralized, von Kossa’s technique for the detection of inorganic phosphate was applied. Note the significantly enhanced layer ofmineralized cementum (cem) on the experimental side (B) in comparison to the control side (A). The entire cellular cementum region waslabeled by von Kossa’s procedure, indicating that the newly formed apical cementum was mineralized. Root dentin (dent) and alveolar bone(ab) were labeled for orientation purposes. (C) High-magnification image of the interface between periodontal ligament (lig) and apicalcementum (cem). The mineralizing cementoblast contours at the ligament/cementum interface resembled patterns of normal apical cemen-togenesis. Bars: A,B 5 100 mm; C 5 20 mm.
Table 2 Transcript levels of extracellular matrix-related genesas revealed via microarray analysisa
Gene Genbank # Ratio day 6 vs control
Collagen I NM_007742 4.4Integin b5 NM_010580 1.9Osteonectin (SPARC) NM_009262 2.3
aExtracellular matrix microarray comparison between 12-day unopposed andcontrol molar apex mRNA revealed differential expression of collagen I, SPARC,and integrin b5 genes before and after axial movement of teeth. Microarraydata suggest a significant increase in collagen I, SPARC, and integrin b5 geneexpression, a finding that was confirmed using RT-PCR (Figure 6) and immuno-reactions (Figure 7 and Figure 8).
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Axial Movement of Teeth 135
by guest on December 10, 2013jhc.sagepub.comDownloaded from

between cementum and alveolar bone, suggesting thatsimilar stimuli might result in similar tissue responses incementum and alveolar bone.
Using the TRAP-staining procedure, we reporteddistinct TRAP-positive demarcation linings containingmultiple osteoclasts along the mesial alveolar walls ofunopposed molars. Secretion of TRAP has been asso-ciated with resorptive behavior of osteoclasts (Kirsteinet al. 2006) and might in this case suggest the presenceof bone resorption at the mesial lining of the alveolarwalls in this model. Bone resorption at the mesial al-veolar walls would be a logical phenomenon associatedwith the distal drift of these molars that occurs in anunopposed situation and that has been previously re-ported (Schneider and Meyer 1965). This finding alsocorresponds with the new bone apposition at the mesialalveolar walls that was detected in our fluorescent la-beling studies using ground sections. Thus, the distaldrift of unopposed rodent molars might be explainedthrough alveolar bone resorption at the mesial alveolarwalls and new alveolar bone deposition at the distalalveolar walls.
Histochemical analysis using von Kossa’s proceduresuggested that the newly formed tissue at the apex ofunopposed molars was mineralized cellular cementum.von Kossa’s stain is a routine procedure to detect inor-ganic calcium phosphate deposits as a marker for min-eralized tissues. Staining newly formed cementum via
von Kossa’s procedure established distinct boundariesbetween mineralized tissues and PDL similar to sharpmineralization boundaries present in normal cellularand acellular cementum. This procedure visualized thata new layer of mineralized cellular cementum wasformed at the apical tip of the unopposed tooth root.
A number of authors have concluded that the PDLmay not be an essential morphological contributortoward tooth eruption, as suggested in earlier studies byGowgiel (1967) and Cahill and Marks (1980) of den-tinal dysplasia type I dentitions. Other studies estab-lished a principal role of the PDL as it pertains to thecontinuous eruption of permanently growing rodentincisors (Berkovitz and Thomas 1969; Berkovitz 1971).Our study resulted in a number of intriguing findingsrelated to the dimensions and possible function of thePDL during posteruptive tooth movement in rodentmolars. Morphometrical analysis indicated that usingthe unopposed molar model, PDL width remained un-affected after axial movement of teeth, in spite of ex-tensive new alveolar bone and cementum deposition.These findings were congruent with previous studiesreporting that PDL width was maintained throughoutthe lifetime of adult mammals (McCulloch andMelcher1983; Melcher 1986). Thus, one of the most remark-able findings of our study was the extraordinary ca-pacity of the PDL to preserve its width even duringtooth movement and even in light of mineralized tissue
Figure 6 RT-PCR amplification of im-portant extracellular matrix (ECM)gene products collagen I and SPARC(osteonectin) and the integrin b5 cellsurface receptor in the unopposedmouse molar model. All three geneproducts, collagen I (A), integrin b5(B), and SPARC (osteonectin, C) dem-onstrated significantly higher expres-sion levels in the unopposed molargroup (Exp) compared with opposedmolar control group (Cont), indicatingan active involvement of ECM genesin the tissue remodeling that occursduring axial tooth movement. Therewas no difference in b-actin controlgene expression levels between theexperimental and the control group(D). The negative control reaction
(Neg) did not yield any product in all four experiments. (E) Densitometric analysis of our semiquantitative RT-PCR results. Note increasedgene expression levels for collagen I, SPARC, and integrin b5 in the 6-day unopposed group as compared with the control group.
’
Figure 7 Immunohistochemistry of integrin b5 and osteonectin (SPARC) in the unopposed mouse molar model. In this set of micrographs,localization and relative staining intensity were compared for integrin b5 (A,B) and osteonectin (C,D). Comparisons were performed betweenthe control group (A,C) and the unopposed molar experimental group (B,D). Both integrin b5 and osteonectin were specifically localized in theperiodontal ligament (lig), whereas surrounding dental tissues such as alveolar bone or cementum exhibited little or no reaction products.Labeling intensity for integrin b5 and osteonectin in the unopposed molar group vs the control group (B,D vs A,C) was dramatically enhancedunder otherwise identical conditions on adjacent same-thickness sections. The following tissues were identified for orientation purposes:alveolar bone (ab), periodontal ligament (lig), root cementum (cem), root (rt), junctional epithelium ( je), enamel demineralized space (en),dentin (dt), and dental pulp (plp). This highly specific localization of key factors suggests a pivotal role of the PDL in the ECM-mediated controland modulation of tooth movement. Bars: A,B 5 100 mm; C,D 5 50 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
136 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from

TheJournal
ofHistoch
emistry&
Cytoch
emistry
Axial Movement of Teeth 137
by guest on December 10, 2013jhc.sagepub.comDownloaded from

TheJournal
ofHistoch
emistry&
Cytoch
emistry
138 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from

deposition from two opposing sides. However, widthpreservation was not the only feature of the PDL thatwas highlighted during our study. A second feature wasthe concentration of integrin-b5 and SPARC immuno-reactive signals in the PDL and the significant enhance-ment of reaction intensity for both gene productsfollowing axial tooth movement. These findings indi-cate that the PDL may play a major role in the coor-dination of the ECM signaling events related to toothmovement while at the same time maintaining its width.
To verify our findings of a potential upregulation ofthe ECM gene products collagen I, SPARC (osteonec-tin), and integrin b5, as initially suggested by micro-array-based results, we confirmed our findings usingsemiquanitative RT-PCR and immunohistochemistry(SPARC and integrin b5 only). We have refrained fromperforming immunoreactions for collagen I because ofthe abundance of this gene product at the tooth apexand because of inherent specificity issues with collagenantibodies. Our findings confirmed our initial micro-array data suggesting that all three ECM gene productswere greatly upregulated in the unopposed molarsituation and that the ECM may play an importantrole during tissue remodeling that occurs duringtooth movement. Based on these findings, we thereforespeculate that matricellular regulators such as SPARCand integrin b5 may act as transducers for mechanicalstressors and affect the synthesis of collagen I and otherECM structural gene products, which in turn may beonce more functionally modulated by SPARC to bindwith collagens or to interact with hydroxyapatite toadapt to constantly changing mechanical stress port-folios imposed on the occluding and non-occludingtooth. Our data suggest that collagen I and SPARCare more concentrated in the unopposed molar PDL,whereas the width of the PDL remains constant. Thisphenomenon might be explained by spatial changes inother ECM components such as proteoglycans, as pre-viously discussed (Cheng et al. 1999; Sato et al. 2002).Although, at present, this remains a speculative model,our data confirmed our hypothesis about changes inECM gene expression following loss of antagonists inthe unopposed mouse molar model.
Acknowledgments
Support for this study was kindly provided by the NationalInstitute of Dental and Craniofacial Research, National
Institutes of Health, Grant R01 DE-15425 (to TGHD) andfrom the Brodie Endowment to the Department of Ortho-dontics, UIC College of Dentistry, Chicago, IL.
Literature Cited
Anneroth G, Ericsson SG (1967) An experimental histologicalstudy of monkey teeth without antagonist. Odontol Revy 18:345–359
Berkovitz BK (1971) The effect of root transection and partial rootresection on the unimpeded eruption rate of the rat incisor. ArchOral Biol 16:1033–1043
Berkovitz BK, Thomas NR (1969) Unimpeded eruption in the root-resected lower incisor of the rat with a preliminary note on roottransection. Arch Oral Biol 14:771–780
Bornstein P (2000) Matricellular proteins: an overview. Matrix Biol19:555–556
Boudreau NJ, Jones PL (1999) Extracellular matrix and integrin sig-nalling: the shape of things to come. Biochem J 339:481–488
Brekken RA, Sage EH (2000) SPARC, a matricellular protein: at thecrossroads of cell-matrix. Matrix Biol 19:569–580
Cahill DR, Marks SC Jr (1980) Tooth eruption: evidence for thecentral role of the dental follicle. J Oral Pathol 9:189–200
Cheng H, Caterson B, Yamauchi M (1999) Identification and immu-nolocalization of chondroitin sulfate proteoglycans in tooth ce-mentum. Connect Tissue Res 40:37–40
Cohn SA (1966) Disuse atrophy of the periodontium in mice follow-ing partial loss of function. Arch Oral Biol 11:95–105
Compagnon D, Woda A (1991) Supraeruption of the unopposedmaxillary first molar. J Prosthet Dent 66:29–34
Diekwisch TGH, Berman BJ, Gentner S, Slavkin HC (1995) Initialenamel crystals are not spatially associated with mineralized den-tine. Cell Tissue Res 279:149–167
Giancotti FG, Ruoslahti E (1999) Integrin signaling. Science 285:1028–1032
Gowgiel JM (1967) Observations on the phenomena of tooth erup-tion. J Dent Res 46:1325–1330
Karimbux NY, Nishimura I (1995) Temporal and spatial expressionsof type XII collagen in the remodeling periodontal ligament duringexperimental tooth movement. J Dent Res 74:313–318
Kirstein B, Chambers TJ, Fuller K (2006) Secretion of tartrate-resistant acid phosphatase by osteoclasts correlates with resorptivebehavior. J Cell Biochem 98:1085–1094
Levy G, Maillard M (1980) Histologic study of the effects of occlu-sal hypofunction following antagonist tooth extraction in the rat.J Periodontol 51:393–399
McCulloch CA, Lekic P, McKeeMD (2000) Role of physical forces inregulating the form and function of the periodontal ligament.Periodontology 24:56–72
McCulloch CA, Melcher AH (1983) Continuous labeling of theperiodontal ligament of mice. J Periodontal Res 18:231–241
Melcher AH (1986) Periodontal ligament. In Bhaskar SN, ed. Orban’sOral Histology and Embryology, 10th ed. Toronto, Mosby,198–231
NakagawaM, Kukita T, Nakasima A, Kurisu K (1994) Expression ofthe type I collagen gene in rat periodontal ligament during toothmovement as revealed by in situ hybridization. Arch Oral Biol39:289–294
Oppenheim A (1911–1912). Tissue changes, particularly of the bone,
’
Figure 8 Localization of integrin b5 and osteonectin (SPARC) in the unopposed molar root apex PDL. These are high-magnification images ofparaffin sections through the apical tips of unopposed mouse molar root apices treated with anti-integrin b5 (A) and anti-osteonectin (B)antibodies. Strong reaction products were detected in the periodontal ligament (lig), whereas adjacent tissues dentin (den), cementum (cem),and alveolar bone (ab) were devoid of reaction products. Careful study of the micrographs revealed that integrin b5 reaction products werelocalized to the cell surfaces of periodontal ligament fibroblasts (A), whereas osteonectin (SPARC) was distributed in ECMs including theadjacent bone and cementum matrix (B). Cellular localization findings were consistent with the normal distribution of integrin b5 andosteonectin and suggest a distinct role of these molecules in tissue remodeling that occurs during axial tooth movement. Bar 5 50 mm.
TheJournal
ofHistoch
emistry&
Cytoch
emistry
Axial Movement of Teeth 139
by guest on December 10, 2013jhc.sagepub.comDownloaded from

incident to tooth movement. American Orthodontist III:57–67,113–132
Remelli M, Luczkowski M, Bonna AM, Mackiewicz Z, Conato C,Kozlowski H (2002) Cu (II) ion coordination to SPARC: a modelstudy on short peptide fragments. N J Chem 27:245–250
Sandstedt C (1904) Einige Beitrage zur Theorie der Zahnregulierung.Nordisk Tandlakare Tidskrift 5:236–256
Sandy JR, Farndale RW, Meikle MC (1993) Recent advances inunderstanding mechanically induced bone remodeling and theirrelevance to orthodontic theory and practice. Am J Orthod Dento-facial Orthop 103:212–222
Sato R, Yamamoto H, Kasai K, Yamauchi M (2002) Distribution
pattern of versican, link protein and hyaluranic acid in the ratperiodontal ligament during experimental tooth movement. JPeriodontal Res 37:15–22
Schneider BJ, Meyer J (1965) Experimental studies on the interrela-tions of condylar growth and alveolar bone formation. AngleOrthod 35:187–199
Shimono M, Ishikawa T, Ishikawa H, Matsuzaki H, Hashimoto S,Muramatsu T, Shima K, et al. (2003) Regulatory mechanismsof periodontal regeneration. Microsc Res Tech 60:491–502
Sringkarnboriboon S, Matsumoto Y, Soma K (2003) Root resorptionrelated to hypofunctional periodontium in experimental toothmovement. J Dent Res 82:486–490
TheJournal
ofHistoch
emistry&
Cytoch
emistry
140 Luan, Ito, Holliday, Walker, Daniel, Galang, Fukui, Yamane, Begole, Evans, Diekwisch
by guest on December 10, 2013jhc.sagepub.comDownloaded from