
Expression pattern of the Tbr2 (Eomesodermin) gene during mouse and chick brain development
Upload
independentCategory
view
2download
0
\ Pergamon Int[ J[ Devl Neuroscience\ Vol[ 04\ No[ 0\ pp[ 016Ð026\ 0886Copyright Þ 0886 ISDN[ Published by Elsevier Science Ltd
Printed in Great Britain[ All rights reserved9625Ð4637:86 ,06[99¦9[99
PII] S9625Ð4637"85#99957Ð7
EXPRESSION PATTERN OF GLUTAMATE DECARBOXYLASE "GAD#IN THE DEVELOPING CORTEX OF THE EMBRYONIC
CHICK BRAIN
PER LUNDGREN\� LENORE JOHANSSON\$ CAMILLA ENGLUND\%A� KE SELLSTRO� M$ and MATS!OLOF MATTSSON�&
�Dept of Cellular and Developmental Biology\ Umea� University\ S!890 76 Umea� Sweden^ $National Defence ResearchEstablishment\ S!890 71 Umea� Sweden^ %Center for Molecular Pathogenesis\ Umea� University\ S!890 76 Umea� Sweden
"Received 14 October 0884^ revised 29 May 0885^ accepted 20 May 0885#
Abstract*The development of the GABAergic system in the chick embryo telencephalon has been studied[Special emphasis was placed on the development of glutamate decarboxylase "GAD# between embryonicday 7 "E7# and E06[ The GABA immunoreactivity and neuron!speci_c enolase expression was detectedsimultaneously in glutardialdehyde _xed sections\ which con_rmed that GABAergic cells exhibit neuronalphenotype[ The GAD expression was studied by means of immunohistochemistry on cryo!sectionedmaterial both at the light and electron microscopic levels[ Furthermore\ the presence and localization ofGAD54 and GAD56 mRNAs were studied with an in situ hybridization technique with digoxigenin!labeledRNA probes[ Protein expression as well as mRNA appearance mostly coincided both temporally andspatially[
In the parahippocampal area\ as well as in other regions of the developing cortex\ GAD staining wasseen from E7 onwards[ The number of positive cells increased as did the intensity of staining up to E03[As observed in the electron microscope\ the GAD protein was co!localized with GABA in most cases\although some GAD!positive cells devoid of GABA!staining also were observed[ The pattern of GADmRNA expression was in general similar to that of GAD immunostaining[ Both GAD54 and GAD56mRNA were detected during the entire period[ Furthermore\ GAD56 mRNA localization spatially wasmore correlated with GAD protein expression[ The study provides evidence for the notion that developmentof the GABAergic system occurs rapidly during embryogenesis and\ as suggested from mRNA data\ thattwo forms of GAD with a slight di}erence in distribution can contribute to this[ Þ 0886 ISDN
Key words] GAD54\ GAD56\ mRNA\ GABA\ NSE\ telencephalon\ immunostaining\ immuno!electronmicroscopy\ in situ hybridization[
g!Aminobutyric acid "GABA# is the major inhibitory transmitter in both the vertebrate and theinvertebrate nervous systems[ The synthesis of GABA is performed predominantly by the enzymeglutamate decarboxylase "GAD#\ although alternative pathways may exist[15 To be consideredGABAergic\ a neuron thus must exhibit GAD!activity\ it must contain GABA\ and this GABAneeds to be released upon depolarization[ After release\ the transmitter is transported back into theneuron[ The GAD enzyme recently has been cloned from several species\ and is found to exist in twoforms at least in mammals[ Each form is encoded by a separate gene4\01 and exhibits dissimilarities interms of molecular mass\ amino acid sequence\ co!factor dependence and possibly subcellularlocalization[ Commonly\ each form is named according to its Mw of 54 and 56 kDa\ respectively"GAD54 and GAD56#[
A number of studies\ particularly in mammals\ has been published concerning the developmentof the GABAergic phenotype[2\8\09 Similar studies also have been performed on the developing chickembryo\ where structures such as the telencephalon are developing considerably earlier than inmammals[05 The time of appearance of GABA!immunoreactive neurons di}ers between structures[Accordingly\ GABA!staining has been documented as early as embryonic day 3 "E3# in the chickspinal cord1 as well as in cranial motor nerves[3 The paraventricular zone of the cortex exhibitsGABA!staining by E7 and structures such as the nucleus magnocellularis and nucleus laminarisdisplay GABAergic cells by E01ÐE03[7 It has been suggested also that part of the GABA!expressionis transient[3\11 This also has been observed in the developing rat spinal cord[12 The reason fortransient GABA expression is unclear\ but may indicate a non!transmitter function for GABA[Possibly\ GABA has trophic actions during embryogenesis[ In rodents\ GABA immunoreactivity
&To whom correspondence should be addressed[
016

P[ Lundgren et al[017
has been documented by E02 in the brainstem and diencephalon07\14 and by E03 in the cerebralcortex[00
We aim for a greater understanding of how transmitter speci_city is obtained[ Previously\ weshowed that GABAergic development in the paraventricular area of the chick telencephalon is arapid process\ GABAergic neurons being present by E7\ and in high numbers only 1 days later[11
How is GAD expression correlated to these _ndings< In particular\ can both forms of GAD beinvolved in this GABA synthesis< The present paper deals with the pattern of GAD expression\ onboth protein and mRNA levels\ and is an attempt to further clarify these matters[
EXPERIMENTAL PROCEDURES
Handlin` of embryos
Fertilized White Leghorn eggs obtained from a local supplier "Agrisera AB\ Va�nna�s\ Sweden#were incubated at 26[7>C in a humidi_ed atmosphere in an Agroswede egg incubator "Malmo� \Sweden# for 7\ 00\ 03 or 06 days[ The eggs were turned automatically every 1 hr[
Preparation for electron microscopy
The brains were recovered on a petri dish _lled with phosphate!bu}ered saline "PBS#\ pH 6[3[The parahippocampus and its adjacent parts were dissected out under microscope and transferredto a _xation solution made of freshly depolymerized 3) paraformaldehyde "Riedel de Hae�n# and9[14) glutardialdehyde "Fluka\ grade I# in PBS[ The material remained in this mixture for 0 hrat room temperature "RT#\ was transferred to a solution made up of 2) newly depolymerizedparaformaldehyde in PBS\ pH 6[3\ and stored at 3>C overnight[
The tissue was cut into 9[2mm thick pieces and in_ltrated in 19) polyvinylpyrolidone in 1[2Msucrose in PBS for 1 hr at RT\ followed by mounting on stubs which subsequently were frozen inliquid nitrogen[
Ultrathin cryo!sections\ 79Ð099 nm\ were made by means of a RMC MT!6999 ultramicrotomeequipped with a cryo!device CR10 at −87>C[ The sections were ~oated on 1[9M sucrose and 9[64)gelatine\ picked up with a platinum loop\ and mounted on Formvar!coated nickel grids with 024hexagonal meshes[
Immuno!electron microscopy
Preincubation of sections was performed in a solution of 9[90) Boehringer Mannheim blockingreagent and 9[64) glycine\ 04min[ The specimens then were incubated in a primary antibody mixcontaining rabbit glutardialdehyde!conjugated GABA antibody "No[ 889#21 and sheep anti!GADantibody "No[ 0339!3#\16 both diluted 0]399 in the same mixture as the preincubation\ for 0[4 hr inRT[ The sections were rinsed thoroughly for 29min in the same bu}er and then incubated with asecondary antibody\ a goat anti!rabbit antibody with 4 nm gold particles "Biocell GAR 4 No[ 8502#\and a donkey anti!sheep antibody conjugated with 09 nm gold particles "Biocell DASS 09 No[8367#\ both diluted 0]19 in the same bu}er[ After the secondary antibody incubation\ the sectionswere rinsed for 29min in PBS and 4min in distilled water and then embedded in 1) methyl celluloseand 9[2) uranyl acetate[ The sections were dried and inspected in a Jeol 099!CX transmissionelectron microscope at 79 kV[ Microphotographs were taken using Kodak electron microscopy E!star Thick Base 3378 _lm[
Immunohistochemistry
Tissue preparation[ Embryonic chick telencephali "E7ÐE06# were dissected out on PBS!_lled petridishes and immediately transferred to cold 3) formaldehyde\ i[e[ newly depolymerized para!formaldehyde "Riedel de Hae�n\ Germany# in PBS[ The _xation was carried out at 3>C for 3Ð7 hrdepending on the brain size[ The brains subsequently were transferred to a mix of 19) sucrose"BDH# and 0) formaldehyde in PBS[ The tissue was allowed to be in_ltrated by the sucrose mixtureat 3>C overnight[ The specimens were mounted with Tissue!Tek O[C[T[ compound!embeddingmedium "Histolab\ Gothenburg\ Sweden# on a piece of cardboard and frozen in liquid propane[The deep!frozen brains were stored in plastic test tubes at −69>C until further treatment[ Sectioning

GAD in the developing chick brain 018
was performed in a Reichert Jung RM 1799E cryo!state at −06>C[ Sections of 19mm were mountedon Super Frost:Plus microscope slides "Histolab# and allowed to dry for at least 29min[ The sectionsthat were not used immediately were stored in plastic containers at −19>C[
Immunostainin`[ The GAD immunostaining was performed on the cryo!sectioned material withsheep anti!GAD antibody "No[ 0339!315\ obtained from NINDS\ Bethesda\ MD\ U[S[A[# diluted0]199 in 9[0) bovine serum albumin "BSA\ Sigma fraction V# in PBS for 1 hr at RT and at 3>Covernight[ Prior to the staining procedure\ the frozen tissue was dried at RT for approximately4min and quickly rinsed in PBS\ pH 6[ 1[ A preincubation mixture made up of 9[0) BSA in PBSwas added for 29min at RT[ After a thorough wash in PBS\ a secondary biotinylated rabbit anti!sheep IgG "Vector#\ diluted 0]199 in the same bu}er that was used in the earlier steps\ was added tothe slices for 0 hr at RT[ Sections were washed thoroughly in PBS and thereafter a tertiary ABCÐElite complex "Vector# was added for 0 hr[ After washing in PBS\ staining was performed using aDAB kit 3990 "Vector# with nickel enhancement prepared according to the manufacturer|s instruc!tions[ Following staining\ sections were dehydrated through ethanol into xylene and mounted withDPX mounting medium "BDH#[ As controls in all the immunostaining protocols\ serum from thesame species\ diluted as the primary antibody\ was used[ For GAD staining\ the proper pre!immunesheep serum acted as control[
The GABA! and neuron!speci_c enolase "NSE# were detected simultaneously by the followingprotocol] para.n!embedded sections "09mm# from _xed material "3) formaldehyde\ 1[4) glu!tardialdehyde in PBS\ 1Ð5 hr# were incubated with a polyclonal rabbit primary antibody directedagainst NSE "DAKOPATTS# "diluted 0]099# in the same bu}er as above plus 9[0) Triton X!099"Sigma# for 1 hr at RT and then at 3>C overnight[ After extensive washing in PBS\ a secondarybiotinylated donkey anti!rabbit IgG "diluted 0]199 in the same bu}er as for the preincubation# wasadded for 0 hr at RT[ Sections were washed thoroughly in PBS and thereafter an ABCÐElite complex"Vector# was added for 0 hr[ After washing in PBS\ the AEC detection system "Vector# was used tovisualize antigenÐantibody complexes[ A red color was obtained after 4Ð09min[ The AEC!processwas stopped by a short rinse in PBS\ followed by a 04min post!_xation in 1[4) glutardialdehydediluted in PBS "TAAB EM!grade# and subsequently a rigorous washing in PBS[ The GABA!stainingwas performed subsequently as described above with the exception of the primary rabbit antibody"GABA No[ 88929\ diluted 0]199^ Incstar\ diluted 0]4999#\ which was applied for 1 hr at RT[ Forimmunological detection\ a gold!conjugated goat anti!rabbit IgG with 4 nm gold particles "BiocellResearch Laboratories#\ diluted 0]199 in BSA:PBS\ was incubated at RT for 1 hr[ Slices were washedand the antigenÐantibody complex was visualized by using a silver!enhancing kit "SEKL 04\ BiocellResearch Laboratories#[ The enhancement procedure was stopped after inspection in microscopeby rinsing in tap water for 4min[
IN SITU HYBRIDIZATION
All solutions used for cryo!sectioning for in situ hybridization were treated with diethyl pyro!carbonate "DEPC# "Sigma#\ to avoid contamination with RNases[17 All glassware and other equip!ment were heated to 079>C for 5 hr[ Sectioned material was obtained as described under Tissuepreparation above[
Probes
Adjacent sections were processed with digoxigenin!labeled rat GAD56 and GAD54 RNA probes03
made according to the manufacturer|s "Boehringer Mannheim# instructions[
Hybridization and detection
This in situ hybridization protocol is modi_ed from the one described by Esclapez et al[03
Sections were rinsed for 29min in PBS and pretreated as follows] PBT "0× PBS ¦9[0) Tween!19\Sigma# for 4min^ 9[14mg:ml proteinase K "GIBCO:BRL# in PBT for 09min^ PBS with 9[1)glycine for 4min^ PBS for 4min\ all at RT[ After these pretreatments\ sections were incubated for0hr at RT in a prehybridization solution containing 49) formamide "Merck#\ 649mM NaCl\ 14mMEDTA\ pH 7[9 "Merck#\ 14mM piperazine!N\N?!bis "1!ethanesulfonic acid# "PIPES\ Boehringer

P[ Lundgren et al[029
Mannheim#\ 9[1) sodium dodecyl sulfate "SDS\ GIBCO:BRL#\ 9[91) Ficoll "Pharmacia#\ 9[91)polyvinylpyrrolidone "Sigma#\ 9[91) BSA "Sigma# and 149mg:ml _sh sperm DNA "Amresco#[
A hybridization solution consisting of the prehybridization solution with the addition of 099mMdithiothreitol "DTT\ BioRad#\ 3) dextran sulfate "Pharmacia# and 9[1 ng:ml digoxigenin!labeledRNA probe then was applied to the slides and a coverslip was lowered on top so that the liquidspread over the sections[ The slides were placed horizontally in a slide box containing tissue soakedwith 4× SSC "0× SSC] 049mM NaCl\ 59mM Na citrate\ pH 6[9#\ the box was sealed and incubatedat 49>C for a minimum of 05 hr[
Following hybridization\ sections were immersed in a 3× SSC solution containing 09mM sodiumthiosulfate "Merck#\ in which coverslips slide o}[ The slides subsequently were incubated at 49>Cfor 29min[ This was followed by rinsing in 1× SSC with 09mM sodium thiosulfate for 1 hr at RT"low stringency condition# and 9[0× SSC with 09mM sodium thiosulfate for 29min at 44>C "highstringency condition#[ After these washes\ the sections were processed for immunodetection[ Thesections were washed in 099mM TrisÐHCl bu}er\ 049mM NaCl\ pH 6[4 "Bu}er 0# for 19min^incubated for 0 hr at RT in the same bu}er supplemented with 9[4) blocking reagent "BoehringerMannheim# and 9[2) Triton!X 099 "Sigma#\ and then incubated overnight at 3>C with the alkalinephosphatase!conjugated digoxigenin antibody "Boehringer Mannheim# diluted 0]0999 in Bu}er 0supplemented with 9[2) Triton!X 099[ On the following day\ the sections were washed for 1 hr inBu}er 0 and 19min in 099mM TrisÐHCl bu}er\ pH 8[4\ 099mM NaCl\ 49mM MgCl1 "Bu}er 1#and levamisole "SIGMA# 139mg:mml "an endogenous alkaline phosphatase inhibitor#[ Then theywere incubated in chromogen solution containing 3[4ml:ml of nitroblue tetrazolium "64mg:mlin 69) dimethylformamide\ Boehringer Mannheim# and 2[4ml:ml of 4!bromo!3!chloro!2!indolylphosphate "49mg:ml in 099) dimethylformamide\ Boehringer Mannheim#\ all in Bu}er 1[ Thechromogen reaction was stopped by rinsing the sections in PBT[
The sections were _xed in 3) formaldehyde in 0× PBS at 3>C overnight\ washed in distilledwater\ quickly taken through an ethanol series\ treated with xylene for 1min and mounted withDPX "BDH#[ All sections were developed for the same length of time in the color reaction\ 0 hr and39min[ Washes and incubations were performed at RT unless otherwise stated[
Controls were treated following the same protocol but omitting the probe in the hybridizationsolution[ The speci_city of the immunological detection was checked by omitting the digoxigenin!labeled probes in the hybridization solution[ No staining was observed under either of theseconditions[
MICROPHOTOGRAPHY
Microphotographs were made using a Wild photoautomat MPS 34 with a Wild MPS 40S cameraconnected to a Leitz DMRBE microscope[ For black!and!white pictures\ Kodak Technical Pan _lmat 099 ASA and AgfaPan 14 were used[
Chemicals
Unless otherwise indicated\ chemicals were of at least analytical grade and obtained from a localsupplier[
RESULTS
We studied previously the GABAergic expression in the parahippocampal region of the developingchick telencephalon[11 The present study describes a development of GAD expression which iscoincident with this expression[ As outlined in Fig[ 0\ the areas where GAD mRNA is expressedgenerally overlap with those of the protein staining[ This is particularly so in the case of GAD56mRNA[
We con_rm the neuronal identity of GABAergic cells\ i[e[ cells staining positive for GABAwere labeled simultaneously for the neuronal marker NSE "data not shown#[ The NSE!positiveGABAergic cells were observed from E00 onwards[ Earlier GABA!immunoreactive cells did notdisplay this neuronal marker[ Immunostaining with the 0339 antiserum\ which recognizes both

GAD in the developing chick brain 020
Fig[ 0[ Drawings made from microscopical observations of di}erent embryonic stages of the developingchicken telencephalon[ Coronal sections were made at the mid!third part of the brain\ which include theprehippocampal and parahippocampal areas[20 Size relationships are according to scale[ The expressionpatterns of GAD protein as detected by immunostaining and GAD mRNAs by in situ hybridization areoutlined[ Older embryos exhibit larger areas of positive staining for the investigated parameters[ Further!
more\ in some regions\ the parameters are superimposed\ rendering a dark color[
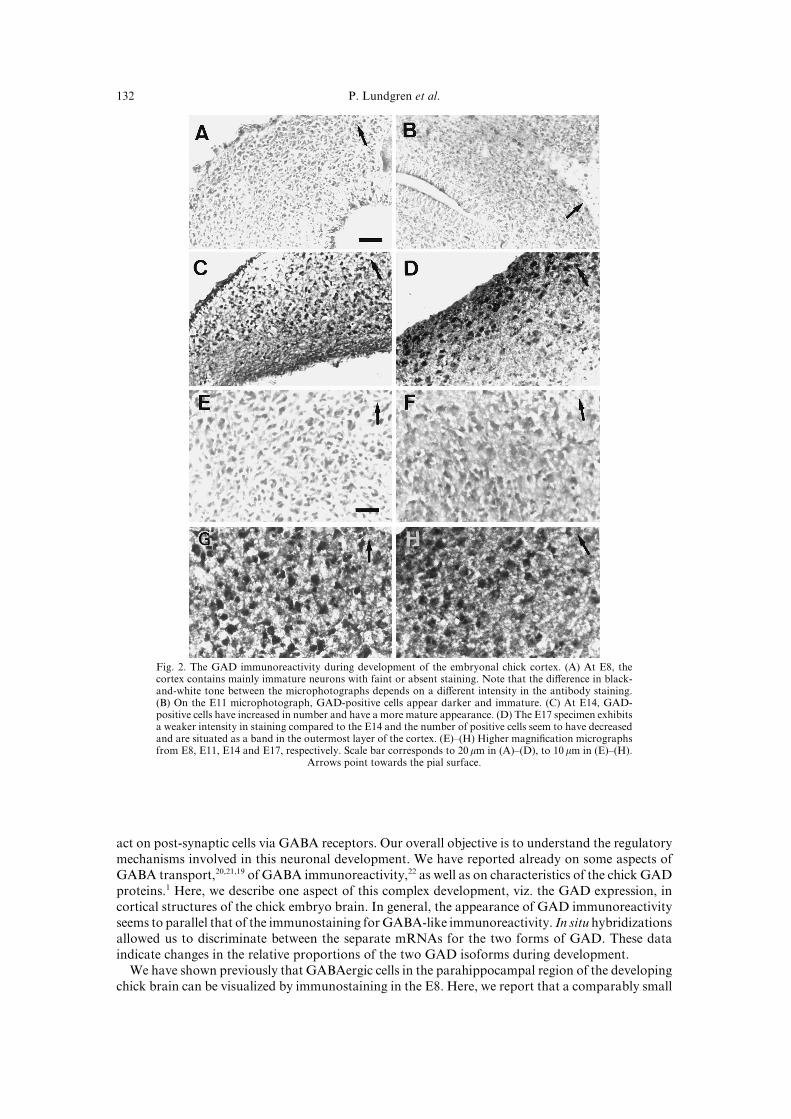
GAD54 and GAD56\ indicates a prominent increase in number of GAD!positive cells from E7 toE06\ with a possible peak at E03[ In the earlier stages\ positive cells are found in the area betweenthe ventricle and the pial surface\ whereas by E06\ the positive cells are more restricted to the surfacelayers "Fig[ 1#[ The intensity of the staining is increasing with development[ This is observed as ahigher degree of contrast between negative and positive cells as illustrated in Fig[ 1[ Furthermore\ thematurity of positive cells becomes more pronounced\ as judged from the appearance of developingprocesses[
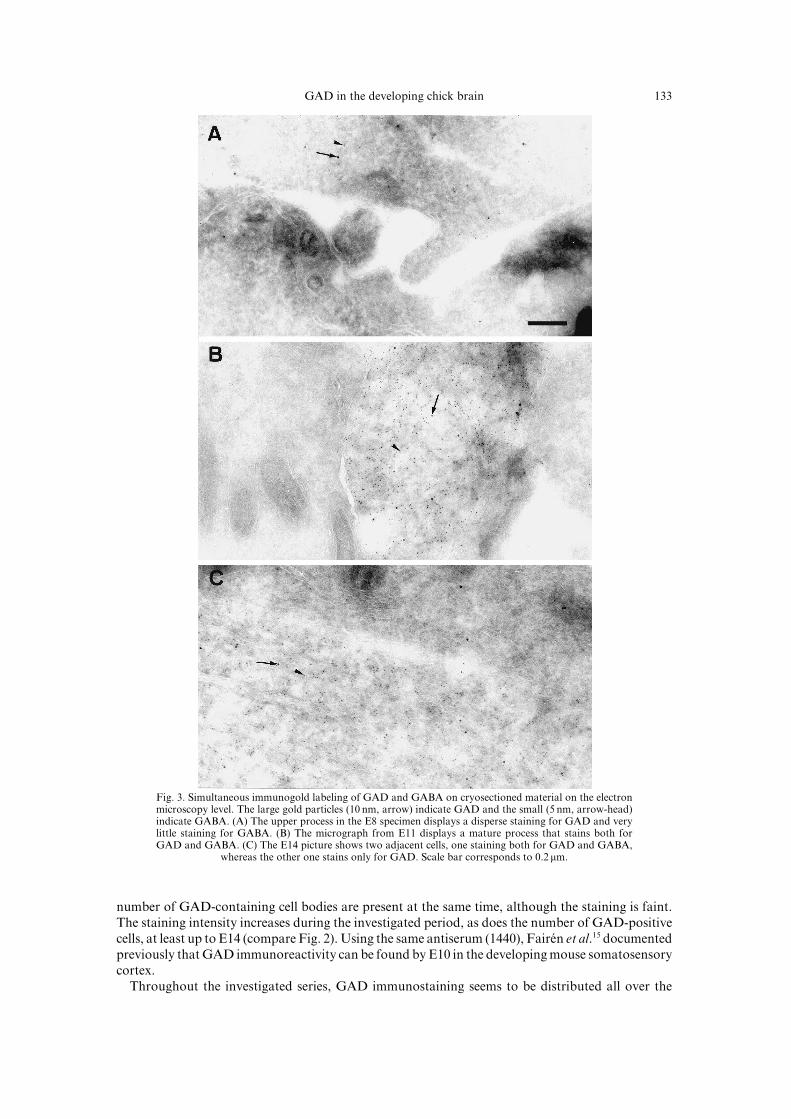
Most cells are clearly positive when co!stained for GABA and GAD on the electron microscopylevel "Fig[ 2#[ In some cases\ though\ GABA immunoreactivity is not distinguishable from thebackground in the controls\ suggesting that GAD!staining is much more prominent in those cellsðFig[ 2"C#Ł[ It should be stressed that the specimens are investigated by means of cryo!sectioningthe material without previous embedding and contrasting\ which makes the tissue appear faint andwith low contrast[
Both GAD54 and GAD56 mRNA are visualized by in situ hybridizations from E7 onwards "Fig[3#[ The staining intensity as well as the number of labeled cells increase during the investigatedperiod\ which seems to be concomitant with the GAD protein expression[ The GAD56 mRNA issomewhat earlier in prominence and also better correlated with the staining of the protein[ In theparahippocampal area by E06\ however\ GAD54 mRNA is expressed equally\ both in terms ofintensity and in number of positive cells\ as the message for GAD56[ An interesting feature is high!lighted at higher magni_cation in Fig[ 4\ where the di}erence in cellular distribution of each mRNAis shown[ The GAD54 message is appearing in the cytoplasm and to some extent in the nuclearregion\ whereas GAD56 mRNA is found predominantly associated to the nucleus[
DISCUSSION
Development of the GABAergic system involves the acquisition of synthetic capacity\ of storagefacilities\ and of the possibility to release and re!uptake GABA[ It also involves the competence to
P[ Lundgren et al[021
Fig[ 1[ The GAD immunoreactivity during development of the embryonal chick cortex[ "A# At E7\ thecortex contains mainly immature neurons with faint or absent staining[ Note that the di}erence in black!and!white tone between the microphotographs depends on a di}erent intensity in the antibody staining["B# On the E00 microphotograph\ GAD!positive cells appear darker and immature[ "C# At E03\ GAD!positive cells have increased in number and have a more mature appearance[ "D# The E06 specimen exhibitsa weaker intensity in staining compared to the E03 and the number of positive cells seem to have decreasedand are situated as a band in the outermost layer of the cortex[ "E#Ð"H# Higher magni_cation micrographsfrom E7\ E00\ E03 and E06\ respectively[ Scale bar corresponds to 19 mm in "A#Ð"D#\ to 09 mm in "E#Ð"H#[
Arrows point towards the pial surface[
act on post!synaptic cells via GABA receptors[ Our overall objective is to understand the regulatorymechanisms involved in this neuronal development[ We have reported already on some aspects ofGABA transport\19\10\08 of GABA immunoreactivity\11 as well as on characteristics of the chick GADproteins[0 Here\ we describe one aspect of this complex development\ viz[ the GAD expression\ incortical structures of the chick embryo brain[ In general\ the appearance of GAD immunoreactivityseems to parallel that of the immunostaining for GABA!like immunoreactivity[ In situ hybridizationsallowed us to discriminate between the separate mRNAs for the two forms of GAD[ These dataindicate changes in the relative proportions of the two GAD isoforms during development[
We have shown previously that GABAergic cells in the parahippocampal region of the developingchick brain can be visualized by immunostaining in the E7[ Here\ we report that a comparably small
GAD in the developing chick brain 022
Fig[ 2[ Simultaneous immunogold labeling of GAD and GABA on cryosectioned material on the electronmicroscopy level[ The large gold particles "09 nm\ arrow# indicate GAD and the small "4 nm\ arrow!head#indicate GABA[ "A# The upper process in the E7 specimen displays a disperse staining for GAD and verylittle staining for GABA[ "B# The micrograph from E00 displays a mature process that stains both forGAD and GABA[ "C# The E03 picture shows two adjacent cells\ one staining both for GAD and GABA\
whereas the other one stains only for GAD[ Scale bar corresponds to 9[1 mm[
number of GAD!containing cell bodies are present at the same time\ although the staining is faint[The staining intensity increases during the investigated period\ as does the number of GAD!positivecells\ at least up to E03 "compare Fig[ 1#[ Using the same antiserum "0339#\ Faire�n et al[04 documentedpreviously that GAD immunoreactivity can be found by E09 in the developing mouse somatosensorycortex[
Throughout the investigated series\ GAD immunostaining seems to be distributed all over the

P[ Lundgren et al[023
Fig[ 3[ In situ hybridization for GAD54 mRNA at the left and for GAD56 at the right panel from E7 toE06[ "A# and "B#�E7[ "C# and "D#�E00[ "E# and "F#�E03[ "G# and "H#�E06[ A weak backgroundstaining and relatively thick "19 mm# cryosections renders somewhat unfocused images from the earlierstages[ At E7\ the hybridization for GAD54 is weak and appears _rst in the outermost layer of theparaventricular zone and in the lateral area of the cortex[ The staining intensity is higher for GAD56mRNA[ At E00\ the number of cells that hybridize for both mRNAs have increased[ There is still a highernumber of cells that express GAD56 mRNA[ By E03\ the cells have grown in size and the staining is moreintense[ At E06\ GAD54 mRNA!containing cells are labeled more intensely and seem to appear at a highernumber than GAD56 mRNA!expressing cells[ Scale bar corresponds to 49 mm[ Arrows point towards the
pial surface[
cell body and only to some extent appearing in a punctate fashion[ As inferred from the NSEstaining as well as from the summary in Table 0\ the neuronal phenotype is forming graduallyduring this period[ The morphological development includes formation of processes and of synapticcontacts[11 One might have expected that GAD!staining would be present to a greater extent inneurites rather than in somata\ at the later stages of the development[ The staining pattern observedby us can\ however\ also be attributed to the fact that the antiserum used "0339#16 does notdiscriminate between the two forms\ GAD54 and GAD56\ of the enzyme[ The possible di}erencesin subcellular localization between the two forms\ which has been reported by Esclapez et al[03 thuscannot be corroborated by us[
The pattern of GAD immunoreactivity seen here corresponds well with the staining of GABA!containing cells in the parahippocampal area previously noted by us[11 It is in accordance also witha study on GABA and GAD localization in the adult pigeon forebrain by Veenman and Reiner\20
where the corresponding region exhibited moderate to dense GABA!staining and GAD presence[These authors used three di}erent anti!GAD antisera\ including the 0339 used by us[ All three

GAD in the developing chick brain 024
Fig[ 4[ High!power magni_cation microphotographs showing the di}erence in intracellular distribution ofthe di}erent GAD mRNAs after in situ hybridization at E06[ These di}erences are apparent throughoutthe investigated embryonic period[ "A# The GAD54 mRNA hybridization\ revealing that the mRNA isdistributed all over the cell[ "B# An adjacent section hybridized against GAD56 mRNA\ where an intensestaining in or near the nucleus can be seen[ Scale bar corresponds to 09 mm in "A# and "B# The insert showsthe E06 GAD56 mRNA in situ hybridization at higher magni_cation "scale bar�4 mm#[ Arrows point
towards the pial surface[
antisera displayed virtually indistinguishable labeling patterns[ The time!frame for GAD appearancein the developing chick nervous system probably is region!speci_c[ Thus\ it has been shown thatGABA!positive cells can be detected as early as by E3 in certain cranial motor nerves3\ which isconcomitant with the _rst GABA immunoreactivity in the ventral spinal cord[1 Assuming thatGABA is synthesized by GAD\ the protein is at least present that early[ We have ourselves0 observedGAD!activity as early as by E2 in the forebrain[ However\ our observations in the above study arenot localized to speci_c regions as is the immunostaining[
The staining intensity for NSE is only consistently apparent from E03 onwards\ although somesporadic staining was observed already by E00[ These observations may mean that the GABApresence is preceding the mature NSE!positive neuronal phenotype[ The function of this earlyGABA content remains to be elucidated[ It has been noted in several studies that GABA expressionin a number of CNS regions can be transient during embryonic development[1\3\6\11\12 Several expla!nations may account for the transient expression[ An interesting hypothesis is that GABA mayexert other e}ects than that of an inhibitory transmitter[ Thus\ it has been suggested that GABAcan act trophically during neuronal development[1\5\29 Also\ in post!hatch chickens\ GABA may\ atleast locally\ perform depolarizing actions\ as shown by Hyson et al[06 who investigated GABAergicinput into neurons in brainstem auditory nuclei[ These authors also indirectly provided evidence fora GABA!receptor subtype functionally di}erent from the common GABAA and GABAB receptors[
Furthermore\ the immuno!electron micrographs presented in this paper display only GAD!
Table 0[ A summary of the expression of investigated parameters of GABA!ergic di}erentiation in the developing chick cortex
GAD54 mRNA ¦ ¦¦ ¦¦¦ ¦¦¦GAD56 mRNA ¦¦ ¦¦¦ ¦¦¦¦ ¦¦¦GAD "0339# ¦¦ ¦¦¦ ¦¦¦¦ ¦¦¦GABA ¦ ¦¦¦¦¦ ¦¦¦ n[dNSE� − ¦� ¦¦ ¦¦
E7 E00 E03 E06
Staining is expressed arbitrarily as {{¦||[ Absence of staining is given by{{−||^ n[ d[ � not determined[
�NSE expression in E00 has been investigated in very few specimens[

P[ Lundgren et al[025
staining cells\ as well as cells co!stained for GABA and GAD[ No quantitation of the relativedistribution of each cell type has been made[ These di}erences could indicate that cells exhibitingsolely GAD!staining have acquired the protein recently\ and have not yet started GABA production[It may also be a consequence of a selective expression of a GAD form\ which is not enzymaticallyactive[ The 0339 antiserum employed in the study does not allow speci_c identi_cation of therespective GAD forms[ This means that we cannot ascertain any existing predominance of either ofthe isozymes[ It has been shown that GAD immunoreactivity can be transient and appear with noGABA presence in the chick[3
Each form of the GAD gene is expressed on the transcriptional level during the investigatedperiod according to the present in situ hybridizations "Figs 3 and 4#[ Using these probes for Northernblot analysis of comparable material\ they hybridize to bands corresponding to transcript sizes of2[8 kb "GAD54# and 4[5 kb "GAD56#[0 The expression patterns indicate that the labeling intensityof GAD56 mRNA somewhat precedes that of GAD54[ In one of the few studies performed on thissubject during development\ Ma et al[13 could show that the transcripts were expressed in a parallelfashion in the rat spinal cord during postnatal development[ Similar results also were reported byothers\18 who found in the embryonic rat spinal cord the transcripts for GAD54\ GAD56 and analternatively spliced form of the latter as early as by E00[ Adult levels are characterized by regionaldi}erences in expression pattern\03\02 although many structures were labeled by probes for eachmRNA[ We interpret the present data as a mostly similar spatial distribution of each message[
An outline of the development of the GABAergic neuron is presented in Table 0[ When comparingthe time!wise appearance of NSE\ GAD and GABA\ this indicates that the mature neuronalphenotype is arrived at later than the GABA synthesis[ Others18 have reported\ however\ that themRNAs for GAD\ GABA and GABA!receptors develop in parallel with another neuronal marker"neuro_lament#[ An interesting picture emerges during comparison of GABA immunoreactivitywith GAD mRNA expression[ In the earlier stages\ GAD56 mRNA is expressed consistently in thesame regions as GABA[ The GAD immunoreactivity also is observed in this vicinity[ This suggeststhat earlier GABA production is primarily a function of translating the GAD56 message into afunctional protein[ Later\ the GAD54 message and the GABA staining are overlapping\ which mayindicate a switch to a more mature:adult expression[ It has been suggested that subcellular local!ization of GAD54 is more con_ned to terminals than GAD56\ which is found in the cell body[ Thisindicates that adult transmitter production is a function of GAD54 activity[02 Whether embryonicGAD roles are similar remains to be investigated[
Acknowled`ements*The _nancial support of the Natural Science Research Council "grant (B!AA:BU 09573!299#\ theHierta!Retzius Foundation and the J[ C[ Kempes Minne Foundation is greatly appreciated[ The cDNAs encoding GAD54and GAD56 were kindly provided by Dr Tillakaratne\ UCLA^ 0339 antiserum against GAD was provided by NINDS\Bethesda^ and 889 antiserum against GABA was a kind gift from Dr O[!P[ Ottersen\ University of Oslo[
REFERENCES
0[ A� hman A[!K[\ Wa�gberg F[ and Mattsson M[!O[ "0885# Two glutamate decarboxylase forms corresponding to themammalian GAD54 and GAD56 are expressed during development of the chick telencephalon[ Eur[ J[ Neurosci[ 7\
1000Ð1006[1[ Antal M[\ Berki A[ C[\ Horvath L[ and O|Donovan M[ J[ "0883# Developmental changes in the distribution of gamma!
aminobutyric acid!immunoreactive neurons in the embryonic chick lumbosacral spinal cord[ J[ Comp[ Neurol[ 232\ 117Ð125[
2[ Balcar V[ J[\ Zetzsche T[ and Wol} J[ R[ "0881# Glutamate decarboxylase in developing rat neocortex] does it correlatewith the di}erentiation of GABAergic neurons and synapses< Neurochem[ Res[ 06\ 142Ð159[
3[ von Bartheld C[ S[ and Rubel E[ W[ "0878# Transient GABA immunoreactivity in cranial nerves of the chick embryo[ JComp[ Neurol[ 175\ 345Ð360[
4[ Bu D[!F[\ Erlander M[ G[\ Hitz B[ C[\ Tillakaratne N[ J[ K[\ Kaufman D[ L[\ Wagner!McPherson C[ B[\ Evans G[ A[and Tobin A[ J[ "0881# Two human glutamate decarboxylases\ 54 kDa GAD and 56 kDa GAD\ are each encoded by asingle gene[ Proc[ Natl Acad[ Sci[ USA 78\ 1004Ð1008[
5[ Cherubini E[\ Gaiarsa J[ L[ and Ben!Ari Y[ "0880# GABA] an excitatory transmitter in early postnatal life[ TrendsNeurosci[ 03\ 404Ð408[
6[ Chun J[ J[ M[ and Shatz C[ "0878# The earliest!generated neurons of the cat cerebral cortex] characterization by MAP1and neurotransmitter immunohistochemistry during fetal life[ J[ Neurosci[ 8\ 0537Ð0556[
7[ Code R[ A[\ Burd G[ D[ and Rubel E[ W[ "0878# Development of GABA immunoreactivity in brainstem auditory nucleiof the chick] ontogeny of gradients in terminal staining[ J[ Comp[ Neurol[ 173\ 493Ð407[
8[ Coyle J[ T[ and Enna S[ J[ "0865# Neurochemical aspect of the ontogenesis of GABAergic neurons in the rat brain[ BrainRes[ 000\ 008Ð022[

GAD in the developing chick brain 026
09[ Del Rio J[ A[\ Soriano E[ and Ferrer I[ "0881# Development of GABA immunoreactivity in the neocortex of the mouse[J[ Comp[ Neurol[ 215\ 490Ð415[
00[ van Eden C[ G[\ Hrzljak L[\ Voorn P[\ Uylings H[ B[ M[ "0878# Prenatal development of GABAergic neurons in theneocortex of the rat[ J[ Comp[ Neurol[ 178\ 102Ð116[
01[ Erlander M[ G[\ Tillakaratne N[ J[ K[\ Feldblum S[\ Patel N[ and Tobin A[ J[ "0880# Two genes encode distinct glutamatedecarboxylases[ Neuron 6\ 80Ð099[
02[ Esclapez M[\ Tillakaratne N[ J[ K[\ Kaufman D[ L[\ Tobin A[ J[ and Houser C[ R[ "0883# Comparative localization oftwo forms of glutamic acid decarboxylase and their mRNAs in rat brain supports the concept of functional di}erencesbetween the two forms[ J[ Neurosci[ 03\ 0723Ð0744[
03[ Esclapez M[\ Tillakaratne N[ J[ K[\ Tobin A[ J[ and Houser C[ R[ "0882# Comparative localization of mRNAs encodingtwo forms of glutamic acid decarboxylase with nonradioactive in situ hybridization methods[ J[ Comp[ Neurol[ 220\ 228Ð251[
04[ Faire�n A[\ Cobas A[ and Fonseca M[ "0875# Times of generation of glutamic acid decarboxylase immunoreactive neuronsin mouse somatosensory cortex[ J[ Comp[ Neurol[ 140\ 56Ð72[
05[ van Gelder N[ M[ and Be�langer F[ "0877# The establishment of essential and metabolically derived amino acid pro_lesin developing chick embryo organs[ Neurochem[ Res[ 02\ 588Ð695[
06[ Hyson R[ L[\ Reyes A[ D[ and Rubel E[ W[ "0884# A depolarizing inhibitory response to GABA in brainstem auditoryneurons of the chick[ Brain Res[ 566\ 006Ð015[
07[ Lauder J[ M[\ Han V[ K[ M[\ Henderson P[\ Verdoorn T[ and Towle A[ C[ "0875# Prenatal ontogeny of the GABAergicsystem in the rat brain] an immunocytochemical study[ Neuroscience 1\ 354Ð382[
08[ Lewin L[\ Mattsson M[!O[\ Grahn B[ and Sellstro�m A� [ "0883# Inhibition of SKF 78865!A of the g!aminobutyric acidrelease from primary neuronal chick cultures[ Acta Phys[ Scand[ 041\ 062Ð068[
19[ Lewin L[\ Mattsson M[!O[\ Rassin D[ K[ and Sellstro�m A� [ "0881# On the activity of g!aminobutyric acid and glutamatetransporters in chick embryonic neurons and rat synaptosomes[ Neurochem[ Res[ 06\ 222Ð226[
10[ Lewin L[\ Mattsson M[!O[ and Sellstro�m A� [ "0881# Inhibition of transporter!mediated g!aminobutyric acid "GABA#release by SKF 78865!A\ a GABA uptake inhibitor\ studied in a primary neuronal culture from chicken[ Neurochem[Res[ 06\ 466Ð473[
11[ Lundgren P[\ Mattsson M[!O[\ Johansson L[\ Ottersen O[!P[ and Sellstro�m A� [ "0884# Morphological and GABA!immunoreactive development of the embryonic chick telencephalon[ Intl J[ Dev[ Neurosci[ 02\ 352Ð361[
12[ Ma W[\ Behar T[ and Barker J[ L[ "0881# Transient expression of GABA immunoreactivity in the developing rat spinalcord[ J[ Comp[ Neurol[ 214\ 160Ð189[
13[ Ma W[\ Behar T[\ Chang L[ and Barker J[ L[ "0883# Transient increase in expression of GAD54 and GAD56 mRNAsduring postnatal development of rat spinal cord[ J[ Comp[ Neurol[ 235\ 040Ð059[
14[ Ma W[\ Behar T[\ Scha}ner A[ E[\ Smith S[ and Barker J[ L[ "0880# Parallel expression of GABAa receptor and GABAin embryonic rat subcortical brain[ Soc[ Neurosci[ Abstr[ 06\ 2991[
15[ Martin D[ L[ and Rimvall K[ "0882# Regulation of g!aminobutyric acid synthesis in the brain[ J[ Neurochem[ 59\ 284Ð396[
16[ Oertel W[ H[\ Schmechel D[ E[\ Tappaz M[ L[ and Kopin I[ J[ "0870# Production of a speci_c antiserum to rat brainglutamic acid decarboxylase by injection of an antigenÐantibody complex[ Neuroscience 5\ 1578Ð1699[
17[ Sambrook J[\ Fritsch E[ F[ and Maniatis T[ "0878# Molecular Clonin` * A Laboratory Manual\ 1nd edn[ Cold SpringHarbor Laboratory Press\ Cold Spring Harbor\ New York[
18[ Somogyi R[\ Wen X[\ Ma W[ and Barker J[ L[ "0884# Developmental kinetics of GAD family mRNAs parallelneurogenesis in the rat spinal cord[ J[ Neurosci[ 04\ 1464Ð1480[
29[ Spoerri P[ E[ and Wol} J[ R[ "0870# E}ect of GABA!administration on murine neuroblastoma cells in culture[ I[Increased membrane dynamics and formation of specialized contacts[ Cell Tissue Res[ 107\ 456Ð468[
20[ Veenman C[ L[ and Reiner A[ "0883# The distribution of GABA!containing perikarya\ _bers\ and terminals in theforebrain and midbrain of pigeons\ with particular reference to the basal ganglia and its projection targets[ J Comp[Neurol[ 228\ 198Ð149[
21[ Walberg F[ and Ottersen O[ P[ "0881# Neuroactive amino acids in the area postrema[ An immunocytochemical investi!gation in rat with some observations in cat and monkey "Macaca fascicularis#[ Anat[ Embryol[ 074\ 418Ð434[
Copyright © 2022 FDOKUMEN




















