
Expression of neuron-associated tumor necrosis factor alpha in the brain is increased during...
10
Expression of Neuron-Associated Tumor Necrosis Factor Alpha in the Brain Is Increased During Persistent Pain William C. Covey, Ph.D., Tracey A. Ignatowski, Ph.D., Amy E. Renauld, Ph.D., Paul R. Knight, M.D., Ph.D., Nader D. Nader, M.D., Ph.D., and Robert N. Spengler, Ph.D. Background and Objectives: Evidence implicates the pleiotropic cytokine tumor necrosis factor alpha (TNF) in the pathogenesis of persistent pain. The present study employs a chronic constriction injury (CCI) model of neuropathic pain to examine TNF production in the central nervous system (CNS) and in the periphery in this pain model. Methods: CCI-induced hyperalgesia is assessed by measuring the nociceptive threshold using the hot- plate test. The development of hyperalgesia is correlated to levels of TNF by assessing: bioactive TNF in homogenates of sciatic nerves, cervical spinal cord, thoracolumbar spinal cord, as well as in plasma using the WEHI-13 variant cytotoxicity bioassay; and mRNA for TNF in sections of locus coeruleus by in situ hybridization. Results: We have previously demonstrated that TNF bioactivity in the region of the brainstem contain- ing the locus coeruleus is increased concurrent with the development of hyperalgesia, returning to baseline values by day 14, when hyperalgesia has ceased. Constitutive levels of TNF are demonstrated in the plasma, sciatic nerves, and cervical and thoracolumbar spinal cord of control rats, sham-operated rats, and rats undergoing CCI. Levels of TNF are significantly elevated in the injured sciatic nerve by day 8 postligature placement, concurrent with maximal hyperalgesia, and remain elevated when hyperalgesia has abated at day 14 postligature placement. Additionally, TNF activity is increased in the thoracolumbar region of the spinal cord by day 4 postligature placement and remains elevated during hyperalgesia (day 8), as well as after hyperalgesia has dissipated (day 14). The increase in TNF expression is specific to discrete regions of the CNS, rather than being the result of a systemic inflammatory response, since TNF bioactivity in plasma is, in fact, decreased in rats undergoing CCI. Additionally, accumulation of mRNA specific for TNF is significantly increased in neurons within a region of the brain containing the locus coeruleus at days 2, 8, and 14 postligature placement, contemporaneous with the development of hyperalgesia. Conclusions: The increases in TNF within regions of the brain and spinal cord that are associated with adrenergic neuron function, as well as with modulation of pain perception, and the time course and distribution of the increases in TNF accumulation support a neuromodulatory role for TNF within the CNS in the development and maintenance of neuropathic pain. Reg Anesth Pain Med 2002;27:357-366. Key Words: Adrenergic, Brain, Central nervous system, Pain, Tumor necrosis factor, Chronic constriction injury. From the Departments of Pathology (W.C.C., T.A.I., R.N.S.) and Anesthesiology (T.A.I., A.E.R., P.R.K., N.D.N., R.N.S), State University of New York at Buffalo, School of Medicine and Biomedical Sciences, Buffalo, New York. Accepted for publication December 11, 2001. Supported by the Charles A. Dana Foundation (R.N.S.), the National Institutes of Health Grant No. NS41352 (R.N.S.), the Paralyzed Veterans of America #RFA008 (R.N.S.), the NARSAD Young Investigator’s Award Grant (T.A.I.). This work is being submitted to the Graduate School of the State University of New York at Buffalo in partial fulfillment of the requirements for the Ph.D. degree in Pathology. Reprint requests: Robert N. Spengler, Ph.D., Department of Pathology, State University of New York at Buffalo, 204 Farber Hall, 3435 Main St, Buffalo, NY 14214. E-mail: [email protected] © 2002 by the American Society of Regional Anesthesia and Pain Medicine. 1098-7339/02/2704-0004$35.00/0 doi:10.1053/rapm.2002.31930 Regional Anesthesia and Pain Medicine, Vol 27, No 4 (July–August), 2002: pp 357–366 357
Transcript of Expression of neuron-associated tumor necrosis factor alpha in the brain is increased during...

Expression of Neuron-Associated Tumor NecrosisFactor Alpha in the Brain Is IncreasedDuring Persistent Pain
William C. Covey, Ph.D., Tracey A. Ignatowski, Ph.D., Amy E. Renauld, Ph.D.,Paul R. Knight, M.D., Ph.D., Nader D. Nader, M.D., Ph.D.,and Robert N. Spengler, Ph.D.
Background and Objectives: Evidence implicates the pleiotropic cytokine tumor necrosis factor alpha (TNF�)in the pathogenesis of persistent pain. The present study employs a chronic constriction injury (CCI) model ofneuropathic pain to examine TNF� production in the central nervous system (CNS) and in the periphery in thispain model.
Methods: CCI-induced hyperalgesia is assessed by measuring the nociceptive threshold using the hot-plate test. The development of hyperalgesia is correlated to levels of TNF� by assessing: bioactive TNF� inhomogenates of sciatic nerves, cervical spinal cord, thoracolumbar spinal cord, as well as in plasma usingthe WEHI-13 variant cytotoxicity bioassay; and mRNA for TNF� in sections of locus coeruleus by in situhybridization.
Results: We have previously demonstrated that TNF� bioactivity in the region of the brainstem contain-ing the locus coeruleus is increased concurrent with the development of hyperalgesia, returning to baselinevalues by day 14, when hyperalgesia has ceased. Constitutive levels of TNF� are demonstrated in theplasma, sciatic nerves, and cervical and thoracolumbar spinal cord of control rats, sham-operated rats, andrats undergoing CCI. Levels of TNF� are significantly elevated in the injured sciatic nerve by day 8postligature placement, concurrent with maximal hyperalgesia, and remain elevated when hyperalgesia hasabated at day 14 postligature placement. Additionally, TNF� activity is increased in the thoracolumbarregion of the spinal cord by day 4 postligature placement and remains elevated during hyperalgesia (day 8),as well as after hyperalgesia has dissipated (day 14). The increase in TNF� expression is specific to discreteregions of the CNS, rather than being the result of a systemic inflammatory response, since TNF� bioactivityin plasma is, in fact, decreased in rats undergoing CCI. Additionally, accumulation of mRNA specific forTNF� is significantly increased in neurons within a region of the brain containing the locus coeruleus atdays 2, 8, and 14 postligature placement, contemporaneous with the development of hyperalgesia.
Conclusions: The increases in TNF� within regions of the brain and spinal cord that are associated withadrenergic neuron function, as well as with modulation of pain perception, and the time course and distributionof the increases in TNF� accumulation support a neuromodulatory role for TNF� within the CNS in thedevelopment and maintenance of neuropathic pain. Reg Anesth Pain Med 2002;27:357-366.
Key Words: Adrenergic, Brain, Central nervous system, Pain, Tumor necrosis factor, Chronic constrictioninjury.
From the Departments of Pathology (W.C.C., T.A.I., R.N.S.) and Anesthesiology (T.A.I., A.E.R., P.R.K., N.D.N., R.N.S), StateUniversity of New York at Buffalo, School of Medicine and Biomedical Sciences, Buffalo, New York.Accepted for publication December 11, 2001.Supported by the Charles A. Dana Foundation (R.N.S.), the National Institutes of Health Grant No. NS41352 (R.N.S.), the Paralyzed
Veterans of America #RFA008 (R.N.S.), the NARSAD Young Investigator’s Award Grant (T.A.I.).This work is being submitted to the Graduate School of the State University of New York at Buffalo in partial fulfillment of the
requirements for the Ph.D. degree in Pathology.Reprint requests: Robert N. Spengler, Ph.D., Department of Pathology, State University of New York at Buffalo, 204 Farber Hall, 3435
Main St, Buffalo, NY 14214. E-mail: [email protected]© 2002 by the American Society of Regional Anesthesia and Pain Medicine.1098-7339/02/2704-0004$35.00/0doi:10.1053/rapm.2002.31930
Regional Anesthesia and Pain Medicine, Vol 27, No 4 (July–August), 2002: pp 357–366 357

Considerable evidence supports the involve-ment of the pleiotropic cytokine tumor necro-
sis factor alpha (TNF�) in the pathogenesis of per-sistent pain. Increases in TNF� are believed tomediate the enhanced responsiveness to painfulstimuli (hyperalgesia) observed following adminis-tration of substances, such as lipopolysaccharide(LPS), that stimulate cytokine production.1 Fur-thermore, intracerebroventricular (icv) microinjec-tion of recombinant TNF� produces hyperalgesia inrats, implicating a site of action within the centralnervous system (CNS) for the nociceptive effects ofTNF�.2 Conversely, systemically administered TNF�binding protein prevents the hyperalgesia associ-ated with LPS administration,1 and preoperativeadministration of thalidomide, an inhibitor of TNF�synthesis, prevents the development of hyperalge-sia in rats undergoing experimental mononeu-ropathy.3
Our laboratory has previously demonstrated in-creased TNF� accumulation in the brain during per-sistent pain,4 as well as a requirement for TNF� inthe brain for the development of hyperalgesia.5
Levels of biologically active TNF� are significantlyelevated in the locus coeruleus and hippocampus ofanimals undergoing chronic constriction injury(CCI) during the manifestation of hyperalgesia.4
Additionally, icv microinfusion of recombinant ratTNF� into the right lateral cerebral ventricle (prox-imal to the hippocampus) produces hyperalgesia innormal rats and exacerbates hyperalgesia in ratsundergoing CCI. Conversely, icv microinfusion ofpolyclonal anti-TNF� antibodies abolishes hyperal-gesia, and, in fact, does produce hypoalgesia insome rats undergoing CCI.5
Bilateral increases in TNF� immunoreactivity aredemonstrated in the dorsal horn of the lumbar spi-nal cord concurrent with the manifestation of ther-mal hyperalgesia characteristic of this pain model.6
This increased TNF� immunostaining is restricted tothe lumbar segments (area of sciatic innervation) ofthe spinal cord and thus suggests a regional processduring CCI, rather than being a generalized inflam-matory process within the gray matter, such as thatobserved following radiation-induced injury to thespinal cord.7 Similarly, TNF� immunoreactivity isincreased in various cell types in the injured sciaticnerve of animals undergoing CCI at a time thatcorrelates with the appearance of hyperalgesia.3
Initiating thalidomide administration prior to sur-gery attenuates this increase in TNF� immunostain-ing and reduces the development of hyperalgesia.However, this treatment is ineffective when initi-ated after hyperalgesia has manifested.3 The directinfluence of TNF� on increased nerve activity canbe demonstrated by applying TNF� onto the sciatic
nerve, which increases spontaneous firing of noci-ceptive afferents in a dose-dependent manner.8
Furthermore, endoneural injection of TNF� intothe sciatic nerve produces allodynia and thermalhyperalgesia similar to that observed following ex-perimental nerve injury.9 These data support acausative role for TNF� in the pathogenesis of per-sistent pain. The purpose of the present study is tocorrelate the alterations in levels of TNF� in theCNS, as well as at the site of injury with the devel-opment of hyperalgesia during CCI. We postulatethat increases in TNF� production by neuronswithin specific regions of the brain during the de-velopment of a central component to neuropathicpain play an important role in the pathogenesis ofpersistent pain.
Methods
Chronic Constriction Injury Model ofNeuropathic Pain
This study was approved by the Institutional An-imal Care and Use Committee (IACUC). MaleSprague-Dawley rats (200 to 250 g, Harlan SpragueDawley, Indianapolis, IN) were used in all experi-ments. Rats were maintained in pathogen-freequarters and given food and water ad libitum. Ev-ery measure was taken to ensure that pain anddiscomfort were limited to that which is unavoid-able in the conduct of scientifically valuable re-search. Animals were anesthetized with ketamine �xylazine (60 mg/kg � 3 mg/kg, intraperitoneal [IP]).The area of incision was shaved and washed withBetadine, followed by 70% ethanol. Sterile proce-dures were observed for the surgery, according toIACUC guidelines. The sciatic nerve was exposed uni-laterally at the midthigh level, and 4 ligatures (chro-mic gut 4.0) were placed around the nerve, 1 mmapart. Ligatures were tied loosely. In sham proce-dures, the nerve is exposed, but no ligatures areplaced.
Quantitation of Hyperalgesia in Operated Rats.Based on our previous findings that TNF� produc-tion in the brain plays a pivotal role in the devel-opment of hyperalgesia, the present investigationwas undertaken to establish that TNF� expressionin neurons is increased contemporaneous with thedevelopment of hyperalgesia during persistent pain.Therefore, these results are presented similarly.5 Atvarious time points (preoperative day 2 throughpostoperative day 14), the thermal nociceptivethreshold was measured using the hot-plate test(58°C).10 Three measurements of paw-withdrawallatency were taken for each rat, at 4-minute inter-vals, as an index of hyperalgesia. Measurementstaken on the 2 consecutive days prior to surgery
358 Regional Anesthesia and Pain Medicine Vol. 27 No. 4 July–August 2002

and on the day of surgery (day 0) are averaged andused as the baseline value for withdrawal latency.Changes in nociceptive thresholds (seconds) are ex-pressed as the percentage of the Maximal PossibleResponse (% MPR) using the equation: % MPR �[(experimental latency – mean basal latency)/(maximal latency – mean basal latency)] � 100.10
Maximum latency is defined as 15 seconds, afterwhich time animals are removed from the hot platein order to prevent tissue damage. Preoperativehind paw-withdrawal latencies (mean basal laten-cies) are subtracted from those taken postopera-tively (experimental latencies) in order to generatea “difference score” that is used as an index ofhyperalgesia.
TNF� Analysis
Animals were killed by decapitation, and theplasma, sciatic nerves, cervical spinal cord (C2-C5segments), and thoracolumbar spinal cord (T7-L6segments) were immediately harvested for bioas-says. All tissue sections were weighed and thenhomogenized in 3 mL RPMI (GIBCO, Grand Island,NY) plus 1% glutamine (GIBCO). Sample homog-enates and blood plasma samples were centrifugedat 13,500 � g for 20 minutes at 4°C. Samples wereassayed for TNF� content using the WEHI-13 vari-ant cytotoxicity bioassay, as previously described.11
Briefly, WEHI-13 variant cells (5 � 105 cells/mL)suspended in culture medium, consisting of 1%glutamine and 10% fetal bovine serum plus 0.5�g/mL Actinomycin-D (Calbiochem, La Jolla, CA)were incubated with 100 �L of sample supernatantin 96-well microtiter plates. Recombinant humanTNF� (Genzyme, Cambridge, MA) in serial dilu-tions ranging from 10,000 pg/mL to 0.01 pg/mL isused as an internal standard. Polyclonal antiserumagainst murine TNF� (1:1,000) was used to deter-mine specificity. The level of TNF� in each sample isquantitated by measuring the amount of TNF�-induced cell lysis, proportional to a color change,using a Titertek Autoreader (Bio-Tek Instruments,Winooski, VT) at 540 nm. Levels of TNF� in thesamples are determined using the regression of theabsorbances of the internal standards.
In Situ Hybridization
In situ hybridization was performed using a mod-ification of a method by Kunkel et al.12 The brain-stem was isolated from control rats, rats undergoingCCI, and sham-operated rats and immediately snapfrozen in liquid nitrogen. This area consists of asection of brainstem 1 mm thick, which is medialand inferior to the superior cerebellar peduncle,superior to the nucleus of the fifth cranial nerve,
and contains the nucleus locus coeruleus. This re-gion was located using morphologic landmarks asdescribed by Paxinos and Watson13 (Bregma:�9.68mm, lateral: �1.5 mm, vertical: –7.5 mm). Frozentissue sections (4 �m thickness) were prepared on acryostat and mounted onto poly-L-lysine–coatedslides (Sigma, St Louis, MO). Mounted sectionswere immediately fixed in ice cold 4% paraformal-dehyde (Sigma) in 1� phosphate-buffered saline(PBS) for 15 minutes. Tissue sections were thenrinsed 3 times in ice cold 70% ethanol, and storedin 70% ethanol at 4°C. At the time of hybridization,sections were postfixed in 4% paraformaldehyde inPBS at room temperature for 10 minutes, washed in0.5� SSC at room temperature for 10 minutes, andpretreated with 5 �g/mL proteinase K (Promega,Madison, WI) in 2� SSC at 37°C for 15 minutes.Sections were rinsed with 2� SSC, and then acety-lated with freshly prepared 0.5% acetic anhydride(Sigma) in 0.1 mol/L triethanolamide (TEA)(Sigma) (pH 7.2) for 5 minutes. Sections wererinsed again in 0.5� SSC and then covered with aprehybridization buffer consisting of 20% deionizedformamide, 5� SSC, 5% dextran sulfate, 10 �g/mLtRNA, 100 �g/mL salmon sperm DNA, 1� Den-hardt’s solution (all from Sigma), and 10 mmol/Ldithiothreitol (DTT) (Boehringer Mannheim, Indi-anapolis, IN). Sections were incubated with theprehybridization buffer at 42°C for 2 hours. Anantisense 30-mer cDNA oligonucleotide probe(5�-GTC-CCC-CTT-CTC-CAG-CTG-GAA-GAC-TCC-TCC-3�) that is complementary to mRNA specificfor TNF� was labeled with 35S-deoxycytidinetriphosphate (dCTP) (NEN, Boston, MA) using aprotocol by Elner et al.14 The probe was diluted inhybridization buffer such that the final activity con-tained within the buffer was 1 � 106 bound cpm/100 �L. Each tissue section was covered with 100�L hybridization buffer and hybridized overnight at42°C. After hybridization, sections were washedtwice with 2� SSC at room temperature for 10minutes to remove nonadherent radioactivity, thenwashed with 1� SSC at 42°C for 60 minutes. Sec-tions were dehydrated in graded concentrations ofethanol (50%, 70%, 95%) with 0.3% ammoniumacetate [Sigma]) and then air dried. Slides werecoated in NTB-2 emulsion (Eastman Kodak, Roch-ester, NY), dried, placed in lightproof boxes, andexposed for 10 weeks at 4°C. Slides were developedwith Kodak D-19 developer (1:1 ddH20), and fixedwith Kodak Rapid Fix Solution A. Tissue sectionswere counterstained with hematoxylin and eosin.Controls run in parallel with experimental sectionsinclude sections hybridized with unlabeled oligonu-cleotide probe, with a complementary sense cDNAoligonucleotide probe, or with an oligonucleotide
TNF in the Brain During Persistent Pain • Covey et al. 359
probe to detect mRNA specific to �-actin mRNA.Grains over 40 neurons from each section werecounted using a Leitz microscope (1,000x magnifi-cation). The neuron phenotype of analyzed cellswas verified by results of immunohistochemicalstaining for neurofilament 200, and results wereexpressed as grains/neuron. Significant differencesbetween control animals and time points postliga-ture placement were determined using analysis ofvariance (ANOVA).
Immunohistochemistry
Frozen brain tissue sections (4�m thickness), takenin series with those used for in situ hybridization,were prepared and mounted onto Histostick-coatedslides (Accurate, Westbury, NY). Sections were fixedin acetone for 10 minutes, air-dried, and stored at–20°C until use. At the time of staining, sections wereair-dried, rehydrated in PBS (pH 7.4), and endoge-nous peroxidase activity was blocked for 50 secondsusing 0.28% periodic acid (Sigma) in ddH2O. Non-specific binding was blocked with normal horseserum (1:10) for 40 minutes. Tissue sections wereincubated overnight at room temperature with amonoclonal antineurofilament (NF)-200 antibody(1:100) (Sigma) or mouse IgG1 (Pharminigen, Min-neapolis, MN) isotype control. Tissue sections ofcerebellum were stained for NF-200 immunoreac-tivity as a positive control. A mouse avidin-biotin-complex secondary antibody system was used (1:800) (Vectastain Elite ABC Kit; Vector Laboratories,Burlingame, CA), and positive immunoreactivitywas visualized with cobalt-enhanced diaminoben-zidine (Sigma). Slides were counterstained with nu-clear fast red (Sigma), dehydrated through an as-cending series of alcohols followed by xylenesubstitute, and coverslipped.
Statistics
Statistical significance of changes in paw-with-drawal latency, alterations in levels of TNF�, andalterations in accumulated mRNA specific for TNF�were determined using paired Student’s t test orANOVA. All results were expressed as mean val-ues � SEM.
Results
Development of Hyperalgesia Following CCI
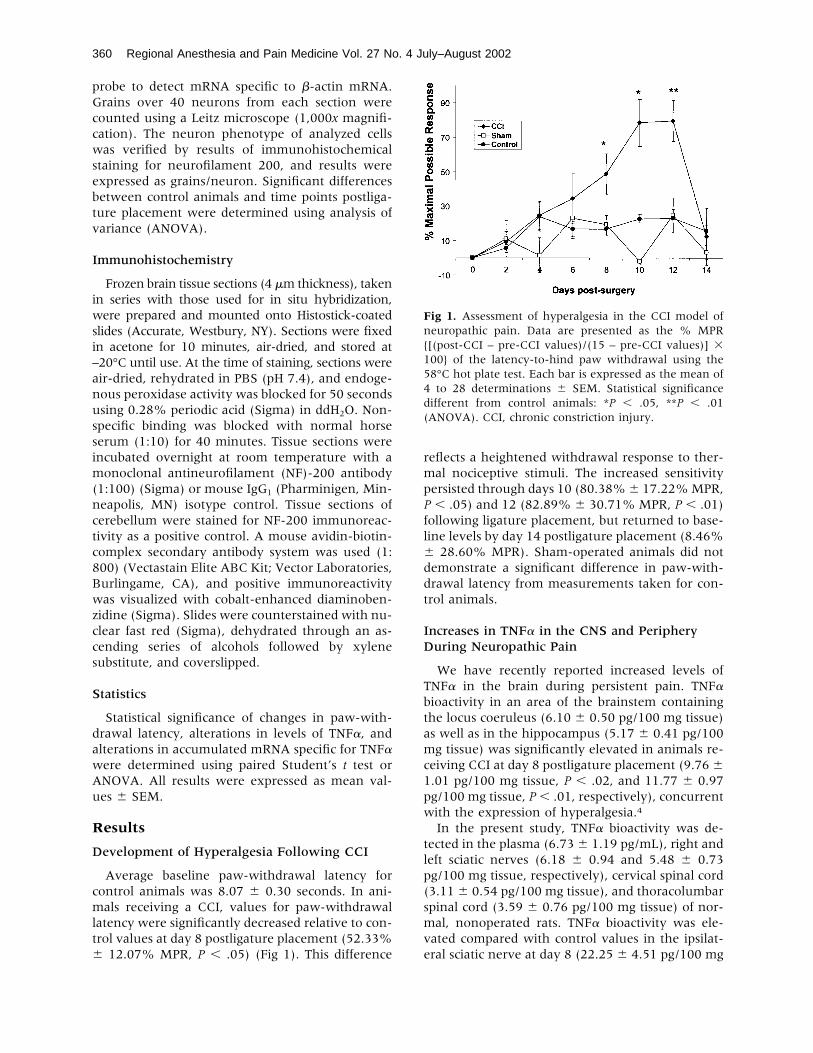
Average baseline paw-withdrawal latency forcontrol animals was 8.07 � 0.30 seconds. In ani-mals receiving a CCI, values for paw-withdrawallatency were significantly decreased relative to con-trol values at day 8 postligature placement (52.33%� 12.07% MPR, P � .05) (Fig 1). This difference
reflects a heightened withdrawal response to ther-mal nociceptive stimuli. The increased sensitivitypersisted through days 10 (80.38%� 17.22%MPR,P � .05) and 12 (82.89% � 30.71% MPR, P � .01)following ligature placement, but returned to base-line levels by day 14 postligature placement (8.46%� 28.60% MPR). Sham-operated animals did notdemonstrate a significant difference in paw-with-drawal latency from measurements taken for con-trol animals.
Increases in TNF� in the CNS and PeripheryDuring Neuropathic Pain
We have recently reported increased levels ofTNF� in the brain during persistent pain. TNF�bioactivity in an area of the brainstem containingthe locus coeruleus (6.10 � 0.50 pg/100 mg tissue)as well as in the hippocampus (5.17 � 0.41 pg/100mg tissue) was significantly elevated in animals re-ceiving CCI at day 8 postligature placement (9.76 �1.01 pg/100 mg tissue, P � .02, and 11.77 � 0.97pg/100 mg tissue, P � .01, respectively), concurrentwith the expression of hyperalgesia.4
In the present study, TNF� bioactivity was de-tected in the plasma (6.73� 1.19 pg/mL), right andleft sciatic nerves (6.18 � 0.94 and 5.48 � 0.73pg/100 mg tissue, respectively), cervical spinal cord(3.11 � 0.54 pg/100 mg tissue), and thoracolumbarspinal cord (3.59 � 0.76 pg/100 mg tissue) of nor-mal, nonoperated rats. TNF� bioactivity was ele-vated compared with control values in the ipsilat-eral sciatic nerve at day 8 (22.25 � 4.51 pg/100 mg
Fig 1. Assessment of hyperalgesia in the CCI model ofneuropathic pain. Data are presented as the % MPR{[(post-CCI – pre-CCI values)/(15 – pre-CCI values)] �100} of the latency-to-hind paw withdrawal using the58°C hot plate test. Each bar is expressed as the mean of4 to 28 determinations � SEM. Statistical significancedifferent from control animals: *P � .05, **P � .01(ANOVA). CCI, chronic constriction injury.
360 Regional Anesthesia and Pain Medicine Vol. 27 No. 4 July–August 2002
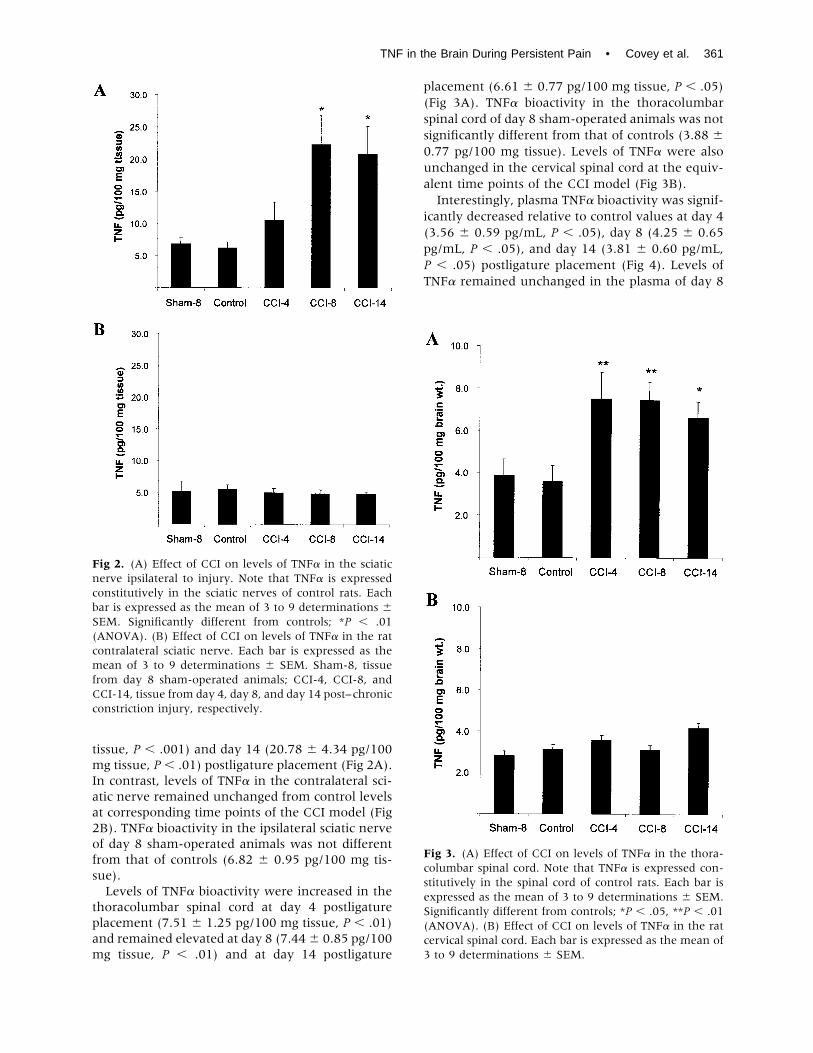
tissue, P � .001) and day 14 (20.78 � 4.34 pg/100mg tissue, P � .01) postligature placement (Fig 2A).In contrast, levels of TNF� in the contralateral sci-atic nerve remained unchanged from control levelsat corresponding time points of the CCI model (Fig2B). TNF� bioactivity in the ipsilateral sciatic nerveof day 8 sham-operated animals was not differentfrom that of controls (6.82 � 0.95 pg/100 mg tis-sue).Levels of TNF� bioactivity were increased in the
thoracolumbar spinal cord at day 4 postligatureplacement (7.51 � 1.25 pg/100 mg tissue, P � .01)and remained elevated at day 8 (7.44� 0.85 pg/100mg tissue, P � .01) and at day 14 postligature
placement (6.61 � 0.77 pg/100 mg tissue, P � .05)(Fig 3A). TNF� bioactivity in the thoracolumbarspinal cord of day 8 sham-operated animals was notsignificantly different from that of controls (3.88 �0.77 pg/100 mg tissue). Levels of TNF� were alsounchanged in the cervical spinal cord at the equiv-alent time points of the CCI model (Fig 3B).Interestingly, plasma TNF� bioactivity was signif-
icantly decreased relative to control values at day 4(3.56 � 0.59 pg/mL, P � .05), day 8 (4.25 � 0.65pg/mL, P � .05), and day 14 (3.81 � 0.60 pg/mL,P � .05) postligature placement (Fig 4). Levels ofTNF� remained unchanged in the plasma of day 8
Fig 2. (A) Effect of CCI on levels of TNF� in the sciaticnerve ipsilateral to injury. Note that TNF� is expressedconstitutively in the sciatic nerves of control rats. Eachbar is expressed as the mean of 3 to 9 determinations �SEM. Significantly different from controls; *P � .01(ANOVA). (B) Effect of CCI on levels of TNF� in the ratcontralateral sciatic nerve. Each bar is expressed as themean of 3 to 9 determinations � SEM. Sham-8, tissuefrom day 8 sham-operated animals; CCI-4, CCI-8, andCCI-14, tissue from day 4, day 8, and day 14 post–chronicconstriction injury, respectively.
Fig 3. (A) Effect of CCI on levels of TNF� in the thora-columbar spinal cord. Note that TNF� is expressed con-stitutively in the spinal cord of control rats. Each bar isexpressed as the mean of 3 to 9 determinations � SEM.Significantly different from controls; *P � .05, **P � .01(ANOVA). (B) Effect of CCI on levels of TNF� in the ratcervical spinal cord. Each bar is expressed as the mean of3 to 9 determinations � SEM.
TNF in the Brain During Persistent Pain • Covey et al. 361
sham-operated rats (8.52 � 0.29 pg/mL, not signif-icant [NS]).
Localization and Quantification of mRNASpecific for TNF�
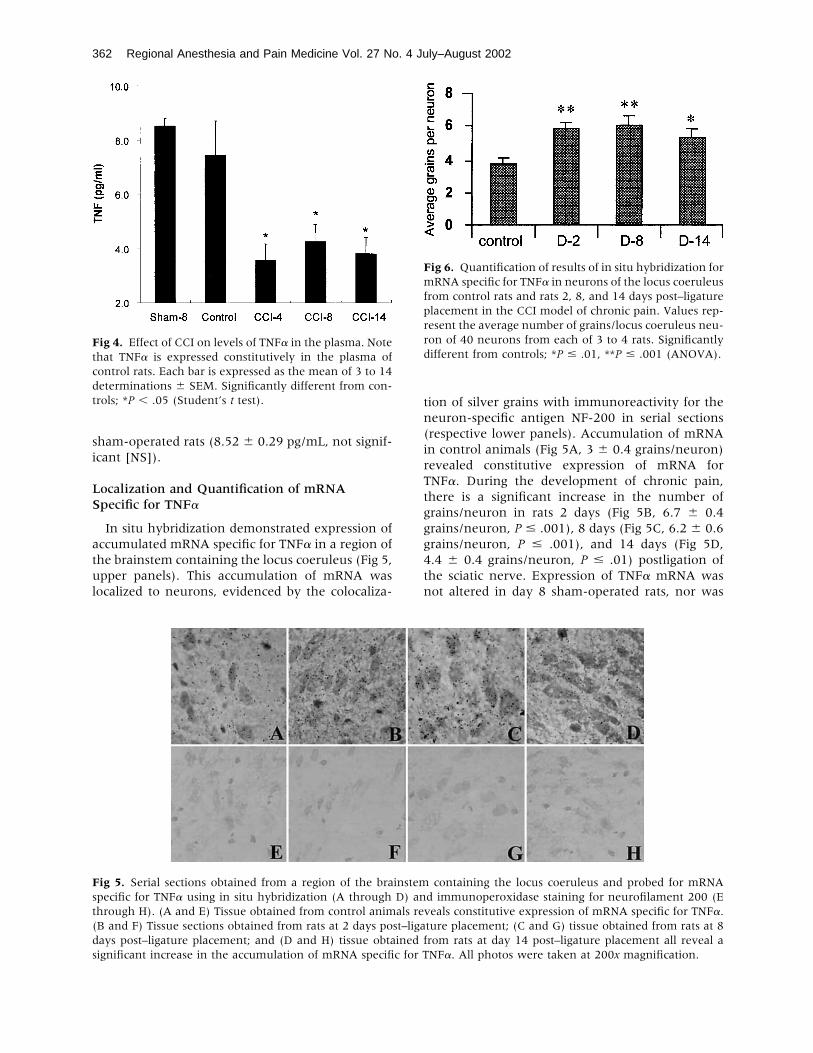
In situ hybridization demonstrated expression ofaccumulated mRNA specific for TNF� in a region ofthe brainstem containing the locus coeruleus (Fig 5,upper panels). This accumulation of mRNA waslocalized to neurons, evidenced by the colocaliza-
tion of silver grains with immunoreactivity for theneuron-specific antigen NF-200 in serial sections(respective lower panels). Accumulation of mRNAin control animals (Fig 5A, 3 � 0.4 grains/neuron)revealed constitutive expression of mRNA forTNF�. During the development of chronic pain,there is a significant increase in the number ofgrains/neuron in rats 2 days (Fig 5B, 6.7 � 0.4grains/neuron, P � .001), 8 days (Fig 5C, 6.2 � 0.6grains/neuron, P � .001), and 14 days (Fig 5D,4.4 � 0.4 grains/neuron, P � .01) postligation ofthe sciatic nerve. Expression of TNF� mRNA wasnot altered in day 8 sham-operated rats, nor was
Fig 4. Effect of CCI on levels of TNF� in the plasma. Notethat TNF� is expressed constitutively in the plasma ofcontrol rats. Each bar is expressed as the mean of 3 to 14determinations � SEM. Significantly different from con-trols; *P � .05 (Student’s t test).
Fig 5. Serial sections obtained from a region of the brainstem containing the locus coeruleus and probed for mRNAspecific for TNF� using in situ hybridization (A through D) and immunoperoxidase staining for neurofilament 200 (Ethrough H). (A and E) Tissue obtained from control animals reveals constitutive expression of mRNA specific for TNF�.(B and F) Tissue sections obtained from rats at 2 days post–ligature placement; (C and G) tissue obtained from rats at 8days post–ligature placement; and (D and H) tissue obtained from rats at day 14 post–ligature placement all reveal asignificant increase in the accumulation of mRNA specific for TNF�. All photos were taken at 200x magnification.
Fig 6. Quantification of results of in situ hybridization formRNA specific for TNF� in neurons of the locus coeruleusfrom control rats and rats 2, 8, and 14 days post–ligatureplacement in the CCI model of chronic pain. Values rep-resent the average number of grains/locus coeruleus neu-ron of 40 neurons from each of 3 to 4 rats. Significantlydifferent from controls; *P � .01, **P � .001 (ANOVA).
362 Regional Anesthesia and Pain Medicine Vol. 27 No. 4 July–August 2002

the pattern of expression of �-actin mRNA alteredby this paradigm (data not shown).
Discussion
Until recently, conventional dogma has stressedthe role of altered neuron function at the level ofthe spinal cord in the pathogenesis of neuropathicpain.15-17 Furthermore, the preponderance of re-search regarding changes in the expression of cyto-kines within the CNS during pain has focused onthis region.6,18,19 We have recently reported in-creased levels of biologically active TNF� in thebrain during persistent pain. TNF� bioactivity in thelocus coeruleus and hippocampus is significantlyelevated above constitutive levels in animals receiv-ing CCI at day 8 post–ligature placement, coincidentwith the appearance of hyperalgesia.4
In addition, our laboratory has demonstrated de-creases in adrenergic neuron activity secondary toincreases in TNF� in the brain during the develop-ment of persistent, neuropathic pain. Specifically,electrically stimulated norepinephrine (NE) releasein vitro is decreased in hippocampal tissue slicesisolated at day 8 post–ligature placement from an-imals undergoing CCI, as well as in spinal cordtissue slices from animals at day 4 and day 8 post–ligature placement.4,5 Similarly, electrically stimu-lated NE release is decreased in hippocampal tissuefrom nonoperated animals that receive icv micro-infusion of recombinant TNF�, contemporaneouswith the development of thermal hyperalgesia.5
This finding further suggests that increases in TNF�may modulate adrenergic neuron activity in theCNS during pain.Interestingly, while �2-adrenergic receptor sensi-
tivity is increased in the brain, it is not increased atthe level of the spinal cord in animals at day 8post–ligature placement.4 We hypothesize that theobserved decrease in NE release in the brain ismediated by elevated �2-adrenergic receptor sensi-tivity within the brain during neuropathic pain.Conversely, we postulate that the coincident reduc-tion in NE release from the spinal cord is not due toelevated �2-adrenergic receptor sensitivity, but oc-curs as a result of greater inhibition of NE release byelevated levels of TNF� bioactivity.The results of the present study also demonstrate
an elevation in TNF� bioactivity in the thoracolum-bar region of the spinal cord at day 4 post–ligatureplacement, prior to the manifestation of thermalhyperalgesia in the CCI model, and remain elevatedcoincident with the development of hyperalgesia atday 8 post–ligature placement. In contrast, TNF�bioactivity in the cervical spinal cord remains un-changed relative to control values during this pe-
riod. These data are consistent with those of otherinvestigators who report that increases in TNF� im-munoreactivity in the spinal cord following CCI arelimited to the lumbar spinal cord.6 The lumbar dor-sal horn is associated with modulation of painprocessing, including segmental antinociceptionfollowing epidural administration of adrenergicagonists, such as epinephrine and clonidine.20 Fur-thermore, neuron degeneration, including pyknosisand hyperchromatosis, is increased specifically inthe lumbar, but not the cervical dorsal horns of thespinal cord of rats undergoing CCI.21 The specificlocalization of these changes in neuron functionand morphology to the lumbar spinal cord follow-ing CCI is associated with the area of innervation ofthe sciatic nerve.6 Many of these changes are foundin the territory of sciatic nerve innervation withinspinal cord laminae I-II.21 The specificity of theincrease in levels of TNF� within this region of thespinal cord, combined with previous data demon-strating decreased NE release from spinal cord tissuefollowing CCI,4 suggests a role for TNF� within theCNS in the pathogenesis of neuropathic pain.The observed increase in TNF� in the injured
sciatic nerve is consistent with other studies thatreport increased TNF� in the sciatic nerve subse-quent to nerve injury.3 Increasing the level of TNF�within the sciatic nerve by direct application ofexogenous TNF� is associated with augmentedspontaneous firing of nociceptive afferents,8 as wellas with the development of thermal hyperalgesiasimilar to that observed following experimentalnerve injury.9 Additionally, other investigatorshave proposed that the elevation in levels of TNF�within the sciatic nerve subsequent to nerve injurycontributes to the afferent barrage to the CNS thatis believed to be causal in producing neuropathicpain.3 Our current findings support this hypothesis.Since levels of TNF� are increased unilaterally inthe injured sciatic nerve, local processes specific tothe site of nerve injury, rather than a global inflam-matory response are likely.Interestingly, the increase in levels of TNF� bio-
activity persists up to day 14 post–ligature place-ment, a time at which hyperalgesia is no longerpresent. Relevant to this is the observation thatpreoperative administration of thalidomide attenu-ates the increase in immunoreactivity to TNF� inthe injured nerve and prevents the development ofhyperalgesia, although this treatment is not effec-tive when the drug is administered after hyperalge-sia has appeared.3 These findings suggest that theincrease in TNF� at the site of nerve injury initiates,but is not required for the maintenance of neuro-pathic pain.The present discovery that TNF� bioactivity is
TNF in the Brain During Persistent Pain • Covey et al. 363

decreased in the plasma during the transient hyper-algesia demonstrates that animals undergoing CCIare not simply manifesting a systemic response tothe nerve injury. This result also demonstrates thatthe increases in levels of TNF� bioactivity observedin the brain and spinal cord are specific to the CNSand do not result from gradient-dependent move-ment of elevated TNF� from the peripheral bloodinto the CNS. This is further supported by the in-crease in mRNA specific for TNF�within neurons inthe brain following CCI, which further implicatesneurons of the CNS as a source of TNF� duringneuropathic pain, and as a site of action by TNF�during the pathogenesis of neuropathic pain. Inter-estingly, given that TNF� mediates numerous hostresponses in the body,22,23 it should be noted thatdecreased systemic levels of this pleiotropic, proin-flammatory cytokine might dramatically affect theantimicrobial host defenses of an animal experienc-ing chronic pain.The increase in levels of TNF� bioactivity in the
thoracolumbar spinal cord correlates with the de-crease in stimulated NE release from tissue slicesisolated from the lumbar region at day 4 post–ligature placement,4 at the time of the initial man-ifestation of hyperalgesia. Both the elevation inTNF� and the decrease in stimulated NE release aremaintained in the spinal cord at day 8 post–ligatureplacement, concurrent with the development of hy-peralgesia. In contrast, both the increase in TNF�levels, as well as the reduction in NE release, doesnot occur within the brain until day 8 post–ligatureplacement, coincident with the manifestation ofhyperalgesia. The temporal progression of thesechanges suggests that a TNF�-induced decrease inneurotransmitter release occurring initially withinthe spinal cord may modulate neuroplastic changesin the brain (increased �2-adrenergic receptor sen-sitivity and concomitantly decreased NE release)that ultimately lead to the initiation of hyperalgesia.It is of interest that levels of TNF� within the
spinal cord are still elevated on day 14 post–ligatureplacement, despite the fact that hyperalgesia is notpresent by this time. These findings suggest thatincreased levels of TNF� in the spinal cord partici-pate in the mechanisms that initiate hyperalgesia,but are not required for the maintenance of hyper-algesia. Conversely, levels of biologically activeTNF� in the brain return to control values by day 14post–ligature placement, coincident with the dissi-pation of hyperalgesia,4 while continued icv micro-infusion of TNF� proximal to the hippocampusresults in sustained hyperalgesia past day 14 post–ligature placement.5 In addition, we have estab-lished the requirement for the increase in TNF� inthe brain for the development of persistent pain,
since icv microinfusion of anti-TNF� antibodies atday 4 post–ligature placement (prior to the appear-ance of increased TNF� within the brain) abolishesthe manifestation of hyperalgesia.5 TNF�-depen-dent alterations in �2-adrenergic receptor sensitiv-ity also occur at the level of the brain (hippocam-pus), but not at the level of the lumbar spinal cordduring neuropathic pain.4 These data support thehypothesis that TNF�-induced alterations in adren-ergic neuron function in the brain, rather than thespinal cord, are required for the perception of hy-peralgesia in the CCI model.The increased accumulation of mRNA specific for
TNF� in neuron cell bodies within a region of thebrain associated with adrenergic neuron function24
and with modulation of nociception25,26 is consis-tent with our earlier findings of elevated levels ofTNF� bioactivity within this region of the brainfollowing CCI.4 Importantly, this increase occurscoincident with the manifestation of hyperalgesia,as well as with changes in adrenergic neuron re-sponsiveness in the brain. However, since increasedaccumulation of mRNA specific for TNF� in neuroncell bodies continues during the dissipation of hy-peralgesia and while TNF� levels decrease to con-trol values, posttranscriptional regulation is likely tobe occurring. The accumulation of TNF�-specificmRNA colocalizes with positive staining for NF-200, a neuron-specific antigen, demonstrating thatthe pain-associated increases in TNF� occur withinneurons.Numerous mechanisms have been proposed by
which TNF�may facilitate the development of painfollowing nerve injury. TNF� produced in the pe-riphery may activate nociceptive pathways in theCNS through transport to the dorsal root ganglia,spinal cord, or brain.27 Alternatively, peripheral cy-tokines, as well as glial or neuron activation, mayinduce increased expression of cytokines in theCNS. One laboratory suggests that TNF� is causativeof neuroplasticity and neuron hyperexcitability byits ability to induce long-term potentiation in neu-rons of the CNS, ultimately leading to central sen-sitization.28 We postulate that increases in TNF�levels in the brain in response to a nociceptiveafferent barrage result in increased autoinhibitoryfunction by �2-adrenergic receptors and a concom-itant decrease in adrenergic neuron activity in thebrain. Activation of spinally projecting neuronswithin adrenergic nulclei of the brain produces an-tinociception mediated by �2-adrenergic receptorsin the spinal cord.25 Neuroplastic changes inducedby increased levels of TNF� within the brain mayultimately culminate in a loss of descending inhibi-tion of nociceptive dorsal horn neurons by adren-ergic supraspinal structures.25,26,29 Nociceptive neu-
364 Regional Anesthesia and Pain Medicine Vol. 27 No. 4 July–August 2002

rotransmission in the dorsal horn may also beenhanced by increases in TNF� at the level of thespinal cord, since TNF� inhibits NE release fromnoradrenergic nerve terminals.30
In summation, TNF� is specifically increased inregions of the brainstem and spinal cord associatedwith modulation of nociception in the CCI painmodel. Accumulation of mRNA specific for TNF� isincreased within neurons in a region of the brainassociated with adrenergic function, as well as withdescending inhibition of dorsal horn neuron activ-ity. Clearly, brain-derived TNF plays an importantrole in the pathogenesis of persistent pain in thismodel. We have postulated that changes in levels ofthis pleiotropic cytokine and alterations in adrener-gic receptor sensitivity participate in a reciprocalfashion, directing the pathogenesis of persistentpain by inducing neuroplastic changes. The findingfrom this study strongly supports our hypothesis.
References
1. Watkins LR, Wiertelak EP, Goehler LE, Smith KP,Martin D, Maier SF. Characterization of cytokine-induced hyperalgesia. Brain Res 1994;654:15-26.
2. Oka T, Wakugawa Y, Hosoi M, Oka K, Hori T. Intra-cerebroventricular injection of tumor necrosis fac-tor-� induces thermal hyperalgesia in rats. Neuroim-munomodulation 1996;3:135-140.
3. Sommer C, Marziniak M, Myers RR. The effect ofthalidomide treatment on vascular pathology and hy-peralgesia caused by chronic constriction injury of ratnerve. Pain 1998;74:83-91.
4. Covey WC, Ignatowski TA, Knight PR, Severin CM,Spengler RN. Brain-derived TNF�: Involvement inneuroplastic changes implicated in the conscious per-ception of persistent pain. Brain Res 2000;859:113-122.
5. Ignatowski TA, Covey WC, Knight PR, Severin CM,Nickola TJ, Spengler RN. Brain-derived TNF alphamediates neuropathic pain. Brain Res 1999;841:70-77.
6. DeLeo JA, Colburn RW, Rickman AJ. Cytokine andgrowth factor immunohistochemical spinal profilesin two animal models of mononeuropathy. Brain Res1997;759:50-57.
7. Hoopes PJ, DeLeo JA, Colburn RW, Rickman AJ,O’Hara JA, Liu KJ, Strawbridge RR. Assessment ofspinal cord oxygen and cytokine (IL-1� and TNF-�)levels following radiation or cryoneurolosis injury.Radiat Res Soc Abstr 1996;P13:233.
8. Sorkin LS, Xiao WH, Wagner R, Myers RR. Tumournecrosis factor-alpha induces ectopic activity in noci-ceptive primary afferent fibres. Neuroscience 1997;81:255-262.
9. Wagner R, Myers RR. Endoneurial injection of TNF-alpha produces neuropathic pain behaviors. Neurore-port 1996;7:2897-2901.
10. Bianchi M, Sacerdote P, Ricciardi-Castagnoli P, Man-
tegazza P, Panerai A. Central effects of tumor necrosisfactor alpha and interleukin-1 alpha on nociceptivethresholds and spontaneous locomotor activity. Neu-rosci Lett 1992;148:76-80.
11. Ignatowski TA, Spengler RN. Tumor necrosis fac-tor-�: Presynaptic sensitivity is modified after antide-pressant drug administration. Brain Res 1994;665:293-299.
12. Kunkel SL, Spengler M, MayMA, Spengler R, LarrickJ, Remick DG. Prostaglandin E2 regulates macro-phage-derived tumor necrosis factor gene expres-sion. J Biol Chem 1988;263:5380-5384.
13. Paxinos G, Watson C. The Rat Brain In StereotaxicCoordinates, ed 3. New York, NY: Academic, 1996.
14. Elner VM, Strieter RM, Pavilack MA, Elner SG, Re-mick DG, Danforth JM, Kunkel SL. Human cornealinterleukin-8, IL-1 and TNF-induced gene expressionand secretion. Am J Pathol 1991;139:977-988.
15. Goff JR, Burkey AR, Goff DJ, Jasmin, L. Reorganiza-tion of the spinal dorsal horn in models of chronicpain: Correlation with behaviour. Neuroscience 1998;82:559-574.
16. Luo L, Puke MJ, Wiesenfeld-Hallin Z. The effects ofintrathecal morphine and clonidine on the preven-tion and reversal of spinal cord hyperexcitability fol-lowing sciatic nerve section in the rat. Pain 1994;58:245-252.
17. Yakhnitsa V, Linderoth B, Meyerson BA. Spinal cordstimulation attenuates dorsal horn neuronal hyper-excitability in a rat model of mononeuropathy. Pain1999;79:223-233.
18. DeLeo JA, Colburn RW, Nichols M, Malhotra A. In-terleukin-6-mediated hyperalgesia/allodynia and in-creased spinal IL-6 expression in a rat mononeuropa-thy model. J Interferon Cytokine Res 1996;16:695-700.
19. Arruda JL, Colburn RW, Rickman AJ, RutkowskiMD, DeLeo JA. Increase of interleukin-6 mRNA inthe spinal cord following peripheral nerve injury inthe rat: Potential role of IL-6 in neuropathic pain.MolBrain Res 1998;62:228-235.
20. Curatolo M, Petersen-Felix S, Arendt-Nielsen L,Zbinden AM. Epidural epinephrine and clonidinesegmental analgesia and effects on different pain mo-dalities. Anesthesiology 1997;87:785-794.
21. Sugimoto T, Bennett GJ, Kajander KC. Transsynapticdegeneration in the superficial dorsal horn after sci-atic nerve injury: Effects of a chronic constrictioninjury, transection, and strychnine. Pain 1990;42:205-213.
22. Ranheim EA, Kipps TJ. Tumor necrosis factor-alphafacilitates induction of CD80 (B7-1) and CD54 onhuman B cells by activated T cells: Complex regula-tion by IL-4, IL-10, and CD40L. Cell Immunol 1995;161:226-235.
23. Douni E, Akassoglou K, Alexopoulou L, Georgopou-los S, Haralambous S, Hill S, Kassiotis G, Kontoyian-nis D, Pasparakis M, Plows D, Probert L, Kollias G.Transgenic and knockout analyses of the role of TNFin immune regulation and disease pathogenesis. J In-flamm 1995-1996;47:27-38.
24. Dahlstrom A, Fuxe K. Evidence for the existence of
TNF in the Brain During Persistent Pain • Covey et al. 365

monoamine containing neurons in the central ner-vous system. I. Demonstration of monoamines in thecell bodies of brainstem neurons. Acta Physiol Scand1964;62:1-155.
25. Jones SL, Gebhart GF. Characterization of coeru-leospinal inhibition of the nociceptive tail-flick reflexin the rat: Mediation by �2-adrenergic receptors.Brain Res 1986;364:315-330.
26. Jones SL, Gebhart GF. Quantitative characteriza-tion of ceruleospinal inhibition of nociceptivetransmission in the rat. J Neurophysiol 1986;56:1397-1410.
27. DeLeo JA, Colburn RW. The role of cytokines innociception and chronic pain. In: Weinstein J, Gor-
don S, eds. Low Back Pain: A Scientific and ClinicalOver-view. Rosemont, IL: American Academy of Orthopae-dic Surgeons; 1996:163-185.
28. Patterson PH, Nawa H. Neuronal differentiation fac-tors/cytokines and synaptic plasticity. Cell 1993;72:123-137.
29. Hodge CJ, Apkarian AV, Stevens RT, Vogelsang GD,Wisnicki HJ. Locus coeruleus modulation of dorsalhorn unit responses to cutaneous stimulation. BrainRes 1980;240:415-420.
30. Elenkov IJ, Kovacs K, Duda E, Stark E, Vizi ES.Presynaptic inhibitory effect of TNF-� on the releaseof noradrenaline in isolated median eminence. J Neu-roimmunol 1992;41:117-120.
366 Regional Anesthesia and Pain Medicine Vol. 27 No. 4 July–August 2002




















