
LIM-Homeobox Gene Lhx5 Is Required for Normal Development of Cajal-Retzius Cells

DEVELOPMENTAL DYNAMICS 19R203-216 (1993)
Expression of Homeobox Genes Msx-I (Hex-7) and Msx-2 (Hox-8) During Cardiac Development in the Chick PENNY S. CHAN-THOMAS, ROBERT P. THOMPSON, BENOIT ROBERT, MAGDI H. YACOUB, AND PAUL J.R. BARTON Department of Cardiothoracic Surgery, National Heart and Lung Institute, London SW3 6LY, United Kingdom (P.S.C.-T.); Department of Cell Biology and Anatomy, Medical University of South Carolina, Charleston, South Carolina (R.P.T.); Dkpartement de Biologie Molkculaire, Institut Pasteur, 75724 Paris, Cedex 15, France (B.R.)
ABSTRACT The vertebrate homeobox genes Msx-1 and Msx-2 are related to the Drosophila msh gene and are expressed in a variety of tissues dur- ing embryogenesis. We have examined their ex- pression by in situ hybridisation during critical stages of cardiac development in the chick from stages 15 + to 37. Msx-1 expression is apparent in a number of non-myocardial cell populations, in- cluding cells undergoing an epithelial to mesen- chymal transformation in the atrioventricular and the outflow tract regions that play an integral role in heart septation and valve formation. Msx-2 ex- pression is restricted to a distinct subpopulation of myocardial cells that, in later stages, coincides morphologically with the cardiac conduction sys- tem. The timing of Msx-2 expression suggests that it plays a role in conduction system tissue forma- tion and that it identifies precursor cells of this specialised myocardium. The pattern of Msx-2 ex- pression is discussed with reference to current models of conduction tissue development. 0 1993 Wiley-Liss, Inc.
Key words: Homeobox genes, Cardiogenesis, Conduction system, Endocardial cushions, In situ hybridisation, Chick embryo
INTRODUCTION Homeobox genes encode a major class of regulatory
proteins believed to play an integral role in pattern formation during early development. Some homeobox genes (Antennapedia-class) occur in clusters at four separate chromosomal sites (reviewed by Krumlauf, 1992) and are expressed in characteristic regional pat- terns along the anterior-posterior axis of the develop- ing embryo (reviewed by Gaunt, 1991). To date, none of these genes has been shown to be expressed in the de- veloping heart although transcription of some (Hox- 1.2, -1.3, -1.4, -1.5, and -3.3) has been detected in the mid-gestation aortic trunk of the mouse (Gaunt et al., 1988). Also, mutation of the Hox-1.5 gene in the mouse can result in variable cardiac malformations in- cluding persistent patency of the arterial duct, aortic valvar stenosis, and defective formation of the pulmo- nary valve (Chisaka and Capecchi, 1991).
0 1993 WILEY-LISS. INC
Two recently isolated homeobox genes, Msx-1 and Msx-2 (formerly referred to as Hox-7 and Hox-8; see footnote) which are related to the Drosophila muscle segment homeobox gene (Gehring, 1987), have been identified in a variety of vertebrate species including Zebrafish (Ekker et al., 19921, Xenopus (Su et al., 19911, chick (Coelho et al., 1991; Yokouchi et al., 1991; Suzuki et al., 1991), quail (Takahashi and Le Douarin, 1990; Takahashi et al., 1991), and mouse (Hill et al., 1989; Robert et al., 1989, 1991; Davidson et al., 1991; MacKenzie et al., 1991a,b; Monaghan et al., 1991). Msx-1 and Msx-2 are not members of the Antennape- dia-class gene clusters (Hill et al., 1989; Robert et al., 1989, 1991) and, in the mouse, are not genetically linked (Monaghan et al., 1991). These genes have been shown to be expressed in a variety of developing tis- sues, including neural tube, branchial arches, limb bud, body wall, and heart but their regional and tem- poral patterns of expression during cardiac develop- ment have not been extensively described.
The heart forms as two concentric tubes of epithelial cells, and subsequently undergoes morphological and inductive events in distinct regions along its length (Icardo and Manasek, 1992) (see Fig. 1). The develop- ment of septa and valves commences in the formation of endocardia1 cushions, which are thought to function as primitive valves in the developing heart as well as contributing to the formation of the mature adult structures. Endocardial cushions are regions where populations of mesenchymal cells form by delamina- tion from the inner layer of the heart tube, the endocar- dium. Cells delaminate into an extracellular matrix, a locally extended basement membrane often referred to as the cardiac jelly, which is secreted principally by the outer, myocardial layer of the heart tube (Krug et al., 1985). Such cell delamination occurs in two regions: at the atrioventricular (AV) junction, where the lumen
Received June 10, 1993; accepted June 22, 1993. Address reprint requestsicorrespondence to Dr. Paul J.R. Barton,
Department of Cardiothoracic Surgery, National Heart and Lung In- stitute, Dovehouse Street, London SW3 6LY, United Kingdom.
We have adopted the nomenclature Msx-1 for Hox-7 and Msr-2 for H o x 8 following the proposals set out by Scott (Cell 71551-553, 1992).

204 CHAN-THOMAS ET AL.
t /delaminatina av
super ior av cushion
Fig. 1. Cartoon depicting stages of heart development, orientation, and tissues described in this paper. In A, the definitions of various terms used to describe the spatial relations of parts of the heart are illustrated. At HH stage 15 + , the onset of septation, the heart consists of a looped tube of myocardium lined by a tube of endocardium. The two are sepa- rated by extracellular matrix (cardiac jelly), which is thickest at the atrio- ventricular (av) junction and the outflow tract (oft). The ventricular (v) myocardial wall is thicker than that of the atria (a) and finger-like projec- tions (trabeculations) are also starting to develop. The atrial and ventric- ular cavities become divided into four chambers by the development of septa. The av septum is formed by the two endocardial "cushions" illus- trated by a transverse section (E) taken through the av junction of B (about HH stage 17). The cells of the single-cell thick endocardium are induced to undergo an epithelial to mesenchyme transformation. The
becomes divided into the right and left AV orifices, and in the outflow tract (OFT), where the single tube along which blood flows from the ventricles to the aortic tree becomes divided longitudinally to form the aorta and pulmonary artery. Valve leaf lets develop to maintain unidirectional blood flow at each of these four orifices.
At the AV junction, delamination of cells from the endocardium is initiated by inducing agents derived from the myocardium and secreted into the cardiac jelly (Krug et al., 1985). Experiments conducted in vitro have demonstrated that cells with the capacity to delaminate are present in the AV endocardium but ab- sent from ventricular endocardium (Runyan and Mark- wald, 1983). Similarly, the AV, but not the ventricular myocardium, secretes factors which induce endocardial cell delamination (Mjaatvedt e t al., 1987). Previous
resulting mesenchymal invasion of the thickened cardiac jelty forms the inferior and superior cushions which eventually meet and fuse (F, about HH stage 27) and contribute to the av valve leaflets. The lateral tubercles (It) also contribute to the latter. The heart undergoes considerable growth during development (not shown) such that the outflow tract comes to lie anterior to the atria. The interventricular septum commences as a coa- lescence of trabeculations and consists of myocardium covered by en- docardium, as does the atrial septum (C, about HH stage 27). The oft becomes divided by the fusion of two spiralling regions of mesenchyme formation into the aorta and pulmonary artery which are separated from the heart by valves. Thus, by HH stage 34 (D), the chick heart consists of four chambers with separate outflow vessels from each ventricle with all junctions guarded by valves.
studies of Msx-1 and Msx-2 expression have suggested that transcription of these genes coincides physically and temporally in tissues subject to pattern forma- tion and inductive processes in limb (Davidson et al., 1991; Coelho et al., 1991; Robert e t al., 1991), tooth (MacKenzie e t al., 1991a), and eye development (Monaghan et al., 1991). It is, therefore, of interest to determine the distribution of Msx-1 and Msx-2 expres- sion in the tissues of the developing heart, particularly during endocardial cushion formation.
This report describes the localisation of Msx-1 and Msx-2 RNA transcripts in chick embryonic hearts by in situ hybridisation from HH stage 15-t (Hamburger and Hamilton, 1951) to HH stage 37. The results dem- onstrate that Msx-1 is restricted during this critical period to certain endocardial and mesenchyme cell pop-

Msx EXPRESSION IN CHICK HEART DEVELOPMENT 205
ulations of the heart and, in particular, that its pattern of expression is consistent with a role during endocar- dial cushion formation both a t the AV junction and in the OFT. Msx-2 expression is restricted to a distinct myocardial cell population. Although partly coincident with myocardial regions involved in the induction of endocardial cushion tissue, the overall pattern of ex- pression is not suggestive of a role in this process. Moreover, in later development (HH stage 34 and 371, myocardial cells expressing Msx-2 coincide morpholog- ically with those regions previously identified as the cardiac conduction tissue. This suggests that Msx-2 plays a role in the formation of the conduction system of the heart, and may provide a useful early marker for conduction tissue.
RESULTS In situ hybridisation was carried out on sections of
staged chick embryos using RNA probes derived from previously characterised cDNAs encoding chick Msx-1 @fox-7) and Msx-2 (Hox-8) (Robert et al., 1991; Yokou- chi et al., 1991). Msx-1- and Msx-2-specific probes were derived by transcription of the non-coding regions of these cDNAs which share no significant sequence ho- mology with each other or other homeobox genes. Spec- ificity was demonstrated by the overall patterns of hy- bridisation of these probes throughout the embryo; for example, the contrasting patterns of expression of Msx-1 and Msx-2 in the neural tube. Labelling of these regions was used as a control for both specificity and efficiency of hybridisation between experiments. Over- all, the level of hybridisation observed in cardiac tis- sues was less than that observed in neural tube and limb. This was particularly the case for Msx-2, which is expressed in a limited number of cells within the heart, and where the level of signal was consistently low.
Expression of Msx-I in the AV Cushions and Lateral Tubercles
Msx-1 expression was first detected in the heart in individual cells delaminating from the AV epithelial endocardium and forming the mesenchymal population of the endocardial cushions. Labelled cells remained present throughout the subsequent delamination, growth, fusion, and incorporation of the AV cushions into the four-chambered structure of the heart. How- ever, there was considerable variation in strength and distribution of labelling during these events.
The first delaminating cells were found at HH stage 15+ (Fig. 2C). At HH stages 16+ to 18, labelled epi- thelial and mesenchymal cells could be found in the sparsely populated cushions, although not all were la- belled (data not shown). By HH stage 21, both inferior and superior cushions contained many mesenchymal cells and, in general, these were labelled strongly (Fig. 3C,E). Regional differences in strength of labelling were apparent, with cells further from the lumen of the heart tube being the more positive (Fig. 3E). There was
less label in the most distal parts of the cushions and a degree of local variation in expression within the ap- parently continuous mesenchymal populations. At HH stage 23, almost all the cushion cells were strongly positive, including those near the lumen, although those more distal were less labelled. Following fusion of the cushions (HH stage 271, labelling remained very strong, especially where the cells were densely packed immediately under the endocardium (Fig. 4C,E). Cells nearer the atrial chamber posteriorly were less la- belled by this stage. By HH stages 34 and 37, when the fused AV cushions were continuous with the atrial and interventricular septa, the cells were less strongly, al- though apparently uniformly, labelled.
The lateral tubercles arise by delamination and con- tribute to the AV valves (see Fig. 1C-F). There is con- siderable difference in structure of the definitive valves at the right and left AV junctions in the avian heart. The right valve consists of a large flap, derived mostly from the lateral myocardial wall, which is apposed by a medial pad. The left valve has a fibrous leaflet struc- ture more similar to that of mammalian AV valves (Vassall-Adams, 1982). At HH stage 23, the single cell- thick endocardium of the right lateral tubercle was la- belled, especially where this was distinctly separated from the myocardial wall (not shown); by HH stage 27, delamination had commenced and both epithelial and mesenchymal cells were labelled (Fig. 4C,E). Cells of the left AV tubercle were more strongly labelled at or immediately underlying the endocardial epithelium. At HH stage 34, expression was detected in all cush- ions of the AV valves. At the right AV junction, posi- tive epithelial and mesenchymal cells could be found, although labelling was largely confined to the epithe- lial layer in distinct regions. A short, non-expressing bridge of tissue divided the positive cells of the lateral cushion into two: a more lateral, proximal part (epithe- lial endocardium only) and a larger cushion opposing the medial AV cushion directly (Fig. 6A). The medial cushion consisted of a positive “cap” of epithelial cells overlying positive and negative populations of mesen- chymal cells (Figs. 6A, 71). Posteriorly, one such posi- tive mesenchymal population was continuous with the positive mesenchymal cells of the fused central cush- ions. Both leaflets of the left AV valve contained la- belled and unlabelled mesenchymal and epithelial cell populations although expression was principally in the endocardium facing the valve lumen (Fig. 6A). By HH stage 37, labelling of the AV valves was principally in the endocardium and the mesenchymal cells immedi- ately underlying i t (Fig. 75). Posteriorly, a positive mesenchymal population was continuous from the right medial cushion to the positive fused central cush- ions.
Expression of Msx-I in the Outflow Tract Cushions
The two ridges of cardiac jelly in the OFT become populated with mesenchymal cells and eventually fuse

206 CHAN-THOMAS ET AL.
A B
Fig. 2. Expression of Msx-1 and Msx-2 RNAs in embryonic chick heart at HH stage 15 + . (A) Cartoon depicting approximate plane of sectioning shown in C and D. (B) Cartoon depicting approximate distribution of Msx-2 expression in the myocardium at this stage. (C) and (D) Dark field images of serial sections of a heart of which alternate sections were hybridised with probes for Msx-1 and Msx-2, respectively. (C) The same few endocardial av cells (arrow) were consistently labelled by the probe
for Msx-1. Bar: 100 prn. (D) Very weak labelling by the probe for Msx-2 probe was consistently detected over myocardium in the lesser curvature of the heart (between large arrows) and along the left margin of the av junction (between fine arrows). The bright cells in the atrium are unla- belled but refracting red blood cells. The body wall and parts of the neural tube are labelled by each probe. a, atrium; v, ventricle; av, atrioventricular junction; oft, outflow tract; nt, neural tube; bw, body wall.
to form separate routes from the left and right ventri- cles, leading to the aorta and the pulmonary artery. Unlike that in the AV cushions, strong expression of Msx-I was restricted to distinct distal cell populations in the OFT. Mesenchyme first appears in the OFT slightly later than in the AV cushions. At HH stage
17-18, mesenchymal cells were present but were not labelled. By HH stage 21, labelled cells were visible, although the majority remained negative (Fig. 7C,D). Labelled cells were distributed unevenly amongst the mesenchymal population. At HH stage 23, delaminat- ing cells forming cushions were evident in the two

Msx EXPRESSION IN CHICK HEART DEVELOPMENT
B
207
Fig. 3. Expression of Msx-1 and Msx-2 RNAs in embryonic chick heart at HH stage 21. (A) Cartoon depicting approximate planes of sec- tioning: frontal (f) in C,D; transverse (1) in E,F. (B) Cartoon depicting approximate distribution of Msx-2 expression. (C,E) Dark field images of in situ hybridisation for Msx-I. Note expression in the av endocardial cushion tissue. (D,F) Dark field images following in situ hybridisation for
Msx-2. Note expression in the myocardial wall near the av junction (be- tween arrows in F). Bar: 100 km; a, atrium; v, ventricle; av, atrioventric- ular junction; oft, outflow tract; s and i, superior and inferior endocardial cushions, respectively; my, myocardium; ba, branchial arch; bw, body wall; Ib, limb bud.

CD9 IN THE RAT NERVOUS SYSTEM 105
TABLE 1. CD9 Expression in the Rat"
Tissue E11.5 E l 2 E14.5 E l 6 E l 9 Neonatal Adult Crest derivatives
Sympathetic neurons + + + + + + +? DRGneurons - -
Chromaffin cells + + + + + Melanocytes -
Glia + + + -c
+ + + +?
Tissue Embryonic Adult CNS:brain
+? Neurons -
Glia & + Neurons ++ +? Glia + + Esophagus + + Skin + Lung + Bladder + Liver +
Bone/ cartilage +
CNS:spinal cord
Epithelia
Hematopoieticb -
a+ = ROCA2 staining; t = weak, almost undetectable staining; - = undetectable staining; +? = uncertain; cannot distinguish neuronal from glial staining; +' = ventral horn of the spinal cord. bPresumed hematopoietic cells in liver.
observed using an inverted fluorescence microscope (Zeiss ICM 405).
Tissue Culture Twenty adrenal glands from 4 day old rat pups were
dissected under sterile conditions, and placed in Ca+ +/ Mg+ + free Hanks'-A solution. The medullae were dis- sected out, and dissociated in 1 mg/ml collagenase in Hanks'-A for 1 hr at 37°C. The enzyme was then blocked by the addition of serum-containing culture medium. Cells were washed free of collagenase by cen- trifuging at 1,000 rpm and resuspending in culture me- dium (repeated twice). Undissociated clumps were bro- ken up by gentle trituration using a flame-polished Pasteur pipette that had been preadsorbed with serum. Cells were plated on polylysine-laminin coated tissue culture dishes (Corning, 35 mm) at a density of lo5 cells per dish.
L15 C02 medium (Hawrot and Patterson, 1979) sup- plemented with 10% FBS was used as the tissue cul- ture medium. For chromaffin cells, dexamethasone (1 mM, Sigma) was added from a 1 , 0 0 0 ~ stock in 95% ethanol. Ara-C (10 mM) was added for 5 days to elim- inate cortical cells. Chromaffin cells grown in this me- dium for 10 days were converted into sympathetic neu- rons by replacing dexamethasone with nerve growth factor (0.1 mg/ml, Boehringer-Mannheim), and cultur- ing for 10 days.
Immunoblotting Embryonic and adult tissues were dissected and fro-
zen directly on dry ice. After homogenization in TBS
(50 mM Tris HC1, 50 mM NaC1, pH 7.5) containing protease inhibitors, the nuclei were removed by cen- trifugation at 1,000 rpm. The supernatant was centri- fuged at 100,OOOg in a Beckman ultracentrifuge. The membrane fraction obtained as the pellet was resus- pended in homogenization buffer containing 1% Triton X-100 and centrifuged at lOO,OOOg, yielding a Triton- extractable component as the supernatant. The protein content was determined using BCA reagent (PIERCE). SDS-PAGE was performed as described by Laemmli (19701, using a 1.5 mm thick, 10% separating gel. Fol- lowing electrophoresis, the gel was equilibrated for 15 min in transfer buffer (12.5 mM Tris, 96 mM glycine, pH 8.5,20% v/v methanol), and the proteins within the gel were transferred to a nitrocellulose membrane (0.45 pm, Schleicher and Schuell) overnight at 50 V. The nitrocellulose strips were then preadsorbed with 5% nonfat dry milk in TBS (50 mM Tris-HC1, 0.9% NaC1, pH 7.5) containing 0.1% Tween-20 (TBST) for 30 min. Incubation with purified ROCA2 (50 pg/ml in 5% nonfat dry milk in TBST) for 5 hr at RT was followed by 3 washes in TBST, and then incubation with a perox- idase-conjugated goat-anti-mouse IgG (Chemicon, 1500) for 1 hr at RT. After 3 washes (the last wash in TBS without Tween-201, immunoreactivity was devel- oped using 0.5 mg/ml 4-chloro-1-naphthol in 0.02% H,O, for 20 min at RT. The nitrocellulose was then air dried and photographed.
ACKNOWLEDGMENTS We thank Li-Ching Lo and Dr. David Anderson for
the mAb 192IgG, Dr. Linda Parysek for the anti-pe-
a
Fig
4 E
xpre
ssio
n of
Msx
-f a
nd M
sx-2
RN
As
in e
mbr
yoni
c ch
ick
hear
t at H
H s
tage
27
(A)
Car
toon
dep
ictin
g tis
sues
pre
sent
in s
ectio
ns s
how
n in
C,D
and
pos
ition
(bo
xed)
of
regi
on e
nlar
ged
in E
-G T
he o
ft tra
ct is
abo
ve th
e pl
ane
of t
he p
age
(9) C
arto
on d
epic
ting
regi
on (r
ing)
of m
yoca
rdiu
m e
xpre
ssin
g M
sx-2
(fin
e st
ippl
ing
and
strip
es)
The
av c
ushi
ons
(circ
led
stip
plin
g) a
re in
clud
ed fo
r or
ient
atio
n (C
) Dar
k fie
ld im
age
follo
win
g hy
brid
isat
ion
for
Msx
-f s
how
ing
labe
lling
of t
he a
v cu
shio
n, la
tera
l tub
ercl
es (
arro
ws)
, and
epi
card
ium
and
su
bepi
card
iurn
(arr
owhe
ads)
(D
) Dar
k fie
ld im
age
follo
win
g hy
brid
isat
ion
for M
sx-Z
show
ing
labe
lling
of t
he c
rest
of t
he in
terv
entri
cula
r sep
tum
and
of a
v m
yoca
rdiu
m (a
rrow
s) B
ar 1
00
)Lm
. (E
) Dar
k fie
ld im
age
of th
e rig
ht a
v ju
nctio
n fo
llow
ing
hybr
idis
atio
n fo
r M
sx-1
sho
win
g de
tail
of la
belli
ng o
f som
e ep
icar
dium
and
und
erly
ing
sube
pica
rdiu
m a
nd v
ery
stro
ng la
bel-
ling
of th
e av
cus
hion
s an
d rig
ht la
tera
l tub
ercl
e. (F
) Dar
k fie
ld im
age
of th
e rig
ht a
v ju
nctio
n fo
llow
ing
hybr
idis
atio
n fo
r M
sx-2
sho
win
g re
stric
tion
of l
abel
ling
to t
he m
yoca
rdiu
m. (
G)
Pha
se c
ontra
st im
age
of rig
ht a
v ju
nctio
n sh
owin
g po
sitio
n of
tis
sues
pre
sent
in E
,F. B
ar:
25pr
n. ra
, la,
righ
t and
left
atriu
m, r
espe
ctiv
ely;
v, v
entri
cle;
avc
, atri
oven
tricu
lar c
ushi
on; o
ft,
outfl
ow tr
act;
e, e
pica
rdiu
rn; l
at,
late
ral t
uber
cle:
ivs,
inte
rven
tricu
lar s
eptu
m; m
y, m
yoca
r-
dium
.

Msx EXPRESSION IN CHICK HEART DEVELOPMENT 209
Fig. 5. Mapping the distribution of Msx-2 RNA expression in the em- bryonic chick heart at HH stage 27. A-D: Dark field images following in situ hybridisation on frontal sections moving from anterior to posterior. Regions of labelling within the heart are indicated by arrows. Parts of the
body wall surrounding the heart are also strongly labelled. E-H: Cartoons depicting tissues present in A-D. a, atrium; v, ventricle; avc, atrioventric- ular cushion; oc, outflow tract cushion; poc, proximal outflow tract cush- ion; pv, posterior ventricular wall; i, crest of the interventricular septum.
main ridges but mesenchymal cells were present in much of the cardiac jelly around the circumference of the OFT lumen. Proximally, the two main regions of cushion formation were labelled, the endocardium and the mesenchymal cells in the cushions between them being negative. More distally, the labelling became much weaker in the regions of delamination and nu- merous mesenchymal cells throughout the cardiac jelly were not labelled. Strong expression was detected in distinct distal cell populations near the point at which the OFT joined the aortic tree (where the aorta divides into branchial arteries). Label was present in both the endocardial epithelial layer and the underlying mes- enchymal cells, many of which were in small groups in otherwise acellular cardiac jelly (Fig. 7E,F). This over- all pattern of expression was maintained at HH Stage 27. Strongly labelled distal populations were present close to the arch mesenchyme, although not all cells were labelled (Fig. 7G,H). Some parts of these cushions contained mixed populations of intensely labelled and unlabelled cells. By HH Stage 34, expression in the OFT was restricted to the valves of the pulmonary ar- tery and aorta. Labelling was strongest in the non- ventricle-facing endocardium a t the mural insertion and in the tips of the cusps themselves (Figs. 6A, 71). Where cusps adjoined, labelling was less intense or ab- sent. The labelling of the rather dense mesenchyme of the valves was weak and diffuse. At HH Stage 37, only weak labelling was present, intermittent around the
endocardial layer on both the proximal and distal faces of the cusps (Fig. 75).
Other Sites of Msx-l Expression Associated With the Heart
Mesothelium from the external surface of the venous sinus starts to cover the heart to form the epicardium from HH stage 17. Expression of Msx-I in the epicar- dium was detected in a stage and region-specific pat- tern. At HH stage 23, weak labelling was present in both the epithelial layer (epicardium) and underlying mesenchymal cells (sub-epicardium) in the right and left AV grooves. Much stronger expression was de- tected in the sub-epicardium beneath the atria but not in the epicardium, except in a region where no mesen- chymal cells were present between it and the atrial wall (not shown). By HH stage 27, the epicardium and sub-epicardium were strongly labelled in the AV re- gion around the heart, including part of the proximal region of the OFT. The transition between positive and negative regions was abrupt (Fig. 4E). Posteriorly, la- belling was limited to the dense epicardium immedi- ately adjacent to the myocardial wall. Such labelling was absent by HH stage 34.
The atrial cavity becomes divided by a myocardial septum which develops from the posterior wall and, like the rest of the atrial myocardium, is lined by epi- thelial endocardium. Some of these endocardial cells and those lining other septa in the atria and inflow
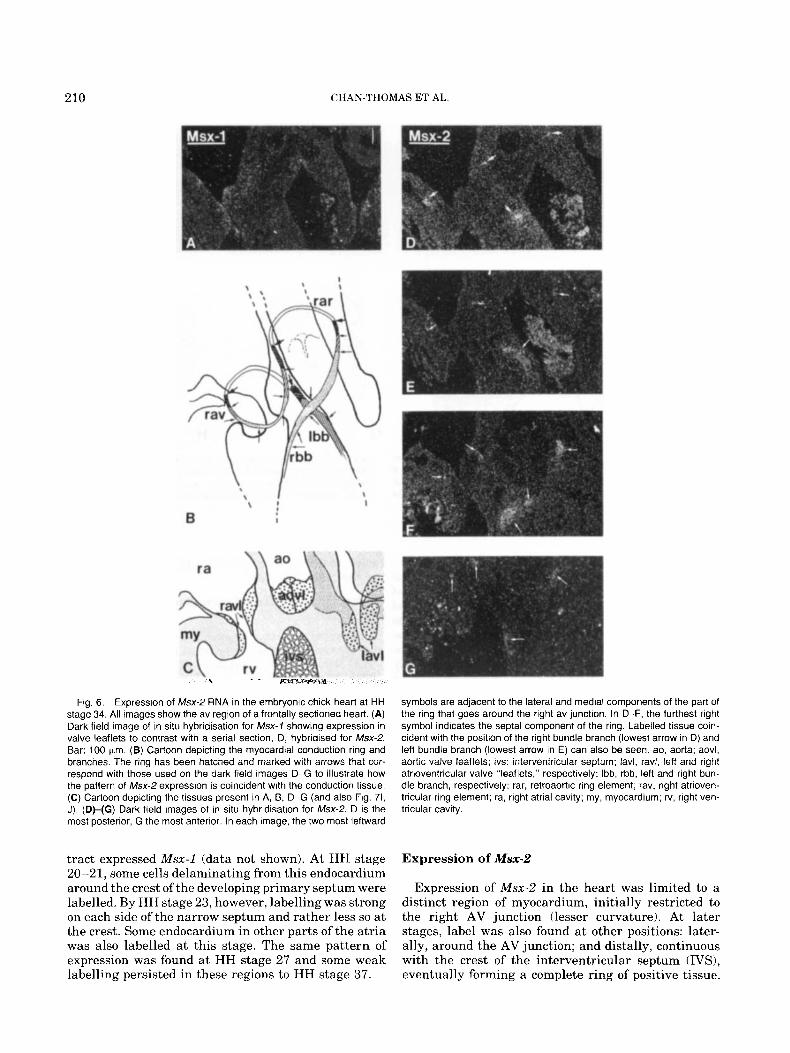
210 CHAN-THOMAS ET AL.
\ I ' : B I
\ I
Fig. 6. Expression of Msx-2 RNA in the embryonic chick heart at HH stage 34. All images show the av region of a frontally sectioned heart. (A) Dark field image of in situ hybridisation for Msx-1 showing expression in valve leaflets to contrast with a serial section, D, hybridised for Msx-2. Bar: 100 Km. (B) Cartoon depicting the myocardial conduction ring and branches. The ring has been hatched and marked with arrows that cor- respond with those used on the dark field images D-G to illustrate how the pattern of Msx-2 expression is coincident with the conduction tissue. (C) Cartoon depicting the tissues present in A, B, D-G (and also Fig. 71, J). (DHG) Dark field images of in situ hybridisation for Msx-2. D is the most posterior, G the most anterior. In each image, the two most leftward
tract expressed Msx-1 (data not shown). At HH stage 20-21, some cells delaminating from this endocardium around the crest of the developing primary septum were labelled. By HH stage 23, however, labelling was strong on each side of the narrow septum and rather less so a t the crest. Some endocardium in other parts of the atria was also labelled at this stage. The same pattern of expression was found a t HH stage 27 and some weak labelling persisted in these regions to HH stage 37.
symbols are adjacent to the lateral and medial components of the part of the ring that goes around the right av junction. In D-F, the furthest right symbol indicates the septa1 component of the ring. Labelled tissue coin- cident with the position of the right bundle branch (lowest arrow in D) and left bundle branch (lowest arrow in E) can also be seen. ao, aorta; aovl, aortic valve leaflets; ivs: interventricular septum; lavl, ravl, left and right atrioventricular valve "leaflets," respectively; Ibb, rbb, left and right bun- dle branch, respectively; rar, retroaortic ring element; rav, right atrioven- tricular ring element; ra, right atrial cavity; my, myocardium; rv, right ven- tricular cavity.
Expression of Msx-2
Expression of Msx-2 in the heart was limited to a distinct region of myocardium, initially restricted to the right AV junction (lesser curvature). At later stages, label was also found at other positions: later- ally, around the AV junction; and distally, continuous with the crest of the interventricular septum (IVS), eventually forming a complete ring of positive tissue.

Msx EXPRESSION IN CHICK HEART DEVELOPMENT 211
Fig. 7. Expression of Msx-1 RNA in the outflow tract. (A,B) Phase contrast and dark field images of a transverse section through an HH stage 21 embryo showing the inferior av cushion. Most mesenchymal cells are labelled. (C,D) Phase contrast and dark field images of a trans- verse section of the same HH stage 21 embryo showing the outflow tract cushion mesenchyme. Very few mesenchymal cells are labelled. The brightness of the myocardium is due to light refraction, not labelling. Bar: 50 pm. (E,F) Phase contrast and dark field images showing proximal and distal parts of the outflow tract in a frontal section of an HH stage 23 embryo. Note strong expression in distal mesenchyme only (between
arrows). Bar: 100 prn. (G,H) Phase and dark field images showing distal outflow tract of an HH stage 27 embryo. Note the strong expression in the valve leaflets (stars) and the clear boundary with the surrounding mes- enchyme (arrows). (I,J) Dark field images of the av region of frontal sections of HH stage 34 ( I ) and 37 (J) chick embryos showing expression in the developing right av valve cushions (solid arrows) and aortic (open arrows) valve leaflets. Bar: 100 pm; a, aorta; av mes, inferior atrioven- tricular cushion mesenchyme; oft mes, outflow tract cushion mesen- chyme; m, my, myocardium; prox, proximal; dist, distal; ra, right atrial cavity.

212 CHAN-THOMAS ET AL.
Where the myocardium was more than one cell layer thick and was labelled, expression was often princi- pally in or restricted to the inner layer (for example, Fig. 4F), which was usually about 25 pm thick.
At HH stage 15+ , expression was weak and re- stricted to an arc around the AV junction from the lesser curvature posteriorly to almost the left lateral limit of the inferior cushion (Fig. 2D). By HH stage 17-18, label was also present around the anterior of the AV groove but strongest expression remained in the lesser curvature. By HH stage 23, strong labelling in the AV groove could be found from a position just me- dial to the inferior cushion and around the right AV junction to beneath the superior cushion. Less positive cells were also present a t the left AV groove but myo- cardium connecting these sites into a complete ring was only very weakly labelled (not shown). By HH stage 27, labelling beneath the superior cushion was weak and diffuse, and no longer continuous with the more positive cells in the inner layer of myocardium underlying the cushion of the left tubercle. Similarly, only weak, diffuse expression could be detected extend- ing posteriorly from this position in the left AV groove towards, but not as far as, the positive cells continuous with the right AV junction. By HH stage 34, no expres- sion was present in the left AV junction. A labelled population of posterior myocardial cells
extended distally from the lesser curvature a t HH stage 21 (Fig. 3D) terminating in or near coalescing trabeculations. Strong labelling of both posterior and anterior myocardium distal to the lesser curvature was found a t HH stage 23; again, the non-coalesced trabe- culations were not labelled. By HH stage 27, coalesced trabeculations forming the developing IVS were con- tinuous from the anterior wall to the posterior wall of the ventricles. Labelled cells were continuous through the crest of the IVS, thus completing a ring of positive myocardium (Figs. 4B, 5A-D). Throughout the ante- rior portion of this ring, the boundaries of expression in the inner myocardial layer were very clear and always to the left of the sharp curvature marking the junction of the OFT and the AV myocardium (Fig. 5B,C). La- belling of the IVS itself was strongest in the apical 50 pm of the crest (Figs. 4D, 5C). Regions where strong expression was present in the lumen-most layer of cells down to the apex of the ventricles were also present (Fig. 5C). Positive cells in the posterior wall formed a band up to 100 pm wide laterally, although the stron- gest expression was limited to a narrower region. Con- tinuous with these, a region of diffusely and weakly labelled myocardium stretched laterally towards the left AV junction.
At HH stage 34, Msx-2-positive cells formed a con- tinuous cord, around the right AV junction, through the IVS and around the aorta, with two positive branches extending to the lumens of each of the right and left ventricular cavities (Fig. GB,D-G). The level of expression and morphology of the positive cells varied around this ring. Labelling was strongest in the por-
tion from the posterior of the right AV junction, medi- ally through the IVS, and diagonally across the ante- rior of the left ventricular outlet. Weaker labelling was detected in the anterior and lateral parts of the right AV ring and retroaortically. Morphologically, the pop- ulations of positive cells were often distinct and recog- nisable as a dense whorl or fascicle of myocytes embed- ded in more loosely packed tissue. The diameter of this ring also varied, being least in the regions of lowest labelling. Label was clearly limited to this distinct pop- ulation of cells apart from two regions: a t the posterior of the right AV junction, where weak and diffuse la- belling was also present in the myocardial walls adja- cent to the strongly positive cells; and in part of the septa1 portion, posterior to the branches, the cord of strongly labelled cells was surrounded by occasional isolated positive cells up to 25 pm away. By HH stage 37, the distribution of positive cells was essentially an incomplete version of that a t HH stage 34; the retroaor- tic part of the ring appeared morphologically distinct but unlabelled (not shown).
DISCUSSION Msx-1 Expression at the AV Junction
The principal site of Msx-1 RNA expression in the heart is in the endocardial cushions of the AV junction. Endocardial cushion tissue plays an integral role in the division of the primitive heart tube and formation of the heart valves, both a t the AV junction and in the OFT (Markwald et al., 1985). Endocardial cushions form by the delamination of epithelial cells from the endocardium into an extended basement matrix where they multiply and form the mesenchymal cell popula- tion of the developing cushion. Msx-1 expression was first detected in the heart with the onset of delamina- tion of cells from AV endocardium at HH stage 15+ (and no expression has been detected in preliminary studies of HH stages 11 and 14). Expression was ap- parent in the majority of both epithelial and mesenchy- ma1 cells in the AV region and persisted in the devel- oping cushion tissue throughout the stages examined here. The pattern of expression is, therefore, consistent with a role during both the epithelial to mesenchymal cell transition characteristic of cushion tissue forma- tion and the subsequent maturation of the developing cushions. Continuous Msx-1 expression cannot be an absolute prerequisite for these processes however, as not all cells of the AV cushions were labelled and, more importantly, considerable portions of the endocardial cushion tissue of the outflow tract were unlabelled (see below).
Expression of Msx-1 was also apparent in the lateral tubercles a t the AV junction which are the forerunners of the lateral valve leaflets. As with the AV cushions, the mesenchymal cells of the tubercles probably form by delamination from the endocardium. There are dif- ferences in morphology and in distribution of expres- sion of Msx-1 between the left and right lateral tuber- cles which may reflect the fact that the morphology of

Mssx EXPRESSION IN CHICK HEART DEVELOPMENT 213
the definitive left and right AV valves differs consid- erably in the chick (Vassall-Adams, 1982).
Msx-I Expression in Cushion Tissue of the OFT Endocardia1 cushion tissue of the OFT was not gen-
erally labelled, suggesting that the mechanism of OFT cushion formation may differ from that seen in the AV region. In the OFT, strong expression was limited to distal regions which probably correspond to the devel- oping OFT valve leaflets.
The difference in Msx-1 expression in AV and OFT cushions may be a reflection of the different develop- mental histories of the cells in these regions. The en- docardium within the heart is believed to be derived from lateral splanchnic mesoderm (Stalsberg and De- Haan, 1969). Recent studies have shown, however, that some precursors of the OFT endocardium and mesen- chyme (but not those of the AV endocardium) migrate to the heart via the cephalic paraxial and lateral me- soderm (Noden, 1991). Furthermore, experiments con- ducted with chick-quail chimeras (Phillips et al., 1987) and in ra t embryos (Fukiishi and Morriss-Kay, 1992) have demonstrated that cells from the neural crest at the level of the first three to four occipital somites also enter the cushion tissue of the OFT. No neural crest contribution has been identified in the AV region. It is not possible to identify the neural crest contribution in the experiments reported here, and the precise origin of the Msx-1 -expressing cells in the OFT remains, there- fore, unknown. Msx-1 has previously been shown to be expressed in certain neural crest-derived cell popula- tions located in other regions of the embryo. In the experiments described here, Msx-1 expression was ap- parent in neural crest-derived populations of the bran- chial arches as has been previously described (e.g., Robert et al., 1989; MacKenzie et al., 1991b).
Other Sites of Msx-I Expression in the Heart Msx-1 expression was detected in a subpopulation of
cells of the epicardium and sub-epicardium located at the AV junction. The epicardium is of extra-cardiac origin and gradually covers the external myocardial surface as an epithelium from the ventral wall of the venous sinus. Mesenchymal cells soon form a sub-epi- cardial layer within which the coronary arterial vas- culogenesis occurs (Hiruma and Hirakow, 1989). In the present study, clear-cut and identical proximal and di- stal boundaries of Msx-1 expression were observed in the epicardial and sub-epicardial layers in the AV grooves consistent with a localised induction event.
Within the atria, the principal site of Msx-1 expres- sion was the endocardium of the primary septum, ini- tially limited to the developing crest (HH stage 20-21) where a region of cell delamination was visible. At later stages, expression was present in the endocar- dium of the walls of the septum and persisted through- out the stages examined here, and may, therefore, be related to the endocardia1 response to the development
of the foramina secunda (Hendrix and Morse, 1977; Morse, 1981).
Msx-2 Expression and the Conduction System Msx-2 RNA expression was detected in a distinct and
consistent myocardial cell population in all the stages studied here, the first reported expression of an Msx gene in vertebrate heart muscle. At HH stage 15 + , cells of the inner layer of myocardium in the lesser curvature of the heart tube were labelled. During sub- sequent stages, expression was detected in an incom- plete AV ring formed by this myocardial cell popula- tion and, additionally, in two prongs extending distally into the inner layer of the myocardium and along the crest of the developing interventricular septum. These latter regions eventually formed a ring structure which, in the later stages, correlates with position of the cardiac conduction tissue (Vassall-Adams, 1982). Additionally, other parts of the conduction system, the left and right bundle branches, were also positive.
In the primitive heart tube, the atrial and ventricu- lar myocardium form a continuum in which electrical impulses pass freely. During subsequent development, they become electrically discontinuous except for the myocardial cells of the conduction system which traverse the atrioventricular junction and transmit the electrical impulses involved in coordinating contrac- tion of the four chambers. Development of the conduc- tion system, therefore, entails the specialisation of a subset of myocardial cells. A number of markers for conduction tissue have been proposed, including co- expression of atrial and ventricular myosin antigens (Sanders et al., 1984); acetyl cholinesterase activity (Lamers et al., 1987); Leu-7 antigen (Ikeda et al., 1991); and GIN2 antigen (Wessels et al., 1992).
The conduction system in the chick can be distin- guished morphologically a t HH stages 34 and 37 as densely grouped cells often surrounded by very loosely packed cells or matrix, giving a fasciculated appear- ance; i t is these densely grouped cells which expressed Msx-2. Prior to these stages, the morphological appear- ance of labelled myocardium differed little from that of other, unlabelled regions. In the present study, conti- nuity in the patterns of Msx-2 expression from stage to stage, however, strongly suggests that the positive cells observed at earlier stages were the precursors to the conduction system. Evidence from other work sup- ports this idea. The distributions of Leu-7 antigen in rat and human embryonic hearts (Ikeda et al., 1990) and of GIN2 antigen in the human embryonic heart (Wessels et al., 1992) are strikingly similar to the dis- tribution of Msx-2-expressing cells described here. Studies using these antibodies show strong binding to the crest of the developing interventricular septum and to part of the right AV myocardium.
Wessels et al. (1992) have proposed a model for the development of conduction tissue from a single “pri- mary conduction ring.” The results obtained here are almost entirely consistent with this model, which is

214 CHAN-THOMAS ET AL.
based on human heart studies, so phylogenetic differ- ences in heart structure (e.g., Robb, 1968) must also be considered. The proposed position of the primary con- duction ring is coincident with that of strong Msx-2 expression a t HH stage 27: from the right AV junction, down the anterior ventricular wall, through the crest of the interventricular septum, and up the posterior ven- tricular wall to the right AV junction once more (Fig. 4B). At a later stage of development (HH stage 34) the ring remains complete both histologically (as i t does in the adult bird) and with respect to Msx-2 expression. At an equivalent stage in the human, however, conduc- tion tissue (and GIN2 expression) is found only in parts of the primary conduction ring, the retroaortic and sep- tal segments already being absent, as they are in the adult. An earlier model (Wenink, 1976) proposed that the conduction system is derived from up to four myo- cardial rings, including an atrioventricular ring (i.e., including both right and left AV junctions). In the ex- periments described here, Msx-2 expression was de- tected not only a t the right AV junction, consistent with Wessels’ primary ring but also in the myocardium of the left AV junction. Although a complete AV ring of strong Msx-2 expression was not observed, there was often no sharp boundary between expressing and non- expressing tissue in this plane, a phenomenon noted with respect to Msx-2 expression in the developing mouse eye by Monaghan et al. (1991).
It is not possible to determine from these experi- ments whether the cells expressing Msx-2 at HH stage 15 + give rise to the entire positive population of cells at the later stages. Given the reduced proliferative ca- pacity of this portion of the myocardium between HH stages 18-20 (Thompson et al., 19901, the more exten- sive region of expression seen a t HH stage 34 may in- volve the recruitment of additional cells rather than only the proliferation of a core population.
Relationship Between Msx-1 and Msx-2 Expression and Induction Events in the Heart
Previous studies have demonstrated that regions of Msx-1 and Msx-2 expression within the embryo are often adjacent or overlapping, and that such regions are often associated with epithelial-mesenchymal cell interactions (e.g., Coelho et al., 1991; MacKenzie et al., 1991a,b, 1992; Monaghan et al., 1991; Robert et al., 1991; Takahashi et al., 1991). It is, therefore, tempting to propose some interdependence of the expression of these genes. Superficially, the expression of Msx-1 and Msx-2 in the endocardial cushions at the AV junction and the underlying myocardium, respectively, supports this view. However, while the region expressing Msx-2 overlaps with the AV myocardium involved in endocar- dial cushion induction, the overall pattern of Msx-2 expression is not easily reconciled with the distribution of cushion tissue formation in this region. Moreover, the distribution of Msx-2 expression in the conduction tissue during later stages of development suggests that
the proximity of Msx-1 and Msx-2 expression during early development may be simply coincidental.
Experiments performed in culture (Krug et al., 1985) have demonstrated that the epithelium to mesenchyme transition at the AV junction is induced by agents se- creted by the myocardium in this region and that only cells of the endocardium located in the AV region are capable of this response (Krug et al., 1987; Mjaatvedt et al., 1987). The results reported here show that mem- bers of this cell population express Msx-1 from the on- set of cell delamination and during maturation of the developing cushions. Studies of the regional expression of members of the TGFP supergene family suggest that they also play an integral role in this process. In par- ticular, expression of TGFP-1 and TGFP-2 (Akhurst et al., 1990; Millan et al., 1991; Schmid et al., 1991) has been detected in AV cushion tissue and the expression of TGFP-2, TGFP-3 (Pelton et al., 1991; Millan et al., 1991; Schmid et al., 1991)) BMP2A (Lyons et al., 19901, and BMP4A (Jones et al., 1991) detected in AV myo- cardium. In the case of TGFP-3, direct involvement has been implied through the use of in vitro endocardial cell culture techniques (Potts and Runyan 1989; Potts et al., 1991). Regions of expression of Msx-1 and of members of the TGFp superfamily also overlap else- where in the embryo. In the limb apex, for example, BMP2A and BMP4A transcripts have been detected and may be involved in the local activation of Msx-1 expression. Regional localisation of FGF receptor gene expression in AV cushion tissue has also been reported (Peters et al., 1992). Evidence of regional distribution of the retinoic acid receptor family in the developing heart has shown that RAR-y is expressed in endocar- dial cushion tissue (Dolle et al., 1990); elsewhere in the mouse embryo, regions that express Msx-1 are subre- gions of RAR-y expression (Lyons et al., 1992). The exact involvement of these gene families in the devel- opment of cushion tissue remains to be determined, although Song et al. (1992) have suggested that mouse Msx-1 promotes cell proliferation and suppresses dif- ferentiation. Experiments using endocardial cell cul- ture, as well as in vivo studies based on cell transfec- tion and transgenic techniques, are now required to address these issues in detail.
Msx-1 and Msx-2 Hybridisation Probes 35S-labelled RNA probes for detection of Msx-1 and
Msx-2 RNA were generated by transcription using pre- viously characterised chick cDNA clones (Robert et al., 1991; Yokouchi et al., 1991). In the case of Msx-1, an additional subclone containing only 3’-non coding se- quence was constructed by the use of site-directed mu- tagenesis in order to introduce an artificial Apal site a t the junction of the coding and non-coding sequence (by altering the sequence TAGGGCCG to TAGGGCCC, where TAG is the translation stop codon of the Msx-l transcript). The resulting clone (SApaIChox7) contains 360 bp of 3’ non-coding sequence with no significant
MATERIALS AND METHODS

Msx EXPRESSION IN CHICK HEART DEVELOPMENT 215
homology to Msx-2. In the case of Msx-2,3’ non-coding probe was generated by linearising a full-length cDNA at the AflII site located at the translation stop codon. This generated a probe corresponding to 200 bp of non- coding sequence. Control (sense strand) probes were generated by transcription of the full-length cDNA Msx-1 and Msx-2 clones.
In Situ Hybridisation The method used is based on those of Wilkinson et al.
(1987) and Sassoon et al. (1988). Briefly, chick em- bryos, staged according to Hamburger and Hamilton (19511, were fixed in 4% paraformaldehyde overnight at 4“C, washed, dehydrated in ethanol, embedded in Paraplast, and sectioned at 5 pm. Sections were mounted on clean 3-aminopropyltriethoxysilane- treated slides. Sections were prepared for hybridisation as follows: After removal of wax in xylene and rehy- dration, they were refixed in 4% paraformaldehyde in PBS for 20 min and washed in PBS. Following treat- ment with Proteinase K (20 pgiml) for 7.5 min, the sections were washed, refixed, washed briefly in water, treated with 0.25% vlv acetic anhydride in 0.1 M tri- ethanolamine twice for 5 min, dehydrated in ethanol, and air-dried. 35S-labelled RNA probe was diluted to 50,000 cpm/pl in hybridisation buffer (50% formamide, 0.3 M NaC1,20 mM Tris-HC1, pH 7.4,5 mM EDTA, 10 mM NaP04, 10% dextran sulphate, 1 x Denhardt’s solution, 50 pg/ml yeast tRNA), heated to 80” for 2 min, and applied to dry sections. Sections were covered with parafilm and incubated at 52°C overnight in a sealed humid box. Following hybridisation, sections were washed as follows: 2 x SSC (50”C, 30 min); 50% for- mamide, 2 x SSC, 10 mM DTT (65”C, 20 rnin); 2 x SSC (37”C, 10 min) twice; 4 x SSC, 20 kg/ml RNase A (37°C) 30 rnin); 2 x SSC (37”C, 10 rnin); and 0.1 x SSC (37°C) 15 min). They were then dehydrated through an ethanol series containing 0.3 M ammonium acetate and air-dried. For autoradiography, the slides were dipped in Ilford K5 emulsion diluted 1: l with water, allowed to air-dry, and exposed for 6 to 10 days at 4°C. Tissues were lightly counterstained, dehydrated, mounted in DPX, and examined under light and dark- field optics on a Zeiss Axioplan microscope.
ACKNOWLEDGMENTS We thank Atsushi Kuroiwa for providing the Msx-2
cDNA clone and Dr. J. Bee for help with chick embryos used in these studies. We are grateful to Professor R.H. Anderson and Dr. Y. Ho for advice and encouragement, to Patricia Germroth and Aldrin Sweeney for assis- tance with embryo preparation and sectioning, to Dr. C.D. Thomas for help with the site-directed mutagen- esis, and to Dr. L. Eisenberg for critical reading of the manuscript. This work was supported by the British Heart Foundation, of which P.B. is a Senior Research Fellow, and by a travel fellowship from the Royal So- ciety to R.P.T..
REFERENCES Akhurst, R.J., Lehnert, S.A., Faissner, A,, and Duflie, E. (1990) TGF
beta in murine morphogenetic processes: The early embryo and cardiogenesis. Development 108:645-656.
Chisaka, O., and Capecchi, M.R. (1991) Regionally restricted devel- opmental defects resulting from targeted disruption of the mouse homeobox gene hox 1.5. Nature 350:473-479.
Coelho, C.N.D., Krabbenhoft, K.M., Upholt, W.B., Fallon, J.F., and Kosher, R.A. (1991) Altered expression of the chicken homeobox- containing genes GHox-7 and GHox-8 in the limb buds of limbless mutant chick embryos. Development 113:1487-1493.
Davidson, D.R., Crawley, A., Hill, R.E., and Tickle, C. (1991) Position- dependent expression of two related homeobox genes in developing vertebrate limbs. Nature 352:429-431.
Dolle, P., Ruberte, E., Leroy, P., Morriss-Kay, G.M., and Chambon, P. (1990) Retinoic acid receptors and cellular retinoid binding proteins I. A systematic study of their differential pattern of transcription during mouse organogenesis. Development 110:1133-1151.
Ekker, M., Akimenko, M.-A., Bremiller, R., and Westerfield, M. (1992) Regional expression of three homeobox transcripts in the inner ear of zebrafish embryos. Neuron 9:27-35.
Fukiishi, Y., and Morriss-Kay, G. (1992) Migration of cranial neural crest cells to the pharyngeal arches and heart in rat embryos. Cell Tissue Res. 268:l-8.
Gaunt, S.J. (1991) Expression patterns of mouse Hox genes: Clues to an understanding of developmental and evolutionary strategies. Bioessays 13505 -513.
Gaunt, S.J., Sharpe P.T., and Duboule, D. (1988) Spatially restricted domains of homeo-gene transcripts in mouse embryos: Relation to a segmented body plan. Development 104 (Suppl):169-179.
Gehring, W.J. (1987) The homeobox: Structural and evolutionary as- pects. In: “Molecular Approaches to Developmental Biology,” Firtel, R.A. and Davidson, E.H. (eds). New York: Alan R. Liss, pp 115- 129.
Hamburger, V., and Hamilton, H.L. (1951) A series of normal stages in the development of the chick embryo. J. Morphology 38:49-92.
Hendrix, M.J.C., and Morse, D.E. (1977) Atrial septation I. Scanning microscopy in the chick. Dev. Biol. 57:345-363.
Hill, R.E., Jones, P.F., Rees, A.R., Sime, C.M., Justice, M.J., Copeland, N.G., Jenkins, N.A., Graham, E., and Davidson, D.R. (1989) A new family of mouse homeobox-containing genes: Molecular structure, chromosomal location, and development expression of Hox7.1. Genes Dev. 3:26-37.
Hiruma, T., and Hirakow, R. (1989) Epicardial formation in embry- onic chick hear t Computer-aided reconstruction, scanning and transmission electron microscopic studies. Am. J . Anat. 184:129- 138.
Icardo, J.M., and Manasek, F.J. (1992) Cardiogenesis: Development mechanisms and embryology. In: “The Heart and Cardiovascular System,” 2nd edition, Fozzard, H.A., Haber, E., Jennings, R.B., Katz, A.M., and Morgan, H.E. (eds). New York: Raven Press, pp 1563-1586.
Ikeda, T., Sakai, H., Shimokawa, I., Iwasaki, K., Matsuo, T. (1990) Expression of Leu-7 antigen in the embryonic h e a r t w i t h special reference to development of the conduction system. In: “Develop- mental Cardiology: Morphogenesis and Function,” Clark, E.B. and Takao, A. (eds). New York: Futura Publishing, Inc., pp 237-256.
Jones, C.M., Lyons, K.M., and Hogan, L.M. (1991) Involvement of Bone Morphogenetic Protein-4 (BMP-4) and Vgr-1 in morphogene- sis and neurogenesis in the mouse. Development 111531-542.
Krug, E.L., Runyan, R.B., and Markwald, R.R. (1985) Protein extracts from early embryonic hearts initiate cardiac endothelial cytodiffer- entiation. Dev. Biol. 112:414-426.
Krug, E.L., Mjaatvedt, C.H., and Markwald, R.R. (1987) Extracellular matrix from embryonic myocardium elicits an early morphogenetic event in cardiac endothelial differentiation. Dev. Biol. 120:348- 345.
Krumlauf, R. (1992) Evolution of the vertebrate Hox genes. Bioessays 14:245-252.
Lamers, W.H., te Kortschot, A,, Los, J.A., and Moorman, A.F.M. (1987) Acetylcholinesterase in prenatal rat hear t A marker for the

216 CHAN-THOMAS ET AL
early development of the cardiac conductive tissue? Anat. Rec. 217: 361-370.
Lyons, G.E., Houzelstein, D., Sassoon, D., Robert, B., and Bucking- ham, M.E. (1992) Multiple sites of Hox-7 expression during mouse embryogenesis: Comparison with retinoic acid receptor mRNA lo- calization. Mol. Reprod. Dev. 32:303-314.
Lyons, K.M., Pelton, R.W., and Hogan, B.L.M. (1990) Organogenesis and pattern formation in the mouse: RNA distribution patterns suggest a role for Bone Morphogenetic Protein 2 fBMP-2AI. Devel- opment 109:833-844.
MacKenzie, A,, Leeming, G., Jowett, A.K., Ferguson, M.W.J., and Sharpe, P.T. (1991a) The homeobox gene Hox-7.1 has specific re- gional and temporal expression patterns during early murine cra- niofacial embryogenesis, especially tooth development in vivo and in vitro. Development 111:269-285.
MacKenzie, A,, Ferguson, M.W.J., and Sharpe, P.T. (1991b) Hox 7.1 expression during murine craniofacial development. Development 113:601-611.
MacKenzie, A,, Ferguson, M.W.J., and Sharpe, P.T. (1992) Expression patterns of the homeobox gene, Hox-8, in the mouse embryo suggest a role in specifying tooth initiation and shape. Development 115: 403-420.
Markwald, R.R., Krug, E.L., Runyan, R.B., and Kitten, G.T. (1985) Proteins in cardiac jelly which induce mesenchyme formation. In: “Cardiac Morphogenesis,” Ferrans, C.V.J., Rosenquist, G. and Weinstein, C. (eds). New York: Elsevier, pp 60-69.
Millan, F.A., Denhez, F., Kondaiah, P., and Akhurst, R.J. (1991) Em- bryonic gene expression patterns of TGF PI, p2 and p3 suggest different developmental functions in vivo. Development 111:131- 144.
Mjaatvedt, C.H., Lepera, R.C., and Markwald, R.R. (1987) Myocardial specificity for initiating endothelial-mesenchymal cell transition in embryonic chick heart correlates with a particular distribution of fibronectin. Dev. Biol. 119:59-67.
Monaghan, A.P., Davidson, D.R., Sime, C., Graham, E., Baldock, R., Bhattacharya, S.S., and Hill, R.E. (1991) The Msh-like homeobox genes define domains in the developing vertebrate eye. Develop- ment 112:1053-1061.
Morse, D.E. (1981) Formation of foramina secunda in the chick. In: “Perspectives in Cardiovascular Research,” vol. 5, Pexieder, T. (ed). New York: Rowen Press, pp 139-149.
Noden, D.M. (1991) Origins and patterning of avian outflow tract. Development 111367-876.
Pelton, R.W., Saxena, B., Jones, M., Moses, H.L., and Gold, L.I. (1991) Irnmunohistochemical localization of TGF PI, TGF (32 and TGF P3 in the mouse embryo: Expression patterns suggest multiple roles during embryonic development. J. Cell Biol. 115:1091-1105.
Peters, K.G., Werner, S., Chen, G., and Williams, L.T. (1992) Two FGF receptor genes are differentially expressed in epithelial and mesenchymal tissues during limb formation and organogenesis in the mouse. Development 114:233-243.
Phillips, M.T., Kirby, M.L., and Forbes, G. (1987) Analysis of cranial neural crest distribution in the developing heart using quail-chick chimeras. Circulation Res. 60:27-30.
Potts, J.D., and Runyan, R.B. (1989) Epithelial-mesenchymal cell transformation in the embryonic heart can be mediated, in part, by transforming growth factor p. Dev. Biol. 134:392-401.
Potts, J.D., Dagle, J.M., Walder, J.A., Weeks, D.L., and Runyan, R.B. (1991) Epithelial-mesenchymal transformation of embryonic car- diac endothelial cells is inhibited by a modified antisense oligode- oxynucleotide to transforming growth factor p3. Proc. Natl. Acad. Sci. U.S.A. 88:1516-1520.
Robb, J.S. (1968) “Comparative Basic Cardiology.” New York and London: Grune and Stratton, p 348.
Robert, B., Sassoon, D., Jacq, B., Gehring, W., and Buckingham, M. (1989) Hox-7, a mouse homeobox gene with a novel pattern of ex- pression during embryogenesis. EMBO J . 8:91-100.
Robert, B., Lyons, G., Simandl, B.K., Kuroiwa, A., and Buckingham, M. (1991) The apical ectodermal ridge regulates Hox-7 and Hox-8 gene expression in developing chick limb buds. Genes Dev. 59363- 2374.
Runyan, R.B., and Markwald, R.R. (1983) Invasion of mesenchyme into three-dimensional collagen gels: A regional and temporal anal- ysis of interaction in embryonic heart tissue. Dev. Biol. 95:108-114.
Sanders, E., Moorman, A.F.M., and Los, J.A. (1984) The local expres- sion of adult chicken heart myosins during development I. The first three days embryonic chicken heart. Anat. Embryol. 169:185-191.
Sassoon, D.A., Garner, I., and Buckingham, M. (1988) Transcripts of a-cardiac and a-skeletal actins are markers for myogenesis in the mouse embryo. Development 104:155-164.
Schmid, P., Cox, D., Bilbe, G., Maier, R., and McMaster, G.K. (1991) Differential expression of TGF 61, 82 and p3 genes during mouse embryogenesis. Development 111:117-130.
Scott, M.P. (1992) Vertebrate homeobox gene nomenclature. Cell 71: 551-553.
Song, K., Wang, Y., and Sassoon, D. (1992) Expression of Hox-7.1 in myoblasts inhibits terminal differentiation and induces cell trans- formation. Nature 360:477-481.
Stalsberg, H., and DeHaan, R.L. (1969) The precardiac areas and formation of the tubular heart in the chick embryo. Dev. Biol. 19: 128-159.
Su, M-W., Suzuki, H.R., Solursh, M., and Ramirez, F. (1991) Progres- sively restricted expression of a new homeobox-containing gene during Xenopus Zaeuis embryogenesis. Development 11 1:1179- 1187.
Suzuki, H.R., Padanilam, B.J., Vitale, E., Ramirez, F., and Solursh, M. (1991) Repeating developmental expression of G-Hox 7, a novel homeobox-containing gene in the chicken. Dev. Biol. 148:375-388.
Takahashi, Y., and Le Douarin, N. (1990) cDNA cloning of a quail homeobox gene and its expression in neural crest-derived mesen- chyme and lateral plate mesoderm. Proc. Natl. Acad. Sci. U.S.A. 87:7482-7486.
Takahashi, Y., Bontoux, M., and Le Douarin, N. (1991) Epithelio- mesenchymal interactions are critical for Quox 7 expression and membrane bone differentiation in the neural crest derived mad ib - ular mesenchyme. EMBO J . 10:2387-2393.
Thompson, R.P., Lindroth, J.R., and Wong, Y.-M.M. (1990) Regional differences in DNA-synthetic activity in the preseptation myocar- dium of the chick. In: “Developmental Cardiology: Morphogenesis and Function,” Clark, E.B. and A. Takao, A. (eds). New York: Fu- tura Publishing, Inc., pp 219-234.
Vassall-Adams, P.R. (1982) The development of the atrioventricular bundle and its branches in the avian heart. J. Anat. 134:169-183.
Wenink, A.C.G. (1976) Development of the human cardiac conducting system. J. Anat. 121:617-631.
Wessels, A., Vermeulen, J.L.M., Verbeek, F.J., Viragh, S.Z., Kalman, F., Lamers, W.H., and Moorman, A.F.M. (1992) Spatial distribution of “tissue-specific” antigens in the developing human heart and skeletal muscle 111. Anat. Rec. 232:97-111.
Wilkinson, D.G., Bailes, J.A., Champion, J.E., and McMahon, A.P. (1987) A molecular analysis of mouse development from 8 to 10 days post coitum detects changes only in embryonic globin. Devel- opment 99:493-500.
Yokouchi, Y., Sasaki, H., and Kuroiwa, A. (1991) Homeobox gene expression correlated with the bifurcation process of limb cartilage development. Nature 353:443 -445.
Copyright © 2022 FDOKUMEN




















