
Expression of chimeric antigen receptors in natural killer cells with a regulatory-compliant...
20
Expression of chimeric antigen receptors in natural killer cells with a regulatory-compliant non-viral method Linhong Li, Ph.D 1 , Linda N Liu, Ph.D 1 , Stephanie Feller, MS 1 , Cornell Allen, MS 1 , Rama Shivakumar, MS 1 , Joseph Fratantoni, MD 1 , Lawrence A. Wolfraim, Ph.D 1 , Hiroyuki Fujisaki, M.D., Ph.D. 2 , Dario Campana, M.D., Ph.D. 2 , Nicholas Chopas, Sergey Dzekunov, Ph.D, and Madhusudan Peshwa, Ph.D 1 1 MaxCyte Inc, 22 Firstfield Rd, Suite 250, Gaithersburg, Maryland, USA 2 Departments of Oncology and Pathology, St Jude Children’s Research Hospital, Memphis, TN, USA Abstract Natural killer 1 cells hold promise for cancer therapy. NK cytotoxicity can be enhanced by expression of chimeric antigen receptors that re-direct specificity toward target cells by engaging cell surface molecules expressed on target cells. We developed a regulatory-compliant, scalable non-viral approach to engineer NK cells to be target-specific based on transfection of mRNA encoding chimeric receptors. Transfection of eGFP mRNA into ex vivo expanded NK cells (N=5) or purified unstimulated NK cells from peripheral blood (N=4) resulted in good cell viability with eGFP expression in 85% ± 6% and 86% ± 4%, 24 hours after transfection, respectively. An mRNA encoding a receptor directed against CD19 (anti-CD19-BB-z) was also transfected into NK cells efficiently. Ex vivo expanded and purified unstimulated NK cells expressing anti-CD19-BB-z exhibited enhanced cytotoxicity against CD19+ target cells resulting in ≥80% lysis of acute lymphoblastic leukemia and B-lineage chronic lymphocytic leukemia cells at effector target ratios lower than 10:1. The target-specific cytotoxicity for anti-CD19-BB-z mRNA-transfected NK cells was observed as early as 3 hours after transfection and persisted for up to 3 days. The method described here should facilitate the clinical development of NK-based antigen-targeted immunotherapy for cancer. Keywords NK cells; chimeric antigen receptor; electroporation; non-viral; transfection; mRNA Introduction The capacity of natural killer1 cells to exert cytotoxicity against a variety of cancer cell types makes them an attractive tool for anti-cancer therapy2 –7 . Data gathered in the setting of allogeneic hematopoietic stem cell transplantation (HSCT) indicate that donor selection based on the degree of mismatch between expression of killer immunoglobulin-like receptors (KIRs) on donor NK cells and HLA Class I molecules expressed by the patient cells should maximize NK cell killing of target cells 4, 8 – 10, hence augmenting the efficacy of HSCT6 , 7 , 11. In Address correspondence to: Linhong Li, Ph. D., MaxCyte Inc., 22 Firstfield Road, Suite 250, Gaithersburg, MD 20878, USA; Phone 301-944-1632, Fax 301-944-1703, [email protected]. MaxCyte employee: Linhong Li, Cornell Allen, Rama Shivakumar, Joseph Fratantoni, Lawrence A. Wolfraim and Madhusudan Peshwa. MaxCyte stock option holder: Linda N Liu and Stephanie Feller. No interest conflict with MaxCyte: Hiroyuki Fujisaki and Dario Campana. NIH Public Access Author Manuscript Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1. Published in final edited form as: Cancer Gene Ther. 2010 March ; 17(3): 147. doi:10.1038/cgt.2009.61. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Expression of chimeric antigen receptors in natural killer cells with a regulatory-compliant...

Expression of chimeric antigen receptors in natural killer cellswith a regulatory-compliant non-viral method
Linhong Li, Ph.D1, Linda N Liu, Ph.D1, Stephanie Feller, MS1, Cornell Allen, MS1, RamaShivakumar, MS1, Joseph Fratantoni, MD1, Lawrence A. Wolfraim, Ph.D1, Hiroyuki Fujisaki,M.D., Ph.D.2, Dario Campana, M.D., Ph.D.2, Nicholas Chopas, Sergey Dzekunov, Ph.D, andMadhusudan Peshwa, Ph.D11 MaxCyte Inc, 22 Firstfield Rd, Suite 250, Gaithersburg, Maryland, USA2 Departments of Oncology and Pathology, St Jude Children’s Research Hospital, Memphis, TN,USA
AbstractNatural killer1 cells hold promise for cancer therapy. NK cytotoxicity can be enhanced by expressionof chimeric antigen receptors that re-direct specificity toward target cells by engaging cell surfacemolecules expressed on target cells. We developed a regulatory-compliant, scalable non-viralapproach to engineer NK cells to be target-specific based on transfection of mRNA encoding chimericreceptors. Transfection of eGFP mRNA into ex vivo expanded NK cells (N=5) or purifiedunstimulated NK cells from peripheral blood (N=4) resulted in good cell viability with eGFPexpression in 85% ± 6% and 86% ± 4%, 24 hours after transfection, respectively. An mRNA encodinga receptor directed against CD19 (anti-CD19-BB-z) was also transfected into NK cells efficiently.Ex vivo expanded and purified unstimulated NK cells expressing anti-CD19-BB-z exhibitedenhanced cytotoxicity against CD19+ target cells resulting in ≥80% lysis of acute lymphoblasticleukemia and B-lineage chronic lymphocytic leukemia cells at effector target ratios lower than 10:1.The target-specific cytotoxicity for anti-CD19-BB-z mRNA-transfected NK cells was observed asearly as 3 hours after transfection and persisted for up to 3 days. The method described here shouldfacilitate the clinical development of NK-based antigen-targeted immunotherapy for cancer.
KeywordsNK cells; chimeric antigen receptor; electroporation; non-viral; transfection; mRNA
IntroductionThe capacity of natural killer1 cells to exert cytotoxicity against a variety of cancer cell typesmakes them an attractive tool for anti-cancer therapy2–7. Data gathered in the setting ofallogeneic hematopoietic stem cell transplantation (HSCT) indicate that donor selection basedon the degree of mismatch between expression of killer immunoglobulin-like receptors (KIRs)on donor NK cells and HLA Class I molecules expressed by the patient cells should maximizeNK cell killing of target cells4,8–10, hence augmenting the efficacy of HSCT6,7,11. In
Address correspondence to: Linhong Li, Ph. D., MaxCyte Inc., 22 Firstfield Road, Suite 250, Gaithersburg, MD 20878, USA; Phone301-944-1632, Fax 301-944-1703, [email protected] employee: Linhong Li, Cornell Allen, Rama Shivakumar, Joseph Fratantoni, Lawrence A. Wolfraim and Madhusudan Peshwa.MaxCyte stock option holder: Linda N Liu and Stephanie Feller.No interest conflict with MaxCyte: Hiroyuki Fujisaki and Dario Campana.
NIH Public AccessAuthor ManuscriptCancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
Published in final edited form as:Cancer Gene Ther. 2010 March ; 17(3): 147. doi:10.1038/cgt.2009.61.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

addition, it was reported that the infusion of haploidentical NK cells in a non-myeloablativetransplant setting could produce remissions in patients with acute myeloid leukemia5. AlthoughNK cell cytotoxicity has a wide spectrum, some cancer cell types appear less susceptible orrefractory to NK cell killing, because of failure to activate NK cells, induction of suppressionor both. Among these relatively NK-resistant cell types are lymphoid malignancies such asacute lymphoblastic leukemia (ALL), B-cell chronic lymphocytic leukemia (B-CLL) and B-cell non-Hodgkin lymphoma. 3,12–16
Chimeric antigen receptor has been studied since late 80’s17–22. It generally contains a singlechain variable fragment (scFv) as the extracellular antigen recognition unit and multiplelymphocyte activation domains as the intracellular activation part. Most work has been focusedin arming T cells with this chimeric antigen receptor for anti-tumor effect21–23. NK cellstransduced with chimeric antigen receptor have also been exploited for anti-tumor effect.Various combinations of intracellular activation domains were studied. The inclusion of bothsignal molecules of CD3ξ and 41BB was found not only to be optimal for mediating moreactivation signals, and producing higher IFN-γ and GM-CSF by NK cells after contacting toantigen-specific target cells18, but also to help NK cells to both direct their reactivity towardsspecific cell types and significantly augment their cytotoxicity19,20,22,24–26, Thus,expression of a receptor binding CD19, a surface molecule widely expressed in B cellmalignancies, and delivering activation signals through CD3z and 4-1BB (anti-CD19-BB-z)could overcome KIR-mediated inhibition and render NK cell highly cytotoxic against ALLcells19. Therefore, the development of a practical and efficient method for large-scaleexpression of chimeric antigen receptors in NK cells should have important clinicalapplications.
Electroporation is an established and efficient non-viral method for loading molecules into awide variety of cell types27–31. We here report a practical scalable method to efficientlytransfect expanded and purified unstimulated NK cells through electroporation of mRNAencoding the anti-CD19-BB-z chimeric antigen receptor. With this method, we observed a hightransfection efficiency and high cell killing of CD19-positive target cells. This method can beapplied to chimeric receptors with different specificities, is suitable for large-scale transfectionand it should be easily translatable to a clinical setting.
Materials and MethodsCells
The CD19+ human B-lineage ALL cell line, OP-132, and the genetically engineered myeloidleukemia cell line, K562, co-expressing 4 1BB ligand and membrane bound IL-1518 weregenerated at St Jude Children’s Research Hospital (Memphis, TN) and were maintained inRPMI-1640 supplemented with 10% fetal bovine serum and antibiotics. Leukemic cells wereobtained from the peripheral blood of patients with B-CLL with appropriate informed consentand approval (George Washington Hospital). Peripheral blood from healthy donor cells wasobtained from BRT Laboratories, Inc (Baltimore, MD). Peripheral blood mononuclear cells(PBMC) were prepared with Ficoll density gradient centrifugation, washed twice withphosphate buffered saline (PBS), and frozen in liquid nitrogen until use.
Purified unstimulated NK cells were selected from PBMC according to the manufacture’sprotocol using a Miltenyi NK cell isolation kit (Auburn, CA) and frozen in liquid nitrogen untiluse. NK cell expansion was achieved as previously described by Imai et al18. Briefly, peripheralblood mononuclear cells were cultured with irradiated and thawed K562 cells co-expressing4-1BB ligand and membrane-bound IL-15. Cultures were performed in the presence of 10 IU/ml-100 U/ml IL-2 in the presence of 10% FBS and antibiotics.
Li et al. Page 2
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Gene constructs used for electroporationA transgene encoding anti-CD19 single chain variable region conjugated with the 4-1BBintracellular domain and the CD3z domain was cloned from the parental plasmid pMSCVanti-CD19BBZ19 into pVAX1 (Invitrogen, Carlsbad, CA). The cloning was performed by digestingthe parental plasmid pMSCVanti-CD19BBZ and the pVAX1 vector with EcoR I and Xho Iand ligating using T4 DNA ligase (NEW ENGLAND BioLab, Beverly, MA). The clonedpVAX1-αCD19BBZ was used as a template for in vitro mRNA transcription after linearizationwith XbaI. The mRNA encoding anti-CD19-BB-z (mRNA anti-CD19-BB-z) was transcribedin vitro with T7 RNA polymerase using the Ambion mMESSAGE mMACHINE T7 Ultra kit(Ambion, Austin, TX). The mRNA quality and quantity was analyzed by 1% agarose gelelectrophoresis after 15 min denaturation at 70°C in mRNA denaturation buffer (Invitrogen,Carlsbad, CA) and quantitation by UV spectrophotometry (OD260/280). The mRNA encodingeGFP (mRNA-GFP) was transcribed in vitro using template of Cla I-linearized pCI-eGFPplasmid cloned into the pCI backbone (Promega, Madison, WI) and the same Ambion kit asmentioned above.
ElectroporationPurified unstimulated NK cells were thawed in a 37°C water bath and incubated for 0.5–1h at37°C in prewarmed fresh complete medium (RPMI-1640+10% FBS+ antibiotics) beforetransfection. Expanded NK cells were harvested for transfection 6–12 days after expansion.After collection, both expanded and purified unstimulated NK cells were washed once withEP buffer (MaxCyte, Gaithersburg, MD), mixed with 100μg/ml mRNA (unless specified, at1-3e8 cells/ml), transferred into MaxCyte processing chamber (Gaithersburg, MD), andtransfected using the program “Expanded-NK#3” and “unstimulated-NK#1” for expanded andpurified unstimulated NK cells, respectively, using a MaxCyte GT system (MaxCyte,Gaithersburg, MD). Immediately after electroporation, cells were recovered from theprocessing chamber and incubated for 20 min at 37°C, and then resuspended and cultured incomplete medium. Expression was analyzed by flow cytometry using a FACSCalibur andCellQuest software (Becton Dickinson, San Jose, CA).
The MaxCyte GT system (MaxCyte, Gaithersburg, MD) has capabilities for both static smallvolume (as mentioned above) and computer-controlled flow large volume electroporation31.The flow electroporation is achieved by computer-controlled repeated cycles ofelectroporation. Each cycle consists of three steps: flowing in a fraction of cells into flowchamber, transfection and flowing out the transfected cells for collection. If mRNA transfectioncould maintain the similar transfection efficiency from fraction to fraction, mRNA transfectioncould be scaled up. About 10–20 transfection cycles with 4ml/cycle (holding capacity forMaxCyte current chamber) could achieve relevant volume for clinical applications. However,considering the cost in primary cell expansion and mRNA production, using not too high cellvolume for transfection to obtain meaningful scale-up result is pursued in this study. Becauseof this, a mini-flow chamber with 0.8ml holding-volume capacity (manuscript in preparation)was designed, aiming at achieving 10–20 fractions in flow electroporation with about 10mlcell volume. NK cells expanded for 10 days were washed twice with 10 ml of EP buffer(MaxCyte, Gaithersburg, MD), resuspended with EP buffer to a diluted cell concentration (5–14ml at 4e7 cells/ml), added with 20μg/ml final concentration of mRNA encoding GFP, andflowed through the mini flow-chamber for transfection with the MaxCyte GT system using theprogram “Expanded-NK#1-Flow”. The transfected cells were collected by fractions with ≅0.8ml/fraction during the flow electroporation, incubated for 20min in the same EP buffer foreach fraction, cultured and analyzed by FACS 6h-1d post transfection. Small volume of themRNA-cell mixture was set aside for regular small volume static electroporation using“expanded-NK#3” program (mentioned above) to compare the scale-up efficiency.
Li et al. Page 3
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Detection of chimeric antigen receptor expression and immunophenotypingTo detect anti-CD19-BB-z expression, NK cells were stained with goat anti-mouse (Fab)2polyclonal antibody conjugated with biotin (Jackson Immuno Research Labs, West Grove, PA)followed by staining with peridinin chlorophyll protein-(PerCp; Becton Dickinson, San Jose,CA) labeled streptavidin. Cells stained with goat biotin-conjugated IgG followed bystreptavidin-PerCp were used for negative gating.
The following antibodies were used for immunophenotypic characterization of NK cells: anti-CD3 conjugated with fluorescein isothiocyanate (FITC), anti-CD19 conjugated withphycoerythrin (PE), anti-CD16-PE, and anti-CD56-PE.
Cell killing assayTo facilitate a high throughput analysis of cell killing, we developed a simple non-radioactivecell killing assay that is based on FACS analysis combined with acetoxymethyl-calcein(calcein-AM, Molecular Probes, Eugene, OR)-labeled target cells (manuscript in preparation).Briefly, calcein-AM pre-labeled target cells (105 cells in 100μl) were co-cultured with 100 μlof either transfected, non-transfected NK cells or just fresh medium at various E:T ratios ineach well of a 96-well U-bottom tissue culture plate in triplicates (Costar, Cambridge, MA).The 96-well plate was centrifuged at 400g for 5 min prior to cell culture in a 37°C, 5% CO2incubator. The cells were resuspended and transferred to 4mL FACS tubes for FACS analysisat the indicated time points.
In some experiments, a previously reported antibody staining method was used for measuringcell killing19. Briefly, unstained target cells (105 cells in 100μl) were co-cultured with 100 μlof either transfected, non-transfected NK cells or just fresh medium at various E:T ratios ineach well of a 96-well U-bottom tissue culture plate in triplicates (Costar, Cambridge, MA).After centrifugation at 400g for 5 min, the cells were cultured for the indicated cell-killingtime. The cells were harvested and stained with anti-CD56-PE and/or anti-CD19-FITCantibodies for 20 min on ice. After washing in PBS, the cells were resuspended with 200 μl ofPBS and analyzed by flow cytometry.
Cell killing assays were performed in the absence of exogenous IL-2. During acquisition, theFACS collection time was kept constant at 15 seconds. The specific cell lysis (%) wascalculated using the following formula:
where NTarget is the number of viable target cells cocultured with NK cells and NControl is thenumber of viable target cells cultured alone, in the absence of NK cells. Cell killing wasassessed at 4 h, unless specified otherwise.
Statistic analysisData was presented as mean ± standard deviation. Unpaired Student’s t-test (Microsoft Excel)with two tails was used to determine the significance of results. Statistical significance wasdetermined as P < 0.05.
ResultseGFP transfection of human NK cells
In our first set of experiments, we tested whether eGFP could be expressed in NK cells aftertransfection of the encoding mRNA mediated by electroporation. Percentage of GFP-positive
Li et al. Page 4
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NK cells after transfection was high, ranging from 81% to 92% (86% ± 5%) in experimentswith purified unstimulated NK cells (n =3) and with NK cell expanded for 6–12 days bycoculture with K562-mb15-41BBL cells (n = 3) (Fig. 1a and b). Transfection did notsignificantly affect cell viability (Fig. 1a and 1b). The mRNA-transfected cells had a decreasedproliferation rate for the first day post transfection, but they typically recovered and exhibitedsimilar proliferation rates as control cells later. The transfected NK cells expanded to 58% ±7% (n = 4) of the cell numbers of control cells during the 4-day culturing period posttransfection. The mRNA concentration in the range of 25μg/ml to 200μg/ml resulted in similarcell post-EP proliferation (data not shown).
Transfection of the anti-CD19-BB-z chimeric receptor in NK cellsThe effect of NK cell transfection on viability and expression efficiency was further studiedusing mRNA encoding a functional transgene, anti-CD19-BB-z. As shown in Fig 1c, anti-CD19-BB-z mRNA was transfected efficiently into expanded and purified unstimulated NKcells. Expression was dose dependent, increasing with the increase of the used mRNAconcentration from 25μg/ml to 200μg/ml (data not shown). Transfection at day 1 posttransfection did not dramatically affect the percentage of viable cells as gated in the R1 regionin this typical transfection result, with viabilities of control and transfected cells of 66% and54% for expanded and 67% and 64% for unstimulated NK cells, respectively. As shown in Fig1c, a clear anti-CD19-BB-z expression was achieved in both expanded and purifiedunstimulated NK cells (Fig 1d). Percentage of anti-CD19BB-z-positive NK cells aftertransfection was high, ranging from 33% to 81% (58% ± 12%) in experiments withunstimulated NK cells (n = 5) and with NK cell expanded for 6–12 days by coculture withK562-mb15-41BBL cells (n = 10) (Fig. 1c and d). Expression of anti-CD19-BB-z in expandedNK cells decreased with time but remained detectable up to day 4 post-transfection (data notshown). In unstimulated NK cells, anti-CD19-BB-z expression for the observed 2 dayexpression was detected with a 20%-30% decrease from day 1 to day 2. Similar cellproliferation characteristics were observed for anti-CD19-BB-z and eGFP transfected cells inthe case of expanded NK cells (data not shown).
To demonstrate the feasibility of scalability of mRNA transfection, we report here a novelcomputer-controlled flow electroporation of mRNA, in which the expanded NK cells wereelectroporated when they were pumped to flow through the flow chamber. The result wasshown in Fig 2. The transfection efficiency for collected fractions after flow electroporationand samples after static electroporation (30μl each, n=4) led to a range of 92%–98% and 94%± 1% in GFP expression respectively. There is basically no decrease in transfection efficiencyfrom fraction to fraction within 18 fractions. Also, the transfection efficiency of each fractionin flow electroporation is comparable to that of samples in 30μl static electroporation. It wasa successful scale up transfection from 30ul to 14ml, a 400–500 folders increase. Our furtherstudy will need to overcome the cost restriction for mRNA production and the primary cellexpansion so that we could exploit further scale up study with chamber of 3.5ml holding-volume capacity and routine cell concentration of 1-3e8 cells/ml, which could lead to≥1×1010 cell processing capacity at the current 18 fraction flow electroporation level.
Leukemic cell killing by NK cells transfected with anti-CD19BB-zExpanded NK cells transfected with anti-CD19-BB-z effectively and specifically lysed CD19+ ALL cells. By contrast, NK cells transfected with GFP and NK cells electroporated withoutmRNA (EP alone) exerted no significant cytotoxicity over that exerted by non-electroporatedNK cells (Fig 3a and b). The degree of ALL cell lysis by NK cells expressing anti-CD19-BB-z receptors depended on the concentration of mRNA used. As shown in Fig 4, cell killing wasseen as early as 3h post transfection (Fig 4a-1 and 2). A 4-hour coculture resulted inapproximately 80% lysis of the OP-1 cells at E: T ratio of 1: 2;16-hour coculture led to almost
Li et al. Page 5
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

100% lysis of the OP-1 cells at the same 1:2 ratio. NK cells transfected with anti-CD19-BB-zmRNA maintained their specific killing activity for up to 3 days post-transfection (Fig 4b). Onday 3 post-transfection, anti-CD19-BB-z mRNA-transfected cells could still lyse 80% to 90%of the target OP-1 cells in a 4-hour assay at E: T ratio of 2: 1 and 4: 1, respectively (Fig 4b).
When purified unstimulated NK cells were transfected with anti-CD19-BB-z mRNA, they alsobecame highly cytotoxic to ALL cells, although to a less extent than expanded activated NKcells (Fig. 5). About 80% of the target cells were lysed in the 4-hour assay at an E: T ratio of8:1. The naive (no EP) and GFP mRNA-transfected unstimulated NK cells did not lyse targetcells. Purified unstimulated NK cells from both donors demonstrated similar killing ability.
We next assessed whether anti-CD19-BB-z-transfected NK cells, either expanded orunstimulated, could kill primary B-lineage leukemia cells. Fig 6 shows the killing of CLL cellsfrom two donors using either expanded NK cells from one donor (Fig 6a) or purifiedunstimulated NK cells from three donors (Fig 6b). The killing efficiency for expanded NKcells was greater than that for purified unstimulated NK cells. At E:T ratio of 2:1 and 1:1, anti-CD19-BB-z-transfected expanded NK cells could lyse about 80%–90% of target CLL cells;whereas, purified unstimulated NK cells could reach this killing level only at an 8:1 E:T ratio.
DiscussionIn this study, we developed a scalable method for efficient mRNA transfection in human NKcells. When an anti-CD19 chimeric antigen receptor was expressed using this method, NK cellsacquired powerful and specific cytotoxicity against CD19+ leukemic cells. These resultsillustrate one of the potential applications of the technology, i.e. redirecting the specificity ofNK cells towards specific tumor cell types and augmenting their specific cytotoxicity.
Even though plasmid DNA electroporation has been most frequently used for electroporation,DNA uptake mediated by electroporation results in toxicity to many cells, includingunstimulated and expanded hematopoietic cells33,34, which hinders its application in primarycells35–42. A great deal of attention has been dedicated to develop a method that could not onlyachieve high transfection efficiency but also preserve cell viability. The utility of mRNAtransfection has recently been recognized in the development of cell-based therapeuticagents43 since it helps maintaining a high cell viability. The successful mRNA transfection ofNK cells in the current report added to the mRNA transfection of dendritic cells41,44, primaryresting leukemia cells45, CD34 hematopoeitic stem cells46 and primary T and B cells47 anddemonstrated the power of this nonviral method41,42,45,47,48. Unlike DNA, mRNA is unableto integrate into the host cell genome. Retroviral vectors can mediate effective transduction ofchimeric antigen receptor in NK and T cells18,23–25. Although effective, this strategy islaborious. With the method described here, high levels of receptor expression can be achievedwithin hours in a single-step procedure. While mRNA transfection avoids integration into thehost cell genome, this advantage is balanced by the possible limitation associated with transientexpression of non-integrated genetic material. Whether transient expression and short-term (upto one week) target cell killing are sufficient to have significant impact on in vivo tumor cellkilling and tumor burden is an important consideration.
The simplicity, safety, cost effectiveness and efficiency of the method described here togenetically modify NK cells is very well suited for clinical applications. An immediateapplication that we envisage is the redirection of NK cells from donors towards moleculesexpressed in the tumor cells of the recipient. Thus, purified NK cells from apheresis productswould be transfected with mRNA encoding a chimeric antigen receptor specific for the targetmolecule (e.g., CD19 in case of B-cell malignancies). The modified NK cells would then be
Li et al. Page 6
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ready for infusion after a few hours. If larger numbers of NK cells are required and/or if multipleinfusions are planned, expanded NK cells could be used instead.
AcknowledgmentsSpecial thanks are due to Nick Chopas for his help in equipment preparation for the study. The authors want to expresstheir gratitude to Dr. Gary Moroff and Ms. Amy Neuschaferand at AMERICAN RED CROSS HOLLAND LAB(15601 Crabbs Branch Way, ROCKVILLE, MD) for training and usage of a Gammacell 1000 irradiator.
References1. Schroers R, Hildebrandt Y, Hasenkamp J, et al. Gene transfer into human T lymphocytes and natural
killer cells by Ad5/F35 chimeric adenoviral vectors. Exp Hematol 2004;32:536–546. [PubMed:15183894]
2. Ruggeri L, Capanni M, Urbani E, et al. Effectiveness of donor natural killer cell alloreactivity inmismatched hematopoietic transplants. Science 2002;295:2097–2100. [PubMed: 11896281]
3. Ruggeri L, Mancusi A, Burchielli E, Aversa F, Martelli MF, Velardi A. Natural killer cell alloreactivityin allogeneic hematopoietic transplantation. Curr Opin Oncol 2007;19:142–147. [PubMed: 17272987]
4. McKenna DH Jr, Sumstad D, Bostrom N, et al. Good manufacturing practices production of naturalkiller cells for immunotherapy: a six-year single-institution experience. Transfusion 2007;47:520–528.[PubMed: 17319835]
5. Miller JS, Soignier Y, Panoskaltsis-Mortari A, et al. Successful adoptive transfer and in vivo expansionof human haploidentical NK cells in patients with cancer. Blood 2005;105:3051–3057. [PubMed:15632206]
6. Farag SS, Fehniger TA, Ruggeri L, Velardi A, Caligiuri MA. Natural killer cell receptors: new biologyand insights into the graft-versus-leukemia effect. Blood 2002;100:1935–1947. [PubMed: 12200350]
7. Chiorean EG, Miller JS. The biology of natural killer cells and implications for therapy of humandisease. J Hematother Stem Cell Res 2001;10:451–463. [PubMed: 11522229]
8. Lanier LL. Activating and inhibitory NK cell receptors. Adv Exp Med Biol 1998;452:13–18. [PubMed:9889954]
9. Passweg JR, Tichelli A, Meyer-Monard S, et al. Purified donor NK-lymphocyte infusion to consolidateengraftment after haploidentical stem cell transplantation. Leukemia 2004;18:1835–1838. [PubMed:15457184]
10. Schellekens J, Rozemuller EH, Petersen EJ, van den Tweel JG, Verdonck LF, Tilanus MG. ActivatingKIRs exert a crucial role on relapse and overall survival after HLA-identical sibling transplantation.Mol Immunol 2008;45:2255–2261. [PubMed: 18243319]
11. Klingemann HG. Natural killer cell-based immunotherapeutic strategies. Cytotherapy 2005;7:16–22.[PubMed: 16040380]
12. Kolb HJ, Schattenberg A, Goldman JM, et al. Graft-versus-leukemia effect of donor lymphocytetransfusions in marrow grafted patients. Blood 1995;86:2041–2050. [PubMed: 7655033]
13. Verdonck LF, Petersen EJ, Lokhorst HM, et al. Donor leukocyte infusions for recurrent hematologicmalignancies after allogeneic bone marrow transplantation: impact of infused and residual donor Tcells. Bone Marrow Transplant 1998;22:1057–1063. [PubMed: 9877267]
14. Collins RH Jr, Goldstein S, Giralt S, et al. Donor leukocyte infusions in acute lymphocytic leukemia.Bone Marrow Transplant 2000;26:511–516. [PubMed: 11019840]
15. Gratwohl A, Brand R, Apperley J, et al. Allogeneic hematopoietic stem cell transplantation for chronicmyeloid leukemia in Europe 2006: transplant activity, long-term data and current results. An analysisby the Chronic Leukemia Working Party of the European Group for Blood and MarrowTransplantation (EBMT). Haematologica 2006;91:513–521. [PubMed: 16533723]
16. Abbott BL. Recent advances in chronic lymphocytic leukemia. Cancer Invest 2006;24:302–309.[PubMed: 16809159]
17. Eshhar Z. The T-body approach: redirecting T cells with antibody specificity. Handb Exp Pharmacol2008:329–342. [PubMed: 18071952]
Li et al. Page 7
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

18. Imai C, Iwamoto S, Campana D. Genetic modification of primary natural killer cells overcomesinhibitory signals and induces specific killing of leukemic cells. Blood 2005;106:376–383. [PubMed:15755898]
19. Imai C, Mihara K, Andreansky M, et al. Chimeric receptors with 4-1BB signaling capacity provokepotent cytotoxicity against acute lymphoblastic leukemia. Leukemia 2004;18:676–684. [PubMed:14961035]
20. Biagi E, Marin V, Giordano Attianese GM, Dander E, D’Amico G, Biondi A. Chimeric T-cellreceptors: new challenges for targeted immunotherapy in hematologic malignancies. Haematologica2007;92:381–388. [PubMed: 17339188]
21. Cooper LJ, Ausubel L, Gutierrez M, et al. Manufacturing of gene-modified cytotoxic T lymphocytesfor autologous cellular therapy for lymphoma. Cytotherapy 2006;8:105–117. [PubMed: 16698684]
22. Finney HM, Akbar AN, Lawson AD. Activation of resting human primary T cells with chimericreceptors: costimulation from CD28, inducible costimulator, CD134, and CD137 in series withsignals from the TCR zeta chain. J Immunol 2004;172:104–113. [PubMed: 14688315]
23. Brentjens RJ, Santos E, Nikhamin Y, et al. Genetically targeted T cells eradicate systemic acutelymphoblastic leukemia xenografts. Clin Cancer Res 2007;13:5426–5435. [PubMed: 17855649]
24. Kowolik CM, Topp MS, Gonzalez S, et al. CD28 costimulation provided through a CD19-specificchimeric antigen receptor enhances in vivo persistence and antitumor efficacy of adoptivelytransferred T cells. Cancer Res 2006;66:10995–11004. [PubMed: 17108138]
25. Brentjens RJ. Novel approaches to immunotherapy for B-cell malignancies. Curr Hematol Rep2005;4:64–72. [PubMed: 15610662]
26. Loskog A, Giandomenico V, Rossig C, Pule M, Dotti G, Brenner MK. Addition of the CD28 signalingdomain to chimeric T-cell receptors enhances chimeric T-cell resistance to T regulatory cells.Leukemia 2006;20:1819–1828. [PubMed: 16932339]
27. Zimmermann U. Electrical breakdown, electropermeabilization and electrofusion. Rev PhysiolBiochem Pharmacol 1986;105:176–256. [PubMed: 3541139]
28. Golzio M, Rols MP, Teissie J. In vitro and in vivo electric field-mediated permeabilization, genetransfer, and expression. Methods 2004;33:126–135. [PubMed: 15121167]
29. Hui, S.; Li, L. In vitro and ex vivo delivery of genes to cells by electroporation. In: Jaroszeski, MJ.;Gilbert, R.; Heller, R., editors. Electrically mediated delivery of molecules to cells:electrochemotherapy, electrogene therapy and transdermal delivery by electroporation. HumanaPress; Totowa, NJ: 2000. p. 157-172.
30. Xie TD, Sun L, Tsong TY. Study of mechanisms of electric field-induced DNA transfection. I. DNAentry by surface binding and diffusion through membrane pores. Biophys J 1990;58:13–19. [PubMed:2200534]
31. Li LH, Shivakumar R, Feller S, et al. Highly efficient, large volume flow electroporation. TechnolCancer Res Treat 2002;1:341–350. [PubMed: 12625759]
32. Manabe A, Coustan-Smith E, Kumagai M, et al. Interleukin-4 induces programmed cell death(apoptosis) in cases of high-risk acute lymphoblastic leukemia. Blood 1994;83:1731–1737.[PubMed: 8142640]
33. Li LH, Sen A, Murphy SP, Jahreis GP, Fuji H, Hui SW. Apoptosis induced by DNA uptake limitstransfection efficiency. Exp Cell Res 1999;253:541–550. [PubMed: 10585278]
34. Li LH, McCarthy P, Hui SW. High-efficiency electrotransfection of human primary hematopoieticstem cells. Faseb J 2001;15:586–588. [PubMed: 11259375]
35. Bell MP, Huntoon CJ, Graham D, McKean DJ. The analysis of costimulatory receptor signalingcascades in normal T lymphocytes using in vitro gene transfer and reporter gene analysis. Nat Med2001;7:1155–1158. [PubMed: 11590441]
36. Van Tendeloo VF, Willems R, Ponsaerts P, et al. High-level transgene expression in primary humanT lymphocytes and adult bone marrow CD34+ cells via electroporation-mediated gene delivery. GeneTher 2000;7:1431–1437. [PubMed: 10981672]
37. Lai W, Chang CH, Farber DL. Gene transfection and expression in resting and activated murine CD4T cell subsets. J Immunol Methods 2003;282:93–102. [PubMed: 14604544]
Li et al. Page 8
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

38. Trompeter HI, Weinhold S, Thiel C, Wernet P, Uhrberg M. Rapid and highly efficient gene transferinto natural killer cells by nucleofection. J Immunol Methods 2003;274:245–256. [PubMed:12609550]
39. Maasho K, Marusina A, Reynolds NM, Coligan JE, Borrego F. Efficient gene transfer into the humannatural killer cell line, NKL, using the Amaxa nucleofection system. J Immunol Methods2004;284:133–140. [PubMed: 14736423]
40. Li LH, Biagi E, Allen C, et al. Rapid and efficient nonviral gene delivery of CD154 to primary chroniclymphocytic leukemia cells. Cancer Gene Ther 2006;13:215–224. [PubMed: 16082377]
41. Landi A, Babiuk LA, van Drunen Littel-van den Hurk S. High transfection efficiency, geneexpression, and viability of monocyte-derived human dendritic cells after nonviral gene transfer. JLeukoc Biol. 2007
42. Van De Parre TJ, Martinet W, Schrijvers DM, Herman AG, De Meyer GR. mRNA but not plasmidDNA is efficiently transfected in murine J774A.1 macrophages. Biochem Biophys Res Commun2005;327:356–360. [PubMed: 15629470]
43. Sullenger BA, Gilboa E. Emerging clinical applications of RNA. Nature 2002;418:252–258.[PubMed: 12110902]
44. Holtkamp S, Kreiter S, Selmi A, et al. Modification of antigen-encoding RNA increases stability,translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 2006;108:4009–4017.[PubMed: 16940422]
45. Li Linhong CA, Shivakumar Rama, Jonathan M. Weiss, Joseph C. Fratantoni, Linda N. Liu. EfficientNon-Viral Gene Delivery to Primary, Leukemia B Cells by mRNA Transfection. Molecular Therapy2004;9:220.
46. Li, LH.; Shivakumar, R.; Allen, C., et al. Engineering stem cell products for enhanced potency andbiological function. Annual Conference of International Society of Cell Therapy; 2006; 2006.
47. Zhao Y, Zheng Z, Cohen CJ, et al. High-efficiency transfection of primary human and mouse Tlymphocytes using RNA electroporation. Mol Ther 2006;13:151–159. [PubMed: 16140584]
48. Rabinovich PM, Komarovskaya ME, Ye ZJ, et al. Synthetic messenger RNA as a tool for gene therapy.Hum Gene Ther 2006;17:1027–1035. [PubMed: 17007566]
Li et al. Page 9
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Li et al. Page 10
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig 1. Transfection of mRNA in expanded and purified unstimulated NK cellsa. FACS analysis of NK cells transfected with mRNA-GFP. Expanded NK cells (left panels)and purified unstimulated (right panels) NK cells are depicted. b. Summary of post-electroporation viability and transfection efficiency of expanded and unstimulated NK cellstransfected with mRNA-GFP. Results were obtained from two donors done in triplicates. cFACS analysis of transfection of mRNA-anti-CD19-BB-z into expanded (left panels) andunstimulated (right panels) NK cells. d. Summary of post-electroporation viability and anti-CD19-BB-z expression of expanded and unstimulated NK cells transfected with mRNAencoding anti-CD19-BB-z. Results of expanded NK cells were obtained from two donors,performed 10 times in total and unstimulated NK cells from four donors, performed 5 times intotal. The expressions of transfected cells are significantly higher than that of control (p<0.01).
Li et al. Page 11
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

The viabilities of transfected cells are insignificantly different from that of control cells(p>0.05), except one indicated in the figure.
Li et al. Page 12
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Li et al. Page 13
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig 2. Scale-up flow electroporation of mRNA encoding eGFPThe result of FACS analysis of 10-day expanded NK cells transfected with mRNA-GFP wasdepicted. NK cells were resuspended in 14 ml, flow electroporated, and collected with 18fractions of 0.8ml each. The viability, GFP+ cell percentage (%) and mean fluorescenceintensity (MFI) were represented for each fraction of the flow electroporated and samples (n=4)of the regularly electroporated (30μl for each electroporation). Results are representative ofthree independent experiments.
Li et al. Page 14
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Li et al. Page 15
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig 3. Specific killing of OP-1 cells by expanded NK cellsa. Cell killing analyzed by Calcein-AM method. b. Cell killing analyzed by CD19-PE antibodystaining method.
Li et al. Page 16
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
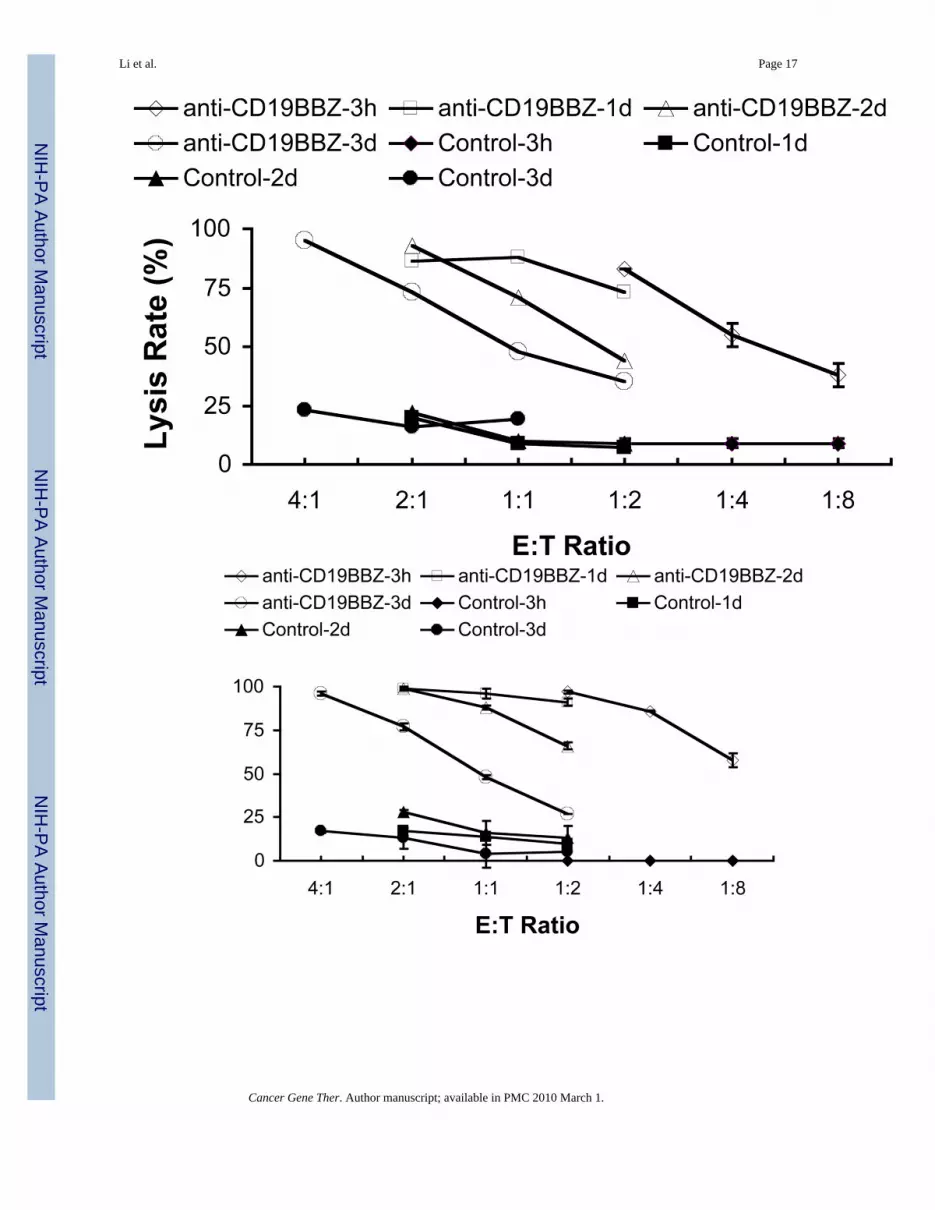
Li et al. Page 17
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig 4. Duration of specific OP-1 killing by expanded NK cells transfected with anti-CD19-BB-zmRNAa-1. Four hour killing assay was performed by calcein-AM method at 3h post transfection. NKcells were transfected with 0, 50, 100 and 200μg/ml anti-CD19-BB-z mRNA. a-2. 1d killingassay was performed by calcein-AM method at 3h post transfection. NK cells were transfectedwith 0, 50, 100 and 200 μg/ml anti-CD19-BB-z mRNA. b-1. Four hour killing assay wasperformed by calcein-AM method at 1d, 2d and 3d post transfection. NK cells were transfectedwith 100 μg/ml anti-CD19-BB-z mRNA. b-2. 1d killing assay was performed by calcein-AMmethod at 1d, 2d and 3d post transfection. NK cells were transfected with 100 μg/ml anti-CD19-BB-z mRNA. Results are representative of three independent experiments.
Li et al. Page 18
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Fig 5. Specific OP-1 cell killing by unstimulated NK cellsTransfected-NK cells (n=2) were tested for OP-1 cell killing 1d post transfection.
Li et al. Page 19
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
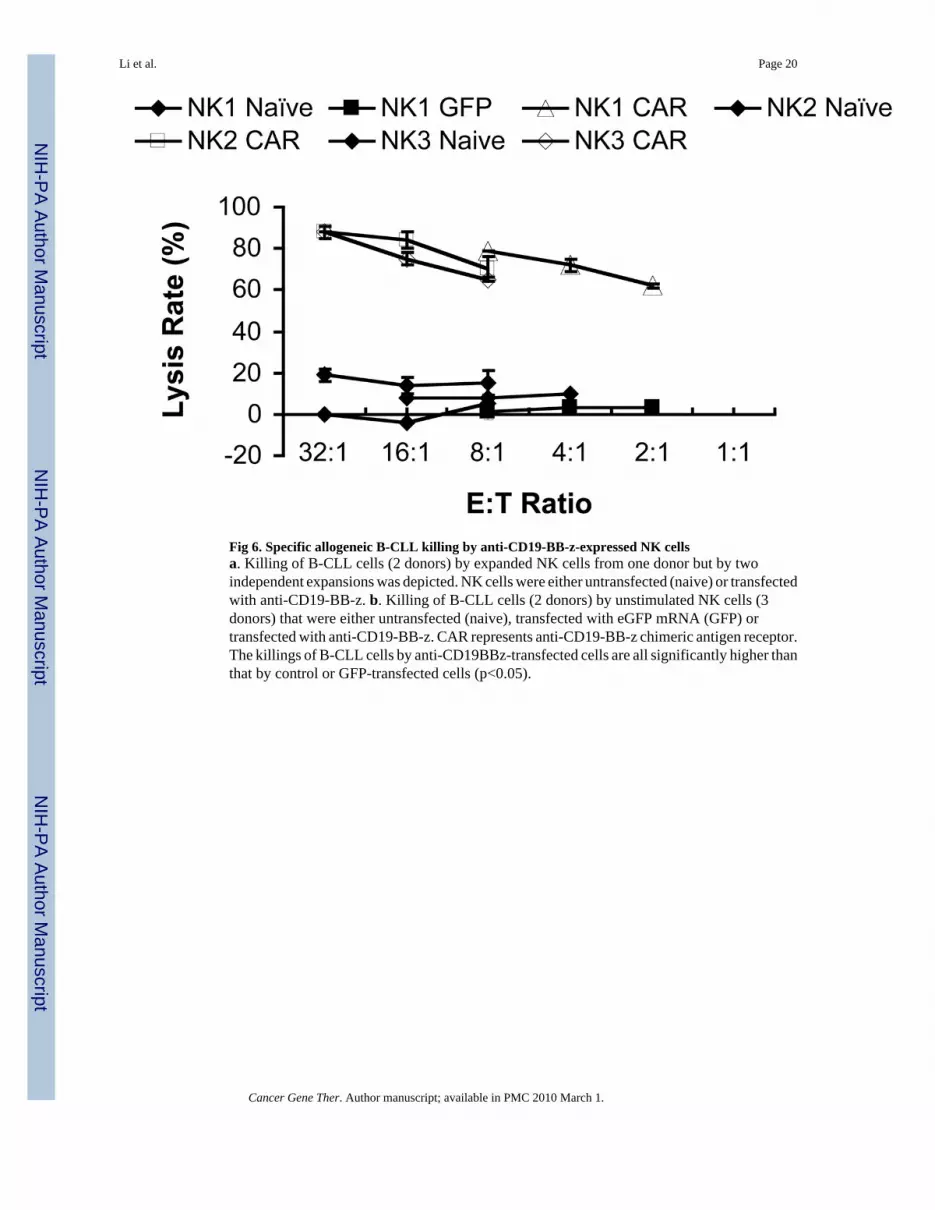
Fig 6. Specific allogeneic B-CLL killing by anti-CD19-BB-z-expressed NK cellsa. Killing of B-CLL cells (2 donors) by expanded NK cells from one donor but by twoindependent expansions was depicted. NK cells were either untransfected (naive) or transfectedwith anti-CD19-BB-z. b. Killing of B-CLL cells (2 donors) by unstimulated NK cells (3donors) that were either untransfected (naive), transfected with eGFP mRNA (GFP) ortransfected with anti-CD19-BB-z. CAR represents anti-CD19-BB-z chimeric antigen receptor.The killings of B-CLL cells by anti-CD19BBz-transfected cells are all significantly higher thanthat by control or GFP-transfected cells (p<0.05).
Li et al. Page 20
Cancer Gene Ther. Author manuscript; available in PMC 2010 March 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















