
Expression in Human Meningioma Cells - NCBI
10
American Journal of Pathology, Vol. 141, No. 3, September 1992 Copyight X) American Association of Pathologists Transforming Growth Factor-r3 and Transforming Growth Factor -receptor Expression in Human Meningioma Cells Mahlon D. Johnson,*t Charles F. Federspiel,t Leslie 1. Gold,11 and Harold L. Mosest From the Division of Neuropathology,* and the Departments of Cell Biologyt and Preventive Medicine,* Vanderbilt University Medical School, Nashville, Tennessee; and the Department of Pathology,11 New York University School of Medicine, New York, New York The transforming growth factor-a (TGFI3) family in mammals includes three closely relatedpeptides that influence proliferation and numerous physiologic processes in most mesenchymal cells. In this study, Northern blots, immunohistochemistry, TGFP ra- dioreceptor assays, TGFI receptor affinity labeling and 13H] thymidine incorporation were used to eval- uate whether primary cell cultures of human men- ingiomas synthesize the three TGFF isoforms, bear TGFP receptors, and respond to TGF. Transcripts for TGFf3l and 2 were detected in the three cases ana- lyzed Transforming growth factor-P1 immunoreac- tivity was detected in three of six cases, and TGFI2 and 3 immunoreactivity were detected in each case analyzed Media conditioned by cells cultured from six meningiomas also contained latent TGF4-like ac- tivity. Transforming growth factor-4 receptor cross- linking studies identified TGF3 binding sites corre- sponding to the type 1, type 2, and type 3 receptors on meningioma cells. Treatment with active TGF43I pro- duced a statistically significant reduction in [3H] thymidine incorporation after stimulation with 10% fetal calf serum and epidermal growth factor in all six cases studied (Am JPathol 1992, 141:633-642) Meningiomas constitute 25% of primary intracranial neo- plasms in humans.1' 2 Although typically benign, perineu- ral growth, growth at inoperable sites, and recurrence complicate traditional surgical management.1' 2 Resec- tion is even less effective as a treatment for malignant variants that characteristically invade brain parenchyma and occasionally metastasize.'12 Development of alterna- tive chemotherapy for these neoplasms has been hin- dered, in part, by the limited understanding of growth regulation in meningioma cells. Progesterone stimulation of meningioma growth has been suggested by numerous clinical studies3'4 and demonstrated in vitro.5 Other growth regulatory hor- mones, however, undoubtedly contribute to arachnoid neoplasia. Epidermal growth factor (EGF) and fibroblast growth factor stimulate human meningioma cell prolifer- ation in vitro.-7 Recent demonstration of EGF in human CSF8 as well as EGF/TGFa receptors, platelet-derived growth factor (PDGF), PDGF receptors, and basic fibro- blast growth factor (bFGF) mRNA in meningiomas9-13 raise the possibility that complex paracrine and autocrine stimulation by growth factors promote meningioma for- mation. Transforming growth factor 4-1 (TGFP1), TGFP2, and TGF433, another family of closely related peptides that regulate cell growth, may also participate. Transforming growth factor-p was originally identified as a peptide producing transformation of non-neoplastic cells in vitro, suggesting a role for this factor in oncogen- esis.1"16 Subsequent studies, however, have described more diverse effects of the TGF,Bs on mesenchymal cell proliferation. Although stimulatory effects have been demonstrated in fibroblasts, osteoblasts, and smooth muscle myocytes,17-18 growth inhibition has been noted in epithelial myeloid and lymphoid cells." The recent demonstration of transcripts and immunoreactivity for TGF,B1, TGF,2, and TGF43 in murine and human me- ninges1921 (M. D. Johnson and H. L. Moses, unpub- lished observations) raises the possibility that TGFp1, TGFB2, or TGF,B3 influence normal arachnoid cell growth. Conceivably, loss or changes in this regulation contribute to neoplastic growth. The current study evalu- ated TGF,B1, TGF,B2, and TGF,B3 expression and TGF,B receptor presence in primary cultures established from six human meningiomas. Additional experiments as- Supported by Veterans Administration Career Development Award (RA- 1793 MJ) and Merit Award (MJ). Accepted for publication March 3, 1992. Address reprint requests to Dr. Mahlon Johnson, Division of Neuro- pathology, Department of Pathology, Vanderbilt University Medical School, C3321 MCN, Nashville, TN 37232. 633
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Expression in Human Meningioma Cells - NCBI

American Journal ofPathology, Vol. 141, No. 3, September 1992Copyight X) American Association of Pathologists
Transforming Growth Factor-r3 andTransforming Growth Factor -receptorExpression in Human Meningioma Cells
Mahlon D. Johnson,*t Charles F. Federspiel,tLeslie 1. Gold,11 and Harold L. MosestFrom the Division ofNeuropathology,* and the Departmentsof Cell Biologyt and Preventive Medicine,* VanderbiltUniversity Medical School, Nashville, Tennessee; and theDepartment of Pathology,11 New York University School ofMedicine, New York, New York
The transforming growth factor-a (TGFI3) family inmammals includes three closely relatedpeptides thatinfluence proliferation and numerous physiologicprocesses in most mesenchymal cells. In this study,Northern blots, immunohistochemistry, TGFP ra-dioreceptor assays, TGFI receptor affinity labelingand 13H] thymidine incorporation were used to eval-uate whether primary cell cultures of human men-ingiomas synthesize the three TGFF isoforms, bear
TGFP receptors, and respond to TGF. TranscriptsforTGFf3l and 2 were detected in the three cases ana-lyzed Transforming growth factor-P1 immunoreac-tivity was detected in three of six cases, and TGFI2and 3 immunoreactivity were detected in each caseanalyzed Media conditioned by cells cultured fromsix meningiomas also contained latent TGF4-like ac-tivity. Transforming growth factor-4 receptor cross-linking studies identified TGF3 binding sites corre-sponding to the type 1, type 2, and type 3 receptors onmeningioma cells. Treatment with active TGF43I pro-duced a statistically significant reduction in [3H]thymidine incorporation after stimulation with 10%fetal calf serum and epidermal growth factor in allsix cases studied (AmJPathol 1992, 141:633-642)
Meningiomas constitute 25% of primary intracranial neo-plasms in humans.1' 2 Although typically benign, perineu-ral growth, growth at inoperable sites, and recurrencecomplicate traditional surgical management.1' 2 Resec-tion is even less effective as a treatment for malignantvariants that characteristically invade brain parenchymaand occasionally metastasize.'12 Development of alterna-
tive chemotherapy for these neoplasms has been hin-dered, in part, by the limited understanding of growthregulation in meningioma cells.
Progesterone stimulation of meningioma growth hasbeen suggested by numerous clinical studies3'4 anddemonstrated in vitro.5 Other growth regulatory hor-mones, however, undoubtedly contribute to arachnoidneoplasia. Epidermal growth factor (EGF) and fibroblastgrowth factor stimulate human meningioma cell prolifer-ation in vitro.-7 Recent demonstration of EGF in humanCSF8 as well as EGF/TGFa receptors, platelet-derivedgrowth factor (PDGF), PDGF receptors, and basic fibro-blast growth factor (bFGF) mRNA in meningiomas9-13raise the possibility that complex paracrine and autocrinestimulation by growth factors promote meningioma for-mation. Transforming growth factor 4-1 (TGFP1), TGFP2,and TGF433, another family of closely related peptidesthat regulate cell growth, may also participate.
Transforming growth factor-p was originally identifiedas a peptide producing transformation of non-neoplasticcells in vitro, suggesting a role for this factor in oncogen-esis.1"16 Subsequent studies, however, have describedmore diverse effects of the TGF,Bs on mesenchymal cellproliferation. Although stimulatory effects have beendemonstrated in fibroblasts, osteoblasts, and smoothmuscle myocytes,17-18 growth inhibition has been notedin epithelial myeloid and lymphoid cells." The recentdemonstration of transcripts and immunoreactivity forTGF,B1, TGF,2, and TGF43 in murine and human me-ninges1921 (M. D. Johnson and H. L. Moses, unpub-lished observations) raises the possibility that TGFp1,TGFB2, or TGF,B3 influence normal arachnoid cellgrowth. Conceivably, loss or changes in this regulationcontribute to neoplastic growth. The current study evalu-ated TGF,B1, TGF,B2, and TGF,B3 expression and TGF,Breceptor presence in primary cultures established fromsix human meningiomas. Additional experiments as-
Supported by Veterans Administration Career Development Award (RA-1793 MJ) and Merit Award (MJ).
Accepted for publication March 3, 1992.Address reprint requests to Dr. Mahlon Johnson, Division of Neuro-
pathology, Department of Pathology, Vanderbilt University MedicalSchool, C3321 MCN, Nashville, TN 37232.
633

634 Johnson et alAJP September 1992, Vol. 141, No. 3
sessed the effects of TGFp1 on quiescent, partially, andmaximally growth-stimulated meningioma cells. Findingsreported here suggest that meningiomas consistentlysynthesize and release all three isoforms of TGFP in alatent form and bear TGF, receptors. Treatment withTGF31 significantly reduced serum or EGF stimulation ofproliferation in these cultures.
Methods
Tumor Characteristics
Meningioma cell cultures were established from six pa-tients after a frozen section diagnosis was made on ad-jacent tissue by one of us (MJ). Tumor 1 was a convexitytransitional meningioma from a 43-year-old woman. Tu-mor 2 was an olfactory groove meningothelial meningio-ma from a 70-year-old woman. Tumor 3 was a menin-gothelial convexity meningioma of a 46-year-old woman.Tumor 4 was a frontal lobe fibroblastic meningioma froman 82-year-old woman. Tumor 5 was an orbital menin-gothelial meningioma from a 47-year-old man. Tumor 6was a convexity fibroblastic meningioma in a 1 6-year-old,previously irradiated boy. Tissue was procured followingprocedures approved by the Vanderbilt University Insti-tutional Review Board.
Meningioma Cell Cultures
Small fragments of tumor were collected in supple-mented Dulbecco's Modified Essential Media, then finelyminced, using 22-gauge needles. Meningioma cellswere seeded onto flasks and maintained in Dulbecco'smodified essential media supplemented with 10% fetalcalf serum (FCS), penicillin (50 units/ml), streptomycin (50,ug/ml), and Fungizone (Squibb, Princeton, NJ; 2.5 ,ug/ml). Cells were fed twice weekly. Additional cells wereplated onto sterile Lab Tech microscope slides (NUNC,Naperville, IL) for immunohistochemical studies.22
Meningioma Cell Characterization
For ultrastructural analysis, meningioma cells grown in35-mm dishes were fixed in 2% glutaraldehyde for 1hour, gently scraped, then pelleted by centrifugation. Thepellet was postfixed in 1% OS04 for 1 hour, then embed-ded in Epon (E. Fullam, Latham, NY). Thin sections werestained with uranyl acetate and lead citrate for ultrastruc-tural analysis on a Philipps 300 electron microscope.22
For immunohistochemical characterization, meningio-
ma cells were plated onto four-well microscope slides(Lab Tech) for 1 day in serum-free media, then fixed for10 minutes in freshly prepared 4% paraformaldehyde inphosphate buffer. Subsequent analysis was performedusing mouse monoclonal antibodies against human ep-ithelial membrane antigen (EMA, DAKO, Carpinteria,CA), and vimentin (DAKO) and the biotin-avidin-horseradish peroxidase method.23 Epithelial membraneantigen and vimentin immunoreactivity are detected inapproximately 50% and 18% of meningiomas, respec-tively.24
RNA Analysis by Northern Blot
Polyadenylated mRNA was derived from cells of conflu-ent meningioma cultures and TGFP-producing HT1080fibrosarcoma cells (American Type Culture Collection,Rockville, MD) using oligo (dT) cellulose as describedpreviously.25 Five micrograms mRNA was separated byelectrophoresis in 1.2% agarose gels containing formal-dehyde and transferred to nitrocellulose.26 Hybridiza-tions with 1 x 106 cpm/ml of labeled probe were per-formed at 420C in 5 x SSC with 50% formamide, 250,ug/ml sheared salmon sperm DNA, hybridization, blotswere washed in 1 x SSC with 0.1% sodium dodecylsulfate (SDS) and 0.1 x SSC and 0.1% SDS for 45 min-utes at 420C. Between hybridizations, the filters werestripped and reexposed to confirm removal of 32p-labeled cDNA.
cDNA probes for TGF13127 and cyclophilin,28 a con-stitutively expressed gene, were labeled using the ran-dom primer extension method.29 cRNA probes forTGF,B230 were transcribed using T7 polymerase.31 Be-cause of loss of RNA after multiple probe strippings, anal-ysis of TGF,33 expression was not undertaken.
Immunohistochemical Localizationof TGFr,-3The distribution of TGFP1-3 immunoreactivity was eval-uated using rabbit polyclonal antibodies against TGFI1-332 and avidin-biotin complex immunohistochemistry3performed on 5-,u-thick sections of the formalin-fixed,paraffin-embedded tumors and meningioma cell cul-tures, as outlined previously.34 Polyclonal antibodieswere produced in rabbits immunized with synthetic pep-tides of TGFP1, TGFP2, or TGF,B3. The TGFP1-3 antiseraexhibit no cross-reactivity between the TGFP isoforms onWestern blots.32 Preabsorption of each antiserum withthe peptide against which they are directed ablates theantiserum's immunoreactivity in tissue sections.32 Addi-

TGFIB1-3 and Receptors in Meningioma Cells 635AJP September 1992, Vol. 141, No. 3
tional characterization of these antisera are describedelsewhere.32
TGFI3 Radioreceptor Assay
Near confluent cultures of early-passage meningiomacells in T75 flasks were washed, then incubated for 72hours with 15 ml serum and growth factor-free 402 MCD,Bmedia. After collection, a portion of each sample was
acidified (pH 1.5) for 2 hours at room temperature, thenrestored to neutral pH. One-, 0.75-, 0.50-, 0.25-, and0.1 25-ml samples of neutral and acid-activated condi-tioned media were evaluated in triplicate for TGFI-competing activity by radioreceptor assay.3536 Briefly,84A cells were plated at a density of 2 x 105 cell per wellin six-well culture dishes and maintained in McCoy's 5amedium with 10% FCS for 1 day. The cells were subse-quently washed three times with phosphate-buffered sa-
line (PBS) containing 0.1% bovine serum albumin (BSA),then rocked at room temperature for 1 hour with buffer(128 mmol/I [millimolar] NaCI, 5.0 mmol/I MgSO4, 1.2mmol/I CaCI, 50.0 mmol/I Hepes, pH 7.5, 2 g/l BSA) be-fore incubation with known concentrations of pureTGF,B2, neutral or acid-activated conditioned media, and0.25 ng/ml 1251 TGFP2. 1251-TGFr31 and -132 can be usedinterchangeably in radioreceptor assays because bothbind all three receptors with essentially equal affinity.35After rocking at room temperature for 2 hours, the cellswere washed three times with PBS containing 0.1% BSA,the solubilized for 10 minutes in PBS with 1% TritonX-100. Results were expressed as percentage 1251
TGFP2 binding after correction for nonspecific binding.36
TGFI Receptor Crosslinking
A TGFI receptor crosslinking assay was performed on
cells from three meningiomas (M-4 through M-6) as de-scribed previously,37 with minor modifications. Briefly,cells were plated and grown to confluence in 100-mmdishes maintained in McCoy's media with 10% FCS as
described above. Ice-cooled plates were washed twicewith PBS then incubated rocking at 40C for 1 hour withbinding buffer. Subsequently, buffer was replaced withbinding buffer containing 2 ng/ml 1251 TGF1-1 without or
with 200 ng/ml unlabeled TGFP-1. After rocking at 40C for4 hours, the cells were washed with binding buffer with-out BSA then rocked at 4°C for 15 minutes with BSA-freebinding buffer containing 0.25 mmol/l disuccinimidylsuberrate. After scraping in dissociation buffer, cell mem-branes were solubilized by rocking at 4°C for 30 minutesin 1% Triton X-1 00. Extracts then were heated to 1 00°C
for 5 minutes in reducing buffer, and then electropho-resed on 5% to 7.5% polyacrylamide gels.
[3H] Thymidine Incorporation
In these studies, we evaluated the effects of TGF,B1 onquiescent, suboptimally, and optimally stimulated menin-gioma cells.6 Meningioma cells from M-1 through M-6were plated at a concentration of approximately 1 x 104cells/ml in 24-well dishes and grown to confluence inMcCoy's medium containing 10% FCS. Cultures thenwere incubated for 24 hours with serum-free MCDB 402media to achieve quiescence.
In the first study, TGFI1 effects were tested on cul-tures optimally stimulated with 10% FCS. Duplicate cellcultures were treated with MCDB 402 media alone orMCDB 402 containing 10% FCS, 10% FCS and TGF,B1(10 ng/ml), or TGFI1 (10 ng/ml) alone. At 12 hours, thesetreatments were replaced with [3H] thymidine 0.5 uCi/ml/well. Cell proliferation was stopped either 12, 24, or 36hours later by addition of 1 mol/l (molar) ascorbic acid.
In a second experiment, TGF1l effects were evalu-ated in suboptimally stimulated duplicate cell culturestreated with MCDB 402 media alone or MCDB 402 con-taining EGF (10 ng/ml) and insulin (500 ng/ml; Collabo-rative Res., New Bedford, MA), EGF, insulin, and TGF,B1(10 ng/ml; R&D, Minneapolis, MN) or TGF,1l (10 ng/ml)alone. At 12 hours, these treatments were replenishedwith identical media also containing [3H] thymidine0.5 ,uCi/ml/well. Cell proliferation was stopped with 1 mol/lascorbic acid as above.
For scintillation counting, the cells were solubilizedwith 1 N NaOH per well and rocked for 30 minutes, thencounted on a scintillation counter. For autoradiography,performed only on M-5 and M-6, cells were fixed in meth-anol (glacial acetic acid 3.1 followed by absolute meth-anol and 10% trichloroacetic acid for 10 minutes each.After washing in distilled H20, the cells were dried, thencoated with NTB2 emulsion (Kodak, Rochester, NY) andexposed for 5 days. The emulsion was developed withMicrodol (Kodak, Rochester, NY).
Percentage of control (MCDB 402 alone) values werecalculated using the logarithm of response for 10% FCS,10% FCS and TGFP1, and TGF,1l alone divided by thelog response for MCDB 402. These percentages of con-trol values were calculated for each of the six tumors ofeach of the three times, yielding the 54 values given in theresults section. Percentages of control values were cal-culated identically in the counterpart experiment, evalu-ating TGF,1l effects on EGF- and insulin-stimulated cells.Determinations for TGF,1l were not available for tumor,M-2; consequently there were 51 values.

636 Johnson et alAJP September 1992, Vol. 141, No. 3
Statistical analysis for the results was conducted us-ing the GLM procedures for analysis of variance, withcontrasts provided through SAS Statistical Software.3e Inaddition, nonparametric tests were used to analyze theeffects of TGFIil when given alone.
Results
Cytologic, Ultrastructural, andImmunohistochemical Characterization
Each meningioma culture was populated by large polyg-onal cells with irregular, often interdigitating borders andcentral oval nuclei with multiple chromocenters. Whorlformation was frequently seen. Typically, by passage 5 to6, senescence occurred, with cells exhibiting markedlydecreased proliferation-expanded cytoplasm and vacu-
olation of the nucleus. These findings are characteristic ofmeningioma cell cultures.22
Ultrastructurally, interdigitation of cell membranes anddesmosomes were identified in each meningioma cul-ture.
Extensive or moderate EMA immunoreactivity and ex-
tensive vimentin immunostaining were detected in thecells cultured from the meningiomas (data not shown).
Northern Blot Analysis
The 2.3- to 2.5-Kb TGF,1 transcript was detected in 5 ,ugpoly A + RNA from the control cells and each of the threemeningiomas tested (M-2, M-4, M-5) (Figure 1). In addi-tion, the 5.1 -Kb TGF,2 transcript was also detected in HT1080 cells and each of the meningiomas (Figure 1). The1.0-Kb cyclophilin transcript was used as a control forRNA loading and integrity.
TGFP Immunohistochemistry
Immunoreactivity detected for TGFP1, TGF02, andTGFPi3 in formalin-fixed meningioma tissue and meningi-oma cells is listed in Table 1. Transforming growth factorbeta-1 immunostaining was detected in tissue sectionsfrom two of six meningiomas examined. Immunoreactiv-ity was confined to the cytoplasm of tumor cells and wasnot seen in the blood vessels or dura. In cultured cells,extensive TGF,B cytoplasmic immunoreactivity was
seen in M-2 and scattered immunostaining was detectedin M4. Transforming growth factor P-1 immunostainingwas confined to the cytoplasm, often distributed in a dis-crete pattern suggesting Golgi localization. Loss of the
C
28S-
B
TGFB2
TGFBI
CYCLO
i8S-
A18S-
'I4 3 I
Figure 1. TGF1 and 32 mRNA in cultured human meningiomacells. Each lane contains approximately 5 pgpoly (A) + RIVAfiromHT1080fibrosarcoma cells known to express TGF1, TGF[2, andTGF133 (lane 1), TGF producing SW620 adenocarcinoma cells(lane 2) andH9 lymphoblasts (lane 3), M-2 (lane 4), M-4 (lane 5)and M-5 (lane 6). Filters were hybridized with 32-labeled cyclo-philin cDNA probe (A), 32P-labeled TGF131 cDNA (B), and 32P-labeled TGMFf2 riboprobe (C).
M-3 meningioma cells precluded immunocytochemicalanalysis of the other positive tumor.
Transforming growth factor beta-2 immunoreactivitywas detected in tissue sections from each of the six tu-mors and in cells cultured from five of the six tumors. Inthe tumors, TGFI2 immunostaining was distributed pri-marily in the cytoplasm of tumor cells. Scattered immu-noreactivity also was detected in the muscularis of largearterioles in one tumor. No distinct immunostaining was
detected in the dura. In cultured meningioma cells, ex-
tensive immunostaining was seen in M-1 and M-2, andscattered immunoreactivity was detected in cells fromM-4-M-6 confined to the cytoplasm, in a pattern similar tothat of TGF,Bl (Figure 2).
Transforming growth factor P-3 immunostaining was
seen also in tumor sections from each of the six menin-giomas and in the cultured meningioma cells from
Table 1. TGF Immunoreactivity in CulturedMeningioma Cells and Meningiomas
Meningioma TGFoI TGFP2 TGF,3
M-1 tumor - + +cells - + +
M-2 tumor + + +cells + + +
M-3 tumor + rare + +cells nd nd nd
M-4 tumor - - +cells + + +
M-5 tumor - + +cells - + +
M-6 tumor - + +cells - + +
+ = positive immunostaining in tumor cells; - = no distinctimmunostaining in tumor cells; nd = not done (no cells available).

TGFI1-3 and Receptors in Meningioma Cells 637AJP September 1992, Vol. 141, No. 3
Figure 2. a: TGF2 immunoreactivity in cellsfrom meningioma culture M-1. Distinct cytoplasmic immunostaining (red) waspresent inmany cells. AEC chromagen, Magnification, x240. b: TGF2 immunoreactivity in meningioma M-1. Cytoplasmic immunoreactivity was seenin cellular whorls throughout the tumor AEC chromagen magnification, X258
each of the five cases evaluated. In the tumor sections,moderate or extensive immunoreactivity was seen in tu-mor cells. Immunostaining was also seen in the media ofone tumor arteriole and in dural arterioles. No distinct im-munoreactivity was detected in the dura. In meningiomacells maintained in culture, TGF,3 immunoreactivity wasalso extensive in M-1 and M-2 and scattered-in, M-4through M-6. In cultured cells, immunoreactivity wasagain confined to the cytoplasm, often perinuclear in dis-tribution.
TGF, Radioreceptor Assays
Neutral conditioned media from each meningioma con-tained no significant TGFO competing activity, suggest-ing that the meningioma cells do not synthesize TGFB1 -3in an active form. Acid activation of the media from eachmeningioma culture, however, produced significantdose-dependent competition with 125TGF12, indicatingthat meningioma cultures synthesize latent TGFps. Ap-proximately 9 ng/ml (M-1), 8 ng/ml (M-2), 2 ng/ml (M-3), 6ng/ml (M4), 1 ng/ml (M-5), and 1 ng/ml (M-6) were se-creted by late passage (P4-5) meningioma cell cultures(Figure 3).
TGF,3 Receptor Crosslinking
In the three meningioma cultures tested, affinity cross-linked bands were identified at the 45- to 60-kd, 70- to90-kd, and the 250-300-kd regions of the gels. Thesebands correspond to the type 1, type 2, and type 3 re-ceptors, respectively.3435 Competitive inhibition using200 ng/ml unlabeled TGFI31 significantly reduced the in-tensity of the 65-kd band and completely displaced 1251
TGF, from binding to the 85 kd (type 11) and 250 to 300kd (type 111) receptors, as demonstrated by the disap-pearance of these bands (Figure 4).
3H-Thymidine Incorporation
Data from the study evaluating TGFi1 effects on [3H]thymidine incorporation after stimulation with 10% FCSare graphed in Figure 5. Incubation with serum-freeMCDB 402 medium was associated with a slight in-crease in [3H] thymidine incorporation 36 and 48 hoursafter addition, indicating continued viability of cells in se-rum-free media. Treatment with 10% FCS significantly in-creased [3H] thymidine incorporation in each meningio-ma culture at each time point. Transforming growth factorbeta-1 (10 ng/ml) in serum-free media had no effect on
z
mm
L._
100 a
90
80 "
70'\
as
60 \ N\ N\ \
50 \.
40 \1
100
90
80
70
60 z
050 m
co4040
30
20
10
0 125 250 500 1000
Condi'lo-nd Modium Added -ul
Figure 3. TGF,3 binding competing activity in meningioma cellconditioned media. Increasing volumes of acid-treated condi-tioned mediafrom M-1 (--- -), M-2 (--), M-4 ( ), M-5 ( . *), andM-6 (-.-) as well as untreated conditioned mediafrom M-4 (--),M-5 (--), and M-6 [-, (M-6 over laps M-5)I were assayed intriplicate.
4 -

638 Johnson et alAJP September 1992, Vol. 141, No. 3
ittoUL
(ID (00F~
280K-
85K-
65K-
Figure 4. Affinity labeling of meningioma cells with "25 TGF;11.Confluent monolayers of meningiomas M-4, M-5, and M-6 wereincubated with 2 ng of1'5I-TGF31, M-6 was also co-incubated withexcess (200 ng) unlabeled TGF1.
M-2, M4, M-5, and M-6, but produced a statistically in-significant attenuation in [3H] thymidine incorporation incells from M-1 at 24, 36, and 48 hours and M-3 at 24hours relative to controls. In contrast, FCS-stimulatedcells treated with TGF,1 consistently exhibited a signifi-cant reduction in [3H] thymidine incorporation comparedwith FCS alone in all six meningiomas at each time point.Autoradiography to determine percent labeled nuclei on
M-5 and M-6 showed similar results (data not shown).Findings from the study evaluating TGFp1 effects on
[3H] thymidine incorporation after suboptimal stimulationwith EGF/lnsulin are illustrated in Figure 6. Incubation withserum-free media was again associated with a slight in-crease in [3H] thymidine incorporation. Administration ofEGF/insulin produced a significant increase in [3H] thy-midine incorporation in all six meningiomas at all timepoints. Transforming growth factor beta-1 alone resultedin an insignificant decrease in incorporation in cells fromM-1, M-3, and M-5 at 24, 36, and 48 hours. Treatment ofcells with EGF/insulin and TGF1 significantly reducedEGF/insulin's stimulatory effect in all six meningiomas ateach time point. Within any given culture, differences ob-served at the different treatment times were not statisti-cally significant.
Discussion
The presence of mRNA and protein of all three mamma-lian isoforms have been demonstrated in the mouse1921and in human leptomeninges (MD Johnson and HLMoses, unpublished observations). These observationsraised the possibility that neoplasms of the meningesmight synthesize, secrete, and respond to members ofthe TGF,B family in an autocrine fashion. Findings pre-sented here indicate that meningioma cells do, in fact,synthesize and secrete all three TGF, isoforms. Althoughthe extent of the TGFP1, TGF,B2, and TGF,3 expressionin various benign and malignant subtypes of meningio-mas is not known, recent detection of TGFp1 in tumortissue from several meningiomas,39 and the present datasuggest consistent expression in these mesenchymal tu-mors. Moreover a recent report that acid/ethanol extractsof one human meningioma exhibited TGFP-like activityon NRK49F fibroblasts also may present latent TGF, ac-tivated by the extraction procedure.40
The extent and pattern of expression of all threeTGFIs in meningiomas has not been reported previ-ously. Takahashi et a139 recently detected TGFo1 tran-scripts in five tumors of unspecified subtype. Analysis ofTGF,2 and TGF,B3 were not performed, however. Find-ings presented here suggest that at least meningiothelialand fibroblastic meningiomas produce all three isoformsof TGF,B. Moreover, TGFI2 or TGF,3 may represent themore abundantly synthesized isoforms. Although the bi-ologic effects of TGF,B2 and TGF,B3 on meningioma cellshave not been reported, all three TGFP isoforms bind thesame surface receptors. Moreover, all three isoforms ap-pear to produce similar biological effects in other celltypes in culture.41 Ongoing analysis comparing expres-sion of the three isoforms in meningioma subtypes mayclarify which species predominate. Nonetheless the cur-rent observations suggest that meningiomas, like theirarachnoid precursors, synthesize multiple members ofthe TGF3 family.
Comparison of TGF,B-competing activity in neutraland acid-activated conditioned media suggests that me-ningioma cells secrete the latent, TGFP precursor in con-centrations ranging from 1 to 10 ng/ml. Similar findingshave been reported in other epithelial cells and fibro-blasts that also synthesize and secrete TGF, in its latentform.4243 Because latent TGF3 does not bind to theTGF, receptors, it is unlikely that the secreted productexerts autocrine regulation of meningioma growth perse.This inability to activate TGFps may represent one facetof defective growth regulation permitting meningioma for-mation.
Transforming growth factor P-i's role, if any, in regu-lation of meningioma cell growth awaits elucidation. Au-tocrine TGF, stimulation of cell growth, however, might
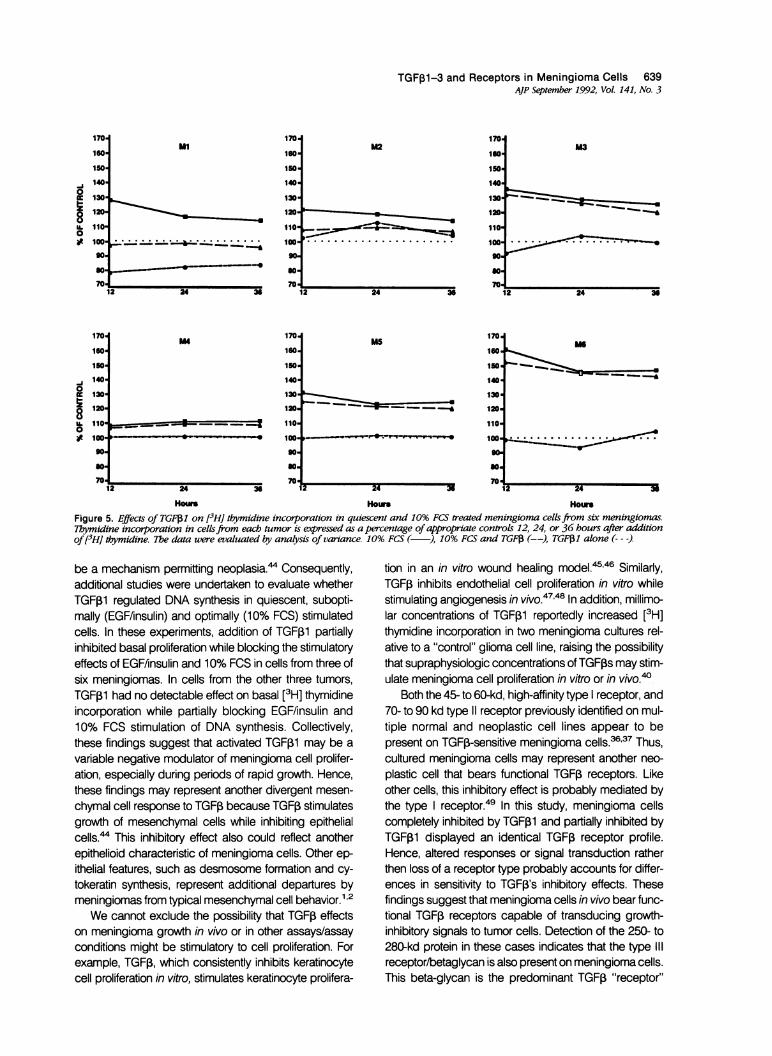
TGFp1-3 and Receptors in Meningioma Cells 639AJP September 1992, Vol. 141, No. 3
ml
- ... *******b**********"
2 24 36
170'
160'
150'
140'
130'
120'
110'
100'90'go.70'
M2
2 24 36
170'
160'
150'140'
130'
120-
110-
100-
90-go.*0'
70'
M3
C====~~~~,=_<--.-_ _A
2 24 36
0
8U.0
170'
160'
150'
140-
130-
120.
110a
M4170.
160.
150'
140'
130a
120.
110'
100'04 ~Ia-*
MS170.
160'
150'
140'
130'
120.
110'
100-
MG
9 90 90
70
12 24 3 12 24 36 2 24 3
Hours Hou HoursFigure 5. Effects of TGF1 on [3HJ thymidine incorporation in quiescent and 10% FCS treated meningioma cellsfrom six meningiomas.7hymidine incorporation in cellsfrom each tumor is expressed as a percentage of appropriate controls 12, 24, or 36 hours after additionof13H] dtymidine. 7Te data were evaluated by analysis of vanance. 10% FCS ( ), 10% FCS and TGFf (--), TGF1 alone (---)
be a mechanism permitting neoplasia.4 Consequently,additional studies were undertaken to evaluate whetherTGFfi1 regulated DNA synthesis in quiescent, subopti-mally (EGF/insulin) and optimally (10% FCS) stimulatedcells. In these experiments, addition of TGF,B1 partiallyinhibited basal proliferation while blocking the stimulatoryeffects of EGFfinsulin and 10% FCS in cells from three ofsix meningiomas. In cells from the other three tumors,TGF,1 had no detectable effect on basal [3H] thymidineincorporation while partially blocking EGF/insulin and10% FCS stimulation of DNA synthesis. Collectively,these findings suggest that activated TGFI1 may be a
variable negative modulator of meningioma cell prolifer-ation, especially during periods of rapid growth. Hence,these findings may represent another divergent mesen-
chymal cell response to TGF, because TGF3 stimulatesgrowth of mesenchymal cells while inhibiting epithelialcells.' This inhibitory effect also could reflect anotherepithelioid characteristic of meningioma cells. Other ep-ithelial features, such as desmosome formation and cy-
tokeratin synthesis, represent additional departures bymeningiomas from typical mesenchymal cell behavior.1' 2
We cannot exclude the possibility that TGFI effectson meningioma growth in vivo or in other assays/assayconditions might be stimulatory to cell proliferation. Forexample, TGFPi, which consistently inhibits keratinocytecell proliferation in vitro, stimulates keratinocyte prolifera-
tion in an in vitro wound healing modeli5'46 Similarly,TGF,B inhibits endothelial cell proliferation in vitro whilestimulating angiogenesis in vivo.47'48 In addition, millimo-lar concentrations of TGFB1 reportedly increased [3H]thymidine incorporation in two meningioma cultures rel-ative to a "control" glioma cell line, raising the possibilitythat supraphysiologic concentrations of TGF,Bs may stim-ulate meningioma cell proliferation in vitro or in vivo.40
Both the 45- to 60-kd, high-affinity type receptor, and70- to 90 kd type 11 receptor previously identified on mul-tiple normal and neoplastic cell lines appear to bepresent on TGF,B-sensitive meningioma cells.36'37 Thus,cultured meningioma cells may represent another neo-
plastic cell that bears functional TGF,B receptors. Likeother cells, this inhibitory effect is probably mediated bythe type receptor.49 In this study, meningioma cellscompletely inhibited by TGF31 and partially inhibited byTGF1 displayed an identical TGFP receptor profile.Hence, altered responses or signal transduction ratherthen loss of a receptor type probably accounts for differ-ences in sensitivity to TGF,B's inhibitory effects. Thesefindings suggest that meningioma cells in vivo bear func-tional TGF, receptors capable of transducing growth-inhibitory signals to tumor cells. Detection of the 250- to280-kd protein in these cases indicates that the type Ill
receptor/betaglycan is also present on meningioma cells.This beta-glycan is the predominant TGFPi "receptor"
170.
160'
150'
140'
130'
120'
110'
100'
90.go.*0
70'
IL0

640 Johnson et alAJP September 1992, Vol. 141, No. 3
140 ml 140 M2 140 M3
130 130 130
120 120 120
d - * -
101 10 110o ~~~~~~~~~~~~~go.so so so
70 70 70
12 24 36 12 24 36 12 24 36
140 M4 140 MS 140 M6
130 130 130
.j 120 >,, ___ _ _ 120 120
0~~~~~~~~~~~~~~~~~~~~~~~~~~~~1
8 100 . ..._ 0 .......... I0 ._ ..
a- - - - - - - -0-
o ~~~~~~~~~~o .4 g
so 90- - - -o - ---- so
70L 70 24_X 70 _
12 24 36 24 36 12 24 36
Hours Hours Hours
Figure 6. Effects of TGFf13 on [3H] thymidine incorporation in quiescent and EGF/Insulin treated meningioma cellsfrom six meningiomas.Thymidine incorporation in cellsfrom each tumor is expressed as a percentage of the appropriate control 12, 24 or 36 h after addition of[3-'H thymidine. The data were evaluated by analysis of variance. EGF/Insulin ( ), EGF/Insulin and TGB3 (--), TGF31 alone (---).
type on several mesenchymal cells (ie, fibroblasts, chon-droblasts, and osteoblasts), but is not present on epithe-lial cells.`051 Hence, its limited expression (relative tofunctional type and 11 receptors) on meningioma cellsmay be another manifestation of the meningiomas mixedmesenchymal/epithelial phenotype. Accumulating evi-dence suggests that the type III receptor/beta-glycan is acell surface protein without signal transducing proper-ties.51 Its role, if any, in meningioma cell physiologyawaits elucidation.
Transforming growth factor beta isoforms could influ-ence a number of other cellular processes in meningio-mas. Transforming growth factor beta-1 stimulates syn-thesis of fibronectin and collagen while inhibiting synthe-sis of collagenase.5254 Thus TGF,s may be animportant modulator of fibronectin and collagen synthe-sis by meningioma cells.1' 22 Angiogenic properties ofTGF,1, including stimulation of endothelial cell prolifera-tion in vivo and induction of capillary formation might con-tribute to the formation and infiltration of leptomeningealblood vessel into these highly vascularized neo-plasms.47'48 In addition, the ability of TGF,B1 to stimulateosteoblasts and influence bone remodeling raises thepossibility that TGFI family members modulate the hy-perostosis that frequently occurs in the skull overlyingmeningiomas.1,1844 Future studies may clarify the role ofTGF,B isoforms, if any, in mediating these changes.
Collectively, these data suggest that meningiomassynthesize one or more isoforms of TGF, that, when ac-tivated, inhibit meningioma cell proliferation. These find-ings raise the possibility that local infusion of degrada-tion-resistant TGF,B agonists, endogenous activators ofTGF,B such as plasmin,43 or other agents that remove thelatency-associated peptide might slow tumor growth orreduce the incidence of recurrence.
References1. Russell DS, Rubinstein U: Pathology of Tumors of the Ner-
vous System. Fifth Edition. Baltimore, Williams & Wilkins,1989, pp 449-532
2. Burger PC, Scheithauer BW, Vogel FS: Surgical Pathologyof the Nervous System and its Coverings. 3rd ed. New York:Wiley, 1991, pp 67-93
3. Bickerstaff ER, Small JM, Guest IA: The relapsing course ofcertain meningiomas in relation to pregnancy and menstru-ation. J Neurol Neurosurg Psychiatry 1958, 21:89-91
4. Chaudhuri PK, Wallenburg HCS: Brain tumours and preg-nancy. Eur J Obstet Gynecol Reprod Biol 1980,11:109-114
5. Jay JR, MacLaughlin DT, Riley KR, Martuza RL: Modulationof meningioma cell growth by sex steroid hormones in vitro.J Neurosurg 1975, 62:757-762
6. Weisman AS, Villemure JG, Kelly PA: Regulation of DNAsynthesis and growth of cells derived from primary humanmeningiomas. Cancer Res 1986, 46:2545-2550

TGF01-3 and Receptors in Meningioma Cells 641AJP September 1992, Vol. 141, No. 3
7. Koper JW, Foekens JA, Braakman R, Lamberts SWJ: Ef-fects of progesterone on the response to epidermal growthfactor and other growth factors in cultured human meningi-oma cells. Cancer 1990, 46:2545-2550
8. Hirata Y, Uchihashi M, Nakajma H, Fujita T, Matsukura S:Presence of human epidermal growth factor in human ce-rebrospinal fluid. J Clin Endocrinol Metab 1982, 55:1174-1177
9. Libermann TA, Nusbaum HR, Razon N, Kris R, Lax I, SoreqH, Whittle N, Waterfield MD, Ullrich A, Schlessinger J: Am-plification enhanced expression and possible rearrange-ment of EGF receptor gene in primary human brain tumorsof glial origin. Nature 1985, 313:144-147
10. Weisman AS, Raguet SS, Kelly PA. Characterization of theepidermal growth factor receptor in human meningioma.Cancer Res 1987, 47:2172-2176
11. Reubi JC, Horisberger U, Lang W, Koper VW, Braakman R,Lamberts SWJ: Coincidence of EGF receptors and somato-statin receptors in meningiomas but inverse differentiation-dependent relationship in glial tumors. Am J Pathol 1989,134:337-344
12. Maxwell M, Galanopoulos T, Hedley-Whyte ET, Black PMcL, Antoniades HN: Human meningiomas co-expressplatelet-derived growth factor (PDGF) and PDGF-receptorgenes and their protein products. Int J Cancer 1990, 46:16-21
13. Kazumoto K, Tamura M, Hoshino H, Yuasa Y: Enhancedexpression of the sis and c-myc oncogenes in human men-ingiomas. J Neurosurg 1990, 72:786-791
14. Moses HL, Branum EB, Proper JA, Robinson RA: Trans-forming growth factor production by chemically transformedcells. Cancer Res 1981, 41:2842-2848
15. Roberts AB, Anzano MA, Lamb LC, Smith JM, Sporn MB:New class of transforming growth factors potentiated by epi-dermal growth factor: Isolation from nonneoplastic tissues.Proc Natl Acad Sci USA 1981, 78:5339-5343
16. Shipley, GD, Tucker RF, Moses HL: Type B-transforminggrowth factor/growth inhibitor stimulates entry into S phasein monolayer cultures of AKR-2B cells after a prolonged pre-replicative interval. Proc Natl Acad Sci USA 1985, 82:4147-4151
17. Battegay EJ, Raines EW, Seifert RA, Bowen-Pope DF, Ross:TGF-Beta induces bimodal proliferation of connective tissuecells via complex control of an autocrine PDGF loop. Cell1990, 63:515-524
18. Moses HL, Yang EY, Pietenpol JA: TGF-4 stimulation andinhibition of cell proliferation: New mechanistic insights. Cell1990, 63:245-247
19. Pelton RW, Nomura S, Moses HL, Hogan BLM: Expressionof transforming growth factor beta-2 RNA during murine em-bryogenesis. Development 1989, 106:759-767
20. Pelton RW, Dickinson ME, Moses HL, Hogan BLM: In situhybridization analysis of TGFp1 and P2. Development 1990,110:609-620
21. Heine VI, Munoy EF, Flanders KC, Ellingsworth LR, LamPHY, Thompson NL, Roberts AB, Spom MB: Role of trans-forming growth factor-p in the development of the mouseembryo. J Cell Biol 1987,105:2861-2876
22. Rutka JT, Giblin J, Dougherty DV, McCulloch JR, DeArmondSV, Rosenblum ML: An ultrastructural and immunocyto-chemical analysis of leptomeningeal and meningioma, cul-tures. J Neuropathol Exp Neurol 1986, 45:285-303
23. Warnke R, Levy R: Detection of T and B cell antigens withhybridoma monoclonal antibodies: A biotin-avidin-horseradish peroxidase method. J Histochem Cytochem1980, 28:771-776
24. Meis JM, Ordoney NG, Bruner JM: Meningioma: An immu-nohistochemical study of 50 cases. Arch Pathol Lab Med1986,10:934-940
25. Aviv H, Leder P: Purification of biologically active globinmessenger RNA by chromatography on oligothymidylicacid-cellulose. Proc Natl Acad Sci USA 1972, 69:1408-1412
26. Thomas PS: Hybridization of denatured RNA transferred ordotted to nitrocellulose paper, Methods in Enzymology. Ed-ited by R Wu, L Grossman, K Moldave. Vol 100. New York,Academic Press, 1987, pp 255-266
27. Derynck R, Jarrett JA, Chen EY, Goeddel DV: The murinetransforming growth factor-beta precursor. J Biol Chem1986, 261:4377-4379
28. Danielson PE, Forss-Petter S, Brow MA, Calavetta L, Doug-lass J, Milner RJ, Sutcliffe JG: p1 B15: A cDNA clone of therat mRNA encoding cyclophilin. DNA 1988, 7:261-267
29. Feinberg AP, Vogelstein B: A technique for radiolabellingDNA restriction endonuclease fragments to high specificactivity. Anal Biochem 1983, 132:6-13
30. Miller DA, Lee A, Pelton RW, Chen EY, Moses HL, DerynckR: Murine transforming growth factor-beta 2 cDNA se-quence and expression in adult tissue and embryos. MolEndocrinol 1989, 3:1108-1114
31. Cox KH, DeLeon DV, Angerer LM, Angerer RC: Detection ofmRNAs in sea urchin embryos by in situ hybridization usingasymmetric RNA probes. Dev Biol 1984,101:485-502
32. Pelton RW, Saxena B, Jones M, Moses HL, Gold LI: Immu-nohistochemical localization of TGF,B1, TGFI2 and TGF,3proteins in the mouse embryo. J Cell Biol 1991, 115:1091-1105
33. Hsu S-M, Raine L: The use of avidin-biotin-peroxidase com-plex (ABC) in diagnostic and research pathology, Ad-vances in Immunochemistry. Edited by RA DeLellis. NewYork, Mason, 1984, pp 31-42
34. Pelton RW, Johnson MD, Saxena B, Perkett BA, Moses HL,Gold LI: Expression of TGF31, 132, and 133 mRNA and pro-tein in the murine lung. Am J Respir Dis Cell Mol Biol 1991,5:522-530
35. Lyons RM, Miller DA, Graycar JL, Moses HL, Derynck R:Differential binding of transforming growth factor -,11, -32and -133 by fibroblasts and epithelial cells measured by af-finity cross-linking of cell surface receptors. Mol Endocrinol1991, 5:1887-1896
36. Tucker RF, Branum EL, Shipley GD, Ryan RJ, Moses HL:Specific binding to cultured cells of 1251-labeled transform-ing growth factor-type 13 from human platelets. Proc NatlAcad Sci USA 1989, 81:6757-6761
37. Massaque J, Like B: Cellular receptors for type B transform-ing growth factor, ligand binding and affinity labeling in hu-

642 Johnson et alAJP September 1992, Vol. 141, No. 3
man and rodent cell lines. J Biol Chem 1985, 260:2636-2645
38. Statistical Users Guide: Statistics. Version 6. Cary, NC: SASInstitute, 1985
39. Takahashi JA, Mori H, Fukumoto M, Igarashi K, Jaye M, OdaY, Kikuchi H, Hatanaka M: Gene expression of fibroblastgrowth factors in human gliomas and meningiomas: Dem-onstration of cellular source of basic fibroblast growth factormRNA and peptide in tumor tissues. Proc NatI Acad SciUSA 1990, 87:5710-5714
40. Nitta T, Sato K, Okumura K: Transforming growth factor(TGF),B-like activity of intracranial meningioma and its effecton cell growth. J Neurol Sci 1991, 101:19-23
41. Graycar JL, Miller DA, Arrick BA, Lyons RM, Moses HL,Derynck R: Human transforming growth factor 133: Recom-binant expression, purification and biological activities incomparison with transforming growth factors-beta 1 andbeta 2. Mol Endocrinol 1989, 3:1977-1986
42. Lawrence DA, Pircher R, Krycena-Martinerie C, Jullien P:Normal embryo fibroblasts release transforming growth fac-tors in a latent form. J Cell Physiol 1984, 121:184-188
43. Lyons RM, Gentry LE, Purchio AF, Moses HL: Mechanism ofactivation of latent recombinant transforming growth factor11 by plasmin, J Cell Biol 1990,110:1361-1367
44. Bamard JA, Lyons RM, Moses HL: The cell biology of trans-forming growth factor 13. Biochim Biophys Acta 1990,1032:79-87
45. Shipley GD, Pittelkow MR, Wille JJ, Scott RE, Moses HL:Reversible inhibition of normal human prokeratinocyte pro-liferation by type 13 transforming growth factor-growth inhib-itor in serum-free medium. Cancer Res 1986, 46:2068-2071
46. Beck SL, Chin TL, Hiraboyashi SE, Deguzman L, Lee WP,McFatridge LL, Xu Y, Bates RL, Ammann AJ: Acceleratedhealing of ulcer wounds in the rabbit ear by recombinant
human transforming growth factor-31. Growth Factors 1990,2:273-282
47. Baird A, Durkin T: Inhibition of endothelial cell proliferation bytype-beta transforming growth factor: Interactions withacidic and basic fibroblast growth factors. Biochem BiophysRes Commun 1986,138:476-482
48. Roberts AB, Spom MB, Assoian RK, Smith JM, Roche NS,Wakefield LM, Heine VI, Liotta LA, Falanga V, Kehil, JH,Fauci AS: Transforming growth factor-beta: Rapid inductionof fibrosis and angiogenesis in vivo and stimulation of col-lagen formation in vitro. Proc Natl Acad Sci 1986, 83:4167-4171
49. Boyd FT, Massague J: Growth inhibitory response to TGF-13linked to expression of a 53 kDa cell surface TGF-,B recep-tor. J Biol Chem 1989, 264:2272-2278
50. Andres JL, Stanley K, Cheifety S, Massaque J. Membrane-anchored and soluble forms of betaglycan, a polymorphicproteoglycan that binds transforming growth-P. J Cell Biol1989,109:3137-3145
51. Segarini PR, Rosen DM, Seyedin SM: Binding of transform-ing growth factor ,B to cell surface proteins varies with celltype. Mol Endocrinol 1989, 3:261-272
52. Ignotz RA, Massaque J: Transforming growth factor 13 stim-ulates the expression of fibronectin and collagen and theirincorporation into the extracellular matrix. J Biol Chem 1986,261:4337-4345
53. Ignotz RA, Takeshi E, Massaque J: Regulation of fibronectinand type collagen nRNA levels by transforming growthfactor-P. J Biol Chem 1987, 262:6443-6446
54. Edwards DR, Murphy G, Reynolds JJ, Whitham SE, Do-cherty J, Angel P, Heath JK: Transforming growth factorbeta modulates the expression of collagenase and metallo-proteinase inhibitor. EMBO J 1987, 6:1889-1904




















