
Human saliva, plasma and breast milk exosomes contain RNA: uptake by macrophages

1999, 73(7):5865. J. Virol.
CrystalFenamore, Kristine Sheridan, Ravi Singh and Ronald G. Stefan Worgall, Ruth Connor, Robert J. Kaner, Elizabeth MacrophagesCoreceptors by Human AlveolarImmunodeficiency Virus Type 1 Expression and Use of Human
http://jvi.asm.org/content/73/7/5865Updated information and services can be found at:
These include:
REFERENCEShttp://jvi.asm.org/content/73/7/5865#ref-list-1at:
This article cites 58 articles, 32 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from
on D
ecember 3, 2013 by guest
http://jvi.asm.org/
Dow
nloaded from

JOURNAL OF VIROLOGY,0022-538X/99/$04.0010
July 1999, p. 5865–5874 Vol. 73, No. 7
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Expression and Use of Human Immunodeficiency Virus Type 1Coreceptors by Human Alveolar Macrophages
STEFAN WORGALL,1 RUTH CONNOR,2 ROBERT J. KANER,1 ELIZABETH FENAMORE,2
KRISTINE SHERIDAN,2 RAVI SINGH,1 AND RONALD G. CRYSTAL1*
Division of Pulmonary and Critical Care Medicine, The New York Hospital–Cornell Medical Center,1 andAaron Diamond AIDS Research Center, The Rockefeller University,2 New York, New York
Received 27 July 1998/Accepted 26 March 1999
Human immunodeficiency virus type 1 (HIV-1) requires, in addition to CD4, coreceptors of the CC or CXCchemokine families for productive infection of T cells and cells of the monocyte-macrophage lineage. Based onthe hypothesis that coreceptor expression on alveolar macrophages (AM) may influence HIV-1 infection of AMin the lung, this study analyzes the expression and utilization of HIV-1 coreceptors on AM of healthyindividuals. AM were productively infected with five different primary isolates of HIV-1. Levels of surfaceexpression of CCR5, CXCR4, and CD4 were low compared to those of blood monocytes, but CCR3 was notdetectable. mRNA for CCR5, CXCR4, CCR2, and CCR3 were all detectable, but to varying degrees and withvariability among donors. Expression of CCR5, CXCR4, and CCR2 mRNA was downregulated followingstimulation with lipopolysaccharide (LPS). In contrast, secretion of the chemokines RANTES, MIP-1a, andMIP-1b was upregulated with LPS stimulation. Interestingly, HIV-1 replication was diminished following LPSstimulation. Infection of AM with HIV-1 in the presence of the CC chemokines demonstrated blocking ofinfection. Together, these studies demonstrate that AM can be infected by a variety of primary HIV-1 isolates,AM express a variety of chemokine receptors, the dominant coreceptor used for HIV entry into AM is CCR5,the expression of these receptors is dependent on the state of activation of AM, and the ability of HIV-1 to infectAM may be modulated by expression of the chemokine receptors and by chemokines per se.
The human immunodeficiency virus type 1 (HIV-1) requiresinteraction of the viral envelope glycoprotein gp120 with CD4and a second coreceptor for productive infection of its targetcell (4, 5, 9, 19, 34). These recently identified coreceptorsinclude the b-chemokine receptors (CCR5, CCR3, andCCR2b) and the a-chemokine receptor CXCR4 (2, 3, 11, 16,20, 21, 23, 24, 46–48). HIV-1 tropism and entry cofactor utili-zation are important determinants of pathogenesis (4, 5, 9, 19,34). During primary HIV-1 infection and throughout theasymptomatic phase of infection, isolates from blood are pre-dominantly macrophagetropic and CCR5 dependent (7, 15, 52,53). In contrast, strains that emerge later in many infectedindividuals can use CXCR4, the main coreceptor for HIV-1infection of T cells (7, 15, 52, 53, 58).
The focus of the present study is to characterize the patternand usage of the HIV-1 coreceptors on healthy human alveolarmacrophages (AM), the pulmonary representative of the mono-nuclear phagocyte system. Other than evidence of productiveinfection of AM in HIV-1-positive individuals (1, 12, 30, 35,38–40, 45, 50), little is known about the interactions of HIV-1with this cell type. Pulmonary infections are a major cause ofthe morbidity and mortality associated with infection withHIV-1, and a majority of individuals with AIDS develop one ormore episodes of pulmonary infection during the course oftheir disease (29, 36). AM represent the major cellular hostdefense against microorganisms on the respiratory epithelialsurface (6, 43). In this context, understanding the mechanismsof HIV-1 infection of AM may be central to understanding theloss of respiratory epithelial surface host defense associatedwith HIV-1 infection.
Based on the knowledge that AM are differentiated fromblood monocytes and that HIV-1 mainly uses CCR5 as a co-receptor on blood monocytes and in vitro monocyte-derivedmacrophages (6, 9, 34, 42, 61, 64) but that the type and level ofcoreceptor expression on monocytes can be influenced by dif-ferentiation and activation (8, 18, 37, 44, 45), it is reasonable toassume that the coreceptors are expressed on AM. Interest-ingly, the data demonstrate that the coreceptor expression onhealthy human AM generally parallels that of autologous bloodmonocytes. However, most coreceptor expression on AM ismarkedly lower and is only mildly influenced by activation.Concomitant production of chemokines such as RANTES,MIP-1a, and MIP-1b may also markedly influence the abilityof HIV-1 to infect AM.
MATERIALS AND METHODS
Cells. Human AM were obtained by bronchoalveolar lavage from healthyvolunteers as previously described (49). The lavage fluid was filtered throughgauze to remove debris and cells were pelleted, washed with phosphate-bufferedsaline (PBS) (pH 7.4) and resuspended in RPMI 1640 medium containing 10%fetal bovine serum, 2 mM glutamine, 100 U of penicillin/ml, and 10 mg ofstreptomycin (GIBCO BRL, Gaithersburg, Md.)/ml. For most experiments, AMwere purified by adherence to plastic (2 h, 37°C). For flow cytometry studies, thecells were cultured in Teflon-coated vials (Savillex Corp., Minnetonka, Minn.)until evaluation. Peripheral blood monocytes (PBM) and peripheral blood lym-phocytes (PBL) were obtained from the blood of the AM donors and purified byFicoll gradients. The monocytes were separated from the lymphocytes by adher-ence and maintained in RPMI 1640 media containing 10% human serum, 100 Uof penicillin/ml, and 10 mg of streptomycin/ml for 18 h. For RNA analysis, PBMand PBL were isolated by using immunomagnetic beads (Dynal, Lake Success,N.Y.) coated with anti-CD14 for the isolation of monocytes and with anti-CD3for the isolation of lymphocytes.
Infection with HIV-1 primary isolates. AM were cultured in 48-well plates andinfected with five different primary HIV-1 isolates with known coreceptor usage(AD2-3 [CCR5], AD2-6 [CCR5 and CXCR4], AD3-3 [CCR5], AD3-7 [CCR5,CCR2b, CCR3, and CXCR4] [15], and JRFL [CCR5]). Twenty-four hours fol-lowing infection, the cells were washed, and fresh medium was added. Productiveinfection was determined by measuring HIV-1 p24 antigen in the supernatant by
* Corresponding author. Mailing address: Weill Medical College ofCornell University-New York Presbyterian Hospital, Starr 505, NewYork, NY 10021. Phone: (212) 746-2258. Fax: (212) 746-8383. E-mail:[email protected].
5865
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

enzyme-linked immunosorbent assay (ELISA) (Abbott Laboratories, North Chi-cago, Ill.) 5, 8, and 14 days after infection.
Flow cytometry. To analyze surface HIV-1 coreceptor expression on AM,PBM, and PBL, the cells were incubated with PBS containing 2% bovine serumalbumin and 10% human serum (4°C for 15 min), followed by incubation withprimary antibodies against CCR5 (2D7), CCR3 (7B11), CXCR4 (12G5) (allantibodies were obtained from the AIDS Research and Reference ReagentProgram, National Institutes of Health, Bethesda, Md.). After washing with PBS,the cells were incubated with fluorescein isothiocyanate (FITC)-conjugated goatanti-mouse immunoglobulin G (IgG) [F(ab9)2] fragments (Boehringer Mann-heim, Indianapolis, Ind.) or FITC-conjugated anti-CD4 (Pharmingen, San Di-ego, Calif.) for 30 min. The cells were then washed and incubated with 10%mouse serum for 15 min, followed by incubation with phycoerythricin (PE)-labeled antibodies against HLA-DR (AM), CD14 (PBM), or CD3 (PBL)(Pharmingen), washed, and then analyzed by flow cytometry. Isotype-matchedunlabeled and PE-labeled antibodies served as negative controls. To analyzeCCR5 surface expression using antibodies other than clone 2D7, the FITC-conjugated primary antibodies recognizing CCR5 (FAB180F, FAB181F,FAB182F, and FAB183F [all from R&D Systems, Minneapolis, Minn.]) wereused to stain AM, PBM, and PBL as described above. FITC-labeled isotype-matched antibody was used as a control.
mRNA analysis. Two strategies were used to evaluate coreceptor mRNA inthe AM in comparison to PBM and PBL, reverse transcription (RT)-PCR andNorthern analysis. For RT-PCR, total RNA was extracted from AM, PBM(CD14 purified), or PBL (CD3 purified) by using Trizol reagent (GIBCO BRL)and reverse transcribed (45 min, 48°C; 2 min, 94°C), and the resulting DNA wasamplified by PCR (9600 Gene Amp; Perkin-Elmer) by 40 cycles of 94°C for 30s, 56°C for 1 min, and 68°C for 2 min by using synthetic oligonucleotide primersspecific for CCR3 (sense primer, TCCACACTCGAGAATGACCATCT; anti-sense primer, ACTGGAAGTTTGAAGGACTGTTTT; product size, 578 bp),CCR5 (sense primer, CAGGGCTGTGAGGCTTATCTT; antisense primer,CCCAGGCTGTGTATGAAAACT; product size, 437 bp), CXCR4 (senseprimer, TTGTCTGAACCCCATCCTCTAT; antisense primer, ACTCCTGAAAACTGAAAAACCA; product size, 626 bp), CCR2B (sense primer, CCAACGAGAGCGGTGAAGAAGT; antisense primer, GGGAGTCCAGAAGAGAAAGTAAACA; product size, 737 bp), and CD4 (sense primer, AGTTGCATCAGGAAGTGAACCT; antisense primer, CTGAGACATCCGCTCTGCTTGG;
product size, 383 bp). Primers for glyceraldehyde 3-phosphate dehydrogenase(GAPDH) (sense primer, CCTTCATTGACCTCAACTACA; antisense primer,GGCAGTGATGGCATGGACTGT; product size, 443 bp) served as an internalcontrol. The PCR products were analyzed on a 1.5% agarose gel. DNA contam-ination was ruled out by pretreatment of the samples with DNase (GIBCO BRL)for 15 min at 37°C and by omitting the reverse transcriptase from the PCR as acontrol.
To analyze coreceptor expression by Northern analysis, total cellular RNA (10mg) was transferred to Duralon membranes (Stratagene, La Jolla, Calif.) afterelectrophoretic separation through a 1% agarose gel under denaturing condi-tions. Probes for CCR5, CCR3, CCR2B, and CXCR4 (kindly provided by NedLandau, Aaron Diamond AIDS Research Center, New York, N.Y.) were gelpurified and labeled with [32P]dCTP by random priming (Stratagene). Hybrid-izations were performed in hybridization solution (Quickhyb; Stratagene) for 2 hat 65°C, followed by sequential washes in 13 SSC (0.15 M NaCl plus 0.015 Msodium citrate)–0.1% sodium dodecyl sulfate (SDS) for 30 min and 0.1% SSC–0.1% 23 SDS for 30 min. Following hybridization, the membranes were analyzedby autoradiography or phosphorimaging.
Influence of AM stimulation on HIV-1 coreceptor expression. To analyze ifstimulation of AM influenced the expression of the HIV coreceptors, AM weretreated with 100 ng of lipopolysaccharide (LPS)/ml for either 4 or 48 h. TotalRNA was extracted and analyzed for coreceptor expression by RT-PCR andNorthern analysis as described above. To determine if LPS stimulation resultedin increased secretion of chemokines, the levels of MIP-1a, MIP-1b, RANTES,and eotaxin in the supernatant were quantified by ELISA (R&D Systems).
To evaluate if stimulation with LPS influences HIV-1 infection and replica-tion, AM were inoculated with 200 50% tissue culture infective doses (TCID50)of five different primary isolates as described above after overnight stimulationwith 100 ng of LPS/ml. HIV-1 p24 levels were measured in the culture superna-tant by ELISA at days 5, 8, and 14 postinfection. To ensure that cell viability wasnot affected by stimulation with LPS, AM were plated in 96-well plates, andviability was assessed in the presence or absence of 100 ng of LPS/ml after 7 and14 days by using an MTT-based cytotoxicity assay (Sigma, St. Louis, Mo.).
Chemokine blocking of HIV-1 infection in AM. To assess the ability of che-mokines to block HIV-1 infection of AM, cells were infected with 100 TCID50 ofHIV-1 AD2-3, AD2-6, AD3-3, AD3-6, or JRFL in the presence of 250 ng ofRANTES, MIP-1a, MIP-1b, or SDF-1/ml, either alone or in combination. After
FIG. 1. Replication of primary HIV-1 isolates in AM from different donors. AM were obtained by bronchoalveolar lavage of healthy individuals and infected in vitrowith five different primary HIV-1 isolates. HIV-1 p24 antigen was measured by ELISA in the supernatant on days 4, 8, and 14. Values are peak p24 levels measuredon day 14. The known coreceptor usage of each primary isolate is listed (15), as is the clinical stage of the individual at the time the virus was isolated. Note that AD2-3and AD2-6 are from the same individual at different stages, as are AD3-3 and AD3-7. Asx, asymptomatic. Each symbol represents data for one individual.
5866 WORGALL ET AL. J. VIROL.
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

48 h, the cells were washed, and the appropriate chemokines were added back tothe wells. p24 levels in the supernatant were measured by ELISA on day 7 afterinfection and were compared to those of control cultures infected in the absenceof added chemokines. Percent inhibition was calculated as (1 2 the mean p24concentration of duplicate wells with chemokines/mean of control wells) 3 100.
RESULTS
Infection of AM with primary HIV-1 isolates. Inoculation ofAM from healthy individuals with primary isolates of HIV-1demonstrated virus replication in AM from all donors for alltested isolates (Fig. 1). Peak p24 levels among the differentdonors ranged from 313 to 778 pg/ml for AD2-3, 25 to 951pg/ml for AD2-6, 341 to 1,516 pg/ml for AD3-3, 86 to 4,408pg/ml for AD3-7, and 604 to 20,000 pg/ml for JRFL.
HIV-coreceptor expression on AM. Flow cytometry analysisof the HIV-1 coreceptors CCR5, CCR3, and CXCR4, as wellas CD4, on the surface of AM demonstrated very low levels ofthese receptors (Fig. 2 and 3). In contrast, surface expressionof HLA-DR, a marker for AM, was detectable on 93 to 98% ofthe cells. Higher surface expression of CCR5, CXCR4, andCD4 was detectable on PBM and PBL from the same individ-uals stained in parallel, while the levels of CCR3 were very low
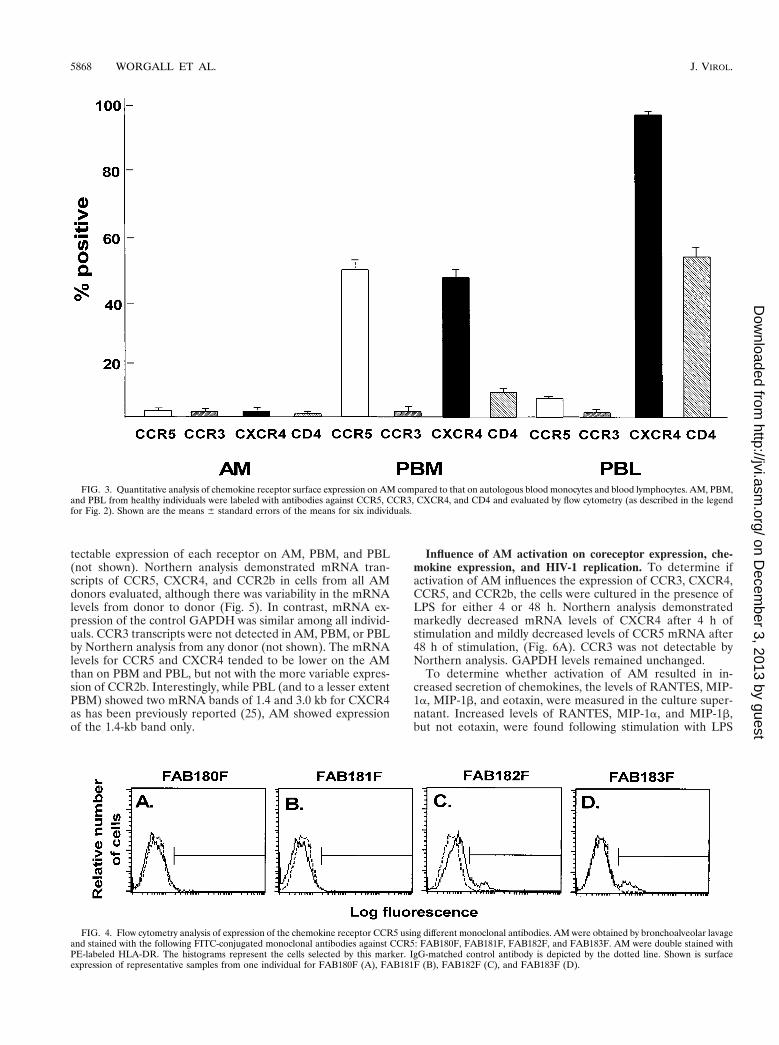
on PBM and PBL. On the average, CCR5 and CXCR4 levelson AM were significantly lower than on autologous PBM andPBL (P , 0.01, all comparisons), while CCR3 levels weresimilar (P . 0.1, all comparisons). Using antibody clonesagainst CCR5 other than clone 2D7 (Fig. 4), AM demon-strated low-to-undetectable levels of cells positive forFAB180F (1.2% 6 0.9%) and FAB181F (0.2% 6 0.2%),whereas some cells stained positive for FAB182F (8.1% 61.0%) and FAB183F (6.2% 6 0.8%). Further testing of theseantibodies on PBM and PBL showed staining similar to theCCR5 antibody 2D7 for clone FAB182F (PBM, 38.5% 66.1%; PBL, 5.9% 6 0.4%), whereas using the other clones,positive cells were less frequently observed or not detectable(clone FAB180F: PBM, 2.3% 6 0.9%, PBL, 2.0% 6 0.7%;clone FAB181F: PBM 6.5% 6 2.1%, PBL, 1.8% 6 0.5%; cloneFAB183F: PBM, 5.3% 6 1.5%, PBL, 2.2% 6 0.3%). Theseresults suggest that CCR5 surface expression levels on AM arelow, although a small subpopulation stained positive with theCCR5 clones FAB182F and FAB183F, suggesting that certainepitopes may be masked by using different antibody clones.
Analysis of CCR5, CCR3, CXCR4, CCR2b, and CD4 ex-pression at the mRNA level using RT-PCR demonstrated de-
FIG. 2. Flow cytometry evaluation of expression of chemokine receptors on AM. AM were obtained by bronchoalveolar lavage and stained with anti-CCR3, CCR5,or CXCR4 monoclonal antibodies (followed by FITC-labeled anti-IgG) or FITC-conjugated anti-CD4. Blood monocytes and lymphocytes obtained from the samedonors were evaluated in parallel. Shown are representative samples from one individual of AM (panels A to D), PBM (panels E to H), and PBL (panels I to M). Inaddition to coreceptor staining, AM were double stained with PE-labeled HLA-DR, blood monocytes were double stained with PE-labeled CD14, and lymphocyteswere double stained with PE-labeled CD3. The histograms shown represent the cells selected by these markers. IgG-irrelevant controls for each antibody are depictedby the dotted lines. Surface expression of CCR3 (panels A, E, and I); CCR5 (panels B, F, and J); CXCR4 (panels C, G, and K); and CD4 (panels D, H, and L) is shown.The solid horizontal line represents the region selected for quantification.
VOL. 73, 1999 ALVEOLAR MACROPHAGES AND HIV-1 CORECEPTORS 5867
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from
tectable expression of each receptor on AM, PBM, and PBL(not shown). Northern analysis demonstrated mRNA tran-scripts of CCR5, CXCR4, and CCR2b in cells from all AMdonors evaluated, although there was variability in the mRNAlevels from donor to donor (Fig. 5). In contrast, mRNA ex-pression of the control GAPDH was similar among all individ-uals. CCR3 transcripts were not detected in AM, PBM, or PBLby Northern analysis from any donor (not shown). The mRNAlevels for CCR5 and CXCR4 tended to be lower on the AMthan on PBM and PBL, but not with the more variable expres-sion of CCR2b. Interestingly, while PBL (and to a lesser extentPBM) showed two mRNA bands of 1.4 and 3.0 kb for CXCR4as has been previously reported (25), AM showed expressionof the 1.4-kb band only.
Influence of AM activation on coreceptor expression, che-mokine expression, and HIV-1 replication. To determine ifactivation of AM influences the expression of CCR3, CXCR4,CCR5, and CCR2b, the cells were cultured in the presence ofLPS for either 4 or 48 h. Northern analysis demonstratedmarkedly decreased mRNA levels of CXCR4 after 4 h ofstimulation and mildly decreased levels of CCR5 mRNA after48 h of stimulation, (Fig. 6A). CCR3 was not detectable byNorthern analysis. GAPDH levels remained unchanged.
To determine whether activation of AM resulted in in-creased secretion of chemokines, the levels of RANTES, MIP-1a, MIP-1b, and eotaxin, were measured in the culture super-natant. Increased levels of RANTES, MIP-1a, and MIP-1b,but not eotaxin, were found following stimulation with LPS
FIG. 3. Quantitative analysis of chemokine receptor surface expression on AM compared to that on autologous blood monocytes and blood lymphocytes. AM, PBM,and PBL from healthy individuals were labeled with antibodies against CCR5, CCR3, CXCR4, and CD4 and evaluated by flow cytometry (as described in the legendfor Fig. 2). Shown are the means 6 standard errors of the means for six individuals.
FIG. 4. Flow cytometry analysis of expression of the chemokine receptor CCR5 using different monoclonal antibodies. AM were obtained by bronchoalveolar lavageand stained with the following FITC-conjugated monoclonal antibodies against CCR5: FAB180F, FAB181F, FAB182F, and FAB183F. AM were double stained withPE-labeled HLA-DR. The histograms represent the cells selected by this marker. IgG-matched control antibody is depicted by the dotted line. Shown is surfaceexpression of representative samples from one individual for FAB180F (A), FAB181F (B), FAB182F (C), and FAB183F (D).
5868 WORGALL ET AL. J. VIROL.
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

(Fig. 6B). Although there was some donor-to-donor variabilityin the response to LPS, there was a marked increase for MIP-1a, MIP-1b, and RANTES following LPS stimulation in allsamples (Table 1).
To analyze if LPS simulation of AM influences infection ofAM with HIV-1, LPS-stimulated cells were inoculated with fivedifferent primary isolates of HIV-1 in the presence of LPS, andthe levels of HIV-1 replication were determined. Strikingly,HIV-1 replication of LPS-stimulated AM was diminished forall of the isolates tested compared to infection of unstimulatedcells (Fig. 6C; P , 0.05, all comparisons). The viability of thecells was not affected by LPS stimulation.
Chemokine blocking of HIV-1 replication in AM. To deter-mine if the ligands for the coreceptors could block HIV-1replication in AM, cells were infected with the primary isolatesAD2-3, AD2-6, AD3-3, AD3-7, or JRFL in the presence of
either RANTES, MIP-1a, MIP-1b, SDF-1, or all four com-bined. HIV-1 replication was inhibited in the presence ofRANTES (44 to 84%), MIP-1a (20 to 62%), and MIP-1b (55to 85%) for all the HIV-1 isolates (Fig. 7). All chemokinescombined had a .80% inhibitory effect on HIV-1 replication.Interestingly, for one HIV-1 isolate which, like CCR5, can useCXCR4 (AD2-6), SDF-1 blocked HIV-1 infection to 67%,whereas for all the other isolates SDF-1 did not block HIV-1infection. However, for AD2-6, the blocking effect of SDF-1did not exceed the effect seen by MIP-1a (92%), MIP-1b(55%), RANTES (44%), or all chemokines combined (86%).
DISCUSSION
The present study analyzes the expression and utilization ofthe major chemokine receptors for HIV-1 entry into normalhuman AM. AM were productively infected with several pri-mary isolates of HIV-1. Expression of the major known HIV-1coreceptors (CCR5 and CXCR4) was detectable at the RNAlevel, whereas surface expression of these receptors occurredat lower levels. However, CCR5-specific chemokines were ableto significantly inhibit HIV-1 replication in AM as did stimu-lation of AM with LPS, which leads to increased expression ofCCR5-specific chemokines. These data suggest that CCR5 isthe predominant coreceptor used by HIV-1 to infect AM, butthat coreceptor expression levels are far lower on AM than onblood monocytes. Importantly, the combined observations thatactivation of normal human AM decreases replication ofHIV-1, downregulates CCR5 expression, and increases therelease of RANTES, MIP-1a, and MIP-1b together suggestthat the interplay of chemokine receptors and chemokine pro-duction plays a major role in the susceptibility of AM to HIV-1infection.
AM and HIV-1 infection. AM play an important role in thepulmonary host defense and are known targets for HIV-1infection (1, 6, 12, 30, 35, 38–40, 43, 45, 50). Although pulmo-nary infections are common in HIV-1-infected individuals andin patients with AIDS, the role of the AM in the progression ofHIV-1-related lung disease is not well defined. Importantly, itis not known if the infection of AM takes place within the lungor is secondary to systemic infection of AM precursors or both(1). AM obtained by bronchoalveolar lavage from HIV-1-in-
FIG. 5. Chemokine receptor expression in AM assessed by Northern analy-sis. RNA obtained from AM of three individuals (lanes A, B, and C) and, as acontrol, from PBM and PBL from individual A. The PBM and PBL were purifiedwith immunomagnetic beads coated with antibodies against CD14 (PBM) andCD3 (PBL), respectively. The cells were analyzed with specific probes for CCR5,CXCR4, and CCR2b. From top to bottom are shown expression of CCR5,CXCR4, CCR2b, and GAPDH. Lanes: 1 to 3, AM for three healthy individuals;4, PBM from individual A; 5, PBL from individual A. The sizes of the mRNAsare indicated in kilobases (kb).
TABLE 1. Secretion of MIP-1a, MIP-1b, RANTES, and eotaxin inculture supernatants following stimulation with LPSa
Chemokineand treatment
Amount (ng/ml) secreted by AM from donor:
1 2 3
MIP-1aLPS2 0.0 0.1 0.1LPS1 5.2 100.1 638.1
MIP-1bLPS2 0.0 0.1 0.2LPS1 4.2 22.5 56.5
RANTESLPS2 0.0 0.0 0.0LPS1 5.8 18.9 17.2
EotaxinLPS2 0.0 0.0 0.0LPS1 0.0 0.0 0.0
a Shown are the means of triplicate samples from the AM from three differentindividuals at 24 h with (1) or without (2) LPS stimulation.
VOL. 73, 1999 ALVEOLAR MACROPHAGES AND HIV-1 CORECEPTORS 5869
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

FIG. 6. Consequences of activation of AM on HIV-1 coreceptor expression, chemokine expression, and the ability of AM to be infected by primary isolates ofHIV-1. (A) AM were stimulated with LPS (100 ng/ml) for either 4 or 48 h and then analyzed for HIV-1 coreceptor expression by Northern analysis. CCR5, CXCR4,and CCR2b expression analyzed by Northern analysis in unstimulated (2) and LPS-stimulated (1) cells. GAPDH expression is used as a control. The sizes of themRNAs are indicated in kilobases (kb). This pattern is representative of three different donors analyzed. (B) RANTES, MIP-1a, MIP-1b, and eotaxin secretion inculture supernatant as measured by ELISA following stimulation of AM with LPS (F) or without LPS stimulation (E). Dashed line represents the limit of detectionof the assay. (C) Influence of LPS stimulation on HIV-1 replication in AM. AM were stimulated with LPS for 12 h and then infected with five primary HIV-1 isolates.After 14 days, HIV-1 p24 antigen levels in the supernatant were measured by ELISA. Shown are data obtained with (1) and without (2) LPS stimulation. The HIV-1isolates are the same as in Fig. 1, and the coreceptor use of these isolates is indicated. Data are means 6 standard errors of the means of triplicate measurements.
5870
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

fected individuals harbor HIV-1 (1, 12, 30, 35, 38–40, 45, 50).In general, the absolute number of HIV-1-infected AM inindividuals with AIDS is lower than that of blood monocytes(35, 38, 39), although a significant increase of HIV-1 in AM,but not monocytes, has been reported in some patients as thedisease progresses (56).
HIV-1 entry in AM. From the results of the present study, itis clear that healthy human AM can be productively infectedwith primary isolates of HIV-1 in vitro. The entry mechanismsfor HIV-1 into AM have not been extensively studied. It isknown that CD4 is critical for HIV infection of AM, and AMare known to express CD4 (26, 32). The discovery that HIV-1
also requires coreceptors of the CC and CXC chemokine fam-ily for entry into lymphocytes and cells of the monocyte-mac-rophage lineage has shed new light on the pathogenesis ofHIV-1 infection (4, 9, 19, 34). Based on a variety of studies ofthe coreceptors for HIV-1 entry into blood monocytes andmonocyte-derived macrophages, it is likely that CCR5 is themain receptor used for entry into these cells (18, 37, 42, 55, 61,64), although recent data obtained by using CCR5-deficientmonocytes demonstrate that CXCR4 can also be used (63).CCR5 seems to play a central role in the transmission of HIV-1in vivo, as individuals homozygous for a 32-bp deletion inCCR5 have increased resistance to HIV-1 infection (33, 51).
FIG. 7. Inhibition of HIV-1 replication in AM by chemokines. AM were infected with primary HIV-1 isolates in the presence of 250 ng of RANTES, MIP-1a,MIP-1b, or SDF-1/ml or all chemokines together. HIV-1 p24 antigen was measured in the culture supernatants on day 10 after virus inoculation. The percent inhibitionwas calculated based on control cultures infected without added chemokines. Data are means of duplicate wells from one (of three) representative experiment. Shownare results for isolates AD2-2 (coreceptor used, CCR5), AD2-6 (coreceptors used, CCR5 and CXCR4), AD3-3 (coreceptor used, CCR5), AD3-7 (coreceptors used,CCR5, CXCR4, CCR2b, and CCR3), and JRFL (coreceptor used, CCR5).
VOL. 73, 1999 ALVEOLAR MACROPHAGES AND HIV-1 CORECEPTORS 5871
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

HIV-1 strains that use CCR5 are present throughout thecourse of the disease, whereas in some individuals, variantsthat use additional coreceptors emerge later in the course ofthe disease (15, 52).
Current knowledge of the pattern of coreceptor expressionon tissue macrophages is limited. Studies with human micro-glial cells of the brain have demonstrated the expression ofCCR3 and CCR5, although recent data suggest a dominantrole for CCR5 in infection (28, 54). Human AM are known toexpress the two orphan seven-transmembrane receptors,GPR-1 and GPR-15, which can be used for simian immuno-deficiency virus entry (22). The present study demonstratesthat primary HIV-1 isolates which solely use CCR5 can repli-cate in AM, in addition to isolates which use more than onecoreceptor. The potential usage of additional coreceptors, in-cluding GPR-1 and/or GPR-15, has not been determined, andthus we cannot rule out the possibility that these receptors playa role in HIV-1 entry into AM.
Coreceptor expression on AM. While all of the evidence isconsistent with the concept that HIV-1 uses predominantlyCCR5 to enter AM, the surface expression of CCR5 and theother major chemokine receptors is much lower on AM thanon autologous blood monocytes, despite the presence ofmRNA. Although there was some variability with differentclones of anti-CCR5 antibodies, the overall detectable surfaceexpression of CCR5 was well below 10% of the cells. Whenusing antibodies against CCR5 other than clone 2D7, a small,distinct, positive subpopulation could be seen with the clonesFAB182F and FAB183F, suggesting that certain CCR5 epitopescould be masked on AM, but this may represent only a smallpercentage of the total population. Likewise, as has been pre-viously shown (32), the expression of CD4 on normal AM islow, suggesting that low-level surface expression of CD4 andcoreceptors is sufficient for infection of HIV-1. It has beenreported that, in retrovirus-modified HeLa cells, CD4 andCCR5 interact in a concentration-dependent manner, i.e., inthe presence of low levels of CD4 expression, high levels ofCCR5 are sufficient for HIV infection and vice versa (41). Inthe present study, however, the levels of surface expression ofboth CD4 and CCR5 were found to be similarly low on AM.Although expression levels of all of the major chemokine re-ceptors are low on healthy AM, the level of expression ofCCR3 is by far the lowest, detectable only by RT-PCR. This issimilar to that previously noted in blood monocytes (18, 37).
Interestingly, the levels of CXCR4 and CCR2 mRNA in AMvary considerably among individuals. As with blood monocyte-derived macrophages (8, 18, 37, 44, 45), the state of activationof AM influences the expression of some HIV-1 coreceptors.Although dependent on time after activation, expression ofCCR2, CXCR4, and CCR5 mRNA is suppressed by activationof AM. CCR2 mRNA levels in monocyte-derived macro-phages and monocytic cell lines are diminished by activation,with moderate decreases in CCR5 mRNA levels (55). Stimu-lation of human endothelial cells with LPS also leads to adecrease in CXCR4 mRNA levels (27).
In addition to receptor mRNA regulation, activation of AMresults in the secretion of the CCR5 ligands RANTES, MIP-1a, and MIP-1b. Chemokines, as the natural ligands of theHIV-1 coreceptors, are able to competitively block HIV-1 in-fection (13, 59). Consistent with these observations, activationof blood-derived monocytes results in decreased replication ofHIV-1 (31) and, as shown in the present study, activation ofAM results in a similar decrease in replication of HIV-1 iso-lates which primarily use CCR5 as the coreceptor for infection.This may be due in part to the decreased expression of thisreceptor following activation and, in part, to the secretion of
CCR5 ligands RANTES, MIP-1a, and MIP-1b, which cancompete with HIV-1 for the coreceptor. Furthermore the de-creased coreceptor expression could have resulted from thesecretion of endogenous beta chemokines, which themselvescould act to downregulate receptor mRNA. RANTES, MIP-1a, and MIP-1b blocked infection of AM with all the fiveprimary HIV-1 isolates tested in our study, a phenomenonwhich has been recently described for infection of AM with theHIV-1 strain BAL (14). SDF-1, the ligand for CXCR4, wasable to block infection to some extent for only one of twoprimary isolates utilizing both CCR5 and CXCR4, but theinhibitory effect was less than that seen for the CCR5-specificchemokines. Although it is possible that CXCR4 may be usedto some degree for HIV-1 entry, indirect effects of SDF-1rather than direct blocking may also be an explanation fordecreased HIV-1 replication in the presence of SDF-1. Forblood monocyte-derived macrophages, HIV-1 replication isinhibited by activation via the release of RANTES, MIP-1a,and MIP-1b (60). Consistent with this concept, MIP-1a hasbeen shown to be produced by AM in increased amounts inHIV-1-infected individuals (17).
Taken as a whole, the present study demonstrates that AMexpress a variety of chemokine receptors relevant for HIV-1entry, that HIV-1 likely enters AM mainly through CCR5, andthat activation of AM can result in decreased infection of thiscell type with HIV-1. Strategies to prevent infection via block-age of chemokine receptors on macrophages, including chem-ically modified chemokines such as AOP-RANTES (57), arecurrently being developed. Intracellular blocking of CCR5 re-ceptor expression via “intrakines” may be equally useful toprevent HIV-1 infection of AM (10, 62).
ACKNOWLEDGMENTS
S.W. and R.C. participated equally in this study.We thank Simon Monard for help with the flow cytometry studies,
Barbara Ferris for technical assistance, Philip L. Leopold and Neil R.Hackett for helpful discussions, and N. Mohamed for help in preparingthe manuscript.
These studies were supported, in part, by NIH grants P01 HL59312and R01 HL59861-01; the Will Rogers Memorial Fund, Los Angeles,Calif.; and GenVec, Inc., Rockville, Md. R.C. was also supported, inpart, by grant AI 41373 from The Aaron Diamond Foundation, NewYork, N.Y.
REFERENCES
1. Agostini, C., L. Trentin, R. Zambello, and G. Semenzato. 1993. HIV-1 andthe lung: infectivity, pathogenic mechanisms, and cellular immune responsestaking place in the lower respiratory tract. Am. Rev. Respir. Dis. 147:1038–1049.
2. Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M.Murphy, and E. A. Berger. 1996. CC CKR5: a RANTES, MIP1a, MIP1breceptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272:1955–1958.
3. Alkhatib, G., E. A. Berger, P. M. Murphy, and J. E. Pease. 1997. Determi-nants of HIV-1 co-receptor function on CC chemokine receptor 3. Impor-tance of both extracellular and transmembrane/cytoplasmic regions. J. Biol.Chem. 272:20420–20426.
4. Bazan, H. A., G. Alkhatib, C. C. Broder, and E. A. Berger. 1998. Patterns ofCCR5, CXCR4, and CCR3 usage by envelope glycoproteins from humanimmunodeficiency type 1 isolates. J. Virol. 72:4485–4491.
5. Berger, E. A. 1997. HIV entry and tropism: the chemokine receptor connec-tion. AIDS 11(Suppl. A):S3–S16.
6. Bezdicek, P., and R. G. Crystal. 1997. Pulmonary macrophages, p. 859–875.In R. G. Crystal, J. B. West, E. R. Weibel, and P. J. Barnes (ed.), The lung:scientific foundations. Lippincott-Raven, Inc., Philadelphia, Pa.
7. Bjorndal, A., H. Deng, M. Jansson, J. R. Fiore, C. Colognesi, A. Karlsson, J.Albert, G. Scarlatti, D. R. Littman, and E. M. Fenyo. 1997. Coreceptor usageof primary human immunodeficiency virus type 1 isolates varies according tobiological phenotype. J. Virol. 71:7478–7487.
8. Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. Mackay. 1997. TheHIV coreceptors CXCR4 and CCR5 are differentially expressed and regu-lated on human T lymphocytes. Proc. Natl. Acad. Sci. USA 94:1925–1930.
5872 WORGALL ET AL. J. VIROL.
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

9. Broder, C. C., and R. G. Collman. 1997. Chemokine receptors and HIV. J.Leukoc. Biol. 62:20–29.
10. Chen, J. D., X. Bai, A. G. Yang, Y. Cong, and S. Y. Chen. 1997. Inactivationof HIV-1 chemokine co-receptor CXCR-4 by a novel intrakine strategy. Nat.Med. 3:1110–1116.
11. Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu,C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, G. Gerard, and J.Sodroski. 1996. The b-chemokine receptors CCR3 and CCR5 facilitateinfection by primary HIV-1 isolates. Cell 85:1135–1148.
12. Clarke, J. R., V. Krishnan, J. Bennett, D. Mitchell, and D. J. Jeffries. 1990.Detection of HIV-1 in human lung macrophages using the polymerase chainreaction. AIDS 4:1133–1136.
13. Cocchi, F., A. L. De Vico, A. Garzino-Demo, S. K. Arya, R. C. Gallo, and P.Lusso. 1995. Identification of RANTES, MIP-1 alpha, and MIP-1 beta as themajor HIV-suppressive factors produced by CD81 T cells. Science 270:1811–1815.
14. Coffey, M. J., C. Woffendin, S. M. Phare, R. M. Strieter, and D. M. Marko-vitz. 1997. Rantes inhibits HIV-1 replication in human peripheral bloodmonocytes and alveolar macrophages. Am. J. Physiol. 272:L1025–L1029.
15. Connor, R. I., K. E. Sheridan, D. Ceradini, S. Choe, and N. R. Landau. 1997.Change in coreceptor use correlates with disease progression in HIV-1-infected individuals. J. Exp. Med. 185:621–628.
16. Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. DiMarzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J.Schall, D. R. Littman, and N. R. Landau. 1996. Identification of a majorco-receptor for primary isolates of HIV-1. Nature 381:661–666.
17. Denis, M., and E. Ghadirian. 1994. Alveolar macrophages from subjectsinfected with HIV-1 express macrophage inflammatory protein-1 alpha(MIP-1 alpha): contribution to the CD81 alveolitis. Clin. Exp. Immunol.96:187–192.
18. Di Marzio, P., J. Tse, and N. R. Landau. 1998. Chemokine receptor regu-lation and HIV type 1 tropism in monocyte-macrophages. AIDS Res. Hum.Retroviruses 14:129–138.
19. Doms, R. W., and S. C. Peiper. 1997. Unwelcome guests with master keys:how HIV uses chemokine receptors for cellular entry. Virology 235:179–190.
20. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. Peiper, M.Parmentier, R. G. Collman, R. W. Doms. 1996. A dual-tropic, primary HIV-1isolate that uses fusin and the b-chemokine receptors CKR-5, CKR-3,CKR-2b as fusion cofactors. Cell 85:1149–1158.
21. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Naga-shima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A.Paxton. 1996. HIV-1 entry into CD41 cells is mediated by the chemokinereceptor CC-CKR-5. Nature 381:667–673.
22. Farzan, M., H. Choe, K. Martin, L. Marcon, W. Hofmann, G. Karlsson, Y.Sun, P. Barrett, N. Marchand, N. Sullivan, N. Gerard, C. Gerard, and J.Sodroski. 1997. Two orphan seven-transmembrane segment receptors whichare expressed in CD4-positive cells support simian immunodeficiency virusinfection. J. Exp. Med. 186:405–411.
23. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger. 1996. HIV-1 entrycofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science 272:872–877.
24. Frade, J. M. R., M. Llorente, M. Mellado, J. Alcami, J. C. Gutierrez-Ramos,A. Zaballos, G. del Real, and A. C. Martinez. 1997. The amino-terminaldomain of the CCR2 chemokine receptor acts as a coreceptor for HIV-1infection. J. Clin. Investig. 100:497–502.
25. Granelli-Piperno, A., B. Moser, M. Pope, D. Chen, Y. Wei, F. Isdell, U.O’Doherty, W. Paxton, R. Koup, S. Mojsov, N. Bhardwaj, I. Clark-Lewis, M.Baggiolini, and R. M. Steinman. 1996. Efficient interaction of HIV-1 withpurified dendritic cells via multiple chemokine coreceptors. J. Exp. Med.184:2433–2438.
26. Guay, L. A., J. G. Sierra-Madero, C. K. Finegan, and E. A. Rich. 1997.Mediation of entry of human immunodeficiency virus-1 into alveolar mac-rophages by CD4 without facilitation by surfactant-associated protein-a.Am. J. Respir. Cell Mol. Biol. 16:421–428.
27. Gupta, S. K., P. G. Lysko, K. Pillarisetti, E. Ohlstein, and J. M. Stadel. 1998.Chemokine receptors in human endothelial cells. Functional expression ofCXCR4 and its transcriptional regulation by inflammatory cytokines. J. Biol.Chem. 273:4282–4287.
28. He, J., Y. Chen, M. Farzan, H. Choe, A. Ohagen, S. Gartner, J. Busciglio, X.Yang, W. Hofmann, W. Newman, C. R. Mackay, J. Sodroski, and D.Gabuzda. 1997. CCR3 and CCR5 are co-receptors for HIV-1 infection ofmicroglia. Nature 385:645–649.
29. Janoff, E. N., R. F. Breiman, C. L. Daley, and P. C. Hopewell. 1992. Pneu-mococcal disease during HIV infection: epidemiologic, clinical, and immu-nologic perspectives. Ann. Intern. Med. 117:314–324.
30. Jeffrey, A. A., D. Israel-Biet, J. M. Andrieu, P. Even, and A. Venet. 1991. HIVisolation from pulmonary cells derived from bronchoalveolar lavage. Clin.Exp. Immunol. 84:488–492.
31. Kornbluth, R. S., P. S. Oh, J. R. Munis, P. H. Cleveland, and D. D. Richman.1989. Interferons and bacterial lipopolysaccharide protect macrophagesfrom productive infection by human immunodeficiency virus in vitro. J. Exp.Med. 169:1137–1151.
32. Lewin, S. R., S. Sonza, L. B. Irving, C. F. McDonald, J. Mills, and S. M.Crowe. 1996. Surface CD4 is critical to in vitro HIV infection of humanalveolar macrophages. AIDS Res. Hum. Retroviruses 12:877–883.
33. Liu, R., W. A. Paxton, S. Choe, D. Ceradini, S. R. Martin, R. Horuk, M. E.MacDonald, H. Stuhlmann, R. A. Koup, and N. R. Landau. 1996. Homozy-gous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell 86:367–377.
34. Moore, J. P., A. Trkola, and T. Dragic. 1997. Co-receptors for HIV-1 entry.Curr. Opin. Immunol. 9:551–562.
35. Moreno, P., M. J. Rebollo, F. Pulido, R. Rubio, A. R. Noriega, and R.Delgado. 1996. Alveolar macrophages are not an important source of viralproduction in HIV-1 infected patients. AIDS 10:682–684.
36. Murray, J. F., and J. Mills. 1990. Pulmonary infectious complications ofhuman immunodeficiency virus infection. Part I. Am. Rev. Respir. Dis.141:1356–1372.
37. Naif, H. M., S. Li, M. Alali, A. Sloane, L. Wu, M. Kelly, G. Lynch, A. Lloyd,and A. L. Cunningham. 1998. CCR5 expression correlates with susceptibilityof maturing monocytes to human immunodeficiency virus type 1 infection.J. Virol. 72:830–836.
38. Nakata, K., M. Weiden, T. Harkin, D. Ho, and W. N. Rom. 1995. Low copynumber and limited variability of proviral DNA in alveolar macrophagesfrom HIV-1-infected patients: evidence for genetic differences in HIV-1between lung and blood macrophage populations. Mol. Med. 1:744–757.
39. Pearce, T. E., M. Nowakowski, E. Eden, Z. B. Huang, P. Steiner, M. Sha-habuddin, M. J. Potash, and D. J. Volsky. 1993. Uniform detection of HIV-1in alveolar macrophages of pediatric but not adult AIDS patients. J. Leukoc.Biol. 53:722–726.
40. Plata, F., F. Garcia-Pons, A. Ryter, F. Lebargy, M. M. Goodenow, M. H. Dat,B. Autran, and C. Mayaud. 1990. HIV-1 infection of lung alveolar fibroblastsand macrophages in humans. AIDS Res. Hum. Retroviruses 6:979–986.
41. Platt, E. J., K. Wehrly, S. E. Kuhmann, B. Chesebro, and D. Kabat. 1998.Effects of CCR5 and CD4 cell surface concentrations on infections by mac-rophagetropic isolates of human immunodeficiency virus type 1. J. Virol.72:2855–2864.
42. Rana, S., G. Besson, D. G. Cook, J. Rucker, R. J. Smyth, Y. Yi, J. D. Turner,H.-H. Guo, J.-G. Du, S. C. Peiper, E. Lavi, M. Samson, F. Libert, C. Lies-nard, G. Vassart, R. W. Doms, M. Parmentier, and R. G. Collman. 1997.Role of CCR5 in infection of primary macrophages and lymphocytes bymacrophage-tropic strains of human immunodeficiency virus: resistance topatient-derived and prototype isolates resulting from the Dccr5 mutation.J. Virol. 71:3219–3227.
43. Reynolds, H. Y. 1997. Integrated host defense against infections, p. 2353–2365. In R. G. Crystal, J. B. West, E. R. Weibel, and P. J. Barnes (ed.), Thelung: scientific foundations. Lippincott-Raven, Inc., Philadelphia, Pa.
44. Rich, E. A., I. S. Chen, J. A. Zack, M. L. Leonard, and W. A. O’Brien. 1992.Increased susceptibility of differentiated mononuclear phagocytes to produc-tive infection with human immunodeficiency virus-1 (HIV-1). J. Clin. Inves-tig. 89:176–183.
45. Rose, R. M., A. Krivine, P. Pinkston, J. M. Gillis, A. Huang, and S. M.Hammer. 1991. Frequent identification of HIV-1 DNA in bronchoalveolarlavage cells obtained from individuals with the acquired immunodeficiencysyndrome. Am. Rev. Respir. Dis. 143:850–854.
46. Ross, T. M., and B. R. Cullen. 1998. The ability of HIV type 1 to CCR-3 asa coreceptor is controlled by envelope V1/V2 sequences acting in conjunc-tion with a CCR-5 tropic V3 loop. Proc. Natl. Acad. Sci. USA 95:7682–7686.
47. Rubbet, A., C. Combadiere, M. Ostrowski, J. Arthos, M. Dybul, E. Machado,M. A. Cohen, J. A. Hoxie, P. M. Murphy, A. S. Fauchi, and D. Weisman.1998. Dendritic cells express multiple chemokine receptors used as corecep-tors for HIV entry. J. Immunol. 160:3933–3941.
48. Rucker, J., A. L. Edinger, M. Sharron, M. Samson, B. Lee, J. F. Berson, Y.Yi, B. Marguilies, R. G. Collman, B. J. Doranz, M. Parmentier, and R. W.Doms. 1997. Utilization of chemokine receptors, orphan receptors, and her-pesvirus-encoded receptors by diverse human and simian immunodeficiencyviruses. J. Virol. 71:8999–9007.
49. Russi, T. J., and R. G. Crystal. 1997. Bronchoalveolar lavage, p. 371. In R. G.Crystal, J. B. West, E. R. Weibel, and P. J. Barnes (ed.), The lung: scientificfoundations. Lippincott-Raven, Inc., Philadelphia, Pa.
50. Salahuddin, S. Z., R. M. Rose, J. E. Groopman, P. D. Markham, and R. C.Gallo. 1986. Human T lymphotropic virus type III infection of human alve-olar macrophages. Blood 68:281–284.
51. Samson, M., F. Libert, B. J. Doranz, J. Rucker, C. Liesnard, C. M. Farber,S. Saragosti, C. Lapoumeroulie, J. Cognaux, C. Forceille, G. Muyldermans,C. Verhofstede, G. Burtonboy, M. Georges, T. Imai, S. Rana, Y. Yi, R. J.Smyth, R. G. Collman, R. W. Doms, G. Vassart, and M. Parmentier. 1996.Resistance to HIV-1 infection in caucasian individuals bearing mutant allelesof the CCR-5 chemokine receptor gene. Nature 382:722–725.
52. Scarlatti, G., E. Tresoldi, A. Bjorndal, R. Fredriksson, C. Colognesi, H. K.Deng, M. S. Malnati, A. Plebani, A. G. Siccardi, D. R. Littman, E. M. Fenyo,and P. Lusso. 1997. In vivo evolution of HIV-1 co-receptor usage andsensitivity to chemokine-mediated suppression. Nat. Med. 3:1259–1265.
53. Schuitemaker, H., M. Koot, N. A. Kootstra, M. W. Dercksen, R. E. de Goede,R. P. van Steenwijk, J. M. Lange, J. K. Schattenkerk, F. Miedema, and M.
VOL. 73, 1999 ALVEOLAR MACROPHAGES AND HIV-1 CORECEPTORS 5873
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from

Tersmette. 1992. Biological phenotype of human immunodeficiency virustype 1 clones at different stages of infection: progression of disease is asso-ciated with a shift from monocytotropic to T-cell-tropic virus population.J. Virol. 66:1354–1360.
54. Shieh, J. T., A. V. Albright, M. Sharron, S. Gartner, J. Strizki, R. W. Doms,and F. Gonzalez-Scarano. 1998. Chemokine receptor utilization by humanimmunodeficiency virus type 1 isolates that replicate in microglia. J. Virol.72:4243–4249.
55. Sica, A., A. Saccani, A. Borsatti, C. A. Power, T. N. Wells, W. Luini, N.Polentarutti, S. Sozzani, and A. Mantovani. 1997. Bacterial lipopolysaccha-ride rapidly inhibits expression of C-C chemokine receptors in human mono-cytes. J. Exp. Med. 185:969–974.
56. Sierra-Madero, J. G., Z. Toossi, D. L. Hom, C. K. Finegan, E. Hoenig, andE. A. Rich. 1994. Relationship between load of virus in alveolar macrophagesfrom human immunodeficiency virus type 1-infected persons, production ofcytokines, and clinical status. J. Infect. Dis. 169:18–27.
57. Simmons, G., P. R. Clapham, L. Picard, R. E. Offord, M. M. Rosenkilde,T. W. Schwartz, R. Buser, T. N. C. Wells, and A. E. Proudfoot. 1997. Potentinhibition of HIV-1 infectivity in macrophages and lymphocytes by a novelCCR5 antagonist. Science 276:276–279.
58. Simmons, G., D. Wilkinson, J. D. Reeves, M. T. Dittmar, S. Beddows, J.Weber, G. Carnegie, U. Desselberger, P. W. Gray, R. A. Weiss, and P. R.Clapham. 1996. Primary, syncytium-inducing human immunodeficiency virustype 1 isolates are dual-tropic and most can use either LESTR or CCR5 ascoreceptors for virus entry. J. Virol. 70:8355–8360.
59. Trkola, A., W. A. Paxton, S. P. Monard, J. A. Hoxie, M. A. Siani, D. A.Thompson, L. Wu, C. R. Mackay, R. Horuk, and J. P. Moore. 1998. Geneticsubtype-independent inhibition of human immunodeficiency virus type 1replication by CC and CXC chemokines. J. Virol. 72:396–404.
60. Verani, A., G. Scarlatti, M. Comar, E. Tresoldi, S. Polo, M. Giacca, P. Lusso,A. G. Siccardi, and D. Vercelli. 1997. C-C chemokines released by lipopoly-saccharide (LPS)-stimulated human macrophages suppress HIV-1 infectionin both macrophages and T cells. J. Exp. Med. 185:805–816.
61. Wu, L., W. A. Paxton, N. Kassam, N. Ruffing, J. B. Rottman, N. Sullivan, H.Choe, J. Sodroski, W. Newman, R. A. Koup, and C. R. Mackay. 1997. CCR5levels and expression pattern correlate with infectability by macrophage-tropic HIV-1, in vitro. J. Exp. Med. 185:1681–1691.
62. Yang, A. G., X. Bai, X. F. Huang, C. Yao, and S. Chen. 1997. Phenotypicknockout of HIV type 1 chemokine coreceptor CCR-5 by intrakines aspotential therapeutic approach for HIV-1 infection. Proc. Natl. Acad. Sci.USA 94:11567–11572.
63. Yi, Y., S. Rana, J. D. Turner, N. Gaddis, and R. G. Collman. 1998. CXCR-4is expressed by primary macrophages and supports CCR5-independent in-fection by dual-tropic but not T-tropic isolates of human immunodeficiencyvirus type 1. J. Virol. 72:772–777.
64. Zaitseva, M., A. Blauvelt, S. Lee, C. K. Lapham, V. Klaus-Kovtun, H.Mostowski, J. Manischewitz, and H. Golding. 1997. Expression and functionof CCR5 and CXCR4 on human Langerhans cells and macrophages: impli-cations for HIV primary infection. Nat. Med. 3:1369–1375.
5874 WORGALL ET AL. J. VIROL.
on Decem
ber 3, 2013 by guesthttp://jvi.asm
.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















