
Evaluation of Salmonella Enteritidis and Escherichia coli ...

Expression and morphology of enterotoxigenicEscherichia coli surface antigen CS31A in E. coliK12 and Vibrio cholerae
Kathrin Kuehni-Boghenbor, Helene A. Jordi, Joachim Frey, Edy M. Vilei,Didier Favre, and Michael H. Stoffel
Abstract: Enterotoxigenic Escherichia coli (ETEC) is known as a worldwide cause of diarrheal disease. The pathogenesisinvolves the attachment of the microorganisms to the mucosa and the production of enterotoxins. Surface expression ofCS31A fimbriae was assessed by Western blots, dot blots, immunofluorescence, and electron microscopy using negativestaining and immunogold labeling. These investigations revealed significant differences in both the morphology of thewild-type and recombinant strains and the antigen exposure of CS31A in the wild-type and recombinant strains. In thewild-type ETEC strain, expression of CS31A was subject to phase variation. The recombinant E. coli strain producedCS31A but was prone to epitope shedding. In Vibrio cholerae vaccine strain CVD 103-HgR, the recombinant CS31Aantigen was expressed but was only found intracellularly. Thus, E. coli strains seem to lend themselves better to the devel-opment of recombinant vaccines expressing ETEC-specific antigens at the cell’s surface than strains from other orders orgenera such as V. cholerae.
Key words: ETEC, CS31A, fimbriae, Vibrio cholerae.
Résumé : Escherichia coli entérotoxinogène (ETEC) est une cause reconnue de maladie diarrhéique à travers le monde. Lapathogenèse implique l’attachement des microorganismes à la muqueuse et la production d’entérotoxines. L’expression desurface des fimbriae CS31A a été évaluée par buvardage Western, transfert en point, immunofluorescence et microscopieélectronique à l’aide d’une coloration négative et par marquage par immunogold. Ces études ont révélé l’existence de diffé-rences significatives tant au plan de la morphologie des souches sauvage et recombinante, que de l’exposition de l’antigèneCS31A par les souches sauvage et recombinante. Chez la souche ETEC sauvage, l’expression de CS31A était sujette à unevariation périodique. La souche recombinante d’E. coli produisait CS31A mais elle était sujette à un délestage d’épitopes.Chez la souche de vaccin CVD 103-HgR de Vibrio cholerae, l’antigène CS31A recombinant était exprimé mais de façon in-tracellulaire seulement. Ainsi, les souches d’E. coli semblent elles-mêmes se prêter davantage au développement de vaccinsrecombinants exprimant des antigènes spécifiques aux ETEC à la surface cellulaire que des souches d’autres ordres ou d’au-tres genres, comme V. cholerae.
Mots‐clés : ETEC, CS31A, fimbriae, Vibrio cholerae.
[Traduit par la Rédaction]
Introduction
Infections with enterotoxigenic Escherichia coli (ETEC)represent a continuous problem. They account globally foran estimated 840 million cases of human gastrointestinal in-fections and some 380 000 casualties per year (Gupta et al.2008). Since natural infection provides a partial immunity(Tacket and Levine 1997), a vaccine strategy may be ex-pected to make a decisive contribution to disease control to-gether with dietary precautions.
The rationale for developing safe and effective ETEC vac-cines relies on knowledge of the main factors involved in thepathogenesis of ETEC diarrhea, which involves 2 crucialsteps: (i) the attachment of the bacteria to the mucosa of thesmall intestine and (ii) the production of enterotoxins. Colo-nization of the intestine is mediated through a variety of col-onization factors (CFs), which are classified as coli surfaceantigens (CS) or colonization factor antigens (CFA). Theirmorphological equivalents are termed fimbriae or pili. Todate, over 25 different types of CFA have been reported
Received 30 January 2012. Accepted 21 February 2012. Published at www.nrcresearchpress.com/cjm on 18 May 2012.
K. Kuehni-Boghenbor,* H.A. Jordi,* and M.H. Stoffel.* Division of Veterinary Anatomy, University of Berne, Veterinary School,P.O. Box 8466 CH-3001 Bern, Switzerland.
J. Frey* and E.M. Vilei.* Institute of Veterinary Bacteriology, University of Berne, P.O. Box 8466 CH-3001 Bern, Switzerland.D. Favre. Crucell Switzerland Limited, Department of Regulatory Affairs, Rehhagstrasse 79, 3018 Bern, Switzerland.
Corresponding author: Michael H. Stoffel (e-mail: [email protected]).
*These authors contributed equally to this work.
728
Can. J. Microbiol. 58: 728–737 (2012) doi:10.1139/W2012-047 Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

(Girard et al. 2006) and epidemiological studies have shownthat the expression of the different human ETEC CFs variesaccording to geographical location and time (Gupta et al.2008). Therefore, immunization against selected CFs will benecessary but will provide only partial protection.Besides their ability to colonize the intestinal mucosa,
ETEC are capable of producing a heat-labile (LT) and aheat-stable (ST) toxin, both of which elicit a hypersecretionof water and salts by the enterocytes. These toxins are poly-peptides and, thus, are potential targets for the immune sys-tem. With respect to the heat-stable toxin, ETEC strainsisolated from humans usually produce STa (Wolf 1997),whereas ETEC strains isolated from animals predominantlyproduce STb. As for STa, it encompasses STh and STp.These designations reflect the previous contention that SThwas mainly of human origin and STp of porcine origin. Dueto its small size of only 19 amino acids (Aimoto et al. 1982),STh is a hapten and does not elicit a proper immune re-sponse on its own. However, when combined with a carriermolecule, anti-STh antibodies are produced (Batisson andDer Vartanian 2000). Thus, a vaccine combining a surfaceantigen with STh might be expected to elicit an immune re-sponse against both pathogenic factors. The rationale of theapproach chosen in the present study was to assess the ex-pression of ETEC CS31A fimbriae and to study their mor-phology in view of their potential suitability as carriers ofSTh. CS31A is a plasmid-encoded fibrillar protein associatedwith animal and human pathogenic isolates of ETEC(Batisson et al. 2000; Girardeau et al. 1988). The appearanceof CS31A fimbriae is described as a capsule-like zonearound the bacteria. It consists of very fine, wiry, fibrillarstructures of 2 nm in diameter (Girardeau et al. 1988). Themajor protein subunit of E. coli CS31A ClpG has beenshown to be a good candidate for the presentation of heterol-ogous epitopes to the immune system (Bousquet et al. 1994).Escherichia coli K12 strain DH10B and Vibrio cholerae
CVD 103-HgR were chosen as host strains. Laboratorystrains of E. coli K12 are biologically safe (Kuhnert et al.
1997), whereas V. cholerae CVD 103-HgR is highly effectiveand a safe live vaccine against cholera. Furthermore, the mo-lecular mechanisms of pathogenicity of ETEC and V. chol-erae are very similar, thus making CVD 103-HgR apromising carrier strain for the development of new vaccines(Favre et al. 2006).The goal of the present study was to investigate the ex-
pression and localization of CS31A in V. cholerae CVD103-HgR and in E. coli DH10B as compared with the ho-mologous CS31A-producing ETEC strain 31A. Surface ex-pression was assessed by negative staining followed bytransmission electron microscopy. In addition, antigenicitywas judged by immunofluorescence and immunogold label-ing.
Material and methods
Strains and culture conditionsInformation pertaining to all strains used is listed in Ta-
ble 1.Bacteria were grown on Luria–Bertani or Minca agar at
37 °C overnight. Antibiotics were included as appropriate(34 µg/mL chloramphenicol).
Production of antibodiesTwo polyclonal antibodies against CS31A were used: anti-
serum MDV (kindly provided by M. Der Vartanian, Labora-toire de Microbiologie, Institut National de la RechercheAgronomique, Centre de Recherche de Clermont-Ferrand-Theix, Saint-Genès-Champanelle, France) and the mono-specific antiserum EV. To produce the monospecific, poly-clonal anti-CS31A antiserum EV, a poly-histidine-tailedClpG protein was constructed by amplification of the clpGgene sequence encoding the 257 amino acids constituting themature fimbrial subunit of CS31A using the primers CS31A-NcoI-L and CS31A-BamHI-R. The PCR product was di-gested by NcoI and BamHI for cloning into pETHIS-1(Schaller et al. 1999). The construct was analyzed by restric-
Table 1. Properties of strains used.
StrainAdhesinantigena
Toxinantigenb Reference
E. coli wild typeH10407 CFA/I LT, ST Evans et al. 197931A CS31A — Gift from M. Der Vartanian
E. coli K12DH10B — — Grant et al. 1990DH10B (pEH524)c CS31A — This study
V. choleraeCVD 103-HgR None CT-B Ketley et al. 1993CVD 103-HgR (pEH524)c CS31A CT-B D. Favre, unpublishedCVD 103-HgR (pEHProSTC28)d CS31A CT-B, ST D. Favre, unpublished
aCFA/I, colonization factor antigen.bLT, heat labile toxin; ST, heat stable toxin; CT-B, cholera toxin subunit B.cpEH524: pHSG575 carrying the CS31A fimbria-encoding clp operon; kindly provided by Dr. M. Der Vartanian.dpEHProSTC28: pEH524 in which the clpG gene was replaced by the clpG::STh fusion gene of pProSTC28 (Ba-
tisson and Der Vartanian 2000); kindly provided by Dr. M. Der Vartanian.
Kuehni-Boghenbor et al. 729
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

tion enzyme digestion and partial DNA sequencing and thenintroduced by transformation into E. coli BL21 (DE3) for ex-pression of the protein. Crude E. coli extract was centrifuged(15 000g for 20 min) and resuspended several times in PNbuffer (50 mmol/L NaH2PO4, pH 8.0, 300 mmol/L NaCl).Analysis of the sonicated fractions on SDS – 10% acrylamidegels showed that the induced protein was contained in thepellet (i.e., insoluble fraction). Guanidine hydrochloride wasadded to a final concentration of 6 mol/L to the insolublefraction, and the mixture was loaded onto a Ni–nitrilotriaceticacid–agarose column (QIAGEN) and washed with PNG buf-fer (50 mmol/L NaH2PO4, pH 8.0, 300 mmol/L NaCl, 6 mol/Lguanidine hydrochloride). Step elution of the protein wasperformed with PNG buffer at different pH values (7.0,6.0, 5.5, 5.0, and 4.5), and fractions of 1 mL were col-lected. The fractions were dialyzed overnight against PNbuffer and analyzed on SDS – 10% acrylamide gels. Thepurified ClpG-6xHis protein eluted at pH 5.0. The polyclo-nal monospecific serum directed against poly-histidine-tailedprotein CS31A was obtained by 3 subcutaneous immuniza-tions of rabbits with ∼80 µg of purified protein at intervalsof 3 weeks. Animals were bled and sera were adsorbedagainst inactivated E. coli cells to remove irrelevant cross-reacting antibodies.Monoclonal antibodies against CFA/I were generated in
BALB/c mice according to standard methods (Köhler andMilstein 1975). Briefly, BALB/c mice were immunized3 times subcutaneously with 10 µg of purified CFA/I and afourth time intraperitoneally with 100 µg. For fusion, spleencells were mixed with Ag8 fusion partner cells in a ratio of2:1 in a PEG–DMSO (polyethylene glycol – dimethyl sulfox-ide) solution. Following fusion, the cells were cultivated inthe presence of feeder cells (mouse peritoneal macrophages)under selection pressure of azaserine and hypoxanthine (bothfrom Sigma, Buchs, Switzerland). Following a primaryscreening, positive cells were expanded and finally clonedtwice by limiting dilution. The monoclonal anti-CFA/I anti-body named 2E5 is of the IgG1 isotype. MAb2E5 was ableto efficiently agglutinate a CFA/I-expressing ETEC wild-type strain. All animal experiments were carried out in ac-cordance with national regulations of Switzerland and ap-proved by the State Veterinary Service of the Canton ofBerne.
Western blotsFor protein extracts, cells were rinsed off the culture plates
with 2 mL of phosphate-buffered saline (PBS). Cell suspen-sions were mixed with an equal volume of SDS–PAGE sam-ple buffer. The samples were boiled for 10 min, and 5–10 µLwas loaded onto a 3× 12% SDS–PAGE gel. Gels were trans-ferred to nitrocellulose membranes (Trans-Blot Transfer Me-dium, Bio-Rad Laboratories AG, Reinach, Switzerland).Blots were blocked in 1% powdered milk for 30 min and in-cubated with the corresponding primary antibodies overnightat 4 °C. Affinity-purified phosphatase-labeled goat anti-rabbitIgG antibodies (Kirkegaard & Perry Laboratories/KPL Pro-tein Research Products, BioConcept AG, Allschwil, Switzer-land) were used as secondary antibodies and were developedwith BCIP–NBT (5-bromo-4-chloro-3-indolylphosphate 4-toluidine salt – 4-nitroblue tetrazolium chloride, Roche Diag-nostics GmbH, Mannheim, Germany) for 3–5 min.
Dot blotsEither 10 µL of bacterial suspension were spotted on a ni-
trocellulose membrane (Trans-Blot Transfer Medium) or acolony from the culture plate was transferred directly ontothe membrane and dried. Dot blots were then performed asdescribed for Western blots.
ImmunofluorescenceA colony was collected from the culture plate, streaked
onto a coverslip, and air-dried for a few hours. The bacteriawere incubated with Baclight (BacLight Green bacterialstain, Molecular Probes, Invitrogen AG, Basel, Switzerland)for 15 min at room temperature, labeled with correspondingprimary antibodies for 1 h, and incubated for 1 h with eitherTRITC-conjugated goat anti-mouse IgG (Sigma-AldrichChemie GmbH, Buchs, Switzerland) or Cy3-conjugated goatanti-rabbit IgG (Jackson Immunoresearch/Milan Analytica,La Roche, Switzerland). The coverslips were examined witha Zeiss AxioImager Z1. Micrographs were taken with a 63×/1.4 Plan-Apochromat oil immersion objective and recordedwith a Zeiss AxioCam MRm digital camera controlled withthe Zeiss software AxioVision 4.6.3.
Transmission electron microscopyBacteria were rinsed off the culture plates with 0.85%
NaCl pre-warmed to 37 °C.They were adsorbed onto carbon-stabilized parlodion-
coated nickel grids for 30 min by placing a grid upsidedown onto a drop of bacterial suspension. Likewise, adsorbedbacteria were washed once on a drop of dH2O for 5 min.Negative staining was performed by floating the grids on adrop of 0.25% phosphotungstic acid in dH2O (pH 7.2) for 5–10 s. Excess liquid was blotted off with filter paper and sam-ples were air-dried.For immunogold labeling, grids were floated on correspond-
ing primary antibodies diluted 1:8 in PBS–BT (0.9% NaCl in0.01 mol/L PBS added with 0.2% bovine serum albumin(BSA) and 0.05% Tween 20) for 60 min. Grids were washedin PBS–BT for 5 min and incubated for another 60 min on adrop of either goat 15 nm gold-conjugated anti-mouse IgG di-luted 1:100 in PBS or goat 15 nm gold-conjugated anti-rabbitIgG (both from British Biocell International, Brunschwig,Germany). Finally, the grids were washed twice in dH2Ofor 5 min. After blotting off excess liquid with filter paper,negative staining, and air-drying, grids were examined in aZeiss 109 transmission electron microscope (Zeiss, Oberko-chen, Germany) with a GATAN wide-angle slow-scan CCDCamera 689 (GATAN GmbH, Munich, Germany).
Control experimentsWild-type strain E. coli 31A, which expresses the corre-
sponding CS31A surface antigen, was used as a positive con-trol strain to corroborate the suitability of antibodies and theirspecificity. Control reactions for immunogold labeling in-cluded omission of primary antibodies and use of inappropri-ate antibodies as substitutes for pertinent primary antibodies,i.e., rabbit anti-calcitonin (Anawa Biomedical Services andProducts, Zurich, Switzerland) (data not shown). Absence ofsurface antigens in host strains used (E. coli DH10B, CVD103-HgR) was demonstrated by negative staining andimmunogold labeling with polyclonal anti-CS31A antibodies.
730 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

Results
Western blot assaysThe expression of CS31A fimbriae was tested on Western
blots using 2 different preparations of anti-CS31A antibodies(Figs. 1a and 1b). Both antisera revealed the purified CS31Aprotein as 2 distinct bands near the computed molecular massof 28.5 kDa. The lower band was stronger. In wild-type
strain E. coli 31A and in recombinant strain DH10B(pEH524), only the lower molecular mass protein was de-tected with either serum. In strain CVD 103-HgR (pEH524),antiserum MDV revealed a feeble but clear band, whereasantiserum EV yielded an even weaker signal. In recombinantstrain CVD 103-HgR (pEHProSTC28), the fusion peptide ofCS31A, ClpG::STh, produced a similar double-band patternas the purified antigen. However, the upper band was indica-
Fig. 1. Expression of CS31A fimbrial protein. Western blot assays of different recombinant Escherichia coli and Vibrio cholera strains with 2different antibodies (a) MDV (b) EV. Escherichia coli K12 (DH10B) and CVD 103-HgR were negative controls. As positive control theimmunogen (purified CS31A Protein) for antiserum EV was used.
Kuehni-Boghenbor et al. 731
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.
tive of a higher molecular mass of approximately 30 kDa.The signal for CS31A was stronger in both the wild-typeE. coli 31A and the recombinant E. coli DH10B than in therecombinant V. cholerae strains. As for the immunoreactivityof V. cholerae (pEHProSTC28), it was again stronger withantiserum MDV than with antiserum EV (Fig. 1).No specific signal was detected in negative control strains
DH10B and CVD 103-HgR. However, the polyclonal anti-bodies of antiserum MDV uncovered nonspecific bands inall strains.
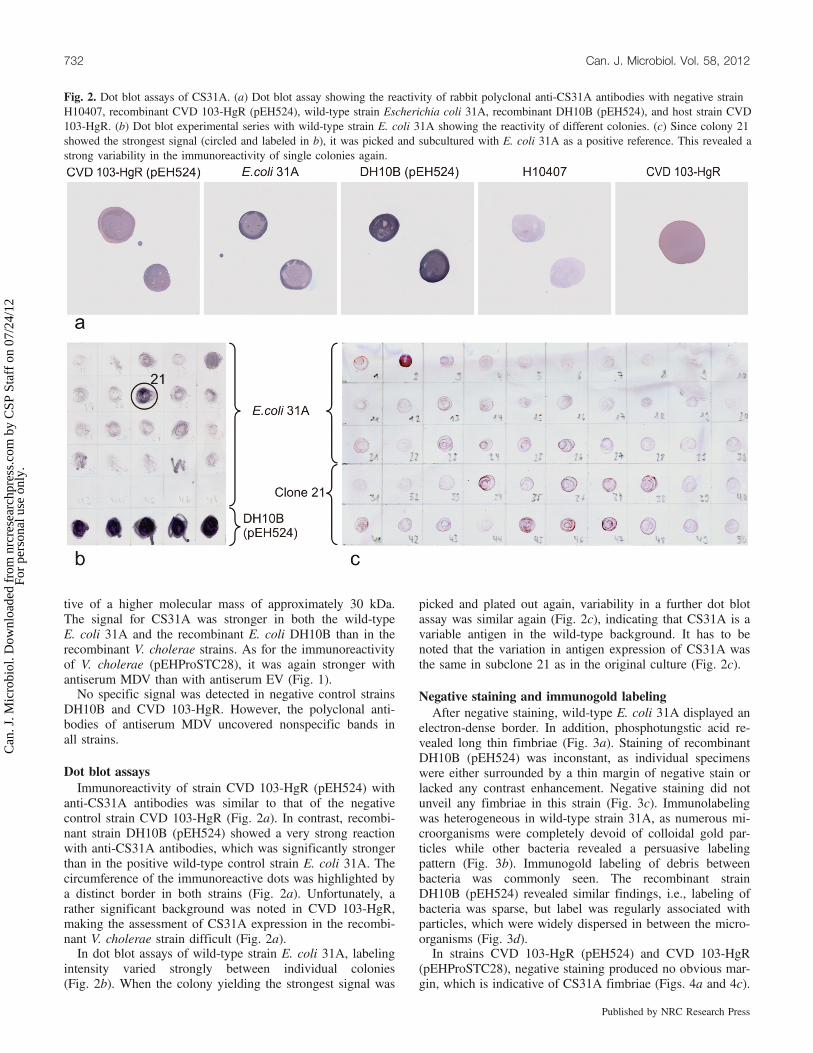
Dot blot assaysImmunoreactivity of strain CVD 103-HgR (pEH524) with
anti-CS31A antibodies was similar to that of the negativecontrol strain CVD 103-HgR (Fig. 2a). In contrast, recombi-nant strain DH10B (pEH524) showed a very strong reactionwith anti-CS31A antibodies, which was significantly strongerthan in the positive wild-type control strain E. coli 31A. Thecircumference of the immunoreactive dots was highlighted bya distinct border in both strains (Fig. 2a). Unfortunately, arather significant background was noted in CVD 103-HgR,making the assessment of CS31A expression in the recombi-nant V. cholerae strain difficult (Fig. 2a).In dot blot assays of wild-type strain E. coli 31A, labeling
intensity varied strongly between individual colonies(Fig. 2b). When the colony yielding the strongest signal was
picked and plated out again, variability in a further dot blotassay was similar again (Fig. 2c), indicating that CS31A is avariable antigen in the wild-type background. It has to benoted that the variation in antigen expression of CS31A wasthe same in subclone 21 as in the original culture (Fig. 2c).
Negative staining and immunogold labelingAfter negative staining, wild-type E. coli 31A displayed an
electron-dense border. In addition, phosphotungstic acid re-vealed long thin fimbriae (Fig. 3a). Staining of recombinantDH10B (pEH524) was inconstant, as individual specimenswere either surrounded by a thin margin of negative stain orlacked any contrast enhancement. Negative staining did notunveil any fimbriae in this strain (Fig. 3c). Immunolabelingwas heterogeneous in wild-type strain 31A, as numerous mi-croorganisms were completely devoid of colloidal gold par-ticles while other bacteria revealed a persuasive labelingpattern (Fig. 3b). Immunogold labeling of debris betweenbacteria was commonly seen. The recombinant strainDH10B (pEH524) revealed similar findings, i.e., labeling ofbacteria was sparse, but label was regularly associated withparticles, which were widely dispersed in between the micro-organisms (Fig. 3d).In strains CVD 103-HgR (pEH524) and CVD 103-HgR
(pEHProSTC28), negative staining produced no obvious mar-gin, which is indicative of CS31A fimbriae (Figs. 4a and 4c).
Fig. 2. Dot blot assays of CS31A. (a) Dot blot assay showing the reactivity of rabbit polyclonal anti-CS31A antibodies with negative strainH10407, recombinant CVD 103-HgR (pEH524), wild-type strain Escherichia coli 31A, recombinant DH10B (pEH524), and host strain CVD103-HgR. (b) Dot blot experimental series with wild-type strain E. coli 31A showing the reactivity of different colonies. (c) Since colony 21showed the strongest signal (circled and labeled in b), it was picked and subcultured with E. coli 31A as a positive reference. This revealed astrong variability in the immunoreactivity of single colonies again.
732 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

Recombinant strains CVD 103-HgR (pEH524) and CVD103-HgR (pEHProSTC28) were devoid of any colloidal goldlabeling (Figs. 4b and 4d).Negative staining and immunogold labeling of unaltered
host strains DH10B (Figs. 3e and 3f) and CVD 103-HgR
(Figs. 4e and 4f) with polyclonal CS31A antibodies yieldedno specific labeling signal. No relevant signal was noticed ei-ther when rabbit anti-calcitonin was used as an irrelevant sub-stitute for pertinent polyclonal primary antisera (data notshown).
Fig. 3. Expression of CS31A in Escherichia coli wild-type and recombinant strains. Escherichia coli strains: wild-type strain 31A (a and b),recombinant DH10B (pEH524) (c and d), and negative control strain DH10B (e and f), as revealed by negative staining (a, c, and e) andimmunogold labeling with polyclonal anti-CS31A antibodies (b, d, and f). Scale bar = 0.5 µm.
Kuehni-Boghenbor et al. 733
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

Immunofluorescence of E. coliAs a positive reference to validate the protocol, strain
H10407 was labeled with anti-CFA/I antibodies. Double im-munofluorescence with Baclight revealed a red labeling of
virtually every single microorganism (Figs. 5a–5d). In wild-type strain E. coli 31A, immunoreactivity was nonhomogene-ous as not all bacteria were labeled (Figs. 5e–5h). As for therecombinant strain DH10B (pEH524), morphology was even
Fig. 4. Expression of CS31A in recombinant CVD 103-HgR strains. Expression of CS31A antigen in Vibrio cholerae strains: recombinantCVD 103-HgR (pEH524) (a and b) and recombinant CVD 103-HgR (pEHProSTC28) (c and d), and negative control strain CVD 103-HgR (eand f), as revealed by negative staining (a, c, and e) and immunogold labeling with polyclonal anti-CS31A antibodies (b, d, and f). Scalebar = 0.5 µm.
734 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.
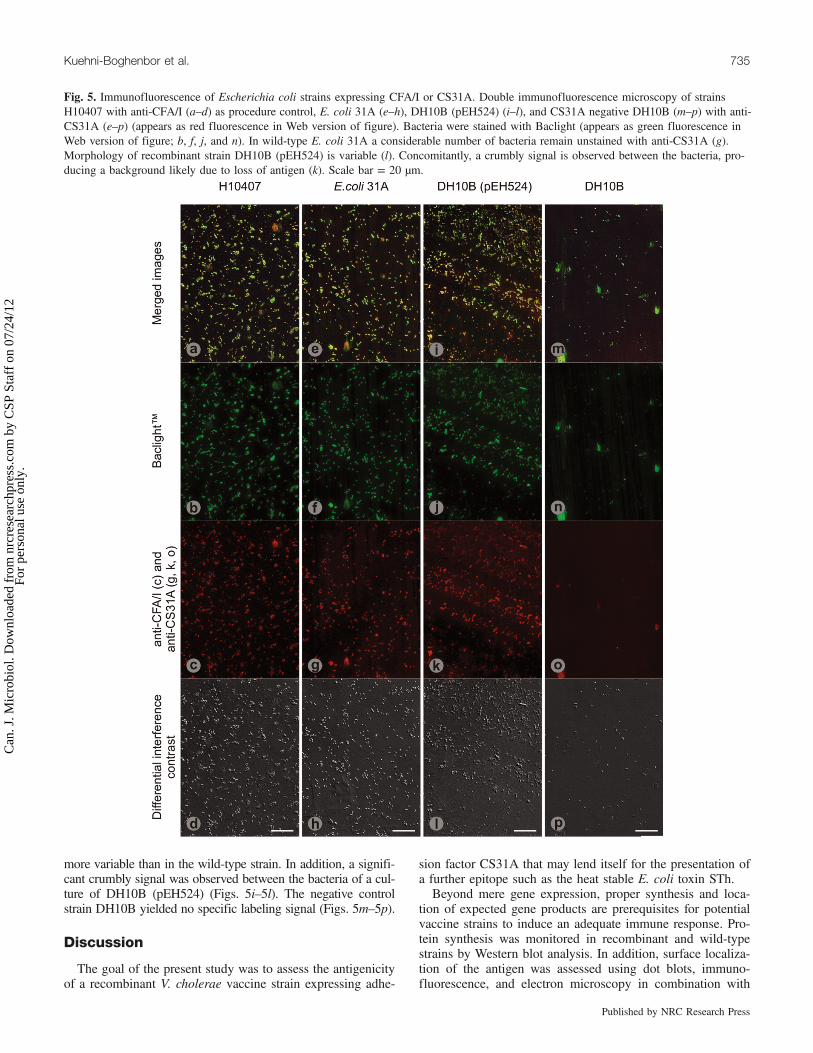
more variable than in the wild-type strain. In addition, a signifi-cant crumbly signal was observed between the bacteria of a cul-ture of DH10B (pEH524) (Figs. 5i–5l). The negative controlstrain DH10B yielded no specific labeling signal (Figs. 5m–5p).
Discussion
The goal of the present study was to assess the antigenicityof a recombinant V. cholerae vaccine strain expressing adhe-
sion factor CS31A that may lend itself for the presentation ofa further epitope such as the heat stable E. coli toxin STh.Beyond mere gene expression, proper synthesis and loca-
tion of expected gene products are prerequisites for potentialvaccine strains to induce an adequate immune response. Pro-tein synthesis was monitored in recombinant and wild-typestrains by Western blot analysis. In addition, surface localiza-tion of the antigen was assessed using dot blots, immuno-fluorescence, and electron microscopy in combination with
Fig. 5. Immunofluorescence of Escherichia coli strains expressing CFA/I or CS31A. Double immunofluorescence microscopy of strainsH10407 with anti-CFA/I (a–d) as procedure control, E. coli 31A (e–h), DH10B (pEH524) (i–l), and CS31A negative DH10B (m–p) with anti-CS31A (e–p) (appears as red fluorescence in Web version of figure). Bacteria were stained with Baclight (appears as green fluorescence inWeb version of figure; b, f, j, and n). In wild-type E. coli 31A a considerable number of bacteria remain unstained with anti-CS31A (g).Morphology of recombinant strain DH10B (pEH524) is variable (l). Concomitantly, a crumbly signal is observed between the bacteria, pro-ducing a background likely due to loss of antigen (k). Scale bar = 20 µm.
Kuehni-Boghenbor et al. 735
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

negative staining and immunogold labeling. This approachaimed at substantiating the presence of the recombinant pro-tein ClpG of CS31A fimbriae at the cell surface and at vali-dating its interaction with antibodies in view of its suitabilityas an immunogen.Western blots of the purified antigen showed expression of
CS31A, as revealed by twin bands near the expected molec-ular mass. This is most likely a result of partial processing ofthe precursor of protein ClpG. As shown by Western blotanalysis, expression of CS31A is higher in the recombinantE. coli strain than in the heterologous host V. cholerae.Although the genetic material introduced into the host strainsencompasses all known regulatory and structural compo-nents, synthesis or correct processing of coli-specific proteinsseems to be less effective in V. cholerae strains. One draw-back of antiserum MDV was its cross-reactivity with numer-ous proteins in both E. coli and V. cholerae strains, makingthe use of this serum inappropriate for microscopic immuno-labeling.We were unable to visualize distinct pili beyond the indi-
rect evidence of an electron-dense margin of phosphotungsticacid surrounding single bacteria. This is consistent with aprevious report showing the difficulty of demonstrating thepresence of CS31A fimbriae by negative staining (Girardeauet al. 1988). Regardless of this limitation, several experimen-tal lines of evidence indicated a substantial variability withrespect to the expression of CS31A by individual micro-organisms: (i) disparate retention of negative stain in wild-type strain 31A cells, (ii) heterogeneity of wild-type strain31A with respect to immunogold reactivity, and (iii) strikingvariability of daughter colonies after subcultivation from asingle strongly immunopositive parent colony. Taken to-gether, these experiments support the concept of a substantialphase variation occurring in E. coli 31A wild-type strain. In-deed, Crost et al. (2003) have described the existence ofphase-ON and phase-OFF states in cells coding for CS31A.In the phase-ON state, CS31A was expressed, whereas thiswas not the case in the phase-OFF state. This phase variationwas controlled by the presence of amino acids, particularly L-alanine and L-leucine.Irrespective of the variability in labeling intensity of single
bacteria, immunogold labeling of the recombinant E. colistrain revealed labeling of small particles being dissociatedfrom the bacteria. Similar observations were made inimmunofluorescence experiments. As opposed to the sharpdelineation of H10407 bacteria with anti-CFA/I antibodies,recombinant E. coli strain were weakly labeled with anti-CS31A serum but a diffuse background signal was presentin between the microorganisms. The conception that CS31Afimbriae seem to detach very easily from the bacteria is alsofully congruent with the dark rim in dot blots, with the label-ing of isolated particles in fluorescence and electron micros-copy and the inconsistent labeling of individual bacteria inimmunofluorescence. Taken together, these observations indi-cate that irrespective of the efficiency of protein synthesis inthe recombinant hosts, CS31A fimbriae are prone to be lostby the bacteria. However, this does not preclude that expres-sion of fimbriae might be promoted to some extent by furthermodification of growth conditions, such as by removingagents leading to a phase-OFF state. Notwithstanding, thepresence of small immunoreactive particles indicates that
CS31A fimbriae-like structures can be produced by recombi-nant gene technology. Although they were detached from thebacteria, these particles are potentially valuable antigens forimmunization.With respect to recombinant V. cholerae strains, the
present study indicates that proper gene expression at thecell surface is difficult to achieve. CS31A is synthesized viathe chaperone–usher pathway (Bertin et al. 1993), and theclpE gene is involved in the transport of the major subunitClpG across the periplasm (Bertin et al. 1993). Although theplasmid pEH524 is supposed to encompass all the genes re-quired for proper expression of CS31A, the complexity of thesynthetic machinery obviously is very susceptible to disrup-tion. This is consistent with previous findings indicating thatproper expression of E. coli-specific fimbriae in V. choleraCVD 103-HgR and its derivatives is intricate to achieve(Ziethlow et al. 2008). Expression of the recombinant proteinClpG::STh representing a fusion between the CS31A core pro-tein and the short heat stable toxin STh could be achieved inV. cholerae CVD 103-HgR (pEHProSTC28). But as for ClpGalone, the fusion protein was not exposed at the cell surface.In the present study, recombinant V. cholerae strains did
not express CS31A in adequate amounts. As for recombinantDH10B strains, protein synthesis of CS31A was even higherthan in the donor strains. However, in the wild-type and re-combinant E. coli strains being studied, the antigen was onlyloosely associated with the microorganisms. Thus, whileCS31A fimbriae antigens are readily expressed in heterolo-gous strains by means of recombinant DNA technology andmight be used for immunization, the use of fimbriae as po-tential carriers for low molecular mass haptens such as theSTh of E. coli requires further optimization.
AcknowledgementsThis study was supported by grant Nr. KTI-7304.1 LSPP-
LS of the Swiss Federal Office for Industrial Education andTechnology. The technical support of Yvonne Schlatter andSimon König is gratefully acknowledged.
ReferencesAimoto, S., Takao, T., Shimonishi, Y., Hara, S., Takeda, T., Takeda,
Y., and Miwatani, T. 1982. Amino-acid sequence of a heat-stableenterotoxin produced by human enterotoxigenic Escherichia coli.Eur. J. Biochem. 129(2): 257–263. doi:10.1111/j.1432-1033.1982.tb07047.x. PMID:6759126.
Batisson, I., and Der Vartanian, M. 2000. Contribution of definedamino acid residues to the immunogenicity of recombinantEscherichia coli heat-stable enterotoxin fusion proteins. FEMSMicrobiol. Lett. 192(2): 223–229. doi:10.1111/j.1574-6968.2000.tb09386.x. PMID:11064199.
Batisson, I., Der Vartanian, M., Gaillard-Martinie, B., and Con-trepois, M. 2000. Full capacity of recombinant Escherichia coliheat-stable enterotoxin fusion proteins for extracellular secretion,antigenicity, disulfide bond formation, and activity. Infect. Immun.68(7): 4064–4074. doi:10.1128/IAI.68.7.4064-4074.2000. PMID:10858223.
Bertin, Y., Girardeau, J.P., Der Vartanian, M., and Martin, C. 1993.The ClpE protein involved in biogenesis of the CS31A capsule-like antigen is a member of a periplasmic chaperone family ingram-negative bacteria. FEMS Microbiol. Lett. 108(1): 59–67.doi:10.1111/j.1574-6968.1993.tb06074.x. PMID:8097176.
736 Can. J. Microbiol. Vol. 58, 2012
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.

Bousquet, F., Martin, C., Girardeau, J.P., Mechin, M.C., DerVartanian, M., Laude, H., and Contrepois, M. 1994. CS31Acapsule-like antigen as an exposure vector for heterologousantigenic determinants. Infect. Immun. 62(6): 2553–2561.PMID:7514578.
Crost, C., Garrivier, A., Harel, J., and Martin, C. 2003. Leucine-responsive regulatory protein-mediated repression of clp (encod-ing CS31A) expression by L-leucine and L-alanine in Escherichiacoli. J. Bacteriol. 185(6): 1886–1894. doi:10.1128/JB.185.6.1886-1894.2003. PMID:12618452.
Evans, D.G., Evans, D.J., Jr, Clegg, S., and Pauley, J.A. 1979.Purification and characterization of the CFA/I antigen ofenterotoxigenic Escherichia coli. Infect. Immun. 25(2): 738–748.PMID:39896.
Favre, D., Lüdi, S., Stoffel, M., Frey, J., Horn, M.P., Dietrich, G., etal. 2006. Expression of enterotoxigenic Escherichia coli coloniza-tion factors in Vibrio cholerae. Vaccine, 24(20): 4354–4368.doi:10.1016/j.vaccine.2006.02.052. PMID:16581160.
Girard, M.P., Steele, D., Chaignat, C.L., and Kieny, M.P. 2006. Areview of vaccine research and development: human entericinfections. Vaccine, 24(15): 2732–2750. doi:10.1016/j.vaccine.2005.10.014. PMID:16483695.
Girardeau, J.P., Der Vartanian, M., Ollier, J.L., and Contrepois, M.1988. CS31A, a new K88-related fimbrial antigen on bovineenterotoxigenic and septicemic Escherichia coli strains. Infect.Immun. 56(8): 2180–2188. PMID:2899553.
Grant, S.G., Jessee, J., Bloom, F.R., and Hanahan, D. 1990.Differential plasmid rescue from transgenic mouse DNAs intoEscherichia coli methylation-restriction mutants. Proc. Natl. Acad.Sci. U.S.A. 87(12): 4645–4649. doi:10.1073/pnas.87.12.4645.PMID:2162051.
Gupta, S.K., Keck, J., Ram, P.K., Crump, J.A., Miller, M.A., andMintz, E.D. 2008. Part III. Analysis of data gaps pertaining toenterotoxigenic Escherichia coli infections in low and mediumhuman development index countries, 1984–2005. Epidemiol.
Infect. 136(6): 721–738. doi:10.1017/S095026880700934X.PMID:17686197.
Ketley, J.M., Michalski, J., Galen, J., Levine, M.M., and Kaper, J.B.1993. Construction of genetically marked Vibrio cholerae O1vaccine strains. FEMS Microbiol. Lett. 111(1): 15–21. doi:10.1111/j.1574-6968.1993.tb06355.x. PMID:8359676.
Köhler, G., and Milstein, C. 1975. Continuous cultures of fused cellssecreting antibody of predefined specificity. Nature, 256(5517):495–497. doi:10.1038/256495a0. PMID:1172191.
Kuhnert, P., Hacker, J., Muhldorfer, I., Burnens, A.P., Nicolet, J., andFrey, J. 1997. Detection system for Escherichia coli-specificvirulence genes: absence of virulence determinants in B and Cstrains. Appl. Environ. Microbiol. 63(2): 703–709. PMID:9023948.
Schaller, A., Kuhn, R., Kuhnert, P., Nicolet, J., Anderson, T.J.,MacInnes, J.I., et al. 1999. Characterization of apxIVA, a new RTXdeterminant of Actinobacillus pleuropneumoniae. Microbiology,145(8): 2105–2116. doi:10.1099/13500872-145-8-2105. PMID:10463177.
Tacket, C.O., and Levine, M.M. 1997. Vaccines against enterotoxi-genic Escherichia coli infections. Part II. Live oral vaccines andsubunits (Purified fimbriae and toxin subunits) vaccines. In Newgeneration vaccines. Edited by M.M. Levine, G.C. Woodrow, J.B.Kaper, and G.S. Cobon. Marcel Dekker Inc., New York. pp. 875–883.
Wolf, M.K. 1997. Occurrence, distribution, and associations of O andH serogroups, colonization factor antigens, and toxins ofenterotoxigenic Escherichia coli. Clin. Microbiol. Rev. 10(4):569–584. PMID:9336662.
Ziethlow, V., Favre, D., Viret, J.F., Frey, J., and Stoffel, M.H. 2008.Assessment by electron-microscopy of recombinant Vibriocholerae and Salmonella vaccine strains expressing enterotoxi-genic Escherichia coli-specific surface antigens. Clin. Microbiol.Infect. 14(3): 282–286. doi:10.1111/j.1469-0691.2007.01908.x.PMID:18093230.
Kuehni-Boghenbor et al. 737
Published by NRC Research Press
Can
. J. M
icro
biol
. Dow
nloa
ded
from
nrc
rese
arch
pres
s.co
m b
y C
SP S
taff
on
07/2
4/12
For
pers
onal
use
onl
y.
Copyright © 2022 FDOKUMEN




















