
Exploring composition and behaviour of fish fauna by in situ observations in the Bari Canyon...
16
ORIGINAL ARTICLE Exploring composition and behaviour of fish fauna by in situ observations in the Bari Canyon (Southern Adriatic Sea, Central Mediterranean) Gianfranco D’Onghia, Francesca Capezzuto, Angela Carluccio, Roberto Carlucci, Agnese Giove, Francesco Mastrototaro, Michele Panza, Letizia Sion, Angelo Tursi & Porzia Maiorano Department of Biology and CoNISMa LRU, University of Bari Aldo Moro, Bari,Italy Keywords Behaviour; canyon; feeding habit; fish fauna; Mediterranean. Correspondence Gianfranco D’Onghia, Department of Biology, University of Bari Aldo Moro – Via E. Orabona 4, 70125, Bari, Italy. E-mail: [email protected] Accepted: 8 February 2014 doi: 10.1111/maec.12162 Abstract Canyons play a fundamental role in enhancing the abundance and diversity of marine organisms through the transport of organic matter and food resources, the presence of complex physical habitats and the absence of trawl fishing. During four baited lander deployments carried out in the Bari Canyon (South- ern Adriatic Sea, Central Mediterranean), at depths of 443–788 m, about 43 h of video records were taken, for a total of 619,200 video frames. A total of 12 benthopelagic fish species (five chondrichthyes and seven osteichthyes) were identified. The blackspot seabream (Pagellus bogaraveo) was the most often observed fish species. The depth of 787 m represents a new depth record for this fish in the Adriatic Sea. Groups of up to 40 individuals of P. bogaraveo were attracted to the bait and were shown in single frames. The individuals were observed both exploring the area and feeding actively on the bait. The European conger (Conger conger) was recorded at each deployment. Clear scavenger behaviour was also observed in this teleost fish and in the shark Etmopterus spinax. The shark species Centrophorus granulosus and Hexanchus griseus, which are considered ‘vulnerable’ on the published IUCN Mediterra- nean Regional Red List, were also recorded but, although attracted by the bait, they were never seen feeding on it. Other fish species, harvested on fishing grounds, such as Merluccius merluccius, Helicolenus dactylopterus and Polyprion americanus, were also recorded. This study represents the first in situ documen- tation, at very low impact, of the fish fauna in the Bari Canyon, providing new insights into its small scale distribution and behaviour, the first in situ direct observation of the variable feeding behaviour of P. bogaraveo and its gregarious habits, as well as indicating that this canyon could act as a refuge area for species that are vulnerable to fishing on the open slope. Introduction Submarine canyons are complex structures capable of enhancing the heterogeneity of the slope and therefore play an important role in the functioning of the deep sea ecosystem (Danovaro et al. 2010; Wu rtz 2012). In fact, canyons act as trapping areas for sediments and provide a direct pathway for sediment transport and dense water cascading from the continental shelf to the deep basin, enhancing the transport and availability of particles and organic matter (Turchetto et al. 2007; Tesi et al. 2008; Canals et al. 2009). The effect of canyons on local circula- tion can enhance the abundance and the diversity of mar- ine organisms, through the transport of sediment and phytodetritus and the presence of complex physical habi- tats (Vetter & Dayton 1998; Ramirez-Llodra et al. 2010). Higher macro- and megafauna biomass as well as spawn- ing and recruitment sites have been detected in the Medi- terranean canyons (e.g. Company et al. 2012; Wu rtz 2012; Tecchio et al. 2013 and references therein). Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 1 Marine Ecology. ISSN 0173-9565
Transcript of Exploring composition and behaviour of fish fauna by in situ observations in the Bari Canyon...

ORIGINAL ARTICLE
Exploring composition and behaviour of fish fauna byin situ observations in the Bari Canyon (Southern AdriaticSea, Central Mediterranean)Gianfranco D’Onghia, Francesca Capezzuto, Angela Carluccio, Roberto Carlucci, Agnese Giove,Francesco Mastrototaro, Michele Panza, Letizia Sion, Angelo Tursi & Porzia Maiorano
Department of Biology and CoNISMa LRU, University of Bari Aldo Moro, Bari,Italy
Keywords
Behaviour; canyon; feeding habit; fish fauna;
Mediterranean.
Correspondence
Gianfranco D’Onghia, Department of
Biology, University of Bari Aldo Moro – Via E.
Orabona 4, 70125, Bari, Italy.
E-mail: [email protected]
Accepted: 8 February 2014
doi: 10.1111/maec.12162
Abstract
Canyons play a fundamental role in enhancing the abundance and diversity of
marine organisms through the transport of organic matter and food resources,
the presence of complex physical habitats and the absence of trawl fishing.
During four baited lander deployments carried out in the Bari Canyon (South-
ern Adriatic Sea, Central Mediterranean), at depths of 443–788 m, about 43 h
of video records were taken, for a total of 619,200 video frames. A total of 12
benthopelagic fish species (five chondrichthyes and seven osteichthyes) were
identified. The blackspot seabream (Pagellus bogaraveo) was the most often
observed fish species. The depth of 787 m represents a new depth record for
this fish in the Adriatic Sea. Groups of up to 40 individuals of P. bogaraveo
were attracted to the bait and were shown in single frames. The individuals
were observed both exploring the area and feeding actively on the bait. The
European conger (Conger conger) was recorded at each deployment. Clear
scavenger behaviour was also observed in this teleost fish and in the shark
Etmopterus spinax. The shark species Centrophorus granulosus and Hexanchus
griseus, which are considered ‘vulnerable’ on the published IUCN Mediterra-
nean Regional Red List, were also recorded but, although attracted by the bait,
they were never seen feeding on it. Other fish species, harvested on fishing
grounds, such as Merluccius merluccius, Helicolenus dactylopterus and Polyprion
americanus, were also recorded. This study represents the first in situ documen-
tation, at very low impact, of the fish fauna in the Bari Canyon, providing new
insights into its small scale distribution and behaviour, the first in situ direct
observation of the variable feeding behaviour of P. bogaraveo and its gregarious
habits, as well as indicating that this canyon could act as a refuge area for
species that are vulnerable to fishing on the open slope.
Introduction
Submarine canyons are complex structures capable of
enhancing the heterogeneity of the slope and therefore
play an important role in the functioning of the deep sea
ecosystem (Danovaro et al. 2010; Wurtz 2012). In fact,
canyons act as trapping areas for sediments and provide a
direct pathway for sediment transport and dense water
cascading from the continental shelf to the deep basin,
enhancing the transport and availability of particles and
organic matter (Turchetto et al. 2007; Tesi et al. 2008;
Canals et al. 2009). The effect of canyons on local circula-
tion can enhance the abundance and the diversity of mar-
ine organisms, through the transport of sediment and
phytodetritus and the presence of complex physical habi-
tats (Vetter & Dayton 1998; Ramirez-Llodra et al. 2010).
Higher macro- and megafauna biomass as well as spawn-
ing and recruitment sites have been detected in the Medi-
terranean canyons (e.g. Company et al. 2012; Wurtz
2012; Tecchio et al. 2013 and references therein).
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 1
Marine Ecology. ISSN 0173-9565

Moreover, canyons can be considered as biodiversity hot-
spots with high megafauna diversity and high rates of
endemism (Danovaro et al. 2010 and references therein;
Ramirez-Llodra et al. 2010 and references therein).
The enhancement of trophic resources, accelerated cur-
rents and structural heterogeneity also make these canyons
favourable habitats for suspension feeders such as cold-
water corals (CWC) (Freiwald et al. 2009; Orejas et al.
2009). CWC have also been recorded in the Bari Canyon
System (Southern Adriatic Sea, Central Mediterranean)
using a remotely operated vehicle (ROV) (Freiwald et al.
2009). This canyon has been investigated on geomorpho-
logical, oceanographic and sedimentological aspects (e.g.
Ridente et al. 2007; Trincardi et al. 2007; Turchetto et al.
2007; Tesi et al. 2008; Rubino et al. 2010; Carniel et al.
2012). Information on the distribution and abundance of
megafauna in the Bari Canyon is mainly related to benthic
sessile species (Freiwald et al. 2009; Bo et al. 2012; Sanfi-
lippo et al. 2013; Angeletti et al. 2014) while knowledge of
the fish fauna is limited to the recording of four teleost
fishes using ROV (Angeletti et al. 2014). This lack of
knowledge can be explained by the inaccessibility of the
area to traditional sampling gear and trawl nets, making
the sampling of mobile megafauna quite difficult. Longline
fishing only occurs on the margins of the Bari Canyon,
mostly during summer. The walls and sea floor of the can-
yon are very rough three-dimensional structures that form
habitats, such as hard ground, boulders and coral branches,
which can tear or entangle longlines.
Video inspections are generally less invasive for fragile
deep-sea ecosystems and can provide information on
small scale species distribution, activity and behaviour
(e.g. Bailey & Priede 2002; Uiblein et al. 2003; Costello
et al. 2005 and references therein; Ross & Quattrini 2007;
King et al. 2006, 2008 and references therein; Capezzuto
et al. 2012). However, several difficulties and limitations
should be taken into consideration during the analysis of
fish from deep sea environments. ROV sampling shows
limitations, among which are those mainly related to
noise and light created by the ROV that may either scare
away or attract fish, thus causing under- or overesti-
mations of the true abundances (Trenkel et al. 2004;
Costello et al. 2005; Lorance & Trenkel 2006; Stoner et al.
2008; Ryer et al. 2009). Such underestimations of species
that avoid the ROV and over estimates of species that are
attracted by the presence of the ROV are problems that
should be considered, in particular for species which are
rarely seen. Benthic landers produce less disturbance in
gathering similar data but are vulnerable to their own
disadvantages. Landers will only attract scavenging fish
and so are highly selective. The time required to explore
unknown deep-sea bottoms suitable for setting up and
precisely positioning the equipment, as well as the time
required at a single station between deployment and
recovery and the many assumptions made in abundance
estimates are other sampling limitations involved in the
use of landers (Priede & Merrett 1996; Roberts et al.
2005; Jamieson et al. 2006; Bailey et al. 2007; Cousins
et al. 2013).
During a survey carried out as part of the RITMARE
Italian project aimed at studying biodiversity and the rela-
tive ecosystem services in the Bari Canyon, data on the fish
fauna were collected using a baited lander equipped with
two digital cameras. In this work, the video records were
used: (i) to explore the composition of the benthopelagic
fauna in this canyon; (ii) to investigate their small-scale
distribution and activity; and (iii) to examine the behav-
iour of the individuals by species. Due to the difficulties
involved in the study of mobile bentopelagic fauna in a
habitat with complex and rugged bathymetry such as the
Bari Canyon, the work is explorative in character.
Material and Methods
Study area
The Bari Canyon System (BCS) is a complex morphologi-
cal structure that breaches the Southwestern Adriatic shelf
with a west–east trend. It is 10 km wide and 30 km long
and is between 200 and 1700 m in depth (Ridente et al.
2007; Trincardi et al. 2007). The BCS is characterized by
two main branches separated by a mounded relief, a
slope-confined trough between 500 and 700 m in depth.
The southern branch is wider (about 4 km), markedly
asymmetric and U-shaped with a very steep wall (Ridente
et al. 2007; Trincardi et al. 2007).
As part of the RITMARE project funded by the Italian
Ministry of University and Research, the MEMO (Marine
Environment MOnitoring system) lander was deployed in
the southern branch of the Bari Canyon (Southern Adri-
atic Sea, Central Mediterranean) in May–June 2012 using
the R/V MINERVA UNO. The four deployments are pre-
sented in Fig. 1 with indications of the deployment
depths.
Baited lander deployments
The MEMO lander, developed as part of the CoralFISH
EU_7FP project, consists of a stainless steel metal frame,
two digital cameras (Sony ICX414) with two LED lights
(12 V, 700 mA, 360 lumen), an electronic compass, incli-
nometer and altimeter, a multiparametric probe, a dopp-
ler current meter, four deep-sea batteries for power
supply, and an ICT infrastructure capable of managing
the entire system (Fig. 2). On the sea bed the lander is
linked by a zinc-coated steel cable to buoys which keep
2 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

the cable under tension and then to a surface floating sig-
nalling buoy. Continuous connection is maintained via
an acoustic modem with an onboard PC software plat-
form, making images and sensor data available.
A total of four deployments between 443 and 788 m
were carried out, for a total time of about 43 h of video
recordings (Table 1). As the MEMO system takes four
video frames each second, a total of 619,200 video frames
were taken by each camera. During each deployment the
lander was baited with eight specimens of Scomber scom-
brus (~700 g). The bait produces an odour plume stretch-
ing downstream from the lander and attracting animals
in view of the camera (Bailey et al. 2007). The hydro-
graphic parameters at each station were also recorded by
means of CTD and a current meter. The mean salinity
recorded on the bottom was 38.72 psu, the mean temper-
ature values varied from 13.32 to 13.38 °C, and those of
the current speed from 0.06 to 0.14 m�s�1.
Data collection and analysis
The different species were identified by their morphologi-
cal characteristics observed in the video frames. In
addition, with the exception of Pagellus bogaraveo, the
single individuals of all recorded species could often be
distinguished by their different sizes and morphological
features, such as scars on the skin. Thus, with the excep-
tion of P. bogaraveo, a total number by species was
recorded for each deployment. According to King et al.
(2006, 2008), the arrival time of the first individual (1st
arrival time) and maximum number of individuals
observed at the bait per deployment (MaxN) were
recorded. Since fish numbers gradually increased after
lander touchdown but with individuals moving in and
out of camera view, the number of fish visible at any
given time during a sequence varied. Thus, the percentage
of frames with individuals was recorded in each deploy-
ment for each species.
As significant numbers of P. bogaraveo individuals were
observed, the variations in the visual count detections
between day and night hours and current velocity were
examined following Aguzzi et al. (2003, 2006) and Doya
et al. (2014). Thus, the maximum numbers of individuals
in the field of view were recorded every 30 s (MaxN30s).
The mean of MaxN30s for each hour of deployment, with
standard deviation, was computed to obtain a mean
17°4'0'' E 17°8'0'' E 17°12'0'' E 17°16'0'' E 17°20'0'' E 17°24'0'' E 17°28'0'' E
17°4'0'' E 17°8'0'' E 17°12'0'' E 17°16'0'' E 17°20'0'' E 17°24'0'' E 17°28'0'' E
17°32'0'' E
17°32'0'' E
41°2
4'0'
' N41
°20'
0'' N
41°1
6'0'
' N41
°12'
0'' N
41°2
4'0'
' N41
°20'
0'' N
41°1
6'0'
' N41
°12'
0'' N
Fig. 1. Study area: location within the Mediterranean of the Bari Canyon and bathymetric map with indication of the MEMO lander deployments.
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 3
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations

fluctuation plot. Phase duration and timing was assessed
using the MESOR (midline estimating statistic of rhythm)
estimated by re-averaging all hourly values and represent-
ing the resulting mean as a horizontal threshold line on
the fluctuation plots (Aguzzi et al. 2003, 2006; Doya et al.
2014). The fluctuation of the hourly mean current veloc-
ity (m�s�1) during the deployment was superimposed on
the fluctuation plot of the visual fish count.
Sizes of fishes were measured from images using
IMAGE J 1.46q, a Java-based public domain program
developed at the USA National Institutes of Health, Beth-
esda, MD, USA (available at, http://rsb.info.nih.gov/ij/
index.html). The software was calibrated using the
within-image reference scale (bait plate, 25 cm diameter).
Fishes were only measured if the full length of the body
was visible at the level of the reference scale. The size was
expressed as total length (TL, cm). To avoid re-measuring
the same individual, the fishes were only measured at
peak abundance in the field of view (MaxN) in the differ-
ent deployments. However, specimens of most species
were often distinguished in relation to their different
sizes, colour and scars on the skin. For P. bogaraveo the
length–frequency distributions were also computed at the
depths where individuals with significant numbers were
detected and measured.
Position with respect to the bottom, reaction to the
lander and activity of the individual organisms were clas-
sified in terms of ‘Fauna Position’, ‘Fauna Reaction’ and
‘Fauna Activity’, respectively, as described in Uiblein
et al. (2003), Trenkel et al. (2004) and Costello et al.
(2005). In particular, Fauna Position was recorded in
three categories: water column (1–2 m above the
Fig. 2. The CoNISMa MEMO lander equipment with indication of the different components.
Table 1. Data related to the MEMO lander deployments in the Bari Canyon.
Date Station Latitude Longitude Depth (m)
Salinity
(psu) T(°C)
Mean
current (m/s)
Start
video
End
video
Time of
video record
(h, m)
29/5/2012 St. 1 41°18.40850 N 17°09, 06220 E 443 38.72 13.37 0.12 11:23 18:37 7.14
30/5/2012 St. 2 41°17.70930 N 17°17,49490 E 787 38.72 13.32 0.06 09:58 17:37 7.39
30–31/05/2012 St. 3 41°17.71270 N 17°17,71260 E 788 38.72 13.34 0.14 18:50 07:35 12.45
31/05–1/06/2012 St. 4 41°14.48760 N 17°20,34150 E 577 38.72 13.38 0.06 11:35 04:01 16.26
4 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

bottom); near the sea bed (within 0.5–1 m); and on the
sea bed. Fauna Reaction was classified in three categories:
no reaction; avoidance-disturbed; and attraction. Fauna
Activity was divided into seven categories: actively swim-
ming; quietly swimming; hovering; moving on the sea
bed; resting on the sea bed; feeding; and feeding on bait.
Results
Species composition
A total of 12 benthopelagic fish species (five chondrich-
thyes and seven osteichthyes) were identified (Table 2,
Fig. 3). The highest number of species (8) was observed
at the shallowest station (443 m). Most species were
recorded during daylight hours. Only one individual was
observed during one deployment for the species Cen-
trophorus granulosus, Galeus melastomus, Hoplostethus
mediterraneus and Polyprion americanus.
Pagellus bogaraveo was the fish most frequently
observed at the two shallowest stations: in 85.16% of the
video frames at 443 m and 51.98% at 577 m. One indi-
vidual of this fish was observed at 787 m, which is a new
depth record of this fish in the Adriatic Sea. Its total
number per deployment is not reported in the Table 2
due to the impossibility of distinguishing individuals.
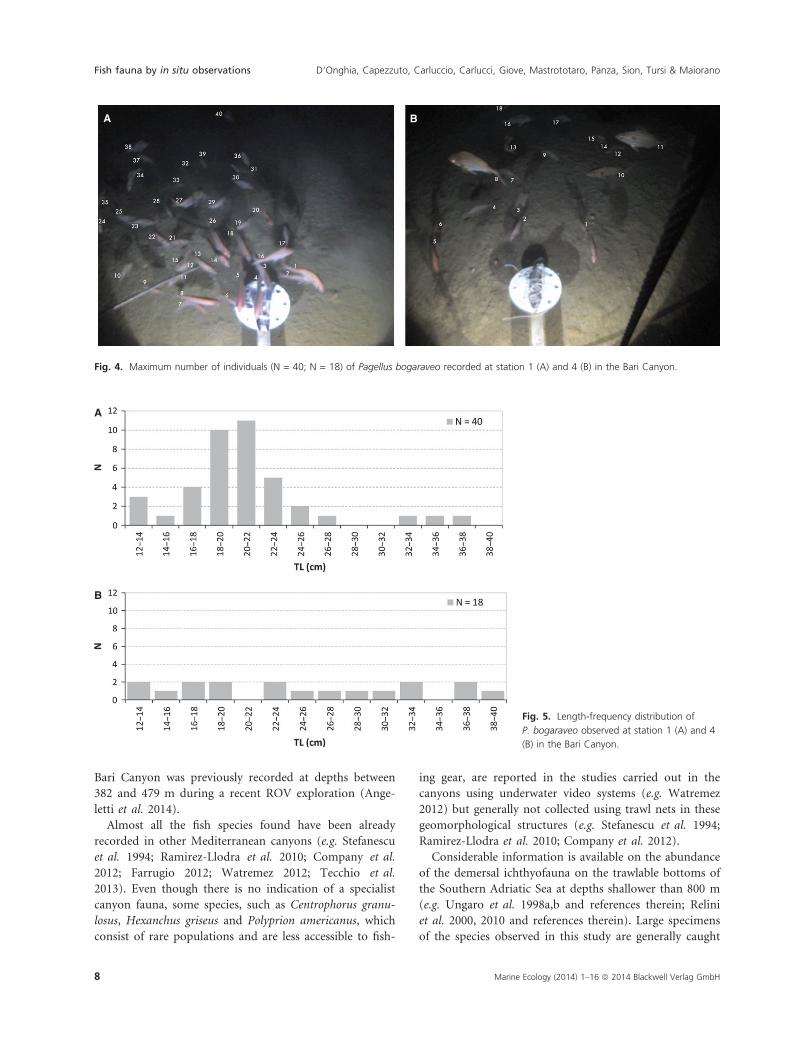
First arrival time for P. bogaraveo was 9 min at 443 m
with a maximum number of 40 individuals in the frames
and 90 min at 577 m with a maximum number of 18
individuals in the frames (Fig. 4). The individuals mea-
sured at 443 m had sizes between 12.5 and 37.5 cm TL
with a mean length of 21 � 5.1 cm TL; those measured
at 577 m ranged from 12.5 to 39 cm TL and the mean
length was 25 � 8.7 cm TL (Fig. 5).
A significant increase in the visual counts of P. bogaraveo
was detected during the afternoon at 443 m and near sun-
set at 577 m. Superimposing the current velocity on the
fluctuation in the visual fish count seems to indicate that
animal detections increase significantly when water speed
decreases at both stations (Fig. 6). Water speed at 577 m,
when day and night hours were investigated, showed a
diurnal oscillation with a higher peak at night and a smaller
one in daytime.
Other fishes frequently observed at the shallowest
deployment were the teleosts Helicolenus dactylopterus
(25.98%) and Conger conger (9.97%). Both species were
recorded during day and night hours. The latter was the
only fish recorded in all four deployments. The maxi-
mum number (7) was observed at 788 m, where it
arrived sooner (12 min) than at the other stations. The
greatest sizes were shown at the greatest depths. The
other teleosts and the cartilagineous fishes were observed
less frequently. The two shark species G. melastomus
(49 cm TL) and Etmopterus spinax (33 and 45 cm TL)
arrived at the bait in a comparable time, 28 min at
443 m and 26 min at 787 m, respectively.
Large individuals (61 and 62 cm TL) of the commer-
cial target species Merluccius merluccius were recorded at
the two shallowest stations during daytime.
Species behaviour
The list of the species recorded with indication of type of
behaviour (position, reaction and activity) is presented in
Table 3. All the species were observed swimming near the
sea bed. However, Pagellus bogaraveo was mainly
recorded swimming, both actively and quietly, in the
water column. It was often observed hovering in front of
the lander near the bait. Galeus melastomus, Conger con-
ger, Helicolenus dactylopetrus and Merluccius merluccius
were also recorded on the sea bed. None of the observed
species seemed to be clearly disturbed by the lander. Indi-
viduals of most species either showed no reaction or were
attracted to the lander. Etmopterus spinax, C. conger and
P. bogaraveo were observed investigating, attacking and
ingesting the bait (Fig. 7). Individuals of P. bogaraveo
were mostly observed in groups swimming in the water
column and near the sea bed attracted to the baited
lander. Members of the group were observed forming
irregular group shapes. The groups appeared to be rather
disorganised near the lander, with most individuals
actively swimming and investigating, others quietly swim-
ming in a circle, and others hovering. They sometimes
showed no reaction. Individuals in the groups were some-
times observed swimming together in a coordinated fash-
ion (Fig. 8). Fish were mostly observed staying on the
bait for a long time, accumulating in numbers within the
camera’s view.
The first specimens ahead of the group often attacked
the bait, removing flesh and even single fish from the bait
plate (Fig. 9). The other individuals generally continued
to attack the same fish primarily utilized on the bait
plate, exhibiting a feeding frenzy. Some individuals were
observed ingesting the bait and others also feeding on the
fast-moving plankton in the water column and near the
bottom. The fish often remained within the camera’s view
even after all the bait had been totally consumed.
Helicolenus dactylopterus was also observed feeding on
the plankton and small crustaceans moving or resting on
the sea bed. It was never observed to feed on the bait.
Lepidopus caudatus was shown feeding in the sediment.
Apart from E. spinax, the other shark species were only
observed being attracted by the bait and investigating it
but without feeding. The wreckfish P. americanus was
observed quietly swimming near the sea bed with no
reaction to the lander.
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 5
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations

Table 2. First arrival time on bait, total number of individuals observed per deployment (Total N), maximum number of individuals observed at
the bait per deployment (Max N), percentage of frames in which each species was visible, total length (TL) and presence during day or night
hours (Day-Night). (n.m. = not measured)
Species
Deployment 1 2 3 4
Depth (m) 443 787 788 577
Chondrichthyes
Centrophorus granulosus First arrival time (min) 116
Tot N 1
Max N 1
% of frames 1.08
TL (cm) 89
Day-Night D
Dalatias licha First arrival time (min) 129
Tot N 2
Max N 1
% of frames 1.44
TL (cm) 96
Day-Night D
Etmopterus spinax First arrival time (min) 26
Tot N 2
Max N 1
% of frames 0.84
TL (cm) 33–45
Day-Night D
Galeus melastomus First arrival time (min) 28
Tot N 1
Max N 1
% of frames 0.57
TL (cm) 49
Day-Night D
Hexanchus griseus First arrival time (min) 268 219
Tot N 1 1
Max N 1 1
% of frames 0.04 0.12
TL (cm) n.m. n.m.
Day-Night D D
Osteichthyes
Conger conger First arrival time (min) 44 47 12 174
Tot N 5 3 7 2
Max N 2 1 2 1
% of frames 9.97 2.74 5.98 1.4
TL (cm) 66 � 17 106 � 20 117 � 12 67–110
Day-Night D D D-N D-N
Helicolenus dactylopterus First arrival time (min) 305 57 76
Tot N 2 2 2
Max N 2 1 1
% of frames 25.98 5.47 3.20
TL (cm) 16–25 22–29 25–29
Day-Night D D D-N
Hoplostethus mediterraneus First arrival time (min) 2
Tot N 1
Max N 1
% of frames 0.40
TL (cm) 19
Day-Night D
6 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

Discussion
The present results represent the first in situ documenta-
tion on the fish fauna composition and behaviour in the
Bari Canyon. The fish fauna distributed in this canyon is
not taxonomically different from the overall regional
fauna. Apart from Pagellus bogaraveo, which was shown
at the depth record of 787 m for Adriatic Sea, the vertical
distribution observed in the species falls within the depth
range known for each of them (Ungaro et al. 1998a,b;
Relini et al. 2000, 2010; Vacchi & Serena 2010). The pres-
ence of Helicolenus dactylopterus and P. bogaraveo in the
Table 2. Continued
Species
Deployment 1 2 3 4
Depth (m) 443 787 788 577
Lepidopus caudatus First arrival time (min) 408 259
Tot N 1 2
Max N 1 1
% of frames 0.003 0.03
TL (cm) n.m. n.m.
Day-Night D D
Merluccius merluccius First arrival time (min) 107 362
Tot N 1 1
Max N 1 1
% of frames 0.66 0.41
TL (cm) 61 62
Day-Night D D
Pagellus bogaraveo First arrival time (min) 9 214 90
Tot N – – –
Max N 40 1 18
% of frames 85.16 0.11 51.98
TL (cm) 21 � 5.1 25 � 8.7
Day-Night D D D-N
Polyprion americanus First arrival time (min) 268
Tot N 1
Max N 1
% of frames 0.13
TL (cm) n.m.
Day-Night D
Fig. 3. Images of most benthopelagic fish species recorded by MEMO lander in the Bari Canyon.
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 7
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations
Bari Canyon was previously recorded at depths between
382 and 479 m during a recent ROV exploration (Ange-
letti et al. 2014).
Almost all the fish species found have been already
recorded in other Mediterranean canyons (e.g. Stefanescu
et al. 1994; Ramirez-Llodra et al. 2010; Company et al.
2012; Farrugio 2012; Watremez 2012; Tecchio et al.
2013). Even though there is no indication of a specialist
canyon fauna, some species, such as Centrophorus granu-
losus, Hexanchus griseus and Polyprion americanus, which
consist of rare populations and are less accessible to fish-
ing gear, are reported in the studies carried out in the
canyons using underwater video systems (e.g. Watremez
2012) but generally not collected using trawl nets in these
geomorphological structures (e.g. Stefanescu et al. 1994;
Ramirez-Llodra et al. 2010; Company et al. 2012).
Considerable information is available on the abundance
of the demersal ichthyofauna on the trawlable bottoms of
the Southern Adriatic Sea at depths shallower than 800 m
(e.g. Ungaro et al. 1998a,b and references therein; Relini
et al. 2000, 2010 and references therein). Large specimens
of the species observed in this study are generally caught
A B
Fig. 4. Maximum number of individuals (N = 40; N = 18) of Pagellus bogaraveo recorded at station 1 (A) and 4 (B) in the Bari Canyon.
A
B
Fig. 5. Length-frequency distribution of
P. bogaraveo observed at station 1 (A) and 4
(B) in the Bari Canyon.
8 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

during offshore bottom longline fishing at depths shal-
lower than 400 m in the Southern Adriatic, of which the
hake (Merluccius merluccius) is the primary target species,
representing 40–70% of the total catch. The secondary
target species fished with noteworthy yields are the Euro-
pean conger (Conger conger) (10–20%), rockfish (H.
dactylopterus) (5–10%), blackspot seabream (P. bogarave-
o) (2–4%) while the shark species and wreckfish (P.
americanus) are rarely caught (De Zio et al. 1998; Ungaro
et al. 2005).
From experimental trawl catches carried out on the
slope (500–800 m) in the Southern Adriatic, Ungaro
et al. (1998a) reported the following abundance indices:
0.1 N�h�1 for C. conger; 2.5 N�h�1 for H. dactylopterus;
0.9 N�h�1 for M. merluccius. The abundance of these
three teleost fishes decreased markedly at depths >400 m.
Lepidopus caudatus and P. bogaraveo were only captured
on the shelf (Ungaro et al. 1998a).
The abundance of most species collected in this study
was also evaluated during the trawl surveys carried out as
part of the MEDITS project (Bertrand et al. 2002). In the
Southern Adriatic Sea, for the period 1994–99 and at a
depth range of 200–800 m, Spedicato et al. (2002)
reported density indices between 2 and 105 N�km�2 for
P. bogaraveo. For the period 1994–2009, Relini et al.
(2010) reported density indices between 0.6 and
0.8 N�km�2 for C. granulosus, 0.8 and 2.9 N�km�2 for
Dalatias licha, 39.5 and 274.3 N�km�2 for Etmopterus
spinax and 39.8 and 371.3 N�km�2 for Galeus melastomus.
Although the blackspot seabream is not among the
most abundant fish species in the Southern Adriatic Sea,
it was the most often observed fish with the highest num-
bers in the Bari Canyon. It is an appreciated fishery
resource mostly caught using longline and gillnet on the
slope (Morato et al. 2001; Chilari et al. 2006; Mytilineou
et al. 2013). Ontogenetic changes of habitat are character-
istic of this fish, with the juveniles appearing mainly in
shallower waters on muddy bottoms and larger fishes
often associated with offshore outcrops, banks and habi-
tats less accessible to trawling (Morato et al. 2001; Mene-
zes et al. 2006; Afonso et al. 2012; Mytilineou et al.
2013). Studies carried out using towed cameras and long-
line in the Santa Maria di Leuca (SML) cold-water coral
province showed this fish to be significantly associated
with the coral habitat (D’Onghia et al. 2011, 2012). The
residency of sub-adult and adult blackspot seabream at
the Condor seamount (Azores, mid-North Atlantic) was
ascertained using acoustic telemetry (Afonso et al. 2012).
Thus, the high numbers observed in the Bari Canyon
could be due to a preferential distribution of this fish in
A
B
Fig. 6. Fluctuations of visual count
(expressed as mean Max N with standard
deviation) and mean current velocity (ms�1
with standard deviation) at 443 m (A) and
577 m (B) in the Bari Canyon. Black bar:
night duration; horizontal line: MESOR of
visual count (9.8 in A and 3.2 in B).
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 9
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations

a complex and heterogeneous habitat. Although the
lander was deployed on a sedimentary bioturbated sea
bed, P. bogaraveo is a very mobile fish species (Afonso
et al. 2012; Mytilineou et al. 2013) and at the depths and
areas where it was detected in the Bari Canyon, the exis-
tence of a megabenthic community dominated by a vari-
ety of cnidarians as major habitat-forming taxa, often in
association with sponges and serpulids has been revealed
using ROVs (Freiwald et al. 2009; Bo et al. 2012; Sanfi-
lippo et al. 2013; Angeletti et al. 2014).
The gregarious behavioural pattern known for blackspot
seabream is derived almost exclusively from fishery data
(Relini et al. 1999; Fabi et al. 2002, 2004; Chilari et al.
2006). Although specific patterns of shoaling and/or
schooling behaviour in the blackspot seabream have not
been investigated in detail here, at the depths where this
fish was observed, groups of individuals aggregate near the
bait, spending extensive amounts of time exploring the site
and remaining together. Schooling is considered an effi-
cient way of swimming which benefits the individual par-
ticipants in defeating and confounding predators, in faster
food localisation and more effective feeding as well as in
hydrodynamic advantages and in increasing reproductive
success (Pitcher 1993; Fr�eon & Misund 1999). The rhythms
in feeding activity in fish have often been explained under
various environmental conditions such as light level and
current speed (e.g. Lokkeborg et al. 2000 and references
therein; Doya et al. 2014 and references therein). At the
station where day–night variations in the visual counts
were investigated in the Bari Canyon, that is, at 577 m,
most fish were detected during the crepuscular period. This
is in agreement with Afonso et al. (2012) who detected
most fish during crepuscular and nocturnal periods over
the Condor seamount summits and reported that local
fishermen prefer to fish during these periods, avoiding day-
time fishing when fish are at deeper levels and less accessi-
ble. Further investigations are required to understand the
spatio-temporal variations in activity patterns of P. bogara-
veo in the Bari Canyon, as an increase in the visual counts
was also detected during afternoon at 443 m and when
water speed decreases at both stations. However, groups of
individuals of P. bogaraveo were observed to be attracted to
the bait, investigating, attacking and ingesting it both dur-
ing day and night hours. In the sharpsnout seabream (Dipl-
odus puntazzo), a fish species of the same Sparidae family,
both diurnal and nocturnal locomotor activities were
detected but feeding behaviour remained strictly diurnal
(Vera et al. 2006).
Although P. bogaraveo feeds both near the bottom on
benthic prey and in the water column on pelagic species
(Morato et al. 2001; Stergiou & Karpouzi 2002; Mytili-
neou et al. 2013), it can behave as a predator or scaven-
ger according its opportunistic encounters in theTable
3.Po
sition,reactionan
dactivity
byspeciesobserved
byMEM
Olander
intheBariCan
yon.
Species
Position
Reaction
Activity
Water
column
Near
seab
ed
On
seab
ed
No
reaction
Avoidan
ce-disturbed
Attraction
Actively
swim
ming
Quietly
swim
ming
Hovering
Moving
onseab
ed
Resting
onseab
edFeed
ing
Feed
ingbait
Chondrich
thyes
Cen
trophorusgranulosus
XX
X
Dalatiaslicha
XX
XX
Etmopterusspinax
XX
XX
Galeu
smelastomus
XX
XX
XX
X
Hexan
chusgriseus
XX
XX
Osteichthyes
Conger
conger
XX
XX
XX
X
Helicolenusdactylopterus
XX
XX
XX
XX
X
Hoplostethusmed
iterraneu
sX
XX
Lepidopuscaudatus
XX
XX
Merlucciusmerluccius
XX
XX
XX
Pagellusbogaraveo
XX
XX
XX
XX
X
Polyprionam
erican
us
XX
X
10 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

environment. The present study is the first in situ direct
observation of the variable feeding behaviour of P. bog-
araveo as well as of its gregarious habits.
Scavenger activity has also been observed in C. conger
which is considered a large opportunistic predator living
and foraging close to rocky areas, where it finds refuge
A B
Fig. 8. Individuals of P. bogaraveo actively swimming near the lander in a circle (A) and in a coordinated fashion (B).
Fig. 9. Individuals of P. bogaraveo feeding on bait.
A B C
Fig. 7. Individuals of Etmopetrus spinax (A), Conger conger (B) and Pagellus bogaraveo (C) feeding on bait.
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 11
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations

during the day (Morato et al. 1999; Xavier et al. 2010).
Conger conger was collected in the Santa Maria di Leuca
coral area using fishing gears (D’Onghia et al. 2010) and
towed cameras, these latter revealing a swimming behav-
iour near the sea bed (D’Onghia et al. 2011). Using the
MEMO lander, the European conger was observed roam-
ing on the bottom and scavenging on the bait in the
SML cwc province (unpublished data). It could be
another species which would have a preferential distribu-
tion in structurally complex habitats like those built by
deep-sea corals. In this respect, Sulak et al. 2007 reported
Conger oceanicus burrowing into the base of Lophelia
bushes.
The teleost H. dactylopterus exhibited a clear behaviour-
al pattern of resting on the sea bed. This fish uses a wide
range of habitats, among which are cwc habitats and sub-
marine canyons, and is closely associated with the bottom
(e.g. Bourcier & Zibrowius 1973; Uiblein et al. 2003; Cos-
tello et al. 2005; Ross & Quattrini 2007; D’Onghia et al.
2010, 2011). Helicolenus dactylopterus seems to be a typical
sit-and-wait ambush predator feeding mainly on benthic
crustaceans and fish as well as on plankton organisms (Mi-
anzan et al. 1996; Nouar & Maurin 2000; Uiblein et al.
2003; Consoli et al. 2010). As it can also be a potential
prey of larger deep-sea predators, the observed behaviour
associated to its orange-red colour, undetectable to preda-
tors, can be considered a trade off between feeding strategy
and predator avoidance. Although this fish is frequently
caught using longlines, the observed behaviour and the
fact that it was never attracted by the bait do not make it a
typical scavenger fish.
The lower presence observed in the other common
species, such as M. merluccius, also deserves consideration
related to their distribution and behaviour. Even though
M. merluccius is among the most abundant demersal spe-
cies in the Southern Adriatic Sea (e.g. Marano et al. 1998;
Carlucci et al. 2009) the investigated sites in this study
are deeper than those where the species is mainly distrib-
uted. In fact, the greatest abundance of this fish is gener-
ally recorded on the shelf and shelf edge (e.g. Marano
et al. 1998; Carlucci et al. 2009). Most probably, M. mer-
luccius was only recorded during daytime because it
migrates up the water column during the night hours in
pursuit of pelagic prey (Carpentieri et al. 2005a,b),
Lepidopus caudatus individuals were also observed dur-
ing daylight hours feeding in the sediment, confirming
observations made in the Santa Maria di Leuca cwc prov-
ince (D’Onghia et al. 2011).
Adult individuals of P. americanus are usually solitary
swimmers and seem to have a preferential distribution
on larger lithoherms and hardgrounds (Messing et al.
1990) as well as in caves and on shipwrecks (Bianchi
et al. 1999). Using a longline and a baited lander,
respectively, the wreckfish was collected and observed in
the complex habitat of the Santa Maria di Leuca cwc
province (D’Onghia et al. 2012; Maiorano et al. in
press).
The deep-water velvet shark E. spinax is an active pred-
ator feeding on a wide range of benthic and free swim-
ming organisms, mostly represented by cephalopods
(Wurtz & Vacchi 1978; Carrass�on et al. 1992). E. spinax
was observed to investigate and feed on the bait in agree-
ment with observations by Gilat & Gelman (1984). These
authors reported that the diurnal distribution of E. spinax
was similar during the day and at night. During observa-
tions carried out in the SML cwc province using the same
baited lander, the bait was not consumed by the attracted
individuals of E. spinax (Sion et al. in press).
The shark G. melastomus has a diet based on epiben-
thic (mostly Calocaris macandreae) and benthopelagic
species (euphasids, ommastrephidae, Pasiphaea multiden-
tata, mesopelagic fishes) (Carrass�on et al. 1992). In the
Bay of Biscay G. melastomus was observed displaying a
high rate of forward locomotion close to the bottom but
no scavenger behaviour was observed (Uiblein et al.
2003).
With regard to C. granulosus, D. licha and H. griseus,
the present observations together with those carried out
in the Eastern and Central Mediterranean (Gilat & Gel-
man 1984; Jones et al. 2003; Sion et al. in press), indi-
cate that the dominant behaviour of deep-sea sharks
seems to be investigation at the foot of the lander with-
out ingesting the bait, suggesting that it is unlikely that
scavenging activity is an important feeding pattern for
these deep-sea sharks. Gilat & Gelman (1984) reported
that at 510 m C. granulosus was observed from noon to
late evening and was probably unaffected by fluctuations
in light. The small numbers of sharks observed in the
Bari Canyon could be related to their rare populations,
set at the top of the marine food web. However, the spo-
radic catches carried out in the Southern Adriatic Sea
have been suggested to be due to the intense fishing
pressure exercised on these k-selected deep-sea species
(Ungaro et al. 1996).
The presence of rare top predators, such as C. granulo-
sus and H. griseus as well as M. merluccius, which are
considered ‘vulnerable’ on the published IUCN Mediter-
ranean Regional Red List (Abdul Malak et al. 2011), and
large individuals of commercial species, such as P.
bogaraveo and M. merluccius, would suggest that this can-
yon, as observed in other marine regions (e.g. Yoklavich
et al. 2000; Farrugio 2012; Tecchio et al. 2013), acts as a
refuge area for several species impacted by fishing. How-
ever, all the above aspects require further investigations
to obtain additional information on fish distribution,
abundance and behaviour.
12 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

Acknowledgements
This study was carried out as part of the Italian RIT-
MARE project funded by the Italian Ministry of Univer-
sity and Research – MIUR. The authors acknowledge the
EU_7FP CoCoNet project. Thanks to Federica Foglini
and Franco Curci for the production of Fig. 1. Thanks
also to Daniela Potenza for the production of the figures
from video frames.
References
Abdul Malak D., Livingstone S.R., Pollard D., Polidoro B.A.,
Cuttelod A., Bariche M., Bilecenoglu M., Carpenter K.E.,
Collette B.B., Francour P., Goren M., Hichem Kara M.,
Massut�ı E., Papaconstantinou C., Tunesi L. (2011) Overview
of the Conservation Status of the Marine Fishes of the
Mediterranean Sea. IUCN, Gland, Switzerland and Malaga,
Spain: vii + 61.
Afonso P., Grac�a G., Berke G., Fontes J. (2012) First
observations on seamount habitat use of blackspot
seabream (Pagellus bogaraveo) using acoustic telemetry.
Journal of Experimental Marine Biology and Ecology, 436–
437, 1–10.
Aguzzi J., Company J.B., Sard�a F., Abello P. (2003) Circadian
oxygen consumption patterns in continental slope Nephrops
norvegicus (Decapoda: Nephropidae) in the Western
Mediterranean. Journal of Crustacean Biology, 23, 749–757.
Aguzzi J., Bullock N.M., Tosini G. (2006) Spontaneous
internal desynchronization of locomotor activity and body
temperature rhythms from plasma melatonin rhythm in rats
exposed to constant dim light. Journal of Circadian
Rhythms, 4, 1–6.
Angeletti L., Taviani M., Canese S., Foglini F., Mastrototaro F.,
Argnani A., Trincardi F., Bakran-Petricioli T., Ceregato A.,
Chimienti G., Macic V., Poliseno A. (2014) New deep-water
cnidarian sites in the southern Adriatic Sea. Mediterranean
Marine Science, 15/2, 1–11.
Bailey D.M., Priede I.G. (2002) Predicting fish behaviour in
response to abyssal food falls. Marine Biology, 141, 831–840.
Bailey D.M., King N.J., Priede I.G. (2007) Cameras and
carcasses: historical and current methods for using artificial
food falls to study deep-water animals. Marine Ecology
Progress Series, 350, 179–191.
Bertrand J.A., Gil de Sola L., Papaconstantinou C., Relini G.,
Souplet A. (2002) The general specifications of the MEDITS
surveys. Scientia Marina, 66, 9–17.
Bianchi G., Carpenter K.E., Roux J.-P., Molloy F.J., Boyer D.,
Boyer H.J. (1999) FAO Species Identification Field Guide for
Fishery Purposes. The Living Marine Resources of Namibia.
FAO, Rome: 250.
Bo M., Bertolino M., Bavestrello G., Canese S., Giusti M.,
Angiolillo M., Pansini M., Taviani M. (2012) Role of deep
sponge grounds in the Mediterranean Sea: a case study in
southern Italy. Hydrobiologia, 687, 163–177.
Bourcier M., Zibrowius H. (1973) Les «boues rouges»d�evers�ees dans le canyon de la Cassidaigne (r�egion de
Marseille). Observations en soucoupe plongeante SP 350
(juin 1971) et r�esultats de dragages. Tethys, 4, 811–842.
Canals M., Danovaro R., Heussner S., Lykousis V., Puig P.,
Trincardi F., Calafat A.M., Durrieu de Madron X., Palanques
A., S�anchez-Vidal A. (2009) Cascades in Mediterranean
marine grand canyon. Oceanography, 22, 26–43.
Capezzuto F., Maiorano P., Panza M., Indennidate A., Sion L.,
D’Onghia G. (2012) Occurrence and behaviour of Paromola
cuvieri (Crustacea, Decapoda) in the Santa Maria di Leuca
cold-water coral community (Mediterranean Sea). Deep-Sea
Research I, 59, 1–7.
Carlucci R., Lembo G., Maiorano P., Capezzuto F., Marano
C.A., Sion L., Spedicato M.T., Ungaro N., Tursi A.,
D’Onghia G. (2009) Nursery areas of red mullet (Mullus
barbatus), hake (Merluccius merluccius) and deep-water rose
shrimp (Parapenaeus longirostris) in Eastern-Central
Mediterranean Sea. Estuarine, Coastal and Shelf Science, 83,
529–538.
Carniel S., Bergamasco A., Book J.W., Hobbs R.W., Sclavo M.,
Wood T. (2012) Tracking bottom waters in the Southern
Adriatic Sea applying seismic oceanography techniques.
Continental Shelf Research, 44, 30–38.
Carpentieri P., Colloca F., Ardizzone G.D. (2005a) Day-night
variations in the demersal nekton assemblage on the
Mediterranean shelf-break. Estuarine, Coastal and Shelf
Science, 63, 577–588.
Carpentieri P., Colloca F., Cardinale M., Belluscio A.,
Ardizzone G.D. (2005b) Feeding habits of European hake
(Merluccius merluccius) in the central Mediterranean Sea.
Fishery Bulletin, 103, 411–416.
Carrass�on M., Stefanescu C., Cartes J.E. (1992) Diets and
bathymetric distributions of two bathyal sharks of the
Catalan deep sea (western Mediterranean). Marine Ecology
Progress Series, 82, 21–30.
Chilari A., Petrakis G., Tsamis E. (2006) Aspects of the biology
of blackspot seabream (Pagellus bogaraveo) in the Ionian
Sea, Greece. Fishery Research, 77, 84–91.
Company J.B., Ramirez-Llodra E., Sard�a F., Aguzzi J., Puig P.,
Canals M., Calafat A., Palanques A., Sol�e M., Sanchez-Vidal
A., Mart�ın J., Lastras G., Tecchio S., Koenig S.,
Fernandez-Arcaya U., Mech�o A., Fern�andez P. (2012)
Submarine canyons in the Catalan Sea (NW
Mediterranean): megafaunal biodiversity patterns and
anthropogenic threats. In: Wurtz M. (Ed.), Mediterranean
Submarine Canyons: Ecology and Governance. IUCN, Gland,
Switzerland and M�alaga, Spain: 133–144.
Consoli P., Battaglia P., Castriota L., Esposito V., Romeo T.,
Andaloro F. (2010) Age, growth and feeding habits of the
bluemouth rockfish, Helicolenus dactylopterus dactylopterus
(Delaroche 1809) in the central Mediterranean (southern
Tyrrhenian Sea). Journal of Applied Ichthyology, 26, 583–591.
Costello M.J., McCrea M., Freiwald A., Lund€alv T., Jonsson L.,
Bett B.J., van Weering T.C.E., de Haas H., Roberts J.M.,
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 13
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations

Allen D. (2005) Role of cold-water Lophelia pertusa coral
reefs as fish habitat in the NE Atlantic. In: Freiwald A.,
Roberts J.M. (Eds), Cold-Water Corals and Ecosystems.
Springer-Verlag, Berlin: 771–805.
Cousins N.J., Linley T.D., Jamieson A.J., Bagley P.M., Blades
H., Box T., Chambers R., Ford A., Shields M.A., Priede I.G.
(2013) Bathyal demersal fishes of Charlie-Gibbs Fracture
Zone region (49-54°N) of the Mid-Atlantic Ridge: II. Baited
camera lander observations. Deep-Sea Research II, 98, 397–
406.
Danovaro R., Company J.B., Corinaldesi C., D’Onghia G.,
Galil B., Gambi C., Gooday A.J., Lampadariou N., Luna
G.M., Morigi C., Olu K., Polymenakou P., Ramirez-Llodra
E., Sabbatini A., Sard�a F., Sibuet M., Tselepides A. (2010)
Deep-Sea biodiversity in the Mediterranean Sea: the known,
the unknown, and the unknowable. PLoS ONE, 5, e11832.
De Zio V., Ungaro N., Vlora A., Strippoli G. (1998) Lo stock
di nasello del basso Adriatico: struttura demografica e
rendimenti di pesca della frazione catturata con palangaro
di fondale. Biologia Marina Mediterranea, 5, 128–135.
D’Onghia G., Maiorano P., Sion L., Giove A., Capezzuto F.,
Carlucci R., Tursi A. (2010) Effects of deep-water coral
banks on the abundance and size structure of the
megafauna in the Mediterranean Sea. Deep-Sea Research II,
57, 397–411.
D’Onghia G., Indennidate A., Giove A., Savini A., Capezzuto
F., Sion L., Vertino A., Maiorano P. (2011) Distribution
and behaviour of the deep-sea benthopelagic fauna
observed using towed cameras in the Santa Maria di Leuca
cold water coral province. Marine Ecology Progress Series,
443, 95–110.
D’Onghia G., Maiorano P., Carlucci R., Capezzuto F.,
Carluccio A., Tursi A., Sion L. (2012) Comparing deep-sea
fish fauna between coral and non-coral ‘megahabitats’ in the
Santa Maria di Leuca cold-water coral province
(Mediterranean Sea). PLoS ONE, 7, e44509.
Doya C., Aguzzi J., Pardo M., Matabos M., Company J.B.,
Costa C., Mihaly S., Canals M. (2014) Diel behavioral
rhythms in sablefish (Anoplopoma fimbria) and other
benthic species, as recorded by the deep-sea cabled
observatories in Barkley canyon (NEPTUNE-Canada).
Journal of Marine Systems, 130, 69–78.
Fabi G., Grati F., Lucchetti A., Trovarelli L. (2002) Evolution
of the fish assemblage around a gas platforms in the
northern Adriatic Sea. ICES Journal of Marine Science, 59,
309–315.
Fabi G., Grati F., Puletti M., Scarcella G. (2004) Effects on fish
community induced by installation of two gas platforms in
the Adriatic Sea. Marine Ecology Progress Series, 273, 187–
197.
Farrugio H. (2012) A refugium for the spawners of exploited
Mediterranean marine species: the canyons of the
continental slope of the Gulf of Lion. In: Wurtz M. (Ed.),
Mediterranean Submarine Canyons: Ecology and Governance.
IUCN, Gland, Switzerland and M�alaga, Spain: 45–49.
Freiwald A., Beuck L., Ruggeberg A., Taviani M., Hebbelln D.
(2009) The white coral community in the Central
Mediterranean Sea revealed by ROV surveys. Oceanography,
22, 36–52.
Fr�eon P., Misund O.A. (1999) Dynamics of Pelagic Fish
Distribution and Behaviour: Effects on Fisheries and Stock
Assessment. Fishing News Books, Blackwell Science Ltd,
Oxford: 348.
Gilat E., Gelman A. (1984) On the sharks and fishes observed
using underwater photography during a deep-water cruise
in the Eastern Mediterranean. Fisheries Research, 2, 257–271.
Jamieson A.J., Bailey D.M., Wagner H.-J., Bagley P.M., Priede
I.G. (2006) Behavioural responses to structures on the
seafloor by the deep-sea fish Coryphaenoides armatus:
implications for the use of baited landers. Deep-Sea
Research, 53, 1157–1166.
Jones E.G., Tselepides A., Bagley P.M., Collins M.A., Priede
I.G. (2003) Bathymetric distribution of some benthic and
benthopelagic species attracted to baited cameras and traps
in the deep eastern Mediterranean. Marine Ecology Progress
Series, 251, 75–86.
King N.J., Bagley P.M., Priede I.G. (2006) Depth zonation and
latitudinal distribution of deep-sea scavenging demersal
fishes of the Mid-Atlantic Ridge, 42 to 53 N°. Marine
Ecology Progress Series, 319, 263–274.
King N.J., Jamieson A.J., Bagley P.M., Priede I.G. (2008)
Deep-sea scavenging demersal fish fauna of the Nazar�e
Canyon system, Iberian Coast, northeast Atlantic Ocean.
Journal of Fish Biology, 72, 1804–1814.
Lokkeborg S., Skajaa K., Ferno A. (2000) Food-search strategy
in ling (Molva molva L.): crepuscular activity and use of
space. Journal of Experimental Marine Biology and Ecology,
247, 195–208.
Lorance P., Trenkel V.M. (2006) Variability in natural
behaviour, and observed reactions to an ROV, by mid-slope
fish species. Journal of Experimental Marine Biology and
Ecology, 332, 106–119.
Maiorano P., Sion L., Capezzuto F., Carlucci R., Mastrototaro
F., Panza M., Tursi A., D’Onghia G. (in press) Exploring
deep-sea benthopelagic fauna using a baited lander in the
Santa Maria di Leuca cold-water coral province. 40th
CIESM Congress, 2013.
Marano G., Ungaro N., Marano C.A., Marsan R. (1998) La
ricerca sulle risorse demersali del bacino Adriatico
sud-occidentale (anni 1985–97): sintesi dei risultati. Biologia
Marina Mediterranea, 5, 109–119.
Menezes G.M., Sigler M.F., Silva H.M., Pinho M.R. (2006)
Structure and zonation of demersal fish assemblages off the
Azores Archipelago (mid-Atlantic). Marine Ecology Progress
Series, 324, 241–260.
Messing C., Neumann A., Lang J. (1990) Biozonation of
deep-water lithoherms and associated hardgrounds in the
North-eastern Straits of Florida. Palaios, 5, 15–33.
Mianzan H.W., Mari N., Prenski B., Sanchez F. (1996) Fish
predation on neritic ctenophores from the Argentine
14 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano

continental shelf: a neglected food resource? Fisheries
Research, 27, 69–79.
Morato T., Sola E., Gros M.P., Menezes G. (1999) Diets of
forkbeard (Phycis phycis) and conger eel (Conger conger) off
the Azores during spring of 1996 and 1997, Arquip�elago.
Life Marine Science, 17A, 51–64.
Morato T., Sol�a Gr�os M.P., Menezes G. (2001) Feeding habits
of two congener species of seabreams, Pagellus bogaraveo
and Pagellus acarne, during spring of 1996 and 1997.
Bulletin of Marine Sciences, 69, 1073–1087.
Mytilineou Ch., Tsagarakis K., Bekas P., Anastasopoulou A.,
Kavadas S., Machias A., Haralabous J., Smith C.J., Petrakis
G., Dokos J., Kapandagakis A. (2013) Spatial distribution
and life-history aspects of blackspot seabream Pagellus
bogaraveo (Osteichthyes: Sparidae). Journal of Fish Biology,
83, 1551–1575.
Nouar A., Maurin C. (2000) Feeding habits of Helicolenus
dactylopterus dactylopterus (Pisces: Scorpaenidae) of the
Algerian coast. Cahiers de Biologie Marine, 41, 313–320.
Orejas C., Gori A., Lo Iacono C., Puig P., Gili J.-M., Dale
M.R.T. (2009) Cold-water corals in the Cap de Creus
canyon, northwestern Mediterranean: spatial distribution,
density and anthropogenic impact. Marine Ecology Progress
Series, 397, 37–51.
Pitcher T.J. (1993). Behaviour of Teleost Fishes. Chapman and
Hall, London: 735.
Priede I.G., Merrett N.R. (1996) Estimation of abundance of
abyssal demersal fishes; a comparison of data from trawls and
baited cameras. Journal of Fish Biology, 49(suppl A), 207–216.
Ramirez-Llodra E., Company J.B., Sard�a F., Rotllant G. (2010)
Megabenthic diversity patterns and community structure of
the Blanes submarine canyon and adjacent slope in the
Northwestern Mediterranean: a human overprint? Marine
Ecology, 31, 167–182.
Relini G., Bertrand J., Zamboni A. (1999) Synthesis of the
knowledge on bottom fishery resources in Central
Mediterranean, Italy and Corsica. Biologia Marina
Mediterranea, 6, 868–pp.
Relini G., Biagi F., Serena F., Belluscio A., Spedicato M.T.,
Rinelli P., Follesa M.C., Piccinetti C., Ungaro N., Sion L.,
Levi D. (2000) I selaci pescati con lo strascico nei mari
italiani. Biologia Marina Mediterranea, 7, 347–384.
Relini G., Mannini A., De Ranieri S., Bitetto I., Follesa M.C.,
Gancitano V., Manfredi C., Casciaro L., Sion L. (2010)
Chondrichthyes caught during the Medits surveys in the
Italian waters. Biologia Marina Mediterranea, 17, 186–204.
Ridente D., Minisini D., Trincardi F., Verdicchio G. (2007)
The onset of the Bari Canyon system on the South-Western
Adriatic Margin, Central Mediterranean. Marine Geology,
246, 193–207.
Roberts J.M., Peppe O.C., Dodds L.A., Mercer D.J., Thomson
W.T., Gage J.D., Meldrum D.T. (2005) Monitoring
environmental variability around cold-water coral reefs: the
use of a benthic photolander and the potential of seafloor
observatories. In: Freiwald A., Roberts J.M. (Eds), Cold-Water
Corals and Ecosystems. Springer-Verlag, Berlin: 483–502.
Ross S.W., Quattrini A.M. (2007) The fish fauna associated
with deep coral banks off the southeastern United States.
Deep-Sea Research I, 54, 975–1007.
Rubino A., Romanenkov D., Zanchettin D., Cardin V.,
Hainbucher D., Bensi M., Boldrin A., Langone L.,
Miserocchi S., Turchetto M. (2010) On the descent of dense
water on a complex canyon system in the southern Adriatic
basin. Continental Shelf Research, 44, 20–29.
Ryer C.H., Stoner A.W., Iseri P.J., Spencer M.L. (2009) Effects
of simulated underwater vehicle lighting on fish behaviour.
Marine Ecology Progress Series, 391, 97–106.
Sanfilippo R., Vertino A., Rosso A., Beuck L., Freiwald A.,
Taviani M. (2013) Serpula aggregates and their role in
deep-sea coral communities in the southern Adriatic Sea.
Facies, 59, 663–677.
Sion L., Capezzuto F., Carluccio A., Indennidate A., Maiorano
P., D’Onghia G. (in press) Behaviour of deep-sea sharks
observed in the Santa Maria di Leuca cold-water coral
province. 40th CIESM Congress, 2013.
Spedicato M.T., Greco S., Sophronidis K., Lembo G.,
Giordano D., Argyri A. (2002) Geographical distribution,
abundance and some population characteristics of the
species of the genus Pagellus (Osteichthyes: Perciformes) in
different areas of the Mediterranean. Scientia Marina, 66,
65–82.
StefanescuC.,Morales-NinB.,MassutiE. (1994)Fishassemblages
ontheslope in theCatalanSea (WesternMediterranean)–
influenceofa submarinecanyon. Journal of theMarineBiological
Associationof theUnitedKingdom,74, 499–512.
Stergiou K.I., Karpouzi V.S. (2002) Feeding habits and trophic
levels of Mediterranean fish. Reviews in Fish Biology and
Fisheries, 11, 217–254.
Stoner A.W., Ryer C.H., Parker S.J., Auster P.J., Wakefield
W.W. (2008) Evaluating the role of fish behavior in surveys
conducted with underwater vehicles. Canadian Journal of
Fisheries and Aquatic Science, 65, 1230–1243.
Sulak K.J., Brooks R.A., Luke K.E., Norem A.D., Randall M.T.,
Quaid A.J., Yeargin G.E., Miller J.M., Harden W.M., Caruso
J.H., Ross S.W. (2007) Demersal fishes associated with
Lophelia pertusa coral and hard-substrate biotopes on the
Continental Slope, Northern Gulf of Mexico. In: George
R.Y., Cairns S.D. (Eds), Conservation and Adaptive
Management of Seamount and Deep-Sea Coral Ecosystems.
University of Miami, Miami: 65–92.
Tecchio S., Ramirez-Llodra E., Aguzzi J., Sanchez-Vidal A.,
Flexas M.M., Sard�a F., Company J.B. (2013) Seasonal
fluctuations of deep megabenthos: finding evidence of
standing stock accumulation in a flux-rich continental
slope. Progress in Oceanography, 118, 188–198.
Tesi T., Langone L., Goni M.A., Turchetto M., Miserocchi S.,
Boldrin A. (2008) Source and composition of organic
matter in the Bari canyon (Italy): dense water cascading
Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH 15
D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano Fish fauna by in situ observations

versus particulate export from the upper ocean. Deep-Sea
Research I, 55, 813–831.
Trenkel V.M., Lorance P., Mah�evas S. (2004) Do visual transects
provide true population density estimates for deepwater fish?
ICES Journal of Marine Science, 61, 1050–1056.
Trincardi F., Foglini F., Verdicchio G., Asioli A., Corregiari A.,
Minisini D., Piva A., Remia A., Ridente D., Taviani M.
(2007) The impact of cascading currents on the Bari
Canyon System, SW-Adriatic Margin (Central
Mediterranean). Marine Geology, 246, 208–230.
Turchetto M., Boldrin A., Langone L., Miserocchi S., Tesi T.,
Foglini F. (2007) Particle transport in the Bari Canyon
(southern Adriatic Sea). Marine Geology, 246, 231–247.
Uiblein F., Lorance P., Latrouite D. (2003) Behaviour and
habitat utilization of seven demersal fish species on the Bay
of Biscay continental slope, NE Atlantic. Marine Ecology
Progress Series, 257, 223–232.
Ungaro N., Marano G., Rizzi E., Marzano M.C. (1996)
Demersal squaliformes and rajiformes in the South-Western
Adriatic Sea: trawl surveys 1985-1994. FAO Fisheries Report
N, 533(Suppl.), 87–92.
Ungaro N., Marano G., Marsan R. (1998a) Demersal fish
assemblage biodiversity as an index of fishery resources
exploitation. Italian Journal of Zoology, 65, 511–516.
Ungaro N., Marano G., Vlora A., Martino M. (1998b)
Space-time variations of demersal fish assemblages in the
south-western Adriatic Sea. Vie Milieu, 48, 191–201.
Ungaro N., Marano G., Pastorelli A.M., Rositani L. (2005)
Some information on offshore bottom longline fishery in
the southern Adriatic Sea (GFCM Geographical Sub-Area
18). Annex of GCP/RER/010/ITA/TD-15/AdriaMed
Technical Documents n. 15.
Vacchi M., Serena F. (2010) Checklist della flora e della fauna
dei mari italiani (Parte II). Chondrichthyes. Biologia Marina
Mediterranea, 17, 642–648.
Vera L.M., Madrid J.A., S�anchez-V�azquez F.J. (2006)
Locomotor, feeding and melatonin daily rhythms in
sharpsnout seabream (Diplodus puntazzo). Physiology &
Behavior, 88, 167–172.
Vetter E.W., Dayton P.K. (1998) Macrofaunal communities
within and adjacent to a detritus-rich submarine canyon
system. Deep-Sea Research II, 45, 25–54.
Watremez P. (2012) Canyon heads in the French
Mediterranean: overview of results from the MEDSEACAN
and CORSEACAN campaigns (2008–2010). In: Wurtz M.
(Ed.), Mediterranean Submarine Canyons: Ecology and
Governance. IUCN, Gland, Switzerland and M�alaga, Spain:
105–112.
Wurtz M. (2012) Submarine canyons and their role in the
Mediterranean ecosystem. In: Wurtz M. (Ed.),
Mediterranean Submarine Canyons: Ecology and Governance.
IUCN, Gland, Switzerland and M�alaga, Spain: 11–26.
Wurtz M., Vacchi M. (1978) Ricerca di cicli nictemerali
nell’alimentazione di selaci batiali. Quaderni del Laboratorio
di Tecnologia della Pesca, Ancona, 3, 155–164.
Xavier J.C., Cherel Y., Assis C.A., Send~ao J., Borges T. (2010)
Feeding ecology of conger eels (Conger conger) in north-east
Atlantic waters. Journal of the Marine Biological Association
of the United Kingdom, 90, 493–501.
Yoklavich M.M., Greene H.G., Cailliet G.M., Sullivan D.E., Lea
N., Love M.S. (2000) Habitat associations of deep-water
rockfishes in a submarine canyon: an example of a natural
refuge. Fishery Bulletin, 98, 625–641.
16 Marine Ecology (2014) 1–16 ª 2014 Blackwell Verlag GmbH
Fish fauna by in situ observations D’Onghia, Capezzuto, Carluccio, Carlucci, Giove, Mastrototaro, Panza, Sion, Tursi & Maiorano




















