
Exon B of human surfactant protein A2 mRNA, alone or within its surrounding sequences, interacts...
14
CALL FOR PAPERS Translational Research in Acute Lung Injury and Pulmonary Fibrosis Exon B of human surfactant protein A2 mRNA, alone or within its surrounding sequences, interacts with 14-3-3; role of cis-elements and secondary structure Georgios T. Noutsios, 1 Patricia Silveyra, 1 Faizah Bhatti, 1 and Joanna Floros 1,2 1 Center for Host Defense, Inflammation, and Lung Disease (CHILD) Research Department of Pediatrics, College of Medicine The Pennsylvania State University, Hershey, Pennsylvania; and 2 Department of Obstetrics and Gynecology, College of Medicine The Pennsylvania State University, Hershey, Pennsylvania Submitted 21 September 2012; accepted in final form 3 March 2013 Noutsios GT, Silveyra P, Bhatti F, Floros J. Exon B of human surfactant protein A2 mRNA, alone or within its surrounding sequences, interacts with 14-3-3; role of cis-elements and secondary structure. Am J Physiol Lung Cell Mol Physiol 304: L722–L735, 2013. First published March 22, 2013; doi:10.1152/ajplung.00324.2012.—Human surfactant protein A, an innate immunity molecule, is encoded by two genes: SFTPA1 (SP-A1) and SFTPA2 (SP-A2). The 5= untranslated (5=UTR) splice variant of SP-A2 (ABD), but not of SP-A1 (AD), contains exon B (eB), which is an enhancer for transcription and translation. We investigated whether eB contains cis-regulatory elements that bind trans-acting factors in a sequence-specific manner as well as the role of the eB mRNA secondary structure. Binding of cytoplasmic NCI- H441 proteins to wild-type eB, eB mutant, AD, and ABD 5=UTR mRNAs were studied by RNA electromobility shift assays (REMSAs). The bound proteins were identified by mass spectroscopy and specific antibodies (Abs). We found that 1) proteins bind eB mRNA in a sequence-specific manner, with two cis-elements identified within eB to be important; 2) eB secondary structure is necessary for binding; 3) mass spectroscopy and specific Abs in REMSAs identified 14-3-3 proteins to bind (directly or indirectly) eB and the natural SP-A2 (ABD) splice variant but not the SP-A1 (AD) splice variant; 4) other ribosomal and cytoskeletal proteins, and translation factors, are also present in the eB mRNA-protein complex; 5) knockdown of 14-3-3 / isoform resulted in a downregulation of SP-A2 expression. In conclusion, proteins including the 14-3-3 family bind two cis-ele- ments within eB of hSP-A2 mRNA in a sequence- and secondary structure-specific manner. Differential regulation of SP-A1 and SP-A2 is mediated by the 14-3-3 protein family as well as by a number of other proteins that bind UTRs with or without eB mRNA. 5= untranslated regions; translation; trans-acting factors; 14-3-3 fam- ily of proteins; electromobility shift assays INTERINDIVIDUAL VARIANCES in predisposition and severity of pulmonary infections and inflammatory lung disease are poorly understood. Alveolar macrophages (AM), the sentinel cells of lung innate immunity (52), play an indispensable role against infection. Inappropriate control or imbalance of AM-mediated immune responses in the lung can cause acute or chronic inflammation. Surfactant protein A (SP-A), aside from its surfactant-related functions (i.e., stability, structure, surfactant secretion), has an essential role in innate host defense. SP-A modulates the phenotype of AM (64) and enhances phagocy- tosis of pathogens, as well as affects cytokine production and other host defense functions (14, 23, 63, 105). Human SP-A is encoded by two homologous genes (SFTPA1: SP-A1, and SFTPA2: SP-A2) with several genetic and 5= untranslated region (5=UTR) splice variants identified for each gene (Fig. 1A). Functional, structural, biophysical, and bio- chemical differences have been observed between the two SP-A proteins (26, 57, 58, 91, 92, 96 –98). Genetic heteroge- neity in SP-A genes has been shown to associate with suscep- tibility to lung infections and predisposition to lung inflam- matory diseases in neonates, children, and adults (22, 78). Alterations in total SP-A levels have been observed for several pulmonary diseases: reduced levels in airway infec- tions (7) and inflammatory lung diseases, including cystic fibrosis (55, 65) and lung transplantation (9), and higher levels in sarcoidosis (34) and hypersensitivity pneumonitis (33, 34). The ratio of SP-A1 to total SP-A protein has been shown to differ in bronchoalveolar lavage among individuals, as a func- tion of age and lung health status (85, 100), supporting the notion that imbalance of SP-A1 and SP-A2 content contributes to susceptibility to lung disease. In human SP-A mRNAs, a number of untranslated exons (A, B, B=, C, D, D=) splice in different configurations (ABD, AD=, etc.) to give rise to several 5=UTR splice variants (Fig. 1A) (44), with the AD= variant being the most common for SP-A1 and the ABD and ABD= for SP-A2. SP-A mRNAs with different 5=UTR splice variants are translated both in vitro and in vivo (44, 45). For the latter, differences were observed among individuals (45) in the relative amount of a given 5=UTR variant in the polysomal (translated) and nonpolysomal (nontranslated) fractions, indicating that SP-A 5=UTR splice variants are subject to differential translational control. Tran- sient transfection of constructs containing SP-A 5=UTR vari- ants (93) showed that constructs with the SP-A2 ABD 5=UTR are translated more efficiently than those with SP-A1 5=UTRs (AD=, AB=D=, or ACD=), indicating a role of 5=UTRs in translational control of SP-A variants. In addition, the SP-A 5=UTR ABD mRNA variant was shown to have a lower rate of Address for reprint requests and other correspondence: J. Floros, Evan Pugh Professor of Pediatrics and Obstetrics and Gynecology, Penn State CHILD Research, 500 University Dr., P.O. Box 850, Hershey, PA 17033-0850 (e-mail: jfl[email protected]). Am J Physiol Lung Cell Mol Physiol 304: L722–L735, 2013. First published March 22, 2013; doi:10.1152/ajplung.00324.2012. 1040-0605/13 Copyright © 2013 the American Physiological Society http://www.ajplung.org L722
Transcript of Exon B of human surfactant protein A2 mRNA, alone or within its surrounding sequences, interacts...

CALL FOR PAPERS Translational Research in Acute Lung Injury and Pulmonary
Fibrosis
Exon B of human surfactant protein A2 mRNA, alone or within itssurrounding sequences, interacts with 14-3-3; role of cis-elements andsecondary structure
Georgios T. Noutsios,1 Patricia Silveyra,1 Faizah Bhatti,1 and Joanna Floros1,2
1Center for Host Defense, Inflammation, and Lung Disease (CHILD) Research Department of Pediatrics, College of MedicineThe Pennsylvania State University, Hershey, Pennsylvania; and 2Department of Obstetrics and Gynecology, College ofMedicine The Pennsylvania State University, Hershey, Pennsylvania
Submitted 21 September 2012; accepted in final form 3 March 2013
Noutsios GT, Silveyra P, Bhatti F, Floros J. Exon B of humansurfactant protein A2 mRNA, alone or within its surrounding sequences,interacts with 14-3-3; role of cis-elements and secondary structure. Am JPhysiol Lung Cell Mol Physiol 304: L722–L735, 2013. First publishedMarch 22, 2013; doi:10.1152/ajplung.00324.2012.—Human surfactantprotein A, an innate immunity molecule, is encoded by two genes:SFTPA1 (SP-A1) and SFTPA2 (SP-A2). The 5= untranslated (5=UTR)splice variant of SP-A2 (ABD), but not of SP-A1 (AD), contains exonB (eB), which is an enhancer for transcription and translation. Weinvestigated whether eB contains cis-regulatory elements that bindtrans-acting factors in a sequence-specific manner as well as the roleof the eB mRNA secondary structure. Binding of cytoplasmic NCI-H441 proteins to wild-type eB, eB mutant, AD, and ABD 5=UTRmRNAs were studied by RNA electromobility shift assays (REMSAs).The bound proteins were identified by mass spectroscopy and specificantibodies (Abs). We found that 1) proteins bind eB mRNA in asequence-specific manner, with two cis-elements identified within eBto be important; 2) eB secondary structure is necessary for binding;3) mass spectroscopy and specific Abs in REMSAs identified 14-3-3proteins to bind (directly or indirectly) eB and the natural SP-A2(ABD) splice variant but not the SP-A1 (AD) splice variant; 4) otherribosomal and cytoskeletal proteins, and translation factors, are alsopresent in the eB mRNA-protein complex; 5) knockdown of 14-3-3�/� isoform resulted in a downregulation of SP-A2 expression. Inconclusion, proteins including the 14-3-3 family bind two cis-ele-ments within eB of hSP-A2 mRNA in a sequence- and secondarystructure-specific manner. Differential regulation of SP-A1 and SP-A2is mediated by the 14-3-3 protein family as well as by a number ofother proteins that bind UTRs with or without eB mRNA.
5= untranslated regions; translation; trans-acting factors; 14-3-3 fam-ily of proteins; electromobility shift assays
INTERINDIVIDUAL VARIANCES in predisposition and severity ofpulmonary infections and inflammatory lung disease are poorlyunderstood. Alveolar macrophages (AM), the sentinel cells oflung innate immunity (52), play an indispensable role againstinfection. Inappropriate control or imbalance of AM-mediatedimmune responses in the lung can cause acute or chronic
inflammation. Surfactant protein A (SP-A), aside from itssurfactant-related functions (i.e., stability, structure, surfactantsecretion), has an essential role in innate host defense. SP-Amodulates the phenotype of AM (64) and enhances phagocy-tosis of pathogens, as well as affects cytokine production andother host defense functions (14, 23, 63, 105).
Human SP-A is encoded by two homologous genes (SFTPA1:SP-A1, and SFTPA2: SP-A2) with several genetic and 5=untranslated region (5=UTR) splice variants identified for eachgene (Fig. 1A). Functional, structural, biophysical, and bio-chemical differences have been observed between the twoSP-A proteins (26, 57, 58, 91, 92, 96–98). Genetic heteroge-neity in SP-A genes has been shown to associate with suscep-tibility to lung infections and predisposition to lung inflam-matory diseases in neonates, children, and adults (22, 78).Alterations in total SP-A levels have been observed forseveral pulmonary diseases: reduced levels in airway infec-tions (7) and inflammatory lung diseases, including cysticfibrosis (55, 65) and lung transplantation (9), and higher levelsin sarcoidosis (34) and hypersensitivity pneumonitis (33, 34).The ratio of SP-A1 to total SP-A protein has been shown todiffer in bronchoalveolar lavage among individuals, as a func-tion of age and lung health status (85, 100), supporting thenotion that imbalance of SP-A1 and SP-A2 content contributesto susceptibility to lung disease.
In human SP-A mRNAs, a number of untranslated exons (A,B, B=, C, D, D=) splice in different configurations (ABD, AD=,etc.) to give rise to several 5=UTR splice variants (Fig. 1A)(44), with the AD= variant being the most common for SP-A1and the ABD and ABD= for SP-A2. SP-A mRNAs withdifferent 5=UTR splice variants are translated both in vitro andin vivo (44, 45). For the latter, differences were observedamong individuals (45) in the relative amount of a given5=UTR variant in the polysomal (translated) and nonpolysomal(nontranslated) fractions, indicating that SP-A 5=UTR splicevariants are subject to differential translational control. Tran-sient transfection of constructs containing SP-A 5=UTR vari-ants (93) showed that constructs with the SP-A2 ABD 5=UTRare translated more efficiently than those with SP-A1 5=UTRs(AD=, AB=D=, or ACD=), indicating a role of 5=UTRs intranslational control of SP-A variants. In addition, the SP-A5=UTR ABD mRNA variant was shown to have a lower rate of
Address for reprint requests and other correspondence: J. Floros, Evan PughProfessor of Pediatrics and Obstetrics and Gynecology, Penn State CHILDResearch, 500 University Dr., P.O. Box 850, Hershey, PA 17033-0850 (e-mail:[email protected]).
Am J Physiol Lung Cell Mol Physiol 304: L722–L735, 2013.First published March 22, 2013; doi:10.1152/ajplung.00324.2012.
1040-0605/13 Copyright © 2013 the American Physiological Society http://www.ajplung.orgL722

mRNA decay, compared with SP-A 5=UTR splice variants thatlack the untranslated exon B (eB). eB is included in the5=UTRs of the common SP-A2 splice variants, but not in thoseof SP-A1 variants. Furthermore, the eB was shown to containspecific regions that mediate cap-independent translation (95).Recently, eB was shown to be a transcriptional and transla-tional enhancer, and to contain regulatory elements for bothtranscription and translation (79). eB increases mRNA contentregardless of position and orientation, enhances translationwhen placed in either orientation within its natural 5=UTRsequence or in heterologous 5=UTRs.
SP-A1 and SP-A2 gene regulation is a dynamic, multifaceted,and complex biological process (80, 93, 94) that is achievedthrough a multitude of mechanisms, most of which are still not
well understood. In many genes translational regulation is medi-ated by many mRNA regulatory elements including the UTRs (2,29, 47, 49). Important roles have been identified for 5=UTRs inposttranscriptional RNA processing, mRNA stability, and local-ization, along with ribosomal recycling and translation initiation(62). Differences in SP-A expression among individuals, and/oramong variants in model systems, have been described (80, 85,93, 100). These differences, singly or synergistically, may accountfor individual susceptibilities to environmental cues, oxidativestress, disease, or other insults.
The translation initiation complex requires the concerted actionof numerous initiation factors/proteins to bind correctly and po-sition mRNAs on the 40S ribosome. Specific cis-regulatory ele-ments, frequently located primarily within the untranslated re-gions, and trans-acting factors (often proteins) bind mRNA orribosomal subunits to generate the RNA ribosomal complex andenable the translation initiation process usually via binding tospecific cis-elements. Regulatory trans-acting proteins may en-hance or attenuate translation of certain mRNAs. For example, the14-3-3 � isoform has been shown to be a regulator of mitotictranslation by binding directly eukaryotic translation/initiationfactors such as eIF4A1, eIF4B, eIF4G, eIF3, eIF2�, and eukary-otic elongation factor 1� (104). The 14-3-3 proteins are con-served, abundant, 28- to 33-kDa acidic polypeptides widely ex-pressed in all eukaryotic organisms and are involved in severalcellular processes, including apoptosis, intracellular signaling/trafficking, response to environmental cues and actin dynamics,and other interactions (1, 3, 24, 60, 81, 86, 89). In humans, �200proteins have been found to bind seven isoforms of 14-3-3 proteindirectly or as part of multiprotein complexes (68). Homo- orheterodimerization of 14-3-3 proteins is important for their func-tion and they bind a large number of partners usually by recog-nition of a phosphoserine- or phosphothreonine-rich containingmotif, resulting in modulation of the activity of the binding partnerby conformational changes, masking of phosphorylated regions,binding with other proteins, and phosphorylation (5, 6, 16, 51, 61).
In the present work, we studied the hypothesis that eBcontains cis-elements that trans-acting factors bind in a se-quence-specific manner. Toward this, 1) we generated severalin vitro mRNAs [eB WT, mRNAs containing eB (SP-A25=UTR: ABD) or lacking eB (SP-A1 5=UTR: AD)] and usedthese in shift assays to study whether specific protein factorsbind these mRNAs; 2) we generated eB deletion mutantmRNAs as well as eB mutant transcripts maintaining thesecondary structure characteristics of the WT eB to studycis-regulatory elements; 3) we used mass spectroscopy andspecific antibody (Ab) competition shift assays to identifybinding of 14-3-3; other ribosomal, cytoskeletal, and transla-tional proteins were also identified; and 4) we used shRNAs toknock down the YWHAB gene (14-3-3 �/�) in NCI-H441 cellline and correlate 14-3-3 inhibition with SP-A2 levels.
MATERIALS AND METHODS
Recombinant Constructs
Four different SP-A 5=UTRs were cloned into EcoRI and PstI sites ofpGEM-3Z vector (Promega, Madison, WI) downstream of T7 RNApolymerase promoter. The 5=UTR inserts included 1) the natural SP-A25=UTR ABD (100 nt) (44); 2) the SP-A1 5=UTR AD (70 nt) (44); 3) eB(GUC GCU GAU UUC UUG GAG CCU GAA AAG AAG); and 4) arandom (R) sequence (UCG GGC CUC AUC CCG ACG CAG UUCUUC GUC) of the same length as eB (30 nt) obtained from “shuffle
eB R
R 4
00x
Free probe
S *
NS
NS: non-specific shift; S: specific shift
1 2 3 4 5 6 7 8 9 10
S
Probe Extract - + + + + + + + + -
Competition (unlabeled
probe) - - eB - - -
lanes
5’
5’ UTR
C*AATG
AA’A’’
BB’
CC’
DD’
B D 3’
A
B
Fig. 1. 5= Untranslated region (5=UTR) structure of SP-A1 and SP-A2 and exonB (eB) sequence-specific shifts. A: exons A, B, C, D differ in length and arerepresented by lines and boxes. The sizes of each untranslated exon are asfollows: A (44 nt), A= (39 nt), A� (34 nt), B (30 nt), B= (70 nt), C (60 nt), C=(63 nt), D (26 nt), and D= (23 nt). Alternative splicing of 5=UTR exons givesrise to several SP-A1 and SP-A2 variants; the most frequent variant for SP-A1is AD= and for SP-A2 are ABD and ABD=. B: RNA electromobility shift assays(REMSA) of eB biotinylated probe with cytoplasmic extract of NCI-H441cells. Lane 1, free probe (negative control); lanes 2 and 7, 200 nM eBriboprobe incubated with 8.5 �g extract (positive controls); lanes 3–6, com-petition of eB-mediated shifts with 50-, 100-, 200-, and 400-fold molar excessof unlabeled eB RNA, respectively; lane 8, competition of eB shifts with400-fold molar excess of unlabeled random (R) RNA; lane 9, R biotinylatedprobe incubated with extract; lane 10, R probe; (�) presence of NCI-H441cytoplasmic extract; (�) absence of cytoplasmic extract or competitor RNA.The top shift band is a nonspecific shift (NS) band because it appears in therandom probe (R) (lane 9) and is not competed. Lanes 1–10 are from the samefilm but some lanes between lanes 6 and 7 were removed.
L723hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

DNA” software (84) having as template a coding sequence of the humanSP-B gene. After cloning, the EcoRI restriction site was removed and thePstI restriction endonuclease site was substituted with HindIII (to providea 5= protruding end necessary for in vitro transcription) by QuikChangeLightning Site-Directed Mutagenesis Kit (Agilent Technologies, SantaClara, CA) following the manufacturer’s protocol.
Deletion Mutant Constructs
The eB 30-nt sequence was modified by site-directed mutagenesiswith specific primers. We generated deletion (Del) mutants by a sequen-tial and overlapping deletion of 10 nucleotides (n 5) as shown inRESULTS. The primers, listed in Table 1, were synthesized in the macromo-lecular core facility of Pennsylvania State University College of Medicine.
eB Mutant Constructs With Characteristics of Secondary StructureMaintained
The QuikChange Lightning Site-Directed Mutagenesis Kit was usedto generate a series of mutant (Mut) constructs (n 6), by changingsequences corresponding to different domains of eB secondary structure
with random (R) sequences (Table 2). RNA secondary structures wereobtained by use of the RNAfold web server (http://rna.tbi.univie.ac.at/cgi-bin/RNAfold.cgi, Institute for Theoretical Chemistry, University ofVienna, Austria) (31). In the present study, we used the secondarystructure with the best possible combination of paired bases and theminimum free energy (dG); dG was normalized for size, by dividing dGby the total number of bases of each experimental construct. Thesequences of all experimental constructs were verified by DNA sequenc-ing (Eurofins MWG Operon, Huntsville, AL, and Molecular GeneticsCore Facility at Pennsylvania State University, College of Medicine).
Cell Culture
The human lung adenocarcinoma cell line NCI-H441 was pur-chased from the American Type Culture Collection (ATCC, Manas-sas, VA). The NCI-H441 cell line expresses endogenous SP-A pro-teins and thus these cells contain factors required for SP-A proteintranslation. Cells were grown in RPMI 1640 medium (Invitrogen LifeTechnologies, Carlsbad, CA) with 10% heat-inactivated fetal bovineserum (FBS; Gemini, Atlanta, GA), 1 antimycotic-antibiotic solu-
Table 1. Primers used in this study
Highlighted lower case on the 5= part of each sense (S) primer or on the 3= of antisense (A) primer indicates part of the T7 promoter sequence; lower case(not highlighted) indicates the restriction sites of the vector; the HindIII restriction site or part of it is indicated with underlined lower case; upper case indicatesthe insert cloned in the pGEM-3Z vector; exon B (eB) sequence or part of it is denoted with bold upper case, and underlined upper case indicates the modifiedsequences of stem or loops; - - - denotes a 10-nt deletion of eB sequence (see MATERIALS AND METHODS).
L724 hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

tion (Sigma, St. Louis, MO), and 1 L-glutamine (Sigma). The cellswere cultured at 37°C in 5% (vol/vol) CO2 atmosphere and weresubcultured weekly in 10-cm dishes to 50–80% confluence.
Knockdown of 14-3-3 �/� Isoform in NCI-H441 Cell Line
SureSilencing shRNA plasmids for human 14-3-3 �/� isoform(Qiagen, Valencia, CA) were employed to knock down humanYWHAB gene by RNA interference. Eight different plasmids with adifferent shRNA sequence were used. A plasmid encoding a scram-bled shRNA that does not target any human, mouse, or rat gene wasused as control. NCI-H441 cells were transfected with 0.40 �g of each14-3-3 �/�-specific shRNA plasmid and 3 �l of Attractene transfec-tion reagent (Qiagen) in Opti-MEM reduced-serum medium (LifeTechnologies, Grand Island, NY) and then were incubated for 48 h at37°C in 5% (vol/vol) CO2 atmosphere, followed by 100 �g/mlhygromycin B (Life Technologies) selection for 7 days, per vendor’sprotocol. Stably transfected NCI-H441 cell lines were isolated, ex-panded, and studied for 14-3-3 �/�, 14-3-3 �/�, SP-A2, SP-A1 andactin expression by Western immunoblot. We used rabbit polyclonalAbs for detection of 14-3-3 �/� and 14-3-3 �/� (Cell Signaling,Danvers, MA) in dilution 1:1,000, rabbit polyclonal anti-SP-A2(Aviva, San Diego, CA) in dilution 1:5,000, chicken SP-A1 gene-specific antibody (IgY) (85) in dilution 1:500, mouse monoclonalanti-actin (Sigma) in dilution 1:2,000, and appropriate secondary horse-radish peroxidase-conjugated Abs (Bio-Rad, Hercules, CA). Western Light-ning Plus-ECL Chemiluminescence substrate (PerkinElmer, Waltham, MA)was used to detect proteins.
Cytoplasmic Fractionation
Cytoplasmic extracts were obtained by using the NE-PER Nuclearand Cytoplasmic Extraction kit (Thermo Scientific, Rockford, IL),after harvest with PBS and trypsinization. Protein concentration wasdetermined by the BCA protein assay (Thermo Scientific), followingthe manufacturer’s instructions. Potential cross-contamination be-tween cytosolic and nuclear fractions was checked by immunoblotusing antibodies MEK1/2 (cytoplasmic) and c-Jun and/or histone H3(nuclear) (Cell Signaling).
In Vitro Transcription
Plasmids (n 15) containing WT ABD, experimental AD, eB, R,Del 1–5, and Mut 1–6 were linearized by digestion with HindIII(Invitrogen, Grand Island, NY) and purified by QIAquick PCR puri-fication kit (Qiagen). RNA probes for gel mobility shift experimentswere synthesized in vitro with the AmpliScribe T7-Flash Biotin-RNATranscription Kit (Epicentre, Madison, WI), by using the linearizedplasmid DNA as template following the manufacturer’s protocol. Inbrief, a 20-�l reaction, containing 110 ng of linear plasmid DNA, 2 �lof 10 transcription buffer, 9 mM of NTP/Biotin-UTP PreMix, 10mM DTT, 40 U RiboGuard RNase Inhibitor, and 10 U of AmpliScribeT7-Flash RNA Polymerase (Epicentre, Madison, WI) was incubatedat 37°C for 1 h. One unit of RNase-Free DNase I (Epicentre), was
added and the reaction was incubated at 37°C for 15 min to removethe DNA template. Unlabeled probes were prepared likewise with9 mM of nonbiotinylated NTPs (Epicentre). RNA transcripts werepurified using RNA-bee (TEL-TEST, Friendswood, TX) in con-junction with Direct-zol RNA MiniPrep columns (Zymo Research,Irvine, CA).
RNA Electromobility Shift Assay
Biotinylated RNA transcripts were heated at 80°C for 20 min andrapidly cooled on ice for 1 min prior to incubation with cytoplasmicprotein extracts. Binding reactions (20 �l) contained 10 mM HEPESpH 7.3, 20 mM KCl, 1 mM MgCl2, 1 mM DTT, 1% (vol/vol)glycerol, 40 U of RNaseOUT recombinant ribonuclease inhibitor(Invitrogen, Carlsbad, CA), 2 �g of tRNA nonspecific competitor,200 nM of in vitro transcribed biotinylated RNA, and 8.5 �g ofcytoplasmic NCI-H441 extract. Reaction mixtures were incubated atroom temperature (RT) for 20 min prior to loading on the gel. Forcompetition shift assays a 50- to 400-fold molar excess of unlabeledin vitro transcribed RNA was used to compete specific interactionsbetween RNA and cytoplasmic extracts. Unlabeled probes were in-cubated with protein extracts at RT 10 min prior to addition ofbiotinylated probe.
Samples were fractionated on native 6% (wt/vol) polyacrylamidegels in 1 Tris-borate-EDTA (TBE) and transferred to positivelycharged nylon membranes (Roche, Indianapolis, IN) by use of asemidry Amersham Biosciences Nova blot apparatus in 1 TBE at400 mA for 30 min. Membranes were exposed to UV (254 nm) 120mJ/cm2 for 45 s (Spectrolinker XL-1000 UV cross-linker; Spectron-ics, New Cassel, NY) for cross-linking. Band shifts were detected byuse of the Chemiluminescent Nucleic Acid Detection Module(Thermo Scientific) and autoradiography (HyBlot CL film, DenvilleScientific, Saint-Laurent, QC, Canada).
For identification of 14-3-3 on RNA electromobility shift assay(REMSA), 1–2 �g of 14-3-3 rabbit polyclonal antibody that recog-nizes all 14-3-3 isoforms (Abcam, Cambridge, MA) was added to thebinding reactions and incubated for an additional 20 min at RT, forabrogation of the specific shift and/or supershift formation. As nega-tive control the anti-DDK monoclonal antibody (anti-tag) (Origene,Rockville, MD) was used.
Mass Spectroscopy
RNA-protein complexes were fractionated on native 6% (wt/vol)polyacrylamide gel in 1 TBE buffer and stained with mass spec-troscopy compatible SilverQuest Silver Staining Kit (Invitrogen)following the manufacturer’s protocol. The stained gel was superim-posed on the X-ray film from the REMSA and the bands correspond-ing to RNA-mediated specific shifts were excised. Silver ions wereremoved following the destaining protocol of the manufacturer. Thedestained gel bands were dehydrated in 100% (vol/vol) methanol for5 min at RT, rehydrated in 30% (vol/vol) methanol for 5 min at RT,washed twice with ultrapure water for 10 min, followed by three
Table 2. Sequences of eB mutants with conserved secondary structure
Pseudo-Loop Stem Loop Stem Shift 1 (*) Shift 2 (circled)
Exon B (WT) GUC GCU GAU UUC UU G GAG CCU GAA AAG AAG � �Mut 1 GUC GCU GAU uga au G GAG CCU GAA auu cag � �Mut 2 GUC GCU GAU ccc cc G GAG CCU GAA ggg ggg � �Mut 3 ucg ggc cuu ccc cc u acg cag uuc ggg ggc � �Mut 4 ucg ggc cuu UUC UU g ucg cag uua AAG AAc � �Mut 5 GUC GCU GAU ccc cc u acg cag uuc ggg ggg � �Mut 6 ucg ggc cuu ccc cc u GAG CCU GAu ggg ggc � �
WT, wild-type eB; upper case (bold or regular) indicates eB sequence; upper case and bold indicates the 2 cis-elements within eB; lower case denotes themutant sequence; Mut, mutant. Pseudo-loop and loop are shown, respectively, with asterisk (*) and circle in shift assays of Figs. 1B, 3A, and 4; presence (�)or absence (�) of specific shift.
L725hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

washings with 100 mM ammonium bicarbonate in 30% (vol/vol)acetonitrile for 10 min at RT. Gel was then cut into small pieces andwashed thoroughly in ultrapure water, dried in SpeedVac for 30 min,and resuspended in 50 mM ammonium bicarbonate. Ten nanogramsof trypsin were added to each sample, which was then incubated at37°C overnight and centrifuged at 12,000 g for 1 min, and thesupernatant was transferred to a sterile microcentrifuge tube. Theremaining peptides in the gel pellet were extracted with 20 �l 50%(vol/vol) acetonitrile containing 0.1% (vol/vol) trifluoroacetic acid atRT and combined with the previous supernatant. Samples wereconcentrated to 5 �l by use of SpeedVac and then analyzed withLC-MS/MS (Keck MS and Proteomics Yale University School ofMedicine, New Haven, CT and Proteomics Core Facility at Pennsyl-vania State University, College of Medicine, Hershey, PA). A numberof independent LC-MS/MS analyses were done from differentREMSAs. For eB, n 4; AD, n 3; ABD, n 3; Del 4, n 2; Mut5, n 2; Mut 6, n 2; eB competed with ABD, n 2; eB competedwith AD, n 2.
RESULTS
Our published studies showed that SP-A 5=UTR splicevariants differentially affect translation (45, 93) and that eB isan enhancer of transcription and translation (79). In this study,we found that trans-acting factors bind eB specific sequences,supporting the hypothesis that interactions between SP-A5=UTR splice variants and regulatory proteins differentiallyaffect expression of SP-A1 and SP-A2 (43, 46, 74).
Sequence-Specific Protein Shifts of the Untranslated eB
When eB in vitro transcribed biotinylated RNA as a probeand NCI-H441 cytoplasmic extracts were used, sequence-specific shifts (S) were observed (Fig. 1B, lanes 2 and 7). TheeB shifts were competed (lanes 3-6) with eB-unlabeled probe(50- to 400-fold molar excess). However, the shifts remained(lane 8) when a random sequence probe (R) was used forcompetition. The R probe migrates to the bottom of the gel, andit did not form a sequence-specific shift with NCI-H441 cellextracts (lane 9). A non-sequence-specific (NS) shift wasobserved with all probes used. Similar observations were madewith whole NCI-H441 extract (not shown). We next 1) studiedwhether specific sequences and/or the secondary structure ofeB are required for the formation of the specific shift,2) studied proteins that bind directly or indirectly and,3) carried out a proof-of-principle experiment to determinewhether inhibition of one of the identified proteins, shown tobind eB alone or within its surrounding sequences, correlatedwith reduced SP-A2 levels.
Specific eB Sequence Elements and Secondary StructureContribute to the Sequence-Specific Protein Shifts
Two groups of mutants were generated, as described inMATERIALS AND METHODS. One group included five deletionmutants and the other group included six mutants with eBsecondary structure characteristics maintained.
Deletion mapping of eB cis-acting elements. Figure 2 showsthat, of the five mutants studied, only deletion mutants 3 and 4retained the eB-specific shift (A), indicating that the sequenceinvolved in the shift is located in the first half of eB toward thejunction with exon A (B).
eB mutants with eB secondary structure characteristicsmaintained identify two cis-elements. The predicted secondarystructure of the wild-type (WT) eB is operationally described
here as consisting of a pseudo-loop, a stem, and a loop (Fig.2C). Because the deletion mutants studied above (Fig. 2, A andB) disrupt normal formation of eB RNA secondary structure(Fig. 2C), we generated six mutants (Table 2) where the eBnucleotides were replaced with nucleotides unrelated to eB.These nucleotides were chosen so that the secondary structureof eB WT shown in Fig. 2C was maintained.
A summary of these mutants is shown in Table 2 and thesequences of the cis-elements in pseudo-loop and loop aremarked. REMSAs (Fig. 3A) of Mut 1 and 2 with altered stemsequences produced prominent upper and lower shifts. Incontrast, Mut 3 and 4 with altered loop sequences did not formshifts. The sequence of the pseudo-loop connects to exon Aand the sequence of the loop is close to exon D, indicating thatbinding of protein factors occurs near A-B and B-D junctions.Interestingly, mutants with random sequences in either loop(Mut 5) or pseudo-loop (Mut 6) formed upper and lower shifts,respectively, indicating formation of different eB-cytoplasmicprotein complexes at A and D junctions. The shift generated byMut 5 exhibits a higher size (Fig. 3A, lane 12, marked with *)compared with that generated by Mut 6 (lane 14, marked bycircle). Both of these are generated by eB (Fig. 1B, lanes 2 and7, *, circle). The shifts of the eB mutant RNAs were competedwith 200 molar excess of eB probe (Fig. 3A, lanes 5, 7, 13,and 15). However, when eB-mediated shifts were competedwith 200 molar excess of eB mutant probes (Fig. 3B), thesemutants (with secondary structure maintained as WT eB)eliminated or significantly attenuated the intensity of the shiftregardless of whether they had partial or no sequence similar-ity. These observations are consistent with those of the deletionmutants (Fig. 2) and together support the notion that bothsequence and secondary structure are important for formationand/or competition of the eB-mediated shift.
The sequence of the cis-element 1 (marked with a blue linein Fig. 2C) identified in the present manuscript is present inboth Del 3 and Del 4. These deletion mutants form a large loopstructure, and presumably the nucleotide sequence of the cis-element is free to interact and bind proteins whereas in Del 5the sequence of this cis-element spans from the pseudo-loop tothe stem and the loop (Fig. 2C, Del 5). It is possible that theDel 5 structure does not enable protein interactions as shownby its inability to form a shift. On the other hand, in Del 1 andDel 2 the cis-element 1 is deleted and these did not form anyshifts despite the fact that these deletion constructs contain thenucleotide sequence of the cis-element 2. We speculate that thecis-element 2 alone (i.e., without the presence of the cis-element 1) cannot form a shift. However, Mut 6 (Fig. 3A, lane14) that lacks the cis-element 1 (but contains cis-element 2)forms the lower size shift. Mut 6 in contrast to Del 1 and Del2 maintains a secondary structure for both cis-elements similarto that of the WT eB. We conclude that the sequence and/or thesecondary structure of cis-element 1 are necessary for a shift toform by the cis-element 2.
Mass Spectroscopy Analysis of Bound Proteins
LC-MS/MS analysis of the eB-, eB mutant-, AD-, andABD-protein complexes identified a number of potential pro-tein factors. The most significant expectation scores identifiedhuman 14-3-3 isoforms (�/�, , �/�, ε, �, �/�, �) to interactwith eB, eB mutants, or ABD (Table 3) but not with the AD
L726 hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

riboprobe, indicating that 14-3-3 proteins interact with eB or5=UTRs containing eB. Of note, the following proteins werealso found in REMSA protein shifts of ABD or eB: ribosomal40S and 60S subunit proteins (SA and S3 for 40S subunit, andL5, L7, L11, L15, L22, and L26 for the 60S subunit), elonga-tion factor 1-�1 (EF 1-�1), eukaryotic initiation factors (eIF4A,eIF4H, eIF6), cytokeratin 17 (KRT17), actin, tubulin, and calre-ticulin. However, for the AD-specific shift significantly fewerribosomal or other proteins were present (Table 3 and Supple-mental Table S1). Together, these data indicate that a numberof proteins directly or indirectly bind eB, or ABD and AD, andthat the 14-3-3 family of proteins is one of the groups ofproteins that differentially bind eB or ABD but not AD.
The 14-3-3 Protein Family Is Present in the eB- and theSP-A2 ABD-Mediated RNA-Protein Complex
eB-mediated shifts. To further determine the presence andimportance of the eB-mediated shifts we performed REMSA inthe presence of 14-3-3 Ab. Figure 4 shows that the Ab thatrecognizes all isoforms of 14-3-3 competed the two specificshifts (S) (lanes 3 and 4) whereas an irrelevant control Ab(DDK) did not affect the eB shifts (lane 5). This is consistentwith the mass spectroscopy data and provides further supportof the presence of 14-3-3 in eB-mediated shift.
To further investigate the mass spectroscopy data that iden-tified most human 14-3-3 isoforms and the REMSAs that in the
Del 3-0.42 kcal mol-1
Del 4-0.44 kcal mol-1
Del 5-1.37 kcal mol-1
Del 1-0.92 kcal mol-1
Del 2-0.50 kcal mol-1
eB -2.69 kcal mol-1 l or -0.09 kcal mol-1 nt-1
Loop:GGA GCC UGA A
cis element #2
Pseudo-loop: GUC GCU GAU
cis element #1
stem
Pseudo-loop
Loop
Presence of cis element #2 Presence of cis element #1
eB (WT) Del 2Del 1 Del 3 Del 5
Free probe
S
Del 4
NS
NS:non-specific shift; S:specific shiftPresence (+) or Absence (-) of cytoplasmic H441 extract
+ - + + + + + +
S
pGEM-3Z vector
eB (30 nt)
GTC GCT GAT TTC TTG GAG CCT GAA AAG AAGeB (WT)--------------------------TC TTG GAG CCT GAA AAG AAG
GTC GC-----------------------G GAG CCT GAA AAG AAG
GTC GCT GAT T---------------------------T GAA AAG AAG
GTC GCT GAT TTC TTG-----------------------------AG AAG
GTC GCT GAT TTC TTG GAG CC--------------------------
Del 1Del 2Del 3Del 4Del 5
HindIII
5’UTRT7
A B
C
Fig. 2. Deletion mapping of eB cis-acting elements and predicted secondary structure of WT eB and deletion mutant mRNAs. A: REMSA results of biotinylatedwild-type (WT) and deletion (Del) mutant riboprobes with cytoplasmic extract of NCI-H441 cells. Del 3 and Del 4 formed sequence-specific shifts whereas therest deletion mutants did not. B: the WT eB sequence and the sequences of the deletion mutants. The mutants were generated by site-directed mutagenesis inpGEM-3Z vector by sequentially removing 10 nt. C: the secondary structure of WT eB and deletion mutants, as well as the sequence and location of the twoeB cis-elements. The eB secondary structure is described by a pseudo-loop, a stem, and a loop. The secondary structure of Del 5 differs from that of Del 3 andDel 4. In particular, the secondary structures of both Del 3 and Del 4 are single loops whereas Del 5 results in a small loop, a short stem, and a short pseudo-loop.The RNAfold online software was used to obtain the secondary structure of WT eB and deletion mutants. The minimum free energy secondary structure wasused, with the best possible combination of paired bases, and the minimum free energy (�G) normalized for size, by dividing �G by the total number of basesof each experimental construct. The colors indicate the propensity of individual nucleotides to participate in base pairs. The scale ranges from red (highestprobability) to blue-violet (lower probability).
L727hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org
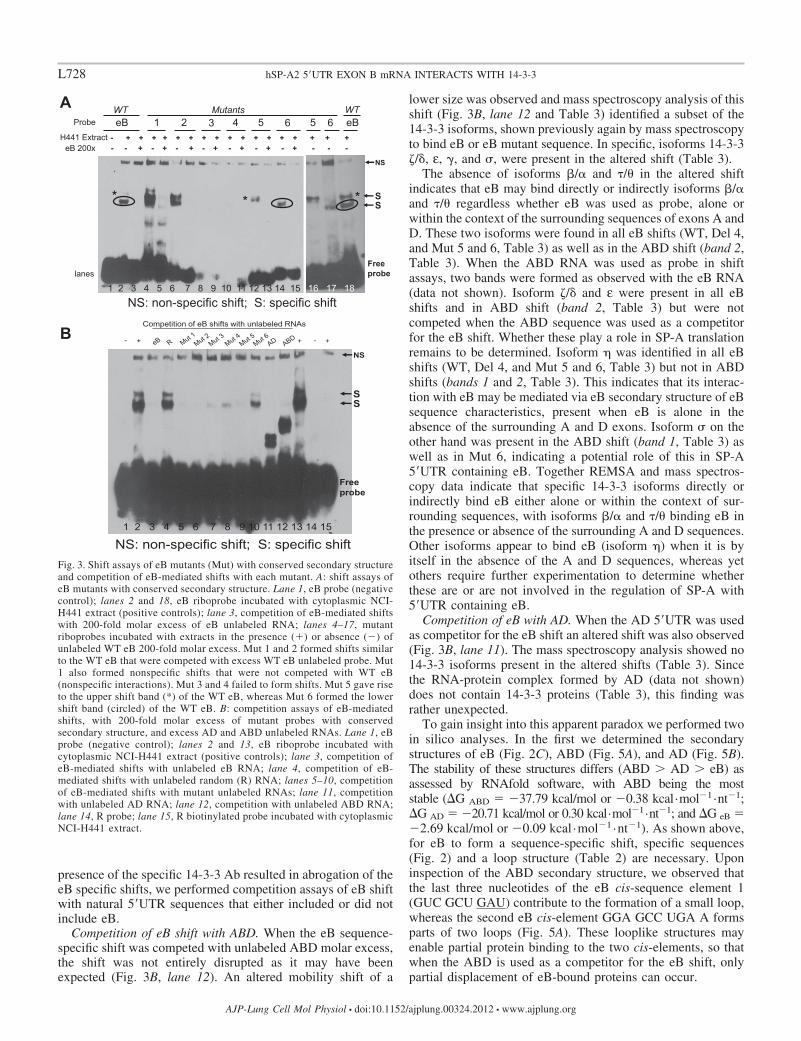
presence of the specific 14-3-3 Ab resulted in abrogation of theeB specific shifts, we performed competition assays of eB shiftwith natural 5=UTR sequences that either included or did notinclude eB.
Competition of eB shift with ABD. When the eB sequence-specific shift was competed with unlabeled ABD molar excess,the shift was not entirely disrupted as it may have beenexpected (Fig. 3B, lane 12). An altered mobility shift of a
lower size was observed and mass spectroscopy analysis of thisshift (Fig. 3B, lane 12 and Table 3) identified a subset of the14-3-3 isoforms, shown previously again by mass spectroscopyto bind eB or eB mutant sequence. In specific, isoforms 14-3-3�/�, ε, , and �, were present in the altered shift (Table 3).
The absence of isoforms �/� and �/� in the altered shiftindicates that eB may bind directly or indirectly isoforms �/�and �/� regardless whether eB was used as probe, alone orwithin the context of the surrounding sequences of exons A andD. These two isoforms were found in all eB shifts (WT, Del 4,and Mut 5 and 6, Table 3) as well as in the ABD shift (band 2,Table 3). When the ABD RNA was used as probe in shiftassays, two bands were formed as observed with the eB RNA(data not shown). Isoform �/� and ε were present in all eBshifts and in ABD shift (band 2, Table 3) but were notcompeted when the ABD sequence was used as a competitorfor the eB shift. Whether these play a role in SP-A translationremains to be determined. Isoform � was identified in all eBshifts (WT, Del 4, and Mut 5 and 6, Table 3) but not in ABDshifts (bands 1 and 2, Table 3). This indicates that its interac-tion with eB may be mediated via eB secondary structure of eBsequence characteristics, present when eB is alone in theabsence of the surrounding A and D exons. Isoform � on theother hand was present in the ABD shift (band 1, Table 3) aswell as in Mut 6, indicating a potential role of this in SP-A5=UTR containing eB. Together REMSA and mass spectros-copy data indicate that specific 14-3-3 isoforms directly orindirectly bind eB either alone or within the context of sur-rounding sequences, with isoforms �/� and �/� binding eB inthe presence or absence of the surrounding A and D sequences.Other isoforms appear to bind eB (isoform �) when it is byitself in the absence of the A and D sequences, whereas yetothers require further experimentation to determine whetherthese are or are not involved in the regulation of SP-A with5=UTR containing eB.
Competition of eB with AD. When the AD 5=UTR was usedas competitor for the eB shift an altered shift was also observed(Fig. 3B, lane 11). The mass spectroscopy analysis showed no14-3-3 isoforms present in the altered shifts (Table 3). Sincethe RNA-protein complex formed by AD (data not shown)does not contain 14-3-3 proteins (Table 3), this finding wasrather unexpected.
To gain insight into this apparent paradox we performed twoin silico analyses. In the first we determined the secondarystructures of eB (Fig. 2C), ABD (Fig. 5A), and AD (Fig. 5B).The stability of these structures differs (ABD � AD � eB) asassessed by RNAfold software, with ABD being the moststable (�G ABD �37.79 kcal/mol or �0.38 kcal·mol�1·nt�1;�G AD �20.71 kcal/mol or 0.30 kcal·mol�1·nt�1; and �G eB �2.69 kcal/mol or �0.09 kcal ·mol�1 ·nt�1). As shown above,for eB to form a sequence-specific shift, specific sequences(Fig. 2) and a loop structure (Table 2) are necessary. Uponinspection of the ABD secondary structure, we observed thatthe last three nucleotides of the eB cis-sequence element 1(GUC GCU GAU) contribute to the formation of a small loop,whereas the second eB cis-element GGA GCC UGA A formsparts of two loops (Fig. 5A). These looplike structures mayenable partial protein binding to the two cis-elements, so thatwhen the ABD is used as a competitor for the eB shift, onlypartial displacement of eB-bound proteins can occur.
1eB
NS: non-specific shift; S: specific shift
NS
Free probe
* * *
2 3 4 5 6 5 6 eBProbe
- + + + + + + + + + + + + + + + + +- - + - + - + - + - + - + - + - - -
WT Mutants WT
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18lanes
SS
H441 ExtracteB 200x
+ +-- +
Competition of eB shifts with unlabeled RNAs
NS
Free probe
SS
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
NS: non-specific shift; S: specific shift
A
B
Fig. 3. Shift assays of eB mutants (Mut) with conserved secondary structureand competition of eB-mediated shifts with each mutant. A: shift assays ofeB mutants with conserved secondary structure. Lane 1, eB probe (negativecontrol); lanes 2 and 18, eB riboprobe incubated with cytoplasmic NCI-H441 extract (positive controls); lane 3, competition of eB-mediated shiftswith 200-fold molar excess of eB unlabeled RNA; lanes 4–17, mutantriboprobes incubated with extracts in the presence (�) or absence (�) ofunlabeled WT eB 200-fold molar excess. Mut 1 and 2 formed shifts similarto the WT eB that were competed with excess WT eB unlabeled probe. Mut1 also formed nonspecific shifts that were not competed with WT eB(nonspecific interactions). Mut 3 and 4 failed to form shifts. Mut 5 gave riseto the upper shift band (*) of the WT eB, whereas Mut 6 formed the lowershift band (circled) of the WT eB. B: competition assays of eB-mediatedshifts, with 200-fold molar excess of mutant probes with conservedsecondary structure, and excess AD and ABD unlabeled RNAs. Lane 1, eBprobe (negative control); lanes 2 and 13, eB riboprobe incubated withcytoplasmic NCI-H441 extract (positive controls); lane 3, competition ofeB-mediated shifts with unlabeled eB RNA; lane 4, competition of eB-mediated shifts with unlabeled random (R) RNA; lanes 5–10, competitionof eB-mediated shifts with mutant unlabeled RNAs; lane 11, competitionwith unlabeled AD RNA; lane 12, competition with unlabeled ABD RNA;lane 14, R probe; lane 15, R biotinylated probe incubated with cytoplasmicNCI-H441 extract.
L728 hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

In the second in silico analysis using a Matrix-based motifdiscovery tool algorithm (17), we sought to determine regula-tory elements within eB, by analyzing the entire ABD se-quence. The eB sequence was identified as an importantregulatory element (except for the last 6 nt AAG AAG that
constitute one part of the eB stem, Table 2, and comparison ofFig. 2C and Fig. 5C), plus 6 nt of exon A immediately adjacentto eB (Fig. 5C). The in silico identification of a cis-elementconsisting of adjacent exon A and eB sequences holds thepotential for factors to bind both exons A (3= end) and B (5=end) within the natural ABD 5=UTR. It is possible that whenAD is used in eB shift assays as a competitor (at 200-foldmolar excess) it partially disrupts the eB-protein complex bydisplacing proteins that could bind both the 3= end of exon Aand the 5= end of eB. On the basis of the collective data above,the AD competition is likely to affect the upper shift (*) of eB(Fig. 1B). The requirement of binding sequences in both exonA and eB may contribute to the instability of this shift whenexon A sequences are absent. It is also possible that the ADcompetition indirectly affects the lower shift band (Fig. 1B,circle) resulting in a shift that lacks 14-3-3 proteins (Fig. 3B,lane 11). Proteins, other than the 14-3-3, ribosomal, andcytoskeleton, were still present in the altered shift, when ADwas used as competitor. Moreover, the AD higher secondarystructure stability (compared with eB, Figs. 2C and 5B) andperhaps other characteristics (yet to be defined) may contributeto this protein displacement and partial dissociation of the eBcomplex.
Correlation of 14-3-3 �/� Isoform Inhibition and SP-A2Expression in NCI-H441 Cell Line
To study the functional role of 14-3-3 �/� protein andcorrelate it with SP-A2 expression and translation in NCI-H441 cells, as proof of principle, we employed shRNAs thattarget the human YWHAB gene. The shRNAs knocked downthe 14-3-3 �/� isoform to a varying degree, as shown byWestern immunoblot (Fig. 6), and this correlated with theexpression of SP-A2. The scrambled shRNA (negative control)did not inhibit 14-3-3 �/� protein and the levels of SP-A2 were
Table 3. Mass-spectroscopy protein identification of WT eB, AD, ABD, and mutant-mediated specific shifts
eB band 1 (*) eB band 2 (circled) Del 4 Mut 5 Mut 6 AD ABD band 1 ABD band 2 eB comp.ABD eB comp. AD
14-3-3 �/� � � � � � � �14-3-3 �/� � � � � � �14-3-3 �/� � � � � � �14-3-3 ε � � � � � � �14-3-3 � � � �14-3-3 � � � �14-3-3 � � � � � �40S ribosomal protein SA � �40S ribosomal protein S3 �ubiquitin-40S ribosomal protein S27a � �60S ribosomal protein L5 � � � � � �60S ribosomal protein L7 �60S ribosomal protein L11 � �60S ribosomal protein L15 �60S ribosomal protein L22 � � � � � � �60S ribosomal protein L26 �Elongation factor 1-�1 � � � � �eIF4H � � �eIF4A �eIF6 � �Actin, cytoplasmic � � � � � � � �tubulin � � � � � � � �KRT17 � �calreticulin � � � � � � �
�, Presence of the particular protein factor; eB comp. ABD, eB shift was competed with excess ABD; eB comp. AD, eB shift was competed with excess AD. Del,deletion.
- +
DD
K
eB
Free
probe
S
S: specific shift
- +
1 2 3 4 5 6 7
* S
Controls
+ + extract Ab
+ - - - - + +
Controls
*
lanes
Fig. 4. eB shift competition with specific 14-3-3 antibody (Ab). REMSA of eBbiotinylated probe with NCI-H441 cytoplasmic extract in the presence of Abagainst all isoforms of the 14-3-3 protein family. Lanes 1 and 7, eB probe(negative controls); lanes 2 and 6, eB riboprobe incubated with extract(positive controls); lanes 3 and 4, 14-3-3 Ab disrupts the eB-specific shifts;lane 5, eB riboprobe incubated with extract and an irrelevant control anti-DDKAb. S, specific shift.
L729hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

high compared with experimental shRNA that inhibited 14-3-3�/�, while the expression of SP-A1 remained unaffected. Thelevels of 14-3-3 �/� and actin (serving as controls) did notchange. These data indicate that 14-3-3 �/� protein plays a rolein the regulation of SP-A2 expression.
DISCUSSION
Human SP-A, an important molecule of lung host defense, isencoded by two functional genes, and a number of geneticvariants have been reported for each gene (19). Several SP-A1and SP-A2 5=UTR splice variants have been identified (44, 54),with differences in mRNA stability and translational efficiency(93). We have shown that eB of the SP-A2 5=UTR mRNA is anenhancer for translation and transcription (79). In the presentstudy, we studied the hypothesis that trans-acting factors bindeB alone or within the context of the surrounding sequences tomodulate the previously observed translation efficiency. Wefound that cytoplasmic NCI-H441 cell extract proteins bind eBRNA in a sequence-specific manner and that both specificcis-elements within eB and eB secondary structure are neces-sary for binding. Members of the 3-monooxygenase/trypto-phan 5-monooxygenase activation protein family (14-3-3) bind(directly or indirectly) eB and the natural SP-A2 (ABD) splicevariant but not the SP-A1 (AD) splice variant that lacks eB.The knockdown of 14-3-3 �/� isoform in NCI-H441 cellsresults in a downregulated expression of SP-A2. Besides the14-3-3, a number of other proteins (ribosomal, cytoskeletal,
and translation factors) are also present in the eB mRNA-14-3-3 complex.
The deletion and mutation mapping analyses identified twocis-elements, element 1 GUC GCU GAU immediately next toexon A and element 2 GGA GCC UGA A near exon D, withineB that are important for protein interactions, as assessed bythe presence of sequence-specific shifts. These shifts failed toform when both cis-elements were missing. However, if thecharacteristics of eB’s secondary structure were maintained,the presence of one or the other cis-element was sufficient togenerate the upper or lower band shift, indicating that bothsequence elements and the secondary structure are necessaryfor the formation of the eB sequence-specific shifts. Thus eBregulatory sequence motifs and eB-mediated secondary struc-ture characteristics may contribute to the higher level of trans-lation observed with transcripts of SP-A 5=UTRs that containthe eB sequence (79, 93). Hence, differences in SP-A1 andSP-A2 translation may result in different immune profiles andresponses to bacterial and inflammatory stressors. Understand-ing these differences is important in determining individualdisease response patterns.
The fact that the motif gugugggucg CUGAUUUCUUggagccugaa, identified by in silico analysis, contained most ofthe eB sequence supports the notion that the eB sequence isneeded for functional and structural integrity of the SP-A2ABD 5=UTR. Furthermore, this in silico-identified regulatoryelement contained both eB cis-elements identified by mutation
ABD
AD -20.71 kcal mol-1 or -0.30 kcal mol-1 nt-1
-37.79 kcal mol-1 or -0.38 kcal mol-1 nt-1 GGA GCC UGA A GUC GCU GAU
A
B
Exon A Exon D
**
eB
Exon D
Exon A **
gugugggucg CUGAUUUCUU ggagccugaa
In silico predicted cis regulatory element
eB AD
** C
Fig. 5. Secondary structure of ABD and AD and cis-elements. A and B depict the secondary structures for ABD and AD, respectively. The sequence and thelocation of the two eB cis-elements is noted (A), with blue and red colors. The RNAfold online software was used to obtain the secondary structures as describedin Fig. 2C. C: the cis-element identified within ABD by the matrix-based motif discovery algorithm. The ** denotes the last 6 nt of exon A that were includedin the identified cis-element. The sequence in blue and red denotes the 2 eB cis-elements, the eB pseudo-loop and loop, respectively. The free energy �G inkcal/mol and the color scale 0 to 1 (red, most stable) are shown.
L730 hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

mapping analysis and shown to be both functionally andstructurally important. Consistent with this is our publishedwork in which deletion of the pseudo-loop or the loop of eB(described in the present study) resulted in reduced internalribosome entry site (IRES) activity (95). The importance ofboth the sequence specificity and the secondary structure inRNA function has been shown to be crucial in other systems ofinnate host defense (69, 101, 102). For example, the 20S RNANarnavirus, via its secondary structure can evade the 5=-exonuclease SKI1/XRN1 host defense in Saccharomycescerevisiae (21), and the human U1 RNA, owing to its double-stranded secondary structure, can induce innate immunity sig-naling by activating TLR-3 (37).
Mass spectroscopy findings and/or shift assays data with14-3-3 specific Ab demonstrated that 5=UTR RNA-proteininteractions include the 14-3-3 protein family. Isoforms �/�and �/�, in particular, bind eB regardless whether eB is foundin the presence or absence of the surrounding sequences ofexons A and D, as shown by competition of eB shift assayswith ABD. Furthermore, knockdown of the 14-3-3 �/� isoformin NCI-H441 cell line results in a downregulation of SP-A2expression, reinforcing the notion that the interaction betweeneB mRNA and 14-3-3 �/� is very important for the efficienttranslation of SP-A.
On the other hand, isoforms �/� and ε, present in all eB shiftsand in the ABD shift 2, failed to be competed when the ABDwas used as competitor. It is possible that differences in thesecondary structure characteristics between eB and ABD,along with potential differences in the other binding proteins,“protect” the binding of these isoforms in eB from the ABDcompetitor. As a result, these 14-3-3 isoforms are still presentin the altered eB shift after competition with ABD.
However, a number of 14-3-3 isoforms ( , �, �) werepresent in shifts when eB WT ( , �) or various mutant
sequences ( , �, �) were used as probes. These ( , �) were notcompeted with ABD and were ( , �) absent in the ABD shift.It is possible that the secondary structure characteristics of eBin the absence of the surrounding sequences of exons A and Dare such that promote binding of other 14-3-3 isoforms such asthe and � isoforms and that these isoforms do not actuallybind eB in its natural sequence context (i.e., ABD sequence).However, further experimentation and use of additional meth-odologies are required to assess the precise interactions and therole of various 14-3-3 isoforms in SP-A variant expression.
The involvement of 14-3-3 in SP-A regulation is ratherintriguing. The 14-3-3 protein family is involved in the innatehost defense of various species. Isoforms �/� and �/� identifiedin the present study to bind eB are evolutionary conservedproteins and are required for phagocytosis and microbial re-sistance in Drosophila, zebrafish larvae, and mouse RAW264.7 cells (88). SP-A also plays a role in phagocytosis andinnate immunity processes, with differences being observed inthe ability of SP-A1 and SP-A2 to modulate phagocytosis (56,57) and regulate inflammatory processes (38, 97, 99) in thealveolar macrophage. Of interest, differences in the ratio ofSP-A1 to total SP-A in bronchoalveolar lavage samples havebeen observed between healthy population and nonhealthyindividuals (infection, cystic fibrosis, asthma) (85, 100), raisingthe possibility that regulatory differences exist between SP-A1and SP-A2 under various conditions. Moreover, levels of14-3-3 have been shown to change in disease, with increasedlevels observed in cerebrospinal fluid after infection with prionin Creutzfeldt-Jakob disease and spongiform encephalopathies(30). It is possible that the 14-3-3 proteins are involved inmechanisms that differentially affect SP-A1 and SP-A2 levels.The fact that inhibition of one of the 14-3-3 isoforms (�/�)shown to differentially bind SP-A1 and SP-A2 5=UTRs resultsin the downregulation of SP-A2 indicates an important role ofthis isoform in SP-A translation.
Moreover, isoform �/� has been shown to be a phagosome-associated protein (8) that dimerizes in response to mitogen-activated protein kinase-dependent inflammatory signaling(67). The tyrosine kinase of the epidermal growth factorreceptor has been shown to regulate SP-A expression andmodulate inflammation (39). Whether 14-3-3 regulates SP-Aexpression via tyrosine kinase pathways remains to be deter-mined. However, the identification of the 14-3-3 proteins asnovel phagosomal proteins (27), and the correlation betweeninhibition of 14-3-3 (�/�) and the reduced levels of SP-A2,provides, in general, further support for their involvement ininnate immunity, and, in specific, the regulation of the innatehost defense molecule, SP-A. The �/� isoform identified in thepresent study as an eB-specific binding protein has been shownto play a regulatory role in Toll-like receptor 2 (TLR2) signal-ing pathway within the innate immune system (75). SP-A hasalso been demonstrated to interact directly with TLR2, modu-lating inflammatory responses against bacterial components(59), indicating a potential role of the 14-3-3 in this process.
Ribosomal and cytoskeletal proteins and initiation and elon-gation translation factors were found by mass spectroscopy tobe included in the WT eB, eB mutants, or ABD shifts. Thesewere virtually all competed when ABD was used as a compet-itor for the eB shift. This indicates that eB, either alone orwithin the context of the surrounding A and D sequences, is animportant element for bringing together protein factors neces-
35 25
kDa 1
scrambled
14-3-3 β/α (28 kDa)
35 25 14-3-3 τ/θ (28 kDa)
shRNAs
40 35
actin (42 kDa)
2 3 4 5 6 ctrl
40 35 SP-A1 (35 kDa)
40 35 SP-A2 (35 kDa)
Fig. 6. Knockdown of 14-3-3 �/� isoform in NCI-H441 cell line and corre-lation with SP-A2 expression. Six different shRNAs (1–6), plus another 2 (notshown), were used to knock down the expression of 14-3-3 �/� isoform byRNA interference. A scrambled shRNA (negative control) that does not targetany human, mouse, or rat gene was used as control. Western immunoblot wasemployed to monitor the expression of 14-3-3 �/� isoform (28 kDa), SP-A2(35 kDa), SP-A1 (35 kDa), 14-3-3 �/� isoform (28 kDa), and actin (42 kDa).The experimental shRNAs inhibited the 14-3-3 �/� isoform to a varyingdegree and this correlated with levels of SP-A2. No differences were observedfor SP-A1 and 14-3-3 �/�; actin was used as internal control.
L731hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

sary for the enhanced translation observed with SP-A UTRsthat contain eB (79, 93).
The AD shift, on the other hand, did not contain any 14-3-3proteins and not as many of the other group of proteinsindicating significant differences in the RNA-protein interac-tions between AD, and eB or ABD. This is consistent withpublished data where significant differences in translation andmRNA stability were observed between SP-As with ADand ABD 5=UTRs (93, 95). The 40S ribosomal proteins, SAand S3, appear to bind exon A and/or D but not eB. These arenecessary for Met-tRNA to bind 40S and translation initiation(13). The S3 is found in ribonucleoprotein granules (42) and inhepatitis C IRES-mediated translation complex (103). How-ever, the absence of the 60S ribosomal protein L5 in the SP-A15=UTR AD splice variant may alter translation efficiency orpose a major disadvantage in terms of translation comparedwith SP-A2 5=UTR ABD splice variant. L5 is required forrRNA maturation and formation of the 60S ribosomal subunits(28). Furthermore, the AD splice variant does not bind the 60Sribosomal protein L7, shown to play a regulatory role intranslation by binding to G-rich structures in mRNAs (35).Another 60S ribosomal protein, L22, a structural constituent ofribosome, was shown to bind all probes except AD and Mut 6,indicating that the nucleotide content of the eB pseudo-loop(GUC GCU GAU) may be the cis-element that elicits thebinding of this trans-acting factor. These observations supportthe notion that differences exist between AD and eB-contain-ing UTRs in their interactions with ribosomal proteins, and thatthese may differentially impact the translation of SP-As withdifferent UTRs.
Cytoskeletal proteins actin and tubulin-� were present vir-tually in all probes indicating that a cytoskeleton-mediatedstabilization of SP-A RNA is required for all SP-A RNAs witheither AD or ABD 5=UTR. This observation is consistent withprevious findings in which cytoskeletal proteins were shown tostabilize the surfactant protein mRNAs in rat alveolar type IIcells (77). Together these indicate that mechanisms of cyto-skeleton-mediated stabilization of AD and ABD mRNAs aresimilar.
Interestingly, eukaryotic initiation factor 4A (eIF4A) inter-acted only with the natural ABD variant, and not the eB alone,or the AD; eIF4H was found in eB and two mutants (Del 4 andMut 5). It seems that this RNA helicase (eIF4A) requires eB tobe within its natural context as in the SP-A2 5=UTR ABDvariant. The group of eIF4A and eIF4H are DEAD box heli-cases (18, 53, 83) that help the scanning process of the 40Ssmall ribosomal unit by unwinding the secondary structure(82). Thus these may enable the SP-A2 5=UTR ABD splicevariant to be more efficiently translated than the SP-A1 5=UTRAD that does not interact with these helicases.
The direct or indirect binding of a multitude of proteins toeB was also seen in the competition assays. Consistently, in allcases competition of an mRNA-protein complex with molarexcess of another mRNA resulted in partial dissociation of thiscomplex. The observations from the competition and massspectroscopy experiments, when ABD and AD RNAs wereused as competitors for the eB-mediated shifts, were unex-pected. These are summarized in Fig. 7. One may have ex-pected that the ABD competitor would have displaced the14-3-3 isoforms, since these were found by mass spectroscopyto be present only in shifts of eB or ABD probes and not of the
AD probe. However, on the basis of the in silico analysis inwhich 6 nt at the 3= end of exon A were identified as part of theeB regulatory element, we speculate that the ABD (but not theAD) competitor somehow provides a certain level of stabilityto A-B junction, maintaining thus a partial displaced RNA-protein complex with most of 14-3-3 still present. Anotherpossibility is that an altered 3D structure formed by ABD andeB together creates an artificial platform upon which thedisrupted 14-3-3 complexes may bind. Future experiments arewarranted to gain insight into these possibilities.
In summary, eB, which was shown previously to be anenhancer for both translation and transcription of SP-A2 (79),contains two cis-elements, and trans-acting factors bind eB ina sequence- and a secondary structure-specific manner. Thebinding factors include 14-3-3 �/� and �/� isoforms, alongwith ribosomal, cytoskeletal, and translation factors and per-haps additional 14-3-3 isoforms. In particular, 14-3-3 �/�isoform, shown to be part of the eB-protein complex eitherwhen eB was alone or in the context of the surroundingsequences of exon A and exon D, is likely to play an importantrole in the translation of SP-A. The trans-acting proteins thatbind eB in the presence or absence of exon A and D sequencesprobably play important roles in the translation and the differ-ential regulation of SP-As with 5=UTRs that contain or lack eB.Future studies are needed to characterize the specific RNA-protein interactions and delineate the underlying molecularmechanisms involved in the SP-A variant differential regula-tion.
ACKNOWLEDGMENTS
The authors thank Dr. D. S. Phelps and T. M. Umstead for input and adviceon mass spectrometry analysis, and L. Murray for typing.
Present address for F. Bhatti: Neonatal-Perinatal Medicine, University ofOklahoma Health Sciences Center, Oklahoma City, OK.
14-3-3 ζ/δ, β/α,
θ
14-3-3 ε, η
14-3-3 ζ/δ, ε
14-3-3 γ, σ
+ competedproteins
+ competed proteins
Fig. 7. Schematic representation of competition of eB-mediated shifts with200-fold molar excess of unlabeled AD and ABD RNAs. The figure depicts thecollective results of eB shift assays, competition shift assays, and massspectroscopy protein identification of the sequence-specific shifts. The 14-3-3protein family (�/�, �/�, ε, �, and �/�), 40S and 60S ribosomal proteins, alongwith cytoskeletal proteins (actin and tubulin) bind directly or indirectly eB.Competition with molar excess of ABD gives rise to an altered eB mRNA-protein complex, which includes 14-3-3 �/�, ε, , � isoforms, L22 60Sribosomal protein, S27a 60S ribosomal protein, and actin; the rest of theproteins (including 14-3-3 isoforms �/� and �/�) are competed by the ABDprobe. Competition with molar excess of AD results in a smaller eB mRNA-protein complex that includes L22 and L26 60S ribosomal proteins, S27a 60Sribosomal protein, eIF6, and actin; the rest of the proteins (including all of the14-3-3 family of proteins) are competed with the AD probe.
L732 hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

GRANTS
This work was supported by National Heart, Lung, and Blood InstituteGrant HL34788.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
G.T.N., P.S., and J.F. conceived and designed the research; G.T.N. and F.B.performed experiments; G.T.N. and J.F. analyzed data; G.T.N. and J.F.interpreted results of experiments; G.T.N. prepared figures; G.T.N. draftedmanuscript; G.T.N. and P.S. edited and revised manuscript; G.T.N., P.S., F.B.,and J.F. approved final version of manuscript.
REFERENCES
1. Aitken A. Functional specificity in 14-3-3 isoform interactions throughdimer formation and phosphorylation. Chromosome location of mamma-lian isoforms and variants. Plant Mol Biol 50: 993–1010, 2002.
2. Andreassi C, Riccio A. To localize or not to localize: mRNA fate is in3=UTR ends. Trends Cell Biol 19: 465–474, 2009.
3. Baldin V. 14-3-3 proteins and growth control. Prog Cell Cycle Res 4:49–60, 2000.
4. Brands JH, Maassen JA, van Hemert FJ, Amons R, Möller W. Theprimary structure of the alpha subunit of human elongation factor 1.Structural aspects of guanine-nucleotide-binding sites. Eur J Biochem155: 167–171, 1986.
5. Braselmann S, McCormick F. Bcr and Raf form a complex in vivo via14-3-3 proteins. EMBO J 14: 4839–4848, 1995.
6. Bridges D, Moorhead GBG. 14-3-3 proteins: a number of functions fora numbered protein. Sci STKE 2004: re10-, 2004.
7. Bruce SR, Atkins CL, Colasurdo GN, Alcorn JL. Respiratory syncy-tial virus infection alters surfactant protein A expression in humanpulmonary epithelial cells by reducing translation efficiency. Am JPhysiol Lung Cell Mol Physiol 297: L559–L567, 2009.
8. Burlak C, Whitney AR, Mead DJ, Hackstadt T, Deleo FR. Maturationof human neutrophil phagosomes includes incorporation of molecularchaperones and endoplasmic reticulum quality control machinery. MolCell Proteomics 5: 620–634, 2006.
9. Casals C, Varela A, Ruano ML, Valiño F, Pérez-Gil J, Torre N,Jorge E, Tendillo F, Castillo-Olivares JL. Increase of C-reactiveprotein and decrease of surfactant protein A in surfactant after lungtransplantation. Am J Respir Crit Care Med 157: 43–49, 1998.
10. Ceci M, Gaviraghi C, Gorrini C, Sala LA, Offenhäuser N, MarchisioPC, Biffo S. Release of eIF6 (p27BBP) from the 60S subunit allows 80Sribosome assembly. Nature 426: 579–584, 2003.
11. Chang JH, Cho YH, Sohn SY, Choi JM, Kim A, Kim YC, Jang SK,Cho Y. Crystal structure of the eIF4A-PDCD4 complex. Proc Natl AcadSci USA 106: 3148–3153, 2009.
12. Cheng ST, Nguyen TQ, Yang YS, Capra JD, Sontheimer RD. Cal-reticulin binds hYRNA and the 52-kDa polypeptide component of theRo/SS-A ribonucleoprotein autoantigen. J Immunol 156: 4484–4491,1996.
13. Clemens MJ, Henshaw EC, Rahamimoff H, London IM. Met-tRNAfMet binding to 40S ribosomal subunits: a site for the regulation ofinitiation of protein synthesis by hemin. Proc Natl Acad Sci USA 71:2946–2950, 1974.
14. Crouch EC. Collectins and pulmonary host defense. Am J Respir CellMol Biol 19: 177–201, 1998.
15. Dabiri GA, Young CL, Rosenbloom J, Southwick FS. Molecularcloning of human macrophage capping protein cDNA. A unique memberof the gelsolin/villin family expressed primarily in macrophages. J BiolChem 267: 16545–16552, 1992.
16. Davezac N, Baldin V, Gabrielli B, Forrest A, Theis-Febvre N,Yashida M, Ducommun B. Regulation of CDC25B phosphatases sub-cellular localization. Oncogene 19: 2179–2185, 2000.
17. Defrance M, van Helden J. info-gibbs: a motif discovery algorithm thatdirectly optimizes information content during sampling. Bioinformatics25: 2715–2722, 2009.
18. Dever TE. Gene-specific regulation by general translation factors. Cell108: 545–556, 2002.
19. DiAngelo S, Lin Z, Wang G, Phillips S, Ramet M, Luo J, Floros J.Novel, non-radioactive, simple and multiplex PCR-cRFLP methods for
genotyping human SP-A and SP-D marker alleles. Dis Markers 15:269–281, 1999.
20. Dubois T, Rommel C, Howell S, Steinhussen U, Soneji Y, Morrice N,Moelling K, Aitken A. 14-3-3 is phosphorylated by casein kinase I onresidue 233. Phosphorylation at this site in vivo regulates Raf/14-3-3interaction. J Biol Chem 272: 28882–28888, 1997.
21. Esteban R, Vega L, Fujimura T. 20S RNA narnavirus defies theantiviral activity of SKI1/XRN1 in Saccharomyces cerevisiae. J BiolChem 283: 25812–25820, 2008.
22. Floros J, Thomas N. Genetic variations of surfactant proteins and lunginjury. In: Surfactant in Pathogenesis and Treatment of Lung Disease,edited by Nakos G, Papathanasiou A. Trivandrum, India: ResearchSignpost, 2009, p. 25–48.
23. Floros J, Wang G, Mikerov AN. Genetic complexity of the humaninnate host defense molecules, surfactant protein A1 (SP-A1) and SP-A2—impact on function. Crit Rev Eukaryot Gene Expr 19: 125–137,2009.
24. Fu H, Subramanian RR, Masters SC. 14-3-3 Proteins: structure,function, regulation. Annu Rev Pharmacol Toxicol 40: 617–647, 2000.
25. Fujita Y, Okamoto T, Noshiro M, McKeehan WL, Crabb JW,Whitney RG, Kato Y, Sato JD, Takada K. A novel heparin-bindingprotein, HBp15, is identified as mammalian ribosomal protein L22.Biochem Biophys Res Commun 199: 706–713, 1994.
26. García-Verdugo I, Wang G, Floros J, Casals C. Structural analysis andlipid-binding properties of recombinant human surfactant protein a de-rived from one or both genes. Biochemistry 41: 14041–14053, 2002.
27. Garin J, Diez R, Kieffer S, Dermine JF, Duclos S, Gagnon E, SadoulR, Rondeau C, Desjardins M. The phagosome proteome: insight intophagosome functions. J Cell Biol 152: 165–180, 2001.
28. Gazda HT, Sheen MR, Vlachos A, Choesmel V, O’Donohue MF,Schneider H, Darras N, Hasman C, Sieff CA, Newburger PE, BallSE, Niewiadomska E, Matysiak M, Zaucha JM, Glader B, NiemeyerC, Meerpohl JJ, Atsidaftos E, Lipton JM, Gleizes PE, Beggs AH.Ribosomal protein L5 and L11 mutations are associated with cleft palateand abnormal thumbs in Diamond-Blackfan anemia patients. Am J HumGenet 83: 769–780, 2008.
29. Gebauer F, Hentze MW. Molecular mechanisms of translational con-trol. Nat Rev Mol Cell Biol 5: 827–835, 2004.
30. Gmitterová K, Heinemann U, Bodemer M, Krasnianski A, MeissnerB, Kretzschmar HA, Zerr I. 14-3-3 CSF levels in sporadic Creutzfeldt-Jakob disease differ across molecular subtypes. Neurobiol Aging 30:1842–1850, 2009.
31. Gruber AR, Lorenz R, Bernhart SH, Neuböck R, Hofacker IL. TheVienna RNA websuite. Nucleic Acids Res 36: W70–W74, 2008.
32. Gu YM, Jin YH, Choi JK, Baek KH, Yeo CY, Lee KY. Protein kinaseA phosphorylates and regulates dimerization of 14-3-3 epsilon. FEBSLett 580: 305–310, 2006.
33. Guzman J, Wang YM, Kalaycioglu O, Schoenfeld B, Hamm H,Bartsch W, Costabel U. Increased surfactant protein A content in humanalveolar macrophages in hypersensitivity pneumonitis. Acta Cytol 36:668–673, 1992.
34. Hamm H, Lührs J, Guzman y Rotaeche J, Costabel U, Fabel H,Bartsch W. Elevated surfactant protein A in bronchoalveolar lavagefluids from sarcoidosis and hypersensitivity pneumonitis patients. Chest106: 1766–1770, 1994.
35. Hemmerich P, von Mikecz A, Neumann F, Sözeri O, Wolff-VorbeckG, Zoebelein R, Krawinkel U. Structural and functional properties ofribosomal protein L7 from humans and rodents. Nucleic Acids Res 21:223–231, 1993.
36. Hermeking H, Lengauer C, Polyak K, He TC, Zhang L, Thiagal-ingam S, Kinzler KW, Vogelstein B. 14-3-3� is a p53-regulatedinhibitor of G2/M progression. Mol Cell 1: 3–11, 1997.
37. Hoffman RW, Gazitt T, Foecking MF, Ortmann RA, Misfeldt M,Jorgenson R, Young SL, Greidinger EL. U1 RNA induces innateimmunity signaling. Arthritis Rheum 50: 2891–2896, 2004.
38. Huang W, Wang G, Phelps DS, Al-Mondhiry H, Floros J. HumanSP-A genetic variants and bleomycin-induced cytokine production byTHP-1 cells: effect of ozone-induced SP-A oxidation. Am J Physiol LungCell Mol Physiol 286: L546–L553, 2004.
39. Inoue A, Xin H, Suzuki T, Kanehira M, Kuroki Y, Fukuhara T,Kikuchi T, Maemondo M, Nukiwa T, Saijo Y. Suppression of surfac-tant protein A by an epidermal growth factor receptor tyrosine kinaseinhibitor exacerbates lung inflammation. Cancer Sci 99: 1679–1684,2008.
L733hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

40. Jang SW, Liu X, Fu H, Rees H, Yepes M, Levey A, Ye K. Interactionof Akt-phosphorylated SRPK2 with 14-3-3 mediates cell cycle and celldeath in neurons. J Biol Chem 284: 24512–24525, 2009.
41. Jin Y, Dai MS, Lu SZ, Xu Y, Luo Z, Zhao Y, Lu H. 14-3-3 binds toMDMX that is phosphorylated by UV-activated Chk1, resulting in p53activation. EMBO J 25: 1207–1218, 2006.
42. Jønson L, Vikesaa J, Krogh A, Nielsen LK, Hansen T, Borup R,Johnsen AH, Christiansen J, Nielsen FC. Molecular composition ofIMP1 ribonucleoprotein granules. Mol Cell Proteomics 6: 798–811,2007.
43. Karinch AM, Deiter G, Ballard PL, Floros J. Regulation of expressionof human SP-A1 and SP-A2 genes in fetal lung explant culture. BiochimBiophys Acta 1398: 192–202, 1998.
44. Karinch AM, Floros J. 5= Splicing and allelic variants of the humanpulmonary surfactant protein A genes. Am J Respir Cell Mol Biol 12:77–88, 1995.
45. Karinch AM, Floros J. Translation in vivo of 5= untranslated-regionsplice variants of human surfactant protein-A. Biochem J 307: 327–330,1995.
46. Kumar AR, Snyder JM. Differential regulation of SP-A1 and SP-A2genes by cAMP, glucocorticoids, and insulin. Am J Physiol Lung CellMol Physiol 274: L177–L185, 1998.
47. Le Quesne JP, Spriggs KA, Bushell M, Willis AE. Dysregulation ofprotein synthesis and disease. J Pathol 220: 140–151, 2010.
48. Liu Y, Ross JF, Bodine PV, Billiard J. Homodimerization of Ror2tyrosine kinase receptor induces 14-3-3� phosphorylation and promotesosteoblast differentiation and bone formation. Mol Endocrinol 21: 3050–3061, 2007.
49. Liu Y, Wimmer E, Paul AV. Cis-acting RNA elements in human andanimal plus-strand RNA viruses. Biochim Biophys Acta 1789: 495–517,2009.
50. Loh PG, Yang HS, Walsh MA, Wang Q, Wang X, Cheng Z, Liu D,Song H. Structural basis for translational inhibition by the tumoursuppressor Pdcd4. EMBO J 28: 274–285, 2009.
51. Margolis SS, Walsh S, Weiser DC, Yoshida M, Shenolikar S, Korn-bluth S. PP1 control of M phase entry exerted through 14-3-3-regulatedCdc25 dephosphorylation. EMBO J 22: 5734–5745, 2003.
52. Martin TR, Frevert CW. Innate immunity in the lungs. Proc AmThorac Soc 2: 403–411, 2005.
53. Mathews MB. Lost in translation. Trends Biochem Sci 27: 267–269,2002.
54. McCormick SM, Boggaram V, Mendelson CR. Characterization ofmRNA transcripts and organization of human SP-A1 and SP-A2 genes.Am J Physiol Lung Cell Mol Physiol 266: L354–L366, 1994.
55. Meyer KC, Sharma A, Brown R, Weatherly M, Moya FR, Lewan-doski J, Zimmerman JJ. Function and composition of pulmonarysurfactant and surfactant-derived fatty acid profiles are altered in youngadults with cystic fibrosis. Chest 118: 164–174, 2000.
56. Mikerov A, Wang G, Umstead T, Zacharatos M, Thomas N, PhelpsD, Floros J. Surfactant protein A2 (SP-A2) variants expressed in CHOcells stimulate phagocytosis of Pseudomonas aeruginosa more than doSP-A1 variants. Infect Immun 75: 1403–1412, 2007.
57. Mikerov AN, Umstead TM, Gan X, Huang W, Guo X, Wang G,Phelps DS, Floros J. Impact of ozone exposure on the phagocyticactivity of human surfactant protein A (SP-A) and SP-A variants. Am JPhysiol Lung Cell Mol Physiol 294: L121–L130, 2008.
58. Mikerov AN, Umstead TM, Huang W, Liu W, Phelps D, Floros J.SP-A1 and SP-A2 variants differentially enhance association of Pseu-domonas aeruginosa with rat alveolar macrophages. Am J Physiol LungCell Mol Physiol 288: L150–L158, 2005.
59. Murakami S, Iwaki D, Mitsuzawa H, Sano H, Takahashi H, VoelkerDR, Akino T, Kuroki Y. Surfactant protein A inhibits peptidoglycan-induced tumor necrosis factor-� secretion in U937 cells and alveolarmacrophages by direct interaction with Toll-like receptor 2. J Biol Chem277: 6830–6837, 2002.
60. Muslin AJ, Xing H. 14-3-3 proteins: regulation of subcellular localiza-tion by molecular interference. Cell Signal 12: 703–709, 2000.
61. Obsil T, Ghirlando R, Klein DC, Ganguly S, Dyda F. Crystal structureof the 14-3-3zeta:serotonin N-acetyltransferase complex. A role forscaffolding in enzyme regulation. Cell 105: 257–267, 2001.
62. Pesole G, Mignone F, Gissi C, Grillo G, Licciulli F, Liuni S. Structuraland functional features of eukaryotic mRNA untranslated regions. Gene276: 73–81, 2001.
63. Phelps DS. Surfactant regulation of host defense function in the lung: aquestion of balance. Pediatr Pathol Mol Med 20: 269–292, 2001.
64. Phelps DS, Umstead TM, Quintero OA, Yengo CM, Floros J. In vivorescue of alveolar macrophages from SP-A knockout mice with exoge-nous SP-A nearly restores a wild type intracellular proteome; actininvolvement. Proteome Sci 9: 67, 2011.
65. Postle AD, Mander A, Reid KB, Wang JY, Wright SM, Moustaki M,Warner JO. Deficient hydrophilic lung surfactant proteins A and D withnormal surfactant phospholipid molecular species in cystic fibrosis. Am JRespir Cell Mol Biol 20: 90–98, 1999.
66. Powell DW, Rane MJ, Chen Q, Singh S, McLeish KR. Identificationof 14-3-3� as a protein kinase B/Akt substrate. J Biol Chem 277:21639–21642, 2002.
67. Powell DW, Rane MJ, Joughin BA, Kalmukova R, Hong JH, TidorB, Dean WL, Pierce WM, Klein JB, Yaffe MB, McLeish KR.Proteomic identification of 14-3-3zeta as a mitogen-activated proteinkinase-activated protein kinase 2 substrate: role in dimer formation andligand binding. Mol Cell Biol 23: 5376–5387, 2003.
68. Pozuelo Rubio M, Geraghty KM, Wong BH, Wood NT, CampbellDG, Morrice N, Mackintosh C. 14-3-3-affinity purification of over 200human phosphoproteins reveals new links to regulation of cellular me-tabolism, proliferation and trafficking. Biochem J 379: 395–408, 2004.
69. Qi Y, Ding B. Inhibition of cell growth and shoot development by aspecific nucleotide sequence in a noncoding viroid RNA. Plant Cell 15:1360–1374, 2003.
70. Richter NJ, Rogers GW, Hensold JO, Merrick WC. Further biochem-ical and kinetic characterization of human eukaryotic initiation factor 4H.J Biol Chem 274: 35415–35424, 1999.
71. Robledo S, Idol RA, Crimmins DL, Ladenson JH, Mason PJ, BesslerM. The role of human ribosomal proteins in the maturation of rRNA andribosome production. RNA 14: 1918–1929, 2008.
72. Rogers GW, Richter NJ, Lima WF, Merrick WC. Modulation of thehelicase activity of eIF4A by eIF4B, eIF4H, and eIF4F. J Biol Chem 276:30914–30922, 2001.
73. Sato S, Fujita N, Tsuruo T. Regulation of kinase activity of 3-phos-phoinositide-dependent protein kinase-1 by binding to 14-3-3. J BiolChem 277: 39360–39367, 2002.
74. Scavo LM, Ertsey R, Gao BQ. Human surfactant proteins A1 and A2are differentially regulated during development and by soluble factors.Am J Physiol Lung Cell Mol Physiol 275: L653–L669, 1998.
75. Schuster TB, Costina V, Findeisen P, Neumaier M, Ahmad-Nejad P.Identification and functional characterization of 14-3-3 in TLR2 signal-ing. J Proteome Res 10: 4661–4670, 2011.
76. Schütz P, Karlberg T, van den Berg S, Collins R, Lehtiö L, HögbomM, Holmberg-Schiavone L, Tempel W, Park HW, Hammarström M,Moche M, Thorsell AG, Schüler H. Comparative structural analysis ofhuman DEAD-box RNA helicases. PLoS One 5, 2010.
77. Shannon JM, Pan T, Edeen KE, Nielsen LD. Influence of the cyto-skeleton on surfactant protein gene expression in cultured rat alveolartype II cells. Am J Physiol Lung Cell Mol Physiol 274: L87–L96, 1998.
78. Silveyra P, Floros J. Genetic variant associations of human SP-A andSP-D with acute and chronic lung injury. Front Biosci 17: 407–429,2012.
79. Silveyra P, Raval M, Simmons B, Diangelo S, Wang G, Floros J. Theuntranslated exon B of human surfactant protein A2 mRNAs is anenhancer for transcription and translation. Am J Physiol Lung Cell MolPhysiol 301: L795–L803, 2011.
80. Silveyra P, Wang G, Floros J. Human SP-A1 (SFTPA1) variant-specific 3= UTRs and poly(A) tail differentially affect the in vitrotranslation of a reporter gene. Am J Physiol Lung Cell Mol Physiol 299:L523–L534, 2010.
81. Skoulakis EM, Davis RL. 14-3-3 proteins in neuronal development andfunction. Mol Neurobiol 16: 269–284, 1998.
82. Sonenberg N. Cap-binding proteins of eukaryotic messenger RNA:functions in initiation and control of translation. Prog Nucleic Acid ResMol Biol 35: 173–207, 1988.
83. Sonenberg N, Dever TE. Eukaryotic translation initiation factors andregulators. Curr Opin Struct Biol 13: 56–63, 2003.
84. Stothard P. The sequence manipulation suite: JavaScript programs foranalyzing and formatting protein and DNA sequences. Biotechniques 28:1102, 1104, 2000.
85. Tagaram HR, Wang G, Umstead TM, Mikerov AN, Thomas NJ,Graff GR, Hess JC, Thomassen MJ, Kavuru MS, Phelps DS, FlorosJ. Characterization of a human surfactant protein A1 (SP-A1) gene-
L734 hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org

specific antibody; SP-A1 content variation among individuals of varyingage and pulmonary health. Am J Physiol Lung Cell Mol Physiol 292:L1052–L1063, 2007.
86. Todd A, Cossons N, Aitken A, Price GB, Zannis-Hadjopoulos M.Human cruciform binding protein belongs to the 14-3-3 family. Biochem-istry 37: 14317–14325, 1998.
87. Troyanovsky SM, Leube RE, Franke WW. Characterization of thehuman gene encoding cytokeratin 17 and its expression pattern. Eur JCell Biol 59: 127–137, 1992.
88. Ulvila J, Vanha-aho LM, Kleino A, Vähä-Mäkilä M, Vuoksio M,Eskelinen S, Hultmark D, Kocks C, Hallman M, Parikka M, RämetM. Cofilin regulator 14-3-3� is an evolutionarily conserved proteinrequired for phagocytosis and microbial resistance. J Leukoc Biol 89:649–659, 2011.
89. van Hemert MJ, Steensma HY, van Heusden GP. 14-3-3 proteins: keyregulators of cell division, signalling and apoptosis. Bioessays 23: 936–946, 2001.
90. Vladimirov SN, Ivanov AV, Karpova GG, Musolyamov AK, EgorovTA, Thiede B, Wittmann-Liebold B, Otto A. Characterization of thehuman small-ribosomal-subunit proteins by N-terminal and internal se-quencing, and mass spectrometry. Eur J Biochem 239: 144–149, 1996.
91. Wang G, Bates-Kenney SR, Tao JQ, Phelps DS, Floros J. Differencesin biochemical properties and in biological function between humanSP-A1 and SP-A2 variants, and the impact of ozone-induced oxidation.Biochemistry 43: 4227–4239, 2004.
92. Wang G, Guo X, Diangelo S, Thomas NJ, Floros J. HumanizedSFTPA1 and SFTPA2 transgenic mice reveal functional divergence ofSP-A1 and SP-A2: formation of tubular myelin in vivo requires bothgene products. J Biol Chem 285: 11998–12010, 2010.
93. Wang G, Guo X, Floros J. Differences in the translation efficiency andmRNA stability mediated by 5=-UTR splice variants of human SP-A1and SP-A2 genes. Am J Physiol Lung Cell Mol Physiol 289: L497–L508,2005.
94. Wang G, Guo X, Floros J. Human SP-A 3=-UTR variants mediatedifferential gene expression in basal levels and in response to dexameth-asone. Am J Physiol Lung Cell Mol Physiol 284: L738–L748, 2003.
95. Wang G, Guo X, Silveyra P, Kimball SR, Floros J. Cap-independenttranslation of human SP-A 5=-UTR variants: a double-loop structure and
cis-element contribution. Am J Physiol Lung Cell Mol Physiol 296:L635–L647, 2009.
96. Wang G, Myers C, Mikerov A, Floros J. Role of Cys85 of human SP-Avariants on their biochemical properties and biological function. FASEBJ 21: A552, 2007.
97. Wang G, Phelps DS, Umstead TM, Floros J. Human SP-A proteinvariants derived from one or both genes stimulate TNF-� production inthe THP-1 cell line. Am J Physiol Lung Cell Mol Physiol 278: L946–L954, 2000.
98. Wang G, Taneva S, Keough KM, Floros J. Differential effects ofhuman SP-A1 and SP-A2 variants on phospholipid monolayers contain-ing surfactant protein B. Biochim Biophys Acta 1768: 2060–2069, 2007.
99. Wang JY, Reid KB. The immunoregulatory roles of lung surfactantcollectins SP-A, and SP-D, in allergen-induced airway inflammation.Immunobiology 212: 417–425, 2007.
100. Wang Y, Voelker DR, Lugogo NL, Wang G, Floros J, Ingram JL,Chu HW, Church TD, Kandasamy P, Fertel D, Wright JR, Kraft M.Surfactant protein A is defective in abrogating inflammation in asthma.Am J Physiol Lung Cell Mol Physiol 301: L598–L606, 2011.
101. Wassenegger M, Spieker RL, Thalmeir S, Gast FU, Riedel L, SängerHL. A single nucleotide substitution converts potato spindle tuber viroid(PSTVd) from a noninfectious to an infectious RNA for Nicotianatabacum. Virology 226: 191–197, 1996.
102. Watts JM, Dang KK, Gorelick RJ, Leonard CW, Bess JW, Swan-strom R, Burch CL, Weeks KM. Architecture and secondary structureof an entire HIV-1 RNA genome. Nature 460: 711–716, 2009.
103. Weinlich S, Hüttelmaier S, Schierhorn A, Behrens SE, Ostareck-Lederer A, Ostareck DH. IGF2BP1 enhances HCV IRES-mediatedtranslation initiation via the 3=UTR. RNA 15: 1528–1542, 2009.
104. Wilker EW, van Vugt MA, Artim SA, Huang PH, Petersen CP,Reinhardt HC, Feng Y, Sharp PA, Sonenberg N, White FM, YaffeMB. 14-3-3� controls mitotic translation to facilitate cytokinesis. Nature446: 329–332, 2007.
105. Wright JR. Immunomodulatory functions of surfactant. Physiol Rev 77:931–962, 1997.
106. Yu Y, Ji H, Doudna JA, Leary JA. Mass spectrometric analysis of thehuman 40S ribosomal subunit: native and HCV IRES-bound complexes.Protein Sci 14: 1438–1446, 2005.
L735hSP-A2 5=UTR EXON B mRNA INTERACTS WITH 14-3-3
AJP-Lung Cell Mol Physiol • doi:10.1152/ajplung.00324.2012 • www.ajplung.org




















