
Exogenous Glycinebetaine and Salicylic Acid Application Improves Water Relations, Allometry and...
12
DROUGHT STRESS Exogenous Glycinebetaine and Salicylic Acid Application Improves Water Relations, Allometry and Quality of Hybrid Sunflower under Water Deficit Conditions M. Hussain 1 , M. A. Malik 1 , M. Farooq 1 , M. B. Khan 2 , M. Akram 1 & M. F. Saleem 1 1 Department of Agronomy, University of Agriculture, Faisalabad, Pakistan 2 University College of Agriculture, Bahauddin Zakariya University, Multan, Pakistan Introduction Water paucity is a severe environmental constraint to crop productivity worldwide. Drought-induced loss in crop yield perhaps exceeds the loss from all other causes, as both severity and duration of stress are critical (Farooq et al. 2008a). Drought stress reduces crop growth rate (CGR) and yield regardless of the growth stage at which it occurs in arable crops including sunflower (Jensen and Mogenson 1984). Under severe water-deficiency, cell elongation of higher plants can be inhibited by interruption of water flow from the xylem to the surrounding elongating cells (Nonami 1998). Impaired mitosis, cell elongation and expansion result in reduced plant height, leaf area and crop growth under drought (Nonami 1998, Kaya et al. 2006). Drought stress disturbs the plant water relations, photosynthesis, assimilate translocation and economic yield in filed crops (Siddique et al. 2000, Atteya 2003, Farooq et al. 2008a,b, Hussain et al. 2008a,b). One of the most common stress tolerance strategies in plants is the overproduction of different types of compatible organic solutes (Serraj and Sinclair 2002). Compatible solutes are of low molecular weight, highly soluble compounds that are usually non-toxic even at high cytosolic concentra- tions. Generally, they protect plants from stress through Keywords drought stress; exogenous application; glycinebetaine; growth; salicylic acid; sunflower; water relations; water use efficiency Correspondence Dr Muhammad Farooq Department of Agronomy, University of Agriculture, Faisalabad 38040, Pakistan Tel.: +92 41 9200161-9/2917 Fax: +92 41 9200605 Email: [email protected] Accepted November 12, 2008 doi:10.1111/j.1439-037X.2008.00354.x Abstract Limited water availability hampers the sustainability of crop production. Exog- enous application of glycinebetaine (GB) and salicylic acid (SA) has been found very effective in reducing the adverse effects of water scarcity. This study was conducted to examine the possible role of exogenous GB and SA application in improving the growth and water relations of hybrid sunflower (Helianthus annuus L.) under different irrigation regimes. There were three levels of irriga- tion, viz. control (normal irrigations), water stress at budding stage (irrigation missing at budding stage) and water stress at flowering stage (FS) (irrigation missing at FS). GB and SA were applied exogenously at 100 and 0.724 mm respectively, each at the budding and FS. Control plants did not receive appli- cation of GB and SA. Water stress reduced the leaf area index (LAI), leaf area duration (LAD), crop growth rate (CGR), leaf relative water contents, water potential, osmotic potential, turgor pressure, achene yield and water use efficiency. Nevertheless, exogenous GB and SA application appreciably improved these attributes under water stress. However, exogenous GB applica- tion at the FS was more effective than other treatments. Net assimilation rate was not affected by water stress as well as application of GB and SA. The protein contents were considerably increased by water stress at different growth stages, but were reduced by exogenous GB and SA application. The effects of water stress and foliar application of GB were more pronounced when applied at FS than at the budding stage. Moreover, exogenous GB application was only advantageous under stress conditions. J. Agronomy & Crop Science (2009) ISSN 0931-2250 ª 2009 The Authors 98 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109
Transcript of Exogenous Glycinebetaine and Salicylic Acid Application Improves Water Relations, Allometry and...

DROUGHT STRESS
Exogenous Glycinebetaine and Salicylic Acid ApplicationImproves Water Relations, Allometry and Quality of HybridSunflower under Water Deficit ConditionsM. Hussain1, M. A. Malik1, M. Farooq1, M. B. Khan2, M. Akram1 & M. F. Saleem1
1 Department of Agronomy, University of Agriculture, Faisalabad, Pakistan
2 University College of Agriculture, Bahauddin Zakariya University, Multan, Pakistan
Introduction
Water paucity is a severe environmental constraint to
crop productivity worldwide. Drought-induced loss in
crop yield perhaps exceeds the loss from all other causes,
as both severity and duration of stress are critical (Farooq
et al. 2008a). Drought stress reduces crop growth rate
(CGR) and yield regardless of the growth stage at which
it occurs in arable crops including sunflower (Jensen and
Mogenson 1984).
Under severe water-deficiency, cell elongation of higher
plants can be inhibited by interruption of water flow
from the xylem to the surrounding elongating cells
(Nonami 1998). Impaired mitosis, cell elongation and
expansion result in reduced plant height, leaf area and
crop growth under drought (Nonami 1998, Kaya et al.
2006). Drought stress disturbs the plant water relations,
photosynthesis, assimilate translocation and economic
yield in filed crops (Siddique et al. 2000, Atteya 2003,
Farooq et al. 2008a,b, Hussain et al. 2008a,b). One of the
most common stress tolerance strategies in plants is the
overproduction of different types of compatible organic
solutes (Serraj and Sinclair 2002). Compatible solutes are
of low molecular weight, highly soluble compounds that
are usually non-toxic even at high cytosolic concentra-
tions. Generally, they protect plants from stress through
Keywords
drought stress; exogenous application;
glycinebetaine; growth; salicylic acid;
sunflower; water relations; water use
efficiency
Correspondence
Dr Muhammad Farooq
Department of Agronomy, University of
Agriculture, Faisalabad 38040, Pakistan
Tel.: +92 41 9200161-9/2917
Fax: +92 41 9200605
Email: [email protected]
Accepted November 12, 2008
doi:10.1111/j.1439-037X.2008.00354.x
Abstract
Limited water availability hampers the sustainability of crop production. Exog-
enous application of glycinebetaine (GB) and salicylic acid (SA) has been found
very effective in reducing the adverse effects of water scarcity. This study was
conducted to examine the possible role of exogenous GB and SA application in
improving the growth and water relations of hybrid sunflower (Helianthus
annuus L.) under different irrigation regimes. There were three levels of irriga-
tion, viz. control (normal irrigations), water stress at budding stage (irrigation
missing at budding stage) and water stress at flowering stage (FS) (irrigation
missing at FS). GB and SA were applied exogenously at 100 and 0.724 mm
respectively, each at the budding and FS. Control plants did not receive appli-
cation of GB and SA. Water stress reduced the leaf area index (LAI), leaf area
duration (LAD), crop growth rate (CGR), leaf relative water contents, water
potential, osmotic potential, turgor pressure, achene yield and water use
efficiency. Nevertheless, exogenous GB and SA application appreciably
improved these attributes under water stress. However, exogenous GB applica-
tion at the FS was more effective than other treatments. Net assimilation rate
was not affected by water stress as well as application of GB and SA. The
protein contents were considerably increased by water stress at different growth
stages, but were reduced by exogenous GB and SA application. The effects of
water stress and foliar application of GB were more pronounced when applied
at FS than at the budding stage. Moreover, exogenous GB application was only
advantageous under stress conditions.
J. Agronomy & Crop Science (2009) ISSN 0931-2250
ª 2009 The Authors98 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109

different means such as contribution towards osmotic
adjustment, detoxification of reactive oxygen species,
stabilization of membranes and native structures of
enzymes and proteins (Farooq et al. 2009). Glycinebetaine
(GB) and proline have been the most extensively studied
compatible organic solutes in this regard. Despite some
controversy, many physiological roles have been assigned
to free proline including stabilization of macromolecules,
a sink for excess reductant and a store of carbon and
nitrogen for use after relief of water deficit (Zhu 2002).
Proline contents were increased under drought stress in
pea cultivars (Alexieva et al. 2001).
Many studies demonstrate that GB plays an important
role in enhancing plant tolerance under a range of abiotic
stresses including drought (Quan et al. 2004). The intro-
duction of genes synthesizing GB into non-accumulators
of GB proved to be effective in increasing tolerance to
various abiotic stresses (Sakamoto and Murata 2002).
Naidu et al. (1998) reported that cotton cultivars adapted
to water-stress conditions accumulated higher GB than
the non-adapted ones under drought. In addition to
direct protective roles of GB either through positive
effects on enzyme and membrane integrity or as an
osmoprotectant, GB may protect cells from environmen-
tal stresses indirectly by participating in signal transduc-
tion pathways (Subbarao et al. 2000).
Glycinebetaine has been shown to protect functional
proteins, enzymes (e.g. Rubisco) and lipids of the photo-
synthetic apparatus and to maintain electron flow
through thylakoid membranes (Xing and Rajashekar
1999, Allakhverdiev et al. 2003). Furthermore, even high
concentration of it does not exert adverse effects on
protein structure, enzyme activities, membrane functions
and metabolic processes occurring within the cell (Rhodes
and Hanson 1993). GB-synthesizing transgenic plants can
accumulate GB to a level much lower than natural accu-
mulators, but confer tolerance to various abiotic stresses
including salt, drought, high and low temperatures
(Rhodes and Hanson 1993). It seems that protective role
of GB does not only depend on its osmoprotective role
but also on its other specific physicochemical and physio-
logical effects on the stress responses of plants.
All higher plants are unable to accumulate GB natu-
rally, but some transgenic plants capable of accumulating
GB have been made successfully (Sakamoto and Murata
2002) at lower levels. Exogenous application of GB to
GB-non-accumulators has been taken as an alternative to
improve the stress tolerance. For example, Ma et al.
(2006) reported that GB-treated plants maintained a
higher net photosynthetic rate during drought stress than
non-GB treated. Farooq et al. (2008b) reported that seed
priming with GB can improve the chilling tolerance in
hybrid maize by activation of antioxidant system. In
another study, exogenous GB application successfully
improved the drought tolerance in rice (Farooq et al.
2008a).
Although the role of endogenously produced salicylic
acid (SA) is still inscrutable, salicylic acid potentiates the
generation of reactive oxygen species in photosynthetic
tissues of Arabidopsis thaliana during osmotic stress
(Borsani et al. 2001). There are several reports available
highlighting the role of SA to induce drought stress toler-
ance in wheat (Singh and Usha 2003, Waseem et al. 2006,
Horvath et al. 2007). Recently, Hussain et al. (2008a)
found that exogenous application of SA can improve
yield and yield related traits in sunflower under drought
stress. While in another study, Farooq et al. (2008c)
reported that seed priming with SA can induce chilling
tolerance in hybrid maize mainly by the activation of
antioxidant system.
In view of gradually depleting irrigation water resources
throughout the world, it is highly imperative to investi-
gate the effects and mechanisms of drought in sunflower.
Although the general effects of drought in sunflower
growth are fairly well known, the primary effects of water
deficit on growth and water relations in sunflower are not
well-understood. It is hypothesized that exogenous GB
and SA application may alleviate the adversities of
drought stress, maintain plant tissue water status and
promote growth and allometric responses under drought
stress.
Materials and Methods
The study was conducted at the Agronomic Research
Area, University of Agriculture, Faisalabad (31.25�N,
73.09�E and 183 m a. s. l.), during two consecutive years
viz. 2006 and 2007. Experimental soil was well drained,
sandy clay loam, with pH 7.8, 0.32 mS cm)1 total
exchangeable salts and 0.75 % organic matter and the soil
belongs to Lyallpur soil series [aridisol-fine-silty, mixed,
hyperthermic Ustalfic, Haplarged in the United States
Department of Agriculture (USDA) classification and
Hyplic Yermosols in the Food and Agriculture Organiza-
tion (FAO) classification].
Sunflower hybrid Hysun-33 was used as experimental
material. The experiment was laid out in randomized
complete block design with split plot arrangement and
was replicated three times with net plot size of
3 m · 5 m. Irrigation levels and GB and SA applications
were randomized in main plots and sub-plots respec-
tively.
Irrigation levels included in the study were standard
[CKI = normal irrigations (no water stress), vegetative
stage (VS) = water stress at budding stage (irrigation
missing at budding stage) and FS = water stress at flower-
Sunflower under Water Deficit Conditions
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109 99
ing stage (irrigation missing at FS)]. GB was applied at
100 mm each at budding (VGB) and FSs (FGB), while SA
was applied at 0.724 mm each at budding (VSA) and
flowering stage (FSA). In control plots (CKF), neither GB
nor SA was applied. The measured amount of water was
applied using cut-throat flume according to following
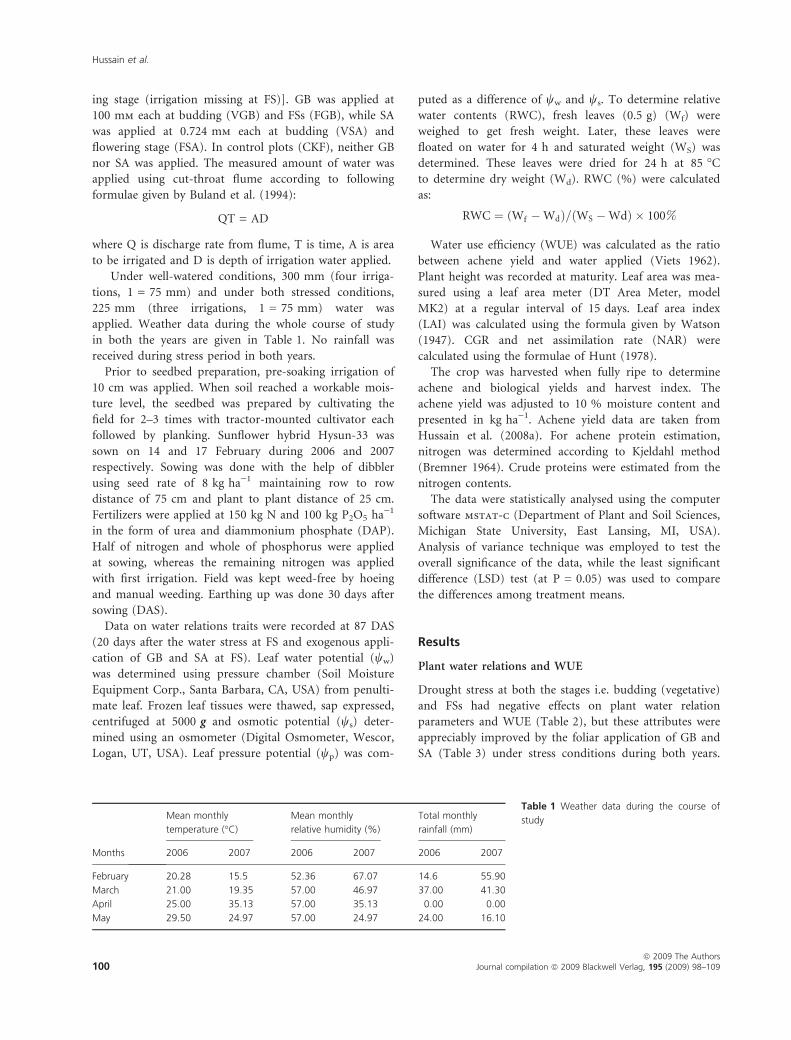
formulae given by Buland et al. (1994):
QT = AD
where Q is discharge rate from flume, T is time, A is area
to be irrigated and D is depth of irrigation water applied.
Under well-watered conditions, 300 mm (four irriga-
tions, 1 = 75 mm) and under both stressed conditions,
225 mm (three irrigations, 1 = 75 mm) water was
applied. Weather data during the whole course of study
in both the years are given in Table 1. No rainfall was
received during stress period in both years.
Prior to seedbed preparation, pre-soaking irrigation of
10 cm was applied. When soil reached a workable mois-
ture level, the seedbed was prepared by cultivating the
field for 2–3 times with tractor-mounted cultivator each
followed by planking. Sunflower hybrid Hysun-33 was
sown on 14 and 17 February during 2006 and 2007
respectively. Sowing was done with the help of dibbler
using seed rate of 8 kg ha)1 maintaining row to row
distance of 75 cm and plant to plant distance of 25 cm.
Fertilizers were applied at 150 kg N and 100 kg P2O5 ha)1
in the form of urea and diammonium phosphate (DAP).
Half of nitrogen and whole of phosphorus were applied
at sowing, whereas the remaining nitrogen was applied
with first irrigation. Field was kept weed-free by hoeing
and manual weeding. Earthing up was done 30 days after
sowing (DAS).
Data on water relations traits were recorded at 87 DAS
(20 days after the water stress at FS and exogenous appli-
cation of GB and SA at FS). Leaf water potential (ww)
was determined using pressure chamber (Soil Moisture
Equipment Corp., Santa Barbara, CA, USA) from penulti-
mate leaf. Frozen leaf tissues were thawed, sap expressed,
centrifuged at 5000 g and osmotic potential (ws) deter-
mined using an osmometer (Digital Osmometer, Wescor,
Logan, UT, USA). Leaf pressure potential (wp) was com-
puted as a difference of ww and ws. To determine relative
water contents (RWC), fresh leaves (0.5 g) (Wf) were
weighed to get fresh weight. Later, these leaves were
floated on water for 4 h and saturated weight (WS) was
determined. These leaves were dried for 24 h at 85 �C
to determine dry weight (Wd). RWC (%) were calculated
as:
RWC ¼ (Wf �WdÞ=ðWS �WdÞ � 100%
Water use efficiency (WUE) was calculated as the ratio
between achene yield and water applied (Viets 1962).
Plant height was recorded at maturity. Leaf area was mea-
sured using a leaf area meter (DT Area Meter, model
MK2) at a regular interval of 15 days. Leaf area index
(LAI) was calculated using the formula given by Watson
(1947). CGR and net assimilation rate (NAR) were
calculated using the formulae of Hunt (1978).
The crop was harvested when fully ripe to determine
achene and biological yields and harvest index. The
achene yield was adjusted to 10 % moisture content and
presented in kg ha)1. Achene yield data are taken from
Hussain et al. (2008a). For achene protein estimation,
nitrogen was determined according to Kjeldahl method
(Bremner 1964). Crude proteins were estimated from the
nitrogen contents.
The data were statistically analysed using the computer
software mstat-c (Department of Plant and Soil Sciences,
Michigan State University, East Lansing, MI, USA).
Analysis of variance technique was employed to test the
overall significance of the data, while the least significant
difference (LSD) test (at P = 0.05) was used to compare
the differences among treatment means.
Results
Plant water relations and WUE
Drought stress at both the stages i.e. budding (vegetative)
and FSs had negative effects on plant water relation
parameters and WUE (Table 2), but these attributes were
appreciably improved by the foliar application of GB and
SA (Table 3) under stress conditions during both years.
Table 1 Weather data during the course of
study
Months
Mean monthly
temperature (�C)
Mean monthly
relative humidity (%)
Total monthly
rainfall (mm)
2006 2007 2006 2007 2006 2007
February 20.28 15.5 52.36 67.07 14.6 55.90
March 21.00 19.35 57.00 46.97 37.00 41.30
April 25.00 35.13 57.00 35.13 0.00 0.00
May 29.50 24.97 57.00 24.97 24.00 16.10
Hussain et al.
ª 2009 The Authors100 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109
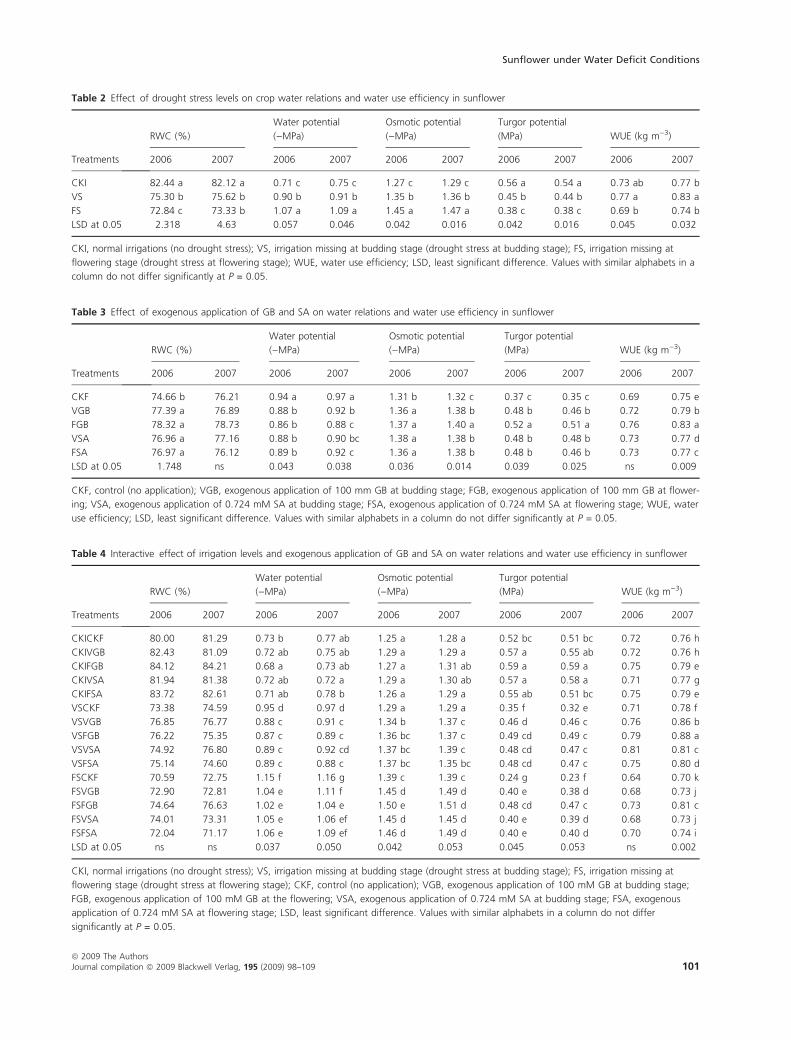
Table 2 Effect of drought stress levels on crop water relations and water use efficiency in sunflower
Treatments
RWC (%)
Water potential
()MPa)
Osmotic potential
()MPa)
Turgor potential
(MPa) WUE (kg m)3)
2006 2007 2006 2007 2006 2007 2006 2007 2006 2007
CKI 82.44 a 82.12 a 0.71 c 0.75 c 1.27 c 1.29 c 0.56 a 0.54 a 0.73 ab 0.77 b
VS 75.30 b 75.62 b 0.90 b 0.91 b 1.35 b 1.36 b 0.45 b 0.44 b 0.77 a 0.83 a
FS 72.84 c 73.33 b 1.07 a 1.09 a 1.45 a 1.47 a 0.38 c 0.38 c 0.69 b 0.74 b
LSD at 0.05 2.318 4.63 0.057 0.046 0.042 0.016 0.042 0.016 0.045 0.032
CKI, normal irrigations (no drought stress); VS, irrigation missing at budding stage (drought stress at budding stage); FS, irrigation missing at
flowering stage (drought stress at flowering stage); WUE, water use efficiency; LSD, least significant difference. Values with similar alphabets in a
column do not differ significantly at P = 0.05.
Table 3 Effect of exogenous application of GB and SA on water relations and water use efficiency in sunflower
Treatments
RWC (%)
Water potential
()MPa)
Osmotic potential
()MPa)
Turgor potential
(MPa) WUE (kg m)3)
2006 2007 2006 2007 2006 2007 2006 2007 2006 2007
CKF 74.66 b 76.21 0.94 a 0.97 a 1.31 b 1.32 c 0.37 c 0.35 c 0.69 0.75 e
VGB 77.39 a 76.89 0.88 b 0.92 b 1.36 a 1.38 b 0.48 b 0.46 b 0.72 0.79 b
FGB 78.32 a 78.73 0.86 b 0.88 c 1.37 a 1.40 a 0.52 a 0.51 a 0.76 0.83 a
VSA 76.96 a 77.16 0.88 b 0.90 bc 1.38 a 1.38 b 0.48 b 0.48 b 0.73 0.77 d
FSA 76.97 a 76.12 0.89 b 0.92 c 1.36 a 1.38 b 0.48 b 0.46 b 0.73 0.77 c
LSD at 0.05 1.748 ns 0.043 0.038 0.036 0.014 0.039 0.025 ns 0.009
CKF, control (no application); VGB, exogenous application of 100 mm GB at budding stage; FGB, exogenous application of 100 mm GB at flower-
ing; VSA, exogenous application of 0.724 mM SA at budding stage; FSA, exogenous application of 0.724 mM SA at flowering stage; WUE, water
use efficiency; LSD, least significant difference. Values with similar alphabets in a column do not differ significantly at P = 0.05.
Table 4 Interactive effect of irrigation levels and exogenous application of GB and SA on water relations and water use efficiency in sunflower
Treatments
RWC (%)
Water potential
()MPa)
Osmotic potential
()MPa)
Turgor potential
(MPa) WUE (kg m)3)
2006 2007 2006 2007 2006 2007 2006 2007 2006 2007
CKICKF 80.00 81.29 0.73 b 0.77 ab 1.25 a 1.28 a 0.52 bc 0.51 bc 0.72 0.76 h
CKIVGB 82.43 81.09 0.72 ab 0.75 ab 1.29 a 1.29 a 0.57 a 0.55 ab 0.72 0.76 h
CKIFGB 84.12 84.21 0.68 a 0.73 ab 1.27 a 1.31 ab 0.59 a 0.59 a 0.75 0.79 e
CKIVSA 81.94 81.38 0.72 ab 0.72 a 1.29 a 1.30 ab 0.57 a 0.58 a 0.71 0.77 g
CKIFSA 83.72 82.61 0.71 ab 0.78 b 1.26 a 1.29 a 0.55 ab 0.51 bc 0.75 0.79 e
VSCKF 73.38 74.59 0.95 d 0.97 d 1.29 a 1.29 a 0.35 f 0.32 e 0.71 0.78 f
VSVGB 76.85 76.77 0.88 c 0.91 c 1.34 b 1.37 c 0.46 d 0.46 c 0.76 0.86 b
VSFGB 76.22 75.35 0.87 c 0.89 c 1.36 bc 1.37 c 0.49 cd 0.49 c 0.79 0.88 a
VSVSA 74.92 76.80 0.89 c 0.92 cd 1.37 bc 1.39 c 0.48 cd 0.47 c 0.81 0.81 c
VSFSA 75.14 74.60 0.89 c 0.88 c 1.37 bc 1.35 bc 0.48 cd 0.47 c 0.75 0.80 d
FSCKF 70.59 72.75 1.15 f 1.16 g 1.39 c 1.39 c 0.24 g 0.23 f 0.64 0.70 k
FSVGB 72.90 72.81 1.04 e 1.11 f 1.45 d 1.49 d 0.40 e 0.38 d 0.68 0.73 j
FSFGB 74.64 76.63 1.02 e 1.04 e 1.50 e 1.51 d 0.48 cd 0.47 c 0.73 0.81 c
FSVSA 74.01 73.31 1.05 e 1.06 ef 1.45 d 1.45 d 0.40 e 0.39 d 0.68 0.73 j
FSFSA 72.04 71.17 1.06 e 1.09 ef 1.46 d 1.49 d 0.40 e 0.40 d 0.70 0.74 i
LSD at 0.05 ns ns 0.037 0.050 0.042 0.053 0.045 0.053 ns 0.002
CKI, normal irrigations (no drought stress); VS, irrigation missing at budding stage (drought stress at budding stage); FS, irrigation missing at
flowering stage (drought stress at flowering stage); CKF, control (no application); VGB, exogenous application of 100 mM GB at budding stage;
FGB, exogenous application of 100 mM GB at the flowering; VSA, exogenous application of 0.724 mM SA at budding stage; FSA, exogenous
application of 0.724 mM SA at flowering stage; LSD, least significant difference. Values with similar alphabets in a column do not differ
significantly at P = 0.05.
Sunflower under Water Deficit Conditions
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109 101
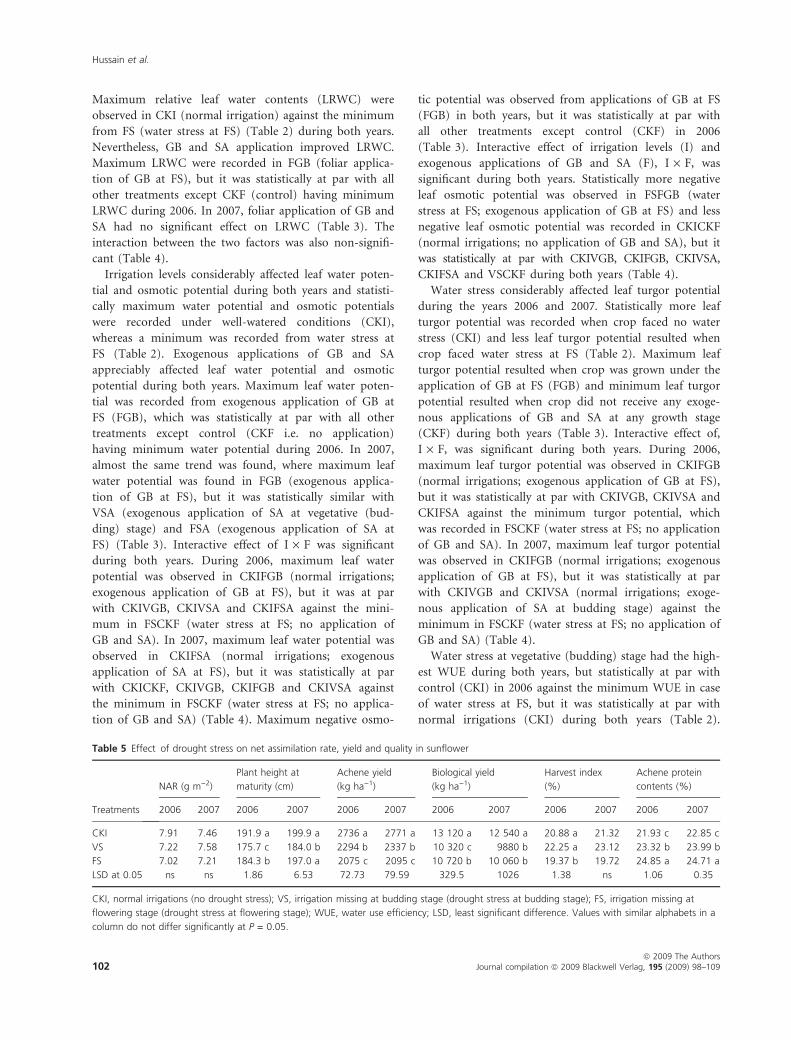
Maximum relative leaf water contents (LRWC) were
observed in CKI (normal irrigation) against the minimum
from FS (water stress at FS) (Table 2) during both years.
Nevertheless, GB and SA application improved LRWC.
Maximum LRWC were recorded in FGB (foliar applica-
tion of GB at FS), but it was statistically at par with all
other treatments except CKF (control) having minimum
LRWC during 2006. In 2007, foliar application of GB and
SA had no significant effect on LRWC (Table 3). The
interaction between the two factors was also non-signifi-
cant (Table 4).
Irrigation levels considerably affected leaf water poten-
tial and osmotic potential during both years and statisti-
cally maximum water potential and osmotic potentials
were recorded under well-watered conditions (CKI),
whereas a minimum was recorded from water stress at
FS (Table 2). Exogenous applications of GB and SA
appreciably affected leaf water potential and osmotic
potential during both years. Maximum leaf water poten-
tial was recorded from exogenous application of GB at
FS (FGB), which was statistically at par with all other
treatments except control (CKF i.e. no application)
having minimum water potential during 2006. In 2007,
almost the same trend was found, where maximum leaf
water potential was found in FGB (exogenous applica-
tion of GB at FS), but it was statistically similar with
VSA (exogenous application of SA at vegetative (bud-
ding) stage) and FSA (exogenous application of SA at
FS) (Table 3). Interactive effect of I · F was significant
during both years. During 2006, maximum leaf water
potential was observed in CKIFGB (normal irrigations;
exogenous application of GB at FS), but it was at par
with CKIVGB, CKIVSA and CKIFSA against the mini-
mum in FSCKF (water stress at FS; no application of
GB and SA). In 2007, maximum leaf water potential was
observed in CKIFSA (normal irrigations; exogenous
application of SA at FS), but it was statistically at par
with CKICKF, CKIVGB, CKIFGB and CKIVSA against
the minimum in FSCKF (water stress at FS; no applica-
tion of GB and SA) (Table 4). Maximum negative osmo-
tic potential was observed from applications of GB at FS
(FGB) in both years, but it was statistically at par with
all other treatments except control (CKF) in 2006
(Table 3). Interactive effect of irrigation levels (I) and
exogenous applications of GB and SA (F), I · F, was
significant during both years. Statistically more negative
leaf osmotic potential was observed in FSFGB (water
stress at FS; exogenous application of GB at FS) and less
negative leaf osmotic potential was recorded in CKICKF
(normal irrigations; no application of GB and SA), but it
was statistically at par with CKIVGB, CKIFGB, CKIVSA,
CKIFSA and VSCKF during both years (Table 4).
Water stress considerably affected leaf turgor potential
during the years 2006 and 2007. Statistically more leaf
turgor potential was recorded when crop faced no water
stress (CKI) and less leaf turgor potential resulted when
crop faced water stress at FS (Table 2). Maximum leaf
turgor potential resulted when crop was grown under the
application of GB at FS (FGB) and minimum leaf turgor
potential resulted when crop did not receive any exoge-
nous applications of GB and SA at any growth stage
(CKF) during both years (Table 3). Interactive effect of,
I · F, was significant during both years. During 2006,
maximum leaf turgor potential was observed in CKIFGB
(normal irrigations; exogenous application of GB at FS),
but it was statistically at par with CKIVGB, CKIVSA and
CKIFSA against the minimum turgor potential, which
was recorded in FSCKF (water stress at FS; no application
of GB and SA). In 2007, maximum leaf turgor potential
was observed in CKIFGB (normal irrigations; exogenous
application of GB at FS), but it was statistically at par
with CKIVGB and CKIVSA (normal irrigations; exoge-
nous application of SA at budding stage) against the
minimum in FSCKF (water stress at FS; no application of
GB and SA) (Table 4).
Water stress at vegetative (budding) stage had the high-
est WUE during both years, but statistically at par with
control (CKI) in 2006 against the minimum WUE in case
of water stress at FS, but it was statistically at par with
normal irrigations (CKI) during both years (Table 2).
Table 5 Effect of drought stress on net assimilation rate, yield and quality in sunflower
Treatments
NAR (g m)2)
Plant height at
maturity (cm)
Achene yield
(kg ha)1)
Biological yield
(kg ha)1)
Harvest index
(%)
Achene protein
contents (%)
2006 2007 2006 2007 2006 2007 2006 2007 2006 2007 2006 2007
CKI 7.91 7.46 191.9 a 199.9 a 2736 a 2771 a 13 120 a 12 540 a 20.88 a 21.32 21.93 c 22.85 c
VS 7.22 7.58 175.7 c 184.0 b 2294 b 2337 b 10 320 c 9880 b 22.25 a 23.12 23.32 b 23.99 b
FS 7.02 7.21 184.3 b 197.0 a 2075 c 2095 c 10 720 b 10 060 b 19.37 b 19.72 24.85 a 24.71 a
LSD at 0.05 ns ns 1.86 6.53 72.73 79.59 329.5 1026 1.38 ns 1.06 0.35
CKI, normal irrigations (no drought stress); VS, irrigation missing at budding stage (drought stress at budding stage); FS, irrigation missing at
flowering stage (drought stress at flowering stage); WUE, water use efficiency; LSD, least significant difference. Values with similar alphabets in a
column do not differ significantly at P = 0.05.
Hussain et al.
ª 2009 The Authors102 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109
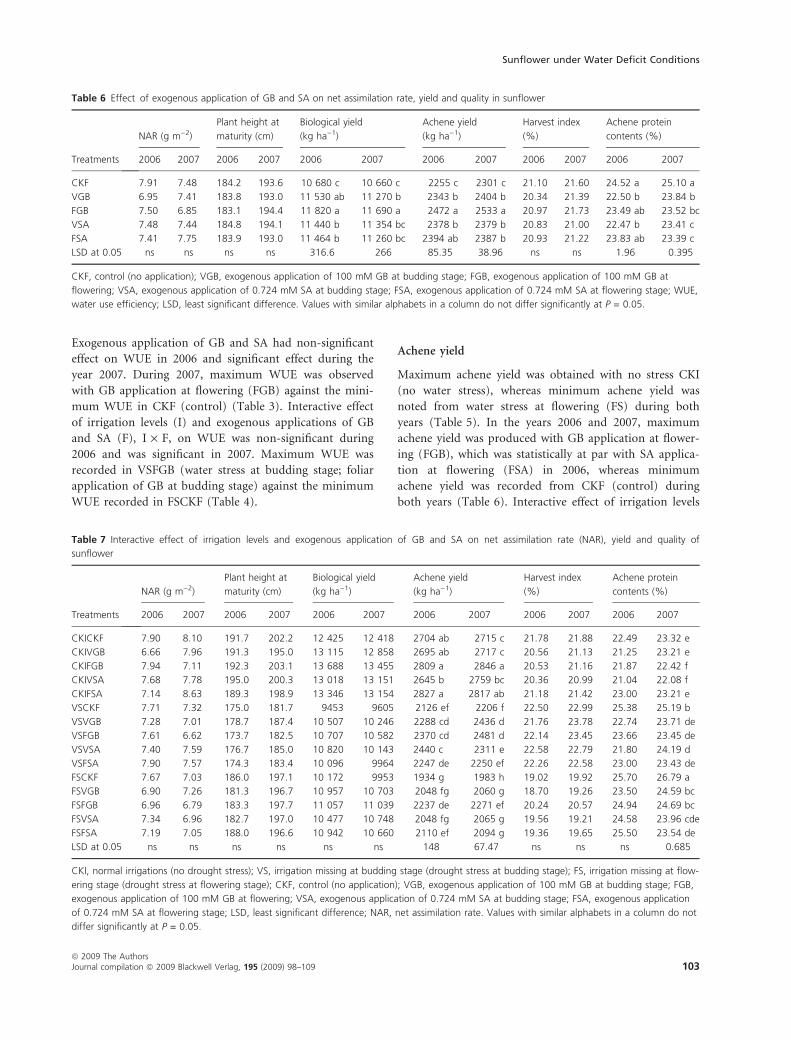
Exogenous application of GB and SA had non-significant
effect on WUE in 2006 and significant effect during the
year 2007. During 2007, maximum WUE was observed
with GB application at flowering (FGB) against the mini-
mum WUE in CKF (control) (Table 3). Interactive effect
of irrigation levels (I) and exogenous applications of GB
and SA (F), I · F, on WUE was non-significant during
2006 and was significant in 2007. Maximum WUE was
recorded in VSFGB (water stress at budding stage; foliar
application of GB at budding stage) against the minimum
WUE recorded in FSCKF (Table 4).
Achene yield
Maximum achene yield was obtained with no stress CKI
(no water stress), whereas minimum achene yield was
noted from water stress at flowering (FS) during both
years (Table 5). In the years 2006 and 2007, maximum
achene yield was produced with GB application at flower-
ing (FGB), which was statistically at par with SA applica-
tion at flowering (FSA) in 2006, whereas minimum
achene yield was recorded from CKF (control) during
both years (Table 6). Interactive effect of irrigation levels
Table 6 Effect of exogenous application of GB and SA on net assimilation rate, yield and quality in sunflower
Treatments
NAR (g m)2)
Plant height at
maturity (cm)
Biological yield
(kg ha)1)
Achene yield
(kg ha)1)
Harvest index
(%)
Achene protein
contents (%)
2006 2007 2006 2007 2006 2007 2006 2007 2006 2007 2006 2007
CKF 7.91 7.48 184.2 193.6 10 680 c 10 660 c 2255 c 2301 c 21.10 21.60 24.52 a 25.10 a
VGB 6.95 7.41 183.8 193.0 11 530 ab 11 270 b 2343 b 2404 b 20.34 21.39 22.50 b 23.84 b
FGB 7.50 6.85 183.1 194.4 11 820 a 11 690 a 2472 a 2533 a 20.97 21.73 23.49 ab 23.52 bc
VSA 7.48 7.44 184.8 194.1 11 440 b 11 354 bc 2378 b 2379 b 20.83 21.00 22.47 b 23.41 c
FSA 7.41 7.75 183.9 193.0 11 464 b 11 260 bc 2394 ab 2387 b 20.93 21.22 23.83 ab 23.39 c
LSD at 0.05 ns ns ns ns 316.6 266 85.35 38.96 ns ns 1.96 0.395
CKF, control (no application); VGB, exogenous application of 100 mM GB at budding stage; FGB, exogenous application of 100 mM GB at
flowering; VSA, exogenous application of 0.724 mM SA at budding stage; FSA, exogenous application of 0.724 mM SA at flowering stage; WUE,
water use efficiency; LSD, least significant difference. Values with similar alphabets in a column do not differ significantly at P = 0.05.
Table 7 Interactive effect of irrigation levels and exogenous application of GB and SA on net assimilation rate (NAR), yield and quality of
sunflower
Treatments
NAR (g m)2)
Plant height at
maturity (cm)
Biological yield
(kg ha)1)
Achene yield
(kg ha)1)
Harvest index
(%)
Achene protein
contents (%)
2006 2007 2006 2007 2006 2007 2006 2007 2006 2007 2006 2007
CKICKF 7.90 8.10 191.7 202.2 12 425 12 418 2704 ab 2715 c 21.78 21.88 22.49 23.32 e
CKIVGB 6.66 7.96 191.3 195.0 13 115 12 858 2695 ab 2717 c 20.56 21.13 21.25 23.21 e
CKIFGB 7.94 7.11 192.3 203.1 13 688 13 455 2809 a 2846 a 20.53 21.16 21.87 22.42 f
CKIVSA 7.68 7.78 195.0 200.3 13 018 13 151 2645 b 2759 bc 20.36 20.99 21.04 22.08 f
CKIFSA 7.14 8.63 189.3 198.9 13 346 13 154 2827 a 2817 ab 21.18 21.42 23.00 23.21 e
VSCKF 7.71 7.32 175.0 181.7 9453 9605 2126 ef 2206 f 22.50 22.99 25.38 25.19 b
VSVGB 7.28 7.01 178.7 187.4 10 507 10 246 2288 cd 2436 d 21.76 23.78 22.74 23.71 de
VSFGB 7.61 6.62 173.7 182.5 10 707 10 582 2370 cd 2481 d 22.14 23.45 23.66 23.45 de
VSVSA 7.40 7.59 176.7 185.0 10 820 10 143 2440 c 2311 e 22.58 22.79 21.80 24.19 d
VSFSA 7.90 7.57 174.3 183.4 10 096 9964 2247 de 2250 ef 22.26 22.58 23.00 23.43 de
FSCKF 7.67 7.03 186.0 197.1 10 172 9953 1934 g 1983 h 19.02 19.92 25.70 26.79 a
FSVGB 6.90 7.26 181.3 196.7 10 957 10 703 2048 fg 2060 g 18.70 19.26 23.50 24.59 bc
FSFGB 6.96 6.79 183.3 197.7 11 057 11 039 2237 de 2271 ef 20.24 20.57 24.94 24.69 bc
FSVSA 7.34 6.96 182.7 197.0 10 477 10 748 2048 fg 2065 g 19.56 19.21 24.58 23.96 cde
FSFSA 7.19 7.05 188.0 196.6 10 942 10 660 2110 ef 2094 g 19.36 19.65 25.50 23.54 de
LSD at 0.05 ns ns ns ns ns ns 148 67.47 ns ns ns 0.685
CKI, normal irrigations (no drought stress); VS, irrigation missing at budding stage (drought stress at budding stage); FS, irrigation missing at flow-
ering stage (drought stress at flowering stage); CKF, control (no application); VGB, exogenous application of 100 mM GB at budding stage; FGB,
exogenous application of 100 mM GB at flowering; VSA, exogenous application of 0.724 mM SA at budding stage; FSA, exogenous application
of 0.724 mM SA at flowering stage; LSD, least significant difference; NAR, net assimilation rate. Values with similar alphabets in a column do not
differ significantly at P = 0.05.
Sunflower under Water Deficit Conditions
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109 103
(I) and exogenous applications of GB and SA (F) was sig-
nificant during both years. During 2006, maximum
achene yield was obtained in CKIFGB (normal irrigations;
exogenous application of GB at FS), but it was statistically
at par with CKICKF, CKIVGB and CKIFSA against the
minimum achene yield from FSCKF (water stress at FS;
no application of GB and SA), which was at par with
FSVGB and FSVSA. During 2007, maximum achene yield
was obtained in CKIFGB (normal irrigations; exogenous
application of GB at FS), which was at par with CKIFSA
against the minimum achene yield from FSCKI (water
stress at FS; no application of GB and SA) (Table 7).
Plant height
Water stress treatments had a significant effect on plant
height. Maximum plant height was recorded from well-
watered control (CKI) during both years; however, it was
at par with water stress at flowering (FS) during 2007,
whereas the minimum plant height was recorded from
water stress at vegetative (budding) stage (VS) during
both years (Table 5). Exogenous applications of GB and
SA did not affect the plant height appreciably (Table 6).
Furthermore, interactive effect of irrigation levels (I) and
exogenous applications of GB and SA (F), I · F, was not
significant during the years 2006 and 2007 (Table 7).
Biological yield
Maximum biological yield was produced when crop was
grown with normal irrigations (CKI). However, minimum
biological yield was harvested from water stress at VS,
which was at par with FS (water stress at FS) in 2007
(Table 5). In 2006, maximum biological yield was
produced by FGB (exogenous application of GB at FS)
that was similar with VGB against the minimum in con-
trol (CKF). Likewise, in 2007, maximum biological yield
was produced in FGB (exogenous application of GB at
FS) treatment against the minimum in control (CKF),
which was statistically at par with VSA and FSA (exoge-
nous application of SA at budding and FS respectively)
(Table 6). Interactive effect of irrigation levels (I) and
exogenous applications of GB and SA (F), I · F, on
biological yield was non-significant during both years
(Table 7).
Harvest index
Irrigation levels had a significant effect on harvest index
during 2006 and non-significant effect during 2007. In
2006, maximum harvest index was observed from
drought stress at VS, which was similar with normal
irrigation (no water stress) and the lowest harvest index
was recorded from water stress at flowering (FS)
(Table 5). Exogenous applications of GB and SA at differ-
ent growth stages and interactive effect of irrigation levels
(I) and exogenous applications of GB and SA (F), I · F,
had non-significant effect on harvest index during both
years (Tables 6 and 7).
Alleometry
Leaf area index progressively increased and reached the
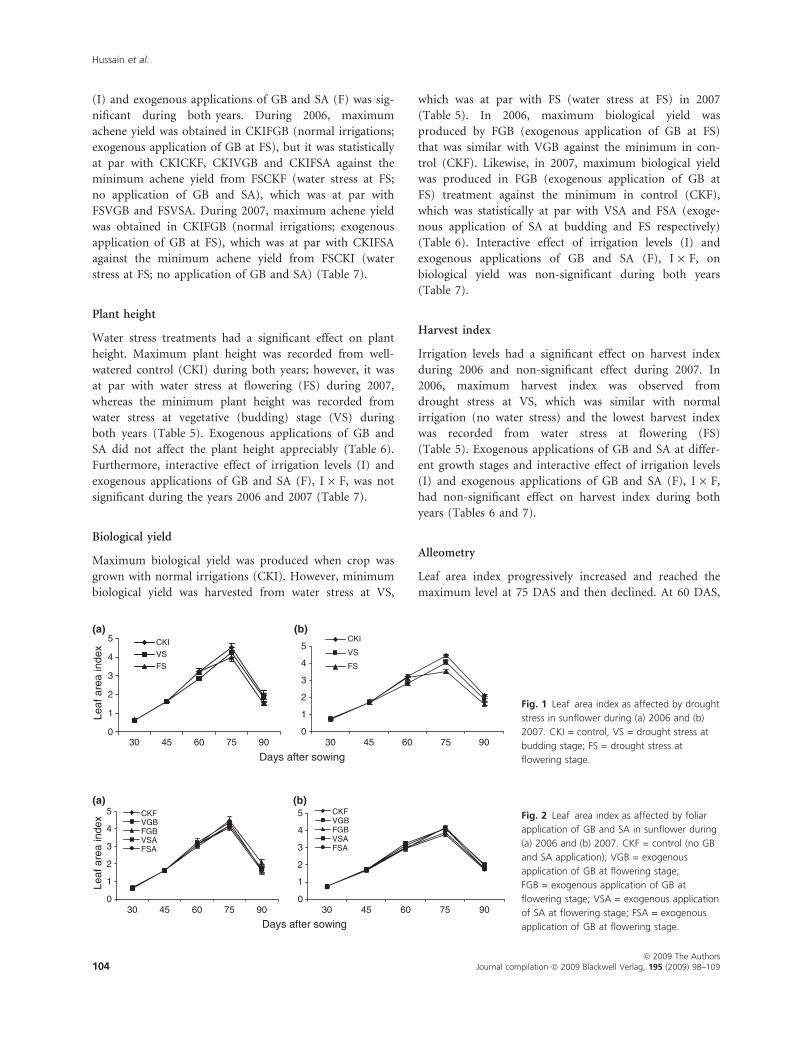
maximum level at 75 DAS and then declined. At 60 DAS,
Leaf
are
a in
dex
0
1
2
3
4
5
30 45 60 75 90
CKI
VS
FS
0
1
2
3
4
5
30 45 60 75 90
CKI(b)(a)
VS
FS
Days after sowing
Fig. 1 Leaf area index as affected by drought
stress in sunflower during (a) 2006 and (b)
2007. CKI = control, VS = drought stress at
budding stage; FS = drought stress at
flowering stage.
Leaf
are
a in
dex
0
1
2
3
4
5
30 45 60 75 90
CKF
(a) (b)
VGB FGB VSA FSA
0
1
2
3
4
5
30 45 60 75 90
CKF VGB FGB VSA FSA
Days after sowing
Fig. 2 Leaf area index as affected by foliar
application of GB and SA in sunflower during
(a) 2006 and (b) 2007. CKF = control (no GB
and SA application); VGB = exogenous
application of GB at flowering stage;
FGB = exogenous application of GB at
flowering stage; VSA = exogenous application
of SA at flowering stage; FSA = exogenous
application of GB at flowering stage.
Hussain et al.
ª 2009 The Authors104 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109
normal irrigation (CKI) and water stress at flowering (FS)
had a higher LAI compared with water stress at budding
stage (VS). At 75 and 90 DAS, CKI (no water stress) had
maximum LAI against the minimum LAI in case of water
stress at flowering (FS) during the years 2006 and 2007
(Fig. 1). At 60 DAS, foliar application of GB at vegetative
(budding) stage (VGB) and at 75 DAS, foliar application
of GB at FS (FGB) had high a LAI against the minimum
LAI from CKF (no application) for both years (Fig. 2).
Leaf area duration (LAD) progressively increased with
increase in growth period (Fig. 3). Drought stress severely
affected total LAD of the crop, whereas higher LAD was
noted when the crop faced no water stress (CKI) against
the minimum LAD that resulted when crop faced water
stress at FS during both years, which was at par with the
LAD from water stress at budding stage (VS) during 2006
(Fig. 3). Nonetheless, exogenous applications of GB and
SA improved the LAD. Maximum LAD was observed
from exogenous application of GB at flowering (FGB),
but it was at par with the LAD from foliar application of
GB at VS (VGB). During 2007, maximum LAD was
observed from exogenous application of GB at FS (F2)
against the minimum from no foliar application, which
was at par with the LAD from application of SA at FS
(F4) (Fig. 4).
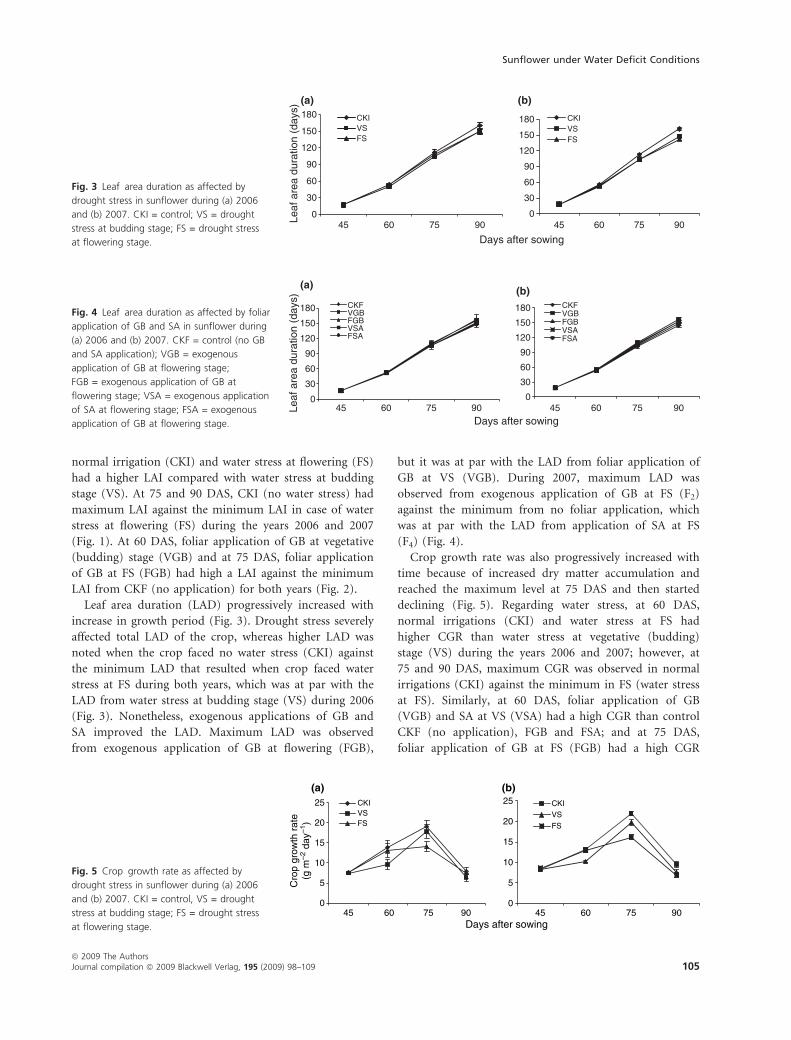
Crop growth rate was also progressively increased with
time because of increased dry matter accumulation and
reached the maximum level at 75 DAS and then started
declining (Fig. 5). Regarding water stress, at 60 DAS,
normal irrigations (CKI) and water stress at FS had
higher CGR than water stress at vegetative (budding)
stage (VS) during the years 2006 and 2007; however, at
75 and 90 DAS, maximum CGR was observed in normal
irrigations (CKI) against the minimum in FS (water stress
at FS). Similarly, at 60 DAS, foliar application of GB
(VGB) and SA at VS (VSA) had a high CGR than control
CKF (no application), FGB and FSA; and at 75 DAS,
foliar application of GB at FS (FGB) had a high CGR
Leaf
are
a du
ratio
n (d
ays)
0
30
60
90
120
150
180
(a) (b)
0
30
60
90
120
150
180
45 60 75 90
CKI VS FS
45 60 75 90
CKI VS FS
Days after sowing
Fig. 3 Leaf area duration as affected by
drought stress in sunflower during (a) 2006
and (b) 2007. CKI = control; VS = drought
stress at budding stage; FS = drought stress
at flowering stage.
Leaf
are
a du
ratio
n (d
ays)
45 60 75 90
CKF VGB FGB VSA FSA
0
30
60
90
120
150
180
0
30
60
90
120
150
180
(a) (b)
45 60 75 90
CKF VGB FGB VSA FSA
Days after sowing
Fig. 4 Leaf area duration as affected by foliar
application of GB and SA in sunflower during
(a) 2006 and (b) 2007. CKF = control (no GB
and SA application); VGB = exogenous
application of GB at flowering stage;
FGB = exogenous application of GB at
flowering stage; VSA = exogenous application
of SA at flowering stage; FSA = exogenous
application of GB at flowering stage.
Cro
p gr
owth
rat
e(g
m–2
day
–1)
0
5
10
15
20
25
45 60 75 90
CKI
(a) (b)
VSFS
0
5
10
15
20
25
45 60 75 90
CKIVS
FS
Days after sowing
Fig. 5 Crop growth rate as affected by
drought stress in sunflower during (a) 2006
and (b) 2007. CKI = control, VS = drought
stress at budding stage; FS = drought stress
at flowering stage.
Sunflower under Water Deficit Conditions
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109 105
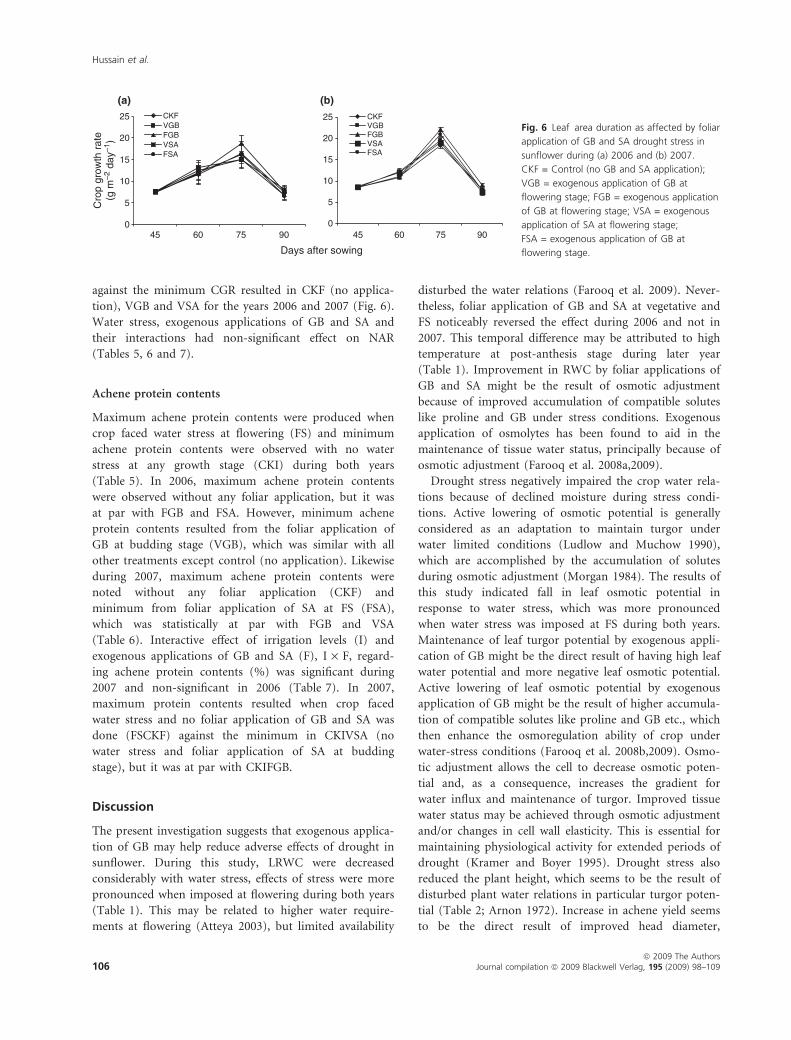
against the minimum CGR resulted in CKF (no applica-
tion), VGB and VSA for the years 2006 and 2007 (Fig. 6).
Water stress, exogenous applications of GB and SA and
their interactions had non-significant effect on NAR
(Tables 5, 6 and 7).
Achene protein contents
Maximum achene protein contents were produced when
crop faced water stress at flowering (FS) and minimum
achene protein contents were observed with no water
stress at any growth stage (CKI) during both years
(Table 5). In 2006, maximum achene protein contents
were observed without any foliar application, but it was
at par with FGB and FSA. However, minimum achene
protein contents resulted from the foliar application of
GB at budding stage (VGB), which was similar with all
other treatments except control (no application). Likewise
during 2007, maximum achene protein contents were
noted without any foliar application (CKF) and
minimum from foliar application of SA at FS (FSA),
which was statistically at par with FGB and VSA
(Table 6). Interactive effect of irrigation levels (I) and
exogenous applications of GB and SA (F), I · F, regard-
ing achene protein contents (%) was significant during
2007 and non-significant in 2006 (Table 7). In 2007,
maximum protein contents resulted when crop faced
water stress and no foliar application of GB and SA was
done (FSCKF) against the minimum in CKIVSA (no
water stress and foliar application of SA at budding
stage), but it was at par with CKIFGB.
Discussion
The present investigation suggests that exogenous applica-
tion of GB may help reduce adverse effects of drought in
sunflower. During this study, LRWC were decreased
considerably with water stress, effects of stress were more
pronounced when imposed at flowering during both years
(Table 1). This may be related to higher water require-
ments at flowering (Atteya 2003), but limited availability
disturbed the water relations (Farooq et al. 2009). Never-
theless, foliar application of GB and SA at vegetative and
FS noticeably reversed the effect during 2006 and not in
2007. This temporal difference may be attributed to high
temperature at post-anthesis stage during later year
(Table 1). Improvement in RWC by foliar applications of
GB and SA might be the result of osmotic adjustment
because of improved accumulation of compatible solutes
like proline and GB under stress conditions. Exogenous
application of osmolytes has been found to aid in the
maintenance of tissue water status, principally because of
osmotic adjustment (Farooq et al. 2008a,2009).
Drought stress negatively impaired the crop water rela-
tions because of declined moisture during stress condi-
tions. Active lowering of osmotic potential is generally
considered as an adaptation to maintain turgor under
water limited conditions (Ludlow and Muchow 1990),
which are accomplished by the accumulation of solutes
during osmotic adjustment (Morgan 1984). The results of
this study indicated fall in leaf osmotic potential in
response to water stress, which was more pronounced
when water stress was imposed at FS during both years.
Maintenance of leaf turgor potential by exogenous appli-
cation of GB might be the direct result of having high leaf
water potential and more negative leaf osmotic potential.
Active lowering of leaf osmotic potential by exogenous
application of GB might be the result of higher accumula-
tion of compatible solutes like proline and GB etc., which
then enhance the osmoregulation ability of crop under
water-stress conditions (Farooq et al. 2008b,2009). Osmo-
tic adjustment allows the cell to decrease osmotic poten-
tial and, as a consequence, increases the gradient for
water influx and maintenance of turgor. Improved tissue
water status may be achieved through osmotic adjustment
and/or changes in cell wall elasticity. This is essential for
maintaining physiological activity for extended periods of
drought (Kramer and Boyer 1995). Drought stress also
reduced the plant height, which seems to be the result of
disturbed plant water relations in particular turgor poten-
tial (Table 2; Arnon 1972). Increase in achene yield seems
to be the direct result of improved head diameter,
Cro
p gr
owth
rat
e(g
m–2
day
–1)
0
5
10
15
20
25
(a) (b)
45 60 75 90
CKFVGBFGBVSAFSA
0
5
10
15
20
25
45 60 75 90
CKFVGBFGBVSAFSA
Days after sowing
Fig. 6 Leaf area duration as affected by foliar
application of GB and SA drought stress in
sunflower during (a) 2006 and (b) 2007.
CKF = Control (no GB and SA application);
VGB = exogenous application of GB at
flowering stage; FGB = exogenous application
of GB at flowering stage; VSA = exogenous
application of SA at flowering stage;
FSA = exogenous application of GB at
flowering stage.
Hussain et al.
ª 2009 The Authors106 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109

number of achenes per had and 1000-achene weight (data
not given). It may be the result of maintenance of photo-
synthetic activity owing to GB application. By means of
osmotic adjustment, the organelles and cytoplasmic activ-
ities take place at about a normal pace and help plants to
perform better in terms of growth, photosynthesis and
assimilate partitioning to grain filling (Ludlow and
Muchow 1990, Subbarao et al. 2000). As a mechanism,
osmotic adjustment has been suggested as an important
trait in postponing the dehydration stress in water scarce
environments (Morgan 1990). Externally applied GB can
rapidly penetrate through leaves and be transported to
other organs, where it would contribute to improved
stress tolerance (Makela et al. 1998).
Reduction in biological yield by water stress might be
because of decreased LAI, LAD and CGR as indicated by
the data (Figs 1–6) resulting in reduced photo-assimila-
tion. It is a common adverse effect of water stress on
crop plants in the reduction of fresh and dry biomass
production (Ashraf and O’Leary 1996) and reduced bio-
mass production resulting from water stress had been
observed in almost all genotypes of sunflower (Tahir and
Mehdi 2001). Manivannan et al. (2007) also reported
reduction in fresh and dry weight in sunflower by
imposing water stress. Exogenous application of GB at
FS improved the biological yield by improving LAI, LAD
and CGR (Figs 1–3). This improvement may be attrib-
uted to maintenance of photosynthetic activity by GB
application.
Harvest index indicates the proportionate translocation
of assimilates into economic yield. Water stress
treatments had a significant effect on harvest index. The
lowest harvest index in case of water stress at FS might be
because water shortage at FS increases the chances of
pollen sterility, abortion, pollen germination and fertiliza-
tion in-compatibility, which directly reduce the number
of achenes per head and achene yield. Exogenous applica-
tions of GB and SA also did not considerably affect the
harvest index. The possible explanation might be that the
treatments producing higher achene yield also resulted in
higher biomass production, biological yield. So, harvest
index remained statistically similar because it is a ratio
between economic yield and biological yield.
Water stress at both the stages (vegetative and FS) ham-
pered the crop growth, but it was considerably improved
by the exogenous applications of GB and SA, particularly
by the exogenous application of GB at FS. Water stress
reduced the LAI, LAD and CGR, which may be attributed
to impaired water relations (Table 1). Sadras et al. (1993)
also reported a decrease in leaf expansion rate because of
decreased water potential under water stress in sunflower.
Likewise, total leaf area was decreased considerably in
sorghum by water stress (Yadav et al. 2005).
Reduced LAI resulted in reduced LAD as LAD is
directly calculated from LAI. Similarly improved CGR
under stress conditions might be the result of mainte-
nance of photosynthetic activity because of GB and SA
application. Earlier, Agboma et al. (1997) found that
exogenous GB application protects the photosynthetic
machinery in maize, wheat and sorghum thereby increas-
ing the final yield. Similarly, Makela et al. (1999) reported
that foliar application of GB increased net photosynthesis
and decreased photorespiration in turnip rape plants
under drought and salt stress. Exogenous application of
GB and SA increased the crop growth. It might be due
maintenance of LAI (assimilatory surface) due to high
turgor and sustained photosynthetic ability of the crop by
protecting the photosynthetic machinery from ROS
produced during drought stress and by increasing RuBP
content under drought condition. One of the interesting
findings of the study was that achene protein was reduced
as a result of GB application and increasing water
application, which might be due to dilution because yield
was increased.
In conclusion, water stress negatively affected the plant
water relations and growth of the crop including achene
yield, but exogenous GB and SA application particularly
GB appreciably improved these attributes under water
stress. However, exogenous GB application at the FS was
more effective than other treatments. NAR was not
considerably affected by water stress as well as application
of GB and SA. WUE was increased by missing irrigation
at budding stage, but considerably decreased by missing
irrigation at FS than control. The protein contents were
appreciably increased by increasing water stress at different
growth stages, but were reduced by exogenous GB and SA
application. The effects of water stress and foliar applica-
tion of GB were more pronounced when applied at FS than
at the budding stage. Moreover, exogenous GB application
was advantageous only under stress conditions.
References
Agboma, P., M. G. K. Jones, P. Peltonen-Sainio, H. Rita, and
E. Pehu, 1997: Exogenous glycinebetaine enhances grain
yield of maize, sorghum and wheat grown under two
watering regimes. J. Agron. Crop Sci. 178, 29–37.
Alexieva, V., I. Sergiev, S. Mapelli, and E. Karanov, 2001: The
effect of drought and ultraviolet radiation on growth and
stress markers in pea and wheat. Plant Cell Environ. 24,
1337–1344.
Allakhverdiev, S. I., H. Hayashi, Y. Nishiyama, A. G. Ivanov, J.
A. Aliev, V. V. Klimov, N. Murata, and R. Carpentier, 2003:
Glycinebetaine protects the D1/D2/Cytb559 complex of
photosystem II against photo-induced and heat-induced
inactivation. J. Plant Physiol. 160, 41–49.
Sunflower under Water Deficit Conditions
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109 107

Arnon, I. 1972. Crop Production in Dry Regions, pp. 203–211.
Leonard Hill Book, London.
Ashraf, M., and J. W. O’Leary, 1996: Effect of drought stress
on growth, water relations and gas exchange of two lines of
sunflower differing in degree of salt tolerance. Int. J. Plant
Sci. 157, 729–732.
Atteya, A. M., 2003: Alteration of water relations and yield
of corn genotypes in response to drought stress. Bulgar.
J. Plant Physiol. 29, 63–76.
Borsani, O., V. Valpuesta, and M. A. Botella, 2001: Evidence
for a role of salicylic acid in the oxidative damage generated
by NaCl and osmotic stress in Arabidopsis seedlings. Plant
Physiol. 126, 1024–1030.
Bremner, J. M., 1964: Organic forms of nitrogen. In: C. A.
Black, ed. Methods of Soil Analysis, Part-2, Vol. 9,
pp. 1235–1255. American Society of Agronomy, Madison,
WI.
Buland, A., A. Hussain, A. H. Ahmad, M. Saleem, N. Meh-
mood, M. Aslam, and K. Ali, 1994: Optimizing irrigation
requirements for maximizing yield of mungbean crop – an
experimental study. Pak. J. Agric. Sci. 31, 331–335.
Farooq, M., S. M. A. Basra, A. Wahid, Z. A. Cheema, M. A.
Cheema, and A. Khaliq, 2008a: Physiological role of exoge-
nously applied glycinebetaine in improving drought toler-
ance of fine grain aromatic rice (Oryza sativa L.). J. Agron.
Crop Sci. 194, 325–333.
Farooq, M., T. Aziz, M. Hussain, H. Rehman, K. Jabran,
and M. B. Khan, 2008b: Glycinebetaine improves chilling
tolerance in hybrid maize. J. Agron. Crop Sci. 194,
152–160.
Farooq, M., T. Aziz, S. M. A. Basra, M. A. Cheema, and H.
Rehamn, 2008c: Chilling tolerance in hybrid maize induced
by seed priming with salicylic acid. J. Agron. Crop Sci. 194,
7, 161–168.
Farooq, M., A. Wahid, N. Kobayashi, D. Fujita, and
S. M. A. Basra, 2009: Plant drought stress: effects,
mechanisms and management. Agron. Sustain. Dev. 29,
185–212.
Horvath, E., M. Pal, G. Szalai, E. Paldi, and T. Janda, 2007:
Exogenous 4-hydroxybenzoic acid and salicylic acid modu-
late the effect of short-term drought and freezing stress on
wheat plants. Biol. Plant. 51, 480–487.
Hunt, R., 1978: Plant growth analysis. Studies in Biology No.
96, pp. 26–38. Edward Arnold, London.
Hussain, M., M. A. Malik, M. Farooq, M. Y. Ashraf, and M.
A. Cheema, 2008a: Improving drought tolerance by exoge-
nous application of glycinebetaine and salicylic acid in
sunflower. J. Agron. Crop Sci. 194, 193–199.
Hussain, M., M. Farooq, K. Jabran, H. Rehman, and
M. Akram, 2008b: Exogenous glycinebetaine application
improves yield under water limited conditions in hybrid
sunflower. Arch. Agron. Soil Sci. 54, 557–567.
Jensen, H. E., and P. Mogenson, 1984: Yield and nutrient
contents of spring wheat subjected to water stress at various
growth stages. Acta Agrica. 34, 527–533.
Kaya, M. D., G. Okcub, M. Ataka, Y. Cıkılıc, and O. Kolsarıcıa,
2006: Seed treatments to overcome salt and drought stress
during germination in sunflower (Helianthus annuus L.).
Eur. J. Agron. 24, 291–295.
Kramer, P. J., and J. S. Boyer, 1995: Water Relations of Plants
and Soils. Academic Press, San Diego, CA.
Ludlow, M. M., and R. C. Muchow, 1990: A critical evaluation
of traits for improving crop yield in water limited environ-
ments. Adv. Agron. 43, 107–153.
Ma, Q.-Q., W. Y.-H. Wang, D. Q. Li, and Q. Zou, 2006:
Alleviation of photoinhibition in drought-stressed wheat
(Triticum aestivum L.) by foliar-applied glycinebetaine.
J. Plant Physiol. 163, 165–175.
Makela, P., K. Jokinen, M. Kontturi, P. Peltonen-Sainio,
E. Pehu, and S. Somersalo, 1998: Foliar application of
glycinebetaine – a novel product from sugar beet – as an
approach to increase tomato yield. Ind. Crops Prod. 7,
139–148.
Makela, P., M. Kontturi, E. Pehu, and S. Somersalo, 1999:
Photosynthetic response of drought and salt-stressed tomato
and turnip rape crops to foliar applied glycinebetaine.
Physiol. Plant. 105, 45–50.
Manivannan, P., C. A. Jaleel, B. Sankar, A. Kishorekumar, R.
G. Somasundaram, M. A. Lakshmanan, and R. Panneersel-
vam, 2007: Growth, biochemical modifications and proline
metabolism in Helianthus annuus L. as induced by drought
stress. Colloids Surf. B Biointerfaces 59, 141–149.
Morgan, J. M., 1984: Osmoregulation and water stress in
higher plants. Ann. Rev. Plant Physiol. 35, 299–319.
Morgan, P. W., 1990: Effects of abiotic stresses on plant
hormone systems. In: R. G. Alscher, and J. R. Cumming,
eds. Stress Responses in Plants: Adaptation and Acclimation
Mechanisms, pp. 113–146. Wiley-Liss, Wilmington, DE.
Naidu, B. P., D. F. Cameron, and S. V. Konduri, 1998:
Improving drought tolerance of cotton by glycinebetaine
application and selection. In: D. Michalk, D. Kemp, P.
Dowling, and J. Kirkegaard, eds. Proceedings of the 9th
Australian Agronomy Conference. Australian Agronomy
Conference, Wagga Wagga.
Nonami, H., 1998: Plant water relations and control of cell
elongation at low water potentials. J. Plant. Res. 111,
373–382.
Quan, R. D., M. Shang, H. Zhang, and J. Zhang, 2004:
Improved chilling tolerance by transformation with betA
gene for the enhancement of glycinebetaine synthesis in
maize. Plant Sci. 166, 141–149.
Rhodes, D., and A. D. Hanson, 1993: Quaternary ammonium
and tertiary sulfonium compound in higher plant. Annu.
Rev. Plant Physiol. Plant Mol. Biol. 44, 357–384.
Sadras, V. O., F. J. Villalobes, and E. Fereres, 1993: Leaf expan-
sion in field-grown sunflower in response to soil and leaf
water status. Agron. J. 85, 564–570.
Sakamoto, A., and N. Murata, 2002: The role of glycinebetaine
in the protection of plants from stress: clues from transgenic
plants. Plant Cell Environ. 25, 163–171.
Hussain et al.
ª 2009 The Authors108 Journal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109

Serraj, R., and T. R. Sinclair, 2002: Osmolyte accumulation:
can it really help increase crop yield under drought condi-
tions? Plant Cell Environ. 25, 333–341.
Siddique, M. R. B., A. Hamid, and M. S. Islam, 2000: Drought
stress effects on water relations of wheat. Bot. Bull. Acad.
Sinica 41, 35–39.
Singh, B., and K. Usha, 2003: Salicylic acid-induced physiologi-
cal and biochemical changes in wheat seedlings under water
stress. Plant Growth Regul. 39, 137–141.
Subbarao, G. V., N. H. Nam, Y. S. Chauhan, and C. Johansen,
2000: Osmotic adjustment, water relations and carbohydrate
remobilization in pigeonpea under water deficits. J. Plant
Physiol. 157, 651–659.
Tahir, M. H. N., and S. S. Mehdi, 2001: Evaluation of open
pollinated sunflower (Helianthus annuus L.) populations
under water stress and normal conditions. Int. J. Agric. Biol.
3, 236–238.
Viets, F. G., 1962: Fertilizers and the efficient use of water.
Adv. Agron. 14, 223–264.
Waseem, M., H. U. Athar, and M. Ashraf, 2006: Effect of
salicylic acid applied through rooting medium on drought
tolerance of wheat. Pak. J. Bot. 38, 1127–1136.
Watson, D. J., 1947: Comparative physiological studies on the
growth of field crops. I. variation in net assimilation rate
and leaf area between species and varieties and within and
between years. Analyt. Bot. 11, 41–76.
Xing, W., and C. B. Rajashekar, 1999: Alleviation of water stress
in beans by exogenous glycinebetaine. Plant Sci. 148, 185–195.
Yadav, S. K., N. J. Lakshmi, M. Maheswari, M. Vanaja, and B.
Venkateswarlu, 2005: Influence of water deficit at vegetative,
anthesis and grain filling stages on water relation and grain
yield in sorghum. Indian J. Plant Physiol. 10, 20–24.
Zhu, J. K., 2002: Salt and drought stress signal transduction in
plants. Annu. Rev. Plant Biol. 53, 247–273.
Sunflower under Water Deficit Conditions
ª 2009 The AuthorsJournal compilation ª 2009 Blackwell Verlag, 195 (2009) 98–109 109




















