
Inhibition of epidermal xenobiotic metabolism in SENCAR mice by naturally occurring plant phenols
Upload
independentCategory
view
0download
0
Hindawi Publishing CorporationISRN GastroenterologyVolume 2013 Article ID 710856 14 pageshttpdxdoiorg1011552013710856
Research ArticleExamination of the Anti-Inflammatory Antioxidant andXenobiotic-Inducing Potential of Broccoli Extract and VariousEssential Oils during a Mild DSS-Induced Colitis in Rats
Kristin Mueller Nicole Michaela Blum and Andreas Stefan Mueller
Institute of Agricultural and Nutritional Sciences Martin Luther University Halle-Wittenberg Von-Danckelmann-Platz 206120 Halle (Saale) Germany
Correspondence should be addressed to Kristin Mueller kristinmuellergmxnet
Received 9 December 2012 Accepted 20 January 2013
Academic Editors A Amedei A K Rishi and N Senninger
Copyright copy 2013 Kristin Mueller et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Phytogenic compounds with antioxidant and anti-inflammatory properties are currently discussed as promising complementaryagents in prevention and treatment of inflammatory bowel disease (IBD) Our study aimed to evaluate possible protective andcurative effects of broccoli extract (BE) and of the essential oils of turmeric (Cuo) thyme (To) and rosemary (Ro) in a rat modelwith a mild dextran sulphate sodium- (DSS-) induced colitis Therefore Wistar rats were fed a diet without an additive (Con) ordiets with the addition of BE Cuo To and Ro during the whole experiment Pretreatment with Ro Cuo and To increased theexpression of the tight junction protein Cldn3 All additives reduced mRNA of VCAM-1 which plays a crucial role in the first stateof inflammatory response Only Ro pretreatment affected the expression of the antioxidant enzymesHO1 GPx2 and of glutathione-S-transferases All additives counteracted the DSS-induced rise in COX2 and VCAM-1 expression Colonic IL-10 was increased byCuo To and Ro During the recovery phase DSS pretreatment increased NF120581B VCAM-1 and MCP-1 This response was counter-regulated by all additives We conclude that the phytogenic additives tested have a promising anti-inflammatory potential in vivoand a particular role in the prevention of IBD
1 Introduction
Inflammatory bowel disease including ulcerative colitis (UC)and Crohnrsquos disease (CD) is a multifactorial relapsing-remitting disorder characterized by intermittent periods ofacute inflammation in the small and in the large intestineThemain difference between CD and UC is the location and thenature of the inflammatory changes CD can affect any part ofthe gastrointestinal tract from mouth to anus although theonset of the majority of cases is located in the terminal ileumIn contrast UC is restricted to the colon and the rectumThe exact pathogenic mechanisms provoking both disordersremain almost unclear However in a number of casesoverreactions of the immune system due to inflammatorystimuli can be observed In this context proinflammatoryimmune modulators like interleukin 1 beta (IL-1120573) mono-cyte chemoattractant protein 1 (MCP-1) and vascular celladhesion molecule 1 (VCAM-1) play an important role in
the development of the disease [1 2] Nuclear factor ldquokappa-light-chain-enhancerrdquo of activated B cells (NF120581B) representsa key transcription factor regulating the synthesis of genesinvolved in immune reactions and inflammatory response Innoninflamed tissues NF120581B is inhibited through linkage to itscytosolic inhibitor protein kappa B (I120581B) The activation ofproteasomal I120581B degradation via phosphorylation of criticalserine residues by proinflammatory stimuli elicits NF120581Btranslocation to the nucleus and the subsequent inductionof its target genes like tumor necrosis factor (TNF)-120572 andother inflammatory mediators (interleukin 2 interleukin 6interleukin 8 VCAM-1 intracellular cell adhesion molecule1 and interferon 120574) [3ndash5]
Dysfunction of the gut barrier accompanied by anincreased intestinal permeability is another characteristicsymptom in the pathophysiology of IBD [6] As a con-sequence of the disordered permeability both antigenicdeterminants derived from food digestion and commensal
2 ISRN Gastroenterology
or pathogenic bacteria can overcome the mucosal barrierunimpeded and provoke a continuous intestinal immuneresponse and tissue damage [7] In this context the increasedpermeability of gastrointestinal epithelial cells frequentlyresults from the destruction of tight junctions This processis triggered by oxidative stress deriving from reactive oxygenspecies (ROS) mycotoxins (eg patulin) bacterial com-ponents (eg lipopolysaccharides LPS) and inflammatorymediators (eg cytokines) [8ndash11]
Due to their generally accepted antioxidant and anti-inflammatory properties the use of plant extracts and inparticular of essential oils represents a promising approachto prevent and cure IBD For instance the essential oil ofOriganum vulgare containing high concentrations of thephenolic terpenes carvacrol and thymol has been shownto efficiently reduce the mRNA levels of the proinflam-matory cytokines TNF120572 IL-1120573 and IL-6 in human THP-1macrophages In contrast in this study the anti-inflammatorycytokine IL-10 was significantly reduced [12] A blend oforegano anise and lemon peel even could be demonstratedto evolve anti-inflammatory effects in piglets in vivo [13]Data from current literature suggest that anti-inflammatoryeffects of plant extracts base on their direct and indirectantioxidant properties which again depend on the chem-ical compounds contained in different plant extracts Theessential oils of thyme and oregano which mainly containthe antioxidant terpene compounds thymol and carvacrol[14 15] could be demonstrated to impair the mRNA andthe protein concentration of the pro-inflammatory cytokinesIL-1120573 and IL-6 in mice with 246-trinitrobenzol (TNBS-)induced colitis [16] Carnosol a terpene of rosemary oilin vitro showed indirect antioxidant effects via inducingnuclear factor erythroid 2-related factor 2- (Nrf2-) regulatedantioxidant enzyme expression and additionally decreasedpro-inflammatory mediators like NF120581B TNF120572 IL-1120573 IL-6 COX2 and ICAM-1 [17ndash19] In another in vitro studytreatment with sulforaphane an isothiocyanate mainly con-tained in broccoli decreased the mRNA concentration ofthe pro-inflammatory cytokines TNF120572 and IL-1120573 in murineRAW2647 macrophages due to a pro-inflammatory stimuluswith bacterial LPS In this experiment the reduction ofinflammation was accompanied by an increase in the expres-sion of the antioxidant enzyme HO1 via the Nrf2Kelch-like ECH-associated protein 1 (Keap1) pathway [20] Similarstrong indirect antioxidant effects through the modificationof Keap1 sulfhydryl groups have also been described for ar-turmeron the main terpene of curcuma oil In addition ar-turmeron possesses also direct antioxidant properties [21]
However until today studies comparing the anti-inflammatory potential of different phytogenic substancesdue to a pro-inflammatory stimulus are not available DSS isa chemical compound routinely used to induce a colitis inmodel animals and therefore to mimic similar inflammatoryconditions as present in IBD
Consequently our study aimed to investigate theconnection between the antioxidant potential and anti-inflammatory effects of broccoli extract turmeric oil thymeoil and rosemary oil in rats with a mild DSS-induced colitis
2 Materials and Methods
21 Animals and Diets The protocol of the rat study wasapproved by the Regional Council of Halle and by theAnimal Welfare Committee of the Martin Luther UniversityHalle-Wittenberg (record token 42502-2-1093-MLUG) 92four=week old male Wistar rats (mean body weight 1862 plusmn945 g) were obtained from Harlan laboratories (Horst TheNetherlands) The rats were fed a standard diet withoutphytogenic feed additives for an acclimatisation period of 14days
At an age of 6 weeks and a mean live weight of 2310 plusmn128 g the rats were assigned to 6 experimental groups of16 rats each During the following experimental periods thecontrol groups (Con and DSS) were fed a basal diet that metthe nutritional demands of the NRC for growing rats Thisbasal diet contained no phytogenic additive (Table 1)
The diet of group BE contained 8750mgkg broccolisprouts extract (JARROW Formulas) 1494mgkg diet ofCurcuma longa oil (Cuo) 618mgkg of Thymus vulgaris oil(To) and 680mgkg of Rosmarinus officinalis oil (Ro) wereadded to the diets of the other groups in order to standardizethe concentration of the isothiocyanate sulforaphane (BE)and of the individual main terpenes (Cuo To and Ro) toa value of 2mmolkg diet The main terpenes consideredwere ar-turmerone for Cuo thymol for To and 18-cineolfor Ro All diets were pelleted with a pellet mill using an8mm die and fed during the whole course of the experimentThe rats had free access to their respective diet and to tapwater Lighting humidity and temperature regime was inaccordance with the recommendations of the Society forLaboratory Animal Science (2004) [22] The trial consistedof 3 phases (1) pretreatment phase (phase 1 7 days) (2) DSS-treatment phase (phase 2 6 days) and (3) recovery phase(phase 3 6 days) Table 2 overviews the feeding protocol indetail
The animal model of a DSS-induced colitis was cho-sen due to several histological and biochemical similaritieswith human IBD [23 24] In the DSS-treatment phase 4DSS (40 kDa Sigma-Aldrich) was administered via drinkingwater to all rats for 6 days with the exception of the Congroup in order to induce amild intestinal inflammation Feedintake and individual live weight were recorded after oneweek daily during the DSS-phase and every other day duringthe recovery phase
At the end of phase 1 four rats were killed for organsampling (liver and colon) and after phases 2 and 3 six ratsper group were sacrificed For the histological examinationand for the determination of relative mRNA concentrationsof antioxidant and xenobiotic enzymes and of inflammationparameters colon samples were prepared from a 10 cmsegment distal to the caecum Liver samples were excisedfrom the middle of lobus sinister lateralis
During treatment with 4 DSS the disease activity index(DAI) was used to assay the severity of the induced colitisDAI was determined daily in phase 2 and every other dayin phase 3 (recovery phase) The scoring system was basedon body weight loss stool consistency andmacroscopic fecal
ISRN Gastroenterology 3
(a) (b)
Figure 1 Representative pictures of proximal colon sections from healthy Con rats (a) and DSS-treated rats (b) fed the control dietHaematoxylin and Eosin (H and E) stained colon cross-sections fromphase 2were analyzedwith inversionmicroscopy (20x) Arrows indicateincreased leucocyte infiltration and arrowheads indicate the loss of goblet cells caused by DSS treatment (b)
(a) (b)
(c) (d)
Figure 2 Representative pictures of proximal colon sections fromDSS-treated rats fed experimental diets with broccoli extract (a) turmericoil (b) thyme oil (c) or rosemary oil (d) Haematoxylin and Eosin (H and E) stained colon cross-sections from phase 2 were analyzed withinversion microscopy (20x)
4 ISRN Gastroenterology
Table 1 Basal diet
Ingredient gkg basal dietWheat (DEUKA GmbH und Co KG KonnernGermany) 2379
Maize (DEUKA GmbH und Co KG KonnernGermany) 2000
Barley (DEUKA GmbH und Co KG KonnernGermany) 1560
Soybean meal 46 CP (DEUKA GmbH und CoKG Konnern Germany) 2200
Wheat bran (DEUKA GmbH und Co KGKonnern Germany) 788
Oat (DEUKA GmbH und Co KG KonnernGermany) 690
Sun flower oil 150Lysine (Feed Grade China) 03dl-methionine (Degussa Duesseldorf Germany) 20Vitamin and mineral premix 121Calcium carbonate (Sigma-Aldrich) 25Calcium phosphate (Mischfutter und LandhandelGmbH Edderitz Germany) 79
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
I120581B120572 degradation
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
Figure 3 NF120581B and Nrf2 crosstalk under balanced anti- andprooxidant conditions
blood debris For each mentioned parameter a scale rangingfrom 0 to 4 was applied as described previously [25]
22 Colonic Histology For the histological examinationfreshly dissected colon samples were washed with 09 (wv) NaCl and cryoconserved in a freezing medium (JungLeica InstrumentsNusslochGermany) Serial cross-sections(7 120583m) were prepared using a microtome (CM 1850 UVmicrotome Jung Leica) and fixed on sterile usual micro-scope slides After staining the samples with Haematoxylin-Eosin full-thickness slices tissue architecture infiltration ofneutrophilic granulocytes into the mucosa and into the sub-mucosa and the formation of crypt abscesses were examinedunder an inversion microscope using a blind protocol
Low oxid stress electrophils and
phytogenic substanceseg SFN curcumin120581BE-driven gene
transkription eg IL-1120573I120581B120572 Keap1
(Cul3-Rbx1)
Nrf2
+
+
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
I120581B120572 degradation
Figure 4 NF120581B and Nrf2 interaction due to feeding phytogeniccompounds or low oxidative stress
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
+
+
+
+
++
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
P
IKK
Progressiveoxid stress
eg DSS H2O2
I120581B120572 degradation
Figure 5 NF120581B and Nrf2 interaction due to DSS treatment orprogressive oxidative stress
23 RNA Preparation and Real-Time RT-PCR Analysis TotalRNA from 100mg of liver and colon tissue was isolatedusing the acid guanidinium thiocyanate-phenol-chloroformextraction method [26] Most recently a strong DSS RNAinterference during acute DSS-treatment resulting in thelack of signals during gene expression analysis has beendescribed [27] To assure the correctness of gene expressionanalyses in our experiment polyA+ mRNA from the colonicsamples of the DSS-phase was purified using the GenElutemRNA Miniprep Kit (Sigma-Aldrich MO USA) accordingto the manufacturersrsquo protocol Following the photometricaldetermination of RNA concentration and purity at 260 nmand 280 nm reverse transcription of 30 120583g of total RNAor of 03 120583g of purified mRNA and real-time RT-PCR wereperformed as described previously [28] Gene bank accession
ISRN Gastroenterology 5
Table 2 Feeding protocol
Group Phytogenicadditive
Koncentration per kgdiet Phase 1 Phase 2 Phase 3
Con None mdash 7 days diets and water adlibitum
6 days diets and water adlibitum
6 days diets and water adlibitum
DSS None mdash 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
BE Broccoli extract 2mmol sulforaphane 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Cuo Turmeric oil 2mmol ar-turmerone 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
To Thyme oil 2mmol thymol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Ro Rosemary oil 2mmol 18-cineol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
numbers and primer sequences (51015840 rarr 31015840) are shown inTable 3
Gene specific mRNA expression was analyzed with theRotor-Gene 6000 series software using the ΔΔCt method[29] The amplification data of the single genes were nor-malized to the expression of the two most stable refer-ence genes in each tissue (liver 120573-actin ribosomal proteinL13A (Rpl13a) colon Rpl13a hypoxanthine phosphoribosyl-transferase 1 (Hprt1)) Relative mRNA expression levels areexpressed as x-fold changes relative to group Con = 10
24 Statistical Analysis Data are presented as means plusmn theirstandard error of the mean (SEM) Statistical differenceswere analyzed with SPSS 190 for Windows (IBM ChicagoUSA) using one-way ANOVA after verifying the normalityof distribution (Shapiro Wilk test and Kolmogorov Smirnovtest) and the homogeneity of variances (Levene test) TheLeast Significant Difference test (LSD) was used to ana-lyze significant differences between means if variances werehomogenous If not the Games Howell test was used Atan error probability of less than 5 (119875 lt 005) differencesbetween means were considered as statistically significant
3 Results
31 Body Weight Development Neither food intake (datanot shown) nor final body weight (Table 4) of the rats wasinfluenced significantly in the different experimental phasesby feeding the specific diets tested Remarkably also DSStreatment in the second phase did not affect the above-mentioned parameters significantly
32 Colitis Severity by Disease Activity Index (DAI) Thecourse of the mild colitis induced by the administration of4 DSS to rats for 6 days was controlled daily in phase2 by measuring the DAI A high number of rats from allDSS-treated groups had a soft stool consistency but severediarrhea accompanied by blood debris could not be observedMoreover a distinct weight loss frequently observed underDSS treatment was present only in some cases and only for
one day Consequently the DAI in all DSS-treated groupsremained below 10 (DSS 012 BE 010 Cuo 010 To007 and Ro 013) and did not differ significantly fromCon rats receiving no DSS In phase 3 obvious changesin stool consistency almost disappeared or were much lesspronounced than in phase 2
33 Histology DSS treatment caused no significant observ-able macroscopic changes in colonic tissue architecturebetween the experimental groups including the untreatedCon rats Furthermore microscopic analysis revealed nosignificant histological damage to the colonic mucosa of ratsexposed to 4 DSS for 6 days However DSS treatmenttended to accelerate initial damage to the mucosa charac-terised by the loss of goblet cells and the occurrence of amorediffuse crypt architecture compared to the colon of healthyCon rats Additionally the accumulation of neutrophils infil-trating the lamina propria could be observedmore frequentlyin colonic slices of DSS-treated rats These mentioned DSS-induced mucosal alterations are shown in Figure 1 DSStreatment in combination with the tested phytogenic extractsalso caused slightmucosal damage (Figure 2) but the severityseemed to be much lower than in group DSS (Figure 1(b))
34 mRNA Expression of NF120581B TNF120572 and VariousInflammatory-Mediating Enzymes in Colon and Liver TissueThe analyzed mRNA expression patterns of pro- and anti-inflammatory genes controlled by NF120581B and TNF120572 differedamong the experimental periods and the tissues investigated(Tables 5 and 6) During the 7-day pretreatment phase 1 themRNA abundance of the colonic pro-inflammatory markersCOX2 and IL-1120573 of the anti-inflammatory cytokine IL-10 ofthe cell adhesion molecules MCP-1 and VCAM-1 and of thetight junction protein Cldn3 showed a high intraindividualvariance In contrast in phase 1 of the trial liver mRNAdata of the above-mentioned genes showed a much betterhomogeneity Nevertheless feeding diets with BE- Cuo-To- and Ro-addition reduced colonic VCAM-1 mRNA by 57to 64 compared to untreated control rats This reductionwas significant for Cuo To and Ro and represented a trend(119875 lt 010) for BE Moreover in phase 1 colonic Cldn3
6 ISRN Gastroenterology
Table 3 Gene bank accession numbers and primer sequences of the genes investigated by real-time RT-PCR
Gene name (abbreviation used) Gene bank accession number Primer sequences (51015840 rarr 31015840)for = forward rev = reverse
Chemokine (C-C motif) ligand 2 (Ccl2) (MCP1) NM 031530 for GTGCGACCCCAATAAGGAArev TGAGGTGGTTGTGGAAAAGA
Claudin 3 (Cldn3) NM 031700 for TATCCTACTGGCAGCCTTCGrev GTTCCCATCTCTCGCTTCTG
Copperzinc superoxide dismutase (SOD1) NM 017050 for CCACTGCAGGACCTCATTTTrev CACCTTTGCCCAAGTCATCT
C-reactive protein (CRP) NM 017096 for GTCTCTATGCCCACGCTGATrev CCGTCAAGCCAAAGCTCTAC
Glutathione S-transferase K1 (GSTK1) NM 181371 for GAGCATGGAGCAACCAGAGATrev AGCTTGCTCTTCACCAGTTCG
Glutathione S-transferase P1 (GSTP1) NM 012577 for GAGGCAAAGCTTTCATTGTGGrev GTTGATGGGACGGTTCAAATG
Glutathione S-transferase T2 (GSTT2) NM 012796 for GAGGAAAAGGTGGAACGGAACrev CGCCCCTCAAACAGATTACAG
Glutathione peroxidase 2 (GPx2) NM 183402 for GTGTGATGTCAATGGGCAGAArev ACGTTTGATGTCAGGCTCGAT
Heme oxygenase 1 (HO1) NM 012580 for AGGCACTGCTGACAGAGGAACrev AGCGGTGTCTGGGATGAACTA
Hypoxanthine phosphoribosyltransferase 1 (Hprt1) NM 012583 for GCAGACTTTGCTTTCCTTGGrev TCCACTTTCGCTGATGACAC
Interleukin 1 beta (IL-1120573) NM 031512 for CTGTGACTCGTGGGATGATGrev GGGATTTTGTCGTTGCTTGT
Interleukin 10 (IL-10) NM 012854 for CTGGAGTGAAGACCAGCAAAGGrev GGAGAAATCGATGACAGCGTCG
Kelch-like ECH-associated protein1 (Keap1) NM 057152 for GTGGCGGATGATTACACCAATrev GAAAAGTGTGGCCATCGTAGC
NAD(P)H dehydrogenase [quinone] 1 (NQO1) NM 017000 for CGCAGAGAGGACATCATTCArev CGCCAGAGATGACTCAACAG
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) NM 031789 for CCAAGGAGCAATTCAACGAAGrev CTCTTGGGAACAAGGAACACG
Nuclear factor kappa B (NF120581B) L26267 for CTTCTCGGAGTCCCTCACTGrev CCAATAGCAGCTGGAAAAGC
Prostaglandin-endoperoxide synthase 2 (Ptgs2) (COX2) NM 017232 for GCTGTACAAGCAGTGGCAAArev CCCCAAAGACAGCATCTGGA
Ribosomal protein L13A (Rpl13a) NM 173340 for CCCTCCACCCTATGACAAGArev CCTTTTCCTTCCGTTTCTCC
Tumor necrosis factor alpha (TNF120572) NM 012675 for GCCAATGGCATGGATCTCAAAGrev AAATCGGCTGACGGTGTGGG
Vascular cell adhesion molecule 1 (VCAM 1) NM 012889 for TGACATCTCCCCTGGATCTCrev CTCCAGTTTCCTTCGCTGAC
120573-actin NM 031144 for ATCGTGCGTGACATTAAAGAGAAGrev GGACAGTGAGGCCAGGATAGAG
expression was significantly higher in Ro-treated rats andtendencially higher in Cuo- and To-treated rats than in theirCon littermates
In phase 1 feeding diets containing BE Cuo and Rosignificantly decreased liver mRNA abundance of the pro-inflammatory cytokine IL-10 by 32 to 44
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 ISRN Gastroenterology
or pathogenic bacteria can overcome the mucosal barrierunimpeded and provoke a continuous intestinal immuneresponse and tissue damage [7] In this context the increasedpermeability of gastrointestinal epithelial cells frequentlyresults from the destruction of tight junctions This processis triggered by oxidative stress deriving from reactive oxygenspecies (ROS) mycotoxins (eg patulin) bacterial com-ponents (eg lipopolysaccharides LPS) and inflammatorymediators (eg cytokines) [8ndash11]
Due to their generally accepted antioxidant and anti-inflammatory properties the use of plant extracts and inparticular of essential oils represents a promising approachto prevent and cure IBD For instance the essential oil ofOriganum vulgare containing high concentrations of thephenolic terpenes carvacrol and thymol has been shownto efficiently reduce the mRNA levels of the proinflam-matory cytokines TNF120572 IL-1120573 and IL-6 in human THP-1macrophages In contrast in this study the anti-inflammatorycytokine IL-10 was significantly reduced [12] A blend oforegano anise and lemon peel even could be demonstratedto evolve anti-inflammatory effects in piglets in vivo [13]Data from current literature suggest that anti-inflammatoryeffects of plant extracts base on their direct and indirectantioxidant properties which again depend on the chem-ical compounds contained in different plant extracts Theessential oils of thyme and oregano which mainly containthe antioxidant terpene compounds thymol and carvacrol[14 15] could be demonstrated to impair the mRNA andthe protein concentration of the pro-inflammatory cytokinesIL-1120573 and IL-6 in mice with 246-trinitrobenzol (TNBS-)induced colitis [16] Carnosol a terpene of rosemary oilin vitro showed indirect antioxidant effects via inducingnuclear factor erythroid 2-related factor 2- (Nrf2-) regulatedantioxidant enzyme expression and additionally decreasedpro-inflammatory mediators like NF120581B TNF120572 IL-1120573 IL-6 COX2 and ICAM-1 [17ndash19] In another in vitro studytreatment with sulforaphane an isothiocyanate mainly con-tained in broccoli decreased the mRNA concentration ofthe pro-inflammatory cytokines TNF120572 and IL-1120573 in murineRAW2647 macrophages due to a pro-inflammatory stimuluswith bacterial LPS In this experiment the reduction ofinflammation was accompanied by an increase in the expres-sion of the antioxidant enzyme HO1 via the Nrf2Kelch-like ECH-associated protein 1 (Keap1) pathway [20] Similarstrong indirect antioxidant effects through the modificationof Keap1 sulfhydryl groups have also been described for ar-turmeron the main terpene of curcuma oil In addition ar-turmeron possesses also direct antioxidant properties [21]
However until today studies comparing the anti-inflammatory potential of different phytogenic substancesdue to a pro-inflammatory stimulus are not available DSS isa chemical compound routinely used to induce a colitis inmodel animals and therefore to mimic similar inflammatoryconditions as present in IBD
Consequently our study aimed to investigate theconnection between the antioxidant potential and anti-inflammatory effects of broccoli extract turmeric oil thymeoil and rosemary oil in rats with a mild DSS-induced colitis
2 Materials and Methods
21 Animals and Diets The protocol of the rat study wasapproved by the Regional Council of Halle and by theAnimal Welfare Committee of the Martin Luther UniversityHalle-Wittenberg (record token 42502-2-1093-MLUG) 92four=week old male Wistar rats (mean body weight 1862 plusmn945 g) were obtained from Harlan laboratories (Horst TheNetherlands) The rats were fed a standard diet withoutphytogenic feed additives for an acclimatisation period of 14days
At an age of 6 weeks and a mean live weight of 2310 plusmn128 g the rats were assigned to 6 experimental groups of16 rats each During the following experimental periods thecontrol groups (Con and DSS) were fed a basal diet that metthe nutritional demands of the NRC for growing rats Thisbasal diet contained no phytogenic additive (Table 1)
The diet of group BE contained 8750mgkg broccolisprouts extract (JARROW Formulas) 1494mgkg diet ofCurcuma longa oil (Cuo) 618mgkg of Thymus vulgaris oil(To) and 680mgkg of Rosmarinus officinalis oil (Ro) wereadded to the diets of the other groups in order to standardizethe concentration of the isothiocyanate sulforaphane (BE)and of the individual main terpenes (Cuo To and Ro) toa value of 2mmolkg diet The main terpenes consideredwere ar-turmerone for Cuo thymol for To and 18-cineolfor Ro All diets were pelleted with a pellet mill using an8mm die and fed during the whole course of the experimentThe rats had free access to their respective diet and to tapwater Lighting humidity and temperature regime was inaccordance with the recommendations of the Society forLaboratory Animal Science (2004) [22] The trial consistedof 3 phases (1) pretreatment phase (phase 1 7 days) (2) DSS-treatment phase (phase 2 6 days) and (3) recovery phase(phase 3 6 days) Table 2 overviews the feeding protocol indetail
The animal model of a DSS-induced colitis was cho-sen due to several histological and biochemical similaritieswith human IBD [23 24] In the DSS-treatment phase 4DSS (40 kDa Sigma-Aldrich) was administered via drinkingwater to all rats for 6 days with the exception of the Congroup in order to induce amild intestinal inflammation Feedintake and individual live weight were recorded after oneweek daily during the DSS-phase and every other day duringthe recovery phase
At the end of phase 1 four rats were killed for organsampling (liver and colon) and after phases 2 and 3 six ratsper group were sacrificed For the histological examinationand for the determination of relative mRNA concentrationsof antioxidant and xenobiotic enzymes and of inflammationparameters colon samples were prepared from a 10 cmsegment distal to the caecum Liver samples were excisedfrom the middle of lobus sinister lateralis
During treatment with 4 DSS the disease activity index(DAI) was used to assay the severity of the induced colitisDAI was determined daily in phase 2 and every other dayin phase 3 (recovery phase) The scoring system was basedon body weight loss stool consistency andmacroscopic fecal
ISRN Gastroenterology 3
(a) (b)
Figure 1 Representative pictures of proximal colon sections from healthy Con rats (a) and DSS-treated rats (b) fed the control dietHaematoxylin and Eosin (H and E) stained colon cross-sections fromphase 2were analyzedwith inversionmicroscopy (20x) Arrows indicateincreased leucocyte infiltration and arrowheads indicate the loss of goblet cells caused by DSS treatment (b)
(a) (b)
(c) (d)
Figure 2 Representative pictures of proximal colon sections fromDSS-treated rats fed experimental diets with broccoli extract (a) turmericoil (b) thyme oil (c) or rosemary oil (d) Haematoxylin and Eosin (H and E) stained colon cross-sections from phase 2 were analyzed withinversion microscopy (20x)
4 ISRN Gastroenterology
Table 1 Basal diet
Ingredient gkg basal dietWheat (DEUKA GmbH und Co KG KonnernGermany) 2379
Maize (DEUKA GmbH und Co KG KonnernGermany) 2000
Barley (DEUKA GmbH und Co KG KonnernGermany) 1560
Soybean meal 46 CP (DEUKA GmbH und CoKG Konnern Germany) 2200
Wheat bran (DEUKA GmbH und Co KGKonnern Germany) 788
Oat (DEUKA GmbH und Co KG KonnernGermany) 690
Sun flower oil 150Lysine (Feed Grade China) 03dl-methionine (Degussa Duesseldorf Germany) 20Vitamin and mineral premix 121Calcium carbonate (Sigma-Aldrich) 25Calcium phosphate (Mischfutter und LandhandelGmbH Edderitz Germany) 79
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
I120581B120572 degradation
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
Figure 3 NF120581B and Nrf2 crosstalk under balanced anti- andprooxidant conditions
blood debris For each mentioned parameter a scale rangingfrom 0 to 4 was applied as described previously [25]
22 Colonic Histology For the histological examinationfreshly dissected colon samples were washed with 09 (wv) NaCl and cryoconserved in a freezing medium (JungLeica InstrumentsNusslochGermany) Serial cross-sections(7 120583m) were prepared using a microtome (CM 1850 UVmicrotome Jung Leica) and fixed on sterile usual micro-scope slides After staining the samples with Haematoxylin-Eosin full-thickness slices tissue architecture infiltration ofneutrophilic granulocytes into the mucosa and into the sub-mucosa and the formation of crypt abscesses were examinedunder an inversion microscope using a blind protocol
Low oxid stress electrophils and
phytogenic substanceseg SFN curcumin120581BE-driven gene
transkription eg IL-1120573I120581B120572 Keap1
(Cul3-Rbx1)
Nrf2
+
+
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
I120581B120572 degradation
Figure 4 NF120581B and Nrf2 interaction due to feeding phytogeniccompounds or low oxidative stress
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
+
+
+
+
++
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
P
IKK
Progressiveoxid stress
eg DSS H2O2
I120581B120572 degradation
Figure 5 NF120581B and Nrf2 interaction due to DSS treatment orprogressive oxidative stress
23 RNA Preparation and Real-Time RT-PCR Analysis TotalRNA from 100mg of liver and colon tissue was isolatedusing the acid guanidinium thiocyanate-phenol-chloroformextraction method [26] Most recently a strong DSS RNAinterference during acute DSS-treatment resulting in thelack of signals during gene expression analysis has beendescribed [27] To assure the correctness of gene expressionanalyses in our experiment polyA+ mRNA from the colonicsamples of the DSS-phase was purified using the GenElutemRNA Miniprep Kit (Sigma-Aldrich MO USA) accordingto the manufacturersrsquo protocol Following the photometricaldetermination of RNA concentration and purity at 260 nmand 280 nm reverse transcription of 30 120583g of total RNAor of 03 120583g of purified mRNA and real-time RT-PCR wereperformed as described previously [28] Gene bank accession
ISRN Gastroenterology 5
Table 2 Feeding protocol
Group Phytogenicadditive
Koncentration per kgdiet Phase 1 Phase 2 Phase 3
Con None mdash 7 days diets and water adlibitum
6 days diets and water adlibitum
6 days diets and water adlibitum
DSS None mdash 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
BE Broccoli extract 2mmol sulforaphane 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Cuo Turmeric oil 2mmol ar-turmerone 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
To Thyme oil 2mmol thymol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Ro Rosemary oil 2mmol 18-cineol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
numbers and primer sequences (51015840 rarr 31015840) are shown inTable 3
Gene specific mRNA expression was analyzed with theRotor-Gene 6000 series software using the ΔΔCt method[29] The amplification data of the single genes were nor-malized to the expression of the two most stable refer-ence genes in each tissue (liver 120573-actin ribosomal proteinL13A (Rpl13a) colon Rpl13a hypoxanthine phosphoribosyl-transferase 1 (Hprt1)) Relative mRNA expression levels areexpressed as x-fold changes relative to group Con = 10
24 Statistical Analysis Data are presented as means plusmn theirstandard error of the mean (SEM) Statistical differenceswere analyzed with SPSS 190 for Windows (IBM ChicagoUSA) using one-way ANOVA after verifying the normalityof distribution (Shapiro Wilk test and Kolmogorov Smirnovtest) and the homogeneity of variances (Levene test) TheLeast Significant Difference test (LSD) was used to ana-lyze significant differences between means if variances werehomogenous If not the Games Howell test was used Atan error probability of less than 5 (119875 lt 005) differencesbetween means were considered as statistically significant
3 Results
31 Body Weight Development Neither food intake (datanot shown) nor final body weight (Table 4) of the rats wasinfluenced significantly in the different experimental phasesby feeding the specific diets tested Remarkably also DSStreatment in the second phase did not affect the above-mentioned parameters significantly
32 Colitis Severity by Disease Activity Index (DAI) Thecourse of the mild colitis induced by the administration of4 DSS to rats for 6 days was controlled daily in phase2 by measuring the DAI A high number of rats from allDSS-treated groups had a soft stool consistency but severediarrhea accompanied by blood debris could not be observedMoreover a distinct weight loss frequently observed underDSS treatment was present only in some cases and only for
one day Consequently the DAI in all DSS-treated groupsremained below 10 (DSS 012 BE 010 Cuo 010 To007 and Ro 013) and did not differ significantly fromCon rats receiving no DSS In phase 3 obvious changesin stool consistency almost disappeared or were much lesspronounced than in phase 2
33 Histology DSS treatment caused no significant observ-able macroscopic changes in colonic tissue architecturebetween the experimental groups including the untreatedCon rats Furthermore microscopic analysis revealed nosignificant histological damage to the colonic mucosa of ratsexposed to 4 DSS for 6 days However DSS treatmenttended to accelerate initial damage to the mucosa charac-terised by the loss of goblet cells and the occurrence of amorediffuse crypt architecture compared to the colon of healthyCon rats Additionally the accumulation of neutrophils infil-trating the lamina propria could be observedmore frequentlyin colonic slices of DSS-treated rats These mentioned DSS-induced mucosal alterations are shown in Figure 1 DSStreatment in combination with the tested phytogenic extractsalso caused slightmucosal damage (Figure 2) but the severityseemed to be much lower than in group DSS (Figure 1(b))
34 mRNA Expression of NF120581B TNF120572 and VariousInflammatory-Mediating Enzymes in Colon and Liver TissueThe analyzed mRNA expression patterns of pro- and anti-inflammatory genes controlled by NF120581B and TNF120572 differedamong the experimental periods and the tissues investigated(Tables 5 and 6) During the 7-day pretreatment phase 1 themRNA abundance of the colonic pro-inflammatory markersCOX2 and IL-1120573 of the anti-inflammatory cytokine IL-10 ofthe cell adhesion molecules MCP-1 and VCAM-1 and of thetight junction protein Cldn3 showed a high intraindividualvariance In contrast in phase 1 of the trial liver mRNAdata of the above-mentioned genes showed a much betterhomogeneity Nevertheless feeding diets with BE- Cuo-To- and Ro-addition reduced colonic VCAM-1 mRNA by 57to 64 compared to untreated control rats This reductionwas significant for Cuo To and Ro and represented a trend(119875 lt 010) for BE Moreover in phase 1 colonic Cldn3
6 ISRN Gastroenterology
Table 3 Gene bank accession numbers and primer sequences of the genes investigated by real-time RT-PCR
Gene name (abbreviation used) Gene bank accession number Primer sequences (51015840 rarr 31015840)for = forward rev = reverse
Chemokine (C-C motif) ligand 2 (Ccl2) (MCP1) NM 031530 for GTGCGACCCCAATAAGGAArev TGAGGTGGTTGTGGAAAAGA
Claudin 3 (Cldn3) NM 031700 for TATCCTACTGGCAGCCTTCGrev GTTCCCATCTCTCGCTTCTG
Copperzinc superoxide dismutase (SOD1) NM 017050 for CCACTGCAGGACCTCATTTTrev CACCTTTGCCCAAGTCATCT
C-reactive protein (CRP) NM 017096 for GTCTCTATGCCCACGCTGATrev CCGTCAAGCCAAAGCTCTAC
Glutathione S-transferase K1 (GSTK1) NM 181371 for GAGCATGGAGCAACCAGAGATrev AGCTTGCTCTTCACCAGTTCG
Glutathione S-transferase P1 (GSTP1) NM 012577 for GAGGCAAAGCTTTCATTGTGGrev GTTGATGGGACGGTTCAAATG
Glutathione S-transferase T2 (GSTT2) NM 012796 for GAGGAAAAGGTGGAACGGAACrev CGCCCCTCAAACAGATTACAG
Glutathione peroxidase 2 (GPx2) NM 183402 for GTGTGATGTCAATGGGCAGAArev ACGTTTGATGTCAGGCTCGAT
Heme oxygenase 1 (HO1) NM 012580 for AGGCACTGCTGACAGAGGAACrev AGCGGTGTCTGGGATGAACTA
Hypoxanthine phosphoribosyltransferase 1 (Hprt1) NM 012583 for GCAGACTTTGCTTTCCTTGGrev TCCACTTTCGCTGATGACAC
Interleukin 1 beta (IL-1120573) NM 031512 for CTGTGACTCGTGGGATGATGrev GGGATTTTGTCGTTGCTTGT
Interleukin 10 (IL-10) NM 012854 for CTGGAGTGAAGACCAGCAAAGGrev GGAGAAATCGATGACAGCGTCG
Kelch-like ECH-associated protein1 (Keap1) NM 057152 for GTGGCGGATGATTACACCAATrev GAAAAGTGTGGCCATCGTAGC
NAD(P)H dehydrogenase [quinone] 1 (NQO1) NM 017000 for CGCAGAGAGGACATCATTCArev CGCCAGAGATGACTCAACAG
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) NM 031789 for CCAAGGAGCAATTCAACGAAGrev CTCTTGGGAACAAGGAACACG
Nuclear factor kappa B (NF120581B) L26267 for CTTCTCGGAGTCCCTCACTGrev CCAATAGCAGCTGGAAAAGC
Prostaglandin-endoperoxide synthase 2 (Ptgs2) (COX2) NM 017232 for GCTGTACAAGCAGTGGCAAArev CCCCAAAGACAGCATCTGGA
Ribosomal protein L13A (Rpl13a) NM 173340 for CCCTCCACCCTATGACAAGArev CCTTTTCCTTCCGTTTCTCC
Tumor necrosis factor alpha (TNF120572) NM 012675 for GCCAATGGCATGGATCTCAAAGrev AAATCGGCTGACGGTGTGGG
Vascular cell adhesion molecule 1 (VCAM 1) NM 012889 for TGACATCTCCCCTGGATCTCrev CTCCAGTTTCCTTCGCTGAC
120573-actin NM 031144 for ATCGTGCGTGACATTAAAGAGAAGrev GGACAGTGAGGCCAGGATAGAG
expression was significantly higher in Ro-treated rats andtendencially higher in Cuo- and To-treated rats than in theirCon littermates
In phase 1 feeding diets containing BE Cuo and Rosignificantly decreased liver mRNA abundance of the pro-inflammatory cytokine IL-10 by 32 to 44
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
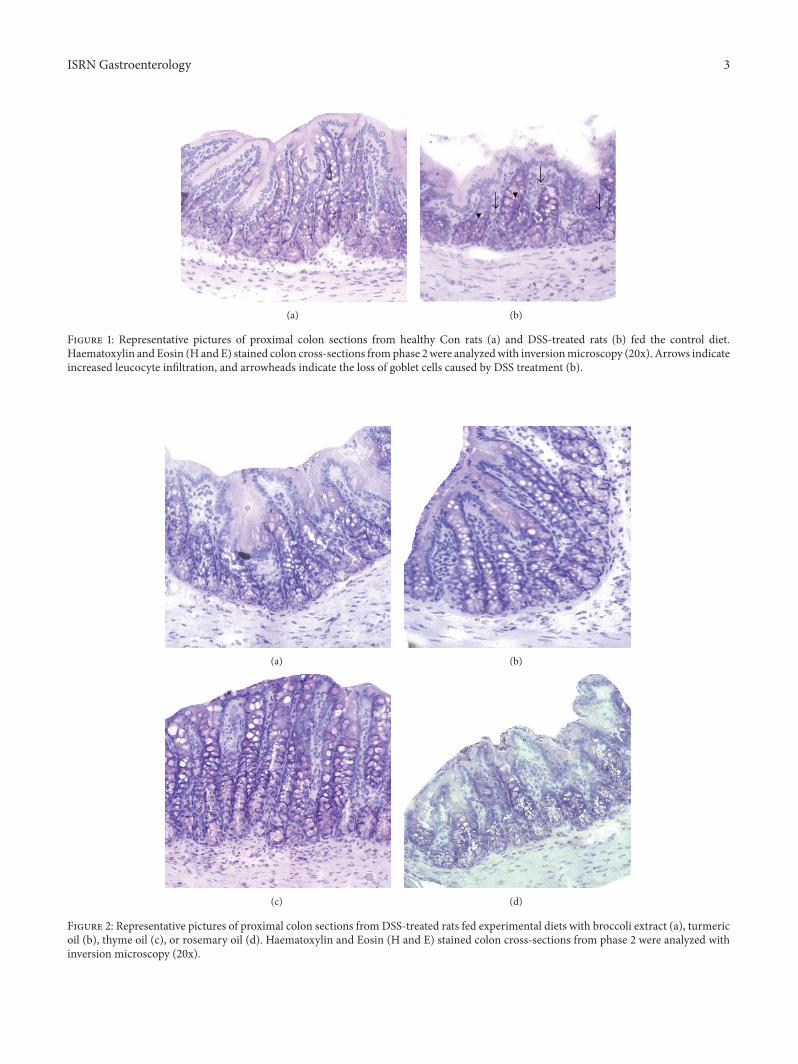
ISRN Gastroenterology 3
(a) (b)
Figure 1 Representative pictures of proximal colon sections from healthy Con rats (a) and DSS-treated rats (b) fed the control dietHaematoxylin and Eosin (H and E) stained colon cross-sections fromphase 2were analyzedwith inversionmicroscopy (20x) Arrows indicateincreased leucocyte infiltration and arrowheads indicate the loss of goblet cells caused by DSS treatment (b)
(a) (b)
(c) (d)
Figure 2 Representative pictures of proximal colon sections fromDSS-treated rats fed experimental diets with broccoli extract (a) turmericoil (b) thyme oil (c) or rosemary oil (d) Haematoxylin and Eosin (H and E) stained colon cross-sections from phase 2 were analyzed withinversion microscopy (20x)
4 ISRN Gastroenterology
Table 1 Basal diet
Ingredient gkg basal dietWheat (DEUKA GmbH und Co KG KonnernGermany) 2379
Maize (DEUKA GmbH und Co KG KonnernGermany) 2000
Barley (DEUKA GmbH und Co KG KonnernGermany) 1560
Soybean meal 46 CP (DEUKA GmbH und CoKG Konnern Germany) 2200
Wheat bran (DEUKA GmbH und Co KGKonnern Germany) 788
Oat (DEUKA GmbH und Co KG KonnernGermany) 690
Sun flower oil 150Lysine (Feed Grade China) 03dl-methionine (Degussa Duesseldorf Germany) 20Vitamin and mineral premix 121Calcium carbonate (Sigma-Aldrich) 25Calcium phosphate (Mischfutter und LandhandelGmbH Edderitz Germany) 79
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
I120581B120572 degradation
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
Figure 3 NF120581B and Nrf2 crosstalk under balanced anti- andprooxidant conditions
blood debris For each mentioned parameter a scale rangingfrom 0 to 4 was applied as described previously [25]
22 Colonic Histology For the histological examinationfreshly dissected colon samples were washed with 09 (wv) NaCl and cryoconserved in a freezing medium (JungLeica InstrumentsNusslochGermany) Serial cross-sections(7 120583m) were prepared using a microtome (CM 1850 UVmicrotome Jung Leica) and fixed on sterile usual micro-scope slides After staining the samples with Haematoxylin-Eosin full-thickness slices tissue architecture infiltration ofneutrophilic granulocytes into the mucosa and into the sub-mucosa and the formation of crypt abscesses were examinedunder an inversion microscope using a blind protocol
Low oxid stress electrophils and
phytogenic substanceseg SFN curcumin120581BE-driven gene
transkription eg IL-1120573I120581B120572 Keap1
(Cul3-Rbx1)
Nrf2
+
+
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
I120581B120572 degradation
Figure 4 NF120581B and Nrf2 interaction due to feeding phytogeniccompounds or low oxidative stress
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
+
+
+
+
++
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
P
IKK
Progressiveoxid stress
eg DSS H2O2
I120581B120572 degradation
Figure 5 NF120581B and Nrf2 interaction due to DSS treatment orprogressive oxidative stress
23 RNA Preparation and Real-Time RT-PCR Analysis TotalRNA from 100mg of liver and colon tissue was isolatedusing the acid guanidinium thiocyanate-phenol-chloroformextraction method [26] Most recently a strong DSS RNAinterference during acute DSS-treatment resulting in thelack of signals during gene expression analysis has beendescribed [27] To assure the correctness of gene expressionanalyses in our experiment polyA+ mRNA from the colonicsamples of the DSS-phase was purified using the GenElutemRNA Miniprep Kit (Sigma-Aldrich MO USA) accordingto the manufacturersrsquo protocol Following the photometricaldetermination of RNA concentration and purity at 260 nmand 280 nm reverse transcription of 30 120583g of total RNAor of 03 120583g of purified mRNA and real-time RT-PCR wereperformed as described previously [28] Gene bank accession
ISRN Gastroenterology 5
Table 2 Feeding protocol
Group Phytogenicadditive
Koncentration per kgdiet Phase 1 Phase 2 Phase 3
Con None mdash 7 days diets and water adlibitum
6 days diets and water adlibitum
6 days diets and water adlibitum
DSS None mdash 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
BE Broccoli extract 2mmol sulforaphane 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Cuo Turmeric oil 2mmol ar-turmerone 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
To Thyme oil 2mmol thymol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Ro Rosemary oil 2mmol 18-cineol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
numbers and primer sequences (51015840 rarr 31015840) are shown inTable 3
Gene specific mRNA expression was analyzed with theRotor-Gene 6000 series software using the ΔΔCt method[29] The amplification data of the single genes were nor-malized to the expression of the two most stable refer-ence genes in each tissue (liver 120573-actin ribosomal proteinL13A (Rpl13a) colon Rpl13a hypoxanthine phosphoribosyl-transferase 1 (Hprt1)) Relative mRNA expression levels areexpressed as x-fold changes relative to group Con = 10
24 Statistical Analysis Data are presented as means plusmn theirstandard error of the mean (SEM) Statistical differenceswere analyzed with SPSS 190 for Windows (IBM ChicagoUSA) using one-way ANOVA after verifying the normalityof distribution (Shapiro Wilk test and Kolmogorov Smirnovtest) and the homogeneity of variances (Levene test) TheLeast Significant Difference test (LSD) was used to ana-lyze significant differences between means if variances werehomogenous If not the Games Howell test was used Atan error probability of less than 5 (119875 lt 005) differencesbetween means were considered as statistically significant
3 Results
31 Body Weight Development Neither food intake (datanot shown) nor final body weight (Table 4) of the rats wasinfluenced significantly in the different experimental phasesby feeding the specific diets tested Remarkably also DSStreatment in the second phase did not affect the above-mentioned parameters significantly
32 Colitis Severity by Disease Activity Index (DAI) Thecourse of the mild colitis induced by the administration of4 DSS to rats for 6 days was controlled daily in phase2 by measuring the DAI A high number of rats from allDSS-treated groups had a soft stool consistency but severediarrhea accompanied by blood debris could not be observedMoreover a distinct weight loss frequently observed underDSS treatment was present only in some cases and only for
one day Consequently the DAI in all DSS-treated groupsremained below 10 (DSS 012 BE 010 Cuo 010 To007 and Ro 013) and did not differ significantly fromCon rats receiving no DSS In phase 3 obvious changesin stool consistency almost disappeared or were much lesspronounced than in phase 2
33 Histology DSS treatment caused no significant observ-able macroscopic changes in colonic tissue architecturebetween the experimental groups including the untreatedCon rats Furthermore microscopic analysis revealed nosignificant histological damage to the colonic mucosa of ratsexposed to 4 DSS for 6 days However DSS treatmenttended to accelerate initial damage to the mucosa charac-terised by the loss of goblet cells and the occurrence of amorediffuse crypt architecture compared to the colon of healthyCon rats Additionally the accumulation of neutrophils infil-trating the lamina propria could be observedmore frequentlyin colonic slices of DSS-treated rats These mentioned DSS-induced mucosal alterations are shown in Figure 1 DSStreatment in combination with the tested phytogenic extractsalso caused slightmucosal damage (Figure 2) but the severityseemed to be much lower than in group DSS (Figure 1(b))
34 mRNA Expression of NF120581B TNF120572 and VariousInflammatory-Mediating Enzymes in Colon and Liver TissueThe analyzed mRNA expression patterns of pro- and anti-inflammatory genes controlled by NF120581B and TNF120572 differedamong the experimental periods and the tissues investigated(Tables 5 and 6) During the 7-day pretreatment phase 1 themRNA abundance of the colonic pro-inflammatory markersCOX2 and IL-1120573 of the anti-inflammatory cytokine IL-10 ofthe cell adhesion molecules MCP-1 and VCAM-1 and of thetight junction protein Cldn3 showed a high intraindividualvariance In contrast in phase 1 of the trial liver mRNAdata of the above-mentioned genes showed a much betterhomogeneity Nevertheless feeding diets with BE- Cuo-To- and Ro-addition reduced colonic VCAM-1 mRNA by 57to 64 compared to untreated control rats This reductionwas significant for Cuo To and Ro and represented a trend(119875 lt 010) for BE Moreover in phase 1 colonic Cldn3
6 ISRN Gastroenterology
Table 3 Gene bank accession numbers and primer sequences of the genes investigated by real-time RT-PCR
Gene name (abbreviation used) Gene bank accession number Primer sequences (51015840 rarr 31015840)for = forward rev = reverse
Chemokine (C-C motif) ligand 2 (Ccl2) (MCP1) NM 031530 for GTGCGACCCCAATAAGGAArev TGAGGTGGTTGTGGAAAAGA
Claudin 3 (Cldn3) NM 031700 for TATCCTACTGGCAGCCTTCGrev GTTCCCATCTCTCGCTTCTG
Copperzinc superoxide dismutase (SOD1) NM 017050 for CCACTGCAGGACCTCATTTTrev CACCTTTGCCCAAGTCATCT
C-reactive protein (CRP) NM 017096 for GTCTCTATGCCCACGCTGATrev CCGTCAAGCCAAAGCTCTAC
Glutathione S-transferase K1 (GSTK1) NM 181371 for GAGCATGGAGCAACCAGAGATrev AGCTTGCTCTTCACCAGTTCG
Glutathione S-transferase P1 (GSTP1) NM 012577 for GAGGCAAAGCTTTCATTGTGGrev GTTGATGGGACGGTTCAAATG
Glutathione S-transferase T2 (GSTT2) NM 012796 for GAGGAAAAGGTGGAACGGAACrev CGCCCCTCAAACAGATTACAG
Glutathione peroxidase 2 (GPx2) NM 183402 for GTGTGATGTCAATGGGCAGAArev ACGTTTGATGTCAGGCTCGAT
Heme oxygenase 1 (HO1) NM 012580 for AGGCACTGCTGACAGAGGAACrev AGCGGTGTCTGGGATGAACTA
Hypoxanthine phosphoribosyltransferase 1 (Hprt1) NM 012583 for GCAGACTTTGCTTTCCTTGGrev TCCACTTTCGCTGATGACAC
Interleukin 1 beta (IL-1120573) NM 031512 for CTGTGACTCGTGGGATGATGrev GGGATTTTGTCGTTGCTTGT
Interleukin 10 (IL-10) NM 012854 for CTGGAGTGAAGACCAGCAAAGGrev GGAGAAATCGATGACAGCGTCG
Kelch-like ECH-associated protein1 (Keap1) NM 057152 for GTGGCGGATGATTACACCAATrev GAAAAGTGTGGCCATCGTAGC
NAD(P)H dehydrogenase [quinone] 1 (NQO1) NM 017000 for CGCAGAGAGGACATCATTCArev CGCCAGAGATGACTCAACAG
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) NM 031789 for CCAAGGAGCAATTCAACGAAGrev CTCTTGGGAACAAGGAACACG
Nuclear factor kappa B (NF120581B) L26267 for CTTCTCGGAGTCCCTCACTGrev CCAATAGCAGCTGGAAAAGC
Prostaglandin-endoperoxide synthase 2 (Ptgs2) (COX2) NM 017232 for GCTGTACAAGCAGTGGCAAArev CCCCAAAGACAGCATCTGGA
Ribosomal protein L13A (Rpl13a) NM 173340 for CCCTCCACCCTATGACAAGArev CCTTTTCCTTCCGTTTCTCC
Tumor necrosis factor alpha (TNF120572) NM 012675 for GCCAATGGCATGGATCTCAAAGrev AAATCGGCTGACGGTGTGGG
Vascular cell adhesion molecule 1 (VCAM 1) NM 012889 for TGACATCTCCCCTGGATCTCrev CTCCAGTTTCCTTCGCTGAC
120573-actin NM 031144 for ATCGTGCGTGACATTAAAGAGAAGrev GGACAGTGAGGCCAGGATAGAG
expression was significantly higher in Ro-treated rats andtendencially higher in Cuo- and To-treated rats than in theirCon littermates
In phase 1 feeding diets containing BE Cuo and Rosignificantly decreased liver mRNA abundance of the pro-inflammatory cytokine IL-10 by 32 to 44
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 ISRN Gastroenterology
Table 1 Basal diet
Ingredient gkg basal dietWheat (DEUKA GmbH und Co KG KonnernGermany) 2379
Maize (DEUKA GmbH und Co KG KonnernGermany) 2000
Barley (DEUKA GmbH und Co KG KonnernGermany) 1560
Soybean meal 46 CP (DEUKA GmbH und CoKG Konnern Germany) 2200
Wheat bran (DEUKA GmbH und Co KGKonnern Germany) 788
Oat (DEUKA GmbH und Co KG KonnernGermany) 690
Sun flower oil 150Lysine (Feed Grade China) 03dl-methionine (Degussa Duesseldorf Germany) 20Vitamin and mineral premix 121Calcium carbonate (Sigma-Aldrich) 25Calcium phosphate (Mischfutter und LandhandelGmbH Edderitz Germany) 79
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
I120581B120572 degradation
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
Figure 3 NF120581B and Nrf2 crosstalk under balanced anti- andprooxidant conditions
blood debris For each mentioned parameter a scale rangingfrom 0 to 4 was applied as described previously [25]
22 Colonic Histology For the histological examinationfreshly dissected colon samples were washed with 09 (wv) NaCl and cryoconserved in a freezing medium (JungLeica InstrumentsNusslochGermany) Serial cross-sections(7 120583m) were prepared using a microtome (CM 1850 UVmicrotome Jung Leica) and fixed on sterile usual micro-scope slides After staining the samples with Haematoxylin-Eosin full-thickness slices tissue architecture infiltration ofneutrophilic granulocytes into the mucosa and into the sub-mucosa and the formation of crypt abscesses were examinedunder an inversion microscope using a blind protocol
Low oxid stress electrophils and
phytogenic substanceseg SFN curcumin120581BE-driven gene
transkription eg IL-1120573I120581B120572 Keap1
(Cul3-Rbx1)
Nrf2
+
+
+
+
+
+
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
I120581B120572 degradation
Figure 4 NF120581B and Nrf2 interaction due to feeding phytogeniccompounds or low oxidative stress
120581BE-driven genetranskription eg IL-1120573
I120581B120572 Keap1(Cul3-Rbx1)
Nrf2
+
+
+
+
++
minus
ARE-driven genetranskription eg
GSTs GPx2NQO1 and HO1
TNF120572 and VCAM-1
TNF120572
NF120581B
P
IKK
Progressiveoxid stress
eg DSS H2O2
I120581B120572 degradation
Figure 5 NF120581B and Nrf2 interaction due to DSS treatment orprogressive oxidative stress
23 RNA Preparation and Real-Time RT-PCR Analysis TotalRNA from 100mg of liver and colon tissue was isolatedusing the acid guanidinium thiocyanate-phenol-chloroformextraction method [26] Most recently a strong DSS RNAinterference during acute DSS-treatment resulting in thelack of signals during gene expression analysis has beendescribed [27] To assure the correctness of gene expressionanalyses in our experiment polyA+ mRNA from the colonicsamples of the DSS-phase was purified using the GenElutemRNA Miniprep Kit (Sigma-Aldrich MO USA) accordingto the manufacturersrsquo protocol Following the photometricaldetermination of RNA concentration and purity at 260 nmand 280 nm reverse transcription of 30 120583g of total RNAor of 03 120583g of purified mRNA and real-time RT-PCR wereperformed as described previously [28] Gene bank accession
ISRN Gastroenterology 5
Table 2 Feeding protocol
Group Phytogenicadditive
Koncentration per kgdiet Phase 1 Phase 2 Phase 3
Con None mdash 7 days diets and water adlibitum
6 days diets and water adlibitum
6 days diets and water adlibitum
DSS None mdash 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
BE Broccoli extract 2mmol sulforaphane 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Cuo Turmeric oil 2mmol ar-turmerone 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
To Thyme oil 2mmol thymol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Ro Rosemary oil 2mmol 18-cineol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
numbers and primer sequences (51015840 rarr 31015840) are shown inTable 3
Gene specific mRNA expression was analyzed with theRotor-Gene 6000 series software using the ΔΔCt method[29] The amplification data of the single genes were nor-malized to the expression of the two most stable refer-ence genes in each tissue (liver 120573-actin ribosomal proteinL13A (Rpl13a) colon Rpl13a hypoxanthine phosphoribosyl-transferase 1 (Hprt1)) Relative mRNA expression levels areexpressed as x-fold changes relative to group Con = 10
24 Statistical Analysis Data are presented as means plusmn theirstandard error of the mean (SEM) Statistical differenceswere analyzed with SPSS 190 for Windows (IBM ChicagoUSA) using one-way ANOVA after verifying the normalityof distribution (Shapiro Wilk test and Kolmogorov Smirnovtest) and the homogeneity of variances (Levene test) TheLeast Significant Difference test (LSD) was used to ana-lyze significant differences between means if variances werehomogenous If not the Games Howell test was used Atan error probability of less than 5 (119875 lt 005) differencesbetween means were considered as statistically significant
3 Results
31 Body Weight Development Neither food intake (datanot shown) nor final body weight (Table 4) of the rats wasinfluenced significantly in the different experimental phasesby feeding the specific diets tested Remarkably also DSStreatment in the second phase did not affect the above-mentioned parameters significantly
32 Colitis Severity by Disease Activity Index (DAI) Thecourse of the mild colitis induced by the administration of4 DSS to rats for 6 days was controlled daily in phase2 by measuring the DAI A high number of rats from allDSS-treated groups had a soft stool consistency but severediarrhea accompanied by blood debris could not be observedMoreover a distinct weight loss frequently observed underDSS treatment was present only in some cases and only for
one day Consequently the DAI in all DSS-treated groupsremained below 10 (DSS 012 BE 010 Cuo 010 To007 and Ro 013) and did not differ significantly fromCon rats receiving no DSS In phase 3 obvious changesin stool consistency almost disappeared or were much lesspronounced than in phase 2
33 Histology DSS treatment caused no significant observ-able macroscopic changes in colonic tissue architecturebetween the experimental groups including the untreatedCon rats Furthermore microscopic analysis revealed nosignificant histological damage to the colonic mucosa of ratsexposed to 4 DSS for 6 days However DSS treatmenttended to accelerate initial damage to the mucosa charac-terised by the loss of goblet cells and the occurrence of amorediffuse crypt architecture compared to the colon of healthyCon rats Additionally the accumulation of neutrophils infil-trating the lamina propria could be observedmore frequentlyin colonic slices of DSS-treated rats These mentioned DSS-induced mucosal alterations are shown in Figure 1 DSStreatment in combination with the tested phytogenic extractsalso caused slightmucosal damage (Figure 2) but the severityseemed to be much lower than in group DSS (Figure 1(b))
34 mRNA Expression of NF120581B TNF120572 and VariousInflammatory-Mediating Enzymes in Colon and Liver TissueThe analyzed mRNA expression patterns of pro- and anti-inflammatory genes controlled by NF120581B and TNF120572 differedamong the experimental periods and the tissues investigated(Tables 5 and 6) During the 7-day pretreatment phase 1 themRNA abundance of the colonic pro-inflammatory markersCOX2 and IL-1120573 of the anti-inflammatory cytokine IL-10 ofthe cell adhesion molecules MCP-1 and VCAM-1 and of thetight junction protein Cldn3 showed a high intraindividualvariance In contrast in phase 1 of the trial liver mRNAdata of the above-mentioned genes showed a much betterhomogeneity Nevertheless feeding diets with BE- Cuo-To- and Ro-addition reduced colonic VCAM-1 mRNA by 57to 64 compared to untreated control rats This reductionwas significant for Cuo To and Ro and represented a trend(119875 lt 010) for BE Moreover in phase 1 colonic Cldn3
6 ISRN Gastroenterology
Table 3 Gene bank accession numbers and primer sequences of the genes investigated by real-time RT-PCR
Gene name (abbreviation used) Gene bank accession number Primer sequences (51015840 rarr 31015840)for = forward rev = reverse
Chemokine (C-C motif) ligand 2 (Ccl2) (MCP1) NM 031530 for GTGCGACCCCAATAAGGAArev TGAGGTGGTTGTGGAAAAGA
Claudin 3 (Cldn3) NM 031700 for TATCCTACTGGCAGCCTTCGrev GTTCCCATCTCTCGCTTCTG
Copperzinc superoxide dismutase (SOD1) NM 017050 for CCACTGCAGGACCTCATTTTrev CACCTTTGCCCAAGTCATCT
C-reactive protein (CRP) NM 017096 for GTCTCTATGCCCACGCTGATrev CCGTCAAGCCAAAGCTCTAC
Glutathione S-transferase K1 (GSTK1) NM 181371 for GAGCATGGAGCAACCAGAGATrev AGCTTGCTCTTCACCAGTTCG
Glutathione S-transferase P1 (GSTP1) NM 012577 for GAGGCAAAGCTTTCATTGTGGrev GTTGATGGGACGGTTCAAATG
Glutathione S-transferase T2 (GSTT2) NM 012796 for GAGGAAAAGGTGGAACGGAACrev CGCCCCTCAAACAGATTACAG
Glutathione peroxidase 2 (GPx2) NM 183402 for GTGTGATGTCAATGGGCAGAArev ACGTTTGATGTCAGGCTCGAT
Heme oxygenase 1 (HO1) NM 012580 for AGGCACTGCTGACAGAGGAACrev AGCGGTGTCTGGGATGAACTA
Hypoxanthine phosphoribosyltransferase 1 (Hprt1) NM 012583 for GCAGACTTTGCTTTCCTTGGrev TCCACTTTCGCTGATGACAC
Interleukin 1 beta (IL-1120573) NM 031512 for CTGTGACTCGTGGGATGATGrev GGGATTTTGTCGTTGCTTGT
Interleukin 10 (IL-10) NM 012854 for CTGGAGTGAAGACCAGCAAAGGrev GGAGAAATCGATGACAGCGTCG
Kelch-like ECH-associated protein1 (Keap1) NM 057152 for GTGGCGGATGATTACACCAATrev GAAAAGTGTGGCCATCGTAGC
NAD(P)H dehydrogenase [quinone] 1 (NQO1) NM 017000 for CGCAGAGAGGACATCATTCArev CGCCAGAGATGACTCAACAG
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) NM 031789 for CCAAGGAGCAATTCAACGAAGrev CTCTTGGGAACAAGGAACACG
Nuclear factor kappa B (NF120581B) L26267 for CTTCTCGGAGTCCCTCACTGrev CCAATAGCAGCTGGAAAAGC
Prostaglandin-endoperoxide synthase 2 (Ptgs2) (COX2) NM 017232 for GCTGTACAAGCAGTGGCAAArev CCCCAAAGACAGCATCTGGA
Ribosomal protein L13A (Rpl13a) NM 173340 for CCCTCCACCCTATGACAAGArev CCTTTTCCTTCCGTTTCTCC
Tumor necrosis factor alpha (TNF120572) NM 012675 for GCCAATGGCATGGATCTCAAAGrev AAATCGGCTGACGGTGTGGG
Vascular cell adhesion molecule 1 (VCAM 1) NM 012889 for TGACATCTCCCCTGGATCTCrev CTCCAGTTTCCTTCGCTGAC
120573-actin NM 031144 for ATCGTGCGTGACATTAAAGAGAAGrev GGACAGTGAGGCCAGGATAGAG
expression was significantly higher in Ro-treated rats andtendencially higher in Cuo- and To-treated rats than in theirCon littermates
In phase 1 feeding diets containing BE Cuo and Rosignificantly decreased liver mRNA abundance of the pro-inflammatory cytokine IL-10 by 32 to 44
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
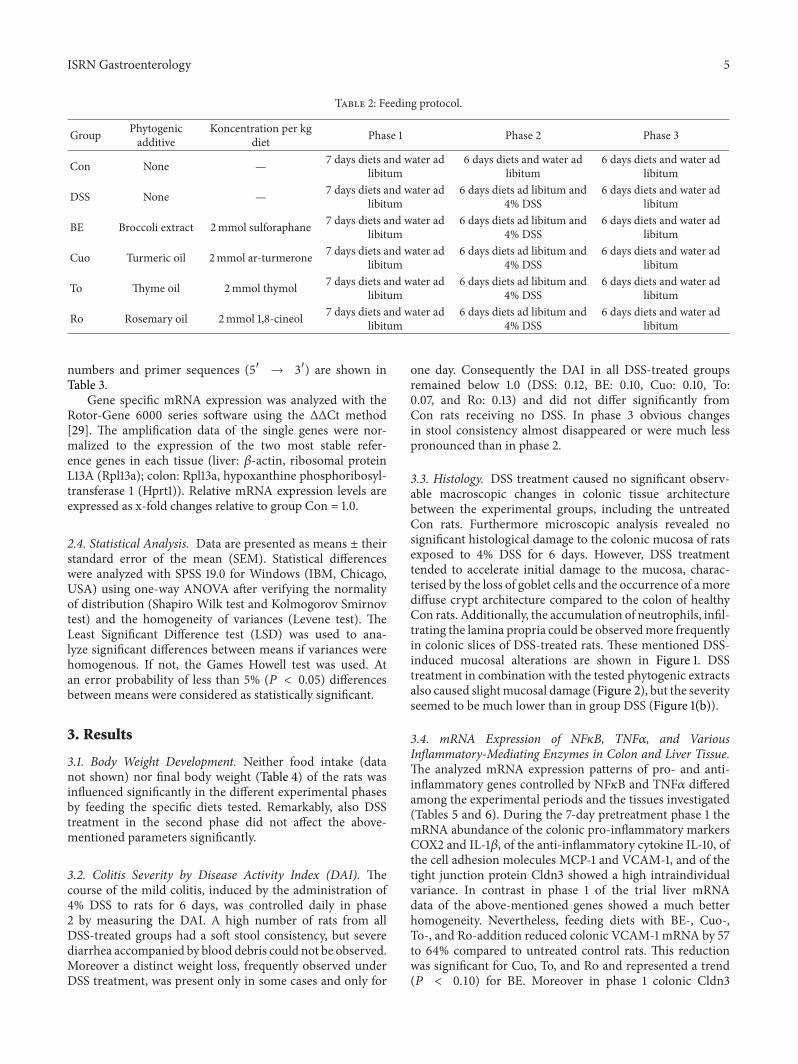
ISRN Gastroenterology 5
Table 2 Feeding protocol
Group Phytogenicadditive
Koncentration per kgdiet Phase 1 Phase 2 Phase 3
Con None mdash 7 days diets and water adlibitum
6 days diets and water adlibitum
6 days diets and water adlibitum
DSS None mdash 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
BE Broccoli extract 2mmol sulforaphane 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Cuo Turmeric oil 2mmol ar-turmerone 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
To Thyme oil 2mmol thymol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
Ro Rosemary oil 2mmol 18-cineol 7 days diets and water adlibitum
6 days diets ad libitum and4 DSS
6 days diets and water adlibitum
numbers and primer sequences (51015840 rarr 31015840) are shown inTable 3
Gene specific mRNA expression was analyzed with theRotor-Gene 6000 series software using the ΔΔCt method[29] The amplification data of the single genes were nor-malized to the expression of the two most stable refer-ence genes in each tissue (liver 120573-actin ribosomal proteinL13A (Rpl13a) colon Rpl13a hypoxanthine phosphoribosyl-transferase 1 (Hprt1)) Relative mRNA expression levels areexpressed as x-fold changes relative to group Con = 10
24 Statistical Analysis Data are presented as means plusmn theirstandard error of the mean (SEM) Statistical differenceswere analyzed with SPSS 190 for Windows (IBM ChicagoUSA) using one-way ANOVA after verifying the normalityof distribution (Shapiro Wilk test and Kolmogorov Smirnovtest) and the homogeneity of variances (Levene test) TheLeast Significant Difference test (LSD) was used to ana-lyze significant differences between means if variances werehomogenous If not the Games Howell test was used Atan error probability of less than 5 (119875 lt 005) differencesbetween means were considered as statistically significant
3 Results
31 Body Weight Development Neither food intake (datanot shown) nor final body weight (Table 4) of the rats wasinfluenced significantly in the different experimental phasesby feeding the specific diets tested Remarkably also DSStreatment in the second phase did not affect the above-mentioned parameters significantly
32 Colitis Severity by Disease Activity Index (DAI) Thecourse of the mild colitis induced by the administration of4 DSS to rats for 6 days was controlled daily in phase2 by measuring the DAI A high number of rats from allDSS-treated groups had a soft stool consistency but severediarrhea accompanied by blood debris could not be observedMoreover a distinct weight loss frequently observed underDSS treatment was present only in some cases and only for
one day Consequently the DAI in all DSS-treated groupsremained below 10 (DSS 012 BE 010 Cuo 010 To007 and Ro 013) and did not differ significantly fromCon rats receiving no DSS In phase 3 obvious changesin stool consistency almost disappeared or were much lesspronounced than in phase 2
33 Histology DSS treatment caused no significant observ-able macroscopic changes in colonic tissue architecturebetween the experimental groups including the untreatedCon rats Furthermore microscopic analysis revealed nosignificant histological damage to the colonic mucosa of ratsexposed to 4 DSS for 6 days However DSS treatmenttended to accelerate initial damage to the mucosa charac-terised by the loss of goblet cells and the occurrence of amorediffuse crypt architecture compared to the colon of healthyCon rats Additionally the accumulation of neutrophils infil-trating the lamina propria could be observedmore frequentlyin colonic slices of DSS-treated rats These mentioned DSS-induced mucosal alterations are shown in Figure 1 DSStreatment in combination with the tested phytogenic extractsalso caused slightmucosal damage (Figure 2) but the severityseemed to be much lower than in group DSS (Figure 1(b))
34 mRNA Expression of NF120581B TNF120572 and VariousInflammatory-Mediating Enzymes in Colon and Liver TissueThe analyzed mRNA expression patterns of pro- and anti-inflammatory genes controlled by NF120581B and TNF120572 differedamong the experimental periods and the tissues investigated(Tables 5 and 6) During the 7-day pretreatment phase 1 themRNA abundance of the colonic pro-inflammatory markersCOX2 and IL-1120573 of the anti-inflammatory cytokine IL-10 ofthe cell adhesion molecules MCP-1 and VCAM-1 and of thetight junction protein Cldn3 showed a high intraindividualvariance In contrast in phase 1 of the trial liver mRNAdata of the above-mentioned genes showed a much betterhomogeneity Nevertheless feeding diets with BE- Cuo-To- and Ro-addition reduced colonic VCAM-1 mRNA by 57to 64 compared to untreated control rats This reductionwas significant for Cuo To and Ro and represented a trend(119875 lt 010) for BE Moreover in phase 1 colonic Cldn3
6 ISRN Gastroenterology
Table 3 Gene bank accession numbers and primer sequences of the genes investigated by real-time RT-PCR
Gene name (abbreviation used) Gene bank accession number Primer sequences (51015840 rarr 31015840)for = forward rev = reverse
Chemokine (C-C motif) ligand 2 (Ccl2) (MCP1) NM 031530 for GTGCGACCCCAATAAGGAArev TGAGGTGGTTGTGGAAAAGA
Claudin 3 (Cldn3) NM 031700 for TATCCTACTGGCAGCCTTCGrev GTTCCCATCTCTCGCTTCTG
Copperzinc superoxide dismutase (SOD1) NM 017050 for CCACTGCAGGACCTCATTTTrev CACCTTTGCCCAAGTCATCT
C-reactive protein (CRP) NM 017096 for GTCTCTATGCCCACGCTGATrev CCGTCAAGCCAAAGCTCTAC
Glutathione S-transferase K1 (GSTK1) NM 181371 for GAGCATGGAGCAACCAGAGATrev AGCTTGCTCTTCACCAGTTCG
Glutathione S-transferase P1 (GSTP1) NM 012577 for GAGGCAAAGCTTTCATTGTGGrev GTTGATGGGACGGTTCAAATG
Glutathione S-transferase T2 (GSTT2) NM 012796 for GAGGAAAAGGTGGAACGGAACrev CGCCCCTCAAACAGATTACAG
Glutathione peroxidase 2 (GPx2) NM 183402 for GTGTGATGTCAATGGGCAGAArev ACGTTTGATGTCAGGCTCGAT
Heme oxygenase 1 (HO1) NM 012580 for AGGCACTGCTGACAGAGGAACrev AGCGGTGTCTGGGATGAACTA
Hypoxanthine phosphoribosyltransferase 1 (Hprt1) NM 012583 for GCAGACTTTGCTTTCCTTGGrev TCCACTTTCGCTGATGACAC
Interleukin 1 beta (IL-1120573) NM 031512 for CTGTGACTCGTGGGATGATGrev GGGATTTTGTCGTTGCTTGT
Interleukin 10 (IL-10) NM 012854 for CTGGAGTGAAGACCAGCAAAGGrev GGAGAAATCGATGACAGCGTCG
Kelch-like ECH-associated protein1 (Keap1) NM 057152 for GTGGCGGATGATTACACCAATrev GAAAAGTGTGGCCATCGTAGC
NAD(P)H dehydrogenase [quinone] 1 (NQO1) NM 017000 for CGCAGAGAGGACATCATTCArev CGCCAGAGATGACTCAACAG
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) NM 031789 for CCAAGGAGCAATTCAACGAAGrev CTCTTGGGAACAAGGAACACG
Nuclear factor kappa B (NF120581B) L26267 for CTTCTCGGAGTCCCTCACTGrev CCAATAGCAGCTGGAAAAGC
Prostaglandin-endoperoxide synthase 2 (Ptgs2) (COX2) NM 017232 for GCTGTACAAGCAGTGGCAAArev CCCCAAAGACAGCATCTGGA
Ribosomal protein L13A (Rpl13a) NM 173340 for CCCTCCACCCTATGACAAGArev CCTTTTCCTTCCGTTTCTCC
Tumor necrosis factor alpha (TNF120572) NM 012675 for GCCAATGGCATGGATCTCAAAGrev AAATCGGCTGACGGTGTGGG
Vascular cell adhesion molecule 1 (VCAM 1) NM 012889 for TGACATCTCCCCTGGATCTCrev CTCCAGTTTCCTTCGCTGAC
120573-actin NM 031144 for ATCGTGCGTGACATTAAAGAGAAGrev GGACAGTGAGGCCAGGATAGAG
expression was significantly higher in Ro-treated rats andtendencially higher in Cuo- and To-treated rats than in theirCon littermates
In phase 1 feeding diets containing BE Cuo and Rosignificantly decreased liver mRNA abundance of the pro-inflammatory cytokine IL-10 by 32 to 44
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
6 ISRN Gastroenterology
Table 3 Gene bank accession numbers and primer sequences of the genes investigated by real-time RT-PCR
Gene name (abbreviation used) Gene bank accession number Primer sequences (51015840 rarr 31015840)for = forward rev = reverse
Chemokine (C-C motif) ligand 2 (Ccl2) (MCP1) NM 031530 for GTGCGACCCCAATAAGGAArev TGAGGTGGTTGTGGAAAAGA
Claudin 3 (Cldn3) NM 031700 for TATCCTACTGGCAGCCTTCGrev GTTCCCATCTCTCGCTTCTG
Copperzinc superoxide dismutase (SOD1) NM 017050 for CCACTGCAGGACCTCATTTTrev CACCTTTGCCCAAGTCATCT
C-reactive protein (CRP) NM 017096 for GTCTCTATGCCCACGCTGATrev CCGTCAAGCCAAAGCTCTAC
Glutathione S-transferase K1 (GSTK1) NM 181371 for GAGCATGGAGCAACCAGAGATrev AGCTTGCTCTTCACCAGTTCG
Glutathione S-transferase P1 (GSTP1) NM 012577 for GAGGCAAAGCTTTCATTGTGGrev GTTGATGGGACGGTTCAAATG
Glutathione S-transferase T2 (GSTT2) NM 012796 for GAGGAAAAGGTGGAACGGAACrev CGCCCCTCAAACAGATTACAG
Glutathione peroxidase 2 (GPx2) NM 183402 for GTGTGATGTCAATGGGCAGAArev ACGTTTGATGTCAGGCTCGAT
Heme oxygenase 1 (HO1) NM 012580 for AGGCACTGCTGACAGAGGAACrev AGCGGTGTCTGGGATGAACTA
Hypoxanthine phosphoribosyltransferase 1 (Hprt1) NM 012583 for GCAGACTTTGCTTTCCTTGGrev TCCACTTTCGCTGATGACAC
Interleukin 1 beta (IL-1120573) NM 031512 for CTGTGACTCGTGGGATGATGrev GGGATTTTGTCGTTGCTTGT
Interleukin 10 (IL-10) NM 012854 for CTGGAGTGAAGACCAGCAAAGGrev GGAGAAATCGATGACAGCGTCG
Kelch-like ECH-associated protein1 (Keap1) NM 057152 for GTGGCGGATGATTACACCAATrev GAAAAGTGTGGCCATCGTAGC
NAD(P)H dehydrogenase [quinone] 1 (NQO1) NM 017000 for CGCAGAGAGGACATCATTCArev CGCCAGAGATGACTCAACAG
Nuclear factor (erythroid-derived 2)-like 2 (Nrf2) NM 031789 for CCAAGGAGCAATTCAACGAAGrev CTCTTGGGAACAAGGAACACG
Nuclear factor kappa B (NF120581B) L26267 for CTTCTCGGAGTCCCTCACTGrev CCAATAGCAGCTGGAAAAGC
Prostaglandin-endoperoxide synthase 2 (Ptgs2) (COX2) NM 017232 for GCTGTACAAGCAGTGGCAAArev CCCCAAAGACAGCATCTGGA
Ribosomal protein L13A (Rpl13a) NM 173340 for CCCTCCACCCTATGACAAGArev CCTTTTCCTTCCGTTTCTCC
Tumor necrosis factor alpha (TNF120572) NM 012675 for GCCAATGGCATGGATCTCAAAGrev AAATCGGCTGACGGTGTGGG
Vascular cell adhesion molecule 1 (VCAM 1) NM 012889 for TGACATCTCCCCTGGATCTCrev CTCCAGTTTCCTTCGCTGAC
120573-actin NM 031144 for ATCGTGCGTGACATTAAAGAGAAGrev GGACAGTGAGGCCAGGATAGAG
expression was significantly higher in Ro-treated rats andtendencially higher in Cuo- and To-treated rats than in theirCon littermates
In phase 1 feeding diets containing BE Cuo and Rosignificantly decreased liver mRNA abundance of the pro-inflammatory cytokine IL-10 by 32 to 44
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
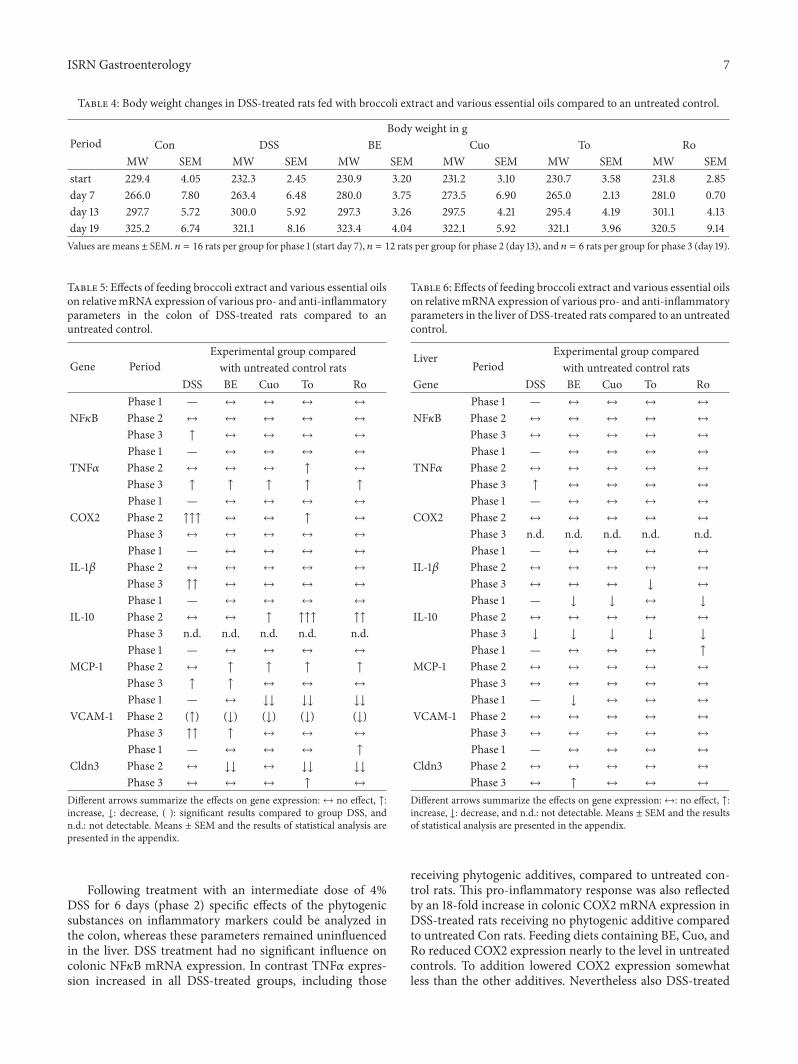
ISRN Gastroenterology 7
Table 4 Body weight changes in DSS-treated rats fed with broccoli extract and various essential oils compared to an untreated control
PeriodBody weight in g
Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
start 2294 405 2323 245 2309 320 2312 310 2307 358 2318 285day 7 2660 780 2634 648 2800 375 2735 690 2650 213 2810 070day 13 2977 572 3000 592 2973 326 2975 421 2954 419 3011 413day 19 3252 674 3211 816 3234 404 3221 592 3211 396 3205 914Values are means plusmn SEM 119899 = 16 rats per group for phase 1 (start day 7) 119899 = 12 rats per group for phase 2 (day 13) and 119899 = 6 rats per group for phase 3 (day 19)
Table 5 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the colon of DSS-treated rats compared to anuntreated control
Gene PeriodExperimental group comparedwith untreated control rats
DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr uarr harr
Phase 3 uarr uarr uarr uarr uarr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 uarruarruarr harr harr uarr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 uarruarr harr harr harr harr
Phase 1 mdash harr harr harr harr
IL-10 Phase 2 harr harr uarr uarruarruarr uarruarr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
MCP-1 Phase 2 harr uarr uarr uarr uarr
Phase 3 uarr uarr harr harr harr
Phase 1 mdash harr darrdarr darrdarr darrdarr
VCAM-1 Phase 2 (uarr) (darr) (darr) (darr) (darr)Phase 3 uarruarr uarr harr harr harr
Phase 1 mdash harr harr harr uarr
Cldn3 Phase 2 harr darrdarr harr darrdarr darrdarr
Phase 3 harr harr harr uarr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease ( ) significant results compared to group DSS andnd not detectable Means plusmn SEM and the results of statistical analysis arepresented in the appendix
Following treatment with an intermediate dose of 4DSS for 6 days (phase 2) specific effects of the phytogenicsubstances on inflammatory markers could be analyzed inthe colon whereas these parameters remained uninfluencedin the liver DSS treatment had no significant influence oncolonic NF120581B mRNA expression In contrast TNF120572 expres-sion increased in all DSS-treated groups including those
Table 6 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreatedcontrol
LiverPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
NF120581B Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
TNF120572 Phase 2 harr harr harr harr harr
Phase 3 uarr harr harr harr harr
Phase 1 mdash harr harr harr harr
COX2 Phase 2 harr harr harr harr harr
Phase 3 nd nd nd nd ndPhase 1 mdash harr harr harr harr
IL-1120573 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr darr harr
Phase 1 mdash darr darr harr darr
IL-10 Phase 2 harr harr harr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr harr harr uarr
MCP-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
VCAM-1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
Cldn3 Phase 2 harr harr harr harr harr
Phase 3 harr uarr harr harr harr
Different arrows summarize the effects on gene expressionharr no effect uarrincrease darr decrease and nd not detectable Means plusmn SEM and the resultsof statistical analysis are presented in the appendix
receiving phytogenic additives compared to untreated con-trol rats This pro-inflammatory response was also reflectedby an 18-fold increase in colonic COX2 mRNA expression inDSS-treated rats receiving no phytogenic additive comparedto untreated Con rats Feeding diets containing BE Cuo andRo reduced COX2 expression nearly to the level in untreatedcontrols To addition lowered COX2 expression somewhatless than the other additives Nevertheless also DSS-treated
8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 ISRN Gastroenterology
Table 7 Effects of feeding broccoli extract and various essential oilson relativemRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreatedcontrol
ColonPeriod
Experimental group comparedwith untreated control rats
Gene DSS BE Cuo To RoPhase 1 mdash harr harr harr harr
Nrf2 Phase 2 darr darr harr darr darr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
Keap1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr harr
HO1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr uarr harr
Phase 1 mdash harr harr harr harr
NQO1 Phase 2 harr harr harr darr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash darr harr harr harr
SOD1 Phase 2 harr harr harr harr harr
Phase 3 harr harr harr harr harr
Phase 1 mdash harr harr harr uarr
GPx2 Phase 2 harr harr uarr harr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash darr harr harr uarr
GSTK1 Phase 2 harr harr harr darr darr
Phase 3 harr darr darr darr darr
Phase 1 mdash harr harr harr uarr
GSTP1 Phase 2 harr harr harr darr harr
Phase 3 darr darr darr darr darr
Phase 1 mdash harr uarr harr uarr
GSTT2 Phase 2 darr darr darr darr darr
Phase 3 harr harr harr harr harr
Different arrows summarize the expression resultsharr no effect uarr increaseand darr decrease For statistically analyzed means plusmn SEM see Table 10
To rats had a colonic COX2 expression which was tenden-cially lower than in their DSS-treated littermates receivingno phytogenic additive Whereas the colonic mRNA con-centration of MCP-1 was not lowered by phytogenic feedadditives during the DSS period all additives tested reducedVCAM-1 expression by 73 to 83 and compared to DSS-treated control rats DSS treatment generally increased theexpression of the anti-inflammatory IL-10 in comparison toCon rats without DSS treatment This effect was relativelysmall in DSS-treated rats receiving no phytogenic additiveand in those receiving the diet containing BE In contrastthe impact of Cuo To and Ro addition on colonic IL-10expression was significant In all DSS-treated rats colonicCldn3 mRNA decreased compared to untreated control ratsThis reduction of Cldn3 expression tended to be lowerin DSS-treated rats without an additive and in the DSS-treated Cuo group and it was significant in their DSS-treatedlittermates receiving diets with the addition of BE To andRo
In the final recovery period (phase 3) NF120581B mRNAstrongly increased in the colon of DSS-treated control ratscompared to their untreated littermates All phytogenicadditives lowered NF120581B response considerably (119875 lt 010)These characteristic changes in NF120581B mRNA were reflectedby similar changes in the expression of the pro-inflammatorycytokine IL-1120573 Nevertheless in all DSS-treated rats includ-ing those receiving phytogenic additives colonic TNF120572mRNA levels remained significantly higher throughout therecovery period In the recovery period also liver TNF120572mRNA level was increased in DSS-treated rats without aphytogenic additive but not in rats fed diets containing anyphytogenic substance compared to untreated controls In therecovery period colonic COX2 mRNA levels of DSS-treatedcontrol rats dropped nearly to the level in untreated controlsand they were further decreased in DSS rats receivingphytogenic additives compared to phase 2 of the experimentIn phase 3 colonic mRNA levels of MCP-1 and VCAM-1in DSS-treated rats without a phytogenic additive furtherincreased compared to phase 2 and they were significantlyhigher than in untreated controls Interestingly a similareffect could be observed in rats fed a BE containing diet Incontrast rats fed diets containing the other additives (CuoTo and Ro) showed MCP-1 and VCAM-1 mRNA levelswhich were comparably low as in untreated controls andsignificantly reduced compared to DSS controls However inthe liver neitherDSS treatment nor combiningDSS treatmentwith feeding phytogenic additives had an influence on MCP-1 and VCAM-1 mRNA abundance in the recovery phaseIn contrast to the acute DSS-treatment period Cldn3 levelswere not lower in the DSS-treated groups than in untreatedcontrols In rats receiving the To diet Cldn3 mRNA was even15ndash20-fold higher than in the other groups
35 mRNA Expression of Nrf2 Keap1 and Various ARE-Regulated Enzymes in the Colon The Nrf2Keap1 systemregulates the expression of antioxidant enzymes and xeno-biotic enzymes In the current experiment some character-istic changes could be observed with regard to Nrf2- andKeap1 expression and on the expression of ARE-regulatedantioxidant and phase II enzymes depending on the exper-imental phase and on the treatment of the rats (Table 7) Inthe pretreatment phase the high impact of Ro on colonicKeap1 expression was directly reflected by an increase inthe expression of several ARE-regulated antioxidant enzymeslike HO1 and GPx2 and on phase II enzymes like GSTK1 P1and T2
In the initial phase all other additives tested had neithera distinct and directed influence on Keap1 mRNA nor onthe expression of the above-mentioned target genes Duringacute DSS treatment the effects on ARE-regulated enzymeswere relatively small Whereas the antioxidant enzymes HO1and NQO1 tended to be higher in DSS-treated control ratsthan in untreated controls all phytogenic additives reducedthis particular DSS effect to a little extent In contrast theexpression of the ARE-regulated phase II enzymes GSTK1P1 and T2 tended to be lowered by DSS treatment In thesecases Nrf2 mRNA abundance showed a similar alteration
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
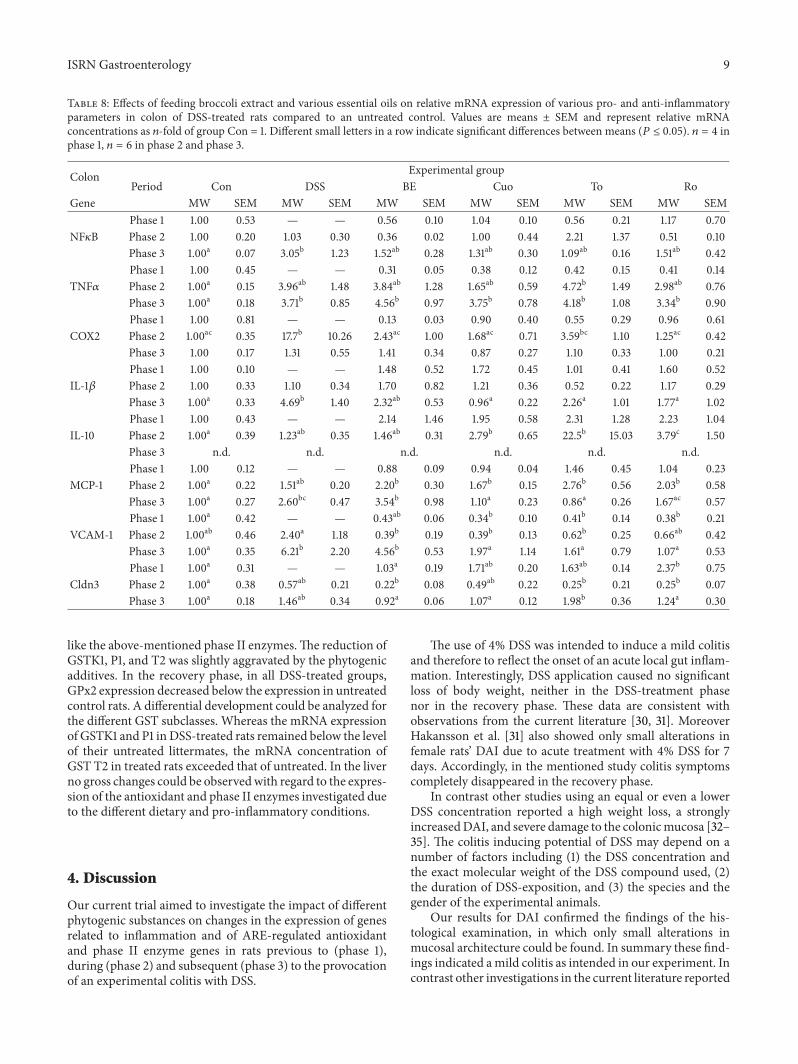
ISRN Gastroenterology 9
Table 8 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in colon of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Colon Experimental groupPeriod Con DSS BE Cuo To Ro
Gene MW SEM MW SEM MW SEM MW SEM MW SEM MW SEMPhase 1 100 053 mdash mdash 056 010 104 010 056 021 117 070
NF120581B Phase 2 100 020 103 030 036 002 100 044 221 137 051 010Phase 3 100a 007 305b 123 152ab 028 131ab 030 109ab 016 151ab 042Phase 1 100 045 mdash mdash 031 005 038 012 042 015 041 014
TNF120572 Phase 2 100a 015 396ab 148 384ab 128 165ab 059 472b 149 298ab 076Phase 3 100a 018 371b 085 456b 097 375b 078 418b 108 334b 090Phase 1 100 081 mdash mdash 013 003 090 040 055 029 096 061
COX2 Phase 2 100ac 035 177b 1026 243ac 100 168ac 071 359bc 110 125ac 042Phase 3 100 017 131 055 141 034 087 027 110 033 100 021Phase 1 100 010 mdash mdash 148 052 172 045 101 041 160 052
IL-1120573 Phase 2 100 033 110 034 170 082 121 036 052 022 117 029Phase 3 100a 033 469b 140 232ab 053 096a 022 226a 101 177a 102Phase 1 100 043 mdash mdash 214 146 195 058 231 128 223 104
IL-10 Phase 2 100a 039 123ab 035 146ab 031 279b 065 225b 1503 379c 150Phase 3 nd nd nd nd nd ndPhase 1 100 012 mdash mdash 088 009 094 004 146 045 104 023
MCP-1 Phase 2 100a 022 151ab 020 220b 030 167b 015 276b 056 203b 058Phase 3 100a 027 260bc 047 354b 098 110a 023 086a 026 167ac 057Phase 1 100a 042 mdash mdash 043ab 006 034b 010 041b 014 038b 021
VCAM-1 Phase 2 100ab 046 240a 118 039b 019 039b 013 062b 025 066ab 042Phase 3 100a 035 621b 220 456b 053 197a 114 161a 079 107a 053Phase 1 100a 031 mdash mdash 103a 019 171ab 020 163ab 014 237b 075
Cldn3 Phase 2 100a 038 057ab 021 022b 008 049ab 022 025b 021 025b 007Phase 3 100a 018 146ab 034 092a 006 107a 012 198b 036 124a 030
like the above-mentioned phase II enzymesThe reduction ofGSTK1 P1 and T2 was slightly aggravated by the phytogenicadditives In the recovery phase in all DSS-treated groupsGPx2 expression decreased below the expression in untreatedcontrol rats A differential development could be analyzed forthe different GST subclasses Whereas the mRNA expressionof GSTK1 and P1 inDSS-treated rats remained below the levelof their untreated littermates the mRNA concentration ofGST T2 in treated rats exceeded that of untreated In the liverno gross changes could be observedwith regard to the expres-sion of the antioxidant and phase II enzymes investigated dueto the different dietary and pro-inflammatory conditions
4 Discussion
Our current trial aimed to investigate the impact of differentphytogenic substances on changes in the expression of genesrelated to inflammation and of ARE-regulated antioxidantand phase II enzyme genes in rats previous to (phase 1)during (phase 2) and subsequent (phase 3) to the provocationof an experimental colitis with DSS
The use of 4 DSS was intended to induce a mild colitisand therefore to reflect the onset of an acute local gut inflam-mation Interestingly DSS application caused no significantloss of body weight neither in the DSS-treatment phasenor in the recovery phase These data are consistent withobservations from the current literature [30 31] MoreoverHakansson et al [31] also showed only small alterations infemale ratsrsquo DAI due to acute treatment with 4 DSS for 7days Accordingly in the mentioned study colitis symptomscompletely disappeared in the recovery phase
In contrast other studies using an equal or even a lowerDSS concentration reported a high weight loss a stronglyincreasedDAI and severe damage to the colonicmucosa [32ndash35] The colitis inducing potential of DSS may depend on anumber of factors including (1) the DSS concentration andthe exact molecular weight of the DSS compound used (2)the duration of DSS-exposition and (3) the species and thegender of the experimental animals
Our results for DAI confirmed the findings of the his-tological examination in which only small alterations inmucosal architecture could be found In summary these find-ings indicated amild colitis as intended in our experiment Incontrast other investigations in the current literature reported
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
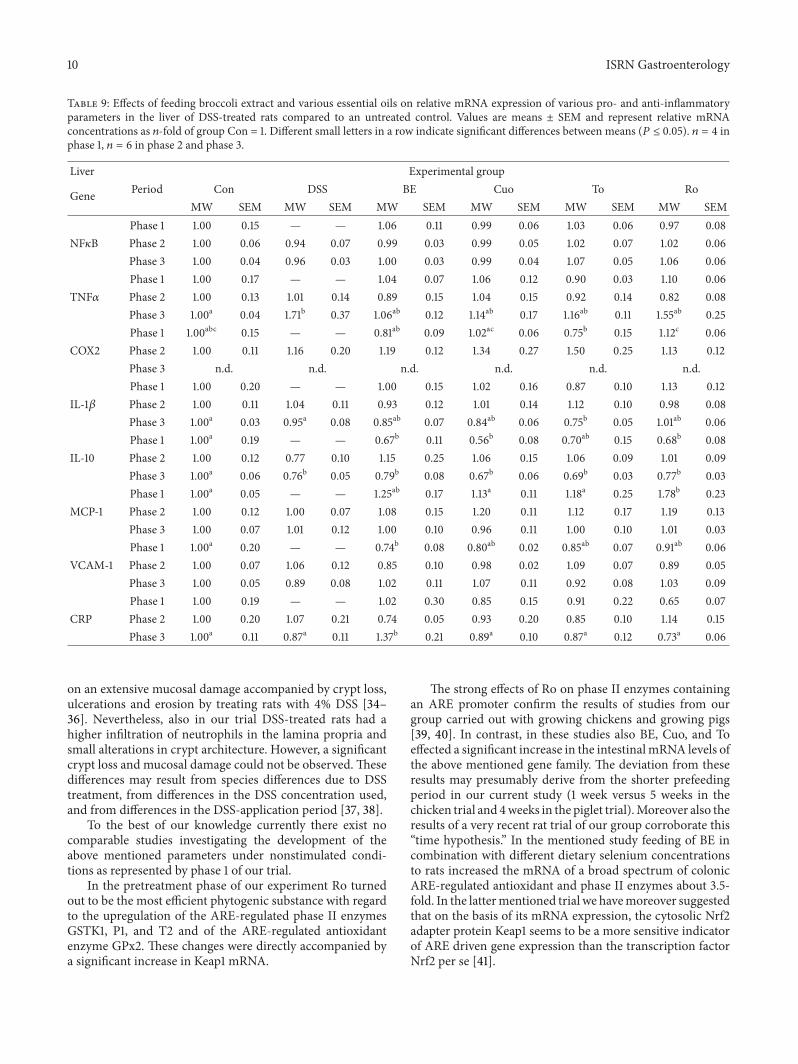
10 ISRN Gastroenterology
Table 9 Effects of feeding broccoli extract and various essential oils on relative mRNA expression of various pro- and anti-inflammatoryparameters in the liver of DSS-treated rats compared to an untreated control Values are means plusmn SEM and represent relative mRNAconcentrations as 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 inphase 1 119899 = 6 in phase 2 and phase 3
Liver Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 015 mdash mdash 106 011 099 006 103 006 097 008NF120581B Phase 2 100 006 094 007 099 003 099 005 102 007 102 006
Phase 3 100 004 096 003 100 003 099 004 107 005 106 006Phase 1 100 017 mdash mdash 104 007 106 012 090 003 110 006
TNF120572 Phase 2 100 013 101 014 089 015 104 015 092 014 082 008Phase 3 100a 004 171b 037 106ab 012 114ab 017 116ab 011 155ab 025Phase 1 100abc 015 mdash mdash 081ab 009 102ac 006 075b 015 112c 006
COX2 Phase 2 100 011 116 020 119 012 134 027 150 025 113 012Phase 3 nd nd nd nd nd ndPhase 1 100 020 mdash mdash 100 015 102 016 087 010 113 012
IL-1120573 Phase 2 100 011 104 011 093 012 101 014 112 010 098 008Phase 3 100a 003 095a 008 085ab 007 084ab 006 075b 005 101ab 006Phase 1 100a 019 mdash mdash 067b 011 056b 008 070ab 015 068b 008
IL-10 Phase 2 100 012 077 010 115 025 106 015 106 009 101 009Phase 3 100a 006 076b 005 079b 008 067b 006 069b 003 077b 003Phase 1 100a 005 mdash mdash 125ab 017 113a 011 118a 025 178b 023
MCP-1 Phase 2 100 012 100 007 108 015 120 011 112 017 119 013Phase 3 100 007 101 012 100 010 096 011 100 010 101 003Phase 1 100a 020 mdash mdash 074b 008 080ab 002 085ab 007 091ab 006
VCAM-1 Phase 2 100 007 106 012 085 010 098 002 109 007 089 005Phase 3 100 005 089 008 102 011 107 011 092 008 103 009Phase 1 100 019 mdash mdash 102 030 085 015 091 022 065 007
CRP Phase 2 100 020 107 021 074 005 093 020 085 010 114 015Phase 3 100a 011 087a 011 137b 021 089a 010 087a 012 073a 006
on an extensive mucosal damage accompanied by crypt lossulcerations and erosion by treating rats with 4 DSS [34ndash36] Nevertheless also in our trial DSS-treated rats had ahigher infiltration of neutrophils in the lamina propria andsmall alterations in crypt architecture However a significantcrypt loss and mucosal damage could not be observed Thesedifferences may result from species differences due to DSStreatment from differences in the DSS concentration usedand from differences in the DSS-application period [37 38]
To the best of our knowledge currently there exist nocomparable studies investigating the development of theabove mentioned parameters under nonstimulated condi-tions as represented by phase 1 of our trial
In the pretreatment phase of our experiment Ro turnedout to be the most efficient phytogenic substance with regardto the upregulation of the ARE-regulated phase II enzymesGSTK1 P1 and T2 and of the ARE-regulated antioxidantenzyme GPx2 These changes were directly accompanied bya significant increase in Keap1 mRNA
The strong effects of Ro on phase II enzymes containingan ARE promoter confirm the results of studies from ourgroup carried out with growing chickens and growing pigs[39 40] In contrast in these studies also BE Cuo and Toeffected a significant increase in the intestinalmRNA levels ofthe above mentioned gene family The deviation from theseresults may presumably derive from the shorter prefeedingperiod in our current study (1 week versus 5 weeks in thechicken trial and 4weeks in the piglet trial)Moreover also theresults of a very recent rat trial of our group corroborate thisldquotime hypothesisrdquo In the mentioned study feeding of BE incombination with different dietary selenium concentrationsto rats increased the mRNA of a broad spectrum of colonicARE-regulated antioxidant and phase II enzymes about 35-fold In the lattermentioned trial we havemoreover suggestedthat on the basis of its mRNA expression the cytosolic Nrf2adapter protein Keap1 seems to be a more sensitive indicatorof ARE driven gene expression than the transcription factorNrf2 per se [41]
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
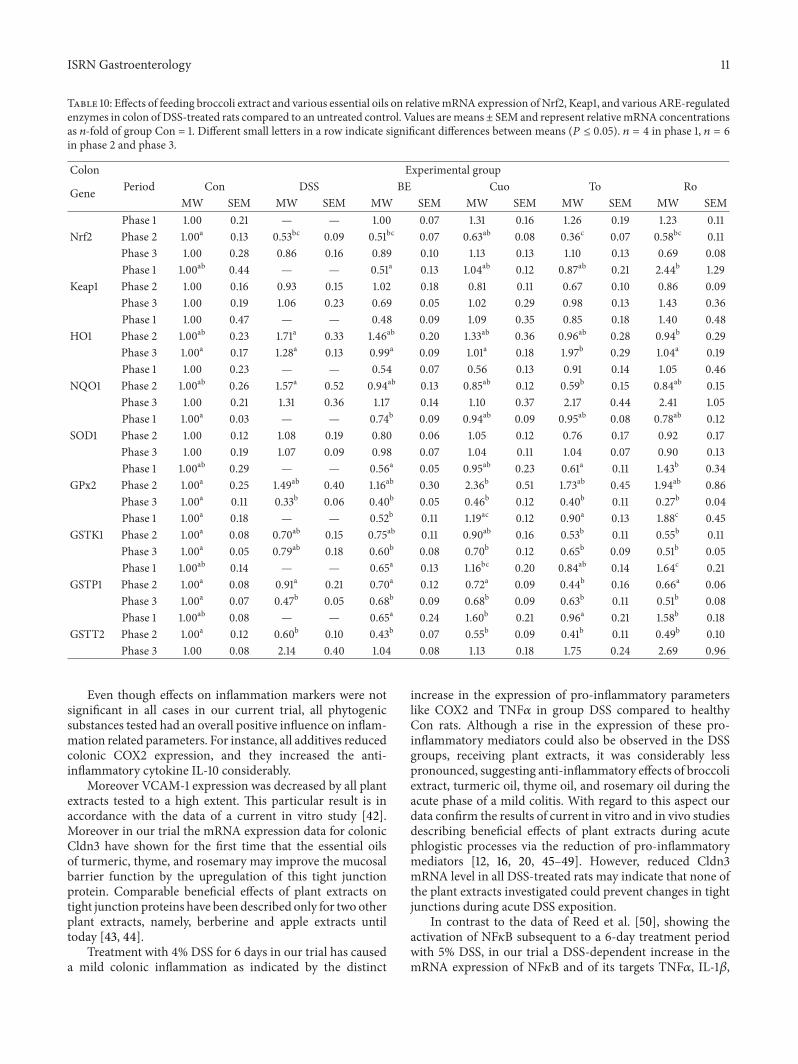
ISRN Gastroenterology 11
Table 10 Effects of feeding broccoli extract and various essential oils on relativemRNA expression of Nrf2 Keap1 and various ARE-regulatedenzymes in colon of DSS-treated rats compared to an untreated control Values aremeansplusmn SEM and represent relativemRNA concentrationsas 119899-fold of group Con = 1 Different small letters in a row indicate significant differences between means (119875 le 005) 119899 = 4 in phase 1 119899 = 6in phase 2 and phase 3
Colon Experimental group
Gene Period Con DSS BE Cuo To RoMW SEM MW SEM MW SEM MW SEM MW SEM MW SEM
Phase 1 100 021 mdash mdash 100 007 131 016 126 019 123 011Nrf2 Phase 2 100a 013 053bc 009 051bc 007 063ab 008 036c 007 058bc 011
Phase 3 100 028 086 016 089 010 113 013 110 013 069 008Phase 1 100ab 044 mdash mdash 051a 013 104ab 012 087ab 021 244b 129
Keap1 Phase 2 100 016 093 015 102 018 081 011 067 010 086 009Phase 3 100 019 106 023 069 005 102 029 098 013 143 036Phase 1 100 047 mdash mdash 048 009 109 035 085 018 140 048
HO1 Phase 2 100ab 023 171a 033 146ab 020 133ab 036 096ab 028 094b 029Phase 3 100a 017 128a 013 099a 009 101a 018 197b 029 104a 019Phase 1 100 023 mdash mdash 054 007 056 013 091 014 105 046
NQO1 Phase 2 100ab 026 157a 052 094ab 013 085ab 012 059b 015 084ab 015Phase 3 100 021 131 036 117 014 110 037 217 044 241 105Phase 1 100a 003 mdash mdash 074b 009 094ab 009 095ab 008 078ab 012
SOD1 Phase 2 100 012 108 019 080 006 105 012 076 017 092 017Phase 3 100 019 107 009 098 007 104 011 104 007 090 013Phase 1 100ab 029 mdash mdash 056a 005 095ab 023 061a 011 143b 034
GPx2 Phase 2 100a 025 149ab 040 116ab 030 236b 051 173ab 045 194ab 086Phase 3 100a 011 033b 006 040b 005 046b 012 040b 011 027b 004Phase 1 100a 018 mdash mdash 052b 011 119ac 012 090a 013 188c 045
GSTK1 Phase 2 100a 008 070ab 015 075ab 011 090ab 016 053b 011 055b 011Phase 3 100a 005 079ab 018 060b 008 070b 012 065b 009 051b 005Phase 1 100ab 014 mdash mdash 065a 013 116bc 020 084ab 014 164c 021
GSTP1 Phase 2 100a 008 091a 021 070a 012 072a 009 044b 016 066a 006Phase 3 100a 007 047b 005 068b 009 068b 009 063b 011 051b 008Phase 1 100ab 008 mdash mdash 065a 024 160b 021 096a 021 158b 018
GSTT2 Phase 2 100a 012 060b 010 043b 007 055b 009 041b 011 049b 010Phase 3 100 008 214 040 104 008 113 018 175 024 269 096
Even though effects on inflammation markers were notsignificant in all cases in our current trial all phytogenicsubstances tested had an overall positive influence on inflam-mation related parameters For instance all additives reducedcolonic COX2 expression and they increased the anti-inflammatory cytokine IL-10 considerably
Moreover VCAM-1 expression was decreased by all plantextracts tested to a high extent This particular result is inaccordance with the data of a current in vitro study [42]Moreover in our trial the mRNA expression data for colonicCldn3 have shown for the first time that the essential oilsof turmeric thyme and rosemary may improve the mucosalbarrier function by the upregulation of this tight junctionprotein Comparable beneficial effects of plant extracts ontight junction proteins have been described only for two otherplant extracts namely berberine and apple extracts untiltoday [43 44]
Treatment with 4 DSS for 6 days in our trial has causeda mild colonic inflammation as indicated by the distinct
increase in the expression of pro-inflammatory parameterslike COX2 and TNF120572 in group DSS compared to healthyCon rats Although a rise in the expression of these pro-inflammatory mediators could also be observed in the DSSgroups receiving plant extracts it was considerably lesspronounced suggesting anti-inflammatory effects of broccoliextract turmeric oil thyme oil and rosemary oil during theacute phase of a mild colitis With regard to this aspect ourdata confirm the results of current in vitro and in vivo studiesdescribing beneficial effects of plant extracts during acutephlogistic processes via the reduction of pro-inflammatorymediators [12 16 20 45ndash49] However reduced Cldn3mRNA level in all DSS-treated rats may indicate that none ofthe plant extracts investigated could prevent changes in tightjunctions during acute DSS exposition
In contrast to the data of Reed et al [50] showing theactivation of NF120581B subsequent to a 6-day treatment periodwith 5 DSS in our trial a DSS-dependent increase in themRNA expression of NF120581B and of its targets TNF120572 IL-1120573
12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 ISRN Gastroenterology
MCP-1 and VCAM-1 did not occur until the end of phase 3(6 days after final DSS treatment) These results may derivefrom the lower DSS concentration (4) used in our trialWith the exception of group To in phase 3 no changes inCldn3 mRNA expression could be observed in the otherexperimental groups compared to Con rats suggesting thatin particular To may improve intestinal barrier function [51]
Data from current literature suggest that the transcriptionfactor Nrf2 responsible for the induction of antioxidant andxenobiotic enzymes is of importance in the control of NF120581Bdependent inflammatory processes (Figure 3) [52] Vice versait is speculated that an increase in NF120581B may inhibit Nrf2signaling Several well-known Nrf2 activators for examplecurcumin resveratrol and sulforaphane are believed tosuppress LPS- and DSS-induced NF120581B activation [53ndash56]whereas the direct relationship between Nrf2 manipulationand NF120581B-inhibition has not been proven until today Ourresults for the acute inflammatory phase 2 cannot confirman association between Nrf2 and NF120581B However a distinctnegative association exists between Nrf2 and TNF120572 whichagain is believed to be the strongest first trigger of NF120581Bactivation (Figures 3ndash5) [57] Thus due to acute DSS exposi-tion Nrf2 expression was distinctly decreased whereas TNF120572expression was strongly elevated In contrast no differencesin the relation between the mentioned transcription factorsexisted under noninflamed conditions (phase 1) Moderateoxidative stress seems to activate Nrf2 and its target genesaccompanied by the simultaneous inhibition of NF120581B signal-ing (Figure 4) This hypothesis seems to be confirmed by thegeneral decrease of VCAM-1 mRNA (NF120581B target) throughapplication of all plant extracts and the increase inGSTK1 andT2 mRNA (Nrf2 targets) by Cuo and Ro application in phase1 of our study (Figure 4)
In phase 2 DSS treatment alone obviously has inducedconsiderable oxidative stress in the colon accompanied by asignificant inhibition of Nrf2 expression finally leading toan increase in NF120581B mRNA in phases 2 and 3 of our trial(Figure 5) In contrast phytogenic additives seem to attenuateboth NF120581B induction and the upregulation of its pro-inflammatory targets This phenomenon may be the resultof the rise in Nrf2-dependent antioxidant and xenobioticenzymes
Our liver data indicate thatDSS-dependent inflammatoryprocesses seem to be restricted to the colon initially but thatthey can also provoke a systemic inflammatory response asindicated by the increase in TNF120572 expression in DSS-treatedrats in phase 3
5 Conclusions
Our experimental design has allowed for the simultaneousexamination of (1) preventive effects of various plant extractson intestinal health (phase 1) of (2) their anti-inflammatoryand antioxidant potential during acute DSS-induced colitis(phase 2) and of (3) their influence on parameters related toinflammation and the antioxidant systemduring the recoveryprocess (phase 3)
We conclude the following
(i) The treatment of rats under noninflamed conditionswith broccoli extract and the essential oils of turmericthyme and rosemary promotes intestinal health byreducing the pro-inflammatory adhesion moleculeVCAM-1 and by increasing the tight junction markerCldn3 leading to an improved gut barrier
(ii) The phytogenic additives investigated have anti-inflammatory properties as indicated by the reductionof the DSS-induced increase in pro-inflammatorymediators like NF120581B VCAM-1 MCP-1 and COX2 toa greater or lesser extent
Further research is needed to evaluate the role of phy-togenic additives in modulating NF120581B-Nrf2 interactions inmore detail Moreover detailed investigations with regardto the time-dependent regulation of inflammatory andantioxidant responses during an acute gut inflammationare urgently needed Nevertheless plant extracts and inparticular essential oils may represent promising substancesin the complementary therapy of IBD with a particular focusin the prevention
Appendix
Thecomplete results (meansplusmn SEM) of themRNAexpressionanalyses including statistical evaluation are shown in Tables8 9 and 10
Conflict of Interests
All authors declare that they have no conflict of interests
Acknowledgments
This study was partially funded by Delacon BiotechnikGesmbH Steyregg (Austria) to investigate fundamentalmechanisms of phytogenic feed additives under inflamma-tory conditions The authors thank Devakumari Hiller (Jar-rowDeutschland GmbH Berlin Germany) for arranging thecontact with Mr Jarrow L Rogovin from Jarrow FormulasLos Angeles USA who provided us with the broccoli seedextract Thank is also addressed to our master students NinaWodrich Lisa Schering and Anne Lehmann for their helpwith analyses
References
[1] G Rogler and T Andus ldquoCytokines in inflammatory boweldiseaserdquo World Journal of Surgery vol 22 no 4 pp 382ndash3891998
[2] I Arijs G deHertogh KMachiels et al ldquoMucosal gene expres-sion of cell adhesion molecules chemokines and chemokinereceptors in patients with inflammatory bowel disease beforeand after infliximab treatmentrdquo The American Journal of Gas-troenterology vol 106 no 4 pp 748ndash776 2011
[3] A S Baldwin ldquoThe NF-120581B and I120581B proteins new discoveriesand insightsrdquo Annual Review of Immunology vol 14 pp 649ndash681 1996
ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

ISRN Gastroenterology 13
[4] S Ghosh M J May and E B Kopp ldquoNF-120581B and rel proteinsevolutionarily conserved mediators of immune responsesrdquoAnnual Review of Immunology vol 16 pp 225ndash260 1998
[5] A S Baldwin Jr ldquoThe transcription factor NF-120581B and humandiseaserdquo The Journal of Clinical Investigation vol 107 no 1 pp3ndash6 2001
[6] M Amasheh I Grotjohann S Amasheh et al ldquoRegulation ofmucosal structure and barrier function in rat colon exposedto tumor necrosis factor alpha and interferon gamma invitro a novel model for studying the pathomechanisms ofinflammatory bowel disease cytokinesrdquo Scandinavian Journal ofGastroenterology vol 44 no 10 pp 1226ndash1235 2009
[7] M F Neurath and G Schurmann ldquoImmunopathogenesis ofinflammatory bowel diseasesrdquo Chirurg vol 71 no 1 pp 30ndash402000
[8] Y Xu B Gong Y Yang Y C Awasthi MWoods and P J BoorldquoGlutathione-S-transferase protects against oxidative injury ofendothelial cell tight junctionsrdquo Endothelium vol 14 no 6 pp333ndash343 2007
[9] P Sheth N Delos Santos A Seth N F LaRusso and R KRao ldquoLipopolysaccharide disrupts tight junctions in cholan-giocyte monolayers by a c-Src- TLR4- and LBP-dependentmechanismrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 293 no 1 pp G308ndashG318 2007
[10] T Kawauchiyaa R Takumia Y Kudoa et al ldquoCorrelationbetween the destruction of tight junction by patulin treatmentand increase of phosphorylation of ZO-1 in Caco-2 humancolon cancer cellsrdquo Toxicology Letters vol 205 no 2 pp 196ndash202 2011
[11] L Petecchia F Sabatini C Usai E Caci L Varesio andG A Rossi ldquoCytokines induce tight junction disassembly inairway cells via an EGFR-dependent MAPKERK12-pathwayrdquoLaboratory Investigation vol 92 pp 1140ndash1148 2012
[12] A Ocana-Fuentes E Arranz-Gutierrez F J Senorans and GReglero ldquoSupercritical fluid extraction of oregano (Origanumvulgare) essentials oils anti-inflammatory properties based oncytokine response on THP-1 macrophagesrdquo Food and ChemicalToxicology vol 48 no 6 pp 1568ndash1575 2010
[13] A Kroismayr J Sehm MW Pfaffl K Schedle C Plitzner andW M Windisch ldquoEffects of avilamycin and essential oils onmRNA expression of apoptotic and inflammatory markers andgutmorphology of pigletsrdquoCzech Journal of Animal Science vol53 no 9 pp 377ndash387 2008
[14] M Viuda-Martos Y Ruiz-Navajas J Fernandez-Lopez and JA Perez-Alvarez ldquoEffect of added citrus fibre and spice essentialoils on quality characteristics and shelf-life of mortadellardquoMeatScience vol 85 no 3 pp 568ndash576 2010
[15] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no3 pp 78ndash98 2012
[16] A Bukovska S Cikos S Juhas G Ilrsquokova P Rehak andJ Koppel ldquoEffects of a combination of thyme and oreganoessential oils on TNBS-induced colitis in micerdquo Mediators ofInflammation vol 2007 Article ID 23296 9 pages 2007
[17] K C Lian J J Chuang C W Hsieh et al ldquoDual mechanismsof NF-120581B inhibition in carnosol-treated endothelial cellsrdquoToxicology and Applied Pharmacology vol 245 no 1 pp 21ndash352010
[18] X F Tian J H Yao X S Zhang et al ldquoProtectiveeffect of carnosol on lung injury induced by intestinal
ischemiareperfusionrdquo Surgery Today vol 40 no 9 pp 858ndash865 2010
[19] E S Mengoni G Vichera L A Rigano et al ldquoSuppression ofCOX-2 IL-1120573 and TNF-120572 expression and leukocyte infiltrationin inflamed skin by bioactive compounds from Rosmarinusofficinalis Lrdquo Fitoterapia vol 82 no 3 pp 414ndash421 2011
[20] A E Wagner C ldquoAnti-inflammatory potential of allyl-isothiocyanate-role of Nrf2 NF120581B and microRNA-155rdquo Journalof Cellular and Molecular Medicine vol 16 no 4 pp 836ndash8432011
[21] Anonymous ldquoldquoEvonik Industries Product InformationrdquoTEGO Turmerone the distilled fraction of tumeric oilextracted from 12 the roots of Curcuma longa by supercriticalcarbon dioxiderdquo 2012 httpwwwcenterchemcomPDFsTEGO20Turmerone20Tech20Lit200208pdf
[22] Society for Laboratory Animal Science Ausschuss fur Tierg-erechte Labortierhaltung Tiergerechte Haltung von Laborratte2004
[23] L A Dieleman M J H J Palmen H Akol et al ldquoChronicexperimental colitis induced by dextran sulphate sodium (DSS)is characterized by Th1 and Th2 cytokinesrdquo Clinical and Exper-imental Immunology vol 114 no 3 pp 385ndash391 1998
[24] I Okayasu M Yamada T Mikami T Yoshida J Kannoand T Ohkusa ldquoDysplasia and carcinoma development in arepeated dextran sulfate sodium-induced colitismodelrdquo JournalofGastroenterology andHepatology vol 17 no 10 pp 1078ndash10832002
[25] D Camuesco M Comalada A Concha et al ldquoIntestinal anti-inflammatory activity of combined quercitrin and dietary oliveoil supplemented with fish oil rich in EPA and DHA (n-3)polyunsaturated fatty acids in rats with DSS-induced colitisrdquoClinical Nutrition vol 25 no 3 pp 466ndash476 2006
[26] P Chomczynski andN Sacchi ldquoThe single-stepmethod of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction twenty-something years onrdquoNature Protocols vol 1no 2 pp 581ndash585 2006
[27] T A Kerr M A Ciorba H Matsumoto et al ldquoDextran sodiumsulfate inhibition of real-time polymerase chain reaction ampli-fication a poly-A purification solutionrdquo Inflammatory BowelDiseases vol 18 no 2 pp 344ndash348 2012
[28] N M Blum K Mueller D Lippmann J Pallauf T Linnand A S Mueller ldquoGlucoraphanin does not reduce plasmahomocysteine in rats with sufficient Se supply via the inductionof liver ARE-regulated glutathione biosynthesis enzymesrdquo Foodamp Function vol 2 pp 654ndash664 2011
[29] K J Livak and T D Schmittgen ldquoAnalysis of relative geneexpression data using real-time quantitative PCR and the 2-ΔΔCT methodrdquoMethods vol 25 no 4 pp 402ndash408 2001
[30] M Vicario C Amat M Rivero M Moreto and C PelegrıldquoDietary glutamine affects mucosal functions in rats with mildDSS-induced colitisrdquo Journal of Nutrition vol 137 no 8 pp1931ndash1937 2007
[31] A Hakansson C Branning G Molin et al ldquoColorectal onco-genesis and inflammation in a rat model based on chronicinflammation due to cycling DSS treatmentsrdquo GastroenterologyResearch and Practice vol 2011 Article ID 924045 15 pages2011
[32] A Venkatraman B S Ramakrishna A B Pulimood S Patraand S Murthy ldquoIncreased permeability in dextran sulphate col-itis in rats time course of development and effect of butyraterdquoScandinavian Journal of Gastroenterology vol 35 no 10 pp1053ndash1059 2000
14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

14 ISRN Gastroenterology
[33] R C Sprong A J Schonewille and R van der Meer ldquoDietarycheese whey protein protects rats against mild dextran sulfatesodium-induced colitis role of mucin and microbiotardquo Journalof Dairy Science vol 93 no 4 pp 1364ndash1371 2010
[34] L Marquez B G Perez-Nievas I Garate et al ldquoAnti-inflammatory effects of Mangifera indica L extract in a modelof colitisrdquoWorld Journal of Gastroenterology vol 16 no 39 pp4922ndash4931 2010
[35] A Tyagi U Kumar S Reddy et al ldquoAttenuation of colonicinflammation by partial replacement of dietary linoleic acidwith 120572-linolenic acid in a rat model of inflammatory boweldiseaserdquo British Journal of Nutrition vol 108 no 9 pp 1612ndash1622 2012
[36] E Gaudio G Taddei A Vetuschi et al ldquoDextran sulfate sodium(DSS) colitis in rats clinical structural and ultrastructuralaspectsrdquoDigestive Diseases and Sciences vol 44 no 7 pp 1458ndash1475 1999
[37] L Pouyet C Roisin-Bouffay A Clement et al ldquoEpithelialvanin-1 controls inflammation-driven carcinogenesis in thecolitis-associated colon cancer modelrdquo Inflammatory BowelDiseases vol 16 no 1 pp 96ndash104 2010
[38] D K Zhang J J Yu Y M Li et al ldquoA Picrorhiza kurroa deriva-tive picroliv attenuates the development of dextran-sulfate-sodium-induced colitis in micerdquo Mediators of Inflammationvol 2012 Article ID 751629 9 pages 2012
[39] K Mueller N M Blum H Kluge and A S Mueller ldquoInfluenceof broccoli extract and various essential oils on performanceand expression of xenobiotic- and antioxidant enzymes inbroiler chickensrdquo British Journal of Nutrition vol 108 no 4 pp588ndash602 2012
[40] K Mueller N M Blum H Kluge et al ldquoEffects of broccoliextract and various essential oils on intestinal and faecalmicroflora and on xenobiotic enzymes and the antioxidantsystem of pigletsrdquo Open Journal of Animal Sciences vol 2 no2 pp 78ndash98 2012
[41] N M Blum K Mueller D Lippmann et al ldquoKeap 1 inmind feeding of selenium alone or in combination withglucoraphanin differentially affects intestinal and hepatic AREregulated antioxidant and phase II enzymes in growing ratsrdquoBiological Trace Element Research vol 151 no 3 pp 384ndash3992013
[42] D G Binion J Heidemann M S Li V M Nelson M FOtterson and P Rafiee ldquoVascular cell adhesion molecule-1expression in human intestinal microvascular endothelial cellsis regulated by PI 3-kinaseAktMAPKNF- 120581B Inhibitory roleof curcuminrdquo American Journal of PhysiologymdashGastrointestinaland Liver Physiology vol 297 no 2 pp G259ndashG268 2009
[43] L Gu N Li Q Li et al ldquoThe effect of berberine in vitroon tight junctions in human Caco-2 intestinal epithelial cellsrdquoFitoterapia vol 80 no 4 pp 241ndash248 2009
[44] R A Vreeburg E E van Wezel F Ocana-Calahorro and J JMes ldquoApple extract induces increased epithelial resistance andclaudin 4 expression in Caco-2 cellsrdquo Journal of the Science ofFood and Agriculture vol 92 no 2 pp 439ndash444 2012
[45] S Juhas S Cikos S Czikkova et al ldquoEffects of borneoland thymoquinone on TNBS-induced colitis in micerdquo FoliaBiologica (Praha) vol 54 no 1 pp 1ndash7 2008
[46] T YWu TO Khor C L L Saw et al ldquoAnti-inflammatoryanti-oxidative stress activities and differential regulation of Nrf2-mediated genes by non-polar fractions of tea Chrysanthemumzawadskii and licorice Glycyrrhiza uralensisrdquo AAPS Journalvol 13 no 1 pp 1ndash13 2011
[47] X Liu and J Wang ldquoAnti-inflammatory effects of iridoidglycosides fraction of Folium syringae leaves on TNBS-inducedcolitis in ratsrdquo Journal of Ethnopharmacology vol 133 no 2 pp780ndash787 2011
[48] D K Gessner R Ringseis M Siebers et al ldquoInhibition ofthe pro-inflammatory NF-120581B pathway by a grape seed andgrape marc meal extract in intestinal epithelial cellsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 6 pp1074ndash1083 2011
[49] J Y Kim H J Park S H Um et al ldquoSulforaphane suppressesvascular adhesion molecule-1 expression in TNF-120572-stimulatedmouse vascular smoothmuscle cells involvement of theMAPKNF-120581B and AP-1 signaling pathwaysrdquo Vascular Pharmacologyvol 56 no 3-4 pp 131ndash141 2012
[50] K L Reed A B Fruin A C Gower et al ldquoNF-120581B activationprecedes increases in mRNA encoding neurokinin-1 receptorproinflammatory cytokines and adhesionmolecules in dextransulfate sodium-induced colitis in ratsrdquo Digestive Diseases andSciences vol 50 no 12 pp 2366ndash2378 2005
[51] A GMarkov A VeshnyakovaM FrommM Amasheh and SAmasheh ldquoSegmental expression of claudin proteins correlateswith tight junction barrier properties in rat intestinerdquo Journal ofComparative Physiology B vol 180 no 4 pp 591ndash598 2010
[52] T O Khor M T Huang K H Kwon J Y Chan B SReddy and A N Kong ldquoNrf2-deficient mice have an increasedsusceptibility to dextran sulfate sodium-induced colitisrdquoCancerResearch vol 66 no 24 pp 11580ndash11584 2006
[53] M Larrosa M J Yanez-Gascon M V Selma et al ldquoEffectof a low dose of dietary resveratrol on colon microbiotainflammation and tissue damage in a DSS-induced colitis ratmodelrdquo Journal of Agricultural and Food Chemistry vol 57 no6 pp 2211ndash2220 2009
[54] L O Brandenburg M Kipp R Lucius T Pufe and C JWruck ldquoSulforaphane suppresses LPS-induced inflammation inprimary ratmicrogliardquo Inflammation Research vol 59 no 6 pp443ndash450 2010
[55] M Nishida S Nishiumi Y Mizushina et al ldquoMonoacetylcur-cumin strongly regulates inflammatory responses through inhi-bition of NF-120581B activationrdquo International Journal of MolecularMedicine vol 25 no 5 pp 761ndash767 2010
[56] Q Jia I Ivanov Z Z Zlatev et al ldquoDietary fish oil and curcumincombine to modulate colonic cytokinetics and gene expressionin dextran sodium sulphate-treated micerdquo British Journal ofNutrition vol 106 no 4 pp 519ndash529 2011
[57] Y H Zhou J P Yu Y F Liu et al ldquoEffects of Ginkgo bilobaextract on inflammatory mediators (SOD MDA TNF-120572 NF-120581Bp65 IL-6) in TNBS-induced colitis in ratsrdquo Mediators ofInflammation vol 2006 Article ID 92642 9 pages 2006
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Copyright © 2022 FDOKUMEN




















