
Generic Priors Yield Competition Between Independently-Occurring Preventive Causes

[CANCER RESEARCH 47, 760-766, February 1, 1987)
Inhibition of Epidermal Xenobiotic Metabolism in SENCAR Mice by NaturallyOccurring Plant Phenols1
Mukul Das,2 Hasan Mukhtar,3 Daniel P. Bik, and David R. Bickers
Departments of Dermatology, University Hospitals of Cleveland, Case Western Reserve University, and Veterans Administration Medical Center, Cleveland, Ohio 44106
ABSTRACT
Naturally occurring plant phenols such as tannic acid, quercetin,myricetin, and anthraflavic acid have been shown to inhibit the mutagen-icity of several bay-region diol-epoxides of polycyclic aromatic hydrocarbons including benzo(a)pyrene (BP). The present study was designed todetermine whether these plant phenols can alter epidermal cytochromeP-450-dependent monooxygenases in SENCAR mice. In vitro additionof these plant phenols to epidermal microsomal preparations inhibitedaryl hydrocarbon hydroxylase (AHH) activity in a concentration-dependent manner. The 50% inhibitory concentrations for tannic acid, myricetin,quercetin, and anthraflavic acid ranged from 4.4 X 10~5Mto 12.4 x 10 '
M in microsomes prepared from control and 3-methylcholanthrene-pre-treated animals. Of the plant phenols studied tannic acid was found to bethe most potent inhibitor of epidermal AHH activity. Tannic acid, quercetin, myricetin, and anthraflavic acid exhibited a mixed type of inhibitoryeffect with A, values of 81, 63, 135, and 165 /JM, respectively. In vitroaddition of these plant phenols (240 MM) to the incubation mixtureprepared from control and 3-methylcholanthrene-treated animals resultedin varying degrees of inhibition of epidermal microsomal AHH (57-92%), ethoxycoumarin 0-deethylase (19-58%), and ethoxyresorufin O-deethylase (33-85%) activities. High pressure liquid Chromatographieanalysis of the organic solvent-soluble metabolites of BP produced byepidermal microsomes indicated a substantial decrease in the formationof BP-diols (23-67%) and BP-phenols (29-57%) by each of the plantphenols. The formation of BP-7,8-diol was substantially inhibited (29-52%) by each of the plant phenols. Further in vivo studies showed thata single topical application of tannic acid, quercetin, and myricetin greatlydiminished epidermal AHH (53-65%), ethoxycoumarin 0-deethylase(30-68%), and ethoxyresorufin 0-deethylase (66-97%) activitieswhereas anthraflavic acid was ineffective in this regard even when repeatedly applied. Our results indicate that plant phenols have substantialthough variable inhibitory effects on epidermal monooxygenase activitiesand BP metabolism suggesting that these compounds may be capable ofinhibiting the carcinogenic effects of polycyclic aromatic hydrocarbons inthe skin.
INTRODUCTIONThe hemeprotein cytochrome P-450 is a heterogeneous mi
crosomal enzyme system found in liver and many extrahepatictissues including skin (1,2). This monooxygenase system functions in the oxidative biotransformation of many drugs andforeign chemicals converting them to polar metabolites, therebyfacilitating both their pharmacological inactivation and theirelimination from the body (1-3). However, this enzyme systemis also capable of transforming certain chemicals into highlyreactive toxic metabolites that may produce a variety of pathological effects. For example, carcinogenic PAHs4 are them-
Received 3/31/86; revised 9/11/86; accepted 10/21/86.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported in part by NIH Grants ES-1900, CA-38028, and AM-34368; by
American Institute for Cancer Research Grant 86A61; and by research fundsfrom the Veterans Administration.
1 Recipient of Schering-Plough Foundation fellowship award from the Der
matology Foundation.' To whom requests for reprints should be addressed, at Veterans Administra
tion Medical Center, 10701 East Boulevard, Cleveland, OH 44106.4The abbreviations used are: PAH, polycyclic aromatic hydrocarbon; AHH,
aryl hydrocarbon hydroxylase; ECD, 7-ethoxycoumarin O-deethylase; ERD, 7-ethoxyresorufin O-deethylase; BP, benzo(a)pyrene; BP-7,8-diol, (±)-/rani-7,8-dihydro-7,8-dihydroxybenzo(a)pyrene; MCA, 3-methylcholanthrene; HPLC,high pressure liquid chromatography; I.,,. 50% inhibitory concentration.
selves relatively inert compounds and essentially act as precar-cinogens that must first undergo metabolic activation to biologically active metabolites that are the ultimate carcinogens (1,4). The first step in the metabolic activation of PAHs is catalyzed by the cytochrome P-450-dependent enzyme commonlyknown as AHH (1, 4). Knowledge of the importance of thisenzyme in the metabolic activation of PAHs has led to thesuggestion that inhibitors of AHH activity could diminish thecarcinogenicity and mutagenicity of PAHs in target tissues suchas the skin. For example a synthetic flavonoid, 7,8-benzofla-vone, is an inhibitor of skin AHH activity, diminishes bindingof 7,12-dimethylbenz(a)anthracene metabolites to cellular mac-romolecules, and reduces skin tumor initiation by 7,12-dimeth-ylbenz(a)anthracene (5, 6). In previous studies our laboratoryhas shown that the antifungal imidazole, clotrimazole, inhibitsepidermal AHH activity, decreases binding of BP metabolitesto epidermal DNA, and protects against skin tumor inductionby topically applied MCA (7).
Recent attention has been focused on the identification ofnaturally occurring plant phenols as possible cancer chemopre-ventive agents. One promising approach to decreasing the riskof developing chemically induced tumors might be the modulation of the activity of enzymes that participate in metabolicactivation of carcinogens. Several recent studies have shownthat a naturally occurring plant polyphenol, ellagic acid, inhibitsthe cytochrome P-450-dependent metabolism, the DNA binding, and the tumorigenicity of PAHs in skin and lung (8-12).In the present study we provide evidence for the in vitro and invivo inhibition of cutaneous monooxygenases and BP metabolism in SENCAR mice by a series of plant phenols includingtannic acid, flavonoids, and anthraquinones (for structures seeFig. 1).
MATERIALS AND METHODS
Chemicals. Gold label BP, 7-ethoxycoumarin, resorufin, tannic acid,digallic acid, myricetin, and anthraflavic acid were obtained from Aid-rich Chemical Co (Milwaukee, WI). 7-Ethoxyresonifin was purchasedfrom Pierce Chemicals. Quercetin, MCA, NADPH, NADH, and bovineserum albumin were obtained from Sigma Chemical Co. (St. Louis,MO). [7,10-I4C]BP (specific activity, 58.5 mCi/mmol) was purchased
from Amersham Searle (Chicago, IL). Prior to use, radiolabeled BPwas purified on a silica gel (l'ari¡siilO-jtm; Waters Associates) columnwith hexane as the eluting solvent and subsequently by reverse-phaseHPLC using a Dupont Zorbax ODS column (6.2 mm x 25 cm) elutedwith methanokwater (19:1 v/v). The purity of BP was greater than 99%as judged by HPLC. BP reference standards were provided by theCancer Research Program, Division of Cancer Cause and Prevention,National Cancer Institute (Bethesda, MD). All other chemicals wereobtained in the purest form commercially available.
Treatment of Animals. Six-week-old female SENCAR mice, obtainedfrom the NCI-Frederick Cancer Research Facility, Bethesda, MD, wereused. The mice were shaved with electric clippers and Nair depilatorywas applied 1 day prior to the beginning of the experiment. Forevaluation of the in vitro effects of tannic acid, quercetin, myricetin,and anthraflavic acid, one group of animals received a single topicalapplication of MCA (50 mg/kg) in 0.1 ml of acetone while controlanimals received an identical volume of the vehicle. For studies of the
760

INHIBITION OF EPIDERMAL MONOOXYGENASES BY PLANT PHENOLS
in vivo effects of plant phenols on epidermal monooxygenases, animalswere treated topically with different doses of the compound (50-400
¿tmol/kgbody weight) in 0.1 ml of acetone and/or dimethyl sulfoxide.Controls were treated with identical volumes of the vehicle.
Preparation of Microsomal Fraction. The mice were killed 24 h aftertreatment by decapitation with surgical scissors. The skin was excisedand immediately placed in an ice-cold solution of KC1 (0.15 M). Eachskin sample was placed epidermal side down on a covered glass Petridish containing crushed ice. The skin was scraped with a sharp scalpelblade (Bard-Parker No. 20) to remove dermis, subcutaneous fat, andmuscle and epidermal microsomes were prepared as described earlier(13). The microsomal pellets were overlaid with buffer A [100 mMpotassium phosphate, pH 7.4, containing 10 mM dithiothreitol, 10 mMEDTA, and 20% (v/v) glycerol] and frozen at -170'C under nitrogen.
For the determination of microsomal enzyme activities the frozenpellets were slowly thawed (within 3-5 days of tissue preparation) inan ice bucket, suspended in buffer A, and used as the enzyme source.Enzyme activities were stable under these storage conditions for at least3 weeks.
Enzyme Assays. AHH activity was determined by a modification ofthe method of Nebert and Gelboin (14), the details of which have beendescribed earlier (13). The quantitation of phenolic BP metabolites wasbased on comparison of fluorescence with a 3-hydroxy-BP standard.ECD activity was determined according to a slight modification of theprocedure of Greenlee and Poland (15), the adaptations of which havebeen described earlier (13). ERD activity was determined according tothe procedure of Pohl and Fouts (16). The reaction was initiated by theaddition of 1.5 pM ethoxyresorufin in 5 pi of dimethyl sulfoxide andwas incubated aerobically at 37*C for 30 min in a Dubnoff metabolic
shaker. The reaction was terminated by the addition of 2.0 ml ofmethanol. Fluorescence of the deethylated metabolite was measured atexcitation and emission wavelengths of 550 and 585 nm, respectively.The quantitation of the deethylated metabolite was based on comparison of its fluorescence with resorufin as standard. Protein was determined, after precipitation with trichloroacetic acid, by the procedure ofLowry et al. (17) using bovine serum albumin as reference standard.
in Vitro System of BP Metabolism. The incubation mixture in a finalvolume of 1.0 ml contained 1.0 mg of microsomal protein, 0.10 mmolof phosphate buffer (pH 7.4), 3 /<mol of MgCl2, and 1.3 /¿molofNADPH. The reaction was initiated by the addition of 80 nmol of[I4C]BP in 20 /il of acetone. The samples were incubated for 30 min at37°Cin a Dubnoff metabolic shaker in the presence or absence of 60
ti\\ plant phenol. The reaction was terminated by the addition of 1 mlof cold acetone followed by 2 ml of ethyl acetate. The mixture wasvortexed for 1 min to extract any unreacted BP as well as the metabolitesinto the organic phase. The organic and aqueous layers were separatedby centrifugation at 1500 rpm for 10 min. The radioactivity in the
HOOC OH
DIGALLIC ACID
CH20-R
/Ho 0-RV \J\0-R/ov-rR 0-R
TANNIC ACID
( R - Digol I ic acid )
OH 0ANTHRAFLAVIC ACID
Fig. 1. Structure of the plant phenols.
aqueous phase ranged from 0.3 to 0.4% of the total radioactivity andwas proportional to the extent of BP metabolized. The organic phasewas dried under a stream of nitrogen and dissolved in 100 ¿ilof methanolfor HPLC analysis as described earlier (8). All operations were performed under subdued yellow light.
HPLC Analysis of Formation of Metabolites. A Waters Associatesmodel 204 liquid Chromatograph, fitted with a Waters Associate ( ',K-
fiBondapak column (4.6 mm x 25 cm) was used for the analysis ofradiolabeled metabolite mixtures of BP. Identification of the metabolites was based on reference standards. The column was eluted atambient temperature with a 20min linear gradient of methanol:water(65:35, v/v) to methanol at a solvent flow rate of 0.6 ml/min. Theeluates were monitored at 254 nm, fractions of 0.2 ml were collecteddropwise, and the radioactivity of each fraction was determined on aPackard Tri-Carb 460 CD liquid scintillation spectrometer.
RESULTS
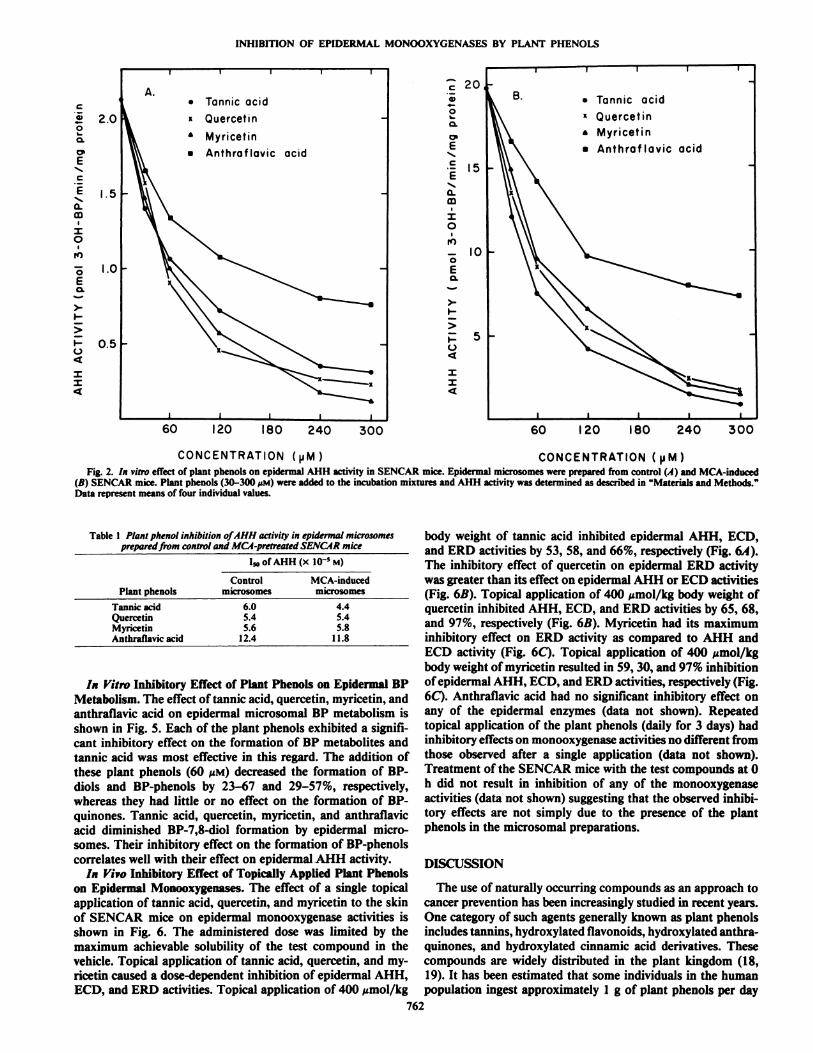
In Vitro Inhibitory Effects of Plant Phenols on EpidermalMonooxygenases. The effect of tannic acid, quercetin, myrice-tin, and anthraflavic acid on epidermal microsomal AHH activity in control and MCA-treated SENCAR mice is shown inFig. 2. Each of the four plant phenols exhibited a concentration-dependent inhibitory effect on AHH activity in microsomesprepared from control as well as MCA-induced animals (Fig.2). The extent of the inhibitory effect on epidermal AHH bythese plant phenols was in the following order: tannic acid >quercetin > myricetin > anthraflavic acid (Fig. 2). It is important to point out that digallic acid, which occurs on five differentpositions of the glucose moiety in the structure of tannic acid,had no inhibitory effect on epidermal AHH activity (data notshown). Table 1 shows the I-,,,for these plant phenols againstAHH activity in epidermal microsomal prepared from controland MCA-pretreated animals. Quercetin was the most effectiveinhibitor with an I5o of 5.4 xlO~5 M for AHH activity in
epidermal microsomes prepared from control animals followedin decreasing order by myricetin (5.6 x 10~5M), tannic acid (6.0x Ifr5 M), and anthraflavic acid (12.4 x 10~5 M) (Table 1). In
contrast, tannic acid was the most effective inhibitor of AHHactivity in microsomes prepared from MCA-treated animalsshowing a Iso of 4.4 x 10~5M followed by quercetin (5.4 x 10~5M), myricetin (5.8 x 10~5M), anthraflavic acid (11.8 x 10~5M),
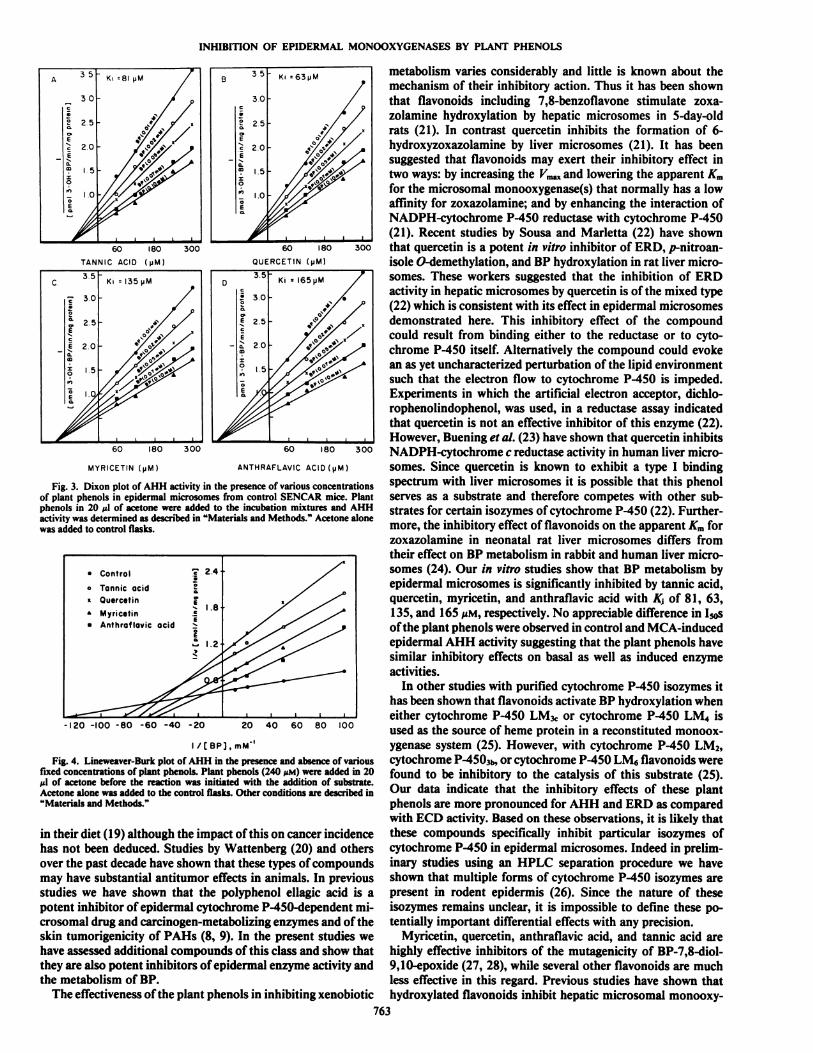
and digallic acid (no inhibition). The kinetics of inhibition ofthese plant phenols on epidermal AHH activity is depicted inFig. 3. Tannic acid, quercetin, myricetin, and anthraflavic acidinhibited epidermal AHH activity with K, of 81, 63, 135, and165 MM,respectively (Fig. 3). Lineweaver-Burk plots of epidermal AHH activity in the presence and absence of fixed concentrations of the plant phenols are shown in Fig. 4. The Kmvaluefor AHH activity in epidermal microsomes prepared fromcontrol animals was found to be 7.9 MMwhile in the presenceof 240 MMtannic acid, quercetin, myricetin, or anthraflavic acidit was 12.2, 14.5, 16.4, and 18.5 MM,respectively. The ym,f wasalso altered in the presence of each of the plant phenols suggesting that the inhibitory effect is of a mixed type. The effectof these plant phenols on epidermal AHH, ECD, and ERDactivities is shown in Table 2. Tannic acid, quercetin, myricetin,and anthraflavic acid at a concentration of 240 MMresulted in57-92% inhibition of AHH activity in epidermal microsomesprepared from control and MCA-induced animals (Table 2).These plant phenols were also found to inhibit ERD activity(33-85%) and ECD activity (19-59%) (Table 2) in epidermalmicrosomes of control as well as MCA-induced animals. Eachof the plant phenols consistently exerted a greater inhibitoryeffect on ERD and AHH activities than on ECD activity.
761
INHIBITION OF EPIDERMAL MONOOXYGENASES BY PLANT PHENOLS
•Tannic acid» Quercetin•Myricetin
•Anthraflavic acid
•Tannic acid«Quercetin»Myricetin•Anthraflavic acid
120 180 240 300 120 180 240 300
CONCENTRATION ((jM) CONCENTRATION ( \¡M )Fig. 2. In vitro effect of plant phenols on epidermal AHH activity in SENCAR mice. Epidermal microsomes were prepared from control (A) and MCA-induced
l /( I SENCAR mice. Plant phenols (30-300 />M)were added to the incubation mixtures and AHH activity was determined as described in "Materials and Methods."
Data represent means of four individual values.
Table 1 Plant phenol inhibition of AHH activity in epidermal microsomesprepared from control and MCA-pretreated SENCAR mice
PlantphenolsTannic
acidQuercetinMyricetinAnthraflavic acidI»
of AHH (x10-5M)Control
MCA-inducedmicrosomesmicrosomes6.0
4.45.4 5.45.6 5.8
12.4 11.8
In Vitro Inhibitory Effect of Plant Phenols on Epidermal BPMetabolism. The effect of tannic acid, quercetin, myricetin, andanthraflavic acid on epidermal microsomal BP metabolism isshown in Fig. S. Each of the plant phenols exhibited a significant inhibitory effect on the formation of BP metabolites andtannic acid was most effective in this regard. The addition ofthese plant phenols (60 /AI) decreased the formation of BPdiols and BP-phenols by 23-67 and 29-57%, respectively,whereas they had little or no effect on the formation of BP-quinones. Tannic acid, quercetin, myricetin, and anthraflavicacid diminished BP-7,8-diol formation by epidermal microsomes. Their inhibitory effect on the formation of BP-phenolscorrelates well with their effect on epidermal AHH activity.
In Vivo Inhibitory Effect of Topically Applied Plant Phenolson Epidermal Monooxygenases. The effect of a single topicalapplication of tannic acid, quercetin, and myricetin to the skinof SENCAR mice on epidermal monooxygenase activities isshown in Fig. 6. The administered dose was limited by themaximum achievable solubility of the test compound in thevehicle. Topical application of tannic acid, quercetin, and myricetin caused a dose-dependent inhibition of epidermal AHH,ECD, and ERD activities. Topical application of 400 /<mol/kg
body weight of tannic acid inhibited epidermal AHH, ECD,and ERD activities by 53, 58, and 66%, respectively (Fig. 6/Õ).The inhibitory effect of quercetin on epidermal ERD activitywas greater than its effect on epidermal AHH or ECD activities(Fig. (ìli).Topical application of 400 ¿¿mol/kgbody weight ofquercetin inhibited AHH, ECD, and ERD activities by 65, 68,and 97%, respectively (Fig. 6B). Myricetin had its maximuminhibitory effect on ERD activity as compared to AHH andECD activity (Fig. 6C). Topical application of 400 /imol/kgbody weight of myricetin resulted in 59, 30, and 97% inhibitionof epidermal AHH, ECD, and ERD activities, respectively (Fig.6f). Anthraflavic acid had no significant inhibitory effect onany of the epidermal enzymes (data not shown). Repeatedtopical application of the plant phenols (daily for 3 days) hadinhibitory effects on monooxygenase activities no different fromthose observed after a single application (data not shown).Treatment of the SENCAR mice with the test compounds at 0h did not result in inhibition of any of the monooxygenaseactivities (data not shown) suggesting that the observed inhibitory effects are not simply due to the presence of the plantphenols in the microsomal preparations.
DISCUSSION
The use of naturally occurring compounds as an approach tocancer prevention has been increasingly studied in recent years.One category of such agents generally known as plant phenolsincludes tannins, hydroxylated flavonoids, hydroxylated anthra-quinones, and hydroxylated cinnamic acid derivatives. Thesecompounds are widely distributed in the plant kingdom (18,19). It has been estimated that some individuals in the humanpopulation ingest approximately l g of plant phenols per day
762
INHIBITION OF EPIDERMAL MONOOXYGENASES BY PLANT PHENOLS
3 0 -
S 25
20 -
1.5
30
2 0
. 1.0
60TANNIC ACID
60QUERCETIN
ISO 300
Ki = 135 JJM
60 ISO 300 60 180 300
MYRICETIN (pM) ANTHRAFLAVIC ACID(pM)
Fig. 3. Dixon plot of AHH activity in the presence of various concentrationsof plant phenols in epidermal microsomes from control SENCAR mice. Plantphenols in 20 j<l of acetone were added to the incubation mixtures and AHHactivity was determined as described in "Materials and Methods." Acetone alone
was added to control flasks.
Control
Tonnic acidOuercetin
MyricetinAnthraflavic acid
24
-120 -100 -80 -60 -40 -20 40 60 80 100
I/[BP], mM"
Fig. 4. Lineweaver-Burk plot of AHH in the presence and absence of variousfixed concentrations of plant phenols. Plant phenols (240 MM)were added in 20«Iof acetone before the reaction was initiated with the addition of substrate.Acetone alone was added to the control flasks. Other conditions are described in"Materials and Methods."
in their diet (19) although the impact of this on cancer incidencehas not been deduced. Studies by Wattenberg (20) and othersover the past decade have shown that these types of compoundsmay have substantial antitumor effects in animals. In previousstudies we have shown that the polyphenol ellagic acid is apotent inhibitor of epidermal cytochrome P-450-dependent mi-crosomal drug and carcinogen-metabolizing enzymes and of theskin tumorigenicity of PAHs (8, 9). In the present studies wehave assessed additional compounds of this class and show thatthey are also potent inhibitors of epidermal enzyme activity andthe metabolism of BP.
The effectiveness of the plant phenols in inhibiting xenobiotic
metabolism varies considerably and little is known about themechanism of their inhibitory action. Thus it has been shownthat flavonoids including 7,8-benzoflavone stimulate zoxa-zolamine hydroxylation by hepatic microsomes in 5-day-oldrats (21). In contrast quercetin inhibits the formation of 6-hydroxyzoxazolamine by liver microsomes (21). It has beensuggested that flavonoids may exert their inhibitory effect intwo ways: by increasing the Vm^and lowering the apparent Kmfor the microsomal monooxygenase(s) that normally has a lowaffinity for zoxazolamine; and by enhancing the interaction ofNADPH-cytochrome P-450 reducÃasewith cytochrome P-450(21). Recent studies by Sousa and Marietta (22) have shownthat quercetin is a potent in vitro inhibitor of ERD, p-nitroan-isole O-demethylation, and BP hydroxylation in rat liver microsomes. These workers suggested that the inhibition of ERDactivity in hepatic microsomes by quercetin is of the mixed type(22) which is consistent with its effect in epidermal microsomesdemonstrated here. This inhibitory effect of the compoundcould result from binding either to the reducÃaseor to cylo-chrome P-450 ilself. Allernalively the compound could evokean as yet uncharacterized perlurbalion of Ihe lipid environmenlsuch lhal the electron flow lo cylochrome P-450 is impeded.Experimenls in which Ihe artificial electron acceptor, dichlo-rophenolindophenol, was used, in a reducÃaseassay indicatedthai quercelin is noi an effeclive inhibitor of Ihis enzyme (22).However, Buening et al. (23) have shown lhal quercelin inhibilsNADPH-cylochrome c reducÃaseaclivily in human liver microsomes. Since quercelin is known lo exhibil a type I bindingspeclrum wilh liver microsomes il is possible lhal Ihis phenolserves as a subslrale and therefore competes with other substrates for certain isozymes of cylochrome P-450 (22). Furthermore, Ihe inhibilory effecl of flavonoids on Ihe apparent Kmforzoxazolamine in neonalal ral liver microsomes differs fromIheir effecl on BP metabolism in rabbil and human liver microsomes (24). Our in vitro sludies show thai BP metabolism byepidermal microsomes is significantly inhibited by tannic acid,quercetin, myricetin, and anlhraflavic acid wilh K¡of 81, 63,135, and 165 n\i, respeclively. No appreciable difference in I50sof Ihe plani phenols were observed in conlrol and MCA-inducedepidermal AHH aclivily suggesling lhat the plant phenols havesimilar inhibitory effects on basal as well as induced enzymeactivities.
In olher sludies wilh purified cylochrome P-450 isozymes ilhas been shown lhal flavonoids activate BP hydroxylalion wheneilher cylochrome P-450 LM& or cytochrome P-450 LM, isused as the source of heme protein in a reconstiluled monoox-ygenase syslem (25). However, wilh cylochrome P-450 LMZ,cylochrome P-4503b, or cylochrome P-450 LM6 flavonoids werefound lo be inhibilory lo Ihe catalysis of Ihis substrate (25).Our data indicate that the inhibitory effects of these planiphenols are more pronounced for AHH and ERD as comparedwilh ECD aclivily. Based on these observations, il is likely lhalIhese compounds specifically inhibil particular isozymes ofcylochrome P-450 in epidermal microsomes. Indeed in preliminary sludies using an HPLC separation procedure we haveshown that multiple forms of cytochrome P-450 isozymes arepresent in rodent epidermis (26). Since the nature of theseisozymes remains unclear, it is impossible lo define Ihese potentially important differential effects wilh any precision.
Myricetin, quercelin, anlhraflavic acid, and lannic acid arehighly effeclive inhibitors of the mutagenicily of BP-7,8-diol-9,10-epoxide (27, 28), while several olher flavonoids are muchless effeclive in this regard. Previous studies have shown thaihydroxylaled flavonoids inhibil hepalic microsomal monooxy-
763

INHIBITION OF EPIDERMAL MONOOXYGENASES BY PLANT PHENOLS
Table 2 In vitro effect of plant phenols on epidermal monooxygenases in SENCAR miceAHHactivity*Plant
phenol*Acetone
Dimethyl sulfoxideTannic acidQuercetinMyricetinAnthraflavic acidControl3.30
±0.29e
2.98 ±0.260.55 ±0.04 (83)0.41 ±0.03 (88)0.28 ±0.03 (92)1.1 1 ±0.08(63)MCA38.86
±3.3036.69 ±3.21
2.81 ±0.30 (92)4.83 ±0.36 (88)4.00 ±0.33 (90)
15.83 ±0.93 (57)ECD
activityControl5.94
±0.386.03 ±0.433.57 ±0.31 (40)2.89 ±0.21 (51)2.96 ±0.30 (50)4.89 ±0.31 (19)MCA17.82
±1.4317.98 ±1.36
9.98 ±0.84 (44)7.63 ±0.74 (57)7.43 ±0.80 (58)
13.58 ±1.10 (25)ERD
activityControl0.55
±0.040.58 ±0.040.16 ±0.02 (71)0.09 ±0.01 (84)0.13 ±0.01 (76)0.39 ±0.02 (33)MCA4.78
±0.284.86 ±0.331.16 ±0.09 (76)0.87 ±0.06 (82)1.08 ±0.11 (77)3.17 ±0.09 (35)
* Enzyme assays were performed in either the absence or the presence of plant phenols (240 ¡M)in the incubation mixtures.* Enzyme specific activity is represented as pmol metabolite/min/mg protein. Percent inhibition is indicated in parentheses.' Mean ±SE of 4 values.
Fig. 5. In vitro effect of plant phenols onepidermal BP metabolism in SENCAR mice.Plant phenols (60 UM)were added to incubation flasks containing epidermal microsomesprepared from control animals. BP metaboliteswere separated on HPLC as described in "Materials and Methods." Data represent mean of
three individual values with a variance of lessthan 10%.
oaO)
eE
ô
a
0.5
CONTROL
TANNIC ACID
OUERCETIN
MYRICETIN
ANTHRAFLAVIC ACID
I 1 19,10-DIOL 4,5-DIOL 7.8-DIOL 1,6-OUINONE 3,6-OUINONE 9-HYDROXY 3-HYDROXY
genäsesystems (5, 23). An examination of the structure-activityrelationship of the hydroxylated anthraquinones, cinnamic acidderivatives, and flavonoids has indicated that phenolic hydroxylgroups on the phenyl substituent are essential for their anti-mutagenic activity (27, 28). Huang et al. (27, 28) have proposedthat the antimutagenic activity of these compounds is due tothe disappearance of bay-region diol-epoxides of PAHs as aresult of their interaction with the reactive metabolites (27, 28).The half-life for the disappearance of BP-7,8-diol-9,10-epoxidefrom aqueous solution was 43 min in the absence of inhibitorand 9, 4.5, and 1.2 min in the presence of 10 pM quercetin,myricetin, and tannic acid, respectively (27, 28). Previous studies have shown that ellagic acid possesses strong antimutagenicand antitumorigenic activity towards PAHs (9, 10, 29) and is astrong antagonist of bay region diol-epoxides of the PAHs dueto the formation of adducts (29, 30).
Another class of naturally occurring compounds known asthe ellipticines also prevents chemically induced mutagenesisand carcinogenesis (31,32). This effect of ellipticines is thoughtto relate to their ability to inhibit monooxygenases, such asAHH (33). Ellipticines are metabolized by cytochrome P-450to 9-hydroxyellipticine which is responsible for the inhibitoryeffects of the compound on the monooxygenase enzyme systemand on PAH-DNA adduct formation in vitro (34). In the presentstudy we have shown that selected hydroxylated anthraquinonesand hydroxylated flavonoids exert strong inhibitory effectsagainst carcinogen-metabolizing enzymes. This may be one
mechanism whereby they diminish the mutagenicity of bay-region diol-epoxides of PAHs. However, several of these hydroxylated flavonoids may themselves be mutagenic (35). Inaddition, there is some controversy about the carcinogenicityof quercetin (35, 36). Quercetin also appears to have limitedWonv;iilability because of poor absorption from the gastrointestinal tract (37). Tannic acid has a variety of biological effectsincluding the capacity to inactivate influenza and chickungunyaviruses (38, 39), to antagonize the effects of several snakevenoms and bacterial toxins (40), to reduce the permeability oferythrocytes to anions (41) or nonelectrolytes (42), and toinduce the agglutination of erythrocytes (42). Tannic acid hasbeen shown to possess strong hepatotoxic effects when administered to rodents (43). Many of these effects appear to berelated to the ability of tannic acid to bind indiscriminately toprotein (44).
The requirement for metabolic activation of PAHs into ultimate carcinogenic metabolites suggests that inhibitors of cytochrome P-450-dependent enzyme activity might be capable ofinterfering with this process thereby affording protectionagainst chemically induced carcinogenesis. Such inhibitors aregenerally known as anticarcinogens. One such plant phenol,ellagic acid, has been shown to inhibit the covalent binding ofBP to DNA of mouse skin (9) and lung (12) and subsequentlyinhibit PAH-induced pulmonary and skin tumors in mice (9,10). Other inhibitors of AHH activity such as benzoflavone andclotrimazole have also been found to inhibit PAH-induced
764

INHIBITION OF EPIDERMAL MONOOXYGENASES BY PLANT PHENOLS
2.0 ^
; i.O
3.0
Ì. 2.0
L0
o¡Z 1.5
uïï 1.0¡n
0.5
'ARYL HYDROCARBON HYDROXYLASE B
ETHOXYCOUMARIN 0 •DEETHYLASE
lililÃETHOXYRESORUFIN 0-DEETHYLASE
2.0 -
0 50 100 200 300 400
TANNIC ACID ( pmol/kg)
0 50 100 200 300 400
OUERCETIN ((imol/kg)
ain
2.0
3.0
2.0
1.5
05
T ARYL HYDROCARBON HYDROXYLASE
HimETHOXYCOUMARIN 0-DEETHYLASE
HUMETHOXYRESORUFIN 0-DEETHYLASE
Ih..0 50 100 200 300 400
MYRICETIN (|jmol/kg)Fig. 6. Effect of topical application of tannic acid, quercetin, and myricetin on epidermal monooxygenases in SENCAR mice. Doses of the compounds were
applied topically to SENCAR mice. Monooxygenase activities were measured in microsomes as described in "Materials and Methods." Data represent mean of four
values; bars, SE. Left, tannic acid; middle, quercetin; right, myricetin.
tumorigenicity when applied topically to mice (5-7). In thepresent study tannic acid, quercetin, myricetin, and anthraflavicacid resulted in significant inhibition of the formation of BP-7,8-diol, a precursor of the ultimate carcinogenic metabolite ofthis hydrocarbon. It is therefore possible that these plant phenols or structurally related compounds may prove useful asantagonists for PAH tumorigenicity. In addition our resultssuggest that dietary plant phenols may have potential usefulnessin modulating the pharmacological and/or toxicological effectsof drugs and other foreign chemicals in animals and in humans.
ACKNOWLEDGMENTS
Thanks are due to James D. Steele for technical assistance and toSandra Evans for preparing the manuscript.
REFERENCES
1. Conney, A. H. Induction of microsomal enzymes by foreign chemicals andcarcinogenesis by polycyclic aromatic hydrocarbons: G. H. A. Clowes Memorial Lecture. Cancer Res., 42:4875-4917, 1982.
2. Gram, T. E. Extrahepatic Metabolism of Drugs and Other Foreign Compounds. Jamaica, NY: Spectrum Publications, 1978.
3. Mannering, G. J. Microsomal enzyme systems which catalyze drug metabolism. In: B. N. LaDu, N. G. Mandel, and E. L. Way (eds.). Fundamentals ofDrug Metabolism and Disposition, pp. 206-252. Baltimore: Williams &Wilkins, 1971.
4. Gelboin, H. V. Benzo(a)pyrene metabolism, activation and carcinogenesis:role of regulation of mixed function oxidases and related enzymes. Physiol.Rev., 60: 1107-1166, 1980.
5. Weibel, F. J., Gelboin, H. V., Buu-Hoi, N. P., Stout, M. G., and Burnham,W. S. Flavones and polycyclic aromatic hydrocarbons as modulators of arylhydrocarbon (benzo(a)pyrene] hydroxylase. In: P. O. P. Ts'o and J. T.Dipaolo (eds.), Chemical Carcinogenesis, pp. 249-270. New York: MarcelDekker, Inc., 1974.
6. Slaga, T. J., Thompsom, S., and Berry, D. L. The effect of benzoflavones onpolycyclic aromatic hydrocarbon metabolism and skin tumor initiation.Chem.-Biol. Interact., 17: 297-312, 1977.
7. Mukhtar, H., DelTito, B. J., Das, M., Cherniack, E. P., Cherniack, A. D.,and Bickers, D. R. Clotrimazole, an inhibitor of epidermal benzo(a)pyrenemetabolism, DNA-binding and carcinogenicity of the hydrocarbon. CancerRes., 44: 4233-4240, 1984.
8. Das, M., Bickers, D. R., and Mukhtar, H. Effect of ellagic acid on hepatic
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22,
23.
and pulmonary xenobiotic metabolism in mice: studies on the mechanism ofits anticarcinogenic action. Carcinogenesis (Lond.), 6:1409-1413, 1985.Mukhtar, H., Das, M., DelTito, B. J., and Bickers, D. R. Protection against3-methylcholanthrene induced skin tumorigenesis in BALB/c mice by ellagicacid. Biochem. Biophys. Res. Commun., 119:751-757, 1984.Lesea, P. Protective effects of ellagic acid and other plant phenols onbenzo(a)pyrene induced neoplasia in mice. Carcinogenesis (Lond.), 4:1651-1653, 1983.Chang, R. L., Huang, M. T., Wood, A. W., Wong, C. Q., Newmark, H. L.,Yagi, H., Sayer, J. M., Jerina, D. M., and Conney, A. H. Effect of ellagicacid and hydroxylated flavonoids on the tumorigenicity of benzo(a)pyreneand (±)-7Ì8,8a-dihydroxy-9o,10a-epoxy-7,8,9,10-tetrahydrobenzo(a)pvreneon mouse skin in the newborn mice. Carcinogenesis (Lond.), 6: 1127-1134,1985.Dixit, R., Teel, R. T., Daniel, F. B., and Stoner, G. D. Inhibition ofbenzo(a)pyrene and benzo(o)pyrene-trans-7,8-diol metabolism and DNAbinding in mouse lung expiants by ellagic acid. Cancer Res., 45: 2951-2956,1985.Das, M., Bickers, D. R., Santella, R. M., and Mukhtar, H. Altered patternsof cutaneous xenobiotic metabolism in ultraviolet-B induced squamous cellcarcinoma in SKH-1 hairless mice. J. Invest. Dermatol., 84: 532-536, 1985.Neben, D. W., and Gelboin, H. V. Substrate-inducible microsomal arylhydrocarbon hydroxylase in mammalian cell culture. I. Assay and propertiesof induced enzyme. J. Biol. Chem., 243:6242-6249, 1968.Greenlee, W. F., and Poland, A. An improved assay of 7-ethoxycoumarin O-deethylase activity: induction of hepatic enzyme activity in C57BL/6J andDBA/2J mice by phénobarbital, 3-methylcholanthrene and 2,3,7,8-tetra-chlorodibenzo-p-doxin. J. Pharmacol. Exp. Ther., 205: 596-605, 1978.Pohl, R. J., and Fouts, J. R. A rapid method for assaying the metabolism of7-ethoxyresorufin by microsomal subcellular fractions. Anal. Biochem., 107:150-155, 1980.Lowry, O. H., Rosebrough, N. J., Fair, A. L., and Randall, R. J. Proteinmeasurement with Folin phenol reagent. J. Biol. Chem., 193:265-275,1951.Grisebach, H. Selected topics in flavonoid biosynthesis. In: T. Swain, J. B.Harborne, and C. F. Van Sumere (eds.), Recent Advances in Photochemistry,Vol. 12, pp. 221-248. New York: Plenum Publishing Corp., 1979.Harborne, J. B. Flavonoids and the evolution of the angiosperms. Biochem.Syst. Ecol., 5:7-12, 1977.Wattenberg, L. W. Inhibition of neoplasia by minor dietary constituents.Cancer Res. (Suppl.), 43: 2448s-2453s, 1983.Lasker, J. M., Huang, M. T., and Conney, A. H. In vitro and in vivoactivationof oxidative drug metabolism by flavonoids. J. Pharmacol. Exp. Ther., 229:162-170, 1984.Sousa, R. L., and Marietta, M. A. Inhibition of cytochrome P-450 activityin rat liver microsomes by the naturally occurring flavonoid, quercetin. Arch.Biochem. Biophys., 240: 345-357, 1985.Buening, M. K., Chang, R. L., Huang, M. T., Fortner, J. G., Wood, A. W.,and Conney, A. H. Activation and inhibition of benzo(a)pyrene and aflatoxin
765

INHIBITION OF EPIDERMAL MONOOXYGENASES BY PLANT PHENOLS
B, metabolism in human liver microsomes by naturally occurring flavonoids.Cancer Res., 41:67-72, 1981.
24. Huang, M. T., Chang, R. L., Fortner, J. G., and Conney, A. H. Studies onthe mechanism of activation of microsomal benzo(a)pyrene hydroxylationby flavonoids. J. Biol. Chem., 256:6829-6836, 1981.
25. Huang, M. T., Johnson, E. F., Mueller-Eberhardt, U., Koop, D. R., Coon,M. J., and Conney, A. H. Specificity in the activation and inhibition byflavonoids of benzo(a)pyrene hydroxylation by cytochrome P-450 isozymesfrom rabbit liver microsomes. J. Biol. Chem., 256:10897-10901, 1981.
26. Bickers, D. R., Das, M., Steele, J. D., and Mukhtar, H. Multiplicity ofepidermal cytochrome P 450: HPLC separation of solubili/ed microsomesin neonatal Sprague-Dawley rats. J. Invest. Dermatol., 87: 130, 1986.
27. Huang, M. T., Wood, A. W., Newmark, H. L., Sayer, J. M., Yagi, H., Jerina,D. M., and Conney, A. H. Inhibition of the mutagenicity of bay-region diol-epoxides of polycyclic aromatic hydrocarbons by phenolic plant flavonoids.Carcinogenesis (Lond.) 4: 1631-1637, 1983.
28. Huang, M. T., Chang, R. L., Wood, A. W., Newmark, H. L., Sayer, J. M.,Yagi, H., Jerina, D. M., and Conney, A. H. Inhibition of mutagenicity ofbay-region diol epoxides of polycyclic aromatic hydrocarbons by tannic acid,hydroxylated anthraquinones and hydroxylated cinnamic acid derivatives.Carcinogenesis (Lond.), 6: 237-242, 1985.
29. Wood, A. W., Huang, M. T., Chang, R. L., Newmark, H. L., Lehr, R. E.,Yagi, H., Sayer, J. M., Jerina, D. W., and Conney, A. H. Inhibition ofmutagenicity of bay region diol epoxides of polycyclic aromatic hydrocarbonsby naturally occurring plant phenols: exceptional activity of ellagic acid.Proc. Nati. Acad. Sci. USA, 79: 5513-5517, 1982.
30. Sayer, J. M., Yagi, H., Wood, A. W., Conney, A. H., and Jerina, D. W.Extremely facile reaction between ultimate carcinogen benzo(a)pyrene-7,8-diol-9,10-epoxide and ellagic acid. J. Am. Chem. Soc., 104: 5562-5564,1982.
31. Lesea, P., Lecointe, P., Paoletti, C, and Mansuy, D. Ellipticines as potentinhibitors of microsome dependent chemical mutagenesis. Chem.-Biol. Interact., 25: 279-287, 1979.
32. Lesea, P., and Mansuy, D. 9-Hydroxyellipticine: inhibitory effect on skinCarcinogenesis induced in Swiss mice by 7,12-dimethylbenz(a)anthracene.Chem.-Biol. Interact., 30:181-187, 1980.
33. Lesea, P., Lecointe, P., Paoletti, C., and Mansuy, D. Ellipticines as potent¡inhibitorsof aryl hydrocarbon hydroxylase: their binding to microsomalcytochrome P-450 and protective effect against benzo(a)pyrene mutagenicity.Biochem. Pharmacol., 27: 1203-1209, 1978.
34. Lesea, P., Lecointe, P., Paoletti, C., and Mansuy, D. The hydroxylation ofthe antitumor agent, ellipticine, by liver microsomes from differently pre-treated rats. Biochem. Pharmacol., 26:2169-2173, 1977.
35. Brown, J. P. A review of the genetic effects of naturally occurring flavonoids,anthraquinolines and related compounds. Mutât.Res., 75: 243-277, 1980.
36. Saito, D., Shirai, A., Matsushima, T., Sugimura, T., and Hirono, I. Test ofcarcinogen icity of quercetine, a widely distributed mutagen in food. Teratog.Carcinog. Mutagen. /: 213-221, 1980.
37. Cingler, R., Leschik, M., and Dengler, J. J. Disposition of quercetin in manafter single and intravenous doses, luir. J. Clin. Pharmacol., 9: 229-234,1975.
38. Carson, R. S., and Frisch, A. W. The inactivation of influenza virus by tannicacid and related compounds. J. Bacterio!., 66: 572-575, 1953.
39. Konishi. I:.., and Hot ta. S. Effects of tannic acid and its related compoundsupon chikungunya virus. Microbiol. Immunol., 23:609-667, 1979.
40. Okonogi, T., Hattoriz, Z., Ogiso, A., and Mitsui, S. Detoxification bypersimmon tannin of snake venoms and bacterial toxins. Toxicon, 17: 524-527, 1979.
41. Jacobs, M. H., Stewart, D. R., and Butler, M. K. Some effects of tannic acidon the cell surface. Am. J. Med. Sci., 205: 154-157, 1943.
42. Hunter, F. R. The effect of tannic acid on the permeability of erythrocytes tononelectrolytes. J. Cell. Comp. Physiol., 55:175-188, 1960.
43. Oler, A., Neal, M. W., and Mitchell, E. K. Tannic acid: acute hepatotoxicityfollowing administration by feeding tube. Food Cosmet. Toxicol., 4: 565-569, 1976.
44. Hall, C. B. Inhibition of tannats pectinesterase by tannic acid. Nature (Lond.),2/2:717-718,1966.
766
Copyright © 2022 FDOKUMEN




















