
EXW EXW FCA FCA CPT CPT CIP DAT DAP DDP CIP DAT DAP DDP FAS FOB CFR CIF FAS FOB CFR CIF
Upload
independentCategory
view
3download
0
Ex vivo pediatric brain tumors express Fas(CD95) and FasL (CD95L) and are resistantto apoptosis induction1
Christopher D Riffkin Amy Z Gray Christine J Hawkins CW Chow and David M Ashley2
Department of Hematology and Oncology (CDR AZG CJH DMA) and Department of Anatomical Pathology (CWC) Royal Childrenrsquos Hospital Flemington Road Parkville 3052 Melbourne Victoria Australia
Neuro-Oncology n OCTOBER 200 1 229
Fas (APO-1CD95TNFRSF6) is a member of the tumornecrosisnerve growth factor receptor family that signalsapoptotic cell death in sensitive cells Expression of Fasand its agonistic ligand (FasLTNFSF6) was investigatedin ex vivo pediatric brain tumor specimens of various his-tologic types Fas expression was identied in all of the 18tumors analyzed by ow cytometry and immunohisto-chemistry FasL expression was identied in most of the13 tumors analyzed by both Western analysis andimmunohistochemistry Nine of these tumor specimenswere treated with either the agonistic anti-Fas antibody(APO-1) in combination with protein A or FasL in short-term cytotoxicity assays Sensitivity to apoptosis inducedby the topoisomerase II inhibitor etoposide was alsoassessed Despite the presence of Fas all the specimensanalyzed demonstrated a high degree of resistance to Fas-mediated apoptosis These 9 specimens also showed ahigh degree of resistance to etoposide Only 2 of the 9specimens were susceptible to etoposide-induced celldeath whereas only 3 were sensitive to Fas-mediated apo-ptosis One brain tumor was sensitive to both Fas ligationand etoposide treatment This contrasted with the highdegree of susceptibility to both etoposide- and Fas-
induced apoptosis observed in the reference Jurkat cellline The results suggest that Fas expression may be a gen-eral feature of tumors of the CNS and that a signicantdegree of resistance to Fas-mediated apoptosis may existin ex vivo pediatric brain tumor specimens Neuro-Oncol-ogy 3 229ndash240 2001 (Posted to Neuro-Oncology [serialonline] Doc 01-002 July 19 2001 URL ltneuro-oncol-ogymcdukeedugt)
Brain tumors are the second most frequently occur-ring neoplasms of childhood (Cohen and Duffner1994) and are associated with high rates of mor-
bidity and mortality Current treatment approaches usingcombinations of surgical resection chemotherapy andradiotherapy not only fail to effect a cure in many casesbut also are themselves associated with short- and long-term adverse effects Thus innovative approaches totherapy that reduce morbidity and improve survival aredesperately needed
CD95FasAPO-1TNFRSF6 is a 45-kDa type I inte-gral membrane protein belonging to the tumor necro-sisnerve growth factor superfamily of receptors (Itoh etal 1991 Oehm et al 1992) The physiologic ligand ofthe Fas receptor FasL is a type II integral membrane pro-tein homologous to TNF-a3 (Kayagaki et al 1997 Sudaet al 1993) which may also be released from the cellsurface as a functional soluble 26-kDa molecule (Kaya-gaki et al 1995 Tanaka et al 1995) Fas belongs to anemerging subfamily of TNF receptors including TNF-R1 TRAIL-R1 TRAIL-R2 and APO-3 (Armitage1994) that are characterized by the presence of a deathdomain within the cytoplasmic region (Itoh et al 1993)Multimerization of Fas receptors by FasL or agonisticantibodies leads to aggregation of the intracellular deathdomains (Boldin et al 1995) Association of adapter
Received 10 January 2001 accepted 2 April 2001
1Supported by the James S McDonell Foundation Royal ChildrensResearch Institute and National Health and Medical Research Council
2Address correspondence and reprint requests to David M AshleyDepartment of Haematology and Oncology Royal Childrenrsquos HospitalFlemington Road Parkville 3052 Melbourne Victoria Australia
3Abbreviations used are as follows FACS uorescence-activated cellsorting MFR mean uorescence ratio PBS phosphate-buffered salinePNET primitive neuroectodermal tumor TNF tumor necrosis factor
Neuro-Oncology by guest on M
arch 22 2015httpneuro-oncologyoxfordjournalsorg
Dow
nloaded from
proteins with the death domains (Kischkel et al 1995)initiates an intracellular proteolytic cascade involvingcaspases (Boldin et al 1996 Medema et al 1997Muzio et al 1996 Scafdi et al 1998) that culminatesin cleavage of cellular substrates and apoptosis in sensi-tive cells (Trauth et al 1989 Yonehara et al 1989)
Fas-FasL interactions play important roles in elimi-nation of autoreactive immune cells (Davidson et al1998 Watanabe-Fukunaga et al 1992) activation-induced T-cell death (Singer et al 1994) and killing bycytotoxic T lymphocytes (Hanabuchi et al 1994) andnatural killer cells (Oshimi et al 1996) FasL has beenimplicated in maintaining immune privilege in the ante-rior chamber of the eye (Grifth et al 1995 1996) inthe mouse testis (Bellgrau et al 1995 Duke et al 1996Yagita et al 1996) and in the brain (Saas et al 1997)FasL is believed to deliver a death signal to the Fas-expressing immune cells inltrating these tissues result-ing in suppression of immune activation
Expression of Fas has been identied in glioma(Weller et al 1994) and medulloblastoma cell lines(Weller et al 1998) and in ex vivo astrocytic tumor spec-imens (Frei et al 1998 Tohma et al 1998 Tachibanaet al 1995) Expression of Fas mRNA in astrocytomasin vivo tends to correlate with a high degree of malig-nancy (World Health Organization grades III and IVTachibana et al 1995) However the Fas proteinappears to be predominantly expressed in associationwith large areas of necrosis within tumor tissue (Tohmaet al 1998) Furthermore both human glioma cell lines(Weller et al 1994 1995a 1995b) and ex vivo gliomacells (Frei et al 1998) have been shown to be suscepti-ble to apoptosis induced by FasL or anti-Fas antibodiesPreexposure of cell lines to various cytokines includingTNF-a and interferon-g (Weller et al 1994) andincreased production of endogenous TGF-b1 by gliomacells (Ashley et al 1998) were shown to enhance Fascytotoxicity This and the absence of Fas on normalbrain parenchyma (Leithauser et al 1993) led to inter-est in Fas as the target of a novel immunotherapeuticapproach against these tumors
Malignant gliomas are among a number of solidtumors including colon carcinoma (OrsquoConnell et al1996) hepatocellular carcinoma (Strand et al 1996)esophageal carcinoma (Gratas et al 1998) andmelanoma (Hahne et al 1996) that express FasL (Saaset al 1997 Weller et al 1998) Membrane-boundFasL on cell lines (Hahne et al 1996 Strand et al1996) or ex vivo cells (Saas et al 1997 OrsquoConnell etal 1996 Walker et al 1997) is capable of inducingcell death in Fas-sensitive target cells This is believed torepresent a mechanism by which tumors can counterat-tack tumor-inltrating lymphocytes and cytotoxic Tcells and thereby escape immune surveillance (OrsquoCon-nell et al 1996)
It is not known whether the previously publishedndings from adult malignant glioma can be generalizedto include other tumors of the CNS including pediatricmalignancies In this report Fas expression was identi-ed in all of the pediatric brain tumors tested FasL wascoexpressed in 9 of the 13 tumors tested by both West-ern blot and immunohistochemistry Despite the pres-
ence of the receptor on tumor cells a large degree ofresistance to Fas-mediated apoptosis was observed in allthe tumor specimens
Methods
Cell Lines
Neuro-2A murine neuroblastoma cell lines were pro-vided by Dr Adriano Fontana (University HospitalZurich Switzerland) Neuro-2AFasL cells have beenengineered to produce soluble FasL by stable transfectionwith the BCMGS expression plasmid subcloned withcDNA encoding murine FasL (Rensing-Ehl et al 1995)Murine and human FasL share 769 homology at anamino acid level and are functionally interchangeable(Nagata and Golstein 1995) Neuro-2Aneo cells trans-fected with an empty BCMGS vector served as a controlin experiments using the FasL-containing supernatantuid Cell lines were maintained in zinc option medium(GIBCO BRL Grand Island NY) supplemented with10 heat-inactivated fetal calf serum (Trace BiosciencesCastle Hill New South Wales Victoria Australia)
Dissociation of Brain Tumor Specimens
Primary pediatric tumor specimens that were surplus topathology were collected during surgical procedures atthe Royal Childrenrsquos Hospital Melbourne Victoria Aus-tralia after Ethics in Human Research Committeeapproval Due to ethical and logistical considerations itwas not always possible to obtain a truly representativesample of the tumor If immediate processing of tumorspecimens was not feasible specimens were stored at 4degCfor less than 12 h Tumor samples were maintained inzinc option medium10 fetal calf serum mechanicallydissociated and then triturated by passage through 18-to-25-gauge needles Any solid debris preventing passageof the cell suspension was removed and discarded Suc-cessful passage through a 25-gauge needle indicated asingle cell suspension had been obtained as conrmed bymicroscopic examination Red blood cells were lysed byincubation in ammonium chloride solution (015MNH4Cl 20 mM Tris pH 74) for 2 min at room temper-ature and tumor cells were resuspended in culturemedium After assessment of viable cell number by try-pan blue exclusion (Sigma St Louis Mo) the tumor sus-pension was diluted to obtain a nal concentration of 106
viable cellsml
Labeling Cells for Immunophenotyping
FACS analysis was performed on freshly obtained tumorcell suspensions Cells (106) were centrifuged (500g 5min 4degC) washed in FACS buffer (1 bovine serumalbumin 005 NaN3 in PBS pH 74) and incubatedwith an anti-Fas monoclonal antibody (DX2 mouseIgG1 PharMingen San Diego Calif 1 mgml) or an iso-type control antibody for 30 min on ice The cells werewashed with FACS buffer and incubated with a uores-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001230
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
cein isothiocyanatendashconjugated afnity-isolated sheepanti-mouse secondary antibody [F(ab)2 fragment](Amrad Pharmacia Melbourne Victoria Australiadiluted 140) After incubation on ice for 30 min thecells were washed in FACS buffer resuspended in 200 mlFACS xative (09 NaCl 2 glucose 2 formalde-hyde) and incubated at 4degC until analyzed Referencecells were kept unstained after the rst wash and resus-pended in FACS xative The Jurkat cell line served as apositive control for each experiment and where the vol-ume of tumor suspension allowed the experiment wasconducted in duplicate
FACS Analysis of Tumor Specimens
A total of 10000 events were collected on the FACScan(Becton Dickinson Immunocytometry Systems San JoseCalif) using CYCLOPSTM software (Cytomation IncFort Collins Colo) The laser was tuned at 488 nm withan output power of 15 mW Fluorescein isothio-cyanatendashemitted ldquogreenrdquo uorescence was collected via aDM560-nm225deg dichroic mirror (gt560-nm reection)and a DF53030-nm band pass lter Cellular debris wasexcluded from the analysis
The MFR was calculated as the ratio of the mean u-orescence of DX2 (anti-Fas)-labeled cells and that of cellslabeled with the isotype control antibody
To quantitate the percentage of tumor cells that werepositive for Fas expression we used WinMDI Version27 software (Scripps Research Institute La Jolla Calif)to directly compare the uorescence prole of DX2-stained cells with the prole of cells stained with the iso-type control The DX2- and isotype-stained cell proleswere overlaid on the same histogram A marker was setdened at its lower limit by the point of intersection ofthe 2 proles The percentage of cells considered positivefor Fas was calculated by subtracting the percentage ofisotype-labeled cells from the percentage of DX2-labeledcells uorescing within the region dened by the marker
Western Blotting
Thirteen frozen tumor samples were selected accordingto availability of tumor tissue and coarsely choppedbefore suspension in 5 volumes (wtvol) RIPA buffer (1sodium dodecyl sulfate 1 NP40 in PBS pH 74) withprotease inhibitor cocktail set 1 (Calbiochem La JollaCalif) Tumor samples were homogenized using a Poly-tron homogenizer and boiled in sample buffer before sep-arating the proteins on 10 sodium dodecylsulfatendashpolyacrylamide gels and transferring them toHybondTM-P membranes (Amersham Pharmacia BiotechBuckinghamshire UK) Supernatant uid from Neuro-2AFasL-expressing cells was used as a positive controlfor FasL expression Membranes were blocked for 1 h atroom temperature in 5 nonfat milkTBST (01Tween 20 125 mM NaCl and 25 mM Tris pH 74)Blots were then probed overnight at 4degC with rabbitpolyclonal anti-human FasL antibody (Q20 Santa CruzBiotechnology Santa Cruz Calif 1100) or anti-b2-microtubulin (PharMingen 1100) as a loading controlQ20 anti-FasL antibody was selected because of its min-
imal cross-reactivity when compared with NOK-1 andG247-4 (both PharMingen) Blots were then incubatedwith horseradish peroxidasendashconjugated secondary anti-bodies (1100000) followed by enhanced chemilumines-cence detection with Supersignalreg West Pico Chemilumi-nescent Substrate according to the manufacturerrsquosinstructions (Pierce Rockford Ill) The blots wereexposed to Kodak XR lm (Eastman Kodak RochesterNY) for 2 s to 20 min
Immunohistochemical Detection of Fas and FasL
Formalin-xed parafn-embedded blocks of each tumorwere obtained from the Department of AnatomicalPathology Royal Childrenrsquos Hospital Melbourne Victo-ria Australia Sections 4-mm thick were cut and placedon poly-L-lysinendashcoated slides (Sigma) The sections weredeparaffinized in xylene and rehydrated Antigenretrieval was performed by immersing the slides for 30min in 001 M sodium citrate buffer preheated to 95degC(Kawai et al 1994 Werner et al 1996) Endogenousperoxidase activity was quenched by incubating the sec-tions in 1 hydrogen peroxide for 20 min A blockingstep was performed for 2 h at room temperature withPBS containing 5 normal goat serum (Vector Labora-tories Inc Burlingame Calif) and 10 normal humanserum Sections were incubated overnight at 4degC withrabbit polyclonal anti-human FasL antibody (Q20 SantaCruz Biotechnology 1100) or anti-human Fas antibody(Ab-1 Calbiochem 140) After 3 washes in PBS the sec-tions were incubated with biotin-conjugated goat anti-rabbit antibody (PharMingen 1200) for 30 min at roomtemperature The reaction was visualized using the Vec-tastain Elite ABC kit (Vector Laboratories Inc) and a33 9 -diaminobenzidine substrate (Sigma) The slides werecounterstained in hematoxylin (Shandon Southern Prod-ucts Ltd RuncornCheshire UK) Formalin-fixedparafn-embedded sections of human liver and mousespleen served as positive controls for Fas and FasLrespectively Negative controls consisted of sections with-out primary or secondary antibody and those incubatedwith nonimmunized rabbit serum Antibodies and con-trol rabbit serum were diluted in 1 normal goat serumin PBS Representative sections on which immunohisto-chemistry was not performed were stained with hema-toxylin and eosin (Shandon Southern Products Ltd) foridentication and classication of the tumor
The level of Fas and FasL expression was determined byassessment of the percentage of immunoreactive cells in 10nonadjacent high-powered elds (original magnicationacute400) The average of the 10 elds was calculated andrecorded as either no expression lt5 cells positive 5 to25 cells positive 25 to 50 cells positive or gt50cells positive The staining intensity of tumor cells was ratedas weak moderate or intense
Preparation of Supernatant Fluid from Neuro-2A Cell Lines
Supernatant uid was collected from conuent 10-mlplates of Neuro-2AFasL and Neuro-2Aneo cells andcentrifuged (500g 15 min) to remove cells and debris
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 231
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
The cell-free supernatant uid was then used at the dilu-tions specied or stored at ndash20degC until used
Flow Cytometric Determination of Cell Viability by FACS Analysis
In a 12-well plate 1 ml tumor cells at a viable cell con-centration of 106ml was incubated in triplicate with eachmedium alone etoposide (10 mgml) and APO-1 anti-body (100 ngml) with protein A (110000) for 24 h(37degC 5 CO2 96 humidity) For specimens PN2and MB1 the combination of APO-1 and protein A wasreplaced with Neuro-2AFasL supernatant uid diluted12 and cells exposed to an equivalent dilution of Neuro-2Aneo supernatant uid served as a negative controlWells containing Jurkat cells at 106ml served as positivecontrols for each condition
At 24 h all cells were harvested from each well by vig-orous pipetting Adherent cells were removed by incu-bating with trypsin-EDTA Each well was washed 3 timeswith PBS and lack of residual cells was conrmed bymicroscopic examination The cells were centrifuged(500g 5 min 4degC) resuspended in 300 mL FACS buffercontaining 50 mgml propidium iodide (Sigma) and incu-bated on ice for 1 h
Sub G1 Peak Analysis of Apoptotic Cell Death
After assessing viability by ow cytometry we xed thecells for sub G1 peak analysis (Tounekti et al 1995) byadding 250 ml of 70 ethanol to achieve a nal concen-tration of approximately 35 After overnight incuba-tion at 4degC samples were centrifuged (500g 5 min 4degC)and resuspended in 200 ml propidium iodide at 50 mgmlin PBS
Between 10000 and 20000 events were collected foranalysis on the FACScan using settings described foranalysis of viability Of the events remaining after dou-blet discrimination cells uorescing below 25 of theG1 peak uorescence were assumed to be debris and wereexcluded from analysis Cells uorescing above this limitbut below the intensity of the G1 peak were designatedapoptotic and the percentage of total tumor cells theyrepresented was calculated
Colorimetric Assay for Determining Total Viable Cell Number
Cell viability across a 48-h period was measured usingthe CellTiter 96regAQueous Non-Radioactive Cell Prolifera-tion Assay (Promega Madison Wisc) which detectsmitochondrial activity according to the recommendedprotocol In a 96-well plate 50 ml tumor cell suspensionat 4 acute 105ml was incubated with etoposide (10 mgml)or anti-Fas antibody APO-1 (100 ngml) with protein A(110000) or in the absence of inducers of apoptosis ina nal volume of 100 ml In later experiments (specimensMB1 and PN2) the combination of APO-1 and proteinA was replaced with Neuro-2AFasL supernatant uiddiluted 12 with parallel wells containing Neuro-2Aneosupernatant uid diluted 12 serving as a negative con-trol The experiment was conducted in triplicate and
incubated over a 48-h period (37degC 5 CO2 96humidity) Jurkat cells at a nal concentration of 2 acute105ml were used as a positive control for each treatmentWells containing 100 ml medium and substrate provideda measurement of the background reading at 490 nmresulting from absorbance within the medium at thiswavelength
Substrate (20 mlwell) was added at 0- 24- and 48-htime points after the assay was established and after afurther 3-h incubation the absorbance at 490 nm wasmeasured with an ELISA plate reader (reference wave-length 620 nm) Absorbance readings were adjusted toremove the contribution of background absorbance andviability was calculated at each time point as a percent-age of the average absorbance from wells containing noapoptosis inducers at the same time point
Statistical Analysis
We determined the signicant differences between treat-ments in individual tumor specimens by using Studentrsquos ttest (Altman 1991) We compared the overall differencein viability in the anti-Fas treatment and control groupsin the medulloblastomaPNET specimens using two-wayANOVA on Stata Corporation 1997 (Stata StatisticalSoftware Release 50 College Station Tex)
Results
Tumor Samples
The 18 pediatric brain tumor specimens analyzed forexpression of Fas and FasL are described in Table 1according to histology patient age and sex The speci-mens consisted of 9 PNETs that included 6 medulloblas-tomas 4 glioblastomas multiforme 3 low-grade gliomasa ganglioglioma and a germ-cell tumor
Fas Expression
Immunohistochemistry Giemsa staining of tumor sec-tions revealed morphologic differences between tumorcells as well as necrotic and apoptotic tissue Small num-bers of normal brain cells epidermal cells and bloodcells were also present in the samples Immunohisto-chemically the Fas receptor was detected in all 18 tumorspecimens (Table 2) The cytoplasm and membrane ofpositive tumor cells stained diffusely Walls of blood ves-sels red blood cells large areas of necrotic tissue andareas of desmoplasia were negative and staining wasconsistently absent in negative controls without primaryor secondary antibody
Tumor sections with little variation in cellular mor-phology demonstrated uniform intensity of Fas stainingthroughout In perinecrotic areas of tumor tissue anapparent increase in the number and staining intensity ofFas-positive tumor cells was frequently observed Tumorswere rated according to the percentage of tumor cellsexpressing the Fas receptor and the staining intensity ofthese cells (Table 2)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001232
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
PNETmedulloblastoma Weakly Fas-positive cellswere found to constitute 25 to 50 of cells in 4 of the9 PNETmedulloblastoma specimens and more than50 of cells in another 4 tumors of this group In theremaining medulloblastoma only occasional cellsappeared immunoreactive to Fas Two tumors PN1 andMB2 otherwise demonstrating relatively consistentstaining for Fas contained populations of apoptotic cellswith hyperchromatic condensed nuclei that did notexpress Fas PN2 contained a large cell population sug-gestive of gliomatous differentiation that expressed Fasmore consistently and prominently than did small cellswithin the tumor Against a weak background of faintlystaining tumor cells MB3 demonstrated intense stainingon large cells with prominent nuclei Occasional Fas-pos-itive Purkinje cells were seen within the tumor tissue
Glioblastoma Of the 4 glioblastomas GBM1 (Fig 1)and GBM4 demonstrated relatively intense staining forFas on more than 50 of cells and GBM2 and GBM3demonstrated more moderate staining on 25 to 50of cells Neurons in the tissue surrounding GBM3 alsoappeared positive for Fas expression
Flow cytometry Fas expression was assessed by owcytometric analysis Fluorescence proles were used tocalculate the MFR and the percentage of cells expressingFas (Fig 2) The well-characterized Jurkat lymphoma cellline was included as a positive control for constitutivereceptor expression Almost all Jurkat cells were found to
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 233
Table 1 Summary of ex vivo pediatric brain tumor specimens
Histology of Patient Tumor No tumor specimen age (yr) Sex
PN1 Pineoblastoma (PNET) 17 F
PN2 PNET 4 M
PN3 PNET 15 F
MB1 Medulloblastoma 8 F
MB2 Medulloblastoma 6 M
MB3 Medulloblastoma 10 F
MB4 Medulloblastoma 8 F
MB5 Medulloblastoma 2 M
MB6 Medulloblastoma 3 F
GBM1 Glioblastoma multiforme 112 M
GBM2 Glioblastoma multiforme 14 M
GBM3 Glioblastoma multiforme 16 M
GBM4 Glioblastoma multiforme 912 F
LGG1 Ganglioglioma 4 F
LGG2 Pilocytic astrocytoma (WHO grade I) 16 M
LGG3 Low-grade astrocytoma (WHO grade II) 7 M
LGG4 Pilocytic astrocytoma (WHO grade I) 14 M
GCT1 Germ cell tumor (germinoma) 15 M
Abbreviation WHO World Health Organization
Table 2 Immunohistochemical and FACS detection of Fas in pediatric brain tumors
Immunohistochemistry FACS
Perinecrotic increase Tumor No Frequencya Intensity Homogeneityb in expressionb MFRc Positived
PN1 ++++ Very weak ndash ndash 165 374
PN2 +++ Moderate ndash + 279 466
PN3 ++++ Moderate-weak + ndash 374 742
MB1 ++++ Moderate ndash + 243 545
MB2 ++++ Weak ndash ndash 285 501
MB3 +++ Weak-moderate ndash + 194 311
MB4 + Weak + ndash 110 131
MB5 +++ Very weak + ndash 268 491
MB6 +++ Weak ndash + 226 230
GBM1 ++++ Moderate-intense ndash ndash 149 245
GBM2 +++ Weak-moderate ndash + 25 531
GBM3 +++ Moderate ndash + 136 967
GBM4 ++++ Intense-moderate + ndash 305 611
LGG1 +++ Intense ndash ndash 240 252
LGG2 + Weak ndash + 354 580
LGG3 +++ Moderate-intense + ndash 248 642
LGG4 +++ Weak ndash ndash 213 444
GCT1 +++ Intense ndash ndash 229 391
Jurkat ND 384 930
Abbreviation ND not done
a+ lt5 positive cells ++ 5-25 positive cells +++ 25-50 positive cells ++++ gt50 positive cells
b+ presence of staining characteristic ndash absence of staining characteristic
cRatio of the mean uorescence of cells labeled with the anti-Fas antibody DX2 compared with those labeled with an isotype control
dPercentage of cells determined to be positive for Fas expression A region M1 was dened at its lower limit by the intersection of the uorescence proles of isotype- and anti-Fas
(DX2)-labeled cells The percentage of Fas-positive cells was calculated as the difference between the percentage of isotype-labeled and DX2-labeled cells uorescing within this region
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
be positive and relatively homogeneous for Fas expres-sion Surface expression of Fas was detected in all 18 pri-mary brain tumors The level of Fas expression bothwith regard to the density per cell (MFR) and the per-centage of Fas-positive cells varied considerably acrossthe spectrum of tumors and within each tumor type(Table 2) There was a relationship between the percent-age of Fas-positive cells and MFR (r = 084) Howeverno real difference was observed between tumor subtypes(data not shown) The variation in Fas expression in indi-vidual tumor specimens was generally greater than thatin the Jurkat population This was exemplied by the
proles of two specimens GBM1 (Fig 1) and GBM3which demonstrated small proportions of cells uoresc-ing at levels 1 or 2 log higher than the peak uorescenceof the population
The level of Fas expression as measured both by thepercentage of Fas-positive cells (421 plusmn 181) and theMFR (237 plusmn 070) in tumor samples was considerablyless than that observed in the Jurkat cells There was apoor correlation between surface Fas expression meas-ured by ow cytometry and immunohistochemical posi-tivity suggesting that in some tumors a large proportionof Fas may be intracellular
FasL Expression
Immunohistochemistry Immunohistochemically FasL wasdetected in 17 of the 18 tumors (Table 3) There tended tobe more homogeneity both across tumor subtypes andwithin each individual tumor than for Fas expressionPNETmedulloblastoma specimens all demonstratedexpression of FasL on more than 50 of tumor cells at var-ious staining intensities Apoptotic cells in PNETmedul-loblastoma specimens that did not express Fas were simi-larly not immunoreactive for FasL
Western blotting Western blotting detected FasL in 10 ofthe 13 tumors examined (Table 3) Of the 7 PNETmedul-loblastomas examined 5 were positive for FasL expressionStaining intensity of positive specimens was compared withreference to the b-tubulin loading control (Fig 3) Intensityranged from very weak (MB4) to strong (MB5 and MB6)with MB1 and MB2 being intermediate PNET 2 andPNET3 specimens were negative for FasL expression Of theglioblastomas all expressed FasL with GBM4 having thehighest level of expression of all the tumors tested HoweverGBM1 and GBM3 showed relatively low levels of expres-sion Of the low-grade tumors LGG4 showed a low expres-sion level with the other two having no or barelydetectable FasL expression Many cross-reactivehighndashmolecular-weight bands were seen in most tumor sam-ples and this may account for the poor correlation betweenthe Western blot and immunohistochemical results
Sensitivity of tumor specimens to Fas-mediated apo-ptosis Cell viability was measured by ow cytometry at24 h and by a colorimetric assay for total viable cell num-ber at 24- and 48-h time points Additionally sub G1peak analysis of cells cultured for 24 h in the presence orabsence of apoptotic mediators provided a measure ofthe percentage of apoptotic cells present in each speci-men Cell viabilityapoptosis in the presence of etoposideor anti-Fas treatment was expressed as a percentage rel-ative to the number of viableapoptotic cells in theabsence of any treatment at the same time point
Cell death assays were rst established using theJurkat cell line with APO-1 antibody in conjunction withprotein A or Neuro-2AFasL supernatant uid A colori-metric assay for total viable cell number exposing Jurkatcells to various titrations of Neuro-2AFasL supernatantuid indicated supernatant uid diluted up to 110 wassufcient to kill 100 of Jurkat cells in culture (data notshown) The loss of viability was due to induction ofapoptosis as indicated by sub G1 peak analysis Additionof Neuro-2Aneo control supernatant uid not contain-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001234
Fig 1 Immunohistochemical analysis of Fas expression in glioblas-toma specimen GBM1 A Clusters of tumor cells surrounded by con-nective tissue as demo in a section stained with hematoxylin andeosin B A similar section of tumor incubated with nonimmune rab-bit serum is completely negative for staining C Fas is expressed ontumor cells in the specimen but the connective tissue is negative forFas Original magnication acute150
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

proteins with the death domains (Kischkel et al 1995)initiates an intracellular proteolytic cascade involvingcaspases (Boldin et al 1996 Medema et al 1997Muzio et al 1996 Scafdi et al 1998) that culminatesin cleavage of cellular substrates and apoptosis in sensi-tive cells (Trauth et al 1989 Yonehara et al 1989)
Fas-FasL interactions play important roles in elimi-nation of autoreactive immune cells (Davidson et al1998 Watanabe-Fukunaga et al 1992) activation-induced T-cell death (Singer et al 1994) and killing bycytotoxic T lymphocytes (Hanabuchi et al 1994) andnatural killer cells (Oshimi et al 1996) FasL has beenimplicated in maintaining immune privilege in the ante-rior chamber of the eye (Grifth et al 1995 1996) inthe mouse testis (Bellgrau et al 1995 Duke et al 1996Yagita et al 1996) and in the brain (Saas et al 1997)FasL is believed to deliver a death signal to the Fas-expressing immune cells inltrating these tissues result-ing in suppression of immune activation
Expression of Fas has been identied in glioma(Weller et al 1994) and medulloblastoma cell lines(Weller et al 1998) and in ex vivo astrocytic tumor spec-imens (Frei et al 1998 Tohma et al 1998 Tachibanaet al 1995) Expression of Fas mRNA in astrocytomasin vivo tends to correlate with a high degree of malig-nancy (World Health Organization grades III and IVTachibana et al 1995) However the Fas proteinappears to be predominantly expressed in associationwith large areas of necrosis within tumor tissue (Tohmaet al 1998) Furthermore both human glioma cell lines(Weller et al 1994 1995a 1995b) and ex vivo gliomacells (Frei et al 1998) have been shown to be suscepti-ble to apoptosis induced by FasL or anti-Fas antibodiesPreexposure of cell lines to various cytokines includingTNF-a and interferon-g (Weller et al 1994) andincreased production of endogenous TGF-b1 by gliomacells (Ashley et al 1998) were shown to enhance Fascytotoxicity This and the absence of Fas on normalbrain parenchyma (Leithauser et al 1993) led to inter-est in Fas as the target of a novel immunotherapeuticapproach against these tumors
Malignant gliomas are among a number of solidtumors including colon carcinoma (OrsquoConnell et al1996) hepatocellular carcinoma (Strand et al 1996)esophageal carcinoma (Gratas et al 1998) andmelanoma (Hahne et al 1996) that express FasL (Saaset al 1997 Weller et al 1998) Membrane-boundFasL on cell lines (Hahne et al 1996 Strand et al1996) or ex vivo cells (Saas et al 1997 OrsquoConnell etal 1996 Walker et al 1997) is capable of inducingcell death in Fas-sensitive target cells This is believed torepresent a mechanism by which tumors can counterat-tack tumor-inltrating lymphocytes and cytotoxic Tcells and thereby escape immune surveillance (OrsquoCon-nell et al 1996)
It is not known whether the previously publishedndings from adult malignant glioma can be generalizedto include other tumors of the CNS including pediatricmalignancies In this report Fas expression was identi-ed in all of the pediatric brain tumors tested FasL wascoexpressed in 9 of the 13 tumors tested by both West-ern blot and immunohistochemistry Despite the pres-
ence of the receptor on tumor cells a large degree ofresistance to Fas-mediated apoptosis was observed in allthe tumor specimens
Methods
Cell Lines
Neuro-2A murine neuroblastoma cell lines were pro-vided by Dr Adriano Fontana (University HospitalZurich Switzerland) Neuro-2AFasL cells have beenengineered to produce soluble FasL by stable transfectionwith the BCMGS expression plasmid subcloned withcDNA encoding murine FasL (Rensing-Ehl et al 1995)Murine and human FasL share 769 homology at anamino acid level and are functionally interchangeable(Nagata and Golstein 1995) Neuro-2Aneo cells trans-fected with an empty BCMGS vector served as a controlin experiments using the FasL-containing supernatantuid Cell lines were maintained in zinc option medium(GIBCO BRL Grand Island NY) supplemented with10 heat-inactivated fetal calf serum (Trace BiosciencesCastle Hill New South Wales Victoria Australia)
Dissociation of Brain Tumor Specimens
Primary pediatric tumor specimens that were surplus topathology were collected during surgical procedures atthe Royal Childrenrsquos Hospital Melbourne Victoria Aus-tralia after Ethics in Human Research Committeeapproval Due to ethical and logistical considerations itwas not always possible to obtain a truly representativesample of the tumor If immediate processing of tumorspecimens was not feasible specimens were stored at 4degCfor less than 12 h Tumor samples were maintained inzinc option medium10 fetal calf serum mechanicallydissociated and then triturated by passage through 18-to-25-gauge needles Any solid debris preventing passageof the cell suspension was removed and discarded Suc-cessful passage through a 25-gauge needle indicated asingle cell suspension had been obtained as conrmed bymicroscopic examination Red blood cells were lysed byincubation in ammonium chloride solution (015MNH4Cl 20 mM Tris pH 74) for 2 min at room temper-ature and tumor cells were resuspended in culturemedium After assessment of viable cell number by try-pan blue exclusion (Sigma St Louis Mo) the tumor sus-pension was diluted to obtain a nal concentration of 106
viable cellsml
Labeling Cells for Immunophenotyping
FACS analysis was performed on freshly obtained tumorcell suspensions Cells (106) were centrifuged (500g 5min 4degC) washed in FACS buffer (1 bovine serumalbumin 005 NaN3 in PBS pH 74) and incubatedwith an anti-Fas monoclonal antibody (DX2 mouseIgG1 PharMingen San Diego Calif 1 mgml) or an iso-type control antibody for 30 min on ice The cells werewashed with FACS buffer and incubated with a uores-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001230
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
cein isothiocyanatendashconjugated afnity-isolated sheepanti-mouse secondary antibody [F(ab)2 fragment](Amrad Pharmacia Melbourne Victoria Australiadiluted 140) After incubation on ice for 30 min thecells were washed in FACS buffer resuspended in 200 mlFACS xative (09 NaCl 2 glucose 2 formalde-hyde) and incubated at 4degC until analyzed Referencecells were kept unstained after the rst wash and resus-pended in FACS xative The Jurkat cell line served as apositive control for each experiment and where the vol-ume of tumor suspension allowed the experiment wasconducted in duplicate
FACS Analysis of Tumor Specimens
A total of 10000 events were collected on the FACScan(Becton Dickinson Immunocytometry Systems San JoseCalif) using CYCLOPSTM software (Cytomation IncFort Collins Colo) The laser was tuned at 488 nm withan output power of 15 mW Fluorescein isothio-cyanatendashemitted ldquogreenrdquo uorescence was collected via aDM560-nm225deg dichroic mirror (gt560-nm reection)and a DF53030-nm band pass lter Cellular debris wasexcluded from the analysis
The MFR was calculated as the ratio of the mean u-orescence of DX2 (anti-Fas)-labeled cells and that of cellslabeled with the isotype control antibody
To quantitate the percentage of tumor cells that werepositive for Fas expression we used WinMDI Version27 software (Scripps Research Institute La Jolla Calif)to directly compare the uorescence prole of DX2-stained cells with the prole of cells stained with the iso-type control The DX2- and isotype-stained cell proleswere overlaid on the same histogram A marker was setdened at its lower limit by the point of intersection ofthe 2 proles The percentage of cells considered positivefor Fas was calculated by subtracting the percentage ofisotype-labeled cells from the percentage of DX2-labeledcells uorescing within the region dened by the marker
Western Blotting
Thirteen frozen tumor samples were selected accordingto availability of tumor tissue and coarsely choppedbefore suspension in 5 volumes (wtvol) RIPA buffer (1sodium dodecyl sulfate 1 NP40 in PBS pH 74) withprotease inhibitor cocktail set 1 (Calbiochem La JollaCalif) Tumor samples were homogenized using a Poly-tron homogenizer and boiled in sample buffer before sep-arating the proteins on 10 sodium dodecylsulfatendashpolyacrylamide gels and transferring them toHybondTM-P membranes (Amersham Pharmacia BiotechBuckinghamshire UK) Supernatant uid from Neuro-2AFasL-expressing cells was used as a positive controlfor FasL expression Membranes were blocked for 1 h atroom temperature in 5 nonfat milkTBST (01Tween 20 125 mM NaCl and 25 mM Tris pH 74)Blots were then probed overnight at 4degC with rabbitpolyclonal anti-human FasL antibody (Q20 Santa CruzBiotechnology Santa Cruz Calif 1100) or anti-b2-microtubulin (PharMingen 1100) as a loading controlQ20 anti-FasL antibody was selected because of its min-
imal cross-reactivity when compared with NOK-1 andG247-4 (both PharMingen) Blots were then incubatedwith horseradish peroxidasendashconjugated secondary anti-bodies (1100000) followed by enhanced chemilumines-cence detection with Supersignalreg West Pico Chemilumi-nescent Substrate according to the manufacturerrsquosinstructions (Pierce Rockford Ill) The blots wereexposed to Kodak XR lm (Eastman Kodak RochesterNY) for 2 s to 20 min
Immunohistochemical Detection of Fas and FasL
Formalin-xed parafn-embedded blocks of each tumorwere obtained from the Department of AnatomicalPathology Royal Childrenrsquos Hospital Melbourne Victo-ria Australia Sections 4-mm thick were cut and placedon poly-L-lysinendashcoated slides (Sigma) The sections weredeparaffinized in xylene and rehydrated Antigenretrieval was performed by immersing the slides for 30min in 001 M sodium citrate buffer preheated to 95degC(Kawai et al 1994 Werner et al 1996) Endogenousperoxidase activity was quenched by incubating the sec-tions in 1 hydrogen peroxide for 20 min A blockingstep was performed for 2 h at room temperature withPBS containing 5 normal goat serum (Vector Labora-tories Inc Burlingame Calif) and 10 normal humanserum Sections were incubated overnight at 4degC withrabbit polyclonal anti-human FasL antibody (Q20 SantaCruz Biotechnology 1100) or anti-human Fas antibody(Ab-1 Calbiochem 140) After 3 washes in PBS the sec-tions were incubated with biotin-conjugated goat anti-rabbit antibody (PharMingen 1200) for 30 min at roomtemperature The reaction was visualized using the Vec-tastain Elite ABC kit (Vector Laboratories Inc) and a33 9 -diaminobenzidine substrate (Sigma) The slides werecounterstained in hematoxylin (Shandon Southern Prod-ucts Ltd RuncornCheshire UK) Formalin-fixedparafn-embedded sections of human liver and mousespleen served as positive controls for Fas and FasLrespectively Negative controls consisted of sections with-out primary or secondary antibody and those incubatedwith nonimmunized rabbit serum Antibodies and con-trol rabbit serum were diluted in 1 normal goat serumin PBS Representative sections on which immunohisto-chemistry was not performed were stained with hema-toxylin and eosin (Shandon Southern Products Ltd) foridentication and classication of the tumor
The level of Fas and FasL expression was determined byassessment of the percentage of immunoreactive cells in 10nonadjacent high-powered elds (original magnicationacute400) The average of the 10 elds was calculated andrecorded as either no expression lt5 cells positive 5 to25 cells positive 25 to 50 cells positive or gt50cells positive The staining intensity of tumor cells was ratedas weak moderate or intense
Preparation of Supernatant Fluid from Neuro-2A Cell Lines
Supernatant uid was collected from conuent 10-mlplates of Neuro-2AFasL and Neuro-2Aneo cells andcentrifuged (500g 15 min) to remove cells and debris
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 231
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
The cell-free supernatant uid was then used at the dilu-tions specied or stored at ndash20degC until used
Flow Cytometric Determination of Cell Viability by FACS Analysis
In a 12-well plate 1 ml tumor cells at a viable cell con-centration of 106ml was incubated in triplicate with eachmedium alone etoposide (10 mgml) and APO-1 anti-body (100 ngml) with protein A (110000) for 24 h(37degC 5 CO2 96 humidity) For specimens PN2and MB1 the combination of APO-1 and protein A wasreplaced with Neuro-2AFasL supernatant uid diluted12 and cells exposed to an equivalent dilution of Neuro-2Aneo supernatant uid served as a negative controlWells containing Jurkat cells at 106ml served as positivecontrols for each condition
At 24 h all cells were harvested from each well by vig-orous pipetting Adherent cells were removed by incu-bating with trypsin-EDTA Each well was washed 3 timeswith PBS and lack of residual cells was conrmed bymicroscopic examination The cells were centrifuged(500g 5 min 4degC) resuspended in 300 mL FACS buffercontaining 50 mgml propidium iodide (Sigma) and incu-bated on ice for 1 h
Sub G1 Peak Analysis of Apoptotic Cell Death
After assessing viability by ow cytometry we xed thecells for sub G1 peak analysis (Tounekti et al 1995) byadding 250 ml of 70 ethanol to achieve a nal concen-tration of approximately 35 After overnight incuba-tion at 4degC samples were centrifuged (500g 5 min 4degC)and resuspended in 200 ml propidium iodide at 50 mgmlin PBS
Between 10000 and 20000 events were collected foranalysis on the FACScan using settings described foranalysis of viability Of the events remaining after dou-blet discrimination cells uorescing below 25 of theG1 peak uorescence were assumed to be debris and wereexcluded from analysis Cells uorescing above this limitbut below the intensity of the G1 peak were designatedapoptotic and the percentage of total tumor cells theyrepresented was calculated
Colorimetric Assay for Determining Total Viable Cell Number
Cell viability across a 48-h period was measured usingthe CellTiter 96regAQueous Non-Radioactive Cell Prolifera-tion Assay (Promega Madison Wisc) which detectsmitochondrial activity according to the recommendedprotocol In a 96-well plate 50 ml tumor cell suspensionat 4 acute 105ml was incubated with etoposide (10 mgml)or anti-Fas antibody APO-1 (100 ngml) with protein A(110000) or in the absence of inducers of apoptosis ina nal volume of 100 ml In later experiments (specimensMB1 and PN2) the combination of APO-1 and proteinA was replaced with Neuro-2AFasL supernatant uiddiluted 12 with parallel wells containing Neuro-2Aneosupernatant uid diluted 12 serving as a negative con-trol The experiment was conducted in triplicate and
incubated over a 48-h period (37degC 5 CO2 96humidity) Jurkat cells at a nal concentration of 2 acute105ml were used as a positive control for each treatmentWells containing 100 ml medium and substrate provideda measurement of the background reading at 490 nmresulting from absorbance within the medium at thiswavelength
Substrate (20 mlwell) was added at 0- 24- and 48-htime points after the assay was established and after afurther 3-h incubation the absorbance at 490 nm wasmeasured with an ELISA plate reader (reference wave-length 620 nm) Absorbance readings were adjusted toremove the contribution of background absorbance andviability was calculated at each time point as a percent-age of the average absorbance from wells containing noapoptosis inducers at the same time point
Statistical Analysis
We determined the signicant differences between treat-ments in individual tumor specimens by using Studentrsquos ttest (Altman 1991) We compared the overall differencein viability in the anti-Fas treatment and control groupsin the medulloblastomaPNET specimens using two-wayANOVA on Stata Corporation 1997 (Stata StatisticalSoftware Release 50 College Station Tex)
Results
Tumor Samples
The 18 pediatric brain tumor specimens analyzed forexpression of Fas and FasL are described in Table 1according to histology patient age and sex The speci-mens consisted of 9 PNETs that included 6 medulloblas-tomas 4 glioblastomas multiforme 3 low-grade gliomasa ganglioglioma and a germ-cell tumor
Fas Expression
Immunohistochemistry Giemsa staining of tumor sec-tions revealed morphologic differences between tumorcells as well as necrotic and apoptotic tissue Small num-bers of normal brain cells epidermal cells and bloodcells were also present in the samples Immunohisto-chemically the Fas receptor was detected in all 18 tumorspecimens (Table 2) The cytoplasm and membrane ofpositive tumor cells stained diffusely Walls of blood ves-sels red blood cells large areas of necrotic tissue andareas of desmoplasia were negative and staining wasconsistently absent in negative controls without primaryor secondary antibody
Tumor sections with little variation in cellular mor-phology demonstrated uniform intensity of Fas stainingthroughout In perinecrotic areas of tumor tissue anapparent increase in the number and staining intensity ofFas-positive tumor cells was frequently observed Tumorswere rated according to the percentage of tumor cellsexpressing the Fas receptor and the staining intensity ofthese cells (Table 2)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001232
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
PNETmedulloblastoma Weakly Fas-positive cellswere found to constitute 25 to 50 of cells in 4 of the9 PNETmedulloblastoma specimens and more than50 of cells in another 4 tumors of this group In theremaining medulloblastoma only occasional cellsappeared immunoreactive to Fas Two tumors PN1 andMB2 otherwise demonstrating relatively consistentstaining for Fas contained populations of apoptotic cellswith hyperchromatic condensed nuclei that did notexpress Fas PN2 contained a large cell population sug-gestive of gliomatous differentiation that expressed Fasmore consistently and prominently than did small cellswithin the tumor Against a weak background of faintlystaining tumor cells MB3 demonstrated intense stainingon large cells with prominent nuclei Occasional Fas-pos-itive Purkinje cells were seen within the tumor tissue
Glioblastoma Of the 4 glioblastomas GBM1 (Fig 1)and GBM4 demonstrated relatively intense staining forFas on more than 50 of cells and GBM2 and GBM3demonstrated more moderate staining on 25 to 50of cells Neurons in the tissue surrounding GBM3 alsoappeared positive for Fas expression
Flow cytometry Fas expression was assessed by owcytometric analysis Fluorescence proles were used tocalculate the MFR and the percentage of cells expressingFas (Fig 2) The well-characterized Jurkat lymphoma cellline was included as a positive control for constitutivereceptor expression Almost all Jurkat cells were found to
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 233
Table 1 Summary of ex vivo pediatric brain tumor specimens
Histology of Patient Tumor No tumor specimen age (yr) Sex
PN1 Pineoblastoma (PNET) 17 F
PN2 PNET 4 M
PN3 PNET 15 F
MB1 Medulloblastoma 8 F
MB2 Medulloblastoma 6 M
MB3 Medulloblastoma 10 F
MB4 Medulloblastoma 8 F
MB5 Medulloblastoma 2 M
MB6 Medulloblastoma 3 F
GBM1 Glioblastoma multiforme 112 M
GBM2 Glioblastoma multiforme 14 M
GBM3 Glioblastoma multiforme 16 M
GBM4 Glioblastoma multiforme 912 F
LGG1 Ganglioglioma 4 F
LGG2 Pilocytic astrocytoma (WHO grade I) 16 M
LGG3 Low-grade astrocytoma (WHO grade II) 7 M
LGG4 Pilocytic astrocytoma (WHO grade I) 14 M
GCT1 Germ cell tumor (germinoma) 15 M
Abbreviation WHO World Health Organization
Table 2 Immunohistochemical and FACS detection of Fas in pediatric brain tumors
Immunohistochemistry FACS
Perinecrotic increase Tumor No Frequencya Intensity Homogeneityb in expressionb MFRc Positived
PN1 ++++ Very weak ndash ndash 165 374
PN2 +++ Moderate ndash + 279 466
PN3 ++++ Moderate-weak + ndash 374 742
MB1 ++++ Moderate ndash + 243 545
MB2 ++++ Weak ndash ndash 285 501
MB3 +++ Weak-moderate ndash + 194 311
MB4 + Weak + ndash 110 131
MB5 +++ Very weak + ndash 268 491
MB6 +++ Weak ndash + 226 230
GBM1 ++++ Moderate-intense ndash ndash 149 245
GBM2 +++ Weak-moderate ndash + 25 531
GBM3 +++ Moderate ndash + 136 967
GBM4 ++++ Intense-moderate + ndash 305 611
LGG1 +++ Intense ndash ndash 240 252
LGG2 + Weak ndash + 354 580
LGG3 +++ Moderate-intense + ndash 248 642
LGG4 +++ Weak ndash ndash 213 444
GCT1 +++ Intense ndash ndash 229 391
Jurkat ND 384 930
Abbreviation ND not done
a+ lt5 positive cells ++ 5-25 positive cells +++ 25-50 positive cells ++++ gt50 positive cells
b+ presence of staining characteristic ndash absence of staining characteristic
cRatio of the mean uorescence of cells labeled with the anti-Fas antibody DX2 compared with those labeled with an isotype control
dPercentage of cells determined to be positive for Fas expression A region M1 was dened at its lower limit by the intersection of the uorescence proles of isotype- and anti-Fas
(DX2)-labeled cells The percentage of Fas-positive cells was calculated as the difference between the percentage of isotype-labeled and DX2-labeled cells uorescing within this region
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
be positive and relatively homogeneous for Fas expres-sion Surface expression of Fas was detected in all 18 pri-mary brain tumors The level of Fas expression bothwith regard to the density per cell (MFR) and the per-centage of Fas-positive cells varied considerably acrossthe spectrum of tumors and within each tumor type(Table 2) There was a relationship between the percent-age of Fas-positive cells and MFR (r = 084) Howeverno real difference was observed between tumor subtypes(data not shown) The variation in Fas expression in indi-vidual tumor specimens was generally greater than thatin the Jurkat population This was exemplied by the
proles of two specimens GBM1 (Fig 1) and GBM3which demonstrated small proportions of cells uoresc-ing at levels 1 or 2 log higher than the peak uorescenceof the population
The level of Fas expression as measured both by thepercentage of Fas-positive cells (421 plusmn 181) and theMFR (237 plusmn 070) in tumor samples was considerablyless than that observed in the Jurkat cells There was apoor correlation between surface Fas expression meas-ured by ow cytometry and immunohistochemical posi-tivity suggesting that in some tumors a large proportionof Fas may be intracellular
FasL Expression
Immunohistochemistry Immunohistochemically FasL wasdetected in 17 of the 18 tumors (Table 3) There tended tobe more homogeneity both across tumor subtypes andwithin each individual tumor than for Fas expressionPNETmedulloblastoma specimens all demonstratedexpression of FasL on more than 50 of tumor cells at var-ious staining intensities Apoptotic cells in PNETmedul-loblastoma specimens that did not express Fas were simi-larly not immunoreactive for FasL
Western blotting Western blotting detected FasL in 10 ofthe 13 tumors examined (Table 3) Of the 7 PNETmedul-loblastomas examined 5 were positive for FasL expressionStaining intensity of positive specimens was compared withreference to the b-tubulin loading control (Fig 3) Intensityranged from very weak (MB4) to strong (MB5 and MB6)with MB1 and MB2 being intermediate PNET 2 andPNET3 specimens were negative for FasL expression Of theglioblastomas all expressed FasL with GBM4 having thehighest level of expression of all the tumors tested HoweverGBM1 and GBM3 showed relatively low levels of expres-sion Of the low-grade tumors LGG4 showed a low expres-sion level with the other two having no or barelydetectable FasL expression Many cross-reactivehighndashmolecular-weight bands were seen in most tumor sam-ples and this may account for the poor correlation betweenthe Western blot and immunohistochemical results
Sensitivity of tumor specimens to Fas-mediated apo-ptosis Cell viability was measured by ow cytometry at24 h and by a colorimetric assay for total viable cell num-ber at 24- and 48-h time points Additionally sub G1peak analysis of cells cultured for 24 h in the presence orabsence of apoptotic mediators provided a measure ofthe percentage of apoptotic cells present in each speci-men Cell viabilityapoptosis in the presence of etoposideor anti-Fas treatment was expressed as a percentage rel-ative to the number of viableapoptotic cells in theabsence of any treatment at the same time point
Cell death assays were rst established using theJurkat cell line with APO-1 antibody in conjunction withprotein A or Neuro-2AFasL supernatant uid A colori-metric assay for total viable cell number exposing Jurkatcells to various titrations of Neuro-2AFasL supernatantuid indicated supernatant uid diluted up to 110 wassufcient to kill 100 of Jurkat cells in culture (data notshown) The loss of viability was due to induction ofapoptosis as indicated by sub G1 peak analysis Additionof Neuro-2Aneo control supernatant uid not contain-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001234
Fig 1 Immunohistochemical analysis of Fas expression in glioblas-toma specimen GBM1 A Clusters of tumor cells surrounded by con-nective tissue as demo in a section stained with hematoxylin andeosin B A similar section of tumor incubated with nonimmune rab-bit serum is completely negative for staining C Fas is expressed ontumor cells in the specimen but the connective tissue is negative forFas Original magnication acute150
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

cein isothiocyanatendashconjugated afnity-isolated sheepanti-mouse secondary antibody [F(ab)2 fragment](Amrad Pharmacia Melbourne Victoria Australiadiluted 140) After incubation on ice for 30 min thecells were washed in FACS buffer resuspended in 200 mlFACS xative (09 NaCl 2 glucose 2 formalde-hyde) and incubated at 4degC until analyzed Referencecells were kept unstained after the rst wash and resus-pended in FACS xative The Jurkat cell line served as apositive control for each experiment and where the vol-ume of tumor suspension allowed the experiment wasconducted in duplicate
FACS Analysis of Tumor Specimens
A total of 10000 events were collected on the FACScan(Becton Dickinson Immunocytometry Systems San JoseCalif) using CYCLOPSTM software (Cytomation IncFort Collins Colo) The laser was tuned at 488 nm withan output power of 15 mW Fluorescein isothio-cyanatendashemitted ldquogreenrdquo uorescence was collected via aDM560-nm225deg dichroic mirror (gt560-nm reection)and a DF53030-nm band pass lter Cellular debris wasexcluded from the analysis
The MFR was calculated as the ratio of the mean u-orescence of DX2 (anti-Fas)-labeled cells and that of cellslabeled with the isotype control antibody
To quantitate the percentage of tumor cells that werepositive for Fas expression we used WinMDI Version27 software (Scripps Research Institute La Jolla Calif)to directly compare the uorescence prole of DX2-stained cells with the prole of cells stained with the iso-type control The DX2- and isotype-stained cell proleswere overlaid on the same histogram A marker was setdened at its lower limit by the point of intersection ofthe 2 proles The percentage of cells considered positivefor Fas was calculated by subtracting the percentage ofisotype-labeled cells from the percentage of DX2-labeledcells uorescing within the region dened by the marker
Western Blotting
Thirteen frozen tumor samples were selected accordingto availability of tumor tissue and coarsely choppedbefore suspension in 5 volumes (wtvol) RIPA buffer (1sodium dodecyl sulfate 1 NP40 in PBS pH 74) withprotease inhibitor cocktail set 1 (Calbiochem La JollaCalif) Tumor samples were homogenized using a Poly-tron homogenizer and boiled in sample buffer before sep-arating the proteins on 10 sodium dodecylsulfatendashpolyacrylamide gels and transferring them toHybondTM-P membranes (Amersham Pharmacia BiotechBuckinghamshire UK) Supernatant uid from Neuro-2AFasL-expressing cells was used as a positive controlfor FasL expression Membranes were blocked for 1 h atroom temperature in 5 nonfat milkTBST (01Tween 20 125 mM NaCl and 25 mM Tris pH 74)Blots were then probed overnight at 4degC with rabbitpolyclonal anti-human FasL antibody (Q20 Santa CruzBiotechnology Santa Cruz Calif 1100) or anti-b2-microtubulin (PharMingen 1100) as a loading controlQ20 anti-FasL antibody was selected because of its min-
imal cross-reactivity when compared with NOK-1 andG247-4 (both PharMingen) Blots were then incubatedwith horseradish peroxidasendashconjugated secondary anti-bodies (1100000) followed by enhanced chemilumines-cence detection with Supersignalreg West Pico Chemilumi-nescent Substrate according to the manufacturerrsquosinstructions (Pierce Rockford Ill) The blots wereexposed to Kodak XR lm (Eastman Kodak RochesterNY) for 2 s to 20 min
Immunohistochemical Detection of Fas and FasL
Formalin-xed parafn-embedded blocks of each tumorwere obtained from the Department of AnatomicalPathology Royal Childrenrsquos Hospital Melbourne Victo-ria Australia Sections 4-mm thick were cut and placedon poly-L-lysinendashcoated slides (Sigma) The sections weredeparaffinized in xylene and rehydrated Antigenretrieval was performed by immersing the slides for 30min in 001 M sodium citrate buffer preheated to 95degC(Kawai et al 1994 Werner et al 1996) Endogenousperoxidase activity was quenched by incubating the sec-tions in 1 hydrogen peroxide for 20 min A blockingstep was performed for 2 h at room temperature withPBS containing 5 normal goat serum (Vector Labora-tories Inc Burlingame Calif) and 10 normal humanserum Sections were incubated overnight at 4degC withrabbit polyclonal anti-human FasL antibody (Q20 SantaCruz Biotechnology 1100) or anti-human Fas antibody(Ab-1 Calbiochem 140) After 3 washes in PBS the sec-tions were incubated with biotin-conjugated goat anti-rabbit antibody (PharMingen 1200) for 30 min at roomtemperature The reaction was visualized using the Vec-tastain Elite ABC kit (Vector Laboratories Inc) and a33 9 -diaminobenzidine substrate (Sigma) The slides werecounterstained in hematoxylin (Shandon Southern Prod-ucts Ltd RuncornCheshire UK) Formalin-fixedparafn-embedded sections of human liver and mousespleen served as positive controls for Fas and FasLrespectively Negative controls consisted of sections with-out primary or secondary antibody and those incubatedwith nonimmunized rabbit serum Antibodies and con-trol rabbit serum were diluted in 1 normal goat serumin PBS Representative sections on which immunohisto-chemistry was not performed were stained with hema-toxylin and eosin (Shandon Southern Products Ltd) foridentication and classication of the tumor
The level of Fas and FasL expression was determined byassessment of the percentage of immunoreactive cells in 10nonadjacent high-powered elds (original magnicationacute400) The average of the 10 elds was calculated andrecorded as either no expression lt5 cells positive 5 to25 cells positive 25 to 50 cells positive or gt50cells positive The staining intensity of tumor cells was ratedas weak moderate or intense
Preparation of Supernatant Fluid from Neuro-2A Cell Lines
Supernatant uid was collected from conuent 10-mlplates of Neuro-2AFasL and Neuro-2Aneo cells andcentrifuged (500g 15 min) to remove cells and debris
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 231
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
The cell-free supernatant uid was then used at the dilu-tions specied or stored at ndash20degC until used
Flow Cytometric Determination of Cell Viability by FACS Analysis
In a 12-well plate 1 ml tumor cells at a viable cell con-centration of 106ml was incubated in triplicate with eachmedium alone etoposide (10 mgml) and APO-1 anti-body (100 ngml) with protein A (110000) for 24 h(37degC 5 CO2 96 humidity) For specimens PN2and MB1 the combination of APO-1 and protein A wasreplaced with Neuro-2AFasL supernatant uid diluted12 and cells exposed to an equivalent dilution of Neuro-2Aneo supernatant uid served as a negative controlWells containing Jurkat cells at 106ml served as positivecontrols for each condition
At 24 h all cells were harvested from each well by vig-orous pipetting Adherent cells were removed by incu-bating with trypsin-EDTA Each well was washed 3 timeswith PBS and lack of residual cells was conrmed bymicroscopic examination The cells were centrifuged(500g 5 min 4degC) resuspended in 300 mL FACS buffercontaining 50 mgml propidium iodide (Sigma) and incu-bated on ice for 1 h
Sub G1 Peak Analysis of Apoptotic Cell Death
After assessing viability by ow cytometry we xed thecells for sub G1 peak analysis (Tounekti et al 1995) byadding 250 ml of 70 ethanol to achieve a nal concen-tration of approximately 35 After overnight incuba-tion at 4degC samples were centrifuged (500g 5 min 4degC)and resuspended in 200 ml propidium iodide at 50 mgmlin PBS
Between 10000 and 20000 events were collected foranalysis on the FACScan using settings described foranalysis of viability Of the events remaining after dou-blet discrimination cells uorescing below 25 of theG1 peak uorescence were assumed to be debris and wereexcluded from analysis Cells uorescing above this limitbut below the intensity of the G1 peak were designatedapoptotic and the percentage of total tumor cells theyrepresented was calculated
Colorimetric Assay for Determining Total Viable Cell Number
Cell viability across a 48-h period was measured usingthe CellTiter 96regAQueous Non-Radioactive Cell Prolifera-tion Assay (Promega Madison Wisc) which detectsmitochondrial activity according to the recommendedprotocol In a 96-well plate 50 ml tumor cell suspensionat 4 acute 105ml was incubated with etoposide (10 mgml)or anti-Fas antibody APO-1 (100 ngml) with protein A(110000) or in the absence of inducers of apoptosis ina nal volume of 100 ml In later experiments (specimensMB1 and PN2) the combination of APO-1 and proteinA was replaced with Neuro-2AFasL supernatant uiddiluted 12 with parallel wells containing Neuro-2Aneosupernatant uid diluted 12 serving as a negative con-trol The experiment was conducted in triplicate and
incubated over a 48-h period (37degC 5 CO2 96humidity) Jurkat cells at a nal concentration of 2 acute105ml were used as a positive control for each treatmentWells containing 100 ml medium and substrate provideda measurement of the background reading at 490 nmresulting from absorbance within the medium at thiswavelength
Substrate (20 mlwell) was added at 0- 24- and 48-htime points after the assay was established and after afurther 3-h incubation the absorbance at 490 nm wasmeasured with an ELISA plate reader (reference wave-length 620 nm) Absorbance readings were adjusted toremove the contribution of background absorbance andviability was calculated at each time point as a percent-age of the average absorbance from wells containing noapoptosis inducers at the same time point
Statistical Analysis
We determined the signicant differences between treat-ments in individual tumor specimens by using Studentrsquos ttest (Altman 1991) We compared the overall differencein viability in the anti-Fas treatment and control groupsin the medulloblastomaPNET specimens using two-wayANOVA on Stata Corporation 1997 (Stata StatisticalSoftware Release 50 College Station Tex)
Results
Tumor Samples
The 18 pediatric brain tumor specimens analyzed forexpression of Fas and FasL are described in Table 1according to histology patient age and sex The speci-mens consisted of 9 PNETs that included 6 medulloblas-tomas 4 glioblastomas multiforme 3 low-grade gliomasa ganglioglioma and a germ-cell tumor
Fas Expression
Immunohistochemistry Giemsa staining of tumor sec-tions revealed morphologic differences between tumorcells as well as necrotic and apoptotic tissue Small num-bers of normal brain cells epidermal cells and bloodcells were also present in the samples Immunohisto-chemically the Fas receptor was detected in all 18 tumorspecimens (Table 2) The cytoplasm and membrane ofpositive tumor cells stained diffusely Walls of blood ves-sels red blood cells large areas of necrotic tissue andareas of desmoplasia were negative and staining wasconsistently absent in negative controls without primaryor secondary antibody
Tumor sections with little variation in cellular mor-phology demonstrated uniform intensity of Fas stainingthroughout In perinecrotic areas of tumor tissue anapparent increase in the number and staining intensity ofFas-positive tumor cells was frequently observed Tumorswere rated according to the percentage of tumor cellsexpressing the Fas receptor and the staining intensity ofthese cells (Table 2)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001232
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
PNETmedulloblastoma Weakly Fas-positive cellswere found to constitute 25 to 50 of cells in 4 of the9 PNETmedulloblastoma specimens and more than50 of cells in another 4 tumors of this group In theremaining medulloblastoma only occasional cellsappeared immunoreactive to Fas Two tumors PN1 andMB2 otherwise demonstrating relatively consistentstaining for Fas contained populations of apoptotic cellswith hyperchromatic condensed nuclei that did notexpress Fas PN2 contained a large cell population sug-gestive of gliomatous differentiation that expressed Fasmore consistently and prominently than did small cellswithin the tumor Against a weak background of faintlystaining tumor cells MB3 demonstrated intense stainingon large cells with prominent nuclei Occasional Fas-pos-itive Purkinje cells were seen within the tumor tissue
Glioblastoma Of the 4 glioblastomas GBM1 (Fig 1)and GBM4 demonstrated relatively intense staining forFas on more than 50 of cells and GBM2 and GBM3demonstrated more moderate staining on 25 to 50of cells Neurons in the tissue surrounding GBM3 alsoappeared positive for Fas expression
Flow cytometry Fas expression was assessed by owcytometric analysis Fluorescence proles were used tocalculate the MFR and the percentage of cells expressingFas (Fig 2) The well-characterized Jurkat lymphoma cellline was included as a positive control for constitutivereceptor expression Almost all Jurkat cells were found to
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 233
Table 1 Summary of ex vivo pediatric brain tumor specimens
Histology of Patient Tumor No tumor specimen age (yr) Sex
PN1 Pineoblastoma (PNET) 17 F
PN2 PNET 4 M
PN3 PNET 15 F
MB1 Medulloblastoma 8 F
MB2 Medulloblastoma 6 M
MB3 Medulloblastoma 10 F
MB4 Medulloblastoma 8 F
MB5 Medulloblastoma 2 M
MB6 Medulloblastoma 3 F
GBM1 Glioblastoma multiforme 112 M
GBM2 Glioblastoma multiforme 14 M
GBM3 Glioblastoma multiforme 16 M
GBM4 Glioblastoma multiforme 912 F
LGG1 Ganglioglioma 4 F
LGG2 Pilocytic astrocytoma (WHO grade I) 16 M
LGG3 Low-grade astrocytoma (WHO grade II) 7 M
LGG4 Pilocytic astrocytoma (WHO grade I) 14 M
GCT1 Germ cell tumor (germinoma) 15 M
Abbreviation WHO World Health Organization
Table 2 Immunohistochemical and FACS detection of Fas in pediatric brain tumors
Immunohistochemistry FACS
Perinecrotic increase Tumor No Frequencya Intensity Homogeneityb in expressionb MFRc Positived
PN1 ++++ Very weak ndash ndash 165 374
PN2 +++ Moderate ndash + 279 466
PN3 ++++ Moderate-weak + ndash 374 742
MB1 ++++ Moderate ndash + 243 545
MB2 ++++ Weak ndash ndash 285 501
MB3 +++ Weak-moderate ndash + 194 311
MB4 + Weak + ndash 110 131
MB5 +++ Very weak + ndash 268 491
MB6 +++ Weak ndash + 226 230
GBM1 ++++ Moderate-intense ndash ndash 149 245
GBM2 +++ Weak-moderate ndash + 25 531
GBM3 +++ Moderate ndash + 136 967
GBM4 ++++ Intense-moderate + ndash 305 611
LGG1 +++ Intense ndash ndash 240 252
LGG2 + Weak ndash + 354 580
LGG3 +++ Moderate-intense + ndash 248 642
LGG4 +++ Weak ndash ndash 213 444
GCT1 +++ Intense ndash ndash 229 391
Jurkat ND 384 930
Abbreviation ND not done
a+ lt5 positive cells ++ 5-25 positive cells +++ 25-50 positive cells ++++ gt50 positive cells
b+ presence of staining characteristic ndash absence of staining characteristic
cRatio of the mean uorescence of cells labeled with the anti-Fas antibody DX2 compared with those labeled with an isotype control
dPercentage of cells determined to be positive for Fas expression A region M1 was dened at its lower limit by the intersection of the uorescence proles of isotype- and anti-Fas
(DX2)-labeled cells The percentage of Fas-positive cells was calculated as the difference between the percentage of isotype-labeled and DX2-labeled cells uorescing within this region
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
be positive and relatively homogeneous for Fas expres-sion Surface expression of Fas was detected in all 18 pri-mary brain tumors The level of Fas expression bothwith regard to the density per cell (MFR) and the per-centage of Fas-positive cells varied considerably acrossthe spectrum of tumors and within each tumor type(Table 2) There was a relationship between the percent-age of Fas-positive cells and MFR (r = 084) Howeverno real difference was observed between tumor subtypes(data not shown) The variation in Fas expression in indi-vidual tumor specimens was generally greater than thatin the Jurkat population This was exemplied by the
proles of two specimens GBM1 (Fig 1) and GBM3which demonstrated small proportions of cells uoresc-ing at levels 1 or 2 log higher than the peak uorescenceof the population
The level of Fas expression as measured both by thepercentage of Fas-positive cells (421 plusmn 181) and theMFR (237 plusmn 070) in tumor samples was considerablyless than that observed in the Jurkat cells There was apoor correlation between surface Fas expression meas-ured by ow cytometry and immunohistochemical posi-tivity suggesting that in some tumors a large proportionof Fas may be intracellular
FasL Expression
Immunohistochemistry Immunohistochemically FasL wasdetected in 17 of the 18 tumors (Table 3) There tended tobe more homogeneity both across tumor subtypes andwithin each individual tumor than for Fas expressionPNETmedulloblastoma specimens all demonstratedexpression of FasL on more than 50 of tumor cells at var-ious staining intensities Apoptotic cells in PNETmedul-loblastoma specimens that did not express Fas were simi-larly not immunoreactive for FasL
Western blotting Western blotting detected FasL in 10 ofthe 13 tumors examined (Table 3) Of the 7 PNETmedul-loblastomas examined 5 were positive for FasL expressionStaining intensity of positive specimens was compared withreference to the b-tubulin loading control (Fig 3) Intensityranged from very weak (MB4) to strong (MB5 and MB6)with MB1 and MB2 being intermediate PNET 2 andPNET3 specimens were negative for FasL expression Of theglioblastomas all expressed FasL with GBM4 having thehighest level of expression of all the tumors tested HoweverGBM1 and GBM3 showed relatively low levels of expres-sion Of the low-grade tumors LGG4 showed a low expres-sion level with the other two having no or barelydetectable FasL expression Many cross-reactivehighndashmolecular-weight bands were seen in most tumor sam-ples and this may account for the poor correlation betweenthe Western blot and immunohistochemical results
Sensitivity of tumor specimens to Fas-mediated apo-ptosis Cell viability was measured by ow cytometry at24 h and by a colorimetric assay for total viable cell num-ber at 24- and 48-h time points Additionally sub G1peak analysis of cells cultured for 24 h in the presence orabsence of apoptotic mediators provided a measure ofthe percentage of apoptotic cells present in each speci-men Cell viabilityapoptosis in the presence of etoposideor anti-Fas treatment was expressed as a percentage rel-ative to the number of viableapoptotic cells in theabsence of any treatment at the same time point
Cell death assays were rst established using theJurkat cell line with APO-1 antibody in conjunction withprotein A or Neuro-2AFasL supernatant uid A colori-metric assay for total viable cell number exposing Jurkatcells to various titrations of Neuro-2AFasL supernatantuid indicated supernatant uid diluted up to 110 wassufcient to kill 100 of Jurkat cells in culture (data notshown) The loss of viability was due to induction ofapoptosis as indicated by sub G1 peak analysis Additionof Neuro-2Aneo control supernatant uid not contain-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001234
Fig 1 Immunohistochemical analysis of Fas expression in glioblas-toma specimen GBM1 A Clusters of tumor cells surrounded by con-nective tissue as demo in a section stained with hematoxylin andeosin B A similar section of tumor incubated with nonimmune rab-bit serum is completely negative for staining C Fas is expressed ontumor cells in the specimen but the connective tissue is negative forFas Original magnication acute150
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

The cell-free supernatant uid was then used at the dilu-tions specied or stored at ndash20degC until used
Flow Cytometric Determination of Cell Viability by FACS Analysis
In a 12-well plate 1 ml tumor cells at a viable cell con-centration of 106ml was incubated in triplicate with eachmedium alone etoposide (10 mgml) and APO-1 anti-body (100 ngml) with protein A (110000) for 24 h(37degC 5 CO2 96 humidity) For specimens PN2and MB1 the combination of APO-1 and protein A wasreplaced with Neuro-2AFasL supernatant uid diluted12 and cells exposed to an equivalent dilution of Neuro-2Aneo supernatant uid served as a negative controlWells containing Jurkat cells at 106ml served as positivecontrols for each condition
At 24 h all cells were harvested from each well by vig-orous pipetting Adherent cells were removed by incu-bating with trypsin-EDTA Each well was washed 3 timeswith PBS and lack of residual cells was conrmed bymicroscopic examination The cells were centrifuged(500g 5 min 4degC) resuspended in 300 mL FACS buffercontaining 50 mgml propidium iodide (Sigma) and incu-bated on ice for 1 h
Sub G1 Peak Analysis of Apoptotic Cell Death
After assessing viability by ow cytometry we xed thecells for sub G1 peak analysis (Tounekti et al 1995) byadding 250 ml of 70 ethanol to achieve a nal concen-tration of approximately 35 After overnight incuba-tion at 4degC samples were centrifuged (500g 5 min 4degC)and resuspended in 200 ml propidium iodide at 50 mgmlin PBS
Between 10000 and 20000 events were collected foranalysis on the FACScan using settings described foranalysis of viability Of the events remaining after dou-blet discrimination cells uorescing below 25 of theG1 peak uorescence were assumed to be debris and wereexcluded from analysis Cells uorescing above this limitbut below the intensity of the G1 peak were designatedapoptotic and the percentage of total tumor cells theyrepresented was calculated
Colorimetric Assay for Determining Total Viable Cell Number
Cell viability across a 48-h period was measured usingthe CellTiter 96regAQueous Non-Radioactive Cell Prolifera-tion Assay (Promega Madison Wisc) which detectsmitochondrial activity according to the recommendedprotocol In a 96-well plate 50 ml tumor cell suspensionat 4 acute 105ml was incubated with etoposide (10 mgml)or anti-Fas antibody APO-1 (100 ngml) with protein A(110000) or in the absence of inducers of apoptosis ina nal volume of 100 ml In later experiments (specimensMB1 and PN2) the combination of APO-1 and proteinA was replaced with Neuro-2AFasL supernatant uiddiluted 12 with parallel wells containing Neuro-2Aneosupernatant uid diluted 12 serving as a negative con-trol The experiment was conducted in triplicate and
incubated over a 48-h period (37degC 5 CO2 96humidity) Jurkat cells at a nal concentration of 2 acute105ml were used as a positive control for each treatmentWells containing 100 ml medium and substrate provideda measurement of the background reading at 490 nmresulting from absorbance within the medium at thiswavelength
Substrate (20 mlwell) was added at 0- 24- and 48-htime points after the assay was established and after afurther 3-h incubation the absorbance at 490 nm wasmeasured with an ELISA plate reader (reference wave-length 620 nm) Absorbance readings were adjusted toremove the contribution of background absorbance andviability was calculated at each time point as a percent-age of the average absorbance from wells containing noapoptosis inducers at the same time point
Statistical Analysis
We determined the signicant differences between treat-ments in individual tumor specimens by using Studentrsquos ttest (Altman 1991) We compared the overall differencein viability in the anti-Fas treatment and control groupsin the medulloblastomaPNET specimens using two-wayANOVA on Stata Corporation 1997 (Stata StatisticalSoftware Release 50 College Station Tex)
Results
Tumor Samples
The 18 pediatric brain tumor specimens analyzed forexpression of Fas and FasL are described in Table 1according to histology patient age and sex The speci-mens consisted of 9 PNETs that included 6 medulloblas-tomas 4 glioblastomas multiforme 3 low-grade gliomasa ganglioglioma and a germ-cell tumor
Fas Expression
Immunohistochemistry Giemsa staining of tumor sec-tions revealed morphologic differences between tumorcells as well as necrotic and apoptotic tissue Small num-bers of normal brain cells epidermal cells and bloodcells were also present in the samples Immunohisto-chemically the Fas receptor was detected in all 18 tumorspecimens (Table 2) The cytoplasm and membrane ofpositive tumor cells stained diffusely Walls of blood ves-sels red blood cells large areas of necrotic tissue andareas of desmoplasia were negative and staining wasconsistently absent in negative controls without primaryor secondary antibody
Tumor sections with little variation in cellular mor-phology demonstrated uniform intensity of Fas stainingthroughout In perinecrotic areas of tumor tissue anapparent increase in the number and staining intensity ofFas-positive tumor cells was frequently observed Tumorswere rated according to the percentage of tumor cellsexpressing the Fas receptor and the staining intensity ofthese cells (Table 2)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001232
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
PNETmedulloblastoma Weakly Fas-positive cellswere found to constitute 25 to 50 of cells in 4 of the9 PNETmedulloblastoma specimens and more than50 of cells in another 4 tumors of this group In theremaining medulloblastoma only occasional cellsappeared immunoreactive to Fas Two tumors PN1 andMB2 otherwise demonstrating relatively consistentstaining for Fas contained populations of apoptotic cellswith hyperchromatic condensed nuclei that did notexpress Fas PN2 contained a large cell population sug-gestive of gliomatous differentiation that expressed Fasmore consistently and prominently than did small cellswithin the tumor Against a weak background of faintlystaining tumor cells MB3 demonstrated intense stainingon large cells with prominent nuclei Occasional Fas-pos-itive Purkinje cells were seen within the tumor tissue
Glioblastoma Of the 4 glioblastomas GBM1 (Fig 1)and GBM4 demonstrated relatively intense staining forFas on more than 50 of cells and GBM2 and GBM3demonstrated more moderate staining on 25 to 50of cells Neurons in the tissue surrounding GBM3 alsoappeared positive for Fas expression
Flow cytometry Fas expression was assessed by owcytometric analysis Fluorescence proles were used tocalculate the MFR and the percentage of cells expressingFas (Fig 2) The well-characterized Jurkat lymphoma cellline was included as a positive control for constitutivereceptor expression Almost all Jurkat cells were found to
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 233
Table 1 Summary of ex vivo pediatric brain tumor specimens
Histology of Patient Tumor No tumor specimen age (yr) Sex
PN1 Pineoblastoma (PNET) 17 F
PN2 PNET 4 M
PN3 PNET 15 F
MB1 Medulloblastoma 8 F
MB2 Medulloblastoma 6 M
MB3 Medulloblastoma 10 F
MB4 Medulloblastoma 8 F
MB5 Medulloblastoma 2 M
MB6 Medulloblastoma 3 F
GBM1 Glioblastoma multiforme 112 M
GBM2 Glioblastoma multiforme 14 M
GBM3 Glioblastoma multiforme 16 M
GBM4 Glioblastoma multiforme 912 F
LGG1 Ganglioglioma 4 F
LGG2 Pilocytic astrocytoma (WHO grade I) 16 M
LGG3 Low-grade astrocytoma (WHO grade II) 7 M
LGG4 Pilocytic astrocytoma (WHO grade I) 14 M
GCT1 Germ cell tumor (germinoma) 15 M
Abbreviation WHO World Health Organization
Table 2 Immunohistochemical and FACS detection of Fas in pediatric brain tumors
Immunohistochemistry FACS
Perinecrotic increase Tumor No Frequencya Intensity Homogeneityb in expressionb MFRc Positived
PN1 ++++ Very weak ndash ndash 165 374
PN2 +++ Moderate ndash + 279 466
PN3 ++++ Moderate-weak + ndash 374 742
MB1 ++++ Moderate ndash + 243 545
MB2 ++++ Weak ndash ndash 285 501
MB3 +++ Weak-moderate ndash + 194 311
MB4 + Weak + ndash 110 131
MB5 +++ Very weak + ndash 268 491
MB6 +++ Weak ndash + 226 230
GBM1 ++++ Moderate-intense ndash ndash 149 245
GBM2 +++ Weak-moderate ndash + 25 531
GBM3 +++ Moderate ndash + 136 967
GBM4 ++++ Intense-moderate + ndash 305 611
LGG1 +++ Intense ndash ndash 240 252
LGG2 + Weak ndash + 354 580
LGG3 +++ Moderate-intense + ndash 248 642
LGG4 +++ Weak ndash ndash 213 444
GCT1 +++ Intense ndash ndash 229 391
Jurkat ND 384 930
Abbreviation ND not done
a+ lt5 positive cells ++ 5-25 positive cells +++ 25-50 positive cells ++++ gt50 positive cells
b+ presence of staining characteristic ndash absence of staining characteristic
cRatio of the mean uorescence of cells labeled with the anti-Fas antibody DX2 compared with those labeled with an isotype control
dPercentage of cells determined to be positive for Fas expression A region M1 was dened at its lower limit by the intersection of the uorescence proles of isotype- and anti-Fas
(DX2)-labeled cells The percentage of Fas-positive cells was calculated as the difference between the percentage of isotype-labeled and DX2-labeled cells uorescing within this region
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
be positive and relatively homogeneous for Fas expres-sion Surface expression of Fas was detected in all 18 pri-mary brain tumors The level of Fas expression bothwith regard to the density per cell (MFR) and the per-centage of Fas-positive cells varied considerably acrossthe spectrum of tumors and within each tumor type(Table 2) There was a relationship between the percent-age of Fas-positive cells and MFR (r = 084) Howeverno real difference was observed between tumor subtypes(data not shown) The variation in Fas expression in indi-vidual tumor specimens was generally greater than thatin the Jurkat population This was exemplied by the
proles of two specimens GBM1 (Fig 1) and GBM3which demonstrated small proportions of cells uoresc-ing at levels 1 or 2 log higher than the peak uorescenceof the population
The level of Fas expression as measured both by thepercentage of Fas-positive cells (421 plusmn 181) and theMFR (237 plusmn 070) in tumor samples was considerablyless than that observed in the Jurkat cells There was apoor correlation between surface Fas expression meas-ured by ow cytometry and immunohistochemical posi-tivity suggesting that in some tumors a large proportionof Fas may be intracellular
FasL Expression
Immunohistochemistry Immunohistochemically FasL wasdetected in 17 of the 18 tumors (Table 3) There tended tobe more homogeneity both across tumor subtypes andwithin each individual tumor than for Fas expressionPNETmedulloblastoma specimens all demonstratedexpression of FasL on more than 50 of tumor cells at var-ious staining intensities Apoptotic cells in PNETmedul-loblastoma specimens that did not express Fas were simi-larly not immunoreactive for FasL
Western blotting Western blotting detected FasL in 10 ofthe 13 tumors examined (Table 3) Of the 7 PNETmedul-loblastomas examined 5 were positive for FasL expressionStaining intensity of positive specimens was compared withreference to the b-tubulin loading control (Fig 3) Intensityranged from very weak (MB4) to strong (MB5 and MB6)with MB1 and MB2 being intermediate PNET 2 andPNET3 specimens were negative for FasL expression Of theglioblastomas all expressed FasL with GBM4 having thehighest level of expression of all the tumors tested HoweverGBM1 and GBM3 showed relatively low levels of expres-sion Of the low-grade tumors LGG4 showed a low expres-sion level with the other two having no or barelydetectable FasL expression Many cross-reactivehighndashmolecular-weight bands were seen in most tumor sam-ples and this may account for the poor correlation betweenthe Western blot and immunohistochemical results
Sensitivity of tumor specimens to Fas-mediated apo-ptosis Cell viability was measured by ow cytometry at24 h and by a colorimetric assay for total viable cell num-ber at 24- and 48-h time points Additionally sub G1peak analysis of cells cultured for 24 h in the presence orabsence of apoptotic mediators provided a measure ofthe percentage of apoptotic cells present in each speci-men Cell viabilityapoptosis in the presence of etoposideor anti-Fas treatment was expressed as a percentage rel-ative to the number of viableapoptotic cells in theabsence of any treatment at the same time point
Cell death assays were rst established using theJurkat cell line with APO-1 antibody in conjunction withprotein A or Neuro-2AFasL supernatant uid A colori-metric assay for total viable cell number exposing Jurkatcells to various titrations of Neuro-2AFasL supernatantuid indicated supernatant uid diluted up to 110 wassufcient to kill 100 of Jurkat cells in culture (data notshown) The loss of viability was due to induction ofapoptosis as indicated by sub G1 peak analysis Additionof Neuro-2Aneo control supernatant uid not contain-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001234
Fig 1 Immunohistochemical analysis of Fas expression in glioblas-toma specimen GBM1 A Clusters of tumor cells surrounded by con-nective tissue as demo in a section stained with hematoxylin andeosin B A similar section of tumor incubated with nonimmune rab-bit serum is completely negative for staining C Fas is expressed ontumor cells in the specimen but the connective tissue is negative forFas Original magnication acute150
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
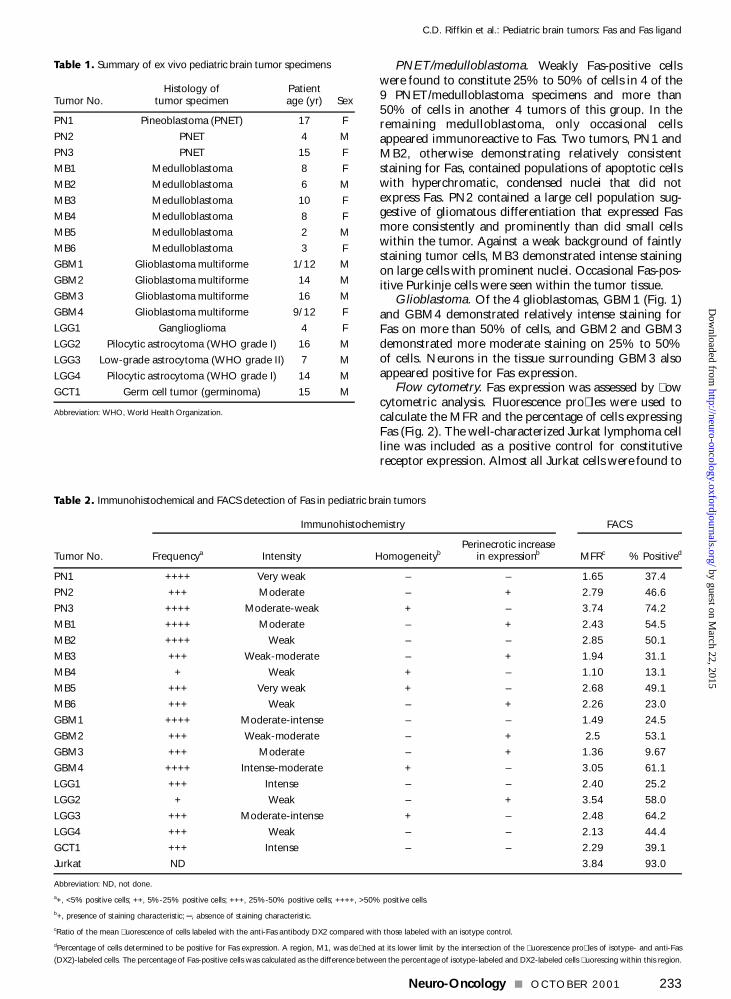
PNETmedulloblastoma Weakly Fas-positive cellswere found to constitute 25 to 50 of cells in 4 of the9 PNETmedulloblastoma specimens and more than50 of cells in another 4 tumors of this group In theremaining medulloblastoma only occasional cellsappeared immunoreactive to Fas Two tumors PN1 andMB2 otherwise demonstrating relatively consistentstaining for Fas contained populations of apoptotic cellswith hyperchromatic condensed nuclei that did notexpress Fas PN2 contained a large cell population sug-gestive of gliomatous differentiation that expressed Fasmore consistently and prominently than did small cellswithin the tumor Against a weak background of faintlystaining tumor cells MB3 demonstrated intense stainingon large cells with prominent nuclei Occasional Fas-pos-itive Purkinje cells were seen within the tumor tissue
Glioblastoma Of the 4 glioblastomas GBM1 (Fig 1)and GBM4 demonstrated relatively intense staining forFas on more than 50 of cells and GBM2 and GBM3demonstrated more moderate staining on 25 to 50of cells Neurons in the tissue surrounding GBM3 alsoappeared positive for Fas expression
Flow cytometry Fas expression was assessed by owcytometric analysis Fluorescence proles were used tocalculate the MFR and the percentage of cells expressingFas (Fig 2) The well-characterized Jurkat lymphoma cellline was included as a positive control for constitutivereceptor expression Almost all Jurkat cells were found to
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 233
Table 1 Summary of ex vivo pediatric brain tumor specimens
Histology of Patient Tumor No tumor specimen age (yr) Sex
PN1 Pineoblastoma (PNET) 17 F
PN2 PNET 4 M
PN3 PNET 15 F
MB1 Medulloblastoma 8 F
MB2 Medulloblastoma 6 M
MB3 Medulloblastoma 10 F
MB4 Medulloblastoma 8 F
MB5 Medulloblastoma 2 M
MB6 Medulloblastoma 3 F
GBM1 Glioblastoma multiforme 112 M
GBM2 Glioblastoma multiforme 14 M
GBM3 Glioblastoma multiforme 16 M
GBM4 Glioblastoma multiforme 912 F
LGG1 Ganglioglioma 4 F
LGG2 Pilocytic astrocytoma (WHO grade I) 16 M
LGG3 Low-grade astrocytoma (WHO grade II) 7 M
LGG4 Pilocytic astrocytoma (WHO grade I) 14 M
GCT1 Germ cell tumor (germinoma) 15 M
Abbreviation WHO World Health Organization
Table 2 Immunohistochemical and FACS detection of Fas in pediatric brain tumors
Immunohistochemistry FACS
Perinecrotic increase Tumor No Frequencya Intensity Homogeneityb in expressionb MFRc Positived
PN1 ++++ Very weak ndash ndash 165 374
PN2 +++ Moderate ndash + 279 466
PN3 ++++ Moderate-weak + ndash 374 742
MB1 ++++ Moderate ndash + 243 545
MB2 ++++ Weak ndash ndash 285 501
MB3 +++ Weak-moderate ndash + 194 311
MB4 + Weak + ndash 110 131
MB5 +++ Very weak + ndash 268 491
MB6 +++ Weak ndash + 226 230
GBM1 ++++ Moderate-intense ndash ndash 149 245
GBM2 +++ Weak-moderate ndash + 25 531
GBM3 +++ Moderate ndash + 136 967
GBM4 ++++ Intense-moderate + ndash 305 611
LGG1 +++ Intense ndash ndash 240 252
LGG2 + Weak ndash + 354 580
LGG3 +++ Moderate-intense + ndash 248 642
LGG4 +++ Weak ndash ndash 213 444
GCT1 +++ Intense ndash ndash 229 391
Jurkat ND 384 930
Abbreviation ND not done
a+ lt5 positive cells ++ 5-25 positive cells +++ 25-50 positive cells ++++ gt50 positive cells
b+ presence of staining characteristic ndash absence of staining characteristic
cRatio of the mean uorescence of cells labeled with the anti-Fas antibody DX2 compared with those labeled with an isotype control
dPercentage of cells determined to be positive for Fas expression A region M1 was dened at its lower limit by the intersection of the uorescence proles of isotype- and anti-Fas
(DX2)-labeled cells The percentage of Fas-positive cells was calculated as the difference between the percentage of isotype-labeled and DX2-labeled cells uorescing within this region
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
be positive and relatively homogeneous for Fas expres-sion Surface expression of Fas was detected in all 18 pri-mary brain tumors The level of Fas expression bothwith regard to the density per cell (MFR) and the per-centage of Fas-positive cells varied considerably acrossthe spectrum of tumors and within each tumor type(Table 2) There was a relationship between the percent-age of Fas-positive cells and MFR (r = 084) Howeverno real difference was observed between tumor subtypes(data not shown) The variation in Fas expression in indi-vidual tumor specimens was generally greater than thatin the Jurkat population This was exemplied by the
proles of two specimens GBM1 (Fig 1) and GBM3which demonstrated small proportions of cells uoresc-ing at levels 1 or 2 log higher than the peak uorescenceof the population
The level of Fas expression as measured both by thepercentage of Fas-positive cells (421 plusmn 181) and theMFR (237 plusmn 070) in tumor samples was considerablyless than that observed in the Jurkat cells There was apoor correlation between surface Fas expression meas-ured by ow cytometry and immunohistochemical posi-tivity suggesting that in some tumors a large proportionof Fas may be intracellular
FasL Expression
Immunohistochemistry Immunohistochemically FasL wasdetected in 17 of the 18 tumors (Table 3) There tended tobe more homogeneity both across tumor subtypes andwithin each individual tumor than for Fas expressionPNETmedulloblastoma specimens all demonstratedexpression of FasL on more than 50 of tumor cells at var-ious staining intensities Apoptotic cells in PNETmedul-loblastoma specimens that did not express Fas were simi-larly not immunoreactive for FasL
Western blotting Western blotting detected FasL in 10 ofthe 13 tumors examined (Table 3) Of the 7 PNETmedul-loblastomas examined 5 were positive for FasL expressionStaining intensity of positive specimens was compared withreference to the b-tubulin loading control (Fig 3) Intensityranged from very weak (MB4) to strong (MB5 and MB6)with MB1 and MB2 being intermediate PNET 2 andPNET3 specimens were negative for FasL expression Of theglioblastomas all expressed FasL with GBM4 having thehighest level of expression of all the tumors tested HoweverGBM1 and GBM3 showed relatively low levels of expres-sion Of the low-grade tumors LGG4 showed a low expres-sion level with the other two having no or barelydetectable FasL expression Many cross-reactivehighndashmolecular-weight bands were seen in most tumor sam-ples and this may account for the poor correlation betweenthe Western blot and immunohistochemical results
Sensitivity of tumor specimens to Fas-mediated apo-ptosis Cell viability was measured by ow cytometry at24 h and by a colorimetric assay for total viable cell num-ber at 24- and 48-h time points Additionally sub G1peak analysis of cells cultured for 24 h in the presence orabsence of apoptotic mediators provided a measure ofthe percentage of apoptotic cells present in each speci-men Cell viabilityapoptosis in the presence of etoposideor anti-Fas treatment was expressed as a percentage rel-ative to the number of viableapoptotic cells in theabsence of any treatment at the same time point
Cell death assays were rst established using theJurkat cell line with APO-1 antibody in conjunction withprotein A or Neuro-2AFasL supernatant uid A colori-metric assay for total viable cell number exposing Jurkatcells to various titrations of Neuro-2AFasL supernatantuid indicated supernatant uid diluted up to 110 wassufcient to kill 100 of Jurkat cells in culture (data notshown) The loss of viability was due to induction ofapoptosis as indicated by sub G1 peak analysis Additionof Neuro-2Aneo control supernatant uid not contain-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001234
Fig 1 Immunohistochemical analysis of Fas expression in glioblas-toma specimen GBM1 A Clusters of tumor cells surrounded by con-nective tissue as demo in a section stained with hematoxylin andeosin B A similar section of tumor incubated with nonimmune rab-bit serum is completely negative for staining C Fas is expressed ontumor cells in the specimen but the connective tissue is negative forFas Original magnication acute150
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
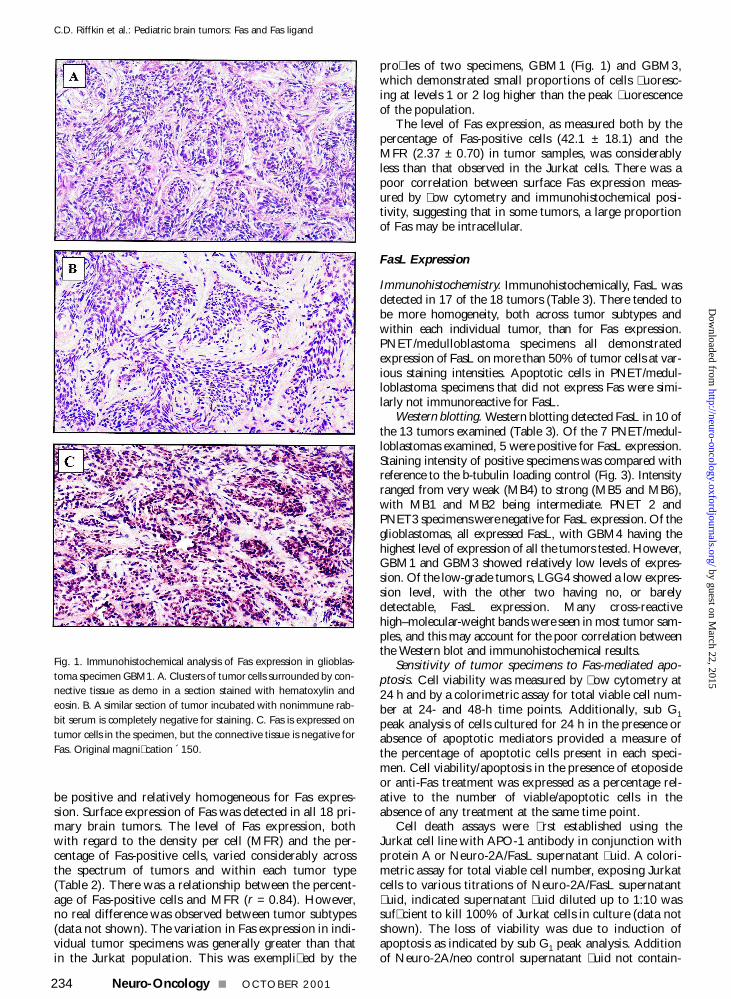
be positive and relatively homogeneous for Fas expres-sion Surface expression of Fas was detected in all 18 pri-mary brain tumors The level of Fas expression bothwith regard to the density per cell (MFR) and the per-centage of Fas-positive cells varied considerably acrossthe spectrum of tumors and within each tumor type(Table 2) There was a relationship between the percent-age of Fas-positive cells and MFR (r = 084) Howeverno real difference was observed between tumor subtypes(data not shown) The variation in Fas expression in indi-vidual tumor specimens was generally greater than thatin the Jurkat population This was exemplied by the
proles of two specimens GBM1 (Fig 1) and GBM3which demonstrated small proportions of cells uoresc-ing at levels 1 or 2 log higher than the peak uorescenceof the population
The level of Fas expression as measured both by thepercentage of Fas-positive cells (421 plusmn 181) and theMFR (237 plusmn 070) in tumor samples was considerablyless than that observed in the Jurkat cells There was apoor correlation between surface Fas expression meas-ured by ow cytometry and immunohistochemical posi-tivity suggesting that in some tumors a large proportionof Fas may be intracellular
FasL Expression
Immunohistochemistry Immunohistochemically FasL wasdetected in 17 of the 18 tumors (Table 3) There tended tobe more homogeneity both across tumor subtypes andwithin each individual tumor than for Fas expressionPNETmedulloblastoma specimens all demonstratedexpression of FasL on more than 50 of tumor cells at var-ious staining intensities Apoptotic cells in PNETmedul-loblastoma specimens that did not express Fas were simi-larly not immunoreactive for FasL
Western blotting Western blotting detected FasL in 10 ofthe 13 tumors examined (Table 3) Of the 7 PNETmedul-loblastomas examined 5 were positive for FasL expressionStaining intensity of positive specimens was compared withreference to the b-tubulin loading control (Fig 3) Intensityranged from very weak (MB4) to strong (MB5 and MB6)with MB1 and MB2 being intermediate PNET 2 andPNET3 specimens were negative for FasL expression Of theglioblastomas all expressed FasL with GBM4 having thehighest level of expression of all the tumors tested HoweverGBM1 and GBM3 showed relatively low levels of expres-sion Of the low-grade tumors LGG4 showed a low expres-sion level with the other two having no or barelydetectable FasL expression Many cross-reactivehighndashmolecular-weight bands were seen in most tumor sam-ples and this may account for the poor correlation betweenthe Western blot and immunohistochemical results
Sensitivity of tumor specimens to Fas-mediated apo-ptosis Cell viability was measured by ow cytometry at24 h and by a colorimetric assay for total viable cell num-ber at 24- and 48-h time points Additionally sub G1peak analysis of cells cultured for 24 h in the presence orabsence of apoptotic mediators provided a measure ofthe percentage of apoptotic cells present in each speci-men Cell viabilityapoptosis in the presence of etoposideor anti-Fas treatment was expressed as a percentage rel-ative to the number of viableapoptotic cells in theabsence of any treatment at the same time point
Cell death assays were rst established using theJurkat cell line with APO-1 antibody in conjunction withprotein A or Neuro-2AFasL supernatant uid A colori-metric assay for total viable cell number exposing Jurkatcells to various titrations of Neuro-2AFasL supernatantuid indicated supernatant uid diluted up to 110 wassufcient to kill 100 of Jurkat cells in culture (data notshown) The loss of viability was due to induction ofapoptosis as indicated by sub G1 peak analysis Additionof Neuro-2Aneo control supernatant uid not contain-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001234
Fig 1 Immunohistochemical analysis of Fas expression in glioblas-toma specimen GBM1 A Clusters of tumor cells surrounded by con-nective tissue as demo in a section stained with hematoxylin andeosin B A similar section of tumor incubated with nonimmune rab-bit serum is completely negative for staining C Fas is expressed ontumor cells in the specimen but the connective tissue is negative forFas Original magnication acute150
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

ing FasL at equivalent dilutions had no effect on viabilitycompared with cells cultured in medium alone
Specicity of killing was conrmed by demonstratingthat the cytotoxicity of Neuro-2AFasL supernatant uidcould be abrogated by the addition of a monoclonal anti-Fas antibody (DX2) that competitively inhibits bindingof FasL to its receptor (data not shown) (Huang et al1999) The cell death rate in Jurkat with APO-1protein Awas the same as with the highly apoptotic Neuro-2AFasL supernatant uid However the death rate incells assayed with APO-1 antibody alone was far lowerAfter this observation Jurkat was used as a positive con-trol in comparing Neuro-2AFasL supernatant uid toAPO-1protein A in MB1 Again both Neuro-2AFasLsupernatant uid and APO-1protein A were highlyapoptotic in the Jurkat cell line and showed the samedegree of cell death in the tumor sample indicating thatNeuro-2AFasL supernatant uid and APO-1protein Aare equivalent in these assays Therefore PNET2 wasassayed using Neuro-2AFasL supernatant uid onlyagain with Jurkat as a control
The reference Jurkat cell line also showed a dramaticdecrease in the percentage of viable cells in the presence ofthe topoisomerase II inhibitor etoposide The corre-
sponding increase in the percentage of apoptotic cellnumbers conrmed the loss of viability was due to induc-tion of apoptosis by this agent
Of the 18 tumor specimens 9 were analyzed for sen-sitivity to Fas-mediated and drug-induced apoptosisincluding 4 PNETs (2 medulloblastomas) 3 glioblas-tomas and 2 low-grade gliomas These tumors wereselected according to the availability of tumor tissue Inall 9 tumor specimens spontaneous cell death occurredand a decrease in the number of viable cells was observedover 48 h of culturing in medium without the addition ofetoposide or anti-Fas antibodyprotein A In most tumors(7 of 9) the loss of viable cell number was less than 50(data not shown) when compared with the viability atthe initial analysis
The measures of viability at 24-h ow cytometry andcolorimetric viability assay were comparable Tumorsresistant to either anti-Fas treatment or etoposide at the24-h time point were similarly resistant at 48 h In tumorsamples sensitive to these agents at 24 h the effectbecame more pronounced at the 48-h time point of thecolorimetric viability assay Of the 9 tumors 4 were sen-sitive to apoptosis mediated by either etoposide or theFas receptor (Table 4)
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 235
Fig 2 A Flow cytometric analysis of Fas expression in the Jurkat cell line B A PNETmedulloblastoma specimen MB5 C A glioblastoma mul-tiforme specimen GBM1 D A low-grade glioma LGG1 comparing the uorescence proles of unstained cells cells labeled with a nonimmunecontrol antibody and cells labeled with the anti-Fas antibody DX2 The difference between the percentage of DX2-labeled and isotype-labeledcells within region M1 denes the percentage of cells positive for Fas expression Histograms are labeled with values for mean uorescence ratioand percentage of cells positive for Fas expression
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
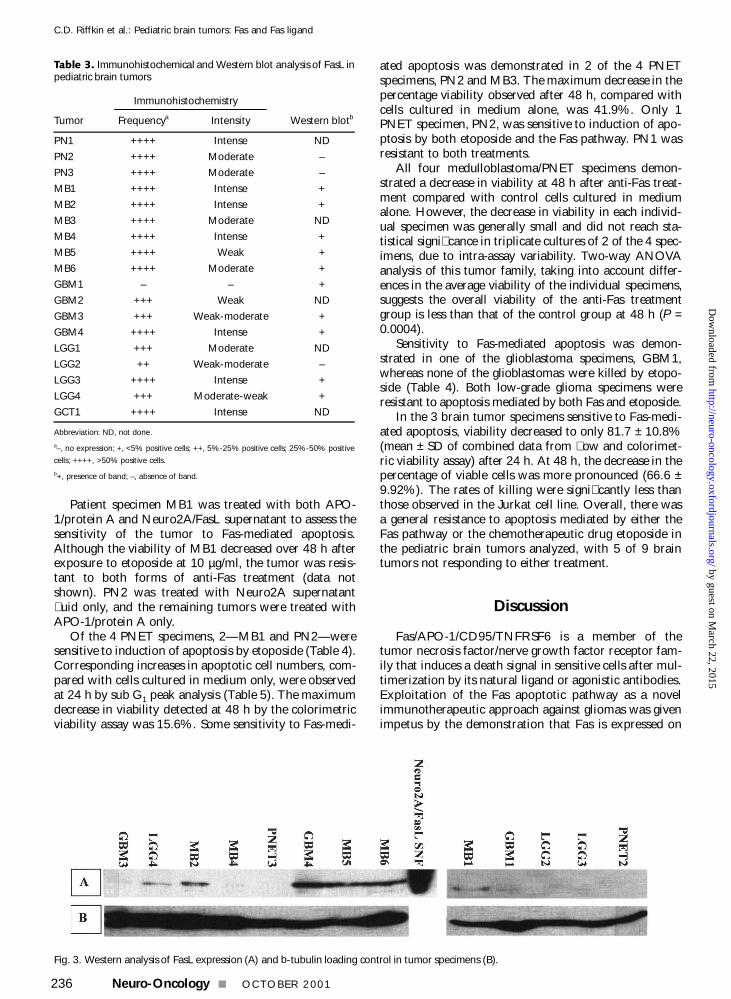
Patient specimen MB1 was treated with both APO-1protein A and Neuro2AFasL supernatant to assess thesensitivity of the tumor to Fas-mediated apoptosisAlthough the viability of MB1 decreased over 48 h afterexposure to etoposide at 10 microgml the tumor was resis-tant to both forms of anti-Fas treatment (data notshown) PN2 was treated with Neuro2A supernatantuid only and the remaining tumors were treated withAPO-1protein A only
Of the 4 PNET specimens 2mdashMB1 and PN2mdashweresensitive to induction of apoptosis by etoposide (Table 4)Corresponding increases in apoptotic cell numbers com-pared with cells cultured in medium only were observedat 24 h by sub G1 peak analysis (Table 5) The maximumdecrease in viability detected at 48 h by the colorimetricviability assay was 156 Some sensitivity to Fas-medi-
ated apoptosis was demonstrated in 2 of the 4 PNETspecimens PN2 and MB3 The maximum decrease in thepercentage viability observed after 48 h compared withcells cultured in medium alone was 419 Only 1PNET specimen PN2 was sensitive to induction of apo-ptosis by both etoposide and the Fas pathway PN1 wasresistant to both treatments
All four medulloblastomaPNET specimens demon-strated a decrease in viability at 48 h after anti-Fas treat-ment compared with control cells cultured in mediumalone However the decrease in viability in each individ-ual specimen was generally small and did not reach sta-tistical signicance in triplicate cultures of 2 of the 4 spec-imens due to intra-assay variability Two-way ANOVAanalysis of this tumor family taking into account differ-ences in the average viability of the individual specimenssuggests the overall viability of the anti-Fas treatmentgroup is less than that of the control group at 48 h (P =00004)
Sensitivity to Fas-mediated apoptosis was demon-strated in one of the glioblastoma specimens GBM1whereas none of the glioblastomas were killed by etopo-side (Table 4) Both low-grade glioma specimens wereresistant to apoptosis mediated by both Fas and etoposide
In the 3 brain tumor specimens sensitive to Fas-medi-ated apoptosis viability decreased to only 817 plusmn 108(mean plusmn SD of combined data from ow and colorimet-ric viability assay) after 24 h At 48 h the decrease in thepercentage of viable cells was more pronounced (666 plusmn992) The rates of killing were signicantly less thanthose observed in the Jurkat cell line Overall there wasa general resistance to apoptosis mediated by either theFas pathway or the chemotherapeutic drug etoposide inthe pediatric brain tumors analyzed with 5 of 9 braintumors not responding to either treatment
Discussion
FasAPO-1CD95TNFRSF6 is a member of thetumor necrosis factornerve growth factor receptor fam-ily that induces a death signal in sensitive cells after mul-timerization by its natural ligand or agonistic antibodiesExploitation of the Fas apoptotic pathway as a novelimmunotherapeutic approach against gliomas was givenimpetus by the demonstration that Fas is expressed on
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001236
Table 3 Immunohistochemical and Western blot analysis of FasL inpediatric brain tumors
Immunohistochemistry
Tumor Frequencya Intensity Western blotb
PN1 ++++ Intense ND
PN2 ++++ Moderate ndash
PN3 ++++ Moderate ndash
MB1 ++++ Intense +
MB2 ++++ Intense +
MB3 ++++ Moderate ND
MB4 ++++ Intense +
MB5 ++++ Weak +
MB6 ++++ Moderate +
GBM1 ndash ndash +
GBM2 +++ Weak ND
GBM3 +++ Weak-moderate +
GBM4 ++++ Intense +
LGG1 +++ Moderate ND
LGG2 ++ Weak-moderate ndash
LGG3 ++++ Intense +
LGG4 +++ Moderate-weak +
GCT1 ++++ Intense ND
Abbreviation ND not done
andash no expression + lt5 positive cells ++ 5-25 positive cells 25-50 positive
cells ++++ gt50 positive cells
b+ presence of band ndash absence of band
Fig 3 Western analysis of FasL expression (A) and b-tubulin loading control in tumor specimens (B)
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
malignant glioma cells both in vitro (Weller et al 1994)and ex vivo (Frei et al 1998 Tachibana et al 1995Tohma et al 1998) that glioma cells are susceptible toFas-induced apoptosis (Weller et al 1994 1995a1995b) and that Fas is absent from normal brainparenchyma (Leithauser et al 1993) Sensitivity to Fasin ex vivo glioma specimens has recently been reported(Frei et al 1998) It has also been shown that somemedulloblastoma cell lines show a low level of Fasexpression coexpression of FasL and sensitivity toFasL-mediated apoptosis However to date there hasbeen no study of ex vivo tissue from pediatric braintumors or brain tumors other than adult gliomas (Welleret al 1998) It is important to study Fas expression andactivity in freshly resected primary tumor specimensbecause due to the selection pressures of culture condi-tions the in vitro phenotype of tumor cells may notreect the in vivo situation (Izumu et al 1993) The exvivo tumors analyzed in this study consisted of varioussubtypes including 9 PNETmedulloblastomas 4glioblastomas multiforme 4 low-grade gliomas and agerm-cell tumor of the CNS
The Fas receptor was detected by ow cytometry in all18 brain tumors Compared with the Jurkat cell line amoderate level of Fas expression was detected in primarypediatric tumors of the CNS There was signicant het-erogeneity in Fas expression within each tumor subtypeand no obvious correlation with grades of malignancy aspreviously reported for expression of Fas mRNA in adultmalignant gliomas (Tachibana et al 1995)
Expression of Fas on tumor cells in each of the 18specimens was confirmed by immunohistochemicalanalysis As observed by ow cytometric immunopheno-typing expression of the Fas receptor again appearedquite heterogeneous both in individual tumor sectionsand within tumor subtypes In concert with previousreports (Tohma et al 1998) there was an apparentincrease in the frequency and intensity of Fas immunore-activity in perinecrotic areas of several sections
Of interest in this study was the observation inPNETmedulloblastoma specimens (PN1 MB2) of Fas-negative or weakly staining apoptotic cells among other-wise homogenous populations of tumor cells expressingFas These apoptotic cells similarly demonstrated mini-mal expression of FasL by immunohistochemistry Thismay indicate that the process of apoptosis is associatedwith down regulation of expression of Fas and FasL orthat proteolysis during apoptosis destroys the Fas andFasL protein Alternatively tumor cells that coexpressthe Fas receptor and ligand may have acquired mecha-nisms of resistance to pathways of apoptosis notrestricted to the Fas pathway to prevent Fas-induced celldeath resulting from contact with surrounding FasL-expressing malignant cells
Finally Fas expression was detected by immunohisto-chemical staining on normal Purkinje cells of the cerebel-lum in a medulloblastoma (MB3) and on neurons sur-rounding a glioblastoma multiforme (GBM3) Previousstudies have demonstrated absence of staining on neu-
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 237
Table 5 Sub G1 peak analysis of apoptosis in tumor specimens aftertreating with anti-Fas or etoposide and culturing for 24 h
Apoptosis with treatment Apoptosis
Tumor in media alonea Etoposide APO-1proteinA
Jurkat 115 plusmn 422 611 plusmn 775b 741 plusmn 105b
PN1 310 314c 312c
PN2 448 plusmn 233 487 plusmn 194b 514 plusmn 299b
MB1 267 plusmn 866 552 plusmn 147b 333 plusmn 167
MB3 410 plusmn 643 3961 plusmn 365 404 plusmn 468
GBM1 208 plusmn 006 248 plusmn 278 273 plusmn 081b
GBM2 578 plusmn 339 572 plusmn 192 562 plusmn 334
GBM3 940 plusmn 084 103 plusmn 144 121 plusmn 137
LGG1 211 plusmn 025 342 plusmn 581 270 plusmn 581
LGG2 373 plusmn 480 300 plusmn 308 388 plusmn 128
Values are mean plusmn SD
aPercentage of apoptotic cells in the specimen after treatment and culturing relative tothe number of apoptotic cells present after culturing in media only at the same time
point
bDifference was statistically signicant from controls lacking APO-1protein A ofNeuro2AFasL treatment (P lt 005)
cNot done in triplicate culture due to size of the available tumor sample
Table 4 FACS and colorimetric assay determination of tumor cellviability after anti-Fas or etoposide treatment and culture over 24or 48 h
FACS analysis Colorimetric assay
Viabilitya Viability Viability Tumor Treatment (24 h) (24 h) (48 h)
Jurkat APO-1PrA 033 plusmn 010bc 139 plusmn 680bc 101 plusmn 202bc
cell line Etoposide 176 plusmn 349bc 724 plusmn 106bc 170 plusmn 833bc
PN1 APO-1PrA 890d 930 884
Etoposide 104d 102 101
PN2 FasL 891c 906e 581c
Etoposide 814c 988 833
MB1 APO-1PrA 125 106 918
Etoposide 258c 703f 744c
MB3 APO-1PrA 937c 750c 642c
Etoposide 966 897 105
GBM1 APO-1PrA 676c 741c 775c
Etoposide 120 103 111
GBM2 APO-1PrA 973 105 974
Etoposide 990 103 102
GBM3 APO-1PrA 129 118 987
Etoposide 109 106 953
LGG1 APO-1PrA 806 111 102
Etoposide 109 126 136
LGG2 APO-1PrA 104 913 961
Etoposide 128 957 110
Abbreviation PrA protein A
aPercentage of viable cells in the specimen after treatment and culturing relative to the
number of viable cells present after culturing in media alone at the same time point
bValues are mean plusmn SD
cDifference was statistically signicant from controls lacking APO-1PrA or Neuro2AFasLtreatment (P lt 005)
dNot done in triplicate culture due to size of the available tumor sample
eP = 009
fP = 007
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

rons in the cerebellar and cerebral cortices (Leithauser etal 1993) However exposure to interferon-g has beenshown to induce low levels of Fas expression on neuronswithout resulting in sensitivity to Fas-mediated apoptosis(Rensing-Ehl et al 1996) Thus Fas expression observedon neurons in this study may be a consequence of theenvironment in and around the tumor tissue The sus-ceptibility of Fas-expressing neurons to Fas-mediatedapoptosis was not explored and indeed receptor expres-sion may not necessarily correlate with Fas-sensitivity Ofthe two specimens containing Fas-expressing neuronsGBM3 demonstrated no decrease in viable cell numberafter anti-Fas treatment whereas a slight decrease wasobserved in MB3 The lack of sensitivity of GBM3 maysuggest that these neurons like the tumor cells are resis-tant to Fas-mediated apoptosis
Of the 9 pediatric brain tumors analyzed in cytotoxi-city assays 6 were resistant to Fas-induced apoptosisdespite the expression of Fas on tumor cells Seven wereresistant to apoptosis induced by the topoisomerase IIinhibitor etoposide Analysis of the overall viability of themedulloblastomaPNET tumors as a group demon-strated a reduced viability of tumor cells 48 h after expo-sure to anti-Fas treatment compared with exposure tomedium alone This observation suggests that this sub-group of embryonal tumors appears to exhibit an overallsensitivity to Fas not observed in the astrocytic pediatrictumors included in this study Furthermore the glioblas-toma GBM1 that appeared sensitive to Fas-inducedapoptosis was derived from an infant
One glioblastoma and 2 PNETs demonstrated sensi-tivity to Fas-mediated apoptosis The loss of viability inthese specimens was signicantly less pronounced thanthat observed in the Jurkat cell line after 48 h The dif-ference in effect may be due in part to the presence ofproliferating Fas-negative cells in short-term cultures ofprimary tumor specimens At each time point viabilityin the presence of either treatment was expressed rela-tive to the number of viable cells in control samplesthat had been cultured in medium alone Jurkat con-trols cultured in medium over a 48-h period wereobserved to proliferate with absorbance readings inthe colorimetric viability assay tripling over the timeperiod (data not shown) The equivalent tumor con-trols demonstrated a decrease in viability over timeThus for an equivalent decrease in viable cell numberthe percentage viability calculated would be relativelylarger in tumor specimens compared with the Jurkatcell line Nonetheless the dramatic loss of Jurkat cellviability demonstrated by forward scatterside scatteranalysis and propidium iodide exclusion was notobserved in tumor specimens
Fas sensitivity among the tumor specimens did notcorrelate with the level of Fas expression For instancepatient specimen PN2 was susceptible to Fas-mediatedapoptosis yet demonstrated a lower percentage of Fas-positive cells and a lower MFR than MB1 which wasresistant to Fas-induced apoptosis
The degree of Fas-resistance observed in this studycontrasts with previous ndings (Frei et al 1998)which demonstrated sensitivity to the highly apoptoticNeuro-2AFasL supernatant uid (Schneider et al
1998) Although APO-1 in combination with protein Agave equivalent results to the FasL-transfected Neuro-2A supernatant uid in Jurkat and MB1 the contrast inndings may reect differences between these twodeath-inducing agents Alternatively differences in thebiology of pediatric compared with adult malignanciesof the CNS or variation between individual tumorsmay contribute to the difference in results
Failure of pediatric brain tumor specimens to undergoFas-induced apoptosis despite signicant levels of Fasexpression suggests the existence of mechanisms of resis-tance affecting the ability of the receptor andor the intra-cellular cascade to transduce the death signal The corre-sponding lack of susceptibility to etoposide suggests theresistance observed is not specic for the Fas pathwayand may involve factors involved in multiple pathways ofapoptosis
The previously reported results in glioma specimensand the results of this study suggest expression of the Fasreceptor and its ligand appear to be general characteris-tics of brain tumors It remains to be shown that FasLexpressed on pediatric brain tumors like that on adultgliomas is indeed capable of transducing an apoptoticsignal to Fas-sensitive target cells and maintainingimmune privilege in the tumor environment
The function of Fas expression on brain tumor cellsremains to be explained It has been reported that Fas sig-naling induces the release of cytokines from astrocyteswhich regulate the inammatory response (Saas et al1999) It is also likely that Fas signaling is associatedwith a proliferative effect in T cells (Strasser and New-ton 1999) The coexpression of Fas and FasL in somepediatric brain tumors may help stimulate the prolifera-tive or immunologic arms of the Fas pathway FasLexpression is perhaps easiest to explain in terms of itsrole in the down regulation of activated T cells In thisway the transformed cells could modulate the immuneresponse to a tumor For tumor cells to express both Fasand FasL the proapoptotic arm of the Fas signaling path-way is likely to be nonfunctional in these cells This couldoccur through loss of function of elements in the apo-ptosis pathway or expression of active inhibitors of thepathway The molecular environment within the tumorand the CNS may also alter the consequence of Fas sig-naling within the cell Subversion of elements of the apo-ptosis pathway to a proliferation or survival pathwaywould provide not only a growth advantage but alsocould account for the resistance to apoptosis in thesepediatric brain tumors
The Fas receptor may represent an attractive targetfor an immunotherapeutic approach However despitedemonstration of Fas-sensitivity in medulloblastomaPNET specimens a high degree of resistance to Fas-mediated apoptosis was observed in ex vivo pediatricbrain tumors despite the presence of signicant levels ofreceptor expression This as well as the presence of sub-stantial subpopulations of Fas-negative tumor cells inmost specimens suggests that an immunotherapeuticapproach targeting the Fas-receptor alone may be inef-fective in signicantly reducing the tumor load and itspotential if any may lie in combination with existingtherapies
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001238
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

Altman DG (1991) Practical Statistics for Medical Research London Chap-
man and Hall
Armitage RJ (1994) Tumor necrosis factor receptor superfamily members
and their ligands Curr Opin Immunol 6 407-413
Ashley DM Kong FM Bigner DD and Hale LP (1998) Endogenous
expression of transforming growth factor beta1 inhibits growth and
tumorigenicity and enhances Fas-mediated apoptosis in a murine high-
grade glioma model Cancer Res 58 302-309
Bellgrau D Gold D Selawry H Moore J Franzusoff A and Duke RC
(1995) A role for CD95 ligand in preventing graft rejection Nature 377
630-632
Boldin MP Mett IL Varfolomeev EE Chumakov I Shemer-Avni Y
Camonis JH and Wallach D (1995) Self-association of the ldquodeath
domainsrdquo of the p55 tumor necrosis factor (TNF) receptor and FasAPO-
1 prompts signaling for TNF and FasAPO-1 effects J Biol Chem 270
387-391
Boldin MP Goncharov TM Goltsev YV and Wallach D (1996) Involve-
ment of MACH a novel MORT1FADD-interacting protease in
FasAPO-1- and TNF receptor-induced cell death Cell 85 803-815
Cohen ME and Duffner PK (1994) Brain Tumors in Children Principles of
Diagnosis and Treatment New York Raven Press
Davidson WF Giese T and Fredrickson TN (1998) Spontaneous develop-
ment of plasmacytoid tumors in mice with defective Fas-Fas ligand inter-
actions J Exp Med 187 1825-1838
Duke RC Franzusoff A and Bellgrau D (1996) CD95 ligand in graft rejec-
tion Nature 379 682-683
Frei K Ambar B Adachi N Yonekawa Y and Fontana A (1998) Ex vivo
malignant glioma cells are sensitive to Fas (CD95APO-1) ligand-medi-
ated apoptosis J Neuroimmunol 87 105-113
Gratas C Tohma Y Barnas C Taniere P Hainaut P and Ohgaki H
(1998) Up-regulation of Fas (APO-1CD95) ligand and down-regulation
of Fas expression in human esophageal cancer Cancer Res 58 2057-
2062
Grifth TS Brunner T Fletcher SM Green DR and Ferguson TA
(1995) Fas ligand-induced apoptosis as a mechanism of immune privilege
Science 270 1189-1192
Grifth TS Yu X Herndon JM Green DR and Ferguson TA (1996)
CD95-induced apoptosis of lymphocytes in an immune privileged site
induces immunological tolerance Immunity 5 7-16
Hahne M Rimoldi D Schroter M Romero P Schreier M French LE
Schneider P Bornand T Fontana A Lienard D Cerottini J and
Tschopp J (1996) Melanoma cell expression of Fas (APO-1CD95)
ligand Implications for tumor immune escape Science 274 1363-1366
Hanabuchi S Koyanagi M Kawasaki A Shinohara N Matsuzawa A
Nishimura Y Kobayashi Y Yonehara S Yagita H and Okumura K
(1994) Fas and its ligand in a general mechanism of T-cell-mediated cyto-
toxicity Proc Natl Acad Sci USA 91 4930-4934
Huang DC Hahne M Schroeter M Frei K Fontana A Villunger A
Newton K Tschopp J and Strasser A (1999) Activation of Fas by FasL
induces apoptosis by a mechanism that cannot be blocked by Bcl-2 or Bcl-
x(L) Proc Natl Acad Sci USA 96 14871-14876
Itoh N Tsujimoto Y and Nagata S (1993) Effect of Bcl-2 on Fas antigen-
mediated cell death J Immunol 151 621-627
Itoh Y Yonehara S Ishii A Yonehara M Mizushima S Sameshima M
Hase A Seto Y and Nagata S (1991) The polypeptide encoded by the
cDNA for human cell surface antigen Fas can mediate apoptosis Cell 66
233-243
Izumu I Mineura K Watanabe K and Kowada M (1993) Characteriza-
tion and chemosensitivity of two cell lines derived from human glioblas-
tomas J Neurooncol 17 111-121
Kawai K Serizawa A Hamana T and Tsutsumi Y (1994) Heat-induced
antigen retrieval of proliferating cell nuclear antigen and p53 protein in
formalin-xed parafn-embedded sections Pathol Int 44 759-764
Kayagaki N Kawasaki A Ebata T Ohmoto H Ikeda S Inoue S
Yoshino K Okumura K and Yagita H (1995) Metaloprotease-medi-
ated release of human Fas ligand J Exp Med 182 1777-1783
Kayagaki N Yamaguchi N Nagao F Matsuo S Maeda H Okumura K
and Yagita H (1997) Polymorphism of murine Fas ligand that affects the
biological activity Proc Natl Acad Sci USA 94 3914-3919
Kischkel FC Hellbardt S Behrmann I Germer M Pawlita M Krammer
PH and Peter ME (1995) Cytotoxicity-dependent APO-1 (FasCD95)-
associated proteins form a death-inducing signaling complex (DISC) with
the receptor EMBO J 14 5579-5588
Leithauser F Dhein J Mechtersheimer G Koretz K Bruderlein S Henne
C Schmidt A Debatin KM Krammer PH and Moller P (1993) Con-
stitutive and induced expression of APO-1 a new member of the nerve
growth factortumor necrosis factor receptor superfamily in normal and
neoplastic cells Lab Invest 69 415-429
Medema JP Scafdi C Kischkel FC Shevchenko A Mann M Kram-
mer PH and Peter ME (1997) FLICE is activated by association with the
CD95 death-inducing signaling complex (DISC) EMBO J 16 2794-2804
Muzio M Chinnaiyan AM Kischkel FC OrsquoRourke K Shevchenko A
Ni J Scafdi C Bretz JD Zhang M Gentz R Mann M Krammer
PH Peter ME and Dixit VM (1996) FLICE a novel FADD-homolo-
gous ICECED-3-like protease is recruited to the CD95 (FasAPO-1)
death-inducing signaling complex Cell 85 817-827
Nagata S and Golstein P (1995) The Fas death factor Science 267 1449-
1456
OrsquoConnell J OrsquoSullivan GC Collins JK and Shanahan F (1996) The Fas
counterattack Fas-mediated T cell killing by colon cancer cells expressing
Fas ligand J Exp Med 184 1075-1082
Oehm A Behrmann I Falk W Pawlita M Maier G Klas C Li-Weber
M Richards S Dhein J Trauth BC Ponsting H and Krammer PH
(1992) Purication and molecular cloning of the APO-1 cell surface anti-
gen a member of the tumor necrosis factornerve growth factor receptor
superfamily sequence identity with Fas antigen J Biochem 267 10709-
10715
Oshimi Y Oda S Honda Y Nagata S and Miyazaki S (1996) Involve-
ment of Fas ligand and Fas-mediated pathway in the cytotoxicity of
human natural killer cells J Immunol 157 2909-2915
Rensing-Ehl A Frei K Flury R Matiba B Mariani SM Weller M Aebis-
cher P Krammer PH and Fontana A (1995) Local FasAPO-1 (CD95)
ligand-mediated tumor cell killing in vivo Eur J Immunol 25 2253-
2258
Rensing-Ehl A Malipiero U Irmler M Tschopp J Constam D and
Fontana A (1996) Neurons induced to express major histocompatibility
complex class I antigen are killed via the perforin and not the Fas (APO-
1CD95) pathway Eur J Immunol 26 2271-2274
Saas P Boucraut J Quiquerez AL Schnuriger V Perrin G Desplat-Jego
S Bernard D Walker PR and Dietrich PY (1999) CD95 (FasAPO-1)
as a receptor governing astrocyte apoptotic or inammatory responses A
key role in brain inammation J Immunol 162 2326-2333
Saas P Walker PR Hahne M Quiquerez AL Schnuriger V Perrin G
French L Van Meir EV de Tribolet ND Tschopp J and Dietrich PY
(1997) Fas ligand expression by astrocytoma in vivo Maintaining immune
privlege in the brain J Clin Invest 99 1173-1178
Scafdi C Fulda S Srinivasen A Friesen C Li F Tomaselli KJ Debatin
KM Krammer PH and Peter ME (1998) Two CD95 (APO-1Fas) sig-
naling pathways EMBO J 17 1675-1687
Schneider P Holler N Bodmer JL Hahne M Frei K Fontana A and
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 20 01 239
References
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from

Tschopp J (1998) Conversion of membrane-bound Fas(CD95) ligand to
its soluble form is associated with downregulation of its proapoptotic
activity and loss of liver toxicity J Exp Med 18 1205-1213
Singer GG Carrera AC Marshak-Rothstein A Martinez C and Abbas
AK (1994) Apoptosis Fas and systemic autoimmunity The MRL-lprlpr
model Curr Opin Immunol 6 913-920
Strand S Hofmann WJ Hug H Muller M Otto G Strand D Mariani
SM Stremmel W Krammer PH and Galle PR (1996) Lymphocyte
apoptosis induced by CD95 (APO-1Fas) ligand-expressing tumor cells mdash
a mechanism of immune invasion Nat Med 2 1361-1366
Strasser A and Newton K (1999) FADDMORT1 a signal transducer that
can promote cell death or cell growth Int J Biochem Cell Biol 31 533-
537
Suda T Takahashi T Goldstein P and Nagata S (1993) Molecular cloning
and expression of the Fas ligand a novel member of the tumor necrosis
factor family Cell 75 1169-1178
Tachibana O Nakazawa H Lampe J Watanabe K Kleihues P and
Ohgaki H (1995) Expression of FasAPO-1 during the progression of
astrocytomas Cancer Res 55 5528-5530
Tanaka M Suda T Takahashi T and Nagata S (1995) Expression of the
functional soluble form of human fas ligand in activated lymphocytes
EMBO J 14 1129-1135
Tohma Y Gratas C Van Meir EG Desbaillets I Tenan M Tachibana O
Kleihues P and Ohgaki H (1998) Necrogenesis and FasAPO-1 (CD95)
expression in primary (de novo) and secondary glioblastomas J Neu-
ropathol Exp Neurol 57 239-245
Tounekti O Belehradek J Jr and Mir LM (1995) Relationships between
DNA fragmentation chromatin condensation and changes in ow
cytometry proles detected during apoptosis Exp Cell Res 217 506-
516
Trauth BC Klas C Peters AM Matzku S Moller P Falk W Debatin
KM and Krammer PH (1989) Monoclonal antibody-mediated tumor
regression by induction of apoptosis Science 245 301-305
Walker PR Saas P and Dietrich PY (1997) Role of Fas ligand (CD95L) in
immune escape The tumor cell strikes back J Immunol 158 4521-
4524
Watanabe-Fukunaga R Brannan CI Copeland NG Jenkins NA and
Nagata S (1992) Lymphoproliferation disorder in mice explained by
defects in Fas antigen that mediates apoptosis Nature 356 314-317
Weller M Frei K Groscurth P Krammer PH Yonekawa Y and Fontana
A (1994) Anti-FasAPO-1 antibody-mediated apoptosis of cultured
human glioma cells Induction and modulation of sensitivity by cytokines
J Clin Invest 94 954-964
Weller M Malipiero U Aguzzi A Reed JC and Fontana A (1995a)
Protooncogene bcl-2 gene transfer abrogates FasAPO-1 antibody-
mediated apoptosis of human malignant glioma cells and confers resis-
tance to chemotherapeutic drugs and therapeutic irradiation J Clin
Invest 95 2633-2643
Weller M Malipiero U Rensing-Ehl A Barr PJ and Fontana A (1995b)
FasAPO-1 gene transfer for human malignant glioma Cancer Res 55
2936-2944
Weller M Schuster M Pietsch T and Schabet M (1998) CD95 ligand-
induced apoptosis of human medulloblastoma cells Cancer Lett 128
121-126
Werner M Von Wasielewski R and Komminoth P (1996) Antigen
retrieval signal amplication and intensication in immunohistochem-
istry Histochem Cell Biol 105 253-260
Yagita H Seino K Kayagaki N and Okumura K (1996) CD95 ligand in
graft rejection Nature 379 682
Yonehara S Ishii A and Yonehara M (1989) A cell-killing monoclonal
antibody (anti-Fas) to a cell surface antigen co-downregulated with the
receptor of tumor necrosis factor J Exp Med 169 1747-1756
CD Riffkin et al Pediatric brain tumors Fas and Fas ligand
Neuro-Oncology n OCTOBER 2 001240
by guest on March 22 2015
httpneuro-oncologyoxfordjournalsorgD
ownloaded from
Copyright © 2022 FDOKUMEN




















