
Evidence that free fatty acids in trophocytes of Periplaneta americana fat body may be regulated by...
12
Insect Biochem. Molec. Biol. Vol. 27, No. 7, pp. 681–692, 1997 1997 Elsevier Science Ltd Pergamon PII: S0965-1748(97)00046-5 All rights reserved. Printed in Great Britain 0965-1748/97 $17.00 + 0.00 Evidence that Free Fatty Acids in Trophocytes of Periplaneta americana Fat Body may be Regulated by the Activity of Phospholipase A 2 and Cyclooxygenase IRSHAD ALI², JOHN E. STEELE*² Received 14 October 1996; revised and accepted 18 June 1997 Previous studies have shown that palmitic, stearic, oleic and linoleic acid levels in trophocytes prepared from the fat body of male Periplaneta americana are increased following treatment of the cells with hypertrehalosemic hormone (HTH). Melittin, an activator of phospholipase A 2 , mimicked the action of HTH by increasing the free fatty acid content in a concentration- dependent manner. The increase caused by HTH could be eliminated by pretreatment of the trophocytes with 1 mM 49-bromophenacyl bromide (BPB), an inhibitor of phospholipase A 2 . BPB also decreases the concentration of free fatty acids in trophocytes not treated with HTH but by a smaller margin. Nordihydroguaiaretic acid (NDGA) and indomethacin, inhibitors of lipoxygenase and cyclooxygenase, respectively, eliminated the increase in free fatty acids evoked by HTH. In the absence of HTH both inhibitors increased the free fatty acid content of the trophocytes, an effect consistent with the known mode of action of these agents. None of the inhibitors tested, all of which blocked HTH activated trehalose synthesis, prevented activation of phosphorylase by HTH. This is taken as evidence that other downstream sites are also important in the regulation of trehalose production by the fat body. It is suggested that the increase in free fatty acids evoked by HTH, or metabolites of those fatty acids, may regulate the synthesis and release of trehalose from the trophocytes because of potential effects on trehalose phosphate synthase, trehalose 6-phosphate phosphatase, and the trehalose trans- port mechanism in the trophocyte membrane. 1997 Elsevier Science Ltd Fat body Cockroach Phospholipase A 2 Cyclooxygenase Hypertrehalosemic hormone Trehalose Fatty acids INTRODUCTION Two hypertrehalosemic hormones (HTH-I and HTH-II) occur in the cockroach corpus cardiacum, from which they are released to the hemolymph and target the fat body to stimulate the synthesis of trehalose. A key step in the synthesis of trehalose is the activation of glycogen phosphorylase (EC 2.4.1.1) to catalyse the release of glu- cosyl residues from glycogen, which then serve as the substrate for synthesis of the sugar (Steele, 1985). That the increased rate of trehalose synthesis is not solely dependent on mass action, as might be expected if phos- *Author for correspondence. Tel: + 1 519 661 3136; Fax: + 1 519 661 2014. ²Department of Zoology, The University of Western Ontario, London, Ontario, Canada N6A 5B7 681 phorylase were the only site of regulation, is apparent from the following observations. Treatment of the cock- roach fat body with cyclic AMP, methyl xanthines and the calcium ionophore A23187 all produce strong acti- vation of phosphorylase, often to levels greater than those obtainable with the HTHs (McClure and Steele, 1981), yet none of these agents significantly increases trehalose output from the fat body (McClure and Steele, 1981; Steele et al., 1988). A possible explanation for these fin- dings is that the agonists change the concentration of allosteric effectors in the trophocytes so that the a form of phosphorylase is unable to express its activity (Ercan et al., 1996). An alternative explanation, explored in the present study, is that downstream sites, as well as phos- phorylase, are regulated to control the production of tre- halose by the fat body. Information now coming to light hints at the possi-
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Evidence that free fatty acids in trophocytes of Periplaneta americana fat body may be regulated by...

Insect Biochem. Molec. Biol.Vol. 27, No. 7, pp. 681–692, 1997 1997 Elsevier Science LtdPergamon PII: S0965-1748(97)00046-5 All rights reserved. Printed in Great Britain0965-1748/97 $17.00+ 0.00
Evidence that Free Fatty Acids in Trophocytesof Periplaneta americanaFat Body may beRegulated by the Activity of Phospholipase A2
and CyclooxygenaseIRSHAD ALI†, JOHN E. STEELE*†
Received 14 October 1996; revised and accepted 18 June 1997
Previous studies have shown that palmitic, stearic, oleic and linoleic acid levels in trophocytesprepared from the fat body of male Periplaneta americanaare increased following treatmentof the cells with hypertrehalosemic hormone (HTH). Melittin, an activator of phospholipaseA2, mimicked the action of HTH by increasing the free fatty acid content in a concentration-dependent manner. The increase caused by HTH could be eliminated by pretreatment of thetrophocytes with 1 mM 49-bromophenacyl bromide (BPB), an inhibitor of phospholipase A2.BPB also decreases the concentration of free fatty acids in trophocytes not treated with HTHbut by a smaller margin. Nordihydroguaiaretic acid (NDGA) and indomethacin, inhibitors oflipoxygenase and cyclooxygenase, respectively, eliminated the increase in free fatty acidsevoked by HTH. In the absence of HTH both inhibitors increased the free fatty acid contentof the trophocytes, an effect consistent with the known mode of action of these agents. Noneof the inhibitors tested, all of which blocked HTH activated trehalose synthesis, preventedactivation of phosphorylase by HTH. This is taken as evidence that other downstream sitesare also important in the regulation of trehalose production by the fat body. It is suggestedthat the increase in free fatty acids evoked by HTH, or metabolites of those fatty acids, mayregulate the synthesis and release of trehalose from the trophocytes because of potential effectson trehalose phosphate synthase, trehalose 6-phosphate phosphatase, and the trehalose trans-port mechanism in the trophocyte membrane. 1997 Elsevier Science Ltd
Fat body Cockroach Phospholipase A2 Cyclooxygenase Hypertrehalosemic hormone TrehaloseFatty acids
INTRODUCTION
Two hypertrehalosemic hormones (HTH-I and HTH-II)occur in the cockroach corpus cardiacum, from whichthey are released to the hemolymph and target the fatbody to stimulate the synthesis of trehalose. A key stepin the synthesis of trehalose is the activation of glycogenphosphorylase (EC 2.4.1.1) to catalyse the release of glu-cosyl residues from glycogen, which then serve as thesubstrate for synthesis of the sugar (Steele, 1985). Thatthe increased rate of trehalose synthesis is not solelydependent on mass action, as might be expected if phos-
*Author for correspondence. Tel:+ 1 519 661 3136; Fax:+ 1 519661 2014.
†Department of Zoology, The University of Western Ontario, London,Ontario, Canada N6A 5B7
681
phorylase were the only site of regulation, is apparentfrom the following observations. Treatment of the cock-roach fat body with cyclic AMP, methyl xanthines andthe calcium ionophore A23187 all produce strong acti-vation of phosphorylase, often to levels greater than thoseobtainable with the HTHs (McClure and Steele, 1981),yet none of these agents significantly increases trehaloseoutput from the fat body (McClure and Steele, 1981;Steeleet al., 1988). A possible explanation for these fin-dings is that the agonists change the concentration ofallosteric effectors in the trophocytes so that thea formof phosphorylase is unable to express its activity (Ercanet al., 1996). An alternative explanation, explored in thepresent study, is that downstream sites, as well as phos-phorylase, are regulated to control the production of tre-halose by the fat body.
Information now coming to light hints at the possi-

682 IRSHAD ALI AND JOHN E. STEELE
bility that fatty acids, or their metabolites, may serve asregulators or modulators of trehalose synthesis or of itsrelease from the cell. In support of this view we havesucceeded in showing that both HTH-I and HTH-IIincrease the palmitic, stearic, oleic and linoleic acid con-tent of trophocytes derived from disaggregated fat body(Ali and Steele, 1997). We have also shown that theefflux of trehalose from the intact fat body in responseto an HTH challenge is blocked by inhibitors of cycloox-ygenase (EC 1.14.99.1) and phospholipase A2 (EC3.1.1.4) (Ali et al., 1997). The possibility that there is arelationship between the activity of cyclooxygenase,phospholipase A2 and the concentration of free fattyacids in the trophocytes does not appear to have beeninvestigated. In this study we provide evidence that arestraining effect of cyclooxygenase and phospholipaseA2 inhibitors on HTH activated trehalose efflux is posi-tively correlated with a failure of the hormones to elevatefree fatty acid levels in the trophocytes.
MATERIALS AND METHODS
Animals
Periplaneta americanawere reared in an insectary at30 ± 2°C with the relative humidity maintained atapproximately 60%. The cockroaches were fed pulver-ized Purina Dog Chow containing 10% (w/w) sucrose.Drinking water was available at all times because at thetemperature of the rearing room the cockroaches rapidlybecame desiccated when water was not present. Onlyadult males 4–6 weeks of age were used in the study.The cockroaches were neck-ligated in late afternoon foruse the following morning. During this period they wereheld in a container in which the relative humidity wasmaintained near saturation to prevent desiccation.
Chemicals
Both the inorganic and organic chemicals used in thisstudy were of reagent grade or better and were obtainedfrom various laboratory suppliers. Solvents wereobtained from British Drug Houses, Toronto, Ontario,Canada and Fisher Scientific Co., Toronto, Ontario, Can-ada. a,a,a,-Trifluoro-m-toluidine, methyl iodide, silveroxide and methyl propionate were purchased from Ald-rich Chemical Co., Milwaukee, WI, U.S.A. Fatty acidsand fatty acid methyl esters were obtained from SigmaChemical Co., St Louis, MO, U.S.A. and BIOMOLResearch Laboratories, Plymouth Meeting, PA, U.S.A.Collagenase A was purchased from BoehringerMannheim Canada, Laval, PQ, Canada. The drugs 49-bromophenacyl bromide (BPB), indomethacin, melittinand nordihydroguaiaretic acid (NDGA) were obtainedfrom Sigma Chemical Co., and the synthetic hormonesHTH-I and HTH-II from Peninsula Laboratories,Belmont, CA, U.S.A. HTH-I and HTH-II were preparedas 1 mM stock solutions in 50% ethanol. Excepting mel-ittin, the drugs were prepared as stock solutions dissolved
in dimethyl sulfoxide: 95% ethanol (50:50, v/v). Melittinwas dissolved in dimethyl sulfoxide. The final concen-tration of carrier solution in the incubation medium didnot exceed 0.5%. An equivalent amount of carrier wasincluded in the medium containing the control tissue.
Physiological saline
The medium used for the disaggregation of the fat bod-ies and the incubation of the trophocytes contained215 mM NaCl, 4.8 mM KCl, 1 mM CaCl2, 1 mM MgCl2,5 mM glucose, 40 mM trehalose and 40 mM HEPESbuffer adjusted to pH 7.4 with 1 M NaOH. The salinewas aerated with filtered air before use.
Preparation of trophocytes
Collagenase was used to disaggregate the fat bodyaccording to the method of Steele and Ireland (1994),using the saline described above instead of that describedin the original study. Generally, eight fat bodies wereused to prepare a batch of cells. The cell yield wasdetermined with a hemocytometer.
Incubation of trophocytes
Normally, 50–100× 103 trophocytes in 1 ml of salinewere pipetted into a 40 ml polypropylene centrifuge tubeand preincubated at 30°C for 30 min in a shaker waterbath. Hormones or other agents were then added and themixture was incubated for an additional 15 min. Thereaction was stopped by pipetting the total reaction mix-ture into a 10 ml plastic syringe barrel fitted with a 20gauge needle and a snug-fitting disc of glass microfibrefilter (Whatman GF/A) at the bottom of the syringe toretain the trophocytes. The needle was passed through arubber stopper fitted into the neck of a 250 ml filter flask.Mild vacuum was used to draw the extracellular mediuminto the flask and away from the cells. The filter contain-ing the cells was quickly removed from the syringe andtransferred to a centrifuge tube containing 1.5 ml of icecold methanol which was then flushed with N2 and sealedwith a teflon-lined screw cap. After thorough mixing thesample was allowed to stand for 20 min at 4°C and wasthen centrifuged at 450g for 10 min. The supernatant wasremoved for analysis of the fatty acid content asdescribed below.
Measurement of fatty acids
Free fatty acids were determined by gas chromatogra-phy using “on column” methylation with trimethyl-(a,a,a-trifluoro-m-tolyl) ammonium hydroxide(TMTFTH) as described by MacGee and Allen (1974).The sample in 1.5 ml of methanol was transferred to aglass stoppered centrifuge tube followed by 5 ml of hex-ane and the mixture was shaken well. After acidificationwith 0.5 ml of 1 M phosphoric acid to drive the fattyacids into the hexane phase the sample was centrifugedat 450g for 2 min. The lower methanol phase wasremoved with a syringe and the hexane phase washedtwice with 2 ml aliquots of 0.1 M phosphoric acid. The

683REGULATION OF FATTY ACIDS IN P. AMERICANA
complete removal of the lower aqueous phase is essentialfor efficient recovery of the fatty acids.
The methylated fatty acids were analysed on a Hew-lett–Packard 5711A gas chromatograph with a flame ion-ization detector and a J and W DB-Wax capillary column(30 m × 0.53 mm i.d.). The splitless injection port andflame ionization detector were maintained at 250°C, andthe oven programmed to operate from 190–230°C at arate of 8°C/min. Helium at 5 ml/min was used as thecarrier gas. Data were recorded with a Hewlett–Packard3380A integrator. Recovery of palmitic, stearic, oleic andlinoleic acids was determined to be 95, 86, 85, and90%, respectively.
Activation of phosphorylase
For the assay, dispersed trophocytes (| 30,000) in250ml of physiological saline were preincubated for30 min at 30°C. Inhibitors, when required, were addedduring preincubation of the cells, 15 min prior to com-pletion of the preincubation period and before additionof the hormone. The effect of HTH on phosphorylaseactivation was routinely determined by incubating thecells with the hormone for 5 min. HTH was addedimmediately after completion of the preincubation per-iod. The reactions were terminated by the addition of750ml of ice-cold 0.1 M sodium fluoride containing0.01 M EDTA followed immediately by sonication of thecells. The samples were then centrifuged at 14,000g.
Glycogen phosphorylase was assayed in the directionof glycogen phosphorolysis using the spectrophotometricassay described by Childress and Sacktor (1970) andmodified slightly by Ziegleret al. (1979). Glycogen, usedas the substrate, was treated with charcoal to remove anycontaminating AMP (Stevenson and Wyatt, 1964). Thereaction mixture was prepared as a concentrated solutionand the pH titrated to 7.4 with 0.1 M NaOH. Afterdilution to 60% of the final volume the solution wasstored at− 20°C. NADP and glucose 1,6-bisphosphatewere prepared fresh in distilled water as 11.4 mM and24 mM solutions, respectively. Phosphorylase activity inthe supernatant solution was determined in a final volumeof 1 ml comprising 600ml of substrate mixture, 50ml each of the NADP and glucose 1,6-bisphosphate sol-utions and 200ml of the sample to be assayed. The reac-tion was started by the addition of 100ml of solutioncontaining the measuring enzymes. After determining thebasal level of phosphorylase activity, the total activity ofthe phosphorylase was estimated by adding 50ml of40 mM 59-AMP to the reaction mixture and redetermin-ing the reaction velocity.
Preparation of trophocyte membranes
The preparation of the trophocyte membranes wasbased on that described by Giudicelliet al. (1982). Tro-phocytes were homogenized with a hand-held homogen-izer (Omni 2000, fitted with 5 mm generator) for 10 s in50 mM glycine-NaOH buffer (pH 9) using freshly dis-persed cells. The homogenate was centrifuged at 20,000g
for 15 min at 4°C. The floating fat cake and the infranat-ant was discarded and the pellet was resuspended in 2 mlof buffer followed by centrifugation at 20,000g for20 min. The pellet was resuspended in fresh buffer and,after centrifugation for 10 min, the final pellet was sus-pended in 1 ml of buffer. Protein was determined accord-ing to the method of Lowryet al. (1951).
Statistical analysis
The significance of differences was tested usingt-tests,analysis of variance and a multiple comparison test(Duncan) as described by Zar (1984).t-tests for unpairedand paired data were used to analyse the phosphorylasedata and fatty acid measurements, respectively.
RESULTS
Both HTH-I and HTH-II, not unexpectedly, havestrong activating effects on the phosphorylase activity ofdispersed trophocytes as shown by the shift from theinactiveb form of the enzyme to the activea form (Table1). This finding is in keeping with the view that phos-phorylase catalyses the rate limiting reaction in trehalosesynthesis. Because a previous study (Aliet al., 1997) hadshown that various inhibitors of phospholipase A2 andcyclooxygenase blocked the increase in trehalose nor-mally produced by the HTHs, it was important to deter-mine whether these agents might cause this effect byblocking the phosphorylase activation reaction. Such aneffect would weaken the argument that changes in othermetabolites are linked to the modulation of trehalose syn-thesis. The data (Table 1) show that none of the agentstested inhibited phosphorylase activation. The data alsoshow that indomethacin and NDGA caused moderate andweak activation of phosphorylase, respectively, whenpresent without HTH, although not to the extent causedby either of the hormones. Furthermore, both inhibitorsenhanced the activating effect of the hormones on trehal-ose efflux from the trophocytes. In contrast to theseresults BPB had no effect on phosphorylase.
Although arachidonic acid could not be detected usingthe methods employed in this study, the synthesis of ara-chidonic acid from linoleic acid and stimulation of thereaction by HTH has been demonstrated in trophocytes(Ali and Steele, 1998). This suggests that other fattyacids may also be potential precursors for the synthesisof arachidonic acid. Because the metabolism of arachi-donic acid is a specific function of cyclooxygenase, itwas of interest to test the possibility that a decrease inthe activity of cyclooxygenase activity might cause anincrease in the concentration of free fatty acids in thetrophocytes. To do this the cells were incubated withindomethacin to lower the activity of the enzyme. Treat-ing the unstimulated trophocytes with the inhibitorcaused a marked increase in the concentration of each ofthe four fatty acids of interest (Fig. 1). The data suggestthat the lowest concentration of indomethacin used(10 mM) was only marginally less effective than the

684 IRSHAD ALI AND JOHN E. STEELE
TABLE 1. The effect of inhibitors of lipoxygenase, cyclooxygenase and phospholipase A2 on the basal phosphorylase activity* of trophocytesand its activation by HTH
Treatment BPB† INDO† NDGA†
Control 29.1± 1.6 (11) 28.5± 1.5 (13) 31.3± 1.0 (12)Inhibitor (I) 33.3 ± 2.3 (6) 52.9± 4.5 (3) 42.4± 1.8 (6)HTH-I 68.9 ± 2.3 (5) 67.2± 2.3 (5) 67.2± 2.3 (5)HTH-I + I 75.2 ± 3.7 (3) 75.6± 1.8 (3) 77.2± 5.8 (3)HTH-II 64.9 ± 1.0 (4) 63.6± 1.2 (4) 69.8± 1.4 (4)HTH-II + I 59.1 ± 2.6 (3) 64.2± 4.1 (6) 81.1± 2.4 (3)
*The values shown for phosphorylase represent the percentage of enzyme in the active (a) form and are expressed as the mean± the standarderror of the mean. Sample size is shown by the bracketed figure. The experimental procedure and determination of phosphorylase activity aredescribed in Materials and methods. The concentration of the inhibitors was 0.1 mM and HTH 40 nM. BPB did not significantly affect phos-phorylase activity. INDO caused a significant increase in the amount of phosphorylasea compared to the controls (P , 0.001), and augmentedthe stimulatory effect of HTH-I on the enzyme. NDGA increased the activity of phosphorylase relative to the controls (P , 0.001) and alsothat of trophocytes challenged with HTH-II (P , 0.005).†Abbreviations used are: BPB, 49-bromophenacyl bromide; INDO, indomethacin; NDGA, nordihydroguaiaretic acid.
FIGURE 1. Data showing that indomethacin increases the level of free fatty acids in dispersed trophocytes. After preincubationfor 30 min at 30°C the trophocytes (| 60,000) in 1 ml of saline were incubated with the indicated concentration of indomethacinfor 15 min, and the free fatty acid content of the trophocytes was then determined as described in Materials and methods.Indomethacin, with one exception (linoleic acid: 10−5 M indomethacin), increased the tissue content of palmitic (P , 0.01),stearic (P , 0.05), oleic (P , 0.01) and linoleic acid (P , 0.05) at all concentrations tested. For all samplesn = 4. Abbrevi-
ations: IND, indomethacin; CON, control.

685REGULATION OF FATTY ACIDS IN P. AMERICANA
highest (1 mM). In common with indomethacin, HTH-IIincreased the level of free fatty acids in the trophocytesby a comparable factor (Fig. 2). However, pretreatmentof the trophocytes with indomethacin followed by a chal-lenge with HTH-II resulted in free fatty acid levels whichwere lower than those that occurred after treatment withthe hormone alone (Fig. 2). Because arachidonic acid,the substrate for the cyclooxygenase reaction, can also bemetabolized by lipoxygenase it was of interest to knowwhether inhibitors of the latter enzyme also increasedfree fatty acid levels to a similar extent as did indome-thacin. NDGA, a potent inhibitor of lipoxygenase in cer-tain mammalian tissues, increased the level of free fattyacids in the trophocytes (Fig. 3), but appeared to do soin a somewhat different manner than did indomethacin.The stimulatory effect of NDGA on the free fatty acidconcentration appeared to occur only within a narrow
FIGURE 2. Data showing that indomethacin blocks the increase in free fatty acids normally evoked by HTH-II. Trophocytes( | 60,000) in 1 ml of saline were allowed to equilibrate for 30 min at 30°C. Indomethacin was then added to three of thesamples to yield the concentrations shown, and the incubation of all samples continued for an additional 15 min. At this timeHTH-II (final concentration 100 pmol/ml) was added to each of the indomethacin-treated samples as well as one of the untreatedsamples. Incubation of all samples was continued for 15 min before stopping the reaction for the determination of the freefatty acid content. The increase in each fatty acid caused by the action of HTH-II was significantly decreased by 0.01 mM
indomethacin (P , 0.05 for each fatty acid,n = 5). Abbreviations as in Fig. 1.
range of inhibitor concentrations in the vicinity of0.1 mM, a 10-fold higher concentration having no effect.The inhibitor blocked the increase in free fatty acids pro-duced by HTH-1 (Fig. 4) with the result that the concen-tration of each of the four fatty acids did not rise abovethe level found in the control trophocytes. This effect iscomparable to that of indomethacin on free fatty acids inHTH-II-treated trophocytes. The low value obtained forlinoleic acid with the lowest concentration of NDGAtested appears to be aberrant.
Although lipoxygenase and cyclooxygenase have thepotential to modulate the concentration of intracellularfree fatty acids by regulating their rate of depletion, theactivity of phospholipase A2 may also be important inthe regulation of intracellular free fatty acids, because ofits role in supplying the cell with free fatty acids aspotential precursors for the synthesis of arachidonic acid.
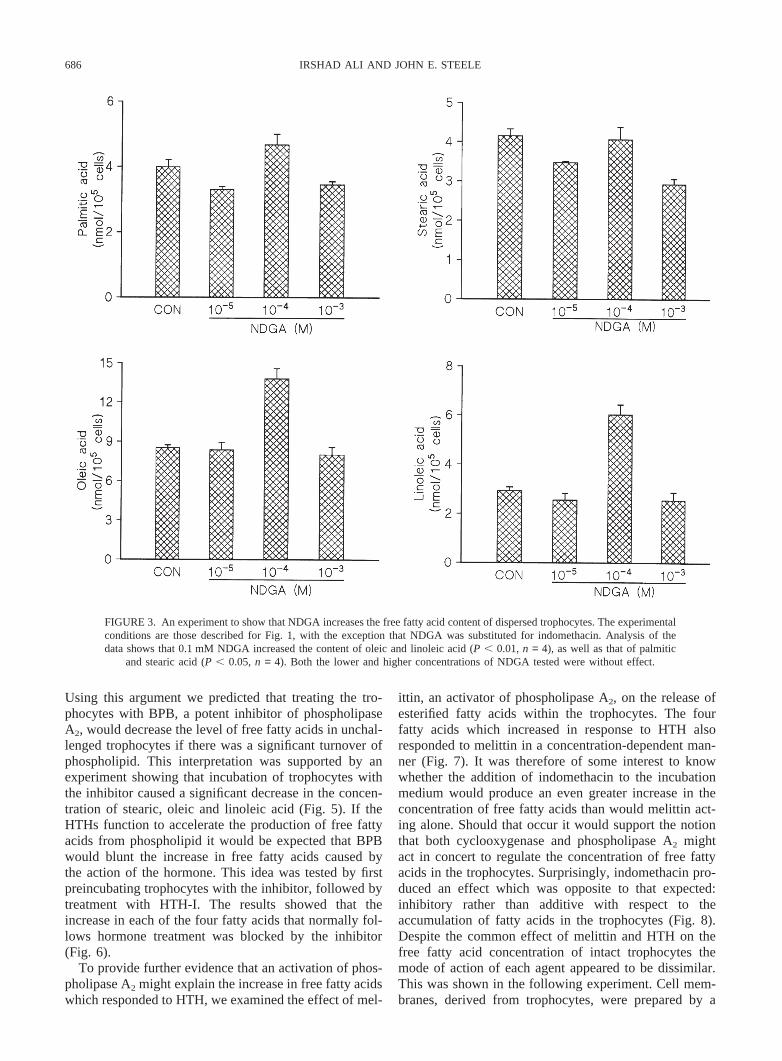
686 IRSHAD ALI AND JOHN E. STEELE
FIGURE 3. An experiment to show that NDGA increases the free fatty acid content of dispersed trophocytes. The experimentalconditions are those described for Fig. 1, with the exception that NDGA was substituted for indomethacin. Analysis of thedata shows that 0.1 mM NDGA increased the content of oleic and linoleic acid (P , 0.01,n = 4), as well as that of palmitic
and stearic acid (P , 0.05,n = 4). Both the lower and higher concentrations of NDGA tested were without effect.
Using this argument we predicted that treating the tro-phocytes with BPB, a potent inhibitor of phospholipaseA2, would decrease the level of free fatty acids in unchal-lenged trophocytes if there was a significant turnover ofphospholipid. This interpretation was supported by anexperiment showing that incubation of trophocytes withthe inhibitor caused a significant decrease in the concen-tration of stearic, oleic and linoleic acid (Fig. 5). If theHTHs function to accelerate the production of free fattyacids from phospholipid it would be expected that BPBwould blunt the increase in free fatty acids caused bythe action of the hormone. This idea was tested by firstpreincubating trophocytes with the inhibitor, followed bytreatment with HTH-I. The results showed that theincrease in each of the four fatty acids that normally fol-lows hormone treatment was blocked by the inhibitor(Fig. 6).
To provide further evidence that an activation of phos-pholipase A2 might explain the increase in free fatty acidswhich responded to HTH, we examined the effect of mel-
ittin, an activator of phospholipase A2, on the release ofesterified fatty acids within the trophocytes. The fourfatty acids which increased in response to HTH alsoresponded to melittin in a concentration-dependent man-ner (Fig. 7). It was therefore of some interest to knowwhether the addition of indomethacin to the incubationmedium would produce an even greater increase in theconcentration of free fatty acids than would melittin act-ing alone. Should that occur it would support the notionthat both cyclooxygenase and phospholipase A2 mightact in concert to regulate the concentration of free fattyacids in the trophocytes. Surprisingly, indomethacin pro-duced an effect which was opposite to that expected:inhibitory rather than additive with respect to theaccumulation of fatty acids in the trophocytes (Fig. 8).Despite the common effect of melittin and HTH on thefree fatty acid concentration of intact trophocytes themode of action of each agent appeared to be dissimilar.This was shown in the following experiment. Cell mem-branes, derived from trophocytes, were prepared by a
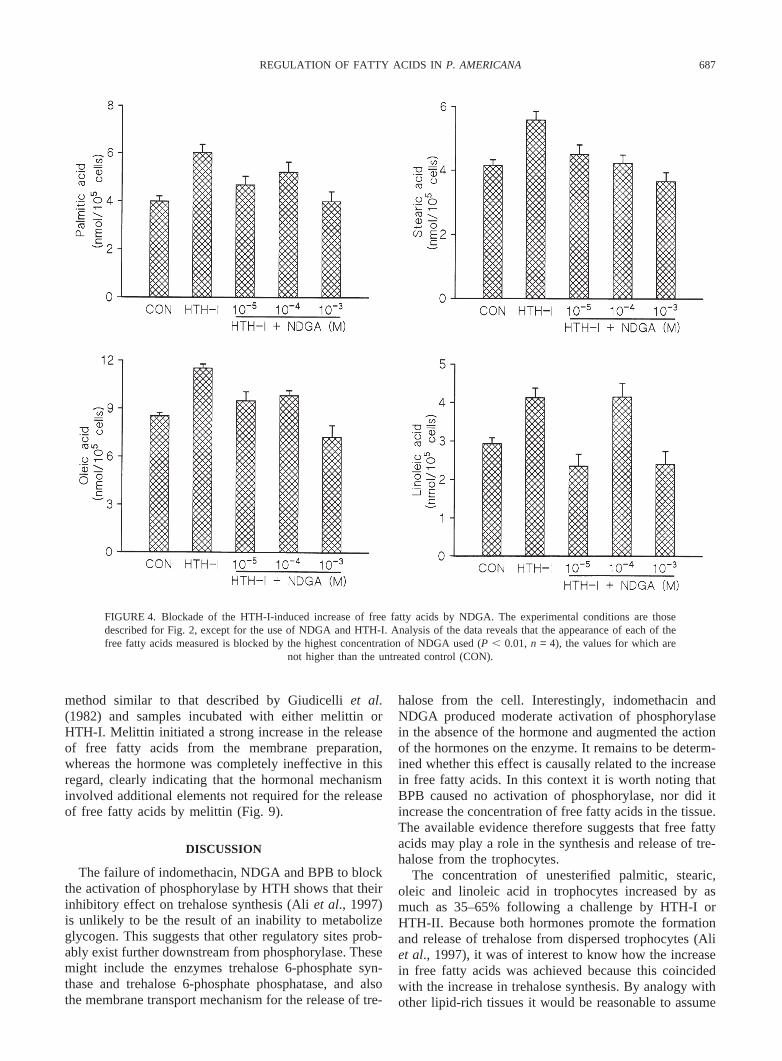
687REGULATION OF FATTY ACIDS IN P. AMERICANA
FIGURE 4. Blockade of the HTH-I-induced increase of free fatty acids by NDGA. The experimental conditions are thosedescribed for Fig. 2, except for the use of NDGA and HTH-I. Analysis of the data reveals that the appearance of each of thefree fatty acids measured is blocked by the highest concentration of NDGA used (P , 0.01,n = 4), the values for which are
not higher than the untreated control (CON).
method similar to that described by Giudicelliet al.(1982) and samples incubated with either melittin orHTH-I. Melittin initiated a strong increase in the releaseof free fatty acids from the membrane preparation,whereas the hormone was completely ineffective in thisregard, clearly indicating that the hormonal mechanisminvolved additional elements not required for the releaseof free fatty acids by melittin (Fig. 9).
DISCUSSION
The failure of indomethacin, NDGA and BPB to blockthe activation of phosphorylase by HTH shows that theirinhibitory effect on trehalose synthesis (Aliet al., 1997)is unlikely to be the result of an inability to metabolizeglycogen. This suggests that other regulatory sites prob-ably exist further downstream from phosphorylase. Thesemight include the enzymes trehalose 6-phosphate syn-thase and trehalose 6-phosphate phosphatase, and alsothe membrane transport mechanism for the release of tre-
halose from the cell. Interestingly, indomethacin andNDGA produced moderate activation of phosphorylasein the absence of the hormone and augmented the actionof the hormones on the enzyme. It remains to be determ-ined whether this effect is causally related to the increasein free fatty acids. In this context it is worth noting thatBPB caused no activation of phosphorylase, nor did itincrease the concentration of free fatty acids in the tissue.The available evidence therefore suggests that free fattyacids may play a role in the synthesis and release of tre-halose from the trophocytes.
The concentration of unesterified palmitic, stearic,oleic and linoleic acid in trophocytes increased by asmuch as 35–65% following a challenge by HTH-I orHTH-II. Because both hormones promote the formationand release of trehalose from dispersed trophocytes (Aliet al., 1997), it was of interest to know how the increasein free fatty acids was achieved because this coincidedwith the increase in trehalose synthesis. By analogy withother lipid-rich tissues it would be reasonable to assume
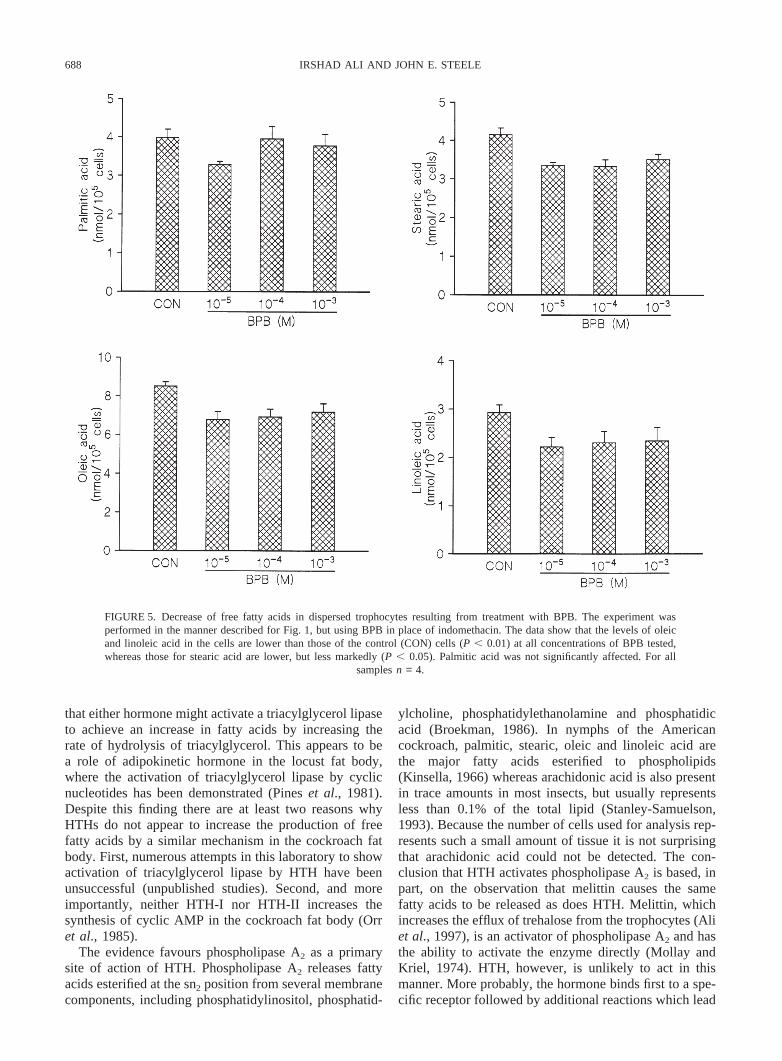
688 IRSHAD ALI AND JOHN E. STEELE
FIGURE 5. Decrease of free fatty acids in dispersed trophocytes resulting from treatment with BPB. The experiment wasperformed in the manner described for Fig. 1, but using BPB in place of indomethacin. The data show that the levels of oleicand linoleic acid in the cells are lower than those of the control (CON) cells (P , 0.01) at all concentrations of BPB tested,whereas those for stearic acid are lower, but less markedly (P , 0.05). Palmitic acid was not significantly affected. For all
samplesn = 4.
that either hormone might activate a triacylglycerol lipaseto achieve an increase in fatty acids by increasing therate of hydrolysis of triacylglycerol. This appears to bea role of adipokinetic hormone in the locust fat body,where the activation of triacylglycerol lipase by cyclicnucleotides has been demonstrated (Pineset al., 1981).Despite this finding there are at least two reasons whyHTHs do not appear to increase the production of freefatty acids by a similar mechanism in the cockroach fatbody. First, numerous attempts in this laboratory to showactivation of triacylglycerol lipase by HTH have beenunsuccessful (unpublished studies). Second, and moreimportantly, neither HTH-I nor HTH-II increases thesynthesis of cyclic AMP in the cockroach fat body (Orret al., 1985).
The evidence favours phospholipase A2 as a primarysite of action of HTH. Phospholipase A2 releases fattyacids esterified at the sn2 position from several membranecomponents, including phosphatidylinositol, phosphatid-
ylcholine, phosphatidylethanolamine and phosphatidicacid (Broekman, 1986). In nymphs of the Americancockroach, palmitic, stearic, oleic and linoleic acid arethe major fatty acids esterified to phospholipids(Kinsella, 1966) whereas arachidonic acid is also presentin trace amounts in most insects, but usually representsless than 0.1% of the total lipid (Stanley-Samuelson,1993). Because the number of cells used for analysis rep-resents such a small amount of tissue it is not surprisingthat arachidonic acid could not be detected. The con-clusion that HTH activates phospholipase A2 is based, inpart, on the observation that melittin causes the samefatty acids to be released as does HTH. Melittin, whichincreases the efflux of trehalose from the trophocytes (Aliet al., 1997), is an activator of phospholipase A2 and hasthe ability to activate the enzyme directly (Mollay andKriel, 1974). HTH, however, is unlikely to act in thismanner. More probably, the hormone binds first to a spe-cific receptor followed by additional reactions which lead

689REGULATION OF FATTY ACIDS IN P. AMERICANA
FIGURE 6. Blockade of the HTH-I-induced increase of free fatty acids by BPB. The experiment was performed as describedfor Fig. 2, but with the substitution of HTH-I for HTH-II and BPB for indomethacin. Analysis of the data shows that BPB,at all concentrations tested, inhibits the HTH-I-evoked increase in palmitic (P , 0.05), stearic (P , 0.05), oleic (P , 0.01)and linoleic acid (P , 0.01). For all samplesn = 4. The action of the inhibitor holds the level of free fatty acids to those
found in the untreated controls (CON).
to the activation of phospholipase A2. That this is a poss-ible mode of action is indicated by the failure of HTHto release fatty acids from trophocyte membranes, aresult which contrasts with the stimulatory action of mel-ittin. Although several examples of receptor-mediatedactivation of phospholipase A2 are known (Sweattet al.,1986; Garciaet al., 1994) there is no substantive evi-dence, as yet, to show that activation of phospholipaseA2 in trophocytes is by means of a receptor-mediatedmechanism. Nevertheless, the model remains an attract-ive possibility.
Further evidence that phospholipase A2 may be a keytarget of HTH is suggested by the observation that BPBprevented the accumulation of free fatty acids in the tro-phocytes. The inhibitor, in the absence of hormone,caused a small decrease in the concentration of the fourfatty acids but in combination with HTH-I was consider-ably more effective in blocking the increase in fatty acidsnormally induced by the action of the hormone. BecauseBPB is a potent inhibitor of phospholipase A2 (Changet
al., 1987) the data are consistent with the view that HTH-I stimulates the release of fatty acids from phospholipid,possibly as a result of activation of phospholipase A2 bythe hormone.
In general, phosphorylase activity and free fatty acidlevels responded similarly to indomethacin and NDGAdespite the fact that the former compound is consideredto be a selective inhibitor of cyclooxygenase and the lat-ter an inhibitor of lipoxygenase. Because arachidonicacid is a substrate for both enzymes it seems likely thateach agent would have a similar effect on free fatty acidaccumulation, but a selective effect on the synthesis ofleukotrienes or prostaglandins. Both drugs block theincrease in HTH-stimulated trehalose synthesis (Alietal., 1997); thus it is questionable whether leukotrienes orprostaglandins might be responsible for this effect. Evi-dence from mammalian studies suggests that prosta-glandins are a more likely candidate. This conclusion isbased on the finding that NDGA is a potent inhibitor ofcyclooxygenase as well as lipoxygenase, whereas indo-
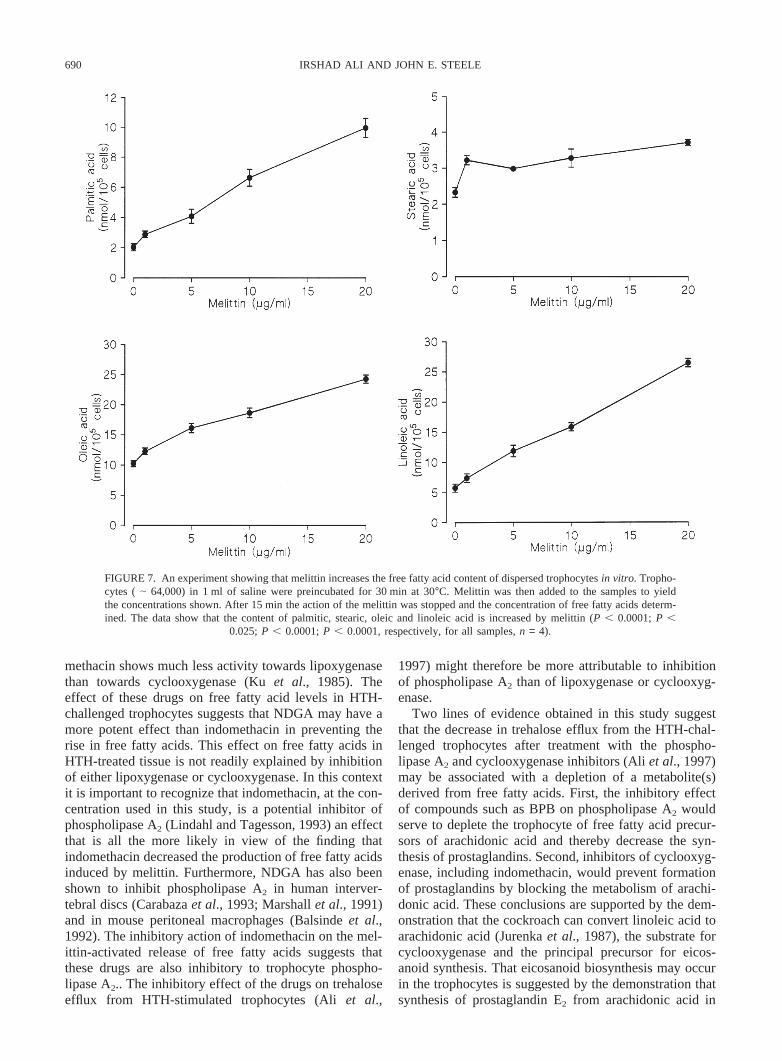
690 IRSHAD ALI AND JOHN E. STEELE
FIGURE 7. An experiment showing that melittin increases the free fatty acid content of dispersed trophocytesin vitro. Tropho-cytes (| 64,000) in 1 ml of saline were preincubated for 30 min at 30°C. Melittin was then added to the samples to yieldthe concentrations shown. After 15 min the action of the melittin was stopped and the concentration of free fatty acids determ-ined. The data show that the content of palmitic, stearic, oleic and linoleic acid is increased by melittin (P , 0.0001;P ,
0.025;P , 0.0001;P , 0.0001, respectively, for all samples,n = 4).
methacin shows much less activity towards lipoxygenasethan towards cyclooxygenase (Kuet al., 1985). Theeffect of these drugs on free fatty acid levels in HTH-challenged trophocytes suggests that NDGA may have amore potent effect than indomethacin in preventing therise in free fatty acids. This effect on free fatty acids inHTH-treated tissue is not readily explained by inhibitionof either lipoxygenase or cyclooxygenase. In this contextit is important to recognize that indomethacin, at the con-centration used in this study, is a potential inhibitor ofphospholipase A2 (Lindahl and Tagesson, 1993) an effectthat is all the more likely in view of the finding thatindomethacin decreased the production of free fatty acidsinduced by melittin. Furthermore, NDGA has also beenshown to inhibit phospholipase A2 in human interver-tebral discs (Carabazaet al., 1993; Marshallet al., 1991)and in mouse peritoneal macrophages (Balsindeet al.,1992). The inhibitory action of indomethacin on the mel-ittin-activated release of free fatty acids suggests thatthese drugs are also inhibitory to trophocyte phospho-lipase A2.. The inhibitory effect of the drugs on trehaloseefflux from HTH-stimulated trophocytes (Aliet al.,
1997) might therefore be more attributable to inhibitionof phospholipase A2 than of lipoxygenase or cyclooxyg-enase.
Two lines of evidence obtained in this study suggestthat the decrease in trehalose efflux from the HTH-chal-lenged trophocytes after treatment with the phospho-lipase A2 and cyclooxygenase inhibitors (Aliet al., 1997)may be associated with a depletion of a metabolite(s)derived from free fatty acids. First, the inhibitory effectof compounds such as BPB on phospholipase A2 wouldserve to deplete the trophocyte of free fatty acid precur-sors of arachidonic acid and thereby decrease the syn-thesis of prostaglandins. Second, inhibitors of cyclooxyg-enase, including indomethacin, would prevent formationof prostaglandins by blocking the metabolism of arachi-donic acid. These conclusions are supported by the dem-onstration that the cockroach can convert linoleic acid toarachidonic acid (Jurenkaet al., 1987), the substrate forcyclooxygenase and the principal precursor for eicos-anoid synthesis. That eicosanoid biosynthesis may occurin the trophocytes is suggested by the demonstration thatsynthesis of prostaglandin E2 from arachidonic acid in
691REGULATION OF FATTY ACIDS IN P. AMERICANA
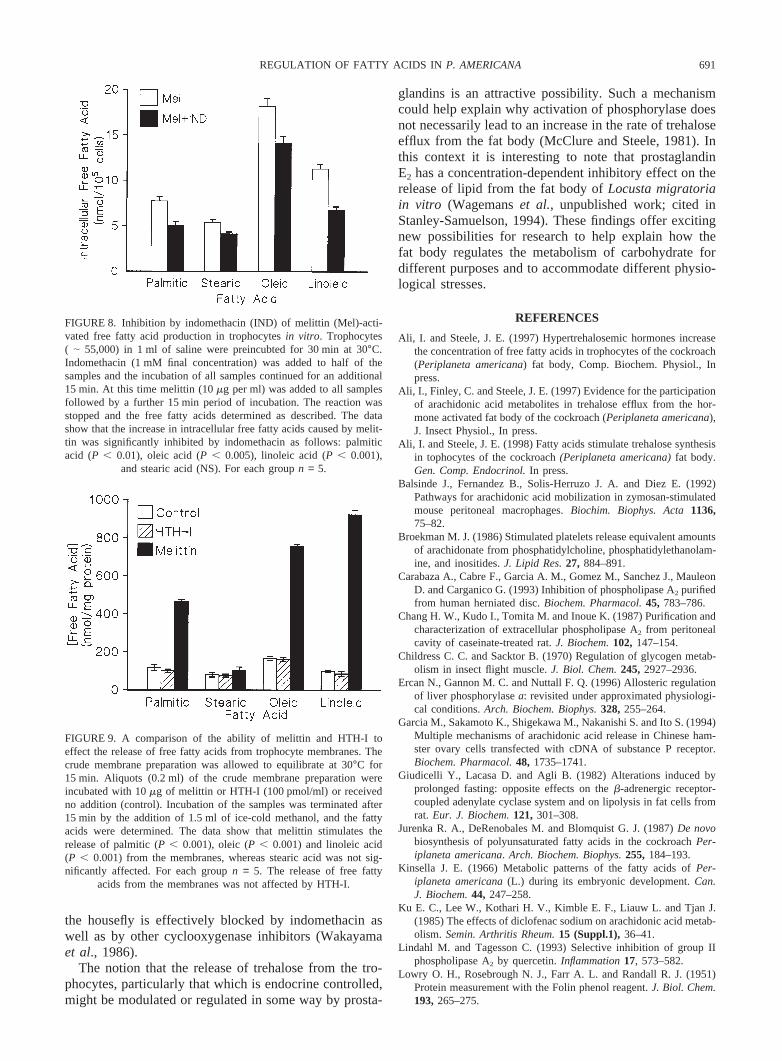
FIGURE 8. Inhibition by indomethacin (IND) of melittin (Mel)-acti-vated free fatty acid production in trophocytesin vitro. Trophocytes( | 55,000) in 1 ml of saline were preincubted for 30 min at 30°C.Indomethacin (1 mM final concentration) was added to half of thesamples and the incubation of all samples continued for an additional15 min. At this time melittin (10mg per ml) was added to all samplesfollowed by a further 15 min period of incubation. The reaction wasstopped and the free fatty acids determined as described. The datashow that the increase in intracellular free fatty acids caused by melit-tin was significantly inhibited by indomethacin as follows: palmiticacid (P , 0.01), oleic acid (P , 0.005), linoleic acid (P , 0.001),
and stearic acid (NS). For each groupn = 5.
FIGURE 9. A comparison of the ability of melittin and HTH-I toeffect the release of free fatty acids from trophocyte membranes. Thecrude membrane preparation was allowed to equilibrate at 30°C for15 min. Aliquots (0.2 ml) of the crude membrane preparation wereincubated with 10mg of melittin or HTH-I (100 pmol/ml) or receivedno addition (control). Incubation of the samples was terminated after15 min by the addition of 1.5 ml of ice-cold methanol, and the fattyacids were determined. The data show that melittin stimulates therelease of palmitic (P , 0.001), oleic (P , 0.001) and linoleic acid(P , 0.001) from the membranes, whereas stearic acid was not sig-nificantly affected. For each groupn = 5. The release of free fatty
acids from the membranes was not affected by HTH-I.
the housefly is effectively blocked by indomethacin aswell as by other cyclooxygenase inhibitors (Wakayamaet al., 1986).
The notion that the release of trehalose from the tro-phocytes, particularly that which is endocrine controlled,might be modulated or regulated in some way by prosta-
glandins is an attractive possibility. Such a mechanismcould help explain why activation of phosphorylase doesnot necessarily lead to an increase in the rate of trehaloseefflux from the fat body (McClure and Steele, 1981). Inthis context it is interesting to note that prostaglandinE2 has a concentration-dependent inhibitory effect on therelease of lipid from the fat body ofLocusta migratoriain vitro (Wagemanset al., unpublished work; cited inStanley-Samuelson, 1994). These findings offer excitingnew possibilities for research to help explain how thefat body regulates the metabolism of carbohydrate fordifferent purposes and to accommodate different physio-logical stresses.
REFERENCES
Ali, I. and Steele, J. E. (1997) Hypertrehalosemic hormones increasethe concentration of free fatty acids in trophocytes of the cockroach(Periplaneta americana) fat body, Comp. Biochem. Physiol., Inpress.
Ali, I., Finley, C. and Steele, J. E. (1997) Evidence for the participationof arachidonic acid metabolites in trehalose efflux from the hor-mone activated fat body of the cockroach (Periplaneta americana),J. Insect Physiol., In press.
Ali, I. and Steele, J. E. (1998) Fatty acids stimulate trehalose synthesisin tophocytes of the cockroach(Periplaneta americana)fat body.Gen. Comp. Endocrinol.In press.
Balsinde J., Fernandez B., Solis-Herruzo J. A. and Diez E. (1992)Pathways for arachidonic acid mobilization in zymosan-stimulatedmouse peritoneal macrophages.Biochim. Biophys. Acta1136,75–82.
Broekman M. J. (1986) Stimulated platelets release equivalent amountsof arachidonate from phosphatidylcholine, phosphatidylethanolam-ine, and inositides.J. Lipid Res.27, 884–891.
Carabaza A., Cabre F., Garcia A. M., Gomez M., Sanchez J., MauleonD. and Carganico G. (1993) Inhibition of phospholipase A2 purifiedfrom human herniated disc.Biochem. Pharmacol.45, 783–786.
Chang H. W., Kudo I., Tomita M. and Inoue K. (1987) Purification andcharacterization of extracellular phospholipase A2 from peritonealcavity of caseinate-treated rat.J. Biochem.102, 147–154.
Childress C. C. and Sacktor B. (1970) Regulation of glycogen metab-olism in insect flight muscle.J. Biol. Chem.245, 2927–2936.
Ercan N., Gannon M. C. and Nuttall F. Q. (1996) Allosteric regulationof liver phosphorylasea: revisited under approximated physiologi-cal conditions.Arch. Biochem. Biophys.328, 255–264.
Garcia M., Sakamoto K., Shigekawa M., Nakanishi S. and Ito S. (1994)Multiple mechanisms of arachidonic acid release in Chinese ham-ster ovary cells transfected with cDNA of substance P receptor.Biochem. Pharmacol.48, 1735–1741.
Giudicelli Y., Lacasa D. and Agli B. (1982) Alterations induced byprolonged fasting: opposite effects on theb-adrenergic receptor-coupled adenylate cyclase system and on lipolysis in fat cells fromrat. Eur. J. Biochem.121, 301–308.
Jurenka R. A., DeRenobales M. and Blomquist G. J. (1987)De novobiosynthesis of polyunsaturated fatty acids in the cockroachPer-iplaneta americana. Arch. Biochem. Biophys.255, 184–193.
Kinsella J. E. (1966) Metabolic patterns of the fatty acids ofPer-iplaneta americana(L.) during its embryonic development.Can.J. Biochem.44, 247–258.
Ku E. C., Lee W., Kothari H. V., Kimble E. F., Liauw L. and Tjan J.(1985) The effects of diclofenac sodium on arachidonic acid metab-olism. Semin. Arthritis Rheum.15 (Suppl.1),36–41.
Lindahl M. and Tagesson C. (1993) Selective inhibition of group IIphospholipase A2 by quercetin.Inflammation17, 573–582.
Lowry O. H., Rosebrough N. J., Farr A. L. and Randall R. J. (1951)Protein measurement with the Folin phenol reagent.J. Biol. Chem.193, 265–275.

692 IRSHAD ALI AND JOHN E. STEELE
MacGee J. and Allen K. G. (1974) Preparation of methyl esters fromthe saponifiable fatty acids in small biological specimens for gas–liquid chromatographic analysis.J. Chromatogr.100, 35–42.
Marshall L. A., Bauer J., Sung M. L. and Chang J. Y. (1991) Evalu-ation of antirheumatic drugs for their effectin vitro on purifiedhuman synovial fluid phospholipase A2. J. Rheumatol.18, 59–65.
McClure J. B. and Steele J. E. (1981) The role of extracellular calciumin hormonal activation of glycogen phosphorylase in cockroach fatbody. Insect Biochem.11, 605–613.
Mollay C. and Kreil G. (1974) Enhancement of bee venom phospho-lipase A2 activity by melittin, direct lytic factor from cobra venomand polymyxin B.FEBS Lett.46, 141–144.
Orr G. L., Gole J. W. D., Jahagirdar A. P., Downer R. G. H. andSteele, J. E. (1985), Cyclic AMP does not mediate the action ofsynthetic hypertrehalosemic peptides from the corpus cardiacum ofPeriplaneta americana. Insect Biochem15, 703–709.
Pines M., Tietz A., Weintraub H., Applebaum S. W. and Josefsson L.(1981) Hormonal activation of protein kinase and lipid mobilizationin the locust fat bodyin vitro. Gen. Comp. Endocrinol.43, 427–431.
Stanley-Samuelson, D. W. (1993) The biological significance ofprostaglandins and related eicosanoids in insects. InInsect Lipids:Chemistry, Biochemistry and Biology, ed D. A. Stanley-Samuelsonand D. R. Nelson, pp. 45–97. University of Nebraska Press, Lin-coln.
Stanley-Samuelson D. W. (1994) Prostaglandins and related eicosano-ids in insects, (ed P. D. Evans).Adv. Insect Physiol.24, 115–212.
Steele, J. E. (1985) Control of metabolic processes. InComprehensiveInsect Physiology Biochemistry and Pharmacology, ed G. A. Ker-kut and L. I. Gilbert, Vol. 8, pp. 99–145. Pergamon Press, Oxford.
Steele J. E. and Ireland R. (1994) The preparation of trophocytes fromdisaggregated fat body of the cockroach (Perilaneta americana).Comp. Biochem. Physiol.107A, 517–522.
Steele J. E., McDougall G. E. and Shadwick R. (1988) Trehalose effluxfrom cockroach fat bodyin vitro: paradoxical effects of the corpuscardiacum and methylxanthines.Insect Biochem.18, 585–590.
Stevenson E. and Wyatt G. R. (1964) Glycogen phosphorylase andits activation in silkmoth fat body.Arch. Biochem. Biophys.108,420–429.
Sweatt J. D., Commolly T., Cragoe E. J. and Linbird L. E. (1986)Evidence that Na+ /H + exchange regulates receptor-mediated phos-pholipase A2 activation in human platelets.J. Biol. Chem.261,8667–8673.
Wakayama E. J., Dillwith J. W. and Blomquist G. J. (1986) Charac-terization of prostaglandin synthesis in the housefly,Musca dom-estica(L.). Insect Biochem.16, 903–909.
Zar, J. H. (1984).Biostatistical Analysis. 2nd edn. Prentice–Hall,Englewood Cliffs, New Jersey.
Ziegler R., Ashida M., Fallon A. M., Wimer L. T., Wyatt S. S. andWyatt G. R. (1979) Regulation of glycogen phosphorylase in fatbody of Cecropia silkmoth pupae.J. Comp. Physiol.131,321–332.
Acknowledgements—This research was supported in part by aResearch Grant from the Natural Sciences and Engineering ResearchCouncil of Canada.





![Bibliotheca americana vetustissima [microform]](https://static.fdokumen.com/doc/165x107/6332978da20c543f370c0e95/bibliotheca-americana-vetustissima-microform.jpg)














