
Evidence of carrier-mediated transport in the penetration of donepezil into the rat brain
19
Evidence of Carrier-Mediated Transport in the Penetration of Donepezil into the Rat Brain MI-HWA KIM, 1 HAN-JOO MAENG, 1 KYUNG-HA YU, 1 KYEONG-RYOON LEE, 1 TAKASHI TSURUO, 2 DAE-DUK KIM, 1 CHANG-KOO SHIM, 1 SUK-JAE CHUNG 1 1 Department of Pharmaceutics, College of Pharmacy, Seoul National University, San 56-1, Shinlim-dong, Kwanak-gu, Seoul 151-742, Korea 2 Graduate School of Pharmaceutical Science, The University of Tokyo, Tokyo, Japan Received 7 April 2009; revised 4 June 2009; accepted 7 July 2009 Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jps.21895 ABSTRACT: The objective of this study was to characterize the mechanism that controls the transport of donepezil into the brain. The apparent brain uptake clearance (CL app,br ) was decreased as a function of the dose of donepezil, suggesting an involvement of a saturable transport process via transporter(s) in the penetration across the blood–brain barrier (BBB). Consistent with in vivo results, the uptake of substrates for organic cation transporters was significantly reduced by donepezil in both MBEC4 cells (i.e., a model for BBB) and HEK 293 cells expressing the transporters found in the brain, indicative of the involvement of organic cation transporters in the transport of the drug. Further- more, donepezil transport was enhanced ( p < 0.01) in HEK 293 cells expressing rOCNT1, rOCTN2, or rCHT1. The CL app,br was reduced up to 52.8% of the control in rats that had been pretreated with choline, while the CL app,br was unaffected with pretreatments with organic cations other than choline, suggesting that choline and donepezil share a common transport mechanism in the penetration across the BBB in vivo. Taken together, these observations suggest that the transport of donepezil across the BBB is mediated by organic cation transporters such as choline transport system(s). ß 2009 Wiley-Liss, Inc. and the American Pharmacists Association J Pharm Sci Keywords: donepezil; MBEC4; in vitro model; BBB; brain uptake clearance; organic cation transporters INTRODUCTION Alzheimer’s disease, a progressive and neuro- degenerative disorder, is characterized by a deficiency in central cholinergic neurotransmis- sion. 1,2 Therefore, cholinesterase inhibitors (e.g., tacrine and donepezil) are likely to enhance the concentration of acetylcholine in the synaptic cleft by stabilizing the neurotransmitter, ultimately leading to increased cholinergic transmission 3 in patients with the neurodegenerative disease. Donepezil hydrochloride, ()-2-(1-benzylpiperi- din-4-yl)methyl]-5,6-dimethoxyindan-1-one monohydrochloride, is a reversible acetylcholines- terase inhibitor that has been reported to have a high potency and selectivity for centrally active cholinesterase. The drug is currently commer- cially available worldwide, and is prescribed for the symptomatic treatment of Alzheimer’s disease in many countries. 4,5 The pharmacokinetic, phar- macodynamic, and clinical characteristics of donepezil have been reported in the literature. 6–9 Based on the physicochemical properties [i.e., molecular weight of 379.5 10 and an n-octanol to water partition coefficient of 3.08–4.11 11 ] of Correspondence to: Suk-Jae Chung (Telephone: 82-2-884- 3170; Fax: 82-2-885-8317; E-mail: [email protected]) Journal of Pharmaceutical Sciences ß 2009 Wiley-Liss, Inc. and the American Pharmacists Association JOURNAL OF PHARMACEUTICAL SCIENCES 2009 1
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Evidence of carrier-mediated transport in the penetration of donepezil into the rat brain

Evidence of Carrier-Mediated Transport in the Penetrationof Donepezil into the Rat Brain
MI-HWA KIM,1 HAN-JOO MAENG,1 KYUNG-HA YU,1 KYEONG-RYOON LEE,1 TAKASHI TSURUO,2 DAE-DUK KIM,1
CHANG-KOO SHIM,1 SUK-JAE CHUNG1
1Department of Pharmaceutics, College of Pharmacy, Seoul National University, San 56-1, Shinlim-dong, Kwanak-gu,Seoul 151-742, Korea
2Graduate School of Pharmaceutical Science, The University of Tokyo, Tokyo, Japan
Received 7 April 2009; revised 4 June 2009; accepted 7 July 2009
Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/jps.21895
Corresponde3170; Fax: 82-2
Journal of Pharm
� 2009 Wiley-Liss
ABSTRACT: The objective of this study was to characterize the mechanism that controlsthe transport of donepezil into the brain. The apparent brain uptake clearance (CLapp,br)was decreased as a function of the dose of donepezil, suggesting an involvement of asaturable transport process via transporter(s) in the penetration across the blood–brainbarrier (BBB). Consistent with in vivo results, the uptake of substrates for organic cationtransporters was significantly reduced by donepezil in both MBEC4 cells (i.e., a modelfor BBB) and HEK 293 cells expressing the transporters found in the brain, indicative ofthe involvement of organic cation transporters in the transport of the drug. Further-more, donepezil transport was enhanced (p< 0.01) in HEK 293 cells expressingrOCNT1, rOCTN2, or rCHT1. The CLapp,br was reduced up to 52.8% of the control inrats that had been pretreated with choline, while the CLapp,br was unaffected withpretreatments with organic cations other than choline, suggesting that choline anddonepezil share a common transport mechanism in the penetration across the BBBin vivo. Taken together, these observations suggest that the transport of donepezilacross the BBB is mediated by organic cation transporters such as choline transportsystem(s). � 2009 Wiley-Liss, Inc. and the American Pharmacists Association J Pharm Sci
Keywords: donepezil; MBEC4; in vitro
model; BBB; brain uptake clearance; organiccation transportersINTRODUCTION
Alzheimer’s disease, a progressive and neuro-degenerative disorder, is characterized by adeficiency in central cholinergic neurotransmis-sion.1,2 Therefore, cholinesterase inhibitors (e.g.,tacrine and donepezil) are likely to enhance theconcentration of acetylcholine in the synaptic cleftby stabilizing the neurotransmitter, ultimatelyleading to increased cholinergic transmission3 in
nce to: Suk-Jae Chung (Telephone: 82-2-884--885-8317; E-mail: [email protected])
aceutical Sciences
, Inc. and the American Pharmacists Association
patients with the neurodegenerative disease.Donepezil hydrochloride, (�)-2-(1-benzylpiperi-din-4-yl)methyl]-5,6-dimethoxyindan-1-onemonohydrochloride, is a reversible acetylcholines-terase inhibitor that has been reported to have ahigh potency and selectivity for centrally activecholinesterase. The drug is currently commer-cially available worldwide, and is prescribed forthe symptomatic treatment of Alzheimer’s diseasein many countries.4,5 The pharmacokinetic, phar-macodynamic, and clinical characteristics ofdonepezil have been reported in the literature.6–9
Based on the physicochemical properties [i.e.,molecular weight of 379.510 and an n-octanol towater partition coefficient of 3.08–4.1111] of
JOURNAL OF PHARMACEUTICAL SCIENCES 2009 1

2 KIM ET AL.
donepezil, its brain permeability by passivediffusion would be expected12 to be high. Despitethis expectation, however, Matsui et al.6 reportedthat the concentration and area under the curve(AUC) of donepezil in the brain were two andseven times higher than those in the plasma.Furthermore, Kang et al.13 recently reported thatthe transport of choline was inhibited by theaddition of a number of organic cations, includingdonepezil, in an in vitro model of the blood–brainbarrier (BBB). These observations are consistentwith the hypothesis that efficient carrier-mediated transport system(s) exist(s) to facilitatethe transport of donepezil across the BBB, inaddition to passive diffusion. Unfortunately,however, the underlying mechanism for thetransport of this cholinesterase inhibitor has notbeen fully delineated.
In the BBB, a variety of transporters, such asglucose transporter 1 (GLUT1), monocarboxylatetransporter 1 (MCT1), organic anion transporters(OATs), organic cation transporters (OCTs), andATP-binding cassette transporters, are expressedto support the movement of endogenous/xenobio-tic compounds into and/or out of the brain.14
Among these, a number of organic cation trans-port systems [e.g., OCT2, OCTN2] are known toplay a role in the BBB in regulating the transportfor tacrine, a cholinesterase inhibitor.15 Mura-kami et al.16 demonstrated that the cholinetransporter (CHT) in the BBB could be an isoformof the OCT family, suggesting that CHT partici-pates in the penetration of organic cations acrossthe BBB. Since donepezil is an organic cation,OCT(s) may be involved in the transport of thecholinesterase inhibitor across the BBB. Further-more, since OCTs are known to have a wide andoverlapping substrate selectivity,15,17 the possi-bility that multiple OCTs participate in the BBBtransport of organic cations (e.g., donepezil)cannot be excluded. However, our current under-standing of this aspect of donepezil transport isincomplete.
The objective of this study, therefore, was tostudy the mechanisms associated with thetransport of donepezil into the brain. Ofparticular interest was the evaluation of thepotential involvement of carrier-mediated trans-port, via OCTs expressed in the brain, in thepenetration of donepezil. We report herein thatthe transport of donepezil across the BBB ismediated by a dose-dependent process, and thatdonepezil/choline apparently shares the trans-port process.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
MATERIALS AND METHODS
Materials
Donepezil hydrochloride was generously sup-plied by the Department of Chemistry in theDaewoong R&D Center (Sungnam, South Korea).Choline, 1-methyl-4-phenylpyridinium (MPP),tetraethylammonium (TEA), carnitine, ergothio-neine, diphenhydramine, cyclosporine A (CsA),digoxin, p-aminohippurate (PAH), benzylpenicillin(BPC), caffeine, Dulbecco’s modified Eagle’s med-ium, Hank’s balanced salt solution, and N-2-hydroxyethylpiperazine-N’-2-ethanesulfonic acid(HEPES) were obtained from Sigma–Aldrich(St. Louis, MO). [3H]choline (specific activity75.7 Ci/mmol), [3H]MPP (specific activity 81 Ci/mmol), [3H]CsA (specific activity 9 Ci/mmol),[3H]digoxin (specific activity 40 Ci/mmol),[3H]PAH (specific activity 3.75 Ci/mmol), and[14C]TEA (specific activity 3.2 mCi/mmol) werepurchased from PerkinElmer Life and AnalyticalSciences (Boston, MA), and [3H]BPC (specificactivity 24 Ci/mmol) was obtained from AmershamLife Science (Buckinghamshire, UK). [3H]ergothio-neine (specific activity 1.1 Ci/mmol) was purchasedfrom Moravek Biochemicals (Brea, CA), and[3H]carnitine (specific activity 85 Ci/mmol), fromAmerican Radiolabeled Chemicals (St. Louis, MO),wasalsousedinthisstudy.Flp-In293cells,modifiedHEK 293 cells capable of site-specific integration ofthe gene of interest, were purchased from Invitro-gen (Carlsbad, CA). MBEC4 cells were previouslygenerated by our research group.18 Solvents wereofHPLC grade, and all other chemicals were ofreagent grade or better. In this study, solvents/chemicals were used without further purification.
Animals
Male Sprague–Dawley (SD) rats (Dae-Han Bio-link, Eumsung, Korea), weighing 270–300 g, wereused in all in vivo experiments and in thecollection of brain samples. Experimental proto-cols involving the animals used in this study werereviewed by the Animal Care and Use Committeeof the College of Pharmacy, Seoul NationalUniversity, according to the National Institutesof Health guidelines (National Institutes ofHealth Publication Number 85-23, revised 1985)in Principles of Laboratory Animal Care.
In Vivo Brain Uptake Study for Donepezil
Male SD rats (270–300 g) were anesthetized byan intramuscular administration of ketamine
DOI 10.1002/jps

MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 3
(Yuhan Ketamine 50 Injection, Yuhan Co., Ltd.,Seoul, Korea, dose: 50 mg/kg) and acepromazine(Cedaject Injection, Samwoo Chemical Co., Ltd.,Seoul, Korea, dose: 10 mg/kg). After confirmingthe induction of anesthesia, the femoral artery (forblood sampling) and vein (for administration)were catheterized with polyethylene tubing (PE-50; Clay Adams, Parsippany, NJ), filled withheparinized saline (25 U/mL). After recovering(approximately 2 h after the surgery) from theanesthesia, the rats received donepezil (dose: 0.01,0.02, 0.05, or 0.1 mg/kg; sterile water for injectionas the vehicle) in the form of a bolus via theintravenous cannula. Blood samples (250mL)were collected from the catheter connected tothe artery at 1, 2, 3, 4, and 5 min, and the plasmafraction obtained by centrifugation. Immediatelyafter the last blood collection, the rat wasdecapitated, and the brain tissue collected/weighed. The plasma and brain samples werefrozen at �808C until used for the analysis ofdonepezil (see below). The apparent brain uptakeclearance (CLapp,br) was estimated by the previousmethod,19,20 in which the apparent uptake clear-ance was estimated based on the drug amount inthe brain at time t and the area under the plasmaconcentration–time curve to the time of the brainsampling for donepezil. In addition, Kp,br, theconcentration ratio between the brain and theplasma, at 5 min after the administration wasdetermined for donepezil.
Uptake of Various Compounds in MBEC4 Cells
MBEC4 cells, immortalized mouse brain capillaryendothelial cells,18,21 were used as an invitromodelof the BBB. The cells were maintained at 378C inDulbecco’s modified Eagle’s medium (low glucose)containing 10% fetal bovine serum, 1% nonessen-tial amino acid solution, 100 U/mL penicillin and0.1 mg/mL streptomycin in an atmosphere of5% CO2 and 90% relative humidity. The cells wereseeded on collagen-coated multiwell dishes at adensity of 1� 106 cells/well and the culturesreached confluence after 3 days. The cells weresubsequently washed twice with phosphate-buf-fered saline (PBS) and preincubated with uptakebuffer. The uptake buffer contained Hank’sbalanced salt solution with 25 mM HEPES and25 mM glucose (pH 7.4). After an appropriateequilibration period (i.e., 60 min), the uptake ofTEA, carnitine, choline, MPP, CsA, digoxin, PAH,or BPC (for all substrates except carnitine, 100 nMand for carnitine, 1mM) was studied at 378C in the
DOI 10.1002/jps
absence or presence of donepezil (1000-fold excessin concentration of the corresponding substrateconcentration) for 10 min. When necessary, theuptake of TEA, MPP, choline, and carnitine wasalso examined at various donepezil concentrationsranging from 0.001 to 1000mM. In this series ofuptake experiments, a trace level (approximately0.1–1% of total concentration) of radiolabeledsubstrates, in addition to the corresponding unla-beled substrates, were used to determine theamounts of tissue-associated substrates. At theend of the incubation, the medium was removed byaspiration and the incubated cells washed threetimes with 1 mL of ice-cold PBS to terminate theuptake. After aspiration of the medium, the cellswere lysed by the addition of 0.1 N NaOH (1 mL),the mixture allowed to stand overnight at roomtemperature, and the radioactivity determined byliquid scintillation counting (Wallac 1409, LKB,Mt. Waverley, Australia) in the presence of ascintillation fluid (Ultima GoldTM, PerkinElmerLife and Analytical Sciences).
Cloning of Organic Cation Transporters and TheirFunctional Expression in Flp-In 293 Cells
To determine whether rat OCTs are involved in thetransport of donepezil, rOCT1, rOCT2, rOCT3,rOCTN1, rOCTN2, and rCHT1 were individuallycloned and stably transfected into Flp-In 293 cells(Invitrogen), a variant of HEK293 cells for site-specific integration of the gene of interest.22,23
rOCT1, rOCT2, rOCTN1, and rOCTN2 were clonedfrom rat kidney premium mRNA (BD BiosciencesClontech, Palo Alto, CA). rOCT3 and rCHT1 werealso cloned from rat placenta total mRNA (BDBiosciences Clontech) and rat brain total mRNA(BD Biosciences Clontech), respectively. In thisstudy, the SMART RACE procedure (BD Bios-ciences Clontech) was applied for the reversetranscription. The specific primers and conditionsfor thepolymerasechainreaction(PCR)proceduresare listed in Table 1.15 The PCR reaction mixture(40mL), containing the corresponding primer(0.2mM, Tab. 1), deoxynucleoside triphosphate(0.2mM), ExTaq DNA polymerase (2 U), and theprovided buffer, was subjected to amplificationusing the above experimental conditions (Tab. 1).All amplification products were cloned intopcDNA5/FRT (Invitrogen), a vector for the Flp-Inexpression system, and the identity of the insertswas confirmed by sequencing. The cloned cDNAs,containing the complete coding region, were underthe control of the T7 promoter of the vector.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009

Table
1.
Lis
tof
Pri
mer
san
dC
ond
itio
ns
Use
din
the
Clo
nin
gof
Rat
Org
an
icC
ati
onT
ran
spor
ters
Gen
eS
ense
Str
an
d(5
0 –30 )
An
tise
nse
Str
an
d(5
0 –30 )
An
nea
lin
gT
emp
eratu
re(8
C)
Pro
du
ctS
ize
(kb)
rOC
T1
CC
CA
AG
CT
TG
CC
GC
CA
CC
GC
AT
GA
GG
TA
TG
AG
GT
GG
AC
TC
CG
TC
TA
GA
TC
AG
GT
AC
TT
GA
GG
AC
TT
GC
C58
1.9
rOC
T2
CC
CA
AG
CT
TG
CC
GC
CA
CC
CC
AG
GG
AC
CA
TG
CC
GC
TC
GA
GT
GA
TG
GA
GT
TC
AG
GG
GT
AA
TG
56
1.8
rOC
T3
CC
CG
GT
AC
CG
CC
GC
CA
CC
AC
CC
TT
TA
GG
GC
AG
GC
TA
TA
AC
CG
GA
TA
TC
TC
TT
TA
CG
GT
TC
CT
TG
GA
AC
G57
1.7
rOC
TN
1C
CC
GG
TA
CC
GC
CG
CC
AC
CA
AG
CA
TG
AG
GG
AC
TA
TG
AC
CT
AC
CG
GA
TA
TC
GC
AG
TT
AT
TA
GA
AC
CT
TG
GG
G60
1.7
rOC
TN
2C
CC
AA
GC
TT
GC
CG
CC
AC
CA
TG
CG
GG
AC
TA
CG
AC
GA
GG
TC
CG
GG
TA
CC
TT
TT
TC
AC
CT
TC
TG
GA
CA
GG
G57
1.7
rCH
T1
CC
CG
GT
AC
CG
CC
GC
CA
CC
TG
TG
GA
AG
GA
TC
TT
CA
CG
GT
AC
CG
GA
TA
TC
TC
AT
CC
AT
AA
TA
TC
TT
AT
TT
C54
1.7
4 KIM ET AL.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
When the functional expression of the trans-porters was needed, the pcDNA5/FRT vector,containing rat OCT1, OCT2, OCT3, OCTN1,OCTN2, or CHT1 (pcDNA5/FRT/rOCT1, pcDNA5/FRT/rOCT2, pcDNA5/FRT/rOCT3, pcDNA5/FRT/rOCTN1, pcDNA5/FRT/rOCTN2, or pcDNA5/FRT/rCHT1) was transfected in Flp-In 293 cellsusing Lipofectamine 2000 (Invitrogen). The trans-fected cells were incubated in culture mediacontaining 0.1 mg/mL hygromycin B (Invitrogen)for several weeks for selection. The expressions ofrOCT1, rOCT2, rOCT3, rOCTN1, rOCTN2, andrCHT1 were verified by reverse transcription-polymerase chain reaction (RT-PCR) at the end ofthe selection period and the function examinedby uptake experiments with corresponding repre-sentative substrates.
Semi-Quantitative RT-PCR Analysis
RT-PCR analyses were carried out to identify theexpression of organic cation transport systems inMBEC4 cells, the rat brain, and Flp-In 293 cellsindividually expressing OCTs. Total mRNA wasextracted from MBEC4 cells or transfected Flp-In293 cells using an RNeasy Mini Kit (QiagenGmbH, Hilden, Germany) according to the man-ufacturer’s instructions. For the isolation of totalmRNA from the rat brain (approximately 1.7 g),the TRIzol reagent (Invitrogen) was used accord-ing to the manufacturer’s instructions. Afterisolation, the concentration and purity of theextracted RNA were confirmed by UV spectro-photometry at 260/280 nm. Reverse transcriptionwas carried out under the following conditionsusing an RNA LA PCR Kit (Takara Shuzo Co.,Ltd., Shiga, Japan); 428C for 30 min, heating to998C for 5 min and subsequently cooling to 48C.The specific primers used in the PCR reactionsand thermocycling conditions are given inTable 2.15,24 The PCR products were separatedby gel electrophoresis on 1.2% (w/v) agarose,inspected under UV light after ethidium bromidestaining and photographed with a digital camera.
Uptake of Organic Cations and Donepezil by rOCTs,rOCTNs, and rCHT1-Transfected Flp-In 293 Cells
Transfected Flp-In 293 cells were maintained at378C in Dulbecco’s modified Eagle’s medium (lowglucose) containing 10% fetal bovine serum, 1%nonessential amino acid solution, 100 U/mL peni-cillin, 0.1 mg/mL streptomycin, and 0.1 mg/mLhygromycin B in an atmosphere of 5% CO2 and
DOI 10.1002/jps

Table 2. List of Primers and Conditions Used in the RT-PCR Procedure
mRNA Sense Strand (50–30) Antisense Strand (50–30)Annealing
Temperature (8C)
mOCT1 ACGATGTCCTGGAGCACGTT AGCCCAAGTTCACACAGGACT 50–60mOCT2 TGCCGACCGTGGATGATATT ACAGCACCGATGAAAAACCC 56mOCT3 ACGGAACTTTGGGGTTTCAC AACTGGAGAACTGGTTGGGG 57mOCTN1 CCTCACCACCTCCCTGTTCT TGTTCATCTTTGCGGCTTTC 56mOCTN2 TGCTTTTTGTCCTTGTTGGC GACTTCAACAGCCGCCAGTA 56mCHT TTTTGCCCTGTCACATCCTG AAACATAGAACTCGCCGACA 55rOCT1 TGCAGACAGGTTTGGCCGTAA TCGAGGCCGCTATTGGGTAGA 57rOCT2 GCTGGTTAATTGGCTACATCC TTCTTGGCCTCTGCATATTC 56rOCT3 CATCGTCAGCCAGTTTGACC TTGACGAGAGGTGTTTCCCA 57rOCTN1 TAGCTGGGGTGCTGTTCTGG TGGGGTTTTCTTCTCTGTCC 57rOCTN2 AGTGTCCTTCTCTTCATACAGC TGTGCTCTTTAGGACCGTTG 57rCHT1 TCGTGGGAGGGCTCTACTCT ACCAAGCCCAAAGAAGGAAA 55
MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 5
90% relative humidity. Cells (5� 105) were seededin poly-L-ornithine (Sigma, St. Louis, MO) coated12-well plates (Corning Glass Works, Corning,NY) and uptake measured 3 days after seeding.After reaching confluence, the cells were washedtwice with PBS and preincubated with uptakebuffer. The uptake buffer contained Hank’sbalanced salt solution with 25 mM HEPES and25 mM glucose (pH 7.4). For Flp-In 293 cellsexpressing rOCT1, rOCT2, or rOCT3, each cellline was separately incubated with MPP (i.e., acommon substrate for the three transporters; thefinal concentration in the uptake mixture of100 nM) for 10 min at 378C in the absence orpresence (0.1, 1, 10, 100, 200, or 500mM, finalconcentration) of donepezil. For Flp-In 293 cellsexpressing rOCTN1 or rCHT1, each cell line wasseparately incubated with 100 nM (final concen-tration) of ergothioneine or choline, respectively,in the absence or presence (0.1, 1, 10, 100, 200, or500mM, final concentration) of donepezil.rOCTN2-expressing Flp-In 293 cells were incu-bated with a carnitine solution (final concentra-tion of 1mM) for 10 min at 378C in the absence orpresence (1, 10, 100, 1000, 2000, or 5000mM, finalconcentration) of donepezil. In this series ofuptake experiments, a trace level (approximately0.1–1% of total concentration) of radiolabeledsubstrates, in addition to the correspondingunlabeled substrates, were used to determinethe amounts of tissue-associated substrates. Atthe end of the incubation, the medium wasremoved by aspiration and the incubated cellswashed three times with 1 mL of ice-cold PBS toterminate the uptake. After aspiration of themedium, the cells were lysed by the addition of0.1 N NaOH (1 mL); the mixture allowed to stand
DOI 10.1002/jps
at room temperature overnight and the radio-activity determined by liquid scintillation count-ing (Wallac 1409, LKB) in the presence of ascintillation fluid (Ultima GoldTM, PerkinElmerLife and Analytical Sciences).
When necessary, the uptake of donepezil wasstudied in Flp-In 293 cells individually expressingthe cationic transporters, to determine whether thetransport of the drug was affected by the transpor-ter(s). Thus, the mock-transfected Flp-In 293 cellsand each of the OCT-transfected Flp-In 293 cellswere separately incubated with donepezil at 10mM(below the expected KM) for 30 min at 378C. At theend of the incubation, the medium was removed byaspiration and the cells washed five times with1 mLof ice-coldPBScontainingorganiccations (i.e.,10 mM each of MPP, carnitine, and choline) tominimize nonspecific binding of the drug in theexperimentalset-up.Apreliminarystudyindicatedthat other washing protocols (e.g., ice-cold PBSwithout the addition of organic cations and ice-coldPBS containing bovine serum albumin) were noteffective in eliminating nonspecific binding. Afteraspirationof themedium,thecellswerelysedbytheaddition of0.1 N NaOH (1 mL); themixture allowedto stand at 48C overnight, and the concentration ofdonepezil determined by a liquid chromatography–mass spectrometry (LC–MS) assay (see below). Thelysis and standing conditions were found to have noeffect on the concentration of donepezil (data notshown).
Inhibition Study for Organic Cations on the Uptakeof Donepezil to the Brain In Vivo
To examine the influence of organic cations on theuptake of donepezil by the brain, MPP, ergothio-
JOURNAL OF PHARMACEUTICAL SCIENCES 2009

6 KIM ET AL.
neine, carnitine, or choline were intravenouslyadministered at doses of 20 mg/kg, respectively,5 min prior to the intravenous administration ofdonepezil (dose: 0.01 mg/kg). Blood collection andsample processing were carried out similar to theprocedure described in the previous section.Sterile water for injection (Daihan Pharm Co.,Ltd., Seoul, Korea) was used as the vehicle fordissolving the potential inhibitors and had noeffect on the uptake of donepezil by the brain (datanot shown).
To determine whether the pretreatment withcholine altered the cerebral blood flow therebyaffecting the uptake of donepezil, the brain uptakeclearance for caffeine, a compound readily perme-able to the brain and a potential marker forchanges in cerebral blood flow, was evaluated.Caffeine, dissolved in sterile water at a concen-tration of 1 mg/mL, was injected intravenously ata dose of 1 mg/kg, 5 min after the intravenousadministration of choline at a dose of 20 mg/kg orof vehicle only (i.e., sterile water for injection). Inthis experiment, the volume of the dosing solution(or vehicle) given to rats was fixed at 1 mL/kg.Blood samples were collected at 1, 2, 3, 4, and5 min after caffeine administration and immedi-ately centrifuged to collect the plasma; brainsamples were also collected at the last bloodsampling time. The plasma and brain sampleswere frozen at �808C until used for the analysis ofcaffeine (see below).
Plasma Protein Binding of Donepezil
Microcon centrifugal filter devices (Microcon YM-3, Millipore Corp., Bedford, MA), having anominal molecular weight limit of 3000, wereused to separate the free drug from its bound formin the rat plasma.25 Rat blank plasma (1 mL) wasspiked with known amounts of donepezil (finalconcentration of 0.01 mg/mL). The sample wasthoroughly mixed by vortexing and the mixtureincubated at 378C in a shaking water bath for12 min. An aliquot (500mL) was then transferredto a Microcon centrifugal filter device (MicroconYM-3) and the sample was centrifuged (5000 rpm,12 min, 378C). The filtrate was weighed and theconcentration of donepezil determined by an LC–MS assay (see below). To examine the effect ofcholine on the binding of donepezil to plasmaproteins, rat blank plasma (1 mL) was spiked withknown amounts of donepezil and choline, to give afinal concentration of 0.01 and 20 mg/mL fordonepezil and choline, respectively. Subsequent
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
procedures were identical to the control sample(i.e., no choline addition). The concentration ofdonepezil was determined by an LC–MS assay fordonepezil (see below) and the free fractioncalculated.
Binding of Donepezil to Brain Tissue
Since the choline pretreatment could have aneffect on the binding of donepezil to brain tissue,the tissue binding of donepezil was studied in thepresence and the absence of choline. To accom-plish this, one volume of rat brain (assuming thedensity of the tissue to be unity) was homogenizedin one volume of PBS (pH 7.4) using a tissuehomogenizer (Ultra-Turrax T25; IKALabortech-nik, Staufen, Germany). Donepezil was added toan aliquot (1 mL) of the brain homogenate to give afinal concentration of the drug of 0.01 mg/mL inthe absence and the presence of choline (finalconcentration of 20 mg/mL). The sample wasthoroughly vortex-mixed and the mixture incu-bated at 378C in a shaking water bath for 10 min.An aliquot (500mL) of the mixture was thentransferred to a Microcon centrifugal filter deviceand the sample centrifuged (10,000 rpm, 30 min,378C). The filtrate was weighed and the concen-tration of donepezil was determined by an LC–MSassay (see below). The free fraction of donepezilwas calculated and compared for samples withand without added choline.
LC–MS Assay for Donepezil
Donepezil concentrations in the plasma and brainsamples were determined by LC–MS usingdiphenhydramine as an internal standard (IS,Ref.26). To a 100mL aliquot of plasma sample, anIS solution (10mL, the concentration of thesolution at 10mg of IS per 1 mL of methanol)and phosphate buffer (100mL, 10 mmol/L, pH 14)were added. The mixture was then brieflyvortexed and ethyl acetate (600mL) added. Theresulting mixture was vortex-mixed, then shakenon a mechanical shaker for 30 min. After centri-fugation at 12,000 rpm for 15 min, the upperorganic layer was transferred to a fresh tubeand evaporated to dryness under a gentle streamof nitrogen. The residue was reconstituted in100mL of acetonitrile, and the mixture vortex-mixed for 10 min. An aliquot (30mL) of themixture was then injected onto an LC–MS system(see below). For processing the brain sample, onevolume of brain tissue (assuming the density ofthe tissue to be unity) was homogenized in two
DOI 10.1002/jps

MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 7
volumes of distilled water using a tissue homo-genizer (Ultra-Turrax T25; IKALabortechnik).The homogenate was then centrifuged at3000 rpm for 30 min and the supernatant col-lected. To a 200mL aliquot of supernatant, ISsolution (20mL, the concentration of the solutionat 10mg of IS per 1 mL of methanol) andphosphate buffer (200mL, 10 mmol/L, pH 14) wereadded. The mixture was then briefly vortexed andethyl acetate (1.2 mL) added. Subsequent proce-dures were identical to that described for theplasma sample.
LC–MS analyses were carried out on a WatersZQ (Milford, MA) single quadruple mass spectro-meter and equipped with an electrospray probeinterfaced to a liquid chromatograph, consisting ofa Waters 2695 separation module (quaternarypump and autoinjector) and a vacuum degasser. ASynergi 4m MAX-RP column (75 mm� 4.6 mm,4mm), supplied by Phenomenex (Torrance, CA)was used. The mobile phase consisted of 90:10 (v/v) mixture of solvent A (50% acetonitrile inmethanol) and solvent B (0.01 M ammoniumacetate, pH 4). The flow rate was maintained at0.4 mL/min. The electrospray interface probeconditions were as follows: 2.81 kV capillaryvoltage, 34 V cone voltage, 2 V extractor voltage,0.2 V RF lens voltage, 1008C source temperature,1508C desolvation temperature, 200 L/h desolva-tion gas flow, and 50 L/h cone gas flow. Quanti-fication was carried out using selected ionmonitoring (SIM) at m/z 380 for donepezil andm/z 256 for diphenhydramine (IS). The response ofthe detector was linear in the concentration rangeexamined (i.e., 0.5–100 nM) for both the plasmasamples and the brain homogenate with inter-/intra-day precisions of less than 15% and anaccuracy within 15% of the theoretical value,indicating the assay was valid in the range ofconcentration studied.
HPLC Analysis for Caffeine
An HPLC assay, essentially as described in ourprevious study,20 was used to determine theconcentration of caffeine in the plasma and brainsamples. In a typical experiment, 300mL of ethylacetate was added to a 100mL aliquot of plasmasample. The mixture was vortex-mixed, thenshaken on a mechanical shaker for 30 min. Aftercentrifugation at 12,000 rpm for 15 min, the upperorganic layer was transferred to a fresh tube andevaporated to dryness under a gentle stream ofnitrogen. The residue was reconstituted in 100mL
DOI 10.1002/jps
of methanol, and vortex-mixed for 10 min. A 50mLaliquot of the resulting solution was injected ontothe HPLC system. For processing the brainsample, one volume of brain tissue was homo-genized in two volumes of distilled water using atissue homogenizer. The homogenate was thencentrifuged at 3,000 rpm for 30 min and thesupernatant collected. To a 100mL aliquot super-natant, 300mL of ethyl acetate was added.Subsequent procedures were identical to thatdescribed for the plasma sample.
The HPLC system consisted of a pump (ModelPU610; GL Sciences, Tokyo, Japan), a UV detector(UV620; GL Sciences), and a C18 reversed-phasecolumn (CAPCELL PAK, 150 mm� 4.6 mm, 5mm,Shiseido). The mobile phase consisted of acetoni-trile–10 mM ammonium acetate buffer (13:87, v/v). The flow rate was 1 mL/min and the eluentmonitored at a wavelength of 270 nm. Thecalibration curves (concentration range of 0.1–5mg/mL), constructed for the plasma sample andthe brain homogenate, were found to be linear.The inter-/intra-day precisions of the assay were<15% and an accuracy within 15% of thetheoretical value, indicating the assay was validin the range of caffeine concentration studied.
Data Analysis
To determine the apparent in vivo brain uptakeclearance (CLapp,br) by the previous method,19,20
the rate of uptake of the compound in the brainwas calculated by Eq. (1),
XbrainðtÞ ¼ CLapp;br � AUCt0 (1)
where Xbrain(t), CLapp,br, and AUCt0 represent the
amount of drug in the brain at time t, the uptakeclearance to the brain, and the area under theplasma concentration time curve from time 0 tothe time of the brain sampling, respectively.
In our inhibition study, the IC50 values werecalculated from the inhibition data using Eq. (2);the kinetic parameter was estimated from thedata by nonlinear regression analysis (Winnonlin3.1, Pharsight, Mountain View, CA) running ona PC
v ¼ V0
1 þ ðCdonepezil=IC50Þn(2)
In this equation, v, n, and V0 represent the rateof uptake in the presence of the inhibitor (i.e.,donepezil), Hill’s coefficient, and the rate ofuptake in the absence of the inhibitor, respec-
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
8 KIM ET AL.
tively. In addition,Cdonepezil is the concentration ofdonepezil as the inhibitor.
In both the plasma protein and the brain tissuebinding studies, the percent of bound drug wascalculated as follows:
1 � Cf
Ct
� �� 100 (3)
where Cf is the drug concentration in the filtrate(i.e., the unbound drug concentration), and Ct isthe total drug concentration expected in theplasma or brain homogenate.
To compare the mean values between thetreatments, one-way ANOVA, followed by Dun-can’s test, was used. In all cases, p< 0.05 wasaccepted as denoting a statistical difference.All data are expressed in the form of themean�SD.
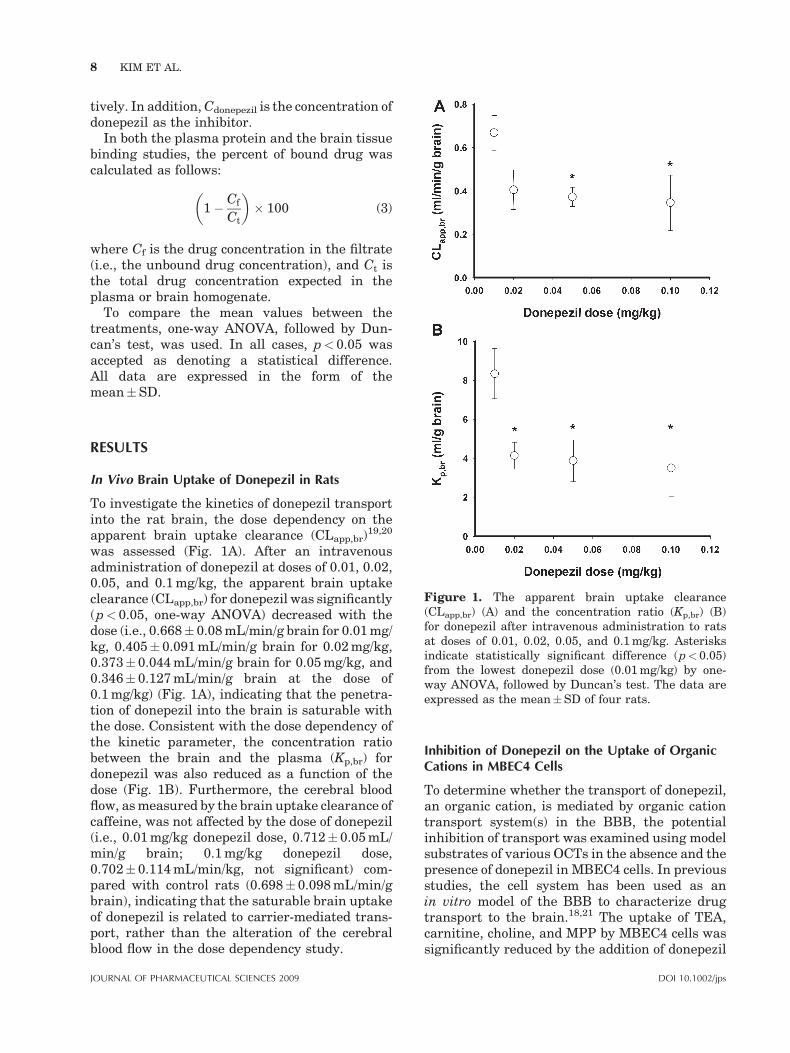
Figure 1. The apparent brain uptake clearance(CLapp,br) (A) and the concentration ratio (Kp,br) (B)for donepezil after intravenous administration to ratsat doses of 0.01, 0.02, 0.05, and 0.1 mg/kg. Asterisksindicate statistically significant difference (p< 0.05)from the lowest donepezil dose (0.01 mg/kg) by one-way ANOVA, followed by Duncan’s test. The data areexpressed as the mean�SD of four rats.
RESULTS
In Vivo Brain Uptake of Donepezil in Rats
To investigate the kinetics of donepezil transportinto the rat brain, the dose dependency on theapparent brain uptake clearance (CLapp,br)
19,20
was assessed (Fig. 1A). After an intravenousadministration of donepezil at doses of 0.01, 0.02,0.05, and 0.1 mg/kg, the apparent brain uptakeclearance (CLapp,br) for donepezil was significantly(p< 0.05, one-way ANOVA) decreased with thedose (i.e., 0.668� 0.08 mL/min/g brain for 0.01 mg/kg, 0.405� 0.091 mL/min/g brain for 0.02 mg/kg,0.373� 0.044 mL/min/g brain for 0.05 mg/kg, and0.346� 0.127 mL/min/g brain at the dose of0.1 mg/kg) (Fig. 1A), indicating that the penetra-tion of donepezil into the brain is saturable withthe dose. Consistent with the dose dependency ofthe kinetic parameter, the concentration ratiobetween the brain and the plasma (Kp,br) fordonepezil was also reduced as a function of thedose (Fig. 1B). Furthermore, the cerebral bloodflow, as measured by the brain uptake clearance ofcaffeine, was not affected by the dose of donepezil(i.e., 0.01 mg/kg donepezil dose, 0.712� 0.05 mL/min/g brain; 0.1 mg/kg donepezil dose,0.702� 0.114 mL/min/kg, not significant) com-pared with control rats (0.698� 0.098 mL/min/gbrain), indicating that the saturable brain uptakeof donepezil is related to carrier-mediated trans-port, rather than the alteration of the cerebralblood flow in the dose dependency study.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
Inhibition of Donepezil on the Uptake of OrganicCations in MBEC4 Cells
To determine whether the transport of donepezil,an organic cation, is mediated by organic cationtransport system(s) in the BBB, the potentialinhibition of transport was examined using modelsubstrates of various OCTs in the absence and thepresence of donepezil in MBEC4 cells. In previousstudies, the cell system has been used as anin vitro model of the BBB to characterize drugtransport to the brain.18,21 The uptake of TEA,carnitine, choline, and MPP by MBEC4 cells wassignificantly reduced by the addition of donepezil
DOI 10.1002/jps

MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 9
(e.g., at 100mM donepezil in the medium, 38.4%,26.3%, 27.8%, and 35.4% reduction for TEA,carnitine, choline, and MPP uptake, respectively;Fig. 2A). To further assess the specificity ofdonepezil transport, the interaction of transportswas examined for neutral/anionic compounds thatare not substrates to OCTs. Thus, the uptake ofCsA [a substrate for P-gp and BCRP27], digoxin [asubstrate for P-gp and Oatp228–30], PAH [OAT1/331,32], or BPC [OAT1/331,32] was examined in the
Figure 2. Uptake of organic cations (A) and neutral/anionic compounds (B) using MBEC4 cells in the absenceand the presence of donepezil (1000mM for carnitineuptake and 100mM for the other substrates). Asterisksindicate a statistically significant difference (p< 0.001)from the control (i.e., the absence of donepezil). Data areexpressed as the mean�SD of triplicate runs.
DOI 10.1002/jps
presence of donepezil (100mM) in the in vitromodel. In contrast to the transport study withorganic cations, the presence of donepezil did notlead to an appreciable reduction in transport(Fig. 2B) for these neutral/anionic compounds,indicating that certain specificity exists for thetransport of donepezil. These observations areconsistent with the hypothesis that donepezil istransported across the BBB via organic cationtransport systems.
The concentration dependency of the inhibitionof transport in the cationic substrates (viz. TEA,MPP, choline, and carnitine) was also assessed inthe presence of donepezil (0.001–1000mM)(Fig. 3A–D). As expected, the uptake of organiccations was significantly inhibited in a dose-dependent manner.
Expression of OCTs, OCTNs, and CHT in MBEC4Cells and the Rat Brain
In this study, the expression of OCTs, asdetermined by the level of mRNA via semi-quantitative RT-PCR, was assessed in MBEC4cells. As shown in Figure 4A, PCR productscorresponding to mOCT2, mOCT3, mOCTN1,mOCTN2, and mCHT were readily detected inMBEC4 cells, while mOCT1 mRNA was unde-tectable in the cell, indicating that mOCT2,mOCT3, mOCTN1, mOCTN2, and mCHT arepresent in our in vitro model of the BBB. Tocompare the expression of OCTs in the rat brainwith that in MBEC4 cells, total mRNA wasextracted from the rat brain and RT-PCR analysescarried out (Fig. 4B). Although the expressionpattern in the rat brain is generally consistent forthe transporters to that of MBEC4, rOCT1 wasdetected in the case of the rat brain. Glyceralde-hyde-3-phosphate dehydrogenase (GAPDH), ahousekeeping gene, was successfully amplifiedin all samples, suggesting that the RT-PCRprocedure is adequate for the detection of themRNA expressions of the transporters in studieswith both MBEC4 cells and the rat brain.
Functional Expression of Organic CationTransporters in Flp-In 293 Cells
Since the six OCTs were found in rat brainsamples, the contribution of the transporters inthe transport of donepezil was studied. Toaccomplish this, the rat OCTs, rOCT1, rOCT2,rOCT3, rOCTN1, rOCTN2, and rCHT1 werecloned and expressed in Flp-In 293 cells. After
JOURNAL OF PHARMACEUTICAL SCIENCES 2009

Figure 3. Uptake of organic cations using MBEC4 cells in the absence and thepresence of donepezil (0.001–1000mM). (A) TEA uptake, (B) MPP uptake, (C) cholineuptake, and (D) carnitine uptake. Asterisks indicate statistically significant difference(p< 0.05) from the control (i.e., the absence of donepezil). Data are expressed as themean�SD of triplicate runs.
Figure 4. Expression of OCTs, OCTNs, and CHT inMBEC4 cells (A) and rat brain tissues (B).
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
10 KIM ET AL.
the selection period, the expression of thetransporters was verified by RT-PCR analysis(Fig. 5) and their functionality determined by aseries of uptake studies using representativesubstrates for organic cation transport systems(Fig. 6). In this study, MPP was used as thesubstrate for rOCT1, rOCT2, and rOCT333;ergothioneine and carnitine as the substrate forrOCTN134 and rOCTN2,35 respectively, and cho-line as the substrate for rCHT1.36 The RT-PCRstudy (Fig. 5) indicated that rOCT1, rOCT2,rOCT3, rOCTN1, rOCTN2, and rCHT1 could bereadily detected in Flp-In 293 cells expressingcorresponding OCTs, indicating that the trans-
DOI 10.1002/jps

Figure 5. Expression of organic cation transporters in Flp-In 293 cells transfectedwith rOCTs, rOCTNs, or rCHT1 and mock-transfected Flp-In 293 cells.
MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 11
porters are individually expressed in the trans-fected cells. Although the native uptake by mock-transfected Flp-In 293 cells was somewhat high,the uptake of MPP (100 nM) in rOCT1, rOCT2,and rOCT3-expressing Flp-In 293 cells wasincreased up to 2.67-, 5.59-, and 2.51-fold,respectively, compared with the control cells(Fig. 6A–C). The uptake of ergothioneine inrOCTN1-expressing Flp-In 293 cells was signifi-cantly increased up to 76.1-fold (Fig. 6D), and theuptake of carnitine in rOCTN2-expressing Flp-In293 cells was also increased up to 3.1-fold (Fig. 6E)compared with mock-transfected Flp-In 293 cells.In addition, the uptake of choline in rCHT1-expressing Flp-In 293 cells was associated with anapproximately 3.75-fold increase (Fig. 6F) com-pared with the control. Collectively, these obser-vations indicate that OCTs found in the rat brainare individually cloned and expressed in thetransfected Flp-In 293 cells.
Inhibitory Effect of Donepezil on the Uptake ofOrganic Cations in Cells Expressing Organic CationTransporters
In this study, the inhibitory effect of donepezil wasdetermined for the uptake of organic cations inFlp-In 293 cells individually expressing the OCTs.In rOCT1, rOCT2, or rOCT3-expressing Flp-In293 cells, the uptake of MPP was reduced by theaddition of donepezil in a dose-dependent manner(Fig. 7A–C). However, a statistically significantreduction was found only for donepezil concentra-tions higher than 100mM for the cells expressing
DOI 10.1002/jps
rOCTs. When the inhibition data were fitted toEq. (2) by a nonlinear regression analysis, theestimated IC50 values were determined to be 107,243, and 135mM for rOCT1, rOCT2, and rOCT3,respectively (Tab. 3). These results indicate that,although donepezil interacts with rOCT1, rOCT2,and rOCT3, the affinity of donepezil for thetransporters appears to be low.
The uptake of standard substrates for OCTNsand CHT1 (i.e., for OCTN1, ergothioneine; forOCTN2, carnitine; for CHT1, choline) was alsoinhibited by the addition of donepezil in cellsexpressing rOCTN1, rOCTN2, or rCHT1 in adose-dependent manner (Fig. 7D–F). The esti-mated IC50 values with donepezil were 168, 138,and 57.4mM for rOCTN1, rOCTN2, and rCHT1(Tab. 3), respectively. A comparison of the ap-parent affinity of donepezil for the transportssuggests that high capacity/low affinity transportsystems exist for the transport of donepezil andthat, among the cation transport systems, rCHT1has a relatively higher affinity than the othercation transport systems. The uptake of MPP,ergothioneine, carnitine, or choline in mock-transfected Flp-In 293 cells was not significantlyaffected by the addition of various concentrationsof donepezil (data not shown), suggesting that thebasal activity in the transport is not high.
Uptake of Donepezil in Flp-In 293 Cells ExpressingOrganic Cation Transporters
The uptake of donepezil was also studied in Flp-Incells individually expressing OCTs. The native
JOURNAL OF PHARMACEUTICAL SCIENCES 2009

Figure 6. Uptake of representative substrates in Flp-In 293 cells expressing rOCTs,rOCTNs, or rCHT1. (A) rOCT1-expressing Flp-In 293 cells (substrate MPP, 100 nM); (B)rOCT2-expressing Flp-In 293 cells (substrate MPP, 100 nM); (C) rOCT3-expressing Flp-In 293 cells (substrate MPP, 100 nM); (D) rOCTN1-expressing Flp-In 293 cells (substrateergothioneine, 100 nM); (E) rOCTN2-expressing Flp-In 293 cells (substrate carnitine,1mM); (F) rCHT1-expressing Flp-In 293 cells (substrate choline, 100 nM). Asterisksindicate statistically significant difference (p< 0.001) from the control (mock-trans-fected Flp-In 293 cells). Data are expressed as the mean�SD of triplicate runs.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009 DOI 10.1002/jps
12 KIM ET AL.

Figure 7. Uptake of representative substrates in the absence and the presence ofdonepezil (0.1–5000mM) in Flp-In 293 cells expressing rOCTs, rOCTNs, or rCHT1. (A)rOCT1-expressing Flp-In 293 cells (substrate MPP, 100 nM); (B) rOCT2-expressing Flp-In 293 cells (substrate MPP, 100 nM); (C) rOCT3-expressing Flp-In 293 cells (substrateMPP, 100 nM); (D) rOCTN1-expressing Flp-In 293 cells (substrate ergothioneine,100 nM); (E) rOCTN2-expressing Flp-In 293 cells (substrate carnitine, 1mM); (F)rCHT1-expressing Flp-In 293 cells (substrate choline, 100 nM). The solid line wasgenerated by the best-fit parameters obtained from the nonlinear regression analysisbased on Eq. (2). Data are expressed as the mean�SD of triplicate runs.
DOI 10.1002/jps JOURNAL OF PHARMACEUTICAL SCIENCES 2009
MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 13
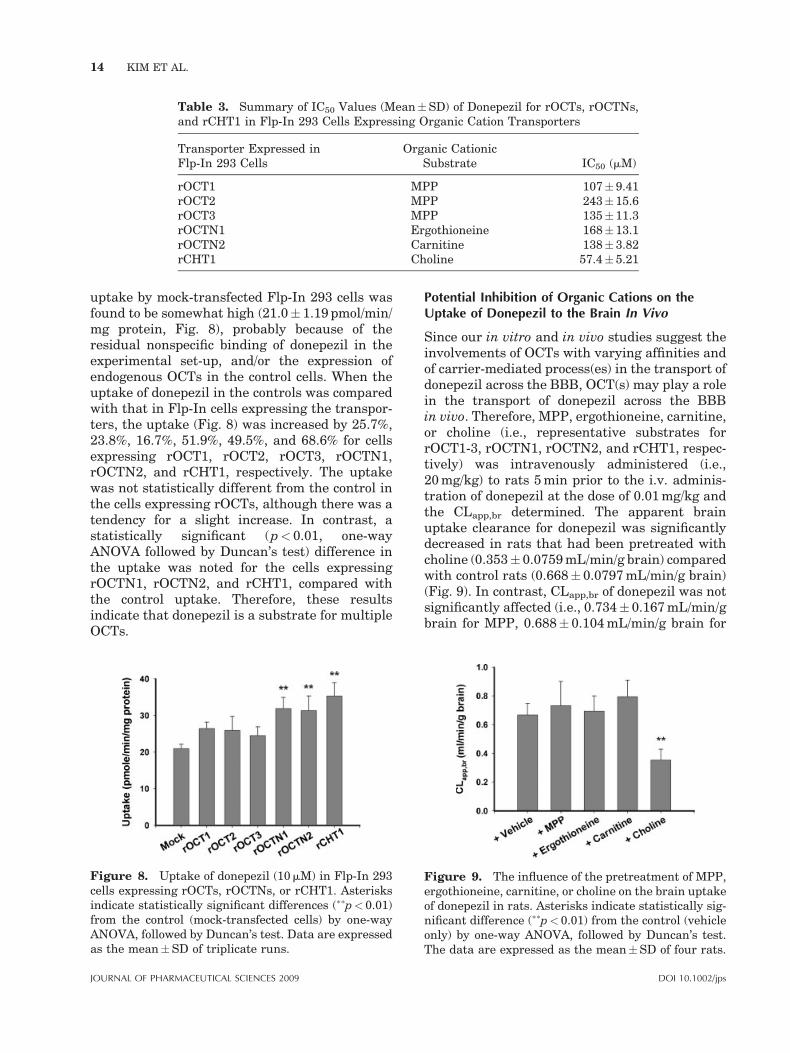
Table 3. Summary of IC50 Values (Mean�SD) of Donepezil for rOCTs, rOCTNs,and rCHT1 in Flp-In 293 Cells Expressing Organic Cation Transporters
Transporter Expressed inFlp-In 293 Cells
Organic CationicSubstrate IC50 (mM)
rOCT1 MPP 107� 9.41rOCT2 MPP 243� 15.6rOCT3 MPP 135� 11.3rOCTN1 Ergothioneine 168� 13.1rOCTN2 Carnitine 138� 3.82rCHT1 Choline 57.4� 5.21
14 KIM ET AL.
uptake by mock-transfected Flp-In 293 cells wasfound to be somewhat high (21.0� 1.19 pmol/min/mg protein, Fig. 8), probably because of theresidual nonspecific binding of donepezil in theexperimental set-up, and/or the expression ofendogenous OCTs in the control cells. When theuptake of donepezil in the controls was comparedwith that in Flp-In cells expressing the transpor-ters, the uptake (Fig. 8) was increased by 25.7%,23.8%, 16.7%, 51.9%, 49.5%, and 68.6% for cellsexpressing rOCT1, rOCT2, rOCT3, rOCTN1,rOCTN2, and rCHT1, respectively. The uptakewas not statistically different from the control inthe cells expressing rOCTs, although there was atendency for a slight increase. In contrast, astatistically significant (p< 0.01, one-wayANOVA followed by Duncan’s test) difference inthe uptake was noted for the cells expressingrOCTN1, rOCTN2, and rCHT1, compared withthe control uptake. Therefore, these resultsindicate that donepezil is a substrate for multipleOCTs.
Figure 8. Uptake of donepezil (10mM) in Flp-In 293cells expressing rOCTs, rOCTNs, or rCHT1. Asterisksindicate statistically significant differences (��p< 0.01)from the control (mock-transfected cells) by one-wayANOVA, followed by Duncan’s test. Data are expressedas the mean�SD of triplicate runs.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
Potential Inhibition of Organic Cations on theUptake of Donepezil to the Brain In Vivo
Since our in vitro and in vivo studies suggest theinvolvements of OCTs with varying affinities andof carrier-mediated process(es) in the transport ofdonepezil across the BBB, OCT(s) may play a rolein the transport of donepezil across the BBBin vivo. Therefore, MPP, ergothioneine, carnitine,or choline (i.e., representative substrates forrOCT1-3, rOCTN1, rOCTN2, and rCHT1, respec-tively) was intravenously administered (i.e.,20 mg/kg) to rats 5 min prior to the i.v. adminis-tration of donepezil at the dose of 0.01 mg/kg andthe CLapp,br determined. The apparent brainuptake clearance for donepezil was significantlydecreased in rats that had been pretreated withcholine (0.353� 0.0759 mL/min/g brain) comparedwith control rats (0.668� 0.0797 mL/min/g brain)(Fig. 9). In contrast, CLapp,br of donepezil was notsignificantly affected (i.e., 0.734� 0.167 mL/min/gbrain for MPP, 0.688� 0.104 mL/min/g brain for
Figure 9. The influence of the pretreatment of MPP,ergothioneine, carnitine, or choline on the brain uptakeof donepezil in rats. Asterisks indicate statistically sig-nificant difference (��p< 0.01) from the control (vehicleonly) by one-way ANOVA, followed by Duncan’s test.The data are expressed as the mean�SD of four rats.
DOI 10.1002/jps

MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 15
ergothioneine, and 0.796� 0.115 mL/min/g brainfor carnitine) by the pretreatment of MPP,ergothioneine, or carnitine (Fig. 9). Furthermore,the CLapp,br of donepezil was not significantlyaltered by the pretreatment with choline at thedoses (i.e., 5 or 10 mg/kg) lower than 20 mg/kg(data not shown). In a preliminary study, weattempted to elevate the dose of organic cations(i.e., choline, MPP, ergothioneine, and carnitine);however, the experimental animals could nottolerate the doses higher than 20 mg/kg. Takenthe experimental difficulty together with thefact the doses (20 mg/kg) of organic cationswere apparently sufficient (viz. approximately2000 times), even accounting for the differences inthe molecular weights, than that of donepezil(0.01 mg/kg), we chose not to purse further theelevation of the dose.
Influence of Cerebral Blood Flow and FreeFraction of Donepezil in Plasma and BrainTissue Homogenates
Since the cerebral blood flow and the free fractionof donepezil can be a determining factor in in vivobrain uptake clearance, the pretreatment withcholine may have caused an alteration in cerebralblood flow and/or the free fraction of donepezilthereby leading to a reduction in donepezilpenetration without any functional change inthe carrier-mediated transport. In this study, thebrain uptake clearance for caffeine, a compoundthat has a high brain uptake clearance20 and,thus, is a reasonable model in studies of cerebralblood flow, was evaluated in the presence and theabsence of choline pretreatment. In particular, a1 mg/kg dose of caffeine was used to eliminate anypossible effect on cerebral blood flow in rats.37 Theapparent brain uptake clearance for caffeine wasnot significantly different in rats pretreated withcholine (0.735� 0.133 mL/min/g brain) comparedwith control rats (0.698� 0.098 mL/min/g brain),suggesting that choline pretreatment does notlead to an alteration of cerebral blood flow. In theliterature, the cerebral blood flow was reported tobe approximately 1.3 mL/min/250 g rat (i.e.,assuming the weight of the brain for 250 g ratto be 1.8 g, blood flow is estimated as 0.722 mL/min/g brain).38
The binding of donepezil to plasma proteins wasdetermined to be approximately 54.6� 3.8% inthe absence of choline, somewhat lower thanthe literature value,39 probably because of thelarge difference in incubation time (viz. 96 h in the
DOI 10.1002/jps
previous study vs. 12 min in the present study).The binding of donepezil to plasma proteins wasapproximately 64.1� 4.42% in the presence ofcholine, not significantly different from thecontrols (i.e., the absence of choline). Further-more, the bound fraction of donepezil was notsignificantly different in the brain tissue homo-genate in the presence (25.3� 5.14%) or absence ofcholine (27.9� 2.87%), indicating that alterationin the binding of donepezil to plasma proteins or tobrain tissue is not a major factor in the reductionin the apparent brain uptake clearance ofdonepezil by choline pretreatment. Given thesefindings along with the choline pretreatmentstudy, it appears that donepezil transport intothe brain, which probably occurs via a carrier-mediated process, is functionally inhibited bycholine in vivo.
DISCUSSION
The site of pharmacological action is the brain forcholinesterase inhibitors such as donepezil, and,thus, the characterization of the transport sys-tem(s) to the brain is likely to have clinicalrelevance for these drugs. In the literature, boththe concentration and the AUC for donepezil inthe brain were significantly higher than those inthe plasma,6 suggesting that carrier-mediatedtransport, in addition to passive diffusion, iskinetically involved in its penetration into thebrain. Consistent with the literature report, wefound that kinetic parameters for the in vivo braintransport for donepezil were apparently saturablein the clinically relevant dose range (i.e., 0.01–0.1 mg/kg) (Fig. 1). Assuming the weight of thebrain to be 1.8 g for a 250 g rat, the uptakeclearance (i.e., 0.668� 0.08 mL/min/g brain) ofdonepezil at a dose of 0.01 mg/kg would bepredicted to be approximately 1.20 mL/min/rat,suggesting that the brain transport of donepezil islimited by the cerebral blood flow. Since thecerebral blood flow remained unchanged in thedonepezil doses used in this study, carrier-mediated transport may be primarily responsiblefor the saturable transport. Furthermore, wefound that the difference in donepezil uptakeclearance was approximately 1.93-fold betweenthe lowest dose (i.e., 0.01 mg/kg, 0.668 mL/min/gbrain) and the highest dose (i.e., 0.1 mg/kg,0.346 mL/min/g brain). Considering the blood flowlimitation in the transport of donepezil to thebrain (i.e., the cerebral blood flow< the intrinsic
JOURNAL OF PHARMACEUTICAL SCIENCES 2009

16 KIM ET AL.
transport clearance of donepezil), the contributionof the saturable component (i.e., the intrinsictransport clearance) is likely to be greater thanthat predicted by the comparison of the uptakeclearances between the two doses.
In this study, we hypothesize that transpor-ter(s) for organic cations is(are) involved in thetransport of donepezil across the BBB, consider-ing the fact that the drug is an organic cation. Inour semi-quantitative RT-PCR study, the profileof expression was generally comparable for OCTs(i.e., OCT2-3, OCTN1-2, and CHT) in the rat brainand in MBEC4 cells, an in vitro model of the BBB.For the case of OCT1, however, a discrepancy inexpression between the rat brain and the cellculture model was observed (Fig. 4A,B). The exactreason(s) for this inconsistency is(are) unknown.Since the rat brain samples contained braintissues in addition to microvessel endothelial cells(i.e., the BBB), the detection of mRNA for rOCT1in the rat brain sample may be related to thepresence of surrounding tissues in the samples. Insupport of this possibility, the expression of OCT1was noted in the cerebellum.40
To further study the underlying mechanism ofdonepezil transport, we functionally expressedeach cation transporter found in the rat brain (viz.including rOCT1) in Flp-In 293 cells, modifiedHEK 293 cells. In this in vitro study, donepezilwas found to inhibit the functions of all of theOCTs (Fig. 7) in the cell systems with IC50 valuesranging from 57.4 to 243mM. Assuming thatcompetitive inhibition is the predominantmechanism and that the substrate concentrations(i.e., in this study, substrate concentrations weremostly 100 nM; for carnitine, the substrate con-centration of 1mM was used) are comparativelylower than the corresponding KM values, the IC50
values are likely to be a reasonable approximationfor the KM of donepezil for the correspondingtransporters. Under this assumption, the affinityof donepezil for the transporters would beexpected to be in the order of (i.e., from thehighest to the lowest affinity) CHT1, OCT1,OCT3, OCTN2, OCTN1, and OCT2. This observa-tion indicates that at least six organic cationtransport systems may participate in the trans-port of donepezil across the BBB. In this study,we found that the IC50 values in MBEC4 cells(Fig. 3) did not appear to correlate well withthe values found for Flp-In 293 cells, probablybecause of the participation of those multipletransporters in the transport (Fig. 8) and/orthe multispecificity of the substrates toward the
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
cationic transporters15,17 in our MBEC4 study.The involvement of multiple transporters (i.e.,OCT2 and OCTN2) was also noted in our previousstudy, in which the mechanism of transport acrossthe BBB was investigated for tacrine, anothercholinesterase inhibitor.15 Similarly, the trans-port of donepezil was enhanced in Flp-In 293 cellsexpressing rOCTN1, rOCTN2, and rCHT1, indi-cative of the multiple involvement of OCTs in thetransport of donepezil in vitro. Consistent withthis hypothesis, the in vivo transport of donepezilto the brain was saturable with the dose andinhibited by pretreatment with choline, a com-pound that is transported by a number of OCTs,including CHT1.
In contrary to the inhibitory effects of donepezilfound in MBEC4 cells (Fig. 3) and the transfectedFlp-In 293 cells (Fig. 7), the uptake of neutral/anionic compounds that were not substrates forOCTs was not affected by donepezil in MBEC4cells (Fig. 2B). These compounds are reported tobe transported by mechanisms (e.g., P-gp, OATs,or Oatps) that are not related to OCTs. Wepreviously reported that the function of P-gp wasdetected in MBEC4 cells41; in the literature, thefunction of OATs and Oatps has been noted in thecell system or in the BBB.42
Murakami et al.16 reported that the CHT issimilar to the OCT family in terms of function andsubstrate selectivity. Consistent with this finding,choline and donepezil appear to share a commontransport system, as evidenced by our in vitro andin vivo results. However, it is not entirely clearwhether CHT1 is the primary mechanism in thebrain transport of donepezil in rats, since it ispossible that unidentified transport system(s),accepting both donepezil and choline, exist(s) inthe BBB thereby facilitating the penetration ofdonepezil to the brain and interacting with theprocess, in vivo, by choline. In addition, thecontribution of OCT1-3 in the brain transport ofdonepezil appeared to be a minor factor consider-ing the fact that MPP, a common substrate forOCT1-3, did not interact with the transport of thedrug in vivo. However, since MPP transport wasreduced by the addition of donepezil in our in vitrostudies, we cannot completely exclude the possi-bility of a partial role of OCTs other than CHT1 inthe transport of donepezil across the BBB. Thisaspect of donepezil warrants further investiga-tion. Alternatively, the absence of an inhibitoryeffect of organic cations other than choline may bean indication that the plasma concentration of thepotential inhibitors was not sufficiently high.
DOI 10.1002/jps

MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 17
However, the elevation of the doses to levelshigher than 20 mg/kg was not feasible because theanimals could not tolerate the elevated doses.
Consistent with our in vitro studies, the brainuptake of donepezil was reduced by pretreatmentwith choline in vivo (Fig. 9). The fraction ofdonepezil bound to rat brain tissue homogenatewas unaffected by the addition of choline,suggesting that the reduced concentration ofdonepezil in the brain caused by choline isprimarily manifested by a reduced transport,rather than the reduced binding/adsorption ofdonepezil to brain tissue. In this study, wedetermined the in vivo uptake of donepezil at5 min after administration. In our previous study,the brain concentration was readily measurableas early as 0.5 min after the administration ofcationic drugs such as caffeine and quinine.20
Furthermore, donepezil was found to be trans-ported to the brain earlier than 5 min43 in apositron emission tomography imaging study with[11C]-donepezil. These observations collectivelysuggest that the transport of certain drugs,including donepezil, to the brain occurs rapidly.In the current study, we found that binding tobrain tissue (i.e., including brain microvessels)was approximately 27.9� 2.87%, suggesting thatthe majority of the donepezil exists as the freeform. Therefore, the amount of donepezil in thebrain measured in 5 min of the administrationmay be primarily free donepezil and a result of thetransport process, rather than the binding of thedrug to brain tissue, including endothelial cells.
Assuming that the brain transport kinetics inhumans for donepezil is comparable to that foundin the rat, the brain transport of donepezil may bealtered in humans when the organic cations arecoadministered with donepezil. Unfortunately,however, this transporter-mediated interactionhas not been systemically studied for the braintransport of donepezil. Since [11C]-donepezil forpositron emission tomography imaging is avail-able,44 a study of drug–drug interactions betweendonepezil and other organic cations in humansmay be feasible. The possibility of inhibition, aswell as the therapeutic impact, warrants furtherstudy, since the therapeutic choice for the treat-ment of Alzheimer’s disease is practically limitedto donepezil.
In conclusion, the transport of representativesubstrates for OCTs expressed in the rat brainwas reduced by the addition of donepezil in cellsindividually expressing the transporters and in anin vitro model of the BBB. The affinities,
DOI 10.1002/jps
estimated from the IC50 values, for donepeziltransport were comparatively low for OCTs andOCTNs while the affinity appeared relatively highfor CHT1. The saturable transport of donepezilacross the brain in vivo was affected by thepretreatment with choline while the transportwas unaffected by MPP, ergothioneine, andcarnitine. These observations indicate that done-pezil and choline share common mechanism(s) inthe transport across the BBB. Organic cationtransport systems other than CHT may also beinvolved as a secondary mechanism. Since anumber of important therapeutic agents areorganic cations, the carrier-mediated transportmay be therapeutically relevant for donepezil.
ACKNOWLEDGMENTS
This study was supported by the Korea Scienceand Engineering Foundation (KOSEF) grantfunded by the Korea Government (MEST) (No.R01-2007-000-11517-0).
REFERENCES
1. Coyle JT, Price DL, DeLong MR. 1983. Alzheimer’sdisease: A disorder of cortical cholinergic innerva-tion. Science 219:1184–1190.
2. Terry AV, Jr., Buccafusco JJ. 2003. The cholinergichypothesis of age and Alzheimer’s disease-relatedcognitive deficits: Recent challenges and theirimplications for novel drug development. J Pharma-col Exp Ther 306:821–827.
3. Kimura M, Akasofu S, Ogura H, Sawada K. 2005.Protective effect of donepezil against Abeta(1-40)neurotoxicity in rat septal neurons. Brain Res1047:72–84.
4. Sugimoto H, Iimura Y, Yamanishi Y, Yamatsu K.1995. Synthesis and structure-activity relation-ships of acetylcholinesterase inhibitors: 1-benzyl-4-[(5,6-dimethoxy-1-oxoindan-2-yl)methyl]piperi-dine hydrochloride and related compounds. J MedChem 38:4821–4829.
5. Nakashima K, Itoh K, Kono M, Nakashima MN,Wada M. 2006. Determination of donepezil hydro-chloride in human and rat plasma, blood and brainmicrodialysates by HPLC with a short C30 column.J Pharm Biomed Anal 41:201–206.
6. Matsui K, Mishima M, Nagai Y, Yuzuriha T, Yoshi-mura T. 1999. Absorption, distribution, metabo-lism, and excretion of donepezil (Aricept) after asingle oral administration to Rat. Drug Metab Dis-pos 27:1406–1414.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009

18 KIM ET AL.
7. Wilkinson DG. 1999. The pharmacology of donepe-zil: A new treatment of Alzheimer’s disease. ExpertOpin Pharmacother 1:121–135.
8. Shigeta M, Homma A. 2001. Donepezil for Alzhei-mer’s disease: Pharmacodynamic, pharmacokinetic,and clinical profiles. CNS Drug Rev 7:353–368.
9. Jann MW, Shirley KL, Small GW. 2002. Clinicalpharmacokinetics and pharmacodynamics of choli-nesterase inhibitors. Clin Pharmacokinet 41:719–739.
10. Thevis M, Wilkens F, Geyer H, Schanzer W. 2006.Determination of therapeutics with growth-hor-mone secretagogue activity in human urine fordoping control purposes. Rapid Commun MassSpectrom 20:3393–3402.
11. Xia Z, Jiang X, Mu X, Chen H. 2008. Improvementof microemulsion electrokinetic chromatographyfor measuring octanol-water partition coefficients.Electrophoresis 29:835–842.
12. Murakami H, Takanaga H, Matsuo H, Ohtani H,Sawada Y. 2000. Comparison of blood-brain barrierpermeability in mice and rats using in situ brainperfusion technique. Am J Physiol Heart Circ Phy-siol 279:H1022–H1028.
13. Kang YS, Lee KE, Lee NY, Terasaki T. 2005. Done-pezil, tacrine and alpha-phenyl-n-tert-butyl nitrone(PBN) inhibit choline transport by conditionallyimmortalized rat brain capillary endothelial celllines (TR-BBB). Arch Pharm Res 28:443–450.
14. Pardridge WM. 2005. Molecular biology of theblood-brain barrier. Mol Biotechnol 30:57–70.
15. Sung JH, Yu KH, Park JS, Tsuruo T, Kim DD, ShimCK, Chung SJ. 2005. Saturable distribution oftacrine into the striatal extracellular fluid of therat: Evidence of involvement of multiple organiccation transporters in the transport. Drug MetabDispos 33:440–448.
16. Murakami H, Sawada N, Koyabu N, Ohtani H,Sawada Y. 2000. Characteristics of choline trans-port across the blood-brain barrier in mice: Correla-tion with in vitro data. Pharm Res 17:1526–1530.
17. Koepsell H, Lips K, Volk C. 2007. Polyspecificorganic cation transporters: Structure, function,physiological roles, and biopharmaceutical implica-tions. Pharm Res 24:1227–1251.
18. Tatsuta T, Naito M, Oh-hara T, Sugawara I, TsuruoT. 1992. Functional involvement of P-glycoproteinin blood-brain barrier. J Biol Chem 267:20383–20391.
19. Kusuhara H, Suzuki H, Terasaki T, Kakee A,Lemaire M, Sugiyama Y. 1997. P-Glycoproteinmediates the efflux of quinidine across the blood-brain barrier. J Pharmacol Exp Ther 283:574–580.
20. Kim KP, Ahn SH, Sung JH, Koo TS, Byun MJ, KimDD, Shim CK, Chung SJ. 2006. Application of asample pooling method for the accelerated assess-ment of the rate of uptake of drugs by the brain inrats. J Pharm Pharmacol 58:837–846.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009
21. Homma M, Suzuki H, Kusuhara H, Naito M,Tsuruo T, Sugiyama Y. 1999. High-affinity effluxtransport system for glutathione conjugates on theluminal membrane of a mouse brain capillaryendothelial cell line (MBEC4). J Pharmacol ExpTher 288:198–203.
22. Wakabayashi K, Nakagawa H, Tamura A, KoshibaS, Hoshijima K, Komada M, Ishikawa T. 2007.Intramolecular disulfide bond is a critical checkpoint determining degradative fates of ATP-bindingcassette (ABC) transporter ABCG2 protein. J BiolChem 282:27841–27846.
23. Nourian Z, Mulvany MJ, Nielsen KB, Pickering DS,Kristensen T. 2008. The antagonistic effect of anti-psychotic drugs on a HEK293 cell line stablyexpressing human alpha1A1-adrenoceptors. Eur JPharmacol 596:32–40.
24. Shang T, Uihlein AV, Van Asten J, Kalyanaraman B,Hillard CJ. 2003. 1-Methyl-4-phenylpyridiniumaccumulates in cerebellar granule neurons viaorganic cation transporter 3. J Neurochem 85:358–367.
25. Li J, Zhou S, Huynh H, Chan E. 2005. Significantintestinal excretion, one source of variability inpharmacokinetics of COL-3, a chemically modifiedtetracycline. Pharm Res 22:397–404.
26. Xie Z, Liao Q, Xu X, Yao M, Wan J, Liu D. 2006.Rapid and sensitive determination of donepezil inhuman plasma by liquid chromatography/tandemmass spectrometry: Application to a pharmacoki-netic study. Rapid Commun Mass Spectrom 20:3193–3198.
27. Silbermann MH, Boersma AW, Janssen AL, Sche-per RJ, Herweijer H, Nooter K. 1989. Effects ofcyclosporin A and verapamil on the intracellulardaunorubicin accumulation in Chinese hamsterovary cells with increasing levels of drug-resis-tance. Int J Cancer 44:722–726.
28. Kurata Y, Ieiri I, Kimura M, Morita T, Irie S, UraeA, Ohdo S, Ohtani H, Sawada Y, Higuchi S, OtsuboK. 2002. Role of human MDR1 gene polymorphismin bioavailability and interaction of digoxin, a sub-strate of P-glycoprotein. Clin Pharmacol Ther72:209–219.
29. Lash LH, Putt DA, Xu F, Matherly LH. 2007. Role ofrat organic anion transporter 3 (Oat3) in the renalbasolateral transport of glutathione. Chem BiolInteract 170:124–134.
30. Kullak-Ublick GA, Ismair MG, Stieger B,Landmann L, Huber R, Pizzagalli F, FattingerK, Meier PJ, Hagenbuch B. 2001. Organicanion-transporting polypeptide B (OATP-B)and its functional comparison with three otherOATPs of human liver. Gastroenterology 120:525–533.
31. Tahara H, Shono M, Kusuhara H, Kinoshita H,Fuse E, Takadate A, Otagiri M, Sugiyama Y.2005. Molecular cloning and functional analyses
DOI 10.1002/jps

MECHANISM OF DONEPEZIL TRANSPORT ACROSS THE BBB 19
of OAT1 and OAT3 from cynomolgus monkey kid-ney. Pharm Res 22:647–660.
32. AsabaH,HosoyaK,TakanagaH,OhtsukiS,TamuraE, Takizawa T, Terasaki T. 2000.Blood-brain barrieris involved in the efflux transport of a neuroactivesteroid, dehydroepiandrosterone sulfate, via organicanion transporting polypeptide 2. J Neurochem75:1907–1916.
33. Dresser MJ, Leabman MK, Giacomini KM. 2001.Transporters involved in the elimination of drugs inthe kidney: Organic anion transporters and organiccation transporters. J Pharm Sci 90:397–421.
34. Nakamura T, Yoshida K, Yabuuchi H, Maeda T,Tamai I. 2008. Functional characterization ofergothioneine transport by rat organic cation/car-nitine transporter Octn1 (slc22a4). Biol Pharm Bull31:1580–1584.
35. Kido Y, Tamai I, Ohnari A, Sai Y, Kagami T, Nezu J,Nikaido H, Hashimoto N, Asano M, Tsuji A. 2001.Functional relevance of carnitine transporterOCTN2 to brain distribution of L-carnitine andacetyl-L-carnitine across the blood-brain barrier.J Neurochem 79:959–969.
36. Okuda T, Haga T, Kanai Y, Endou H, Ishihara T,Katsura I. 2000. Identification and characterizationof the high-affinity choline transporter. Nat Neu-rosci 3:120–125.
37. Meno JR, Nguyen TS, Jensen EM, Alexander WestG, Groysman L, Kung DK, Ngai AC, Britz GW,Winn HR. 2005. Effect of caffeine on cerebral bloodflow response to somatosensory stimulation.J Cereb Blood Flow Metab 25:775–784.
38. Davies B, Morris T. 1993. Physiological parametersin laboratory animals and humans. Pharm Res10:1093–1095.
DOI 10.1002/jps
39. Matsui K, Taniguchi S, Yoshimura T. 1999.Correlation of the intrinsic clearance of donepe-zil (Aricept) between in vivo and in vitro stu-dies in rat, dog and human. Xenobiotica 29:1059–1072.
40. Slitt AL, Cherrington NJ, Hartley DP, Leazer TM,Klaassen CD. 2002. Tissue distribution and renaldevelopmental changes in rat organic cation trans-porter mRNA levels. Drug Metab Dispos 30:212–219.
41. Maeng HJ, Kim MH, Jin HE, Shin SM, Tsuruo T,Kim SG, Kim DD, Shim CK, Chung SJ. 2007.Functional induction of P-glycoprotein in theblood-brain barrier of streptozotocin-induced dia-betic rats: Evidence for the involvement of nuclearfactor-kappaB, a nitrosative stress-sensitive tran-scription factor, in the regulation. Drug MetabDispos 35:1996–2005.
42. Kusuhara H, Sugiyama Y. 2005. Active effluxacross the blood-brain barrier: Role of the solutecarrier family. NeuroRx 2:73–85.
43. Shiraishi T, Kikuchi T, Fukushi K, Shinotoh H,Nagatsuka S, Tanaka N, Ota T, Sato K, HiranoS, Tanada S, Iyo M, Irie T. 2005. Estimation ofplasma IC50 of donepezil hydrochloride for brainacetylcholinesterase inhibition in monkey usingN-[11C]methylpiperidin-4-yl acetate ([11C]MP4A)and PET. Neuropsychopharmacology 30:2154–2161.
44. Okamura N, Funaki Y, Tashiro M, Kato M, Ishi-kawa Y, Maruyama M, Ishikawa H, Meguro K,Iwata R, Yanai K. 2008. In vivo visualization ofdonepezil binding in the brain of patients withAlzheimer’s disease. Br J Clin Pharmacol 65:472–479.
JOURNAL OF PHARMACEUTICAL SCIENCES 2009




















