
Nicotinic receptor modulation of neurotransmitter release in the cerebellum
Upload
independentCategory
view
1download
0
Evidence in Postmortem Brain Tissue for Decreased Numbers of Hippocampal Nicotinic Receptors in Schizophrenia
Robert Freedman, Michael Hall, Lawrence E. Adler, and Sherry Leonard
This study tests the h3pothesis that nicotinic cholinergic receptors, including those sensitive to the antagonist c~-bungarotoxin, are decreased in the hippocampus of schizophrenics. The hypothesis is derived from the finding that e~-bungarotoxin causes a defect in the inhibitory gating of auditor3"-evoked potentials in laboratory animals that resembles a defect in auditory sensory gating observed in schizophrenics. Nicotine transiently normalizes this psychophysio- logical deficit in schizophrenic patients. Postmortem brain tissue was obtained from eight schizophrenic and eight age-matched nonschizophrenic subjects. Sections of the hippocampus were labeled with [~e~l]-c~-bungarotoxin and imagined by autoradiography. Binding of the nicotinic agonist [-~H]-cytisine was determined in tissue homogenates, c~-Bungarotoxin labeled a population of putative interneurons in the hippocampus, primarily in the dentate gyrus and the CA3 region qlAmmon's horn. This labeling was significantly decreased in the tissue from the schizophrenic patients, with seven or eight patients below the range of the nonschizophrenic subjects. There n'as also a significant decrease in the binding qf ~Ttisine. The results were not related to general&ed hippocampal cell loss, drug exposure at time of death, or smoking history. This initial study suggests that schizophrenic patients have jewer nicotinic receptors in the hippocampus, a condition which may lead to fidlure qf cholinergic activation ~( inhibitor3' interneurons, man,lest clinically as decreased ,~ating q/'re~g~onse to sensory stimulation.
Key Words: Nicotinic receptors, schizophrenia, hippocampus, o~-bungarotoxin, cytisine
Introduction Recent findings from several types of investigation suggest a role for nicotinic cholinergic mechanisms in schizophre- nia. Nicotine, a cholinergic agonist, normalizes two psy- chophysiological deficits found in schizophrenia: an audi- tory sensory gating deficit (Adler et al 1992) and disordered
From the Deparlments of Psychiatry (RF, LEA). Pharmacology ( RF. SL I, and Pediat- rics (MHI, Denver Veterans Administration Medical Center and University o~ Colorado Health Sciences Center, Denver, CO.
Address reprint requests lo Robert Freedman, M.D. Depavtmem oi Psychiatr~ C-268-71, University of Colorado Health Sciences Cenler. 4200 East Ninlh Avenue, Denvcr, Colorado 80262.
Received March 7, 1994: revised September 12, 1994.
smooth pursuit eye movements (Klein and Andersen 1991 ). Schizophrenic subjects and many of their relatives fail to diminish the cerebral evoked response to repeated auditory stimuli (Adler et al 1985; Judd et al 1992). This failure of sensory gating may lead to the distractibility and hypervigi- lance characterized clinically in psychotic patients (Ven- ables 1964). Animal models of sensory gating point to a critical role ['or cholinergic innervation from the medial septal nucleus to the hippocampus (Vinogradova 1975). In humans, the hippocampus is a likely source of one of the evoked potential waves, P50, that fails to habituate in schiz- ophrenic subjects (Goff et al 1980). Pharmacological stud-
© 1995 Societ~r of Biological Psychiatry 0006-3223/95/$09.50 SSDI 0006 3223194)00252-X

Nicotinic Receptors in Schizophrenia BIOL PSYCHIATRY 23 1995:38:22-33
ies suggest that the cholinergic receptor in the hippocampus critical for inhibitory gating of auditory response is an o~-bungarotoxin-sensitive nicotinic receptor (Luntz- Leybman et al 1992). oL-B ungarotoxin, isolated from snake venom, binds to the neuromuscular junction and to a subset of low-affinity brain nicotinic cholinergic receptors (Schoepfer et al 1990). Antimuscarinic agents such as sco- polamine and antagonists of high-affinity nicotinic recep- tors, such as K-bungarotoxin and mecamylamine, do not block the habituation of auditory evoked responses in ani- mals, whereas a-bungarotoxin and curare, another antago- nist of neuromuscular transmission and of similar nicotinic receptors in the brain, are both effective blockers (Luntz- Leybman et al., 1992).
Based on these animal data, which indicate that choliner- gic blockade causes a sensory dysfunction in animals simi- lar to that seen in schizophrenia, nicotine gum was adminis- tered in a double-blind, placebo-controlled experiment to relatives of schizophrenics who shared the trait of abnormal auditory sensory gating. Nicotine, which stimulates cholin- ergic receptors, transiently' normalized the subjects" evoked potential gating (Adler et al 1992). Schizophrenics who regularly smoked cigarettes showed a similar normalization of P50 gating alter smoking (Adler et al 1993). Cigarette smoking had earlier been shown to normalize disordered smooth pursuit eye movements, a second psychophysiolog- ical abnormality common in schizophrenics and their rela- tives (Klein and Andersen 1991). Together, these observa- tions raise the possibility that a nicotinic cholinergic dysfunction is responsible liar some of the psychophysio- logical defects observed in schizophrenia.
Cholinergic neurons have been examined directly in brain tissue obtained postmortem from schizophrenic patients to determine if there are pathological changes related to schizophrenia. There are abnormalities in brain- stem cholinergic neurons of the peduncular pontine nu- cleus (Karson et al 1993), but the neurons of the nucleus basalis of Meynert, the principal source of cerebral acetyl- choline, do not appear to be diminished in schizophrenia (EI-Mallakh et al 1991). Choline acetyltransferase, the enzyme that synthesizes acetylcholine, was not found to be diminished in the hippocampus and other lorebrain regions (Karson et al 1993). Thus, cholinergic innervation of the forebrain appears to be normal in schizophrenia. A possible mechanism ofcholinergic dysfunction in schizo- phrenia is a deficit in postsynaptic receptors tbr acetyl- choline on neurons that receive cholinergic innervation. Muscarinic cholinergic receptors have been found to be increased in schizophrenics, an effect perhaps linked to treatment with antimuscarinic drugs for extrapyramidal side effects (Watanabe et al 1983). Nicotinic cholinergic receptors have not been previously examined in postmor- tem brain tissue of schizophrenics.
A growing family of genes lbr cholinergic nicotinic re- ceptor subunits are being cloned. The prototype is the cho- linergic receptor of the neuromuscular junction, which is composed of five subunits: two identical c~-1 subunits, two identical 13-1 subunits, and a ysubunit, arranged in a pen- tamer to form an ion channel in the center. The receptor is activated when two molecules of acetylcholine or nicotine bind to it, probably at the interfaces between the a and 13 subunits. The binding of the cholinergic ligands causes a contbrmational change in the molecule that opens the chan- nel to cations. A series of neuronal nicotinic receptor genes has been cloned, based on pharmacological and genetic similarity to the neuromuscular receptors. It had long been recognized that nicotine and acetylcholine bind with high at~nity to specific receptors in the brain, but that a-bungar- otoxin, the most specific antagonist of cholinergic neuro- transmission at the neuromuscular junction, binds to an- other population of receptors, which has a lower affinity tbr nicotine and acetylcholine. Alpha subunits 2-8 and beta subunits 2-4 have been reported by various authors to form nicotinic cholinergic receptors in neurons. The most com- mon high-affinity nicotinic receptor appears to have three e~-4 subunits and two 13-2 subunits (Luetje et al 1990), The most common lower-affinity nicotinic receptors, which also binds c~-bungarotoxin, appears to be a pentamer composed of five identical e~-7 subunits (Schoepfer et a11990; Seguela et al 1993). o~-8 subunits also form a receptor that binds c~-bungarotoxin, but this receptor subunit has so far been found only in chickens (Schoepfer et al 1990; Gerzanich et al 1994). The majority of high-affinity nicotinic receptors appear to be concerned with the regulation of the release of neurotransmitters from their terminals (Wonnacott et al 1990), They have been found to regulate the release of acetylcholine itself, as well as norepinephrine, dopamine, glutamate, and y-aminobutyric acid (GABA).
Receptors that bind c~-bungarotoxin have a distribution in the brain that is different from these higher-affinity nicotinic receptors (Harfstrand et al 1988). In the hippo- campus, c~-bungarotoxin binds most intensely to inhibit- ory interneurons in the CA3 region of Ammon's horn and in the hilus of the dentate gyrus (Freedman et al 1993). c~-Bungarotoxin-sensitive receptors have been found on inhibitory interneurons in other brain regions as well (de la Garza et al 1987). Inhibitory interneurons with a- bungarotoxin-sensitive receptors are possible candidates /'or mediation of habituation of auditory responses in the hippocampus. Activation of the interneurons via their o~- bungarotoxin-sensitive receptors would increase the in- hibitory synaptic input to pyramidal neurons and thereby diminish the responsiveness of these pyramidal neurons to sensory stimulation. Enhancement of GABA release by activation of the high-affinity nicotinic synapses on the presynaptic terminals of these interneurons would be a

24 BIOL PSYCHIATRY R. Freedman et al 1995;38:22-33
complementary mechanism by which cholinergic neuro- transmission could also facilitate inhibitory gating in the hippocampus.
Another area in which o~-bungarotoxin binding may be relevant to the pathophysiology of schizophrenia is the ven- tral lateral geniculate. The physiological role of o~-bungaro- toxin-sensitive receptors in the ventral lateral geniculate is not known, but this nucleus is one of the neuronal centers responsible for the generation of smooth pursuit eye move- ments (Denny-Brown and Fisher, 1976). Thus, cx-bungaro- toxin-sensitive cholinergic receptors are located in at least two areas where their dysfunction could cause psychophys- iological disturbances commonly observed in schizophre- nia. ot-Bungarotoxin also binds to scattered neurons in many cortical areas, including the cingulate, and in other subcorti- cal areas, including several of the mamillary nuclei, the ventral and dorsal hypothalamic nuclei, the superior colli- culus, the amygdala, the supraoptic nucleus, the endopyri- form nucleus, and the superior colliculus (Harfstrand et al 1988). To what extent these and other areas that contain receptors labeled by cx-bungarotoxin might participate in the pathophysiology of schizophrenia is uncertain.
Clinical importance of these nicotinic receptor mecha- nisms is suggested by the behavior of schizophrenic patients themselves. Although smoking has been associated with other psychiatric disorders, heavy smoking is particularly common in schizophrenics (Goffet al 1992: Masterson and O'Shea 1984: Glassman 1993). The aim of the present study is to determine if there is decreased expression of nicotinic cholinergic receptors in the hippocampus in postmortem brain tissue from schizophrenic patients. Two ligands were examined, a-Bungarotoxin was chosen because of evidence for the role of o~-bungarotoxin-sensitive receptors in psy- chophysiological dysfunction in schizophrenics. The alka- loid cytisine, a ganglionic agonist, was chosen to measure the high-affinity nicotine receptors (Hall et al 1993: Flores et al 1992), although at higher concentrations it activates a-7 receptors as well (Seguela et al 1993; Anand et a11993).
M e t h o d
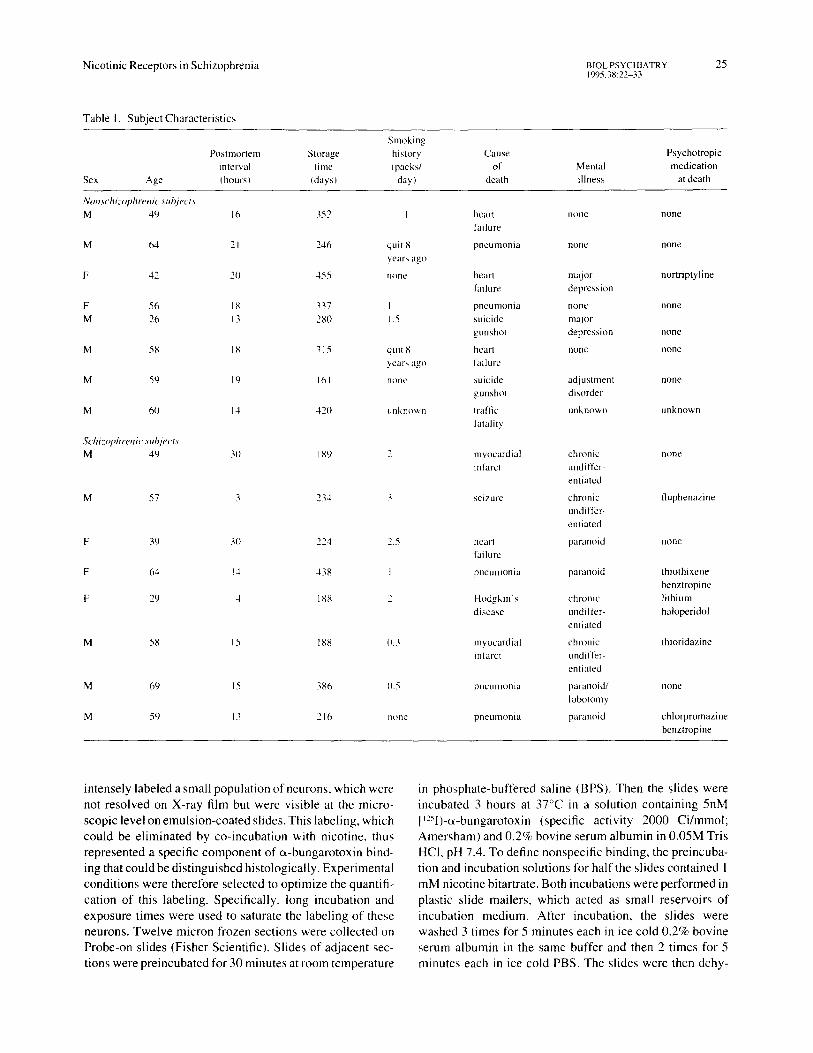
Eight subjects were identified at autopsy who had histories of schizophrenia in their medical record. The diagnosis was confirmed to meed DSM-III-R criteria (APA 1987) through interviews with relatives and treating clinicians and review of other psychiatric records. Other clinical characteristics are shown in Table 1. All the schizophrenic patients had been treated with neuroleptics at some time during their life. One subject had a closed frontal lobotomy 47 years prior to death; typical psychotic symptoms, including auditory hal- lucinations and paranoid delusions, had continued postop- eratively. Postmortem examination showed that the lesion had only affected the caudate nucleus and had spared the frontal lobes entirely.
Eight nonschizophrenic subjects were matched with the schizophrenic subjects for age and, for all but one pair, for sex. For all but one, a relative was interviewed to rule out a personal or family history of schizophrenia. One brain came from an automobile accident fatality through the National Disease Resource Interchange (Philadelphia, PA); no other information was available. Four of the eight nonschi- zophrenic subjects had no known history of mental illness. Diagnoses of the other three are shown in Table 1. The mean age of the nonschizophrenic subjects was 51.8 _+ 12.5 years, not significantly different from the schizophrenic subjects, whose mean age was 53.0 + 13.3 years. The mean age difference between subject pairs was 1.5 _+ 2.2 years.
Tissue Preservation and Examination
No subject was unconscious for more than 1 day before death, and none had any clinical evidence of neurological illness, including stroke, dementia, or head injury. Tissue blocks were taken for neuropathological examination to confirm the absence of degenerative disease or infarct. For two nonschizophrenics and one schizophrenic, sufficient tissue was not available for cytisine binding. Tissue from appropriate, similarly aged subjects was substituted in these three cases.
The brain was removed from the skull within 30 hours of death. The mean postmortem interval was 17.8 -+ 13.9 hours for schizophrenics, which was not significantly dif- ferent from 17.2 + 3.1 hours for nonschizophrenics. The brain was sliced in coronal sections at I-cm intervals and placed on chilled aluminum plates. The hippocampus was then dissected from the fifth coronal slice. One side was selected randomly for histological examination and the other was used for receptor binding assays. The dissected tissue was immediately snap frozen in dry ice snow and stored at -70°C until use. Storage time for the nonschizo- phrenics, 321 + 94 days, was not significantly longer than storage time for the schizophrenics: 258 -+ 98 days. o~-Bun- garotoxin binding has been specifically shown to remain intact in brains collected within these parameters (Davies and Feisullin 1981 ).
[ ~:~I]- c*-Bungarotoxin Labeling
An initial study of a-bungarotoxin binding in homogenates of human hippocampus revealed nonspecific binding often greater than 50% of total binding. Nonspecific binding was defined as binding in the presence of 1 mM nicotine bitar- trate. Another group reported similar results (Lang and Henke 1983). The high percentage of nonsignificant bind- ing precluded quantitative determination of binding sites. Imaging of human hippocampus by conventional quantita- tive autoradiography methods on X-ray film also revealed high nonspecific activity; however, as in an earlier study of rat hippocampus (Freedman et al 1993), o~-bungarotoxin
Nicotinic Receptors in Schizophrenia BIOL PSYCHIATRY 25 1995:38:22-33
Table I. Subject Characteristics
Sex Age
Smoking Postmortem Storage history
interval lime (packs/ (hours) (days) day)
Cause Psychotropic of Mental medication
death illness at death
Nonschi=ophrenic subject~ M 49 16 352
M 64 21 246
F 42 2O 455
F 56 18 337 M 26 13 280
M 58 18 315
M 59 19 161
M 60 14 420
Schiztq#lrenic sut~jects M 49 30 189
M 57 3 234 3 seizure
F 39 30 ~'~4 . . . . 5
F 64 14 438 I
F 29 4 188 2
M 58 15 188 0.3
M 69 15 386 0.5
M 59 13 216 none
] h e i l r l n o n e n o n e
failure
quit 8 pneumonia mine none years ago
none heart major nortriptyline failure depression
I pneurnonia none none 1.5 suicide major
gunshot depression none
quit 8 hearl none none years ago failure
none suicide adjustment none gunshot disorder
unknown traffic unknown unknown fatality
2 myocardial chronic none intktrct undi ffer-
entiated
chronic fluphenazine undHfizr- entiated
hearl paranoid none tailure
pneumonia paranoid thiothixene benztropine
Hodgkin's chronic lithium disease undifter- haloperidol
entiated
chronic undiffer- entiated
pneumonia paranoid/ none Iobotomy
pneumonia paranoid chlorpromazine benztropine
myocardial Ihioridazine inlarcl
in tense ly labeled a smal l popu la t ion of neurons , wh ich were
not reso lved on X-ray film but were v is ib le at the micro-
scopic level on e m u l s i o n - c o a t e d slides. Th i s label ing, wh ich
could be e l imina ted by co - incuba t ion wi th nicot ine , thus
r ep resen ted a specif ic c o m p o n e n t of oL-bungarotoxin bind-
ing that cou ld be d i s t i ngu i shed his to logical ly . Expe r imen ta l
cond i t ions were therefore selected to op t imize the quant i f i -
ca t ion of this label ing. Specif ical ly , long incuba t ion and
exposure t imes were used to saturate the labe l ing o f these
neurons . T w e l v e m i c r o n f rozen sec t ions were co l lec ted on
P robe -on sl ides (F isher Scientif ic) . Sl ides of ad jacen t sec-
t ions were p re incuba t ed for 30 minu tes at room t empera tu re
in phospha t e -bu f f e r ed sal ine (BPS) . T h e n the s l ides were
incuba ted 3 hours at 37°C in a solut ion con ta in ing 5 n M
[~251)-o~-bungarotoxin (specif ic ac t iv i ty 2000 C i /mmol ;
A m e r s h a m ) and 0 .2% b o v i n e se rum a l b u m i n in 0 . 0 5 M Tris
HCI, pH 7.4. To def ine nonspec i f ic b ind ing , the p re incuba -
t ion and incuba t ion so lu t ions for ha l f the sl ides con t a ined 1
m M nico t ine bi tar trate. Bo th incuba t ions were pe r fo rmed in
plast ic slide mai lers , wh ich acted as small rese rvo i r s of
incuba t ion med ium. Af te r incuba t ion , the s l ides were
w a s h e d 3 t imes for 5 m inu t e s each in ice cold 0 .2% b o v i n e
se rum a lbumin in the same buffer and then 2 t imes for 5
minu tes each in ice cold PBS. The sl ides were then dehy-

26 BIOL PSYCHIATRY R. Freedman et al 1995:38 :22 ~
drated in 75% and 95c/, ethanol, dried, and dipped in autora- diographic emulsion diluted 1:1 in water (NTB-2, Kodak, Rochester, New York). The slides were exposed to the emulsion for 5 weeks at -20°C. They were then developed in Kodak D-19 diluted 1:1 in water, fixed, and lightly coun- terstained with cresyl violet to help identify bippocampal regions.
The slides were imaged on a computer-based vide() imag- ing system (Magiscan, Joyce-Loebel Ltd., Tyne and Wear, U.K.) through a Zeiss Universal Microscope (Zeiss Instru- ments, Newark N J), using a 10x. objective. Light intensity and camera settings were chosen to maximize the imaging of labeled neurons. The same settings were used for all subjects. Selection of areas to be imaged was performed blind to subjects" diagnoses. A computer algorithm deter- mined the fraction of the video field labeled by c~-bungaro- toxin. Single-labeled pixels were not counted, to minimize detection of diffuse nonspecific labeling. The values used for the analysis of each subject 's labeling in each hippocam- pal region were measured from five adjacent artifact-free fields (area of each field 1.95 mm~). Adjacent sections were used for total labeling and lk)r nonspecific labeling, for which incubation in 1 mM nicotine was used to block nico- tinic receptors. The difference in values was used as the measure of specific labeling.
Student 's t test was used to determine if the differences between the paired total and nonspecific measurements were statistically significant. Seven of the CA3 region mea- surements and seven of the dentate gyrus measurements were significant for the eight nonschizophrenic subjects (p < .05). A single-tailed probability was used because of the assumption that total binding is greater than nonspecific. Only one of the CA3 region measurements and five of the dentate gyrus measurements were significant for the schiz- ophrenic subjects. Four of the CA I region measurements were significant for both groups. Thus, the measuren)ent strategy was sufficient to detect significant specific c~-bun- garotoxin labeling in the nonschizophrenic subjects. Signif- icant labeling was not detected in most schizophrenic sub- jects, consistent with their much lower levels of labeling: however, all values were used for subsequent statistical analysis, rather than assuming that failure to find a signifi- cant difference represented a true absence of labeling in subjects with low values. The mean proportion of nonspeci- fic labeling was 17.6% + 20.1% of total labeling in the dentate hilus and 9.0% _+ 9.49~- in CA3 for the nonschizo- phrenics, and 66.0c/c _+ 31.6c/c in the dentate hilus and 53.8 _+ 32.4% in CA3 for the schizophrenics; however, the abso- lute amount of nonspecific labeling was more similar be- tween the two groups. For the nonschizophrenics, the mean nonspecific labeling in the dentate hilus, expressed as a percentage of the area of the region of intercst, was 0.19c~ _+ 0.56%, compared to ().2()r7~ _+ 0.249~ for schizophrenics.
For CA3, the mean nonspecific labeling was 0.02% -+ 0.02% for nonschizophrenics and 0.08% _+ 0.11% for schizophrenics.
Cell Counting Cells were counted in the dentate hilus, and the CA I and CA3 regions. The granule cells of the dentate gyms fre- quently overlap and thus cannot be separated for counting. Moreover, they did not appear to be labeled by oL-bungaro- toxin. Cells were counted through a 40x objective in five 0.1 ram-' felds. Only cells with identifiable nucleoli were counted to minimize artifacts due to partial voluming of the cell body. Staining of neurons by cresyl violet is diminished alter treatment of the slides with emulsion; therefore, adja- cent sections that were not used for autoradiography were stained for cell counting. These sections are marked "cre- syl" in Figures I and 2.
[ +H]-Qvtisine Binding
One to 1,5-gram portions of the hippocampus were dis- sected from the brain specimens prior to freezing. Gross inspection confirmed that the tissue contained dentate and Ammon ' s horn. The frozen tissue was homogenized for 10s with a Brinkman polytron in a 4°C homogenization buffer which consisted of 50 mM Tris-HC1 (pH 7.0), 120 mM NaCI, 5 mM KCI, 1 mM MgCI> and 2.5 mM CaCL. The homogenate was centrifuged for 10 rain at 40,000 × g: the resulting pellet was resuspended in buffer and centrifuged again. The wash process was repeated twice, and the pellet was resuspended in sufficient homogenization buffer to give a protein concentration of 5-10 mg/ml. The protein content of the original brain tissue ranged from 2.5~t.5% of wet weight. Saturation analyses were performed in dupli- cate in 250 ixl containing buffer, tissue homogenate, and varying concentrations of [+H]-cytisine (Dupont/New En- gland Nuclear). Nicotine bitartrate (1 raM) was added to halt" the tubes to determine nonspecific association of the ligand. The concentration range of PHI-cytosine was 40 pMM-0 nM. The incubation was al lowed to reach equilib- rium (60 rain at 4c'C) and then filtered over glass fiber filters (Schleicher & Schuell 30), which had been pre- soaked in 0.3% polyethylenimine to reduce associat ion of the [ 'H]-cyt is ine with the filters. The filters were washed twice with 2.5 ml of ice cold buffer, vacuum dried, and counted in a scintil lation counter. The dissociat ion con- stant (K,,) and the B ...... for each subject were determined using nonlinear regression (Graphpad-lnplot , Graphpad Software, San Diego, CA). Protein concentrat ions were determined using a colorimetric assay (Pierce Chemical , Chicago, IL). Other details of this method have been previously descr ibed (Hall et al 1993).
N i c o t i n i c R e c e p t o r s in S c h i z o p h r e n i a BIOL PSYCHIATRY 2 7 1995:38:22 33
t
4r I~ e b
¥
:~"
T %
¢.
.2 ~: 7 ' ~"5 . t o t a l
,~ ?Ir
t ~ J l ~ I I F " " " - * *
|
o : nne
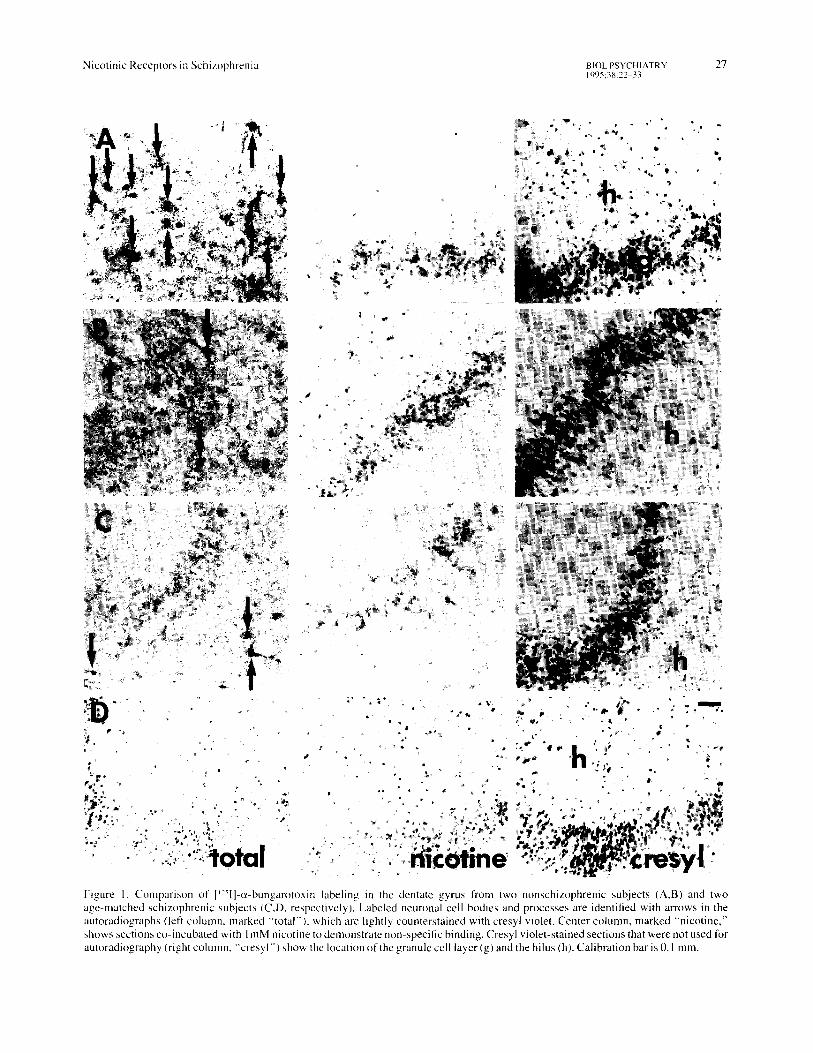
Figure 1. Comparison of ['~'I]-c~-bungarotoxin labeling in the dentate gyrus from two nonschizophrenic subjects (A,B) and two age-matched schizophrenic subjects (C~D, respectively). Labeled neuronal cell bodies and processes are identified with arrows in the autoradiographs (left column, marked "total" ), which are lightly counterstained with cresyl violet. Center column, marked "nicotine," shows sections co-incubated with I mM nicotine to demonstrate non-specific binding. Cresyl violet-stained sections that were not used for autoradiography (right column, "'cresyl'" ) show the location of the granule cell layer (g) and the hilus (h). Calibration bar is 0.1 ram.

28 BIOL PSYCHIATRY R. Freedman et al 1995 ;38:22-33
£
. f
° •
7 =
J ~ 4
\
: ~" a , '-
Figure 2. Comparison of [~25I]-c~-bungarotoxin labeling in the CA3 region from a nonschizophrenic subject (A) and an age-matched schizophrenic subject (B). Labeled cell bodies are identified with the arrows in the autoradiographs ("total"), which are counterstained with cresyl violet• Cresyl violet-stained sections ("'cresyl") that were not used for autoradiography show the location of the pyramidal cell layer (p). Two panels from subject A in the upper right column are 4 times higher power images of individual neurons in other fields from the CA3 region from the same nonschizophrenic subject, which show labeling of principal dendrites as well as the cell body by a-bungarotoxin. The high-power panel from subject B, f:'om the same field shown at lower power in the left column, demonstrates labeling limited to the cell body, which was more commonly observed in the tissue from schizophrenic subjects. Bottom panels marked "nicotine" show sections from both subjects co-incubated with I mM nicotine to demonstrate nonspecific binding• Calibration bar is 0.1 ram.
Nicotinic Receptors in Schizophrenia BIOL PSYCHIATRY 29 1995:38:22-33
Statistical Analyses An analysis of variance (ANOVA) was used to compare (x-bungarotoxin labeling in the two subject groups across the three hippocampal regions. Subsequent comparisons were performed using Tukey 's HSD. Chi-square was used to compare the distribution of schizophrenic values to those of the nonschizophrenics. Student's t test was used to com- pare cytisine binding between the two groups.
R e s u l t s
Histological Description [~25I}-(x-bungarotoxin labeled scattered cells throughout the hippocampus. In nonschizophrenic subjects, the labeled cells were large neurons, typically located in the dentate gyms and in the CA3 region of Ammon's horn. In the dentate, they were often found in the hilus, on the interior margin of the granule cell layer, with a few cells deeper in the hilus (Figure 1: A, B). In the CA3 region, they were more prominent outside stratum pyramidale (Figure 2A). Smaller cells were also found occasionally in the CA I region, but not to the extent seen in the CA3 region and the dentate. Generally, the cell body was prominently labeled, and often heavy labeling could be seen on major dendrites as well (Figure 2A). The granule cells and pyramidal cells them- selves were not prominently labeled by c(-bungarotoxin, nor was any major fiber tract labeled.
e~-Bungarotoxin binds not only to specific nicotinic re- ceptors in the tissue, but to other sites as well. Because there is specific as well as nonspecific binding, the autoradiogra- phic images are marked "total" in Figures 1 and 2. The specific binding is blocked by competition with a high dose of nicotine, so that the slides which are preincubated with 1 rnM nicotine, marked "nicotine" in Figures 1 and 2, show only the nonspecific part of the binding. For both the non- schizophrenics and the schizophrenics, the nonspecific binding was a moderately dense diffuse background over all cell bodies and to a lesser extent over the neuropil, which appears in both the "total" and the "nicotine" sections. Dense labeling of neurons and their processes was seen in the "total" sections, but not in the "nicotine" sections, and thus represents the specific labeling of nicotinic receptors by (~-bungarotoxin,
In schizophrenics, there were generally fewer such la- beled cells. Although some cells could be found in both the dentate and the CA3 region, as well as occasionally in the CA1 region, they were less frequent than those found in the nonschizophrenic subjects (Figure 1C, D; Figure 2B). In addi- tion, the cells appeared to be less heavily labeled, and labeling did not often extend to dendritic processes in schizophrenics as it did in nonschizophrenic subjects (Figure 2B).
( -
o o
¢.-
.El
. E Q.
.El
"10
..Q
E .o
r~
1 . 5 - •
1 . 4 -
1 .3 -
1 . 2 -
1 . 1 - •
1 . 0 -
0 . 9 -
0 . 8 - l i
0 . 7 -
0 . 6 - o
0 . 5 -
0 . 4 -
0 . 3 "
0.2 - • • I
0.1"
0 . 0 -
- - D e n t a t e * *
* n o n - s c h i z o p h r e n i c
o s c h i z o p h r e n i c
~" _.o I • ,,~ ~ '~
CA3* CA1
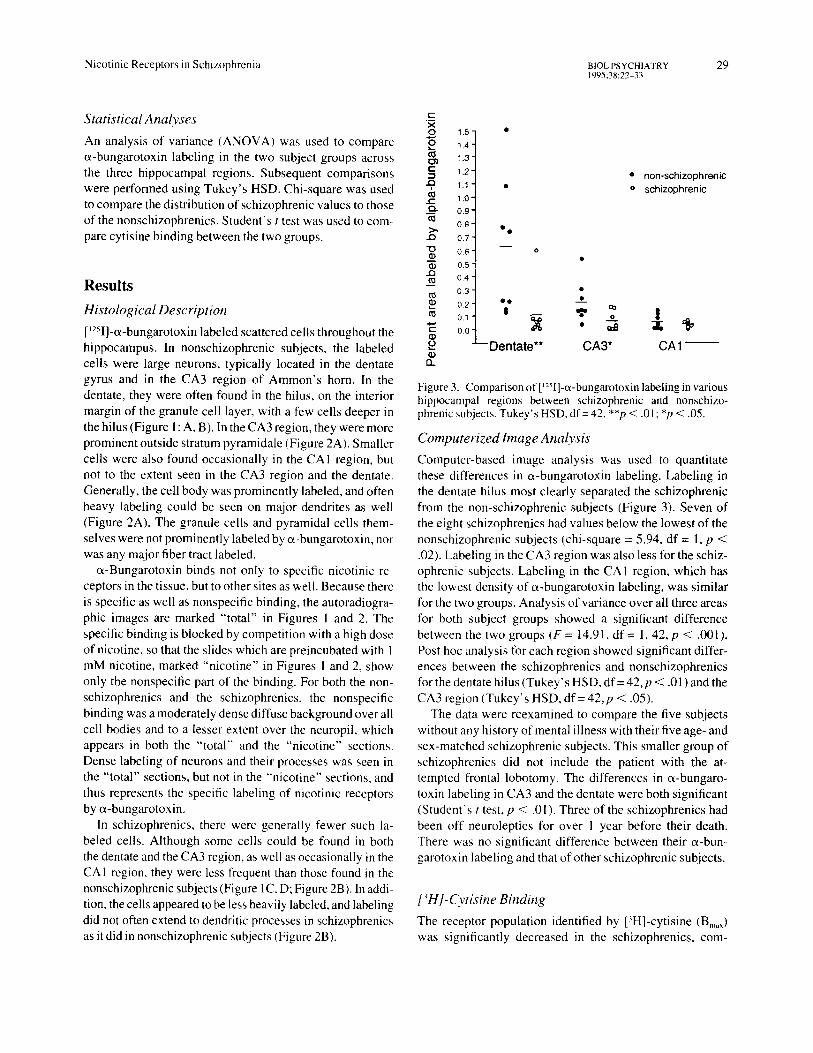
Figure3. Comparison L,~ . " - of [ - I]-(x-bungarotoxm labehng m various hippocampal regions between schizophrenic and nonschizo- phrenic subjects. Tukey's HSD, df= 42, **p < .01 ; *p < .05.
Computerized Image Analysis Computer-based image analysis was used to quantitate these differences in et-bungarotoxin labeling. Labeling in the dentate hilus most clearly separated the schizophrenic from the non-schizophrenic subjects (Figure 3). Seven of the eight schizophrenics had values below the lowest of the nonschizophrenic subjects (chi-square = 5.94, df = 1, p < .02). Labeling in the CA3 region was also less for the schiz- ophrenic subjects. Labeling in the CA1 region, which has the lowest density of e~-bungarotoxin labeling, was similar for the two groups. Analysis of variance over all three areas for both subject groups showed a significant difference between the two groups (F = 14.91, df = 1, 42, p < .001). Post hoc analysis for each region showed significant differ- ences between the schizophrenics and nonschizophrenics for the dentate hi lus (Tukey's HSD, df = 42, p < .01) and the CA3 region (Tukey' s HSD, df = 42, p < .05).
The data were reexamined to compare the five subjects without any history of mental illness with their five age- and sex-matched schizophrenic subjects. This smaller group of schizophrenics did not include the patient with the at- tempted frontal lobotomy. The differences in (x-bungaro- toxin labeling in CA3 and the dentate were both significant (Student's t test, p < .01 ). Three of the schizophrenics had been off neuroleptics for over 1 year before their death. There was no significant difference between their s-bun- garotoxin labeling and that of other schizophrenic subjects.
[-~H]- Cytisine Binding The receptor population identified by [~H]-cytisine (Bin.x) was significantly decreased in the schizophrenics, corn-

3 0 BIOL PSYCHIATRY R. F r e e d m a n et al 1995:38:22-33
7O f - -
*~ 60
50
-~ 40 E
' 4 - -
× 30
E m 20
.E 10
o
o
.Jt_ o
o
> . (D 0 Non-schizophrenic Schizophrenic subjects subjects"
Figure 4. Comparison of [~Hi-cytisine binding between schizo- phrenic and nonschizophrenic subjects: t = 2.51. df = 14,*p < .05.
pared to the nonschizophrenic subjects (t = 2.51, df = 14, p < .05; Figure 4). The K,~ values and Hill coefficients were not significantly different between the two groups. The mean Kd was 2. I 0 + 1.0 nM for the schizophrenics versus 2.2 +_ 1.8 nM for the nonschizophrenic subjects. Mean Hill coefficients were 0.89 _+ 0.09 for the schizophrenics and 0.92 _+ 0.13 for the nonschizophrenics, consistent with a single binding site.
Cell Numbers
To determine if there were differences in the overall neur- onal population between the schizophrenic and the non- schizophrenic brain specimens, sections adjacent to those used for c~-bungarotoxin labeling were examined after fixa- tion and staining with cresyl violet (Figures 1 and 2). Cell numbers in the dentate hilus, the CA3 region, and the CA 1 region were not significantly different between the two groups (Table 2). The cell numbers were not correlated with ~-bungarotoxin labeling or with cytisine binding.
Discuss ion
~-Bungarotoxin- labeled cells in the human hippocampus that appear similar in distribution to the population of inter- neurons labeled in the rat hippocampus (Freedman et al 1993); however, human hippocampus has a low density of labeling, compared to rats. This low density, combined with the nonspecific affinity of c~-bungarotoxin for other proteins, e.g., tachykinin receptors (Utkin 1989), precluded quantita- tive receptor binding studies in hippocampal homogenates. The labeling conditions and computerized image analysis technique used here were therefore directed towards measur- ing the most intensely labeled cells, whose labeling is clearly antagonized by nicotine. As in the rat, these cells were most prominent in the dentate hilus and the CA3 region, the areas of
the hippocampus that receive most of the cholinergic innerva- tion from the medial septal nucleus.
Seven of the eight schizophrenic brain samples showed less a-bungarotoxin labeling than the nonschizophrenics. The limited sample size available for these post mortem analyses tempers conclusions about the applicabili ty of these findings to other groups of schizophrenics. Further- more, within the present sample one schizophrenic subject has normal e~-bungarotoxin labeling, suggesting the pres- ence of heterogeneity with regard to the existence of a defect in expression of nicotinic receptors. Nevertheless, in light of the clinical and psychophysiological evidence for a deficit in nicotinic neurotransmission, this finding may point to a significant pathophysiological mechanism in many schizo- phrenic patients. Cytisine binding in homogenates of hippo- campus was also decreased in the brains from the schizo- phrenic subjects, which suggests that there is also a decrease in high-affinity nicotinic receptors. In many rat brain areas, these high-affinity receptors are associated with modulation of the release of neurotransmitters (Wonnacott et al 1990) Although high-affinity nicotinic receptors are rare in human hippocampus (Perry et al 1992; Hall et al 1993), they pre- sumably have the same role.
There are several possible alternative explanations for the differences in nicotinic receptors found between schizo- phrenic and nonschizophrenic brains. The hippocampus is one of the brain areas most likely to show significant cell loss in schizophrenics (Bogerts et al 1985; Jeste and Lohr 1989: Benes et al 199 ! b). Although findings are somewhat variant, the CA3 region and the dentate hilus contain one of the cell populations that may be decreased (Jeste and Lohr 1989). Therefore, cell counts of the CA3 pyramidal area and the dentate hilus, as well as the CA I pyramidal area, were performed to determine if decreased cell numbers in these areas were related to the decrease in o~-bungarotoxin label- ing. These were not stereological analyses of the entire cell population, but only a measure of the density of cells in tissue sections; however, there was no difference between the schizophrenic and nonschizophrenic group on this mea- sure; nor was there correlation between cell density and o~-bungarotoxin labeling or cytisine binding. Thus, the de- crease in nicotinic receptors does not appear to be a reflec- tion of a generalized loss of cell density.
A second possibili ty is that increased smoking in the schizophrenics decreased the number of e~-bungarotoxin receptors. Only one schizophrenic and two nonschizo- phrenics had never smoked; however, the two nonsmoking,
Table 2. Neuronal Numbers in 0.1 mm z Sections of Hippocampus
Subjects Dentate hilus CA3 region CA I region
Nonschizophrenic 11.2 + 4.0 18.4 + 7.4 14.2 _+ 3.5 Schizophrenic 15.3 + 6.8 16.4 _+ 5.4 15.0 + 4.9

Nicotinic Receptors in Schizophrenia BIOL PSYCHIATRY 31 1995;38:22-33
nonschizophrenic subjects were within the range of the smokers in their group, and the nonsmoking schizophrenic had one of the lowest values observed. Furthermore, smok- ing in humans is associated with increased, rather than decreased levels of high-affinity nicotinic receptors (Ben- well et al 1988). Additionally, animal studies demonstrate that chronic nicotine administration raises levels of c~-bun- garotoxin-sensitive receptors (Morley 1989). Thus, smok- ing behavior is an unlikely explanation tk)r the low c~-bun- garotoxin labeling and cytisine binding observed in the schizophrenics.
A third possible cause of the difference is the effects of the chronic exposure to neuroleptics and anticholinergics experienced by all the schizophrenic subjects. Three of the subjects had been drug free at least 1 year before their death. Like patients on drug at the time of death, these three sub- jects showed lower than normal a-bungarotoxin labeling and cytisine binding; however, because all of the subjects had some history of drug use, the effects of psychotropic medication exposure remains unknown.
A deficit in nicotinic receptors is only one of several abnormalities that has been demonstrated in postmortem examinations of the hippocampus and other brain areas in schizophrenia. Perhaps the most relevant to the results of the present study is the deficit in markers lk)r GABA-containing neurons reported in the hippocampus (Reynolds et al 1990), since these neurons have the highest concentration of c~- bungarotoxin labeling in the rat (Freedman et al 1993). Decreased numbers of GABAergic interneurons have also been noted in the cingulate cortex (Benes et al 1991a), but not in other cortical areas (Daviss and Lewis 1993). These findings raise the possibility that the deficit in nicotinic receptors observed in this study is actually a deficit in neurons that express most of these receptors, i.e., GABA- ergic interneurons. A developmental mechanism for an interneuron deficiency in certain cortical layers has also been proposed, based on postmortem findings that neurons containing nicotinamide-adenine-dinucleotide- phosphate(NADPH)-diaphorase have tailed to migrate from deeper layers of the cortical plate (Akbarian et al 1993). These NADPH-diaphorase-containing neurons are generally GABAergic interneurons (Valtschanoffet al 1993). It is thus possible that the diminished c~-bungaro- toxin-sensitive receptors are reflective of a pervasive de- velopmental deficit, a pathophysiological mechanism often proposed for schizophrenia: however, it also possi- ble that the developmental deficit is caused in part by a deficit in c*-bungarotoxin-sensitive receptors. The recep- tors are expressed early in brain development, particularly in primary sensory cortical areas as they are about to receive thalamic innervation (Fuchs 1989). Activation ofe~-bungar- otoxin-sensitive receptors has been proposed to regulate neurite outgrowth (Vijayaraghavan et al 1992).
c~-Bungarotoxin-sensitive receptors have a high rate of desensitization, i.e., loss of activity as a result of agonist stimulation (Schoepfer et al 1990), which may explain the ineffectiveness of attempts to treat schizophrenia with cho- linergic agonists. Desensitization is not a functional prob- lem if a large pool of receptors is available for activation. For example, less than 1% of the cholinergic receptors in the neuromuscular junction are activated during a muscle con- traction, so that only a small portion are in the desensitized state at any time. The muscle can thus be activated repeat- edly. If the pool of muscle receptors diminishes, as in myasthenia gravis, then the muscle cannot be activated more than a few times before all the receptors are desensi- tized, resulting in failure of muscle activation (Acheson et al 1948). If a similar physiology occurs in schizophrenia, be- cause of decreased numbers of brain c~-bungarotoxin recep- tors, then cigarette smoking may not be an effective treat- ment for schizophrenia because it activates cholinergic receptors only transiently. After this brief activation, the receptors may then become desensitized, so that the net effect is a loss of cholinergic neurotransmission. Attempts to prolong the effect through longer-acting agents such as physostigmine would only lead to more desensitization, which may explain why these treatments have been ineffec- tive in schizophrenia (Berger et al 1979). It is noteworthy that physostigmine appears to be more effective in psy- choses associated with illnesses other than schizophrenia, such as mania and Alzheimer's disease (Janowsky et al 1973: Cummings et al 1993). In Alzheimer's disease, there is a marked decrease in high-affinity nicotinic receptors, but no change in e-bungarotoxin binding (Lang and Henke 1983; Sugaya et al ! 990). A strategy to test if desensitization is responsible for deficit in sensory gating in schizophrenia, by allowing the receptors to resensitize during sleep, has shown preliminary evidence in support of this hypothesis (Griffith et a11993).
Mutations in the receptor also alter the rate of desensiti- zation (Revah et al 1991 ). Whether such mutations occur in humans and whether they might play a pathophysiological role in individuals with otherwise normal numbers of c~- bungarotoxin receptors, such as the schizophrenic patient with normal o~-bungarotoxin labeling found in this study, is unknown. The c~-7 nicotinic receptor subunit has been localized to chromosome 15, band q 14 (Chini et al 1994). Modest, but statistically significant evidence for genetic linkage to the deficit in P50 auditory evoked potential gating had been found previously at the chromosome 15q 14 site. The linkage was found in two of nine families that were selected for having multiple cases of schizo- phrenia and then phenotyped for both schizophrenia and the gating of the P50 auditory evoked potential (Coon et al 1993). A lod score smaller than that obtained for the deficit in P50 gating, but still statistically significant, was

32 BIOL PSYCHIATRY R. Freedman et al 1995:38:22-33
found for the sch izophren ia pheno type as well in these famil ies .
In conclusion, nicotinic receptors in the hippocampus and other brain regions may be involved in the pathophysiology of schizophrenia. Failure of cholinergic neurotransmission, because of alterations in these receptors, may be a mecha- n ism for d iminished habituation to sensory stimuli, ob-
served in schizophrenics psychophysiologically and mani- fested clinically as distractibility and hypervigilance.
This work was supported by the Veterans Administration Medical Re- search Service and USPHS Grants MH44212, MH38321, and MH00728. The assistance of Bernadette Sullivan, Dorothy Dill, Dr. Roberta Payne, DeAnne Torres, and Laura Zerbe is gratefully acknowledged.
R e f e r e n c e s
Acheson GH, Langohr JL, Stanbury JB (1948): Sensitivity of skeletal muscle to intra-arterial acetylcholine in normal and myathenic man. J Clin Invest 27:439-445.
Adler LE, Hoffer LJ, Griffith J, Waldo MC, Freedman R (1992): Normalization by nicotine of deficient auditory sensory gating in the relatives of schizophrenics. Biol Psychiatry 32:607-616.
Adler LE, Hoffer LJ, Wiser A, Freedman R (1993): Cigarette smoking normalizes auditory physiology in schizophrenics. Am J Psychiatr), 150:1856-- 1861.
Adler LE, Pachtman E, Franks R, Pacevich M, Waldo MC, Freed- man R (1985): Neurophysiological evidence for a defect in neuronal mechanisms involved in sensory gating in schizo- phrenia. Biol Psychiato' 17:1284-1296.
Akbarian S, Vinuela A, Kim J J, Potkin SG, Bunncy WE Jr, Jones EG (1993): Distorted distribution of nicotinamide-adenine di- nucleotide phosphate-diaphorase neurons in temporal lobe of schizophrenics implies anomalous cortical development. Arch Gen Psychiatry' 50:178-187.
American Psychiatric Association ( 1987): Diagnostic and Statisti- cal Manual o f Mental Disorders, 3rd ed rev. Washington DC: American Psychiatric Association.
Anand R, Peng X, Lindstrom J ( 1993): Homomeric and native e~ 7 acetylcholine receptors exhibit remarkably similar but non- identical pharmacological properties, suggesting that the na- tive receptor is a heteromeric protein complex. FEBS Lett 327:241-246.
Benes FM, McSparren J, Bird ED, Vincent SL, SanGiovanni JP (1991a): Deficits in small interneurons in schizophrenic cortex. Arch Gen Psychiatry., 48:996-[001.
Benes FM, Sorensen I, Bird ED ( 1991 b): Reduced neuronal size in posterior hippocampus of schizophrenic patients. Schi:ophr Bull 17:597-6-8,
Benwell MEM, Balfour DJK, Anderson JM ( 1988 ): Evidence that tobacco smoking increases the density of (-):-['H]-nicotine binding sites in human brain. JNeurochem 50:1243-1247.
Berger PA, Davis KL, Hollister LE (1979): Pharmacological in- vestigation of cholinergic mechanisms of psychosis in schizo- phrenia and manic psychosis. In Davis KL, Berger PA (eds). Brain Acetvlcholine and Neuropsychiatric Disease. New York: Plenum, pp 15-32.
Bogerts B, Meertz E, Schonfeldt-Bausch R (1985 ): Basal ganglia and limbic system pathology in schizophrenia: A morphome- tric study of brain volume and shrinkage. Arch Gen Psychiatry 42:784-791.
Chini B, Raymond E, Elgoyhen A, Moralli D, Balzaretti M, Heine- mann S ( 1994): Molecular cloning and chromosomal localiza- tion of the human alpha-7 nicotinic receptor subunit gone (CHRNA7). Genomics t 9:379-381.
Coon H, Plaetke R, Holik J, et al ( 1993): Use ofa neurophysiologi- cal trait in linkage analysis of schizophrenia. Biol Psychiatr), 34:277-289.
Cummings JL, Gorman DG, Shapira J (1993): Physostigmine ameliorates the delusions of Alzheimer's disease. Biol Psychi- atry 33:536-541.
de la Garza R, McGuire T J, Freedman R, Hoffer BJ ( 1987): Selec- tive antagonism of nicotinic actions in the rat cerebellum with o~-bungarotoxin. Neuroscience 23:887-891.
Davies P, Feisullin S (1981): Postmortem stability of o~-bungaro- toxin binding sites in mouse and human brain. Brain Res 216:449-454.
Daviss SR, Lewis DA ( 1993): Calbindin- and calretinin-immuno- reactive local circuit neurons are increased in density in the prefrontal cortex of schizophrenic subjects. NeurosciAbstracts 19:201.
Denny-Brown D, Fisher EG (1976): Physiological aspects of visual perception. II. The subcortical visual direction of behav- ior. Arch Neuro133:228-242.
EI-Mallakh RS, Kirch DG, Shelton R, et al (1991): The nucleus basalis of Meynert, senile plaques, and intellectual impairment in schizophrenia. J Neurop.2wqtiatrv 3:383-386.
Flores CM, Rogers SW, Pabreza LA, Wolfe BB, Kellar KJ ( 1992): A subtype of nicotinic cholinergic receptor in rat brain is com- posed of o~4 and [32 subunits and is upregulated by chronic nicotine treatment. Mol Pharmacol 41:31-37.
Freedman R, Wetmore C, Stromberg I, Leonard S, Olson L ( 1993): Alpha-bungarotoxin binding to hippocampal intemeurons: im- munocytochemical characterization and effects on growth fac- tor expression. J Neurosci 13:1965-1975.
Fuchs JL ( 1989): ['~l]c~-bungarotoxin binding marks primary sen- sory area of developing rat neocortex. Brain Res 501:223-234.
Gerzanich V, Anand R, Lindstrom J (1994): Homomers of o~8 and e~7 subunits of nicotinic receptors exhibit similar channel but contrasting binding site properties. Mol Pharmacol 45:212-220.
Glassman AH ( 1993): Cigarette smoking: implications for psychi- atric illness. Ant J Psychiatry 150:546--553.
Goff DC, Henderson DC, Amico E ( 1992): Cigarette smoking in schizophrenia: Relationship to psychopathology and medica- tion side effects. Am J Psychiatry 149:1189-1194.
Goff WR, Williamson PD, VanGilder JC, Allison T, Fisher TC (1980): Neural origins of long latency evoked potentials recorded from the depth and from the cortical surface of the brain in man. Prog Clin Neurophysiol 7:126-145.

Nicotinic Receptors in Schizophrenia BIOL PSYCHIATRY 33 1995:38:22-33
Griffith JM, Waldo M, Adler LE, Freedman R ( 1993): Normaliza- tion of auditory sensory gating in schizophrenics after a brief period for sleep. Psychiatr 3, Res 49:29-39.
Hall M, Zerbe L, Leonard S, Freedman R ( 1993): Characterization of [3H]cytisine binding to human brain membrane prepara- tions. Brain Res 600:127-133.
Harfstrand A, Adem A, Fuxe K, Agnati L, Andersson K, Nordberg A ( 1988): Distribution of nicotinic cholinergic receptors in the rat tel- and diencephalon: a quantitative receptor autoradiogra- phic study using [3H]acetylcholine, [alpha '25I] bungarotoxin and [3H] nicotine. Acta Physiol Scand 132:1-14.
Janowsky DS, el-Yousef K, David JM, Sekerke HJ (1973): Para- sympathetic suppression of manic symptoms by physostig- mine. Arch Gen Psychiatry 28:542-547.
Jeste D, Lohr JB (1989): Hippocampal pathologic findings in schizophrenia. Arch Gen Psychiatry 46: I 019-1024.
Judd LJ, McAdams L, Budnick B, Braff DL (1992): Sensory gating deficits in Schizophrenia: new results. Am J Psychiatry 149:488-493.
Karson CN, Casanova MF, Kleinman JE, Griffin WS (1993): Choline acetyltransferase in schizophrenia. Am J P:~3'chiatr3' 150:454-459.
Klein C, Andersen B (1991): On the influence of smoking upon smooth pursuit eye movements of schizophrenics and normal controls. J P©'chophysiology 5:361-369.
Lang W, Henke H ( 1983): Cholinergic receptor binding and auto- radiography in brains of non-neurological and senile dementia of Alzheimer-type patients. Brain Res 267:271-280.
Luetje CW, Patrick J, Seguela P ( 1990): Nicotinic receptors in the mammalian brain. FASEB J 4:2753-2760.
Luntz-Leybman V, Bickford P, Freedman R (1992): Cholinergic gating of response to auditory stimuli in rat hippocampus. Brain Res 587:130-136.
Masterson E, O'Shea B (1984): Smoking and malignancy in schizophrenia. Br J Ps3'chiatr3, 145:429--432.
Morley BJ (1989): a-bungarotoxin receptors in the CNS. Prog Brain Res 79:10 I- 108.
Perry EK, Court JA, Johnson M, Piggott MA, Perry RH (1992): Autoradiographic distribution of ['H]nicotine binding in human cortex: relative abundance in subicular complex. J Chem Neuroanatom v 5:399-405.
Revah F, Bertrand D, Galzi JL, Changeux JP ( 1991 ): Mutations in the channel domain alter desensitization of a neuronal nicotinic receptor. Nature 353:846-849.
Reynolds P, Czudek C, Andrews HB 1990): Deficit and hemi- spheric asymmetry of GABA uptake sites in the hippocampus of schizophrenics. Biol Psychiatry" 27:1038-1044.
Schoepfer R, Conroy WG, Whiting P, Gore M, Lindstrom J (1990): Brain ot bungarotoxin binding protein cDNAs and mAbs reveal subtypes of this branch of the ligand gated ion channel gene superfamily. Neuron 5:35-48.
Seguela P, Wadiche I, Dineley-Miller K, Dani J, and Patrick JW ( 1993): Molecular cloning, functional properties, and distribu- tion of rat brain c~-7: A nicotinic channel highly permeable to calcium. J Neurosci 13:596~04.
Sugaya K, Giacobini E, Chiappinelli VA ( 1990): Nicotinic acetyl- choline receptor subtypes in human frontal cortex: changes in Alzheimer's disease. J Neurosci Res 27:349-359.
Utkin Y, Lazakovich EM, Kasheverov IE, Tsetlen VI (1989): c~-bungarotoxin interacts with the rat brain tachkinin receptors. FEBS Lett 255: I 11-115.
Vattschanoff JG, Weinberg R, Kharazia VN, Nakane M, Schmidt HHHW (1993): Neurons in rat hippocampus that synthesize nitric oxide. J Comp Neuro1331 : 111-121.
Venables P ( 1964): Input dysfunction in schizophrenia. In Maher BA (ed), Progress in Experimental Personalit3, Research. New York: Academic Press, pp 1-47.
Vijayaraghavan S, Pugh PC, Zhang Z, Rathouz MM, Berg D ( 1992): Nicotinic receptors that bind a-bungarotoxin on neur- ons raise intracellular free Ca-". Neuron 8:353-362.
Vinogradova OS (1975): Functional organization of the limbic system in the process of registration of information: facts and hypothesis. In lsaacson RL, Pribram KH (eds), The Hippocam- pus, 2: Neurophysiology and Behavior. New York: Plenum, pp 3-70.
Watanabe S, Nishikawa T, Takashima M, Toru M (1983): In- creased muscarinic cholinergic receptors in prefrontal cortices of mediated schizophrenics. Life Sci 33:2187-2196.
Wonnacott S, Drasdo A, Sanderson E, Rowell P ( 1990): Presynap- tic nicotinic receptors and the modulation of transmitter re- lease. Ciba Found Syrup 152:87-101.
Copyright © 2022 FDOKUMEN




















