Event-related potentials reflect heterogeneity of developmental prosopagnosia
14
Event-related potentials reflect heterogeneity of developmental prosopagnosia Denise A. Minnebusch, Boris Suchan, Meike Ramon and Irene Daum Institute of Cognitive Neuroscience, Department. of Neuropsychology, Ruhr-University of Bochum, Universita ¨ tsstraße 150, D-44780 Bochum, Germany Keywords: ERP, face processing, N170, prosopagnosia Abstract Event-related potential (ERP) studies of developmental prosopagnosia (DP) are rare. Previous ERP investigations have reported smaller N170 amplitude differences between faces and objects in at least three prosopagnosics. The present study is based on a combination of behavioural and electrophysiological assessment of face processing. The aim was to investigate the face-specifity of the N170 in both healthy subjects and a group of DP individuals (N ¼ 4), using famous and nonfamous faces, caricatures and houses as stimuli. All prosopagnosic subjects showed impairments in recognition of famous faces, memory for faces and learning new faces in behavioural assessment. In healthy subjects the largest effects were found at parieto-occipital electrode positions (PO7 and PO8), along with a familiarity effect at these electrode positions. Thus, parieto-occipital areas appear to play an important role in face processing. Three prosopagnosics showed reliable N170 amplitude differences between faces and nonface stimuli, whereas one DP individual showed significantly reduced amplitude differences between faces and nonface objects. The behavioural and electrophysiological data support the idea that DP reflects a heterogeneous impairment and that face processing deficits are not necessarily correlated with a lack of face-specific N170. Introduction The term prosopagnosia refers to the inability to recognize a familiar individual on the basis of his or her face alone (Bodamer, 1947). Some individuals have had face recognition impairments all their life, without known brain injury at any stage of development (Behrmann et al., 2005). This problem is referred to as ‘congenital prosopagnosia’ (CP). The more general term ‘developmental prosopagnosia’ (DP) includes CP cases as well as individuals who suffered brain damage before birth or in early childhood (Farah et al., 2000; Barton et al., 2003; Behrmann et al., 2005). A recent study of face processing patterns in a larger group of eight DP cases (Le Grand et al., 2006) concluded that DP reflects a heterogeneous impairment which cannot be linked to a deficit on one individual aspect of face processing alone. The underlying mechanisms of face recognition problems in DP have been addressed by functional neuroimaging and electrophysio- logical studies. The fusiform face area (FFA) in humans is known to be more strongly activated during passive viewing of faces than of objects (Kanwisher et al., 1997; McCarty et al., 1997). CP individuals showed normal activation patterns in the fusiform gyrus in response to faces and buildings (Hasson et al., 2003; Avidan et al., 2005). The dissociation between intact FFA activation and impaired face processing in CP suggests that FFA activation is not sufficient for normal face processing. In event-related potential (ERP) studies, the N170 is generally found to be larger for faces than for nonface stimuli and has therefore been considered a face-specific component (Bentin et al., 1996). As the N170 is unaffected by the familiarity of faces, it is thought to reflect processing stages prior to face identification (Bentin & Deouell, 2000; Eimer, 2000a,b). According to Eimer (2000c), the N170 is generated by brain processes involved in late stages of structural encoding, where representations of global face configurations are generated for face recognition. Single-case analyses have reported a smaller N170 amplitude difference between faces and objects in prosopagnosia (Bentin et al., 1999; Kress & Daum, 2003). Prosopagnosics thus appear to process faces and houses in a similar manner, possibly linked to nonspecific structural encoding. However, the use of magnetoencephalography yielded a more complex pattern. Three prosopagnosics showed no amplitude differences between faces and houses whereas the other two subjects showed higher amplitudes to faces than to nonface stimuli, revealed by both magnetoencephalography (M170) and ERP (N170; Harris et al., 2005). These results suggest a neurophysiological heterogeneity of DP. The aim of the present study was to investigate the face-specificity of the N170 in a group of four DP individuals using a broader range of face stimuli to further elucidate the possible functional heterogeneity of the condition. The combination of behavioural and electrophysi- ological assessments should permit further insights into the brain mechanisms underlying face processing and their dysfunction in prosopagnosia. Materials and methods Subjects Four individuals (two female; age 37.3 ± 2.5 years; mean values are quoted ± SD throughout) who subjectively experienced severe face recognition deficits, and nine healthy control subjects (four female; Correspondence: Denise A. Minnebusch, as above. E-mail: [email protected] Received 19 October 2006, revised 17 January 2007, accepted 31 January 2007 European Journal of Neuroscience, Vol. 25, pp. 2234–2247, 2007 doi:10.1111/j.1460-9568.2007.05451.x ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd
Transcript of Event-related potentials reflect heterogeneity of developmental prosopagnosia

Event-related potentials reflect heterogeneity ofdevelopmental prosopagnosia
Denise A. Minnebusch, Boris Suchan, Meike Ramon and Irene DaumInstitute of Cognitive Neuroscience, Department. of Neuropsychology, Ruhr-University of Bochum, Universitatsstraße 150, D-44780Bochum, Germany
Keywords: ERP, face processing, N170, prosopagnosia
Abstract
Event-related potential (ERP) studies of developmental prosopagnosia (DP) are rare. Previous ERP investigations have reportedsmaller N170 amplitude differences between faces and objects in at least three prosopagnosics. The present study is based on acombination of behavioural and electrophysiological assessment of face processing. The aim was to investigate the face-specifity ofthe N170 in both healthy subjects and a group of DP individuals (N ¼ 4), using famous and nonfamous faces, caricatures and housesas stimuli. All prosopagnosic subjects showed impairments in recognition of famous faces, memory for faces and learning new facesin behavioural assessment. In healthy subjects the largest effects were found at parieto-occipital electrode positions (PO7 and PO8),along with a familiarity effect at these electrode positions. Thus, parieto-occipital areas appear to play an important role in faceprocessing. Three prosopagnosics showed reliable N170 amplitude differences between faces and nonface stimuli, whereas one DPindividual showed significantly reduced amplitude differences between faces and nonface objects. The behavioural andelectrophysiological data support the idea that DP reflects a heterogeneous impairment and that face processing deficits are notnecessarily correlated with a lack of face-specific N170.
Introduction
The term prosopagnosia refers to the inability to recognize a familiarindividual on the basis of his or her face alone (Bodamer, 1947). Someindividuals have had face recognition impairments all their life,without known brain injury at any stage of development (Behrmannet al., 2005). This problem is referred to as ‘congenital prosopagnosia’(CP). The more general term ‘developmental prosopagnosia’ (DP)includes CP cases as well as individuals who suffered brain damagebefore birth or in early childhood (Farah et al., 2000; Barton et al.,2003; Behrmann et al., 2005). A recent study of face processingpatterns in a larger group of eight DP cases (Le Grand et al., 2006)concluded that DP reflects a heterogeneous impairment which cannotbe linked to a deficit on one individual aspect of face processing alone.The underlying mechanisms of face recognition problems in DP
have been addressed by functional neuroimaging and electrophysio-logical studies. The fusiform face area (FFA) in humans is known tobe more strongly activated during passive viewing of faces than ofobjects (Kanwisher et al., 1997; McCarty et al., 1997). CP individualsshowed normal activation patterns in the fusiform gyrus in response tofaces and buildings (Hasson et al., 2003; Avidan et al., 2005). Thedissociation between intact FFA activation and impaired faceprocessing in CP suggests that FFA activation is not sufficient fornormal face processing. In event-related potential (ERP) studies, theN170 is generally found to be larger for faces than for nonface stimuliand has therefore been considered a face-specific component (Bentinet al., 1996). As the N170 is unaffected by the familiarity of faces, it isthought to reflect processing stages prior to face identification
(Bentin & Deouell, 2000; Eimer, 2000a,b). According to Eimer(2000c), the N170 is generated by brain processes involved in latestages of structural encoding, where representations of global faceconfigurations are generated for face recognition.Single-case analyses have reported a smaller N170 amplitude
difference between faces and objects in prosopagnosia (Bentin et al.,1999; Kress & Daum, 2003). Prosopagnosics thus appear to processfaces and houses in a similar manner, possibly linked to nonspecificstructural encoding.However, the use of magnetoencephalography yielded a more
complex pattern. Three prosopagnosics showed no amplitudedifferences between faces and houses whereas the other two subjectsshowed higher amplitudes to faces than to nonface stimuli, revealed byboth magnetoencephalography (M170) and ERP (N170; Harris et al.,2005). These results suggest a neurophysiological heterogeneity ofDP.The aim of the present study was to investigate the face-specificity
of the N170 in a group of four DP individuals using a broader range offace stimuli to further elucidate the possible functional heterogeneityof the condition. The combination of behavioural and electrophysi-ological assessments should permit further insights into the brainmechanisms underlying face processing and their dysfunction inprosopagnosia.
Materials and methods
Subjects
Four individuals (two female; age 37.3 ± 2.5 years; mean values arequoted ± SD throughout) who subjectively experienced severe facerecognition deficits, and nine healthy control subjects (four female;
Correspondence: Denise A. Minnebusch, as above.E-mail: [email protected]
Received 19 October 2006, revised 17 January 2007, accepted 31 January 2007
European Journal of Neuroscience, Vol. 25, pp. 2234–2247, 2007 doi:10.1111/j.1460-9568.2007.05451.x
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing Ltd

39.5 ± 12 years) participated in this study; details are given in Table 1.All subjects were right-handed (Oldfield, 1971) and receivedreimbursement for participation. All participants had normal orcorrected-to-normal vision. Magnetic resonance imaging did not yieldany overt brain abnormalities in the DP group, who showed facerecognition deficits in the presence of intact basic visual processingand intact object recognition abilities (see below). The study wasapproved by the Ethics Committee of the Medical Faculty of the Ruhr-University, Bochum, Germany and written informed consent wasobtained from all subjects. The study was performed in accordancewith the guidelines of the Declaration of Helsinki (Varga, 1975).
Basic demographic and clinical variables relating to the DPindividuals are presented in Table 1. In three cases (ET, LT and
NN), the medical history was unremarkable. NN and TP shared theirprosopagnosia with at least two family members, suggesting apotential genetic involvement. Premature delivery and birth compli-cations were reported by TP. All subjects may have some early braindamage or insult to the brain and so they are classifiable potentially asDP cases rather than CP cases (according to Behrmann et al., 2005).
Basic visual processing
Basic level vision was assessed using six subtests of the BirminghamObject Recognition Battery (BORB; Riddoch & Humphreys, 1993;Copying test, Length match, Size match, Orientation match, Gapmatch and Overlapping figure test) and three subtests of the BentonBattery (Benton et al., 1983; Right–left orientation, Judgement of lineorientation and Visual form discrimination). The control data are basedon the BORB and Benton manual. These two test batteries have beenused in several other studies in order to examine visual objectprocessing abilities (e.g. Bentin et al., 1999; DeGelder & Rouw, 2000;Duchaine, 2000; Nunn, Postma & Pearson, 2001; Duchaine et al.,2003a,b; 2004; Humphreys et al., 2006), allowing comparabilityacross studies.Basic-level object recognition was assessed by digitized versions of
100 line drawings from the pictures of Snodgrass & Vanderwart(1980; see Farah & McClelland, 1991; Duchaine et al., 2003a,b;2004). This test version was standardized in our laboratory and a
Table 1. Characterization of the prosopagnosic subjects
Patient Sex
Age attesting(years)
Familialinvolvement
Neurological orpsychiatric disease
ET Male 41 – –LT Female 36 – –NN Male 36 Father, mother
(after meningitis),brother
–
TP Female 36 Parents and uncle Birth at 34th weekof gestation
Table 2. Performance of the prosopagnosic subjects and controls in behavioural assessment
Tests and Subtests
Patients
ControlsET LT NN TP
BentonRight–left orientation 18 20 20 20 NR 17-20Judgement ofline orientation 29 30 29 29 NR 26-24Visual formdiscrimination 32 32 30 30 NR 24-32
BORBLength matching 25 30 28 28 26.9 ± 1.6Size matching 28 26 26 28 27.3 ± 2.4Orientation matching 28 25 27 28 24.8 ± 2.6Gap matching 35 36 38 37 35.1 ± 4
Snodgrass, percentage correct 96 99 99 96 96 ± 2.5Rey figure
Copy 34 36 29.5 36 31.3 ± 4.5Recall 17.5 ⁄ 36 13 ⁄ 36 5 ⁄ 36� 14.5 ⁄ 36 16.4 ± 7.1
TABIdentity discrim. 93.33� 91.6� 91.6� 100 98.8 ± 2.7Affect discrim. 91.6 78.57� 85.71� 92.86 91.6 ± 5.6Affect naming 94.81 86.67� 66.7� 93.33 94.8 ± 5.8Affect ident. 86.6� 86.67� 93.3 93.33 96.5 ± 3.7Affect matching 94.3 86.67� 46.7� 93.33 94.3 ± 5.3
FF Test 1, percentage correct 35.71� 10.5� 13.63� 10� 76 ± 14FF Test 2, percentage correct 44.8� 23.7� 28.95� 13.9� 84 ± 12Famous caricatures, percentage correct 24.1� 28.9� 34.21� 25� 63 ± 15BFRT 45 44 40� 42 NR 43–46RMT-F 38� 35� 33� 34� 43.9 ± 3.7a
44.8 ± 3.3b
Bochum test of face processingd¢ 0.6� 1.9 0.09� 1.3 2.1 ± 0.7RT for hits (ms) 117 436� 380� 430� 187 ± 99
FF, famous face; NR, normal range; d¢, measure of discrimination; RT, reaction time. Mean values are ± SD. �Performance below average (z < 1.65 relative tocontrol group).a,bAge-related test scores: a35–39 years, b40–44 years.
Developmental prosopagnosia 2235
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

group of 100 healthy volunteers (64 female, 36 male; mean age29.51 ± 10.2 years) were able to identify 96 ± 2.5% of the pictures.None of the prosopagnosics differed from the normal range on any ofthese tasks (see Table 2).Visuoconstructive and visual memory abilities were examined by
copy and free recall (after 30 min) of the Rey Complex Figure(Osterrieth, 1944). All prosopagnosics copied this figure without anydifficulty (Mitrushina et al., 1999; Table 2). For delayed recall NNperformed poorly (score of 5).
Face processing
Face processing and recognition abilities were assessed using twonewly developed face recognition tests and two widely used tests, theRecognition Memory Test for faces (RMT-F; Warrington, 1984) andthe Benton Facial Recognition Test (BFRT; Benton et al., 1983)making the data from this study comparable to that from otherinvestigations (Bentin et al., 1999; DeGelder & Rouw, 2000;Duchaine, 2000; Nunn et al., 2001; Kress & Daum, 2003;Humphreys et al., 2006). The control data are based on the RMT-F and BFRT manual. In line with previous studies of DP (Behrmannet al., 2005; Duchaine et al., 2003a,b, 2004; Le Grand et al., 2006),newly developed tests were added to the battery as individuals withsevere prosopagnosia have been reported to score normally on theRMT-F and the BFRT (Duchaine & Weidenfeld, 2003). NN wasimpaired on the BFRT (score 40); all other DP subjects performedwithin the normal range (Table 2). All subjects were impaired on theRMT-F (scores 33–38), indicating a problem with memory for faces(Table 2).The ability to discriminate face identity and to recognize emotions
in facial expressions was assessed using five subtests of the TubingerAffect Battery (TAB, Breitenstein et al., 1996, 1998), the Germanversion of the Florida Affect Battery. In Subtest 1 (Identity discrim-ination), pairs of unfamiliar faces are shown and subjects have todetermine whether the faces belong to the same or to two differentpeople. In Subtest 2 (Affect discrimination), subjects have to deter-
mine whether two faces depict the same or different emotionalexpression and Subtest 3 (Affect naming) requires naming of facialexpressions. Subtest 4 (Affect identification) assessed the ability toselect facial expressions named by the examiner. In Subtest 5 (Affectmatching), subjects are asked to match the picture of an emotional faceto another face with the same emotional expression. The control dataare based on the TAB manual (see Breitenstein et al., 1998).The TAB was also used by Kress & Daum (2003). Although
emotional processing is not a diagnostic criterion in DP, it hasfrequently been pointed out that some prosopagnosics have difficultiesin recognizing facial expressions of emotions (Kress & Daum, 2003;Duchaine et al., 2004) whereas others did not (Nunn et al., 2001;Duchaine et al., 2003b). Humphreys et al. (2006) reported normalfacial expression processing in three CP subjects while two individualswith acquired prosopagnosia performed below average. Our DP groupshowed a heterogeneous pattern: TP performed largely in the normalrange, ET showed evidence of mild impairments and LT and NN wereimpaired on at least four of the five tests of the TAB (Table 2).
Bochum test of face processing skills
This taskwas developed in analogy to the ‘One-in-ten Test’ described byDuchaine et al. (2003a,b) to assess the ability to learn new faces.Subjects are asked to recognize 15 photos of a target individual differingin illumination in a series of 150 photos presented one at a time. Thephotos are black-and-white pictures of women aged between 20 and30 years with only internal face features visible. The stimuli arepresented for 500 ms with an interstimulus interval (ISI) of 1000 ms.Signal detection analysis was used to determine the ability to
discriminate between target and distractor individuals (Snodgrass &Corwin, 1988). The mean measure of discrimination (d¢) was2.10 ± 0.74 for 64 control subjects (35 female; mean age 31.3 ±12.3 years).The data from the prosopagnosics were converted into z-scores
(cutoff )1.65). ET and NN showed impairments in learning new faces(d¢ ¼ 0.60 and 0.09, respectively), whereas d¢ was within the normal
0
10
20
30
40
50
60
70
80
90
100
Per
form
ance
(%
)
Famous faces I Famous faces II Caricatures
ET LT NN TP Controls
Fig. 1. Performance of the prosopagnosic subjects and control group in two famous face recognition tasks and in recognition of caricatures from famous people.
2236 D. A. Minnebusch et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

Fig.2.
Experim
entaldesign.
Developmental prosopagnosia 2237
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247
range for LT and TP (Table 2). In addition, the reaction times (RTs) forhits were significantly slower than control RTs in LT, NN and TP (436,380 and 430 ms, respectively; z ¼ 1.9–2.5).
Recognition of famous faces
The Famous Face Recognition Test was developed to test theprosopagnosics’ ability to recognize famous faces. Grey-scaledphotographs of faces of well-known individuals were shown for10 s each, with little hair or other external features being visible. Thefamous faces were taken from four categories (politicians, comedians,athletes, musicians), with six items in each category. Subjects wereasked to name the individual or provide any other information whichuniquely identifies the person. After test completion, the names of thepersons who served as stimuli were presented and subjects had toassign them into five possible professional categories (politicians,comedians, athletes, musicians and others) to assess the level offamiliarity with the specific stimuli of the test. A sample of 100 controlsubjects (64 female, mean age 30.2 ± 10.2 years) correctly named76 ± 14% of the 24 faces. Age and sex were unrelated to performance.Individual scores were expressed as the proportion of identified
faces relative to the number of faces which were familiar to anindividual subject. Compared to the control group, all DP subjectswere impaired in recognizing famous faces (scores 35.7–10%, seeFig. 1 and Table 2).In further famous face recognition tasks administered in the same
way as the ‘Famous Faces Test’ described above, subjects were shown40 photos of famous people and 40 caricatures of faces of the samepeople used in the subsequent EEG study. One hundred healthy controlsubjects (65 female, mean age 27.9 ± 8.2 years) were able to identify83.7 ± 11.8% of the photos of the famous people used in this study. Allprosopagnosics scored at least 3 SDs below the control mean (scores
13.9–44.8%; see Fig. 1 and Table 2). Fifty-five control subjects (29female; mean age 32 ± 13.2 years) correctly named 63 ± 15% of thepeople presented as caricatures. All prosopagnosic subjects wereimpaired in recognizing celebrities on the basis of caricatures (scores24.1–34.2%). All control subjects performed better with photos thanwith caricatures, whereas three prosopagnosics (LT, NN and TP)showed a trend towards better recognition of caricatures of famousfaces than photographs of famous faces (t3,2 ¼ )3.67, P ¼ 0.067). ETshowed the same pattern as the control group (see Fig. 1 and Table 2).
Stimuli for EEG study
Grey-scaled pictures (140 · 200 pixels) from 16 categories (famousfaces, nonfamous faces, caricatures, houses, faces of horses, horses inmovement, horses-stationary, fruits and vegetables, butterflies, cars-lateral views, cars-frontal views, bicycles, sunglasses, guns, tools,musical instruments) were presented on a white background. Eachcategory involved 40 stimuli, with the exception of ‘houses’ (32items). Photographs of famous and nonfamous faces were shown atfull frontal view, with only internal facial features being visible. Thenonfamous faces comprised 20 females and 20 males, and facialexpressions were neutral. The caricatures consisted of illustrations ofthe 40 celebrities used as famous faces. A further stimulus categorywas 190 pictures of brain slices from high-resolution T1-weightedscans which served as a control task to focus attention. Each stimuluswas presented for 500 ms with an ISI of 1000 ms. During the ISIs, ablack cross appeared in the centre of the monitor (see Fig. 2).
Procedure
Subjects were seated in a dimly lit sound-attenuated and electricallyshielded room. Pictures of the 17 stimulus classes were presented in
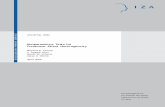
Fig. 3. Grand mean maps for controls for nonfamous faces, famous faces and caricatures.
2238 D. A. Minnebusch et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

random order on a monitor at a distance of 80 cm from the subject’seyes. Participants were instructed that they would see pictures ofdifferent categories and occasionally brain slices would appear. Theywere asked to press one key if a grey dot appeared on the right of thebrain picture and another key if it was on the left (Fig. 2). Subjectswere generally unfamiliar with such pictures, which were thusmeaningless to them. The investigation lasted � 25 min.
EEG recordings and ERP analyses
EEG was recorded from 30 electrodes, mounted in an elastic electrodecap adapted from the 10–20 standard setup, referenced to an electrodeon the tip of the nose. Vertical and horizontal eye movements wererecorded using four additional electrodes placed above and below theleft eye and on the outer canthus of each eye. Electrode impedancewas kept < 5 kW. Data were digitized at a sampling rate of 250 Hz.EEG was epoched off-line in periods of 1100 ms, starting 100 msprior to the stimulus onset. The 100-ms interval before stimulus onsetserved as baseline. Trials with EOG artefacts and amplitudes > 50 lVwere rejected from further analyses. The present analysis focused onface stimuli (famous faces, nonfamous faces and caricatures) andhouses which have typically been used as comparison stimuli inprevious studies (Eimer, 2000a,2b,c). The results for the remainingcategories will be reported elsewhere. ERPs were averaged separatelyfor each category (famous faces, nonfamous faces, caricatures and
houses). The N170 was determined by the mean amplitude within the140–190 ms poststimulus latency window and was analysed forelectrode positions TP7 and TP8, and for PO7 and PO8.
Results
Individual maps for the group of controls and each prosopagnosicsubject (see Figs 3 and 4) indicated that the highest amplitudesemerged on electrode positions PO7 and PO8.Therefore, the analyses focused on these positions, together with
positions TP7 and TP8 for comparison with previous findings (Eimer,2000a,b,c; Kress & Daum, 2003). Because of the very large variabilityand the small sample size, the data of the prosopagnosics are presentedin terms of z-scores relative to the data of the controls.
Electrode positions TP7 and TP8
Control subjects
Grand averages for ERPs elicited by faces, caricatures and houses atleft and right lateral temporal electrodes (TP7 and TP8) are presentedin Fig. 5. N170 amplitudes were analysed with factors StimulusCategory (famous faces, nonfamous faces, caricatures and houses) andHemisphere (left vs. right). There were no significant differences inN170 amplitude between famous faces and nonfamous faces
Fig. 4. Individual maps for each prosopagnosic subject for nonfamous faces, famous faces and caricatures.
Developmental prosopagnosia 2239
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

(P ¼ 0.2); the data were therefore pooled into a single face category.In addition there were no significant amplitude differences betweenfamous faces and caricatures (P ¼ 1.0). Photographs of famous andnonfamous faces are realistic representations of faces whereascaricatures represent unrealistic illustrations of faces, and the categ-ories of face photos and caricatures were therefore analysed separately.The analysis yielded a main effect of Stimulus Category
(F2,9 ¼ 10.2, P ¼ 0.002) with larger N170 amplitudes for facesrelative to houses (P ¼ 0.004), but not for caricatures relative tohouses (P ¼ 0.147). None of the other effects reached significance.
Prosopagnosics
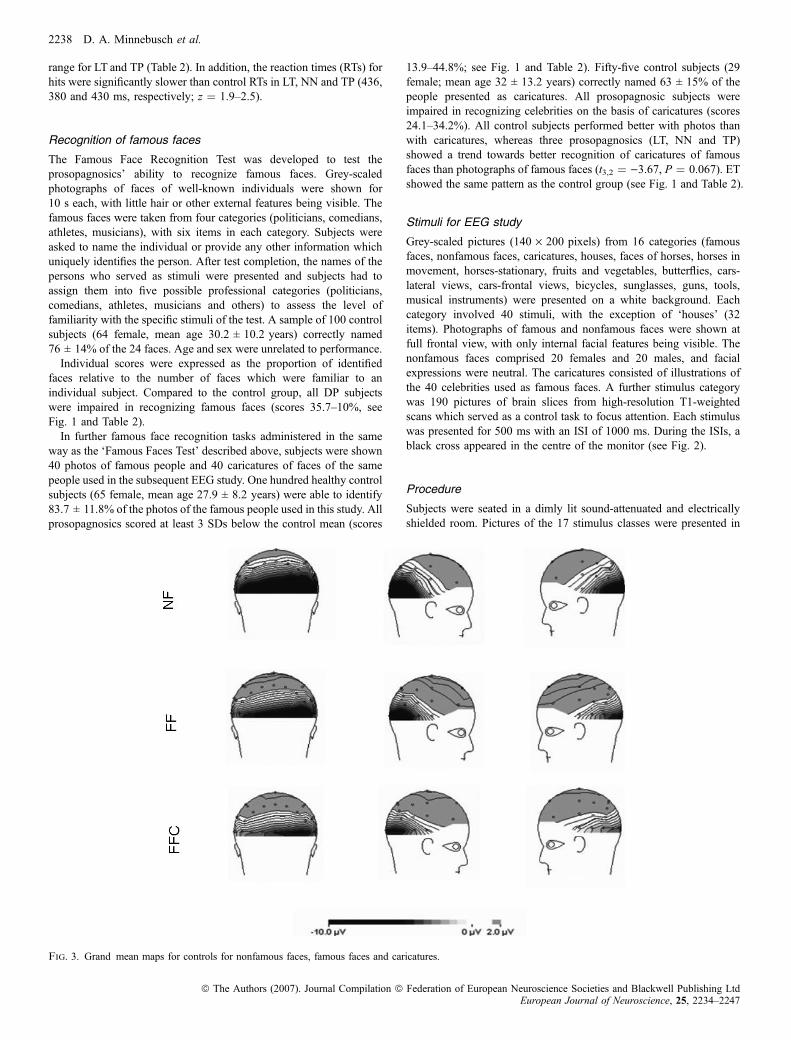
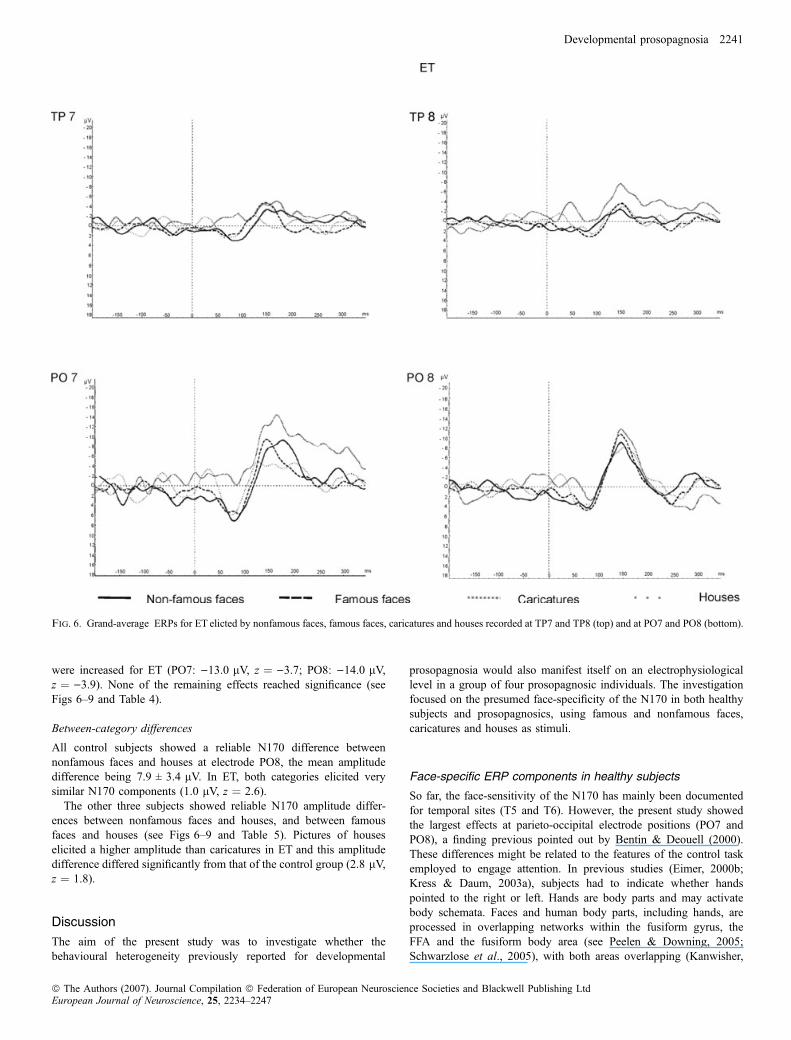
Grand average ERPs for each prosopagnosic subject elicited bynonfamous faces, famous faces, caricatures and houses at TP7 andTP8, and for PO7 and PO8 are presented in Figs 6–9.Compared to the control group, pictures of faces elicited a larger
N170 in NN at TP8 ()9.2 lV, z ¼ )2.0). ET and NN also showed alarger N170 to caricatures than controls in the right hemisphere (ET:)7.4 lV, z ¼ )1.7; NN: )8.3 lV, z ¼ )2.0). At electrode positionTP8 pictures of houses elicited a significantly larger N170 in ET, NNand TP (ET: )6.7 lV, z ¼ )2.9; NN: )5.8 lV, z ¼ )2.4; TP:)5.9 lV, z ¼ )2.5) compared to the control group (see Table 3).
Between-category differences
Amplitude differences between faces and houses are summerized inTable 5. ET showed a larger N170 to houses than to faces (amplitudedifferences 1.3 lV, z ¼ 1.9); this was in contrast to the control group.In addition, TP showed a greater amplitude difference betweencaricatures and houses at TP8 (2.1 lV, z ¼ 1.8). The other twoprosopagnosics did not differ significantly from the control group (seeTable 5).
Electrode positions PO7 and PO8
Control subjects
Grand averages for ERPs at right and left parieto-occipital electrodes(PO7 and PO8) are presented in Fig. 10. Analyses with factorsStimulus Category (famous faces, nonfamous faces, caricatures andhouses) and Hemisphere (left vs. right) yielded main effects ofStimulus Category (F3,9 ¼ 23.2, P < 0.001) and Hemisphere(F1,9 ¼ 45.4, P < 0.001), and a significant interaction (F3,9 ¼ 4.7,P ¼ 0.017). Subsequent paired comparisons revealed higher N170amplitudes for nonfamous faces than for famous faces (P ¼ 0.005),caricatures (P < 0.005) and houses (P ¼ 0.01). In addition, N170amplitudes were higher for famous faces than for houses (P ¼ 0.008),whereas the N170 amplitudes for caricatures did not differ from thosefor houses (P > 0.05). Famous faces (F1,9 ¼ 22.3, P < 0.005),nonfamous faces (F1,9 ¼ 39.1, P < 0.001) and caricatures(F1,9 ¼ 11.2, P ¼ 0.01) elicited a larger N170 over the right thanthe left hemisphere; houses did not yield a significant hemisphereeffect.
Prosopagnosics
Famous faces elicited increased N170 components in ET (PO7:)17.3 lV, z ¼ )3.5; PO8: )17.5 lV, z ¼ )2.3). In ET, N170amplitudes for nonfamous faces were significantly larger over theleft hemisphere (PO7: 19.1 lV, z ¼ )2.8). In LT, the N170 fornonfamous faces was reduced over the right hemisphere ()1.6 lV,z ¼ 2.1, respectively).Caricatures elicited a significantly larger N170 in ET over the left
hemisphere (PO7: )14.4 lV, z ¼ )3.1) and reduced amplitudes in LTat PO8 (1.3 lV, z ¼ 2.0). The N170 was also reduced for houses inLT at PO8 (2.5 lV, z ¼ 1.8). In contrast, N170 amplitudes for houses
Fig. 5. Controls grand-average ERPs elicted by nonfamous faces, famous faces, caricatures and houses recorded at TP7 (left side) and TP8 (right side).
2240 D. A. Minnebusch et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247
were increased for ET (PO7: )13.0 lV, z ¼ )3.7; PO8: )14.0 lV,z ¼ )3.9). None of the remaining effects reached significance (seeFigs 6–9 and Table 4).
Between-category differences
All control subjects showed a reliable N170 difference betweennonfamous faces and houses at electrode PO8, the mean amplitudedifference being 7.9 ± 3.4 lV. In ET, both categories elicited verysimilar N170 components (1.0 lV, z ¼ 2.6).
The other three subjects showed reliable N170 amplitude differ-ences between nonfamous faces and houses, and between famousfaces and houses (see Figs 6–9 and Table 5). Pictures of houseselicited a higher amplitude than caricatures in ET and this amplitudedifference differed significantly from that of the control group (2.8 lV,z ¼ 1.8).
Discussion
The aim of the present study was to investigate whether thebehavioural heterogeneity previously reported for developmental
prosopagnosia would also manifest itself on an electrophysiologicallevel in a group of four prosopagnosic individuals. The investigationfocused on the presumed face-specificity of the N170 in both healthysubjects and prosopagnosics, using famous and nonfamous faces,caricatures and houses as stimuli.
Face-specific ERP components in healthy subjects
So far, the face-sensitivity of the N170 has mainly been documentedfor temporal sites (T5 and T6). However, the present study showedthe largest effects at parieto-occipital electrode positions (PO7 andPO8), a finding previous pointed out by Bentin & Deouell (2000).These differences might be related to the features of the control taskemployed to engage attention. In previous studies (Eimer, 2000b;Kress & Daum, 2003a), subjects had to indicate whether handspointed to the right or left. Hands are body parts and may activatebody schemata. Faces and human body parts, including hands, areprocessed in overlapping networks within the fusiform gyrus, theFFA and the fusiform body area (see Peelen & Downing, 2005;Schwarzlose et al., 2005), with both areas overlapping (Kanwisher,
Fig. 6. Grand-average ERPs for ET elicted by nonfamous faces, famous faces, caricatures and houses recorded at TP7 and TP8 (top) and at PO7 and PO8 (bottom).
Developmental prosopagnosia 2241
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

2000; Downing et al., 2001; Yovel & Kanwisher, 2004). It ispossible that the control task of previous studies activated fusiformgyrus networks which may have affected the ERP correlates of faceprocessing. This potential problem was avoided in the present studywhere brain slices served as control stimuli. In addition, faces werepreviously shown together with external features (hair, etc: Bentinet al., 1999; Eimer, 2000a,b; Kress & Daum, 2003) which may haveinfluenced face recognition and the N170. The face stimuli of thepresent study were therefore presented with external featuresremoved. In previous studies electrode positions T5 and T6 werechosen a priori without visual inspection of the highest amplitude(Eimer & McCarthy, 1999; Kress & Daum, 2003; Harris et al.,2005). In contrast to the present findings, Bentin & Deouell (2000)did not find higher amplitudes at PO7 and PO8 than at TP7 and TP8for passive viewing of famous and nonfamous faces (subjects had tocount butterflies and ignore the faces). However, in a secondexperiment they explored active viewing of famous and nonfamouspeople (subjects had to identify and count famous people) and foundhigher amplitudes at PO7 and PO8 than at TP7 and TP8. This resultis in agreement with our findings. The present experiment thusextends the previous reported findings (Bentin & Deouell, 2000) of
higher amplitudes at parieto-occipital electrodes to passive viewingof famous and nonfamous faces.Consistent with previous findings, face familiarity had no effect on
the N170 component at TP7 and TP8 (Fig. 5), suggesting that theN170 reflects processes prior to face identification (Eimer, 2000a,b).In addition, caricatures of famous people elicited similar N170components as famous and nonfamous faces. At parieto-occipital sites,on the other hand, nonfamous faces evoked significantly larger N170amplitudes than famous faces and caricatures (Fig. 10), which is incontrast to Bentin & Deouell (2000), who showed similar N170amplitudes for famous and nonfamous faces. As the largest N170amplitudes were seen at PO7 and PO8 and as the amplitudedifferences between faces and houses were also most pronouncedover these regions, parieto-occipital areas appear to play a predomi-nant role in face processing. These results also suggest that famousand nonfamous faces are processed differently in these cortical areas.The N170 has been related to an early mechanism operating at earlystages of face processing, related to structural encoding (Bentin et al.,1999). Our results imply that forming an invariant sensory represen-tation of a new face requires a larger effort than the processing of well-known faces for which such representations already exist. It is possible
Fig. 7. Grand-average ERPs for LT elicted by nonfamous faces, famous faces, caricatures and houses recorded at TP7 and TP8 (top) and at PO7 and PO8 (bottom).
2242 D. A. Minnebusch et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247
that face representations are formed in parieto-occipital areas and thatfamous faces, nonfamous faces and caricatures are processed in asimilar way in temporal areas. PET studies (Rossion et al., 2001;Rossion et al., 2003) demonstrated that the right FFA and the rightoccipital face area (OFA) are involved in the discrimination betweenfamiliar and unfamiliar faces. Consistent with our electrophysiologicalfindings, both areas (FFA and OFA) showed higher activations fornonfamous than for famous faces. Previous event-related studiesindicated that the discrimination of faces and objects occurs 170 msafter stimulus onset (Bentin et al., 1996), whereas the discriminationof famous and nonfamous faces does not appear before 250 ms(Eimer, 2000b). Our data support the view that the discriminationbetween famous and nonfamous faces occurs 170 ms after stimulusonset. Non-famous faces are possibly more difficult to process thanfamous faces, which might be reflected in higher amplitudes fornonfamous than for famous faces.
N170 in prosopagnosia
The behavioural and electrophysiological data of the present studysupport the idea that DP reflects a heterogonous impairment and that
face processing deficits are not necessarily correlated with a lack of theface-selective N170. Consistent with previous findings, none of theprosopagnosics was impaired on tests of low level vision and basiclevel object recognition. A general low level object recognition deficitcan thus be excluded in all subjects. In NN, there was evidence ofvisuospatial memory deficits. While face matching assessed by theBFRT was intact in all subjects but NN, all individuals (ET, LT, NNand TP) were impaired on the RMT-F (Old–New face decision task),indicating memory impairments for faces. As all prosopagnosicsexcept NN scored normally on a visual memory task (Rey ComplexFigure), these results cannot be attributed to general visual memoryimpairment in ET, LT and TP. Normal performance on face memorytasks does not exclude an impairment in face processing. In addition,all prosopagnosic subjects had difficulties in recognizing famous facesand ET, LT and NN were impaired in face discrimination (TAB),indicating a broader perceptual basis for the deficits. All prosopag-nosics also performed poorly at learning new faces (Bochum Test ofFace Processing Skills). Two DP individuals (LT and NN) had severedifficulties in identifying emotions in facial expressions. In sum, allprosopagnosics were impaired at memory for faces, learning new facesand recognition of famous faces. These data support the assumptionthat all subjects have deficits in processing and recognizing faces.
Fig. 8. Grand-average ERPs for NN elicted by nonfamous faces, famous faces, caricatures and houses recorded at TP7 and TP8 (top) and at PO7 and PO8 (bottom).
Developmental prosopagnosia 2243
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

As described above, in healthy subjects faces evoked an enhancedN170 compared to nonface stimuli, particularly over the righthemisphere. At right temporal (TP8) and parieto-occipital (PO8) sites,only ET showed a significantly reduced amplitude difference between
faces and houses, which was, similar to previous reports (Bentin et al.,1999; Kress & Daum, 2003), related to a higher N170 amplitude fornonface stimuli compared to controls. These data support the idea thatET processed faces and nonface stimuli in a similar way. It should be
Fig. 9. Grand-average ERPs for TP elicted by nonfamous faces, famous faces, caricatures and houses recorded at TP7 and TP8 (top) and at PO7 and PO8 (bottom).
Table 3. N170 amplitudes for caricatures (FFC), famous faces (FF), nonfamous faces (NF) and houses, for each prosopagnosic subject and for controls, atelectrode positions TP7 and TP8
Patient
N170 amplitudes (lV)
FFC FF NF House
TP7 TP8 TP7 TP8 TP7 TP8 TP7 TP8
ET )5.1 )7.4� )10.4� )7.5 )2.0 )3.3 )2.9 )6.7LT 0.1 0.2 )3.1 )4.7 )1.5 )1.9 )0.1 )1.3NN )3.8 )8.3� )5.4 )6.9 )8.9� )11.4� )3.2 )5.8TP )2.3 )3.8 )5.9 )8.4� )6.4� )7.2� )3.8 )5.9Controls )2.2 ± 2.0 )3.2 ± 2.5 )2.6 ± 2.5 )3.8 ± 2.5 )3.8 ± 2.4 )4.9 ± 2.6 )1.6 ± 2.1 )0.9 ± 2.0
Mean values are ± SD; �z < )1.65 (i.e. �z is more negative than )1.65); �z > 1.65.
2244 D. A. Minnebusch et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

Table 5. Patients’ and control N170 amplitude differences between caricatures (FFC) and houses, faces and houses, famous faces (FF) and houses and non-famousfaces (NF) and houses at electrode positions TP7 and TP8 and ⁄ or PO7 and PO8
Patient
N170 amplitude differences (lV)
FFC ) Houses Faces ) Houses FF ) Houses NF ) Houses
TP7 TP8 PO7 PO8 TP7 TP8 PO7 PO8 PO7 PO8
ET )2.2 )0.6 )1.4 2.8� )3.3 1.3� )4.3 )3.5 )6.1 1.0�
LT 0.3 1.5 )2.7 )1.3 )2.2 )2.0 )5.6 )7.0 )3.9 )4.1NN )0.6 )2.4 )3.1 )2.2 )3.9 )3.3 )5.2 )4.7 )5.7 )7¢.3TP 1.6 2.1� 0.04 0.7 )2.3 )2.0 )1.5 )3.3 )4.7 )4.3Controls )0.65 ± 2.0 )2.2 ± 2.4 )1.8 ± 2.2 )2.9 ± 3.3 )1.6 ± 2.3 )3.4 ± 2.5 )3.2 ± 3.1 )5.5 ± 2.8 )5.84 ± 3.3 )7.8 ± 3.4
Mean values are ± SD;�
z > 1.65.
Fig. 10. Controls’ grand-average ERPs elicted by nonfamous faces, famous faces, caricatures and houses recorded at PO7 (left side) and PO8 (right side).
Table 4. N170 amplitudes for caricatures (FFC), famous faces (FF), nonfamous faces (NF) and houses, for each prosopagnosic subject and for controls, atelectrode positions PO7 and PO8
Patient
N170 amplitudes (lV)
FFC FF NF House
PO7 PO8 PO7 PO8 PO7 PO8 PO7 PO8
ET )14.4� )11.2 )17.3� )17.5� )19.1� )13.1 )13.0� )14.0�
LT )0.7 1.3� )3.6 )4.5 )1.9 )1.6� 2.0 2.5�
NN )7.2 )4.8 )9.3 )7.4 )9.7 )10.0 )4.1 )2.7TP )4.6 )6.7 )6.1 )10.7 )9.3 )11.7 )4.7 )7.4Controls )3.8 ± 3.4 )5.7 ± 3.6 )5.2 ± 3.5 )8.3 ± 4.0 )7.8 ± 4.0 )10.6 ± 4.2 )2.0 ± 3.0 )2.8 ± 2.9
Mean values are ± SD; �z < )1.65 (i.e. �z is more negative than )1.65); �z > 1.65.
Developmental prosopagnosia 2245
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

noted that in spite of the similarity of the general pattern of findings,there were also differences between our study and the two former ERPstudies (Bentin et al., 1999; Kress & Daum, 2003). Bentin et al.(1999) recorded ERPs from one prosopagnosic subject and did notfind laterality effects, whereas Kress & Daum (2003) as well as ourstudy showed larger effects over the right hemisphere. In addition, ETshowed a reduced N170 amplitude difference between faces andhouses; this is similar to Bentin et al. (1999) but in contrast to Kress &Daum (2003), who did not find amplitude differences between facesand nonface stimuli in two prosopagnosics. It is also noteworthy thatthis pattern mainly emerged over parieto-occipital electrodes, a findingwhich was also seen in controls for passive viewing of faces.The other three prosopagnosics (LT, NN and TP) showed normal
amplitude differences between faces and houses at TP8 and PO8. As it isasserted that the N170 is associated with structural encoding of faces(Eimer & McCarthy, 1999; Eimer, 2000c), its absence in ET suggeststhat ET’s impairment in face processing could be attributed to perceptualprocessing deficits. ET’s prosopagnosia is thus based on deficits prior toface identification or recognition, which is supported by behaviouraldata showing an impairment in face discrimination. By contrast, LT andNN performed 2 SD below average in this task, while their amplitudedifferences between faces and houses were indistinguishable from thecontrols. An alternative explanation would be that face processingdeficits, especially face encoding deficits, are not necessarily correlatedwith a lack of face selective N170 (Harris et al., 2005).Three prosopagnosics showed reliable N170 amplitude differences
between faces and nonface stimuli both at TP7 and TP8 and at PO7and PO8. ET exhibited the same pattern on the left side (TP7 andPO7). These data are in contradiction to previous findings (Bentinet al., 1999; Kress & Daum, 2003) of an absence of N170 amplitudedifferences in prosopagnosia, suggesting that the lack of a face-specific N170 is not a general feature of the condition. This later pointis in agreement with a recent combined magnetoencephalography andERP study (Harris et al., 2005), which reported normal M170 andN170 responses to faces and houses in at least two prosopagnosics.Our findings are also similar to those of functional neuroimagingstudies (Hasson et al., 2003; Avidan et al., 2005), of normal FFAactivation in prosopagnosic subjects. The normal electrophysiologicalactivity in parietotemporal and parieto-occipital regions does notnecessarily reflect the functional integrity of face processing. Therewas no systematic pattern of deviation from the control group amongthe DP subjects, nor were the scores on the visual and face processingtests systematically related to the ERP patterns. These findings sharesimilarities with recent functional neuroimaging data of normal face-related activations in ventral occipitotemporal regions in four CPindividuals (Avidan et al., 2005).To our knowledge there is so far no empirical evidence about N170
features elicited by caricatures. Studies examining the ability ofprosopagnosics to recognize caricatures are rare. All prosopagnosicsubjects of the current study were impaired at recognizing famousfaces on the basis of caricatures and photographs (Fig. 1). Thesefindings are similar to those of a study by DeGelder & Rouw (2000),which used face recognition tasks with photographs of famouscelebrities, caricatures and cartoons in a comparison of acquired anddevelopmental prosopagnosia. Both subjects could not recognize facesfrom photographs, caricatures and cartoons. All control subjectsshowed better performance for photos than for caricatures in thisstudy. Caricatures involve the exaggerated display of the specific facialfeatures characteristic of a particular person. It is conceivable thatprosopagnosics who report that they concentrate on characteristicfeatures (such as a large nose) when attempting to identify a personmay benefit from the emphasis on specific features in a caricature
relative to a realistic representation in a photograph. This pattern wasobserved in three prosopagnosics (LT, NN and TP), who showed atrend towards better recognition of famous people on the basis ofcaricatures than on the basis of photographs; this might reflect relianceon exaggerated features in caricatures. In contrast to this pattern, ETwas better at identifying famous people on the basis of photographs(see Fig. 1), which is similar to healthy controls. DP subjects mightdevelop different compensatory strategies because of their lifelongface-processing impairment. It is possible that LT, NN and TPconcentrate on salient cues in order to compensate their face-processing deficit whereas ET might benefit from another strategy todeal with prosopagnosia. The N170 data were not systematicallyrelated to the behavioural results (or compensatory strategies) in theDP group. In control subjects, N170 amplitudes for caricatures did notdiffer from those for houses (Figs 5 and 10). Pictures of houseselicited higher N170 amplitudes than pictures of caricatures in ET andTP (see Tables 3 and 4). The other two subjects did not differ from thecontrol group.It is remarkable, however, that the overall performance of the
prosopagnosics in recognizing famous people on the basis ofcaricatures did not differ greatly from their performance whenidentifying famous people on the basis of photographs (Fig. 1),suggesting that prosopagnosics may indeed benefit from the focus onprominent facial features.In a recent investigation, Le Grand et al. (2006) concluded that DP
is a heterogeneous impairment which cannot be predicted by poorperformance on one measure of face processing alone. This conclusionalso applies when behavioural and electrophysiological data areconsidered jointly. On the other hand, electrophysiological data mayindeed reflect an impairment in processing of faces in some DPindividuals, presumably as a manifestation of an impaired encoding ofconfigural information thought to be mirrored by the N170 (Rossionet al., 2003; Itier & Taylor, 2004; Boutsen et al., 2006).
Abbreviations
CP, congenital prosopagnosia; BFRT, Benton Facial Recognition Test; BORB,Birmingham Object Recognition Battery; d¢, measure of discrimination; DP,developmental prosopagnosia; ERP, event-related potentials; FFA, fusiformface area; RMT-F, Recognition Memory Test for faces; TAB, Tubinger AffectBattery.
References
Avidan, G., Hasson, U., Malach, R. & Behrmann, M. (2005) Detailedexploration of face-related processing in congenital prosopagnosia:2. Functional neuroimaging findings. J. Cogn. Neurosci., 17, 1150–1167.
Barton, J.J.S., Cherkasova, M.V., Press, D.Z., Intrilligator, J.M. & O’Connor,M. (2003) Developmental prosopagnosia: a study of three patients. BrainCogn., 51, 12–30.
Behrmann, M., Avidan, G., Marotta, J.J. & Kimchi, R. (2005) Detailedexploration of face-related processing in congenital prosopagnosia:1. Behavioral findings. J. Cogn. Neurosci., 17, 1130–1149.
Bentin, S., Allison, T., Puce, A., Perez, E. & McCarthy, G. (1996)Electrophysiological studies of face perception in humans. J. Cogn.Neurosci., 8, 551–565.
Bentin, S. & Deouell, L.Y. (2000) Structural encoding and identification in faceprocessing: ERP evidence for separated mechanisms. Cogn. Neuropsychol.,17, 35–54.
Bentin, S., Deouell, L.Y. & Soroker, N. (1999) Selective visual streaming inface recognition: evidence from developmental prosopagnosia. Neuroreport,11, 823–827.
Benton, A.L., Sivan, A.B., Hamsher, K., Varney, N.R. & Spreen, O. (1983)Contribution to Neuropsychological Assessment. Oxford University Press,New York.
2246 D. A. Minnebusch et al.
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247

Bodamer, J. (1947) Die Prosopagnosie. Arch. Psychiatrie Nervenkrankheiten,179, 6–53.
Boutsen, L., Humphreys, G.W., Praamstra, P. & Warbrick, T. (2006)Comparing neural correlates of configural processing in faces and objects:An ERP study of the Thatcher illusion. Neuroimage, 32, 352–367.
Breitenstein, C., Daum, I. & Ackermann, H. (1998) Emotional processingfollowing cortical and subcortical brain damage: contribution of the fronto-striatal circuty. Behav. Neurol., 11, 29–42.
Breitenstein, C., Daum, I., Ackermann, H., Lutgehetmann, R. & Muller, E.(1996) Erfassung der Emotionswahrnehmung bei zentralnervosen Lasionenund Erkrankungen: Psychometrische Gutekriterien der ‘Tubinger AffektBatterie’. Neurologische Rehabilitation, 2, 93–101.
DeGelder, B. & Rouw, R. (2000) Configural face processing in acquired anddevelopmental prosopagnosia: evidence for two separate face systems?Neuroreport, 11, 3145–3150.
Downing, P.E., Jiang, Y., Shuman, M. & Kanwisher, N. (2001) A cortical areaselective for visual processing of the human body. Science, 239, 2470–2473.
Duchaine, B.C. (2000) Developmental prosopagnosia with normal configuralprocessing. Neuroreport, 11, 79–83.
Duchaine, B.C., Dingle, K., Butterworth, E. & Nakayama, K. (2004) Normalgreeble learning in a severe case of developmental prosopagnosia. Neuron,43, 469–473.
Duchaine, B.C., Nieminen-von Wendt, T., New, J. & Kulomaki, T. (2003a)Dissociation of visual recognition in a developmental agnostic. Neurocase, 9,380–389.
Duchaine, B.C., Parker, H. & Nakayama, K. (2003b) Normal recognition ofemotion in a prosopagnosic. Perception, 32, 827–838.
Duchaine, B.C. & Weidenfeld, A. (2003) An evaluation of two commonly usedtests of familiar face recognition. Neuropsychologia, 41, 713–720.
Eimer, M. (2000a) Effect of face inversion on the structural encoding andrecognition of faces evidence from event-related brain potentials. Cogn.Brain. Res., 10, 145–158.
Eimer, M. (2000b) Event- related brain potentials sensitive to face perceptionand recognition. Clin. Neurophysiol., 111, 694–705.
Eimer, M. (2000c) The face- specific N170 component reflects late stages in thestructural encoding of faces. Neuroreport, 11, 2319–2324.
Eimer, M. & McCarthy, A. (1999) Prosopagnosia and structural encoding offaces: Evidence from event-related potentials. Neuroreport, 10, 255–259.
Farah, M.J. & McClelland, J.L. (1991) A computational model of semanticmemory impairment; modality specific and emergent category specificity.J. Exp. Psychol., 120, 339–357.
Farah, M.J., Rabinowitz, C., Quinn, G.E. & Lui, G.T. (2000) Early commitmentof neural substrates for face recognition. Cogn. Neuropsychol., 17, 117–123.
Harris, A.M., Duchaine, B.C. & Nakayama, K. (2005) Normal and abnormalface selectivity in the M170 response in developmental prosopagnosia.Neuropsychologia, 43, 2125–2136.
Hasson, U., Avidan, G., Deouell, L.Y., Bentin, S. & Malach, R. (2003) Face-selective activation in a congenital prosopagnosic subject. J. Cogn.Neurosci., 15, 419–431.
Humphreys, K., Avidan, G. & Behrmann, M. (2006) A detailed investigation offacial expression processing in congenital prosopagnosia as compared toacquired prosopagnosia. Exp. Brain Res., 176, 356–373.
Itier, R.J. & Taylor, M.J. (2004) N170 or N1? Spatiotemporal differencesbetween object and face processing using ERPs. Cereb. Cortex, 14,132–142.
Kanwisher, N. (2000) Domain specificity in face perception. Nat. Neurosci., 3,759–763.
Kanwisher, N., McDermott, J. & Chun, M.M. (1997) The fusiform face area: amodule in human extrastriate cortex specialized for face perception.J. Neurosci., 17, 4302–4311.
Kress, T. & Daum, I. (2003) Event-related potentials reflect impaired facerecognition in patients with congenital prosopagnosia. Neurosci. Lett., 352,133–136.
Le Grand, R., Cooper, P.A., Mondloch, C.J., Lewis, T.L., Sagiv, N., de Gelder,B. & Maurer, D. (2006) What aspects of face processing are impaired indevelopmental prosopagnosia?. Brain Cogn., 61, 139–158.
McCarty, G., Puce, A., Gore, J.C. & Allison, T. (1997) Face-specific processingin the human fusiform gyrus. J. Cogn. Neurosci., 9, 605–610.
Mitrushina, M.N., Boone, K.B. & Louis, F.D. (1999) Handbook of NormativeData for Neuropsychological Assessment. Oxford University Press, NewYork.
Nunn, J.A., Postma, P. & Pearson, R. (2001) Developmental prosopagnosia:should it be taken at face value? Neurocase, 7, 15–27.
Oldfield, R.C. (1971) The assessment and analysis of handedness: theedinburgh inventory. Neuropsychologia, 9, 97–113.
Osterrieth, P.A. (1944) Le test de copie d’une figure complexe. Arch.Psychologie, 30, 206–356.
Peelen, M.V. & Downing, P.E. (2005) Selectivity for the human body in thefusiform gyrus. J. Neurophysiol., 93, 603–608.
Riddoch, M.J. & Humphreys, G.W. (1993) Birmingham object recognitionbattery. LEA, Hove, UK.
Rossion, B., Schitz, C. & Crommelinck, M. (2003) The functionally definedright occipital and fusiform ‘face area’ discriminate novel from visuallyfamiliar faces. Neuroimage, 19, 877–883.
Rossion, B., Schitz, C., Robaye, L., Pirenne, D. & Crommelinck, M. (2001)How does the brain discriminate familiar and unfamiliar faces?: a PETstudy of face categorical perception. J. Cogn. Neurosci., 13,1019–1034.
Schwarzlose, R.F., Baker, C.I. & Kanwisher, N. (2005) Separate face and bodyselectivity on the fusiform gyrus. J. Neurosci., 25, 11055–11059.
Snodgrass, J.G. & Corwin, J. (1988) Pragmatics of measuring recognitionmemory: Applications to dementia and amnesia. J. Exp. Psychol. General.,117, 34–50.
Snodgrass, J.G. & Vanderwart, M. (1980) A standardised set of 260 pictures:Norms for name agreement, familiarity, and visual complexity. J. Exp.Psychol. Gen., 6, 174–215.
Varga, A.C. (1975) Declaration of Helsinki (Adopted by the 18th WorldMedical Assembly in Helsinki, Finland, and revised by the 29th WorldMedical Assembly in Tokyo, 1975). In: The Main Issue in Bioethics, revisededition. Paulist Press, New York, 1984.
Warrington, E.K. (1984) Recognition Memory Test. NFER-Nelson, Windsor,UK.
Yovel, G. & Kanwisher, N. (2004) Face perception: domain specific, notprocess specific. Neuron, 44, 889–898.
Developmental prosopagnosia 2247
ª The Authors (2007). Journal Compilation ª Federation of European Neuroscience Societies and Blackwell Publishing LtdEuropean Journal of Neuroscience, 25, 2234–2247