
Penaeus monodon (Fabricius) production related to water quality in East Java, Indonesia
Upload
mangaloreuniversityCategory
view
2download
0
lable at ScienceDirect
Fish & Shellfish Immunology 29 (2010) 724e732
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate/ fs i
Evaluation of immune response and resistance to diseases in tiger shrimp,Penaeus monodon fed with biofilm of Vibrio alginolyticus
S.R. Krupesha Sharma 1, K.M. Shankar*, M.L. Sathyanarayana 2, A.K. Sahoo, Rajreddy Patil,H.D. Narayanaswamy 2, Suguna Rao 2
Fish Pathology & Biotechnology Laboratory, Department of Aquaculture, College of Fisheries, Mangalore 575002, India
a r t i c l e i n f o
Article history:Received 20 January 2010Received in revised form25 June 2010Accepted 2 July 2010Available online 16 July 2010
Keywords:BiofilmVibrio alginolyticusPenaeus monodonImmune responseDisease resistance
* Corresponding author. Tel.: þ91 8242244356.E-mail address: [email protected] (K.M. S
1 Karwar Research Centre of Central Marine Fisheri581 301, Karnataka, India.
2 Department of Pathology, Veterinary College, BaIndia.
1050-4648/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.fsi.2010.07.016
a b s t r a c t
Immune response in juvenile tiger shrimp, Penaeus monodon fed with biofilm (BF) and free cells (FC) ofVibrio alginolyticus was studied by evaluating the hemocyte count, phenoloxidase activity and antibac-terial activity. The above immune responses were higher in BF fed shrimp than that in FC fed or controlshrimp. Among the different doses of BF of V. alginolyticus tested, 109 cfu g�1 shrimp day�1 for two weekscould evoke higher immune response. BF fed shrimp were more resistant to injection challenge withV. alginolyticus and whitespot syndrome virus (WSSV) with significantly higher RPS compared to thatwith FC fed and control shrimp. Better resistance was also reflected by rapid clearance of V. alginolyticusand WSSV from the hemolymph as confirmed by immunodot and histopathology.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Shrimp culture industry has often been affected by infectiousdiseases mainly of bacterial and viral etiology causing loss ofproduction [1]. As there are limitations in using antibiotics orvaccines for management of diseases in shrimp culture, there isgrowing interest in using immunostimulants. A range of prepara-tions such as live and killed bacteria, glucans, peptidoglycans andlipopolysaccharide (LPS) have been used to stimulate the immunesystem of shrimp [2]. Recently, scope for vaccination of shrimp hasalso been demonstrated [3]. Bacterin prepared from Vibrio harveyiwas found to enhance levels of hemolymph agglutinins in Penaeusmonodon [4]. Use of Vibrio alginolyticus probiotic in Penaeus van-namei culture could effectively improve the survival and growth bycompetitively excluding potential pathogenic bacteria [5].
Route of administration determines the success of immunosti-mulation [6]. Oral route was reported to be most beneficial foradministration of immunostimulants in shrimp [7]. Gut destructionof vaccines administered orally is a common hindrance in fish
hankar).es Research Institute, Karwar
ngalore 560 024, Karnataka,
All rights reserved.
leading to poor response [8]. Therefore, to withstand hostileconditions of stomach or foregut, a gut resistant form of immu-nostimulant or vaccine is always required. In general, compared tofree cells (FC), biofilm (BF) of bacteria is resistant to action ofantibiotics [9], antibodies [10] and disinfectants [11]. The resistantnature of BF of Aeromonas hydrophila has been used for successfuloral vaccination of carp [8]. The better performance of A. hydrophilaBF vaccine in carp by superior antigen delivery to lymphoid tissueshas been demonstrated by antigen localization using monoclonalantibodies [12]. Furthermore, as shrimp require continuousimmune stimulation with antigenic preparations due to theabsence of immune memory [6], a slow and steady releasing formof immunostimulant is needed. Against this background, thepresent study was undertaken to evaluate the immune responseand resistance to experimental microbial infection in P. monodonfed with BF of V. alginolyticus.
2. Materials and methods
2.1. Source, isolation and maintenance of V. alginolyticus
The bacterium was obtained from an infected P. monodon froma culture pond at Vypeen near Kochi, Kerala, India during June2006. The bacteriumwas isolated on 1.5% Tryptone soya broth (TSB;Himedia, Mumbai) containing 2% NaCl and identified based onbiochemical characteristics (Table 1). The isolate was preserved in

Table 1Characteristics of V. alginolyticus isolated from P. monodon.
Gram staining e
Shape Short rodGrowth on TCBS YellowLuminescence e
Swarming þOxidase production þCatalase production þAcid/gas productionGlucose AcidSucrose AcidMannitol AcidMaltose AcidSorbitol e
Lactose e
Galactose e
Arabinose AcidDecarboxylase of:Arginine e
Lysine þOrnithine þ
Growth in peptone with NaCl (%):0 e
1 þ3 þ6 þ8 þ10 þ
S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732 725
1.5% TSB containing 0.15% glycerol at�20 �C. For use, an aliquot wasrevived on nutrient agar slant and stored at 4 �C.
2.2. Source and isolation of WSSV
WSSV from infected P. monodon was purified as previouslydescribed [13]. Briefly, 10 g of infected tissue was homogenized in500 ml of TNE buffer (Tris Hydrochloride 50 mM, Sodium chloride400 mM, EDTA 5 mM, pH 8.5) and centrifuged at 3500�g for 5 minat 4 �C. The supernatant was centrifuged at 30,000�g for 30 min at4 �C. The upper loose pellet was removed carefully and the lowerwhite pellet suspended in 10 ml TM buffer (Tris hydrochloride50mM,Magnesium chloride 10mM, pH 7.5). After centrifugation at30,000�g for 20 min at 4 �C, the pellet was resuspended in 1 ml TMbuffer containing 0.1% Sodium azide. Protein concentration of thepellet was determined using the kit (Bangalore Genei, Bangalore,India).
2.3. Source and maintenance of shrimp
In all the experiments, juvenile P. monodon (1.6 � 0.3 g)procured from Matsyafed Prawn Hatchery, Kannur, India wereused. Shrimp were acclimated in the laboratory in circular FRPtanks containing 1000 l of chlorinated seawater (salinity 30 � 1&,pH 7.6� 0.2, temperature 26� 0.2 �C) with continuous aeration fortwo weeks before the commencement of the experiment. Duringthe acclimation period shrimpwere fed with commercial feed at 5%of body weight.
2.4. Development of BF and FC of V. alginolyticus
BF of V. alginolyticus was developed according to methodpreviously described for A. hydrophila [14]. Briefly, the isolate wasgrown on 0.3% chitin flakes suspended in 0.15% TSB and incubatedfor 3 d. Chitin with BF was harvested and heat inactivated at 80 �Cfor 10 min. FC of V. alginolyticus were grown in 1.5% TSB for 24 h.After harvesting by centrifugation, the cells were heat inactivated at60 �C for 10 min.
2.5. Preparation of experimental diet
Heat inactivated V. alginolyticus BF on chitin flakes was mixedwith standard commercial shrimp feed with the help of a binder(Protogel, Orgavet Pharma, Vijayawada, India). Similarly, heatinactivated FC of V. alginolyticus along with autoclaved chitin flakeswere mixed with feed using the binder. Autoclaved chitin flakesalone mixed with commercial shrimp feed with the binder servedas control feed.
2.6. Immune response in P. monodon fed with BF and FC
Shrimp were maintained in plastic tubs in three groups, eachconsisting of triplicate tubs containing 70 l sea water (salinity30 � 1&, pH 7.6 � 0.2, temperature 26 � 0.2 �C) with continuousaeration. Each tub was stocked with 30 shrimp (1.6 � 0.3 g). Group1 and 2 were fed with 108 cfu g�1 shrimp of BF and FC respectivelythrough feed for 7 d. The dose of BF is based on dose of BF ofA. hydrophila used for oral vaccination of carp (13). Shrimp in group3 which received autoclaved chitin flakes alone through feedserved as control. Hemolymph was collected from shrimp on day0 and 7 in their inter moult stage directly from the heart usinga tuberculin syringe fitted to a 26 gauge needle. Hemolymph fromtwo or more shrimp from a tub was pooled to 300 ml, mixed with700 ml of anticoagulant solution (Trisodium citrate 0.01 M, Sodiumchloride 0.34 M, EDTA 10 mM, Glucose 0.12 M, pH 7.4) and totalhemocyte count, phenoloxidase activity and antibacterial activitydetermined.
2.6.1. Total hemocyte count (THC)A drop of hemolymph-anticoagulant mixture was dispensed on
a Neubaur slide and cells allowed to settle for few seconds. Cellswere counted in five large squares and expressed as cells ml�1 ofhemolymph.
2.6.2. Phenoloxidase (PO) activityPO activity was measured spectrophotometrically by recording
the formation of dopachrome from L-dihydroxyphenylalanine(L-DOPA) as previously described [15] with modifications. Hemo-lymph-anticoagulant mixture was centrifuged at 700�g at 4 �C for10 min. After discarding the supernatant, the pellet was rinsed,resuspended gently in cacodylate-citrate buffer (Sodium cacodylate0.01 M, Sodium chloride 0.45 M, Trisodium citrate 0.10 M, pH 7.0)and centrifuged again. The pellet was then resuspended in 600 mlcacodylate buffer (Sodium cacodylate 0.01 M, Sodium chloride0.45 M, Calcium chloride 0.01 M, Magnesium chloride 0.26 M, pH7.0) and 100 ml of the cell suspension was incubated with 50 ml oftrypsin (1 mgml�1) as elicitor for 10 min at 20 �C. Fifty ml of L-DOPA(3mgml�1) was then added followed by 800 ml of cacodylate buffer5 min later. Optical density at 490 nm was measured using VarionCarry-50 spectrophotometer. PO activity was expressed asd (490)/50 ml of hemolymph. A control solution consisting of 100 mlof cell suspension, 50 ml cacodylate buffer (to replace trypsin) and50 ml of L-DOPA was used for background PO activity. The back-ground PO activity (range: 0.02e0.08) was subtracted from the POactivity of shrimp.
2.6.3. Antibacterial activityAntibacterial activity was assayed as previously described
[16,17] with modifications. V. alginolyticus was grown to log phasein 1.5% TSB supplemented with 2% NaCl. The cells were washedtwice with sterile Phosphate Buffered Saline (PBS; NaCl 137 mM,KCl 2.7 mM, NaH2PO4 1.4 mM, K2HPO4 4.3 mM, pH 7.4) by centri-fuging at 400�g for 10 min and resuspended in PBS to1�107 cfuml�1. The cell suspension (10 ml) was mixed with 80 ml of

S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732726
hemocyte suspension (prepared above) and 10 ml of sterile 1.5% TSB(supplemented with 2% NaCl) in duplicate wells of a microtitreplate. Final concentration of the bacterial cells was 1 �106 ml�1. Ascontrol, 90 ml of 1.5% TSB supplemented with 2% NaCl was incu-bated with 10 ml of bacterial suspension in duplicate wells. A thirdpair of duplicate wells containing 80 ml of cell suspension, 10 ml ofPBS and 10 ml of 1.5% TSB supplemented with 2% NaCl served ascontamination control. The contents of the plate were mixedproperly and from each well 50 ml was transferred to preformedTryptone Soya Agar (TSA) plates in duplicate and spread. Themicrotitre plates and the TSA plates were incubated at roomtemperature for 24 h. After incubation, colonies on the TSA plateswere counted and 50 ml from each well of the microtitre plate wastransferred to preformed TSA plates in duplicate and spread. Theplates were incubated for 24 h at room temperature and colonieswere counted. The plate counts thus obtained were expressed asSurvival Index (SI), SI ¼ cfu at time tt/cfu at time t0 where t0 is theinitial cfu and tt is the final cfu. SI values above one indicatedbacterial growth, whereas, values less than one indicated antibac-terial activity.
2.7. Standardization of BF dose
Four groups of shrimp in triplicate were maintained in circularplastic tubs containing 80 l of chlorinated sea water (salinity30 � 1&, pH 7.6 � 0.2, temperature 26 � 0.2 �C) with continuousaeration. Each tub was stocked with 30 shrimp. Group 1 was fed BFincorporated feed at 108, group 2 at 109 and group 3 at 1010 cfu g�1
shrimp day�1. Shrimp in group 4 which were fed normal shrimpfeed containing autoclaved chitin served as common control. Theshrimp were fed at 8 AM at 5% of body weight for 14 d. The wastewas siphoned out before and 3 h after feeding and 20% waterreplaced daily. Hemolymph collected on day 0, 7 and 14 was sub-jected to estimation of THC, PO activity and antibacterial activity asexplained in Section 2.6.
2.8. Determination of LD50 of V. alginolyticus
LD50 of V. alginolyticus in P. monodon was determined as previ-ously described [18]. Briefly, juveniles of P. monodon (1.6 � 0.3 g)were maintained in eight plastic tubs (80 l capacity), in duplicate,with 10 shrimp in each tub. Prior to the experiment, shrimp wererandomly checked for the presence of bacteria in the hemolymph.V. alginolyticus maintained on nutrient agar slant was cultured in1.5% TSB supplemented with 2% NaCl for 18 h on a shaker. The cellswere harvested, resuspended in PBS and serially diluted withsterile normal saline for enumeration on TSA plates. Shrimp inseven tubs were injected with 103, 104, 105, 106, 107, 108, and 109 cfuin 0.1 ml normal saline respectively on the ventral side on thesecond segment. Control shrimp in tub 8 received 0.1 ml of normalsaline only. Mortality was recorded at the end of 6,12, 24, 48, 72, 96,120, 144 and 168 h post injection. Hemolymph of moribund shrimpwas used for re-isolation of the bacterium using TCBS agar(Himedia, Mumbai).
2.9. Challenge with V. alginolyticus
Acclimated shrimp were maintained in 4 groups in plastic tubscontaining 80 l chlorinated sea water with continuous aeration. Ineach group there were triplicate tubs. Each tub consisted of 30shrimp (1.6 � 0.3 g). Based on dose standardization of BF cells, firstgroupwas fedwith BF incorporated feed at109 cfu g�1 shrimp day�1
for 14 d; second group with FC incorporated feed at109 cfu g�1
shrimp day�1; third group with normal shrimp feed containingautoclaved chitin flakes and the fourth group with normal shrimp
feed. Fromday15 onwards standard shrimp feedwas given to all thegroups. V. alginolyticus was grown to log phase in 1.5% TSB, har-vested and washed thrice with sterile PBS (pH 7.4). On day 15, eachshrimp in groups 1, 2 and 3 was injected with 1 � 106 cfu ofV. alginolyticus in 0.1 ml of normal saline on the second abdominalsegment. Shrimp in fourth group received 0.1 ml of normal saline.Hemolymph (0.1 ml) was aseptically collected from three shrimp ina replicate into 0.9 ml of sterile anticoagulant solution at 3, 6,12, 24,48, 72, 96 and 120 h post challenge (PC) for enumeration ofV. alginolyticus. The hemolymph-anticoagulantmixture (0.1ml)wastransferred and spread on triplicate TSA plates, incubated at roomtemperature for 24 h and cfu ml�1 of hemolymph was calculated.Mortality in remaining two replicates of each group was observedforfive days and relative per cent survival (RPS)was calculated as: 1-(% mortality in BF or FC group/% mortality in control group) � 100.Freshly dead shrimp were preserved in 10% neutral bufferedformalin (NBF) for histopathology. The tissues were processed,embedded in paraffinwax and the sections (3 m) were stained withhaematoxylene & eosin and histologic Gram’s stain of Brown andBrenn [19]. Number of hemocytic nodules (maximum 20) in a crosssection of abdomen was considered to classify the severity oflesions; 13e20 hemocytic nodules of various sizes in a section asgrade A; 7e12 as grade B; and 1e6 as grade C.
2.10. Challenge with WSSV
Acclimated shrimpweremaintained in 4 groups as in Section 2.9.On day 15 post feeding with experimental diets, each shrimp ingroup 1, 2 and 3 was injected with 50 ml (protein concentration:60 mg) of purified WSSV at the second abdominal segment. Theshrimp in group 4were injectedwith 50 ml of TMbuffer. Two shrimpfrom a tub in each group were collected every 24 h up to 7 d andtested forWSSV using Immunodot [20]. Shrimp from the remainingtwo tubs of each group were observed for 7 d for mortality and RPSwas calculated as above. Dead shrimp were immediately preservedin 10% NBF for histopathology. Tissue sections were stained withhaematoxylene & eosin and immunoperoxidase (IP) according tomethod previously described [19] to confirmWSSV. Briefly, sectionswere dewaxed with xylene and hydrated to water. After washingwith PBS (containing 0.3% tween), sections were treated with 1%bovine serum albumin (BSA) for 30 min and washed with PBS. Thesections were reactedwithmonoclonal antibody againstWSSV [20]for 45 min, washed with PBS and reacted with horse radish perox-idase conjugated goat antimouse IgG (1 in 200 in 1% BSA in PBS) for30 min. After washing with PBS, the sections were treated withDiaminobenzidine (Sigma, Bangalore) (0.6 mg/ml in 50 mM trisbuffer containing 150 mM NaCl and 0.03% hydrogen peroxide) for5 min. After washing with tap water, the sections were counterstained with 1% eosin for 3 min. The sections were dehydrated,clearedwith xylene andmounted using DPX. Dark brown colorationin tissue was considered as positive reaction. Severity of lesionsunder H & E staining was graded as A, B and C [21]. If more than 50%cells in the cuticular epithelium contained intranuclear occlusionbodies, the lesionwas considered as grade A; 50e5% as grade B andless than 5% as grade C.
2.11. Statistical analysis
Themean (�SD) values of immune response parameters and themean (�SD) cfu ml�1 hemolymph were statistically analyzed byTwo-Way-ANOVAwith Bonferroni Post-tests using GraphpadPrism(Version:4) for Windows. The LD50 (24 h) was calculated byemploying non linear fit of transformation with BoltzmannSigmoidal Equation using GraphpadPrism (Version: 4).

S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732 727
3. Results
3.1. Characteristics of bacterial isolate
The brief characteristics of V. alginolyticus isolated from theinfected P. monodon are shown in Table 1.
0
20
40
60
80
100
120
140
160
180
200
Control Free cells Biofilm cells
01X(
CH
T5
lm
sllec-1
)
day 0 day 7
a
a
a
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
0.18
0.2
Control Free cells Biofilm cells
(ytivitca
OP
)054/D
O
day 0 day 7
a
a
a
0
0.2
0.4
0.6
0.8
1
1.2
Control Free cells Biofilm cells
xednIlavivruS
day 0 day 7
a
a
a
Δ
a
b
c
Fig. 1. (a). Total hemocyte count of P. monodon fed with BF and FC of V. alginolyticus(Mean � SD, vertical bars indicate standard deviation, means with common super-scripts differ significantly): (b) PO activity of P. monodon fed with BF and FC of V.alginolyticus (Mean � SD, vertical bars indicate standard deviation, means withcommon superscripts differ significantly). (c) Antibacterial activity of P. monodon fedwith BF and FC of V. alginolyticus (Mean � SD, vertical bars indicate standard deviation,means with common superscripts differ significantly).
3.2. Immune response in P. monodon fed with BF and FC
Immune response in P. monodon fed with BF and FC of V. algi-nolyticus is shown in Fig.1(aec). Total hemocyte count on day 7wassignificantly higher (P < 0.001) compared to that on 0 day both inBF and FC groups. However, THC in BF group was significantlyhigher (P< 0.001) than that in FC group on day 7. A similar trend inphenoloxidase and antibacterial activity was recorded where it wassignificantly higher (P< 0.001) on day 7 in BF group compared to FCgroup.
0
20
40
60
80
100
120
140
160
01X(
CH
T5
lm
sllec-1
)
Day 0 Day 7 Day 14
10 cfu 10 cfuControl 10 cfu
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0.16
Day 0 Day 7 Day 14
10 cfu10 cfu10 cfuControl
0
0.2
0.4
0.6
0.8
1
1.2
xednIlavivruS
Day 0 Day 7 Day 14
10 cfu 10 cfuControl 10 cfu
(ytivitca
OP
)054/D
OΔ
a
b
c
Fig. 2. (a) Total hemocyte count of P. monodon fed with different doses of BF cells ofV. alginolyticus (Mean � SD, vertical bars indicate standard deviation). (b). PO activityof P. monodon fed with different doses of BF cells of V. alginolyticus (Mean � SE, verticalbars indicate standard deviation). (c) Antibacterial activity of P. monodon fed withdifferent doses of BF cells of V. alginolyticus (Mean � SD, vertical bars indicate standarddeviation).

0
1
2
3
4
5
6
7
8
9
0 6 12 24 48 72 96 120
01X
3l
mufc
-1
Time (h)
Control
FC
BFa,e
a
b,g
c.h
d, i
b,kc,l
d,m
e
f
f,j
g,k
h,l i,m
j
Fig. 4. V. alginolyticus cfu ml�1 of hemolymph of P.monodon after injection challenge(vertical bars indicate standard deviation; means with common superscripts differsignificantly between experimental groups).
S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732728
3.3. Standardization of BF dose
Immune response with different doses of BF is shown in Fig. 2(aec). Compared to control, THC increased with dosage from 108
to 1010 reaching highest with 1010 cfu on day 14. However, therewas no significant (P > 0.05) difference in THC between 109 and1010 cfu. A similar trend was observed with phenoloxidase andantibacterial activity which was highest in 1010 cfu with nosignificant difference between 109 and 1010 cfu. Also, PO and anti-bacterial activity values in all the dosage groups on day 14 weresignificantly higher (P < 0.001) than that on day 7. THC and POactivity values on day 7 were significantly higher (P < 0.001) thanthat on day 0 in all the experimental groups. In case of antibacterialactivity, the values were significantly higher (P < 0.001) in 109 and1010 cfu groups on day 7 when compared to day 0.
3.4. Determination of LD50 of V. alginolyticus
LD50 of V. alginolyticus was found to be 3.12 � 107 cfu shrimp�1
(Fig. 3). At higher concentrations (109 cfu shrimp�1), all the shrimpdied before 24 h. Therewas nomortality within 7 d post injection at103 cfu. No mortality was observed in shrimp injected with normalsaline.
3.5. Challenge with V. alginolyticus
V. alginolyticus (cfu ml�1) in hemolymph of BF group decreasedsignificantly (P < 0.001) from 24 h post challenge (PC) compared tothat in FC and control group (Fig. 4). The bacterial count in thehemolymph of FC group, although higher than in BF group, weresignificantly lower (P < 0.01) than that in control group. Bacterialcount in the hemolymph declined much earlier in BF groupcompared to that in FC or control group. The bacterial count in BFgroup was near zero from 96 h PC. No bacteria could be isolatedfrom the hemolymph of shrimp administered sterile saline. Re-isolation of the bacteria on TCBS agar revealed yellow, round, raisedand discrete colonies. RPS in experimental V. alginolyticus infectionwas significantly higher (P < 0.001) in BF fed shrimp (56.4)compared to that in FC group (18.8) (Fig. 5). In tissue sectionsstained with H & E, colonization of bacteria along with circum-scribed hemocytic accumulation as nodules forming capsules orplugs in the musculature and sinusoids was evident (Fig. 6). Lesionsconsisted of nodular accumulation of hemocytes around thebacteria, rounding and sloughing of hepatopancreatic cells into thetubular lumen and colonization of bacteria as basophilic amor-phous plaque in abdominal musculature. Sections stained with
3 4 5 6 7 8 9 10
0
25
50
75
100
Log cfu
Pe
r c
en
t m
orta
lity
Fig. 3. LD50 of V. alginolyticus to P. monodon juveniles.
Gram’s stain revealed extensive infiltration of hemocytes alongwith the presence of bacteria as pink short rods in the abdominalmusculature. Severity of lesions (number of hemocytic plugs ina cross section of abdominal tissue) was higher in control group(grade A: 8/10), moderate in FC treated group (grade B: 7/10) andmild in BF group (grade C: 9/10).
3.6. Challenge with WSSV
Shrimp in control group injection challenged with WSSV werepositive to WSSV by immunodot till the end of the experiment.Shrimp in FC and BF group were positive upto 144 h and 96 hrespectively (Table 2). Shrimp which were administered TM bufferonly were negative by immunodot throughout the period ofexperiment. RPS in experimental WSSV infection was significantlyhigher (P < 0.001) in BF group (62.1) compared to that in FC group(20.1) (Fig. 5). In histological sections, lesions were more conspic-uous in the cuticular epithelium. The cuticular epithelial cells oforgans like gills, haematopoietic tissue and antennal gland showedpresence of intra-nuclear occlusions in different stages of devel-opment under H & E staining (Fig. 7). Immunoperoxidase stainingof the infected cells of cuticular epithelium revealed dark browncrystalline material indicating positive reaction. Cuticular lesions asevidenced by the presence of occlusions in the hypertrophied
0
10
20
30
40
50
60
70
V.alginolyticus infection
WSSV infection
BF
FC
Fig. 5. Relative per cent survival of P. monodon after infection challenge with V. algi-nolyticus and WSSV.

Fig. 6. Microphotograph of muscle tissue of P. monodon fed with BF and FC of V. alginolyticus and injection challenged with V. alginolyticus showing hemocytic plugs: a. Controlgroup (H & E X 450); b. FC group (H & E X 450); c. BF group (H & E X 450); d. Control group (Gram’s stain X 1000)
S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732 729
nucleus of the cuticular epithelial cells were severe in controlshrimp (Grade A: 15/15), moderate in FC group (Grade B: 10/15) andmild in BF group (Grade C: 9/15).
4. Discussion
Immunostimulants have been administered to marine fish andshellfish through oral, immersion and injection methods. As
Table 2Results of immunodot of various treatment groups in experimental WSSV infection.
Groups hours post challenge
24 48 72 96 120 144 168
Control þ þ þ þ þ þ þFC þ þ þ þ þ þ e
BF þ þ þ þ e e e
TM buffer e e e e e e e
þ: Positive for WSSV.�: Negative for WSSV.
immersion and injection methods induce severe stress, oraladministration is considered to be an ideal antigen delivery systemto immunostimulate large number of shrimp at any stage of life [2].As shrimp lack memory, to facilitate continuous stimulation,immune response to biofilm of V. alginolyticus developed on chitinand fed through feed was evaluated in this study.
In the present study, effect of biofilm on three immuneparameters e THC, PO activity and antibacterial activity- has beenevaluated. THC has been reported to be a potential indicator ofimmune status in crustaceans [21] and a lower-than-normalnumber of circulating hemocytes in crustaceans correlate well witha reduced resistance to pathogens [22]. Apart from their role inmelanization, components of the PO activity system stimulateseveral cellular defense reactions, including phagocytosis, noduleformation, encapsulation, and hemocyte migration [23]. Activationof PO system has been used by many investigators to measureimmunostimulation in shrimp [6,24]. Antibacterial activity hasbeen evaluated by feeding immunostimulants to tiger shrimp [16].Mean values of THC, PO activity and antibacterial activity wereconspicuously elevated with BF treated shrimp than that with FC or
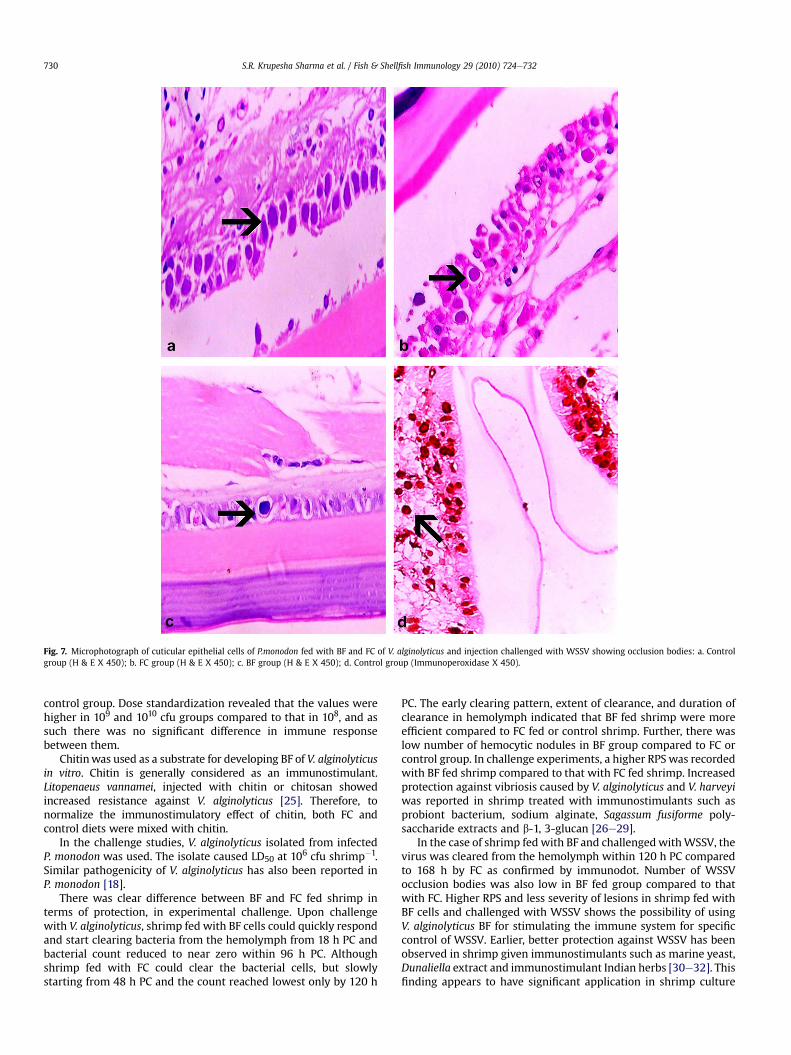
Fig. 7. Microphotograph of cuticular epithelial cells of P.monodon fed with BF and FC of V. alginolyticus and injection challenged with WSSV showing occlusion bodies: a. Controlgroup (H & E X 450); b. FC group (H & E X 450); c. BF group (H & E X 450); d. Control group (Immunoperoxidase X 450).
S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732730
control group. Dose standardization revealed that the values werehigher in 109 and 1010 cfu groups compared to that in 108, and assuch there was no significant difference in immune responsebetween them.
Chitinwas used as a substrate for developing BF of V. alginolyticusin vitro. Chitin is generally considered as an immunostimulant.Litopenaeus vannamei, injected with chitin or chitosan showedincreased resistance against V. alginolyticus [25]. Therefore, tonormalize the immunostimulatory effect of chitin, both FC andcontrol diets were mixed with chitin.
In the challenge studies, V. alginolyticus isolated from infectedP. monodon was used. The isolate caused LD50 at 106 cfu shrimp�1.Similar pathogenicity of V. alginolyticus has also been reported inP. monodon [18].
There was clear difference between BF and FC fed shrimp interms of protection, in experimental challenge. Upon challengewith V. alginolyticus, shrimp fed with BF cells could quickly respondand start clearing bacteria from the hemolymph from 18 h PC andbacterial count reduced to near zero within 96 h PC. Althoughshrimp fed with FC could clear the bacterial cells, but slowlystarting from 48 h PC and the count reached lowest only by 120 h
PC. The early clearing pattern, extent of clearance, and duration ofclearance in hemolymph indicated that BF fed shrimp were moreefficient compared to FC fed or control shrimp. Further, there waslow number of hemocytic nodules in BF group compared to FC orcontrol group. In challenge experiments, a higher RPS was recordedwith BF fed shrimp compared to that with FC fed shrimp. Increasedprotection against vibriosis caused by V. alginolyticus and V. harveyiwas reported in shrimp treated with immunostimulants such asprobiont bacterium, sodium alginate, Sagassum fusiforme poly-saccharide extracts and b-1, 3-glucan [26e29].
In the case of shrimp fed with BF and challengedwithWSSV, thevirus was cleared from the hemolymph within 120 h PC comparedto 168 h by FC as confirmed by immunodot. Number of WSSVocclusion bodies was also low in BF fed group compared to thatwith FC. Higher RPS and less severity of lesions in shrimp fed withBF cells and challenged with WSSV shows the possibility of usingV. alginolyticus BF for stimulating the immune system for specificcontrol of WSSV. Earlier, better protection against WSSV has beenobserved in shrimp given immunostimulants such as marine yeast,Dunaliella extract and immunostimulant Indian herbs [30e32]. Thisfinding appears to have significant application in shrimp culture

S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732 731
system where WSSV infection causes most serious disease thataffects the economy of shrimp farming [33].
Cumulative effect of elevated THC, PO activity and antibacterialactivity coupledwith quicker clearance of bacterium/virus reflectedwith higher RPS in BF fed group compared to that in FC group,suggests that the former is superior in enhancing the immunity inP. monodon. This might be due to better delivery of antigen from BF,since the cells are embedded in the glycocalyx matrix, facilitatinga slow and sustained release of antigens for a continuous immu-nological activation when compared to FC antigens that are nor-mally rapidly digested and released resulting in short-lived andinsufficient immune activation. The glycocalyx being a polymer ofneutral hexoses [34] acts similar to a microcapsule in protecting theantigen against degradation in the stomach as observed in encap-sulated antigen microspheres [35,36]. Improved antigen deliveryand protective responses in carp administered with BF cells ofA. hydrophila by oral route have been amply demonstrated [12].
Biofilm of V. alginolyticus in the present study could effectivelyimprove the immune response and resistance to diseases in P.monodon by invoking various immune responses, such as changesin hemocyte count, PO activity and antibacterial activity which arethe components of the crustacean immune system. In the absenceof an adaptive immune system, the immunostimulants in shrimpmust act on the innate immune system so as to boost the surveil-lance of and reactivity towards potential non-self threats [2]. Earlierstudies have also revealed changes in the above immune compo-nents when crustaceans were administered various immunosti-mulants such as LPS, Bacterins and glucan [37e39].
It is concluded that the BF cells of V. alginolyticus were superiorto FC for a better immunostimulation of shrimp. Oral administra-tion with BF cells of V. alginolyticus at a dose rate of 109 cfu g�1
shrimp day�1 for 14 d could effectively enhance resistance ofshrimp to V. alginolyticus and WSSV.
Acknowledgements
The study formed part of the Ph.D research work of the firstauthor. The first author is indebted to the Director, CMFRI forgranting study leave to pursue higher studies.
References
[1] Lightner DV. Diseases of cultured penaeid shrimp. In: McVey JP, editor. CRChandbook of mariculture. Crustacean aquaculture, vol. 1. Florida: CRC Press;1983. p. 289e320.
[2] Smith VJ, Brown JH, Hauton C. Immunostimulation in crustaceans: does itreally protect against infection? Fish and Shellfish Immunology 2003;15:71e90.
[3] Singh ISB, Manjusha M, Pai SS, Philip R. Fenneropenaeus indicus is protectedfrom white spot disease by oral administration of inactivated white spotsyndrome virus. Diseases of Aquatic Organisms 2005; 66 p. 265e70.
[4] Pais R, Khushivamani R, Karunasagar I, Karunasagar I. Effect of immunosti-mulants on the haemolymph haemagglutinins of tiger shrimp Penaeus mon-odon. Aquaculture Research 2008;39:1339e45.
[5] Garriques D, Arevalo G. An evaluation of the production and use of a livebacterial isolate to manipulate the microbial flora in the commercialproduction of Penaeus vannamei post larvae in Ecuador. In: Browdy CL, Hop-kins JS, editors. Swimming through troubled waters. Proceedings of thespecial session on shrimp farming, Aquaculture ’95. World AquacultureSociety, Baton Rough, p.53e59.
[6] Sung HH, Kou GH, Song YL. Vibriosis resistance induced by glucan treatmentin tiger shrimp (Penaeus monodon). Fish Pathology 1994;29:11e7.
[7] Azad IS, Panigrahi A, Gopal C, Paulpandi P, Mahima C, Ravichandran P. Routesof immunostimulation vis-a-vis survival and growth of Penaeus monodonpostlarvae. Aquaculture 2005;248:227e34.
[8] Azad IS, Shankar KM, Mohan CV, Kalita B. Biofilm vaccine of Aeromonashydrophila-standardization of dose and duration for oral vaccination of carps.Fish and Shellfish Immunology 1999;9:519e28.
[9] Brown MRW, Williams P. The influence of environment on envelop propertiesaffecting survival of bacteria in infections. Annual Review of Microbiology1985;39:527e56.
[10] Kreig DP, Helmke RJ, German VF, Mangos JA. Resistance of mucoid Pseudo-monas aeroginosa to non-opsonic phagocytosis by alveolar macrophagesin vitro. Infection and Immunity 1988;56:3173e9.
[11] Govan JRW. Mucoid strains of Pseudomonas aeruginosa: the influence ofculture medium on the stability of mucus production. Journal of MedicalMicrobiology 1975;8:513e22.
[12] Azad IS, Shankar KM, Mohan CV, Kalita B. Uptake and processing of biofilmand free-cell vaccines of Aeromonas hydrophila in Indian major carps andcommon carp following oral vaccination e antigen localization by a mono-clonal antibody. Diseases of Aquatic Organisms 2000;43:103e8.
[13] Xie X, Li H, Xu L, Yang F. A simple and efficient method for purification ofintact white spot syndrome virus (WSSV) viral particles. Virus Research2005;108:63e7.
[14] Azad IS, Shankar KM, Mohan CV. Evaluation of biofilm of Aeromonas hydro-phila for oral vaccination of carps. In: Flegel TW, Macrae IH, editors. Diseasesin Asian aquaculture III. Manila: Fish Health Section, AFS; 1997. p. 181e6.
[15] Liu CH, Chen JC. Effect of ammonia on the immune response of white shrimpLitopenaeus vannamei and its susceptibility to Vibrio alginolyticus. Fish andShellfish Immunology 2004;16:321e34.
[16] Smith VJ, Findlay C, Schnapp D, Hutton MCD. Detection of antibacterialactivity in the tissue or body fluids of marine invertebrates. In: Stolen JS,Fletcher TC, Smith SA, Zelikoff JT, Kaattari SL, Anderson RS, Soderhall K,Weeks-Perkins BA, editors. Techniques in fish immunology-4: immunologyand pathology of aquatic invertebrates. Fair Haven: SOS Publications; 1995. p.173e81.
[17] Sung HH, Yang YL, Song YL. Enhancement of microbicidal activity in the tigershrimp Penaeus monodon via immunostimulation. Journal of CrustaceanBiology 1996;16:278e84.
[18] Selvin J, Lipton AP. Vibrio alginolyticus associated with white spot disease ofPenaeus monodon. Diseases of Aquatic Organisms 2003;57:147e50.
[19] Luna LG. Manual of histologic staining methods of the armed forces instituteof pathology. London: McGraw Hill Book Co; 1968. p. 258.
[20] Anil TM, Shankar KM, Mohan CV. Monoclonal antibodies developed forsensitive detection and comparison of white spot virus isolates in India.Diseases of Aquatic Organisms 2002;51:67e75.
[21] Le Moullac G, Soyez C, Saulneir D, Ansquer D, Avarre JC, Levy P. Effect ofhypoxic stress on the immune response and the resistance to vibriosis ofthe shrimp Penaeus stylirostris. Fish and Shellfish Immunology 1998;8:621e9.
[22] Le Moullac G, Haffner P. Environmental factors affecting immune responses inCrustacea. Aquaculture 2000;191:121e31.
[23] Direkbusarakom S, Danayadol Y. Effect of oxygen depletion on some param-eters of the immune system of black tiger shrimp (Penaeus monodon). In:Flegel TW, editor. Advances in shrimp biotechnology. Bangkok: NationalCentre for Genetic Engineering and Biotechnology; 1998. p. 147e9.
[24] Devaraja TN, Otta SK, Shubha G, Karunasagar I, Tauro P, Karunasagar I.Immunostimulation of shrimp through oral administration of Vibrio bacterinand yeast glucan. In: Flegel TW, editor. Advances in shrimp biotechnology.Bangkok: National Centre for Genetic Engineering and Biotechnology; 1998.p. 167e70.
[25] Wang SH, Chen JC. The protective effect of chitin and chitosan against Vibrioalginolyticus in white shrimp Litopenaeus vannamei. Fish and ShellfishImmunology 2005;19:191e204.
[26] Rengpipat S, Rukpratanporn S, Piyatiratitivorakul S, Menasaveta P. Immunityenhancement in black tiger shrimp (Penaeus monodon) by a probiont bacte-rium (Bacillus S11). Aquaculture 2000;191:271e88.
[27] Cheng W, Liu CH, Kuo CM, Chen JC. Dietary administration of sodium alginateenhances the immune ability of white shrimp Litopenaeus vannamei and itsresistance against Vibrio alginolyticus. Fish and Shellfish Immunology 2005;18:1e12.
[28] Huang X, Zhou H, Zhang H. The effect of Sargassum fusiforme polysaccharideextracts on vibriosis resistance and immune activity of the shrimp, Fenner-openaeus chinensis. Fish and Shellfish Immunology 2006;20:750e7.
[29] Itami T, Takahashi Y, Nakamura Y. Efficacy of vaccination against vibriosis incultured kuruma prawns Penaeus japonicus. Journal of Aquatic Animal Health1989;1:238e42.
[30] Sajeevan TP, Philip R, Singh ISB. Immunostimulatory effect of a marineyeast Candida sake S165 in Fenneropenaeus indicus. Aquaculture 2006;257:150e5.
[31] Supamattaya K, Kiriratnikom S, Boonyaratpalin M, Borowitzka L. Effect ofa Dunaliella extract on growth performance, health condition, immuneresponse and disease resistance in black tiger shrimp (Penaeus monodon).Aquaculture 2005;248:207e16.
[32] Citarasu T, Sivaram V, Immanuel G, Rout N, Murugan V. Influence of selectedIndian immunostimulant herbs against white spot syndrome virus (WSSV)infection in black tiger shrimp, Penaeus monodon with reference to haema-tological, biochemical and immunological changes. Fish and Shellfish Immu-nology 2006;21:372e84.
[33] Flegel TW. Detection of major penaeid shrimp viruses in Asia, a historicalperspective with emphasis on Thailand. Aquaculture 2006;258:1e33.
[34] Costerton JW, Irvin R, Cheng KJ. The bacterial glycocalyx in nature and disease.Annual Review of Microbiology 1981;35:299e324.
[35] Dalmo RA, Liefson RM, Bogwald J. Microspheres as antigen carriers: studies onintestinal absorption and tissue localization of polysterene microspheres inAtlantic salmon, Salmo salar. Journal of Fish Diseases 1995;18:87e91.

S.R. Krupesha Sharma et al. / Fish & Shellfish Immunology 29 (2010) 724e732732
[36] Polk AE, Amsden B, Scarrat DJ, Gonzal A, Okahmafe AO, Goosen MFA. Oraldelivery in aquaculture: controlled release of proteins from chitosan-alginatemicrocapsules. Aquacultural Engineering 1994;13:311e23.
[37] Lorenzon S, De Guarrini S, Smith VJ, Ferrero EA. Effects of LPS on circulatinghaemocytes in crustaceans invivo. FishandShellfish Immunology1999;9:31e50.
[38] Ashida M, Soderhall K. The prophenoloxidase activating system in crayfish.Comparative Biochemistry and Physiology 1984;77B:21e6.
[39] Smith VJ, Soderhall K, Hamilton M. b 1,3-glucan induced cellular defensereactions in the shore crab, Carcinus maenas. Comparative Biochemistry andPhysiology 1984;77A:635e9.
Copyright © 2022 FDOKUMEN




















