
Comores : développement humain durable et élimination de ...
Upload
independentCategory
view
0download
0
Université Bordeaux 1 UMR PACEA 5199
Institut de Préhistoire et de Géologie du Quaternaire
Mémoire de Master 2 Sciences et Technologies
Mention Anthropologie Biologique, Paléoanthropologie et Préhistoire
Spécialité : Préhistoire
ETUDE DE L’EFFET DU PIETINEMENT HUMAIN SUR DES
OSSEMENTS DE FAUNE MELANGES A UN AMAS DE TAILLE
BARISIC Marilyne
Sous la direction de David Cochard et Véronique Laroulandie
Année 2005-2006

1

2
Remerciements
Je tiens à exprimer toute ma gratitude à mes encadrants, Véronique Laroulandie et David
Cochard, qui ont su être très présents tant pour me donner des conseils que pour m’orienter
dans mes recherches. Mes plus vifs remerciements à Jean Guillaume Bordes et Patrick Michel
qui m’ont aidée à réaliser cette expérience. Un grand et chaleureux merci à l’équipe de
marcheurs sans qui l’expérience n’aurait pas pu se faire : Wilfried Arrondel, François
Bachellerie, Christophe Boisselier, Myriam Boudadi-Maligne, Olivier Huard, Thibault
Morala, Marion Sasias, Johair Temma, Sébastien Verbecq. Je remercie également Cédric
Bauval, Emilie Claud, Isabelle Clouchoud, Arnaud Lenoble, Jean-Baptiste Mallye, William
Rendu pour leurs conseils et leur disponibilité. J’exprime toute ma gratitude aux entreprises
de boucherie qui m’ont gracieusement fourni une grande partie des ossements utilisés : la
société SOVIAGO, BOBIF, les bouchers du marché des Capucins. Mes remerciements vont
aussi à mon employeur qui m’a permis d’aménager du temps sur mes horaires de travail pour
réaliser mes recherches.
Enfin, je remercie ma famille et mes amis qui m’ont soutenue et beaucoup aidée pour la
réalisation de ce mémoire.

3
Sommaire
AVANT-PROPOS ..................................................................................................................... 5
I. ETAT DES CONNAISSANCES ET PROBLEMATIQUE ................................................... 7
1. Etat des connaissances........................................................................................................ 7
1.1. Economie de subsistance............................................................................................. 7
1.2. Comment définir une activité de boucherie ?.............................................................. 8
2. Strie de découpe : Boucherie ou piétinement ? ................................................................ 10
2.1. Une trace, des causes................................................................................................. 10
2.2. Piétinement : expérimentation et effets ..................................................................... 11
3. Objectif de l’étude............................................................................................................ 15
II. MATERIEL ET METHODE............................................................................................... 16
1. Choix des paramètres ....................................................................................................... 16
1.1. Quel matériel osseux ? .............................................................................................. 16
1.2. Quels produits lithiques ?.......................................................................................... 17
1.3. Densité des os par rapport aux outils lithiques :........................................................ 17
1.4. Quel sédiment ?......................................................................................................... 17
1.5. La cellule d’expérimentation :................................................................................... 18
1.6. Qui et comment ? ...................................................................................................... 19
2. Expérience........................................................................................................................ 19
2.1. Matériel ..................................................................................................................... 19
2.2. Réalisation de l’expérience : réorientation et biais ................................................... 22
2.3. Protocole en image .................................................................................................... 22
3. Méthode d’analyse ........................................................................................................... 27
3.1. Nettoyage et séchage du matériel piétiné.................................................................. 27
3.2. Observation, comptabilisation des traces et réalisation des planches ....................... 27
3.3. Création de la base de données ................................................................................. 29
III. RESULTATS ..................................................................................................................... 31
1. Représentativité des espèces et des parties anatomiques ................................................. 31
1.1. Une espèce présente-t-elle plus de stries ? ................................................................ 31
1.2. Y’a-t-il plus de stries sur un ossement ? ................................................................... 32
2. Schéma de répartition....................................................................................................... 34
2.1. Répartition en fonction des segments osseux............................................................ 34

4
2.2. Rôle du relief osseux ? .............................................................................................. 36
2.3. Regroupement ........................................................................................................... 37
3. Morphologie ..................................................................................................................... 38
3.1. Profondeur................................................................................................................. 38
3.2. Orientation................................................................................................................. 39
3.3. Longueur ................................................................................................................... 41
3.4. Particularités morphologiques................................................................................... 41
4. Tableau récapitulatif des résultats .................................................................................... 42
5. Autres effets du piétinement............................................................................................. 43
IV. DISCUSSION : APPORTS ET BIAIS .............................................................................. 44
1. Implications archéozoologiques ....................................................................................... 44
1.1. Similarité entre espèces : schéma de répartition correspondant au piétinement ....... 44
1.2. Différences entre espèces .......................................................................................... 45
1.3. Modèle de reconnaissance pour l’archéozoologie.................................................... 46
2. Biais et restrictions à l’application archéologique ........................................................... 46
2.1. Le terreau................................................................................................................... 46
2.2. L’intensité.................................................................................................................. 47
2.3. Le corpus expérimental ............................................................................................. 47
CONCLUSION ........................................................................................................................ 48
BIBLIOGRAPHIE ................................................................................................................... 49
ANNEXES ............................................................................................................................... 53

5
AVANT-PROPOS
Depuis le début du XXe siècle, il n’est plus ignoré en archéologie que les sites où sont
retrouvés des traces d’occupations humaines ou de passages d’animaux ont pu subir depuis le
dépôt de ces indices et jusqu’à leur étude de nombreuses modifications. La taphonomie est
« l’étude du passage (dans tous ses détails) des restes animaux de la biosphère dans la
lithosphère » (Efremov, 1940 in Costamagno, 1999). Les définitions de la taphonomie sont
multiples, C. Denys la définit comme « l’étude des lois de l’enfouissement et de la
fossilisation, et, par extension, des biais paléoécologiques résultant du passage d’un individu
d’un ensemble vivant à un ensemble fossile » (Denys, 2002).
Les processus de modification des traces d’occupations telles que des restes osseux, des éclats
de taille ou des restes végétaux sont multiples et complexes. Leurs origines sont diverses
(mécaniques, chimiques,...) et on distingue les processus intervenant avant l’enfouissement
des restes de ceux se produisant après. Ils sont principalement dus à des agents biologiques
tels que des champignons, des bactéries, des racines ou des bioturbations par exemple. En ce
qui concerne les agents responsables des modifications pré-enfouissement nous allons nous
focaliser sur les restes osseux. Ceux-ci peuvent être altérés ou modifiés par l’exploitation par
l’homme, le charognage, le piétinement humain ou animal, le « weathering ». Chacun de ces
agents est susceptible de laisser sur les restes osseux des traces plus ou moins spécifiques.
Souvent, les artéfacts sont touchés par plusieurs agents taphonomiques d’origines diverses qui
aident à recréer une « histoire » taphonomique des restes et du site où ils ont été trouvés.
Pourtant l’observation et l’analyse directe des traces portées par les restes osseux ne suffisent
pas à définir avec certitude l’origine même de ces traces.
La taphonomie nécessite d’effectuer en parallèle un travail de reconstitution expérimentale
pour vérifier des hypothèses nées des observations réalisées sur les pièces archéologiques
(Nicholson, 2001). L’expérimentation est « une méthode scientifique fondée sur l’utilisation
systématique d’expériences dans le but de vérifier des hypothèses » (C. Denys, 2002).
De plus elle est basée sur l’uniformité des lois physiques et biologiques à travers le temps.
Pour certains sites archéologiques, les résultats issus des études taphonomiques ou
archéozoologiques sont parfois problématiques. Les marques portées par les ossements telles
que les traces de découpe, de raclage, les fracturations peuvent avoir différentes origines mais
sont parfois trop vite interprétées comme d’origine anthropique sans une réelle étude du
contexte archéologique et sédimentaire (Hill, 1986). Ainsi, on observe de temps à autre dans

6
les sites archéologiques que la fréquence et/ou la morphologie des traces de découpe portées
par les os sont aléatoires ou anarchiques (Oliver, 1989) : seulement quelques os isolés portant
de telles traces ou bien une répartition non spécifique des traces sur les os.
Un des agents taphonomiques pouvant créer de telles marques est le piétinement. Il peut
résulter du passage de faune ou d’humains dont le mouvement provoque l’entrée en contact
des particules ou fragments rocheux contenus dans le sédiment avec l’os.
Jusque là, peu d’auteurs ont mené des expérimentations portant sur les modifications de la
surface des restes osseux entraînées par le piétinement humain (cf. Behrensmeyer et al., 1986
; Olsen et Shipman, 1988).
Ici nous avons voulu réaliser une expérience pour constater les effets du piétinement sur des
os longs du squelette appendiculaire de mésofaune (blaireau, lapin, pigeon) et de macrofaune
(mouton). Ces éléments sont piétinés dans un substrat contenant des éléments lithiques de
type lame, éclat, nucléus de toutes tailles contenus dans un substrat d’origine végétale. Notre
but est d’une part de voir les modifications qu’a pu causer le contact entre éléments lithiques
et éléments osseux sous l’effet du piétinement, en particulier les traces. D’autre part, cette
étude vise à mettre en évidence les particularités de ces marques et les différences qui peuvent
exister entre les espèces.
En première partie de cette étude, nous dresserons un bref état des lieux des connaissances sur
les traces de consommation sur la petite faune à la fin du Paléolithique. Nous ferons ensuite
une revue des recherches et des problématiques ayant porté jusqu’à présent sur les stries de
découpe et le piétinement. Puis nous détaillerons les paramètres choisis pour l’expérience et le
protocole expérimental. Ceci nous mènera à la présentation des résultats préliminaires et des
observations réalisées à l’issu de l’expérience, pour enfin confronter et discuter nos résultats
par rapport à notre problématique et aux expériences précédentes.

7
I. ETAT DES CONNAISSANCES ET PROBLEMATIQUE
Cette première partie a pour objet de faire un bref panorama des savoirs et des expériences
autours de la consommation de la faune pendant le Paléolithique supérieur et de l’état des
restes osseux retrouvés dans les sites archéologiques. Nous verrons que les traces laissées lors
du traitement boucher peuvent être imitées par un autre phénomène : le piétinement. La
similarité de ces traces rend nécessaire des études expérimentales. Nous justifierons la notre
dans la troisième sous partie de ce chapitre.
1. Etat des connaissances
C’est à partir des vestiges fauniques issus des fouilles archéologiques que des hypothèses
sur la faune consommée et utilisée par les hommes sont émises. Bien que l’état de nos
connaissances à ce sujet soit assez vaste, beaucoup de problèmes se posent encore. Ils
concernent la présence, la consommation ou l’utilisation de certaines espèces dans des sites
archéologiques mais aussi les techniques employées pour la découpe des carcasses.
1.1. Economie de subsistance
Le mode de subsistance des hommes du Sud Ouest de la France au Paléolithique supérieur
consiste essentiellement en l’exploitation de grande faune (cheval, cerf,...). A la fin du
Magdalénien intervient une intensification et une diversification du spectre de chasse en
direction de certains petits gibiers : oiseaux, poissons, lagomorphes. L’interprétation des
traces de consommation et des conditions d’accumulation font toujours débat (Costamagno,
1999).
D’après Laroulandie (2004), avant le Tardiglaciaire, la consommation de l’avifaune par les
hommes reste incertaine ou exceptionnelle car la majorité des traces retrouvées sur les
ossements sont des traces de manufacture. A la fin du Magdalénien, les traces de
consommation sont sur la Chouette Harfang ou le lagopède en particulier (Laroulandie, 2004).
En ce qui concerne les mustélidés, leur présence est souvent attestée dans les sites
archéologiques en France (Mallye et Laroulandie, 2004). En Italie, à Grotta della Madonna,
des traces de découpe ont pu être identifiées sur des restes osseux de blaireaux (23,1 %) (Fiore
et al., 2001).
Pour les lapins, en général à la fin du Paléolithique, le pourcentage d’os portant des marques
de découpe est de 2% (Cochard, 2004). Pour certaines études, comme par exemple celle de

8
Manuel Perez Ripoll à la Cova de las Cendres (2004), la quantité de traces de découpe sur les
ossements de lapin est beaucoup plus grande (plus de 50% des restes osseux de lapins). Il est
difficile d’affirmer que les traces en question sont bien toutes dûes à de la boucherie et non à
un autre phénomène.
1.2. Comment définir une activité de boucherie ?
La méthode de découpe va être orientée par ce que l’on veut faire de la carcasse ou de la
partie anatomique (Binford, 1981 ; Guilday et al., 1962), mais aussi par l’anatomie même de
la pièce à découper : parties plus charnues, insertions musculaires, tendons,... (Abe et al.,
2002 ; Fiorillo, 1991). Aussi, il est possible de distinguer une récurrence dans la distribution
des traces laissées par l’outil sur l’os, mais aussi que ces traces sont placées en fonction de
l’anatomie (Egeland, 2003 ; Guilday et al., 1962).
La boucherie faite sur la faune peut modifier la surface de l’os de manières diverses. Les
modifications les plus fréquentes sont des enfoncements de la surface de l’os, des marques de
découpe sur les diaphyses ou les épiphyses, des traces de raclage (Binford, 1981 ; Fisher,
1995 ; Shipman and Rose, 1983). Elles sont considérées depuis les premiers travaux de H.
Martin (1907 in Costamagno, 1999) comme des indices anthropiques certains (Egeland,
2003 ; Fiorillo, 1984 ; Morlan, 1984 ; Shipman and Rose, 1983). Pourtant, on peut souligner
qu’aucune de ces modifications n’est engendrée sciemment par la main de l’homme, Lyman
(1994) parle d’épiphénomène d’origine accidentelle. Etant donnée la diversité des traces
portées par les ossements, certains auteurs ont souligné la nécessité de clarifier la définition et
les critères de reconnaissance de celles-ci (Fisher, 1995 ; Morlan, 1983 ; Shipman and Rose,
1984).
Depuis le début des années 50, des recherches proposant de mieux définir les activités de
boucherie (Lyman, 2005) et leurs conséquences se sont orientées dans 3 voies différentes :
l’observation ethnologique, l’expérimentation et l’observation archéologique.
Tout d’abord, les études ethnoarchéologiques consistent en l’observation de populations
exploitant la faune en vue de la consommer et de l’utiliser. Le but des auteurs réalisant ce type
d’étude est de décrire les techniques de découpe, les traces laissées sur les ossements d’où la
viande a été extraite et globalement l’utilisation des carcasses pour formuler des hypothèses
sur les méthodes d’exploitation des hommes au Paléolithique. On peut citer L.R. Binford qui a
suivi les Eskimo Nunamiut (1978, in Binford, 1981), ou encore les travaux de Bunn and Kroll

9
(1988, in Egeland, 2003) qui ont étudié les bouchers Hadza, Crader (1983, in Egeland, 2003)
qui travailla lui sur les techniques de boucherie sur éléphant des Bisa.
L’observation archéologique réalisée sur des collections d’ossements assez importantes
d’époques plus ou moins récentes, a été développée en vue de reconstituer, de même que pour
l’ethnologie, les pratiques bouchères. Dans cette optique, Guilday (1962) a étudié les restes
fauniques laissés par une tribu amérindienne datant du XVIIe siècle pour reconstituer les
schémas de traitement en fonction des espèces et de leur utilisation.
Enfin, l’expérimentation de boucherie sur carcasses, fraîches ou non, permet en tentant de se
rapprocher au maximum des conditions archéologiques d’envisager des schémas de boucherie
: observation de la récurrence, positions, fréquences des traces et morphologie (Dominguez
Rodrigo, 1997 ; Egeland, 2003 ; Selvaggio, 1994). Par exemple, Egeland (2003) a
expérimentalement découpé des membres postérieurs de grands herbivores. Il a pu observer
que la totalité des éléments portaient au moins une trace et qu’il y avait, pour un même
ossement, une récurrence quant à la localisation des marques de découpe.
Bien que toutes ces études aient donné des résultats, ceux-ci n’en restent pas moins assez
subjectifs et très dépendants du boucher et d’autres conditions. Ainsi, certains biais ont été
soulignés ; pour certains auteurs la quantité des trace de découpe dépend de l’adresse du
boucher, de la rapidité (Guilday et al., 1962), de la fraîcheur de la viande : gel, rigidité
cadavérique(Binford, 1981 ; Egeland, 2003). Guilday (1962) fait aussi remarquer que plus le
boucher est habile et plus l’activité sera précise et sans empressement, moins l’os découpé
portera des traces.
Bien que nombre de chercheurs s’accordent à dire que la trace de découpe est un indice fort
de l’exploitation d’une carcasse par l’homme, la reconnaissance et l’interprétation de traces
d’activité de boucherie dans certains sites sont remises en question. En effet, deux régions du
monde en particulier sont prises dans cette polémique : les sites à premiers hominidés en
Afrique de l’Est et les plus anciens sites d’Amérique du Nord.
En ce qui concerne l’Afrique de l’Est, A. Hill (1986) a tout d’abord souligné le manque de
recul des résultats de Shipman and Rose (1983), puis H. T. Bunn (1989) (le manque de
connaissances sur la formation des accumulations de vestiges fauniques et osseux à Olduvai
interprétés comme d’origine anthropique.
En outre, J. Oliver (1989) confronte ses résultats à Shield Trap Cave, Montana, avec ceux de
Shipman and Rose (Shipman and Rose, 1983) à Olduvai. Les ossements de bisons trouvés
dans la grotte, pourtant inaccessibles à l’homme, portent des pseudo-stries de découpe.

10
Ces polémiques entre auteurs sont révélatrices des manques de clarté des critères quant à la
reconnaissance et à la définition des caractéristiques d’une trace de découpe tant au niveau
morphologique que de la distribution sur l’os. Elles ont ouvert la voie à l’expérimentation et à
l’utilisation de nouveaux outils comme le microscope à balayage dans les années 80 pour
répondre à ces lacunes comme l’ont fait entre autres Shipman et Rose (1983, 1984). Bien que
la polémique reste vive encore aujourd’hui (Dominguez Rodrigo, 1997), les critères les moins
critiqués pour définir un schéma de boucherie restent ceux de E. D. Guilday (1962) :
récurrence dans la distribution des traces et localisation en fonction de l’anatomie.
2. Strie de découpe : boucherie ou piétinement ?
Comme on vient de le voir, de multiples études ont été réalisées sur les activités de
boucherie. Pourtant, aucun critère clair n’a pour l’instant été défini pour distinguer avec
certitude une trace de découpe d’une strie créée par du piétinement. «It is already evident on
simple theoritical grounds that there will be non absolutely diagnostic set of criteria that will
always allow a clear choice between causal agents for single striae» (White, 1985 in Fisher
1995). L’expérimentation apparaît, dans ce contexte, nécessaire pour connaître les
conséquences du piétinement quelles que soient les espèces.
2.1. Une trace, des causes
Pour R.L. Lyman (1994), la «trace ou effet taphonomique» est le résultat :
• de l’action de l’«agent taphonomique» qui est l’origine de l’énergie portée sur l’os,
• et du «processus taphonomique», l’action (ou la force) qui va entraîner la
modification de la surface de l’os par l’agent (transport, chute et glissement d’un
fragment rocheux sur l’os...).
Un même agent peut créer une même trace par un processus différent. De plus, et c’est ce que
souligne D. Gifford-Gonzalez (1991 in Fisher, 1995) des agents divers peuvent être à l’origine
de traces très similaires.
Les études expérimentales sur les traces présentes sur les restes osseux se succèdent dès les
années 60-70 pour tenter de définir les agents causant ces pseudo-traces et des caractères
spécifiques aux traces de découpe ont pu être mis en évidence pour les distinguer des traces
de dents de carnivores, de rongeurs, de sers ou de racines (Binford, 1981 ; Shipman and Rose,
1983). Les études utilisant le microscope électronique à balayage ont été entreprises pour

11
déterminer la morphologie des traces de découpe réalisées expérimentalement (Bromage and
Boyde, 1984 ; Walker and Long, 1977). Reprenant les études de différents auteurs, Fisher
(1995) dresse le portrait type d’une trace de découpe : strie allongée,étroite et linéaire, ayant
souvent une section en V. Elle s’accompagne occasionnellement d’autres figures : effet
d’épaulement, barbes,... Les travaux de Shipman et Rose (1983,1984) ont conclu à une
différence morphologique au niveau microscopique des traces de découpes par rapport aux
traces produites par tous les autres agents responsables des modifications pré-enfouissement.
Pour d’autres auteurs, il est pourtant impossible de distinguer une trace produite par un outil,
d’une trace produite par piétinement dans un sédiment sableux, par le contact d’un grain de
sable sur l’os, et ce même au niveau microscopique (Andrews and Cooks, 1985 ;
Behrensmeyer et al., 1986 ; Fiorillo, 1984 ; Oliver, 1989).
Ainsi, les travaux cités plus haut d’Oliver (1989) et de Shipman et Rose (1983) illustrent bien
la difficulté d’interprétation liée à la similarité des traces produites par un même élément (ici
de nature lithique) mais par un acteur différent : «if any two otherwise distinct agents share
certain mechanics in the manner which they impact energy to bone then similar damages may
be created» (Oliver, 1989, p. 74). Sur le site de Shield Trap Cave, de nombreux ossements de
bison portent notamment des pseudo-marques de découpe créées par la chute du plafond de la
grotte ou le mouvement des ossements contre la paroi ou la roche du sol. La similarité des
traces au niveau morphologique s’explique d’une part par la géologie du site qui est creusé
dans un calcaire contenant des rognons de silex (Oliver, 1989); ces derniers se fracturent en
tombant de la paroi, créant ainsi des géofacts tels que des éclats.
Ce dernier exemple montre qu’il y a vraiment une nécessité de tester les effets de différents
agents causals soumis à diverses forces d’origine non humaine en contact avec des ossements.
Des éléments lithiques tels que des grains de sable ou des éclats de silex mis en contact avec
la surface d’ossements par du piétinement peuvent produire des marques ressemblant
fortement à des traces de découpe. L’expérimentation des effets du piétinement dans de telles
conditions est incontournable pour analyser les traces produites et d’en dégager des critères
diagnostiques.
2.2. Piétinement : expérimentation et effets
L’expérimentation en archéozoologie et en taphonomie répond à des problèmes rencontrés
sur le terrain. Cette démarche scientifique a accompagné l’archéologie depuis ses débuts. On
peut citer Edouard Lartet qui, en 1860, réalisait déjà une expérience de boucherie

12
(Costamagno, 1999) : utilisation d’outils en pierre sur os frais et comparaison avec du
matériel préhistorique. Les expérimentations portant sur les effets du piétinement des objets
archéologiques (matériel osseux, lithique, céramique,...) ont débuté dans les années soixante
et soixante-dix avec F. Bordes (Bordes, 1961 in Mc Brearthy et al., 1998) et E. D. Stockton
(1973). Elles se sont ensuite orientées globalement vers deux pistes de recherche :
• études au niveau spatial, mouvements des artéfacts induits par le piétinement
• études sur les conséquences au niveau physique de l’os telles que les traces de découpe,
fracturation, polis…
Piétinement et déplacements dans le sol :
Divers auteurs (Stockton, 1973 ; Courtin et Villa, 1982 ; Gifford-Gonzalez et al., 1985 ;
Nielsen, 1991 ; Lenoble et Bordes, 2002) ont réalisé des expérimentations dans diverses
conditions pour tester l’effet du piétinement sur le déplacement des objets contenus dans le
sol.
Les objets utilisés pour les expériences réalisées par ces auteurs sont : des outils taillés en
verre (Stockton, 1973), des outils lithiques (Gifford-Gonzalez et al., 1985), des ossements,
des débris de céramique ou encore des pièces lithiques (Courtin et Villa, 1982 ; Nielsen,
1991). Tous ces auteurs ont utilisé un ou plusieurs sédiments pour la réalisation de leur
expérience : sable (Stockton), gravier argileux ou «muddy gravel» (Nielsen), silt sableux
(Gifford-Gonzalez). Enfin, Lenoble et Bordes (2002) ont couplé une activité de ruissellement
au piétinement exercé sur des artéfacts lithiques.
Stockton (1973) décrit l’effet de deux composantes l’une verticale « treadage- vertical action
of the foots » et l’autre horizontale« scuffage – horizontal action of the foots ».
Le piétinement entraîne deux types de déplacement des objets dans le sol :
o des déplacements verticaux
o des déplacements horizontaux : Nielsen décrit une zone périphérique formée par
les éléments les plus volumineux et une zone centrale de « trafic » formée, elle, par
les éléments petits ou moyens parfois enfouis.
Plusieurs paramètres influent sur ces mouvements :
o la nature du substrat : degré de compaction et d’humidité déterminant la
pénétrabilité
o intensité du piétinement
o la taille et la forme de l’objet

13
Conséquences des mouvements spatiaux : Le piétinement peut provoquer des déplacements
verticaux aboutissant au mélange des couches et des déplacements horizontaux des objets, qui
peuvent être importants.
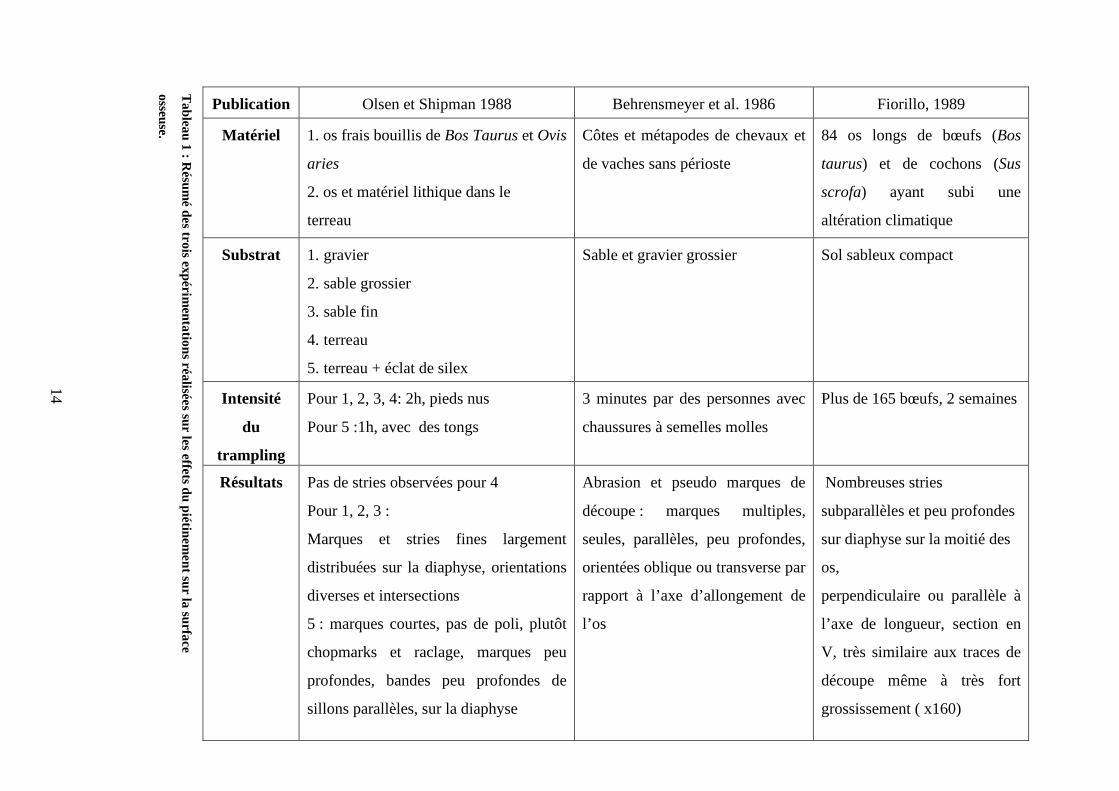
Piétinement et altérations de l’état de surface des ossements (cf. tableau 1)
En ce qui concerne le piétinement sur ossements d’animaux, les auteurs ont utilisé en
général des os de grands herbivores contenus dans différents types de sédiment
(Behrensmeyer et al. 1986 ; Olsen et Shipman,1988 ; Fiorillo, 1984,1989). Les deux
premières expériences ont mis en œuvre du piétinement humain et aboutissent selon
l’interprétation des auteurs à des résultats assez antagonistes en ce qui concerne les critères
morphologiques des traces. Pour (Olsen et Shipman, 1988), les traces issues du mouvement
du sédiment contre la surface osseuse ont des caractères qui diffèrent clairement des traces de
découpe faites expérimentalement ou retrouvées sur du matériel archéologique. Par contre, les
résultats obtenus par Fiorillo (1984, 1989) et Behrensmeyer (1986) tendent à montrer qu’au
contraire, les particules du sédiment créent des traces qui peuvent être confondues et ce même
à fort grossissement avec des traces de découpe. Oliver (Oliver, 1989) avait déjà mis en
évidence avec les restes de bisons de Shield Trap Cave une telle similarité entre traces issues
du piétinement et traces de boucherie. Par contre, globalement les schémas de répartition que
ces auteurs mettent en avant sont similaires et marqués par des caractères aléatoires :
diaphyses touchées en majorité, placements aléatoires par rapport à l’anatomie, orientations
diverses. Pour Behrensmeyer et al. (1986), il y a plus de traces sur la surface de l’os exposée
vers le haut et sur des zones plates, bombées ou convexes, les stries peuvent être isolées ou en
groupe.
On pourra constater dans le tableau ci-dessous qui résume les trois expériences majeures,
que les auteurs ont choisi des sédiments très abrasifs comme du sable pour étudier l’effet des
particules. Olsen et Shipman (1988) ont utilisé du terreau contenant une vingtaine d’éclats de
silex, c’est la seule expérience de ce type réalisée jusqu'à présent. Aussi, il serait très
intéressant de réitérer le même genre d’expérience avec différents paramètres afin
d’approfondir et de développer les pistes de recherches.
14
Tableau 1 : R
ésumé des trois expérim
entations réalisées sur les effets du piétinem
ent sur la surface
osseuse.
Fiorillo, 1989
84 os longs de bœufs (Bos
taurus) et de cochons (Sus
scrofa) ayant subi une
altération climatique
Sol sableux compact
Plus de 165 bœufs, 2 semaines
Nombreuses stries
subparallèles et peu profondes
sur diaphyse sur la moitié des
os,
perpendiculaire ou parallèle à
l’axe de longueur, section en
V, très similaire aux traces de
découpe même à très fort
grossissement ( x160)
Behrensmeyer et al. 1986
Côtes et métapodes de chevaux et
de vaches sans périoste
Sable et gravier grossier
3 minutes par des personnes avec
chaussures à semelles molles
Abrasion et pseudo marques de
découpe : marques multiples,
seules, parallèles, peu profondes,
orientées oblique ou transverse par
rapport à l’axe d’allongement de
l’os
Olsen et Shipman 1988
1. os frais bouillis de Bos Taurus et Ovis
aries
2. os et matériel lithique dans le
terreau
1. gravier
2. sable grossier
3. sable fin
4. terreau
5. terreau + éclat de silex
Pour 1, 2, 3, 4: 2h, pieds nus
Pour 5 :1h, avec des tongs
Pas de stries observées pour 4
Pour 1, 2, 3 :
Marques et stries fines largement
distribuées sur la diaphyse, orientations
diverses et intersections
5 : marques courtes, pas de poli, plutôt
chopmarks et raclage, marques peu
profondes, bandes peu profondes de
sillons parallèles, sur la diaphyse
Publication
Matériel
Substrat
Intensité
du
trampling
Résultats

15
3. Objectif de l’étude
Comme on vient de le voir plus haut les expériences concernent exclusivement des
ossements de grands herbivores. Globalement et jusqu’à présent, l’expérimentation sur le
piétinement n’a jamais concerné les petits mammifères ou les oiseaux bien que présents dans
de nombreux sites de la fin du Paléolithique Supérieur en Europe de l’Ouest. Certains de ces
ossements portent pourtant des traces de découpe. Ces traces pouvant être créées par un agent
taphonomique tel que des outils en pierre, il est important de savoir si ce même agent mis en
action non par la main de l’homme mais par un mouvement de piétinement va produire des
stigmates. En effet, des indices comme la fréquence, la récurrence des traces et les parties
anatomiques concernées sont des indices qui peuvent mener à la conclusion d’une activité de
boucherie sur le squelette. Ainsi, en recréant expérimentalement une couche archéologique
contenant des os longs d’oiseaux et de mammifères de petite et moyenne tailles mélangés à
divers produits lithiques, nous pourrons tester l’effet du piétinement sur la surface osseuse. Le
piétinement va-t-il permettre le contact entre les pièces lithiques et la surface des ossements et
donc la création de marques ? Quelles vont être leurs caractéristiques (localisation,
morphologie, fréquence) ? Pourrons-nous constater des différences interspécifiques ?

16
II. MATERIEL ET METHODE
Dans ce chapitre, nous définirons tout d’abord tous les paramètres et matériaux choisis
ainsi que les raisons de ces choix ; puis, grâce au compte rendu expérimental, nous verrons la
réalisation et les adaptations de certains des choix théoriques.
1. Choix des paramètres
Dans cette sous partie, nous justifions le choix de chacun des paramètres de l’expérience de
la forme de la cellule aux chaussures portées par les expérimentateurs. Il ne faut pas négliger
l’importance de ces choix pour coller au maximum à la réalité archéologique, optimiser les
chances d’arriver à notre but et laisser la plus petite part possible au hasard.
1.1. Quel matériel osseux ?
⊗ Espèces :
Trois espèces de petite faune ont été utilisées : lapin (Orictolagus cuniculus), blaireau (Meles
meles), pigeon (Colombus palumbus). De plus, une espèce d’ongulé a aussi fait partie du
matériel d’expérimentation : le mouton (Ovis aries).
Tous ces taxons ont été choisis tout d’abord pour leur facilité d’accès à la différence du
lagopède, du lièvre ou du mouflon. Puis, chacune peut servir de référentiel à d’autres espèces
de tailles similaires par exemple le mouton pour le bouquetin, le mouflon ou le chamois.
⊗ Parties anatomiques :
Nous avons choisi d’étudier des os longs des membres postérieurs et antérieurs :
Os longs du membre antérieur Os longs du membre postérieur
Humérus
Radius
Ulna
Carpométacarpe (pigeon)
Fémur
Tibia
Tibiotarse (pigeon)
Tarsométatarse (pigeon)
Tableau 2 : Liste des os longs des membres supérieurs et inférieurs utilisés pour l’expérience.
Globalement, c’est le squelette appendiculaire qui porte le plus de traces de découpe ou de
décarnisation (Binford, 1981) ; en outre, les os des membres se conservent relativement bien.

17
⊗ Age
Pour chaque taxon, des os d’individus matures et d’autres un peu plus jeunes sont utilisés sous
les mêmes conditions.
⊗ Le traitement des os avant expérimentation :
Tous les os ont été bouillis partiellement pour éviter de les assécher complètement. Cela pour,
d’une part, une meilleure conservation car avoir tout le matériel frais nécessaire le jour de
l’expérimentation aurait été impossible ; d’autre part, pour éliminer le périoste qui aurait pu
empêcher les artéfacts lithiques de bien marquer les os. Enfin, bouillir partiellement les
ossements ne change pas leur structure externe «what could be considered conventional
boiling time (1-9h) having no or little physico-chemical effects. » (S.J. Roberts et al., 2002).
1.2. Quels produits lithiques ?
Toutes les pièces sont issues de taille expérimentale et ont été fournies par Jean-Guillaume
Bordes.
⊗ Typologie :
Les éléments lithiques utilisés pour l’expérience correspondent, suivant la typologie Bordes,
au matériel produit pendant le Paléolithique supérieur (puisque c’est ici la période sur laquelle
on se base pour notre expérience) : lames, lamelles, éclats, nucléus...
1.3. Densité des os par rapport aux outils lithiques :
D’après le modèle de Glynn Isaac (1971, in Binford, 1984), pour correspondre à un site
d’occupation, la densité des outils par rapport aux ossements doit être très élevée. C’est ce
contexte qui est le plus favorable aux mélanges et à la rencontre des deux types de matériel.
Une forte densité permet d’optimiser l’action du piétinement et donc le mouvement des objets
les uns par rapport aux autres.
1.4. Quel sédiment ?
Les expériences précédentes de piétinement ont démontré que des grains de sables ou des
graviers (c’est-à-dire des grains supérieurs à 0,25 mm) contenus dans du sédiment peuvent

18
laisser des traces similaires à des traces de découpe (cf. partie I 2.2). Le terreau est un
sédiment d’origine végétale et ne contient pas de petits grains minéraux (tels que du sable).
L’expérience réalisée par S. L. Olsen et P. Shipman (1988), a eu pour résultat une absence de
traces sur les ossements piétinés dans une cellule expérimentale contenant uniquement du
terreau. Ainsi, ce sédiment élimine tout risque de produire des traces par piétinement.
1.5. La cellule d’expérimentation :
La forme et la taille de la cellule vont influer sur les déplacements des expérimentateurs. On
a donc deux hypothèses :
- soit une cellule carrée entraînant la possibilité de déplacements au hasard
- soit une cellule beaucoup plus allongée prenant la forme d’un chemin permettant des
déplacements préférentiels suivant l’axe longitudinal de la cellule.
Figure 1 : Schéma des directions de déplacements selon la forme de la cellule.
Comme on l’a déjà dit, ce sont les mouvements induits par le piétinement qui vont mettre en
action outils lithiques et ossements. Pour optimiser cela, il faut donc privilégier une forme qui
fera que les expérimentateurs marchent sur toute la cellule et le plus souvent possible.
Dans la nature, les points de passages privilégiés sont les chemins empruntés régulièrement
ou un point d’eau, pour les hommes une entrée de grotte (Courtin et Villa, 1982). Ainsi en
mettant en place une cellule très allongée, les expérimentateurs se déplaceront suivant un axe
privilégié. Par contre, pour une cellule rectangulaire, ils marcheront sans prendre de trajet
préférentiel. C’est cette deuxième solution qui a été choisie sur le terrain pour maximiser le
piétinement sur l’ensemble de la cellule et s’adapter à la conformation du terrain.

19
La surface du sédiment a été fixée à 2,5 m sur 1,5 m et la cellule d’expérimentation a été
placée dans une cuvette naturelle pour éviter que le terreau et les artéfacts ne s’étalent trop
hors du périmètre fixé.
En outre, la hauteur de la couche de terreau a été définie à environ 15 cm. En effet, la couche
active dans un sédiment comme le terreau est beaucoup plus faible que dans du sable (cf. les
expériences de Olsen et Shipman, 1988, Courtin et Villa, 1982). Celle-ci est inférieure à 2 cm.
Pourtant, bien que la couche minimale de terreau pour occasionner le mouvement des
éléments soit faible, nous avons choisi une hauteur de couche en adéquation avec le volume
de certains ossements, dans le but que ceux-ci puissent y loger et pour isoler tous les objets le
plus possible du sol.
1.6. Qui et comment ?
⊗ Nombre de personnes :
Un nombre restreint de personnes (3 ou 4), vu la superficie de la cellule, semble plus
approprié pour faciliter le déplacement.
⊗ Equipement :
Le matériel consiste en des chaussures à semelles souples. On peut ainsi éliminer le risque
que des chaussures à semelles dures causent plus de dommages et donc entraînent un biais.
⊗ Mode de déplacement :
La manière de se déplacer la plus naturelle est la marche plus ou moins rapide ; c’est donc
celle que nous avons décidé d’utiliser.
2. Expérience
La liste du matériel utilisé lors de l’expérience ainsi que le protocole sont ici donnés en
détail. Certains paramètres théoriques n’ont pas pu être maintenus face à la réalité du terrain.
2.1. Matériel
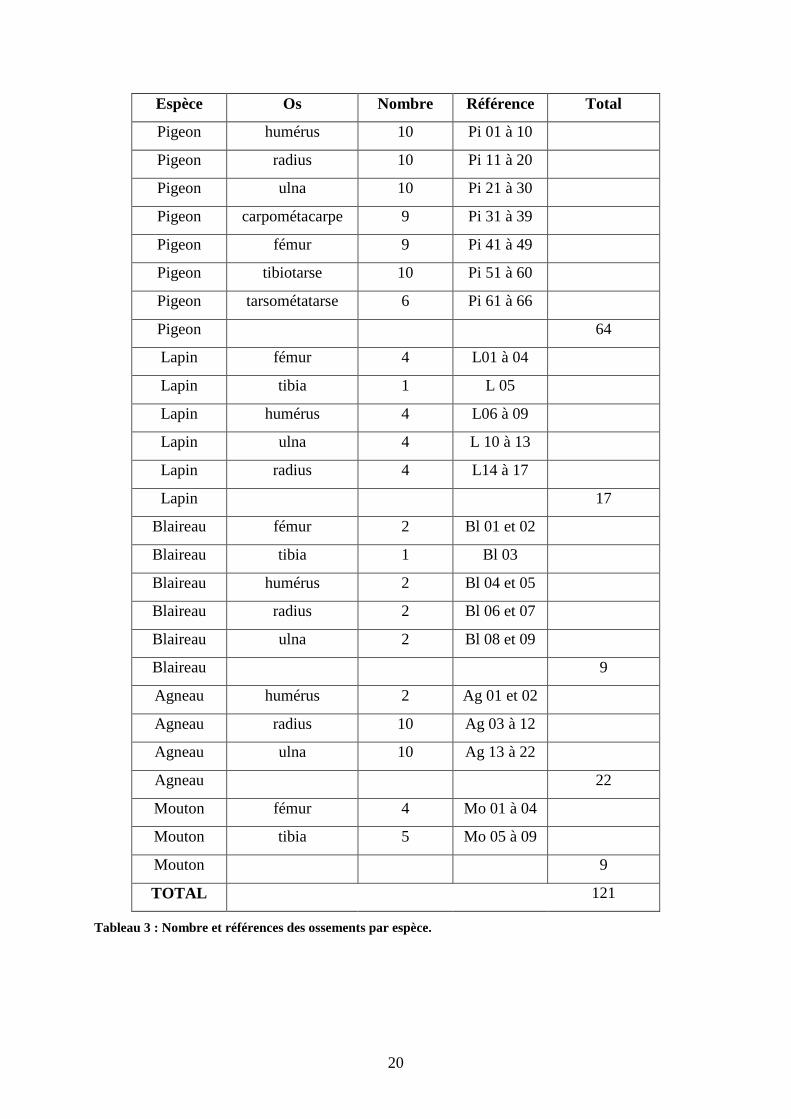
⊗Matériel osseux :
Le poids total du matériel osseux est de 3,1 kg.
20
Espèce Os Nombre Référence Total
Pigeon humérus 10 Pi 01 à 10
Pigeon radius 10 Pi 11 à 20
Pigeon ulna 10 Pi 21 à 30
Pigeon carpométacarpe 9 Pi 31 à 39
Pigeon fémur 9 Pi 41 à 49
Pigeon tibiotarse 10 Pi 51 à 60
Pigeon tarsométatarse 6 Pi 61 à 66
Pigeon 64
Lapin fémur 4 L01 à 04
Lapin tibia 1 L 05
Lapin humérus 4 L06 à 09
Lapin ulna 4 L 10 à 13
Lapin radius 4 L14 à 17
Lapin 17
Blaireau fémur 2 Bl 01 et 02
Blaireau tibia 1 Bl 03
Blaireau humérus 2 Bl 04 et 05
Blaireau radius 2 Bl 06 et 07
Blaireau ulna 2 Bl 08 et 09
Blaireau 9
Agneau humérus 2 Ag 01 et 02
Agneau radius 10 Ag 03 à 12
Agneau ulna 10 Ag 13 à 22
Agneau 22
Mouton fémur 4 Mo 01 à 04
Mouton tibia 5 Mo 05 à 09
Mouton 9
TOTAL 121
Tableau 3 : Nombre et références des ossements par espèce.

21
⊗Matériel lithique :
Au total 1216 pièces lithiques ont été utilisées lors de l’expérience. Des classes de taille ont
été établies suivant la mesure de la longueur maximale de chaque pièce. La majorité est de
petit volume et se compose d’éclats et de débris de taille et de quelques lamelles. Les 2
classes médianes ont une composition très variée : nucléus, lames, éclats,... Enfin, la classe
15-20 cm réunit 3 grandes lames. Le poids total des pièces lithiques est de 12,4 kg.
Figure 2 : Pièces lithiques par classe de taille en centimètre
⊗Autre matériel :
• Bâche 4*5 m pour protéger la cellule du substrat environnant.
• 280 L de terreau soit 158,2 kg
⊗Densité :
La densité du matériel lithique est de 8 g par 100 g de terreau (voir figures 3 et 4).
La densité du matériel osseux est de 2 g par 100 g de terreau (voir figures 3 et 4).

22
2.2. Réalisation de l’expérience : réorientation et biais
⊗Composition du terreau
Une analyse sédimentologique a été menée sur le terreau (voir protocole en annexe 1), sur
44,7 g de sédiment sec. Le poids de la fraction minérale est de 21,07 g, 75,3 % de cette
fraction composé de grains de taille supérieure à 50 µm (cf. annexe 1). Cette partie minérale
du terreau est assez conséquente composée majoritairement de petits galets et de sable. Ces
éléments peuvent causer des traces sur les ossements par piétinement (Behrensmeyer et al.
1986 ; Fiorillo, 1989 ; Olsen et Shipman,1988) et donc entraîner un biais non négligeable.
⊗Réorganisation de la cellule :diminution de la quantité de terreau
Afin de diminuer l’enfoncement des pièces dans le terreau et d’améliorer leurs possibilités de
mouvement, la décision de retirer environ 70 litres de terreau (soit environ 40 kg) a été prise
après 2 heures de piétinement. Le sédiment enlevé a été tamisé à une maille de 5 mm puis les
objets issus du tamisage ont été replacés aléatoirement sur la surface de la cellule. Celle-ci a,
de plus, était réduite à une surface de 2 m sur 1,5 m.
Puis, l’expérience s’est poursuivie encore pendant une heure et quart, en prenant toujours soin
de remuer la cellule régulièrement.
2.3. Protocole en image
L’expérience a duré au total 3h15, on peut noter que des traces de découpe ont été observées
dès la première demi-heure de piétinement.
Figure 3: Part relative de sédiment et d'éléments osseux et lithiques.
Figure 4: Part relative des éléments osseux et lithiques.

23
1. Placement du terreau
Figure 5 : Cellule avant placement des matériels lithiques et osseux
Figure 6: Répartition du terreau
Toute l’équipe d’expérimentateurs a d’abord vidé et réparti le terreau (figure 6), pour former
une couche d’épaisseur uniforme d’environ 10 cm sur une superficie de 2,5 m sur 1,5 m
(figure 5).
2. Placement du matériel osseux puis lithique
Figure 7:Cellule avec les matériels osseux et lithiques.

24
Figure 8 : Répartition des os et des objets lithiques sur la cellule.
Après avoir réparti le terreau de façon uniforme (figures 7 et 8), nous avons d’abord déposé
les ossements afin de bien les répartir sur la surface, puis mis les pièces lithiques en rajoutant
une légère couche de terreau.
3. Piétinement
Figure 9 : 3 expérimentateurs piétinant.

25
Figure 10: Etat de la cellule après 1h30 de marche.
L’équipe d’expérimentateurs se composait de 10 personnes, se relayant de façon à être 3 ou 4
en continu sur la cellule (figure 9). Les mouvements de piétinement se sont échelonnés d’une
marche lente à une marche très rapide, avec quelques petits sauts. Le matériel s’est
rapidement enfoncé et mélangé au terreau (figure 10).
4. Tassement et remaniement du terreau
Figure 11: Mesure de la hauteur de
terreau au centre de la cellule

26
Figure 12 : Remaniement du terreau
Dès le début de l’expérience, le tassement du terreau a été très important (figure 11), de plus
le sédiment a eu tendance à s’accumuler dans le centre de la cuvette. Nous avons donc dû
palier à ces biais en opérant un réétalement régulier (figure 12) toutes les 15 minutes, mais
aussi en remélangeant complètement la cellule pour aérer le terreau et permettre aux matériels
de bouger plus facilement.
5. Tamisage et tri des éléments : tamis de maille 0,5 cm
Figure 13 : Tamisage du terreau Figure 14 : Tri et tamisage du terreau en équipe

27
Au moment du retrait des 40 L de terreau et à la fin de l’expérience, le terreau a été tamisé et
trié de façon à séparer matériels osseux et lithiques (figures 13 et 14).
3. Méthode d’analyse
3.1. Nettoyage et séchage du matériel piétiné
Le matériel a été nettoyé avec beaucoup de précaution pour éviter que des éléments
minéraux contenus dans le substrat puissent faire des traces par frottement sur la surface des
os pendant le lavage. Les éléments ont d’abord été passés sous l’eau afin de retirer la plus
grosse partie du sédiment qui y était accroché. Puis, sous eau courante, l’on a frotté
doucement la surface des os à l’aide d’une éponge. Les parties difficiles et non soumises à
notre observation (intérieur des épiphyses et des diaphyses) ont était nettoyées en partie avec
une brosse à dent en plastique très souple.
Les ossements dont le marquage a disparu au cours de l’expérience ont été marqués à
nouveau à l’encre de Chine après séchage.
3.2. Observation, comptabilisation des traces et réalisation des planches
Le matériel utilisé pour effectuer l’observation des traces consiste seulement en un
macroscope. Le grossissement employé est de x 12,5 à x 25, suivant la taille des traces et des
ossements. Toutes les traces observées 2 ou 3 fois et ont été reportées sur des planches
ostéologiques portant les os vus sous 4 faces et pour le pigeon sous 2 faces. Ces planches ont
été réalisées d’après photo (blaireau et certains os du pigeon) ou d’après dessin : d’après les
planches du Barone (1966) pour le mouton et le lapin, et celles Cohen et Serjeantson (1996)
pour le pigeon (voir annexe 2 planches 1 à 15).
Comme a pu le détailler Lyman (1994), il est important de fixer des critères précis
concernant l’observation des ossements, la définition des traces observées, leur décompte et
leur description (nombre, position, morphologie particulière par exemple). Pour cette étude
nous nous sommes focalisés sur la présence de traces de découpe, leur localisation sur l’os,
leur fréquence. Ainsi nous avons mis en place une grille de lecture de la surface osseuse
permettant de connaître l’état de l’os avec précision.
Les ossements étant tous des os longs, nous avons pu distinguer des zones plus précises, et
non uniquement diaphyse/épiphyse. En effet, comme on a pu le voir dans le premier chapitre,

28
les traces ont des localisations différentes suivant leur causes supposées ; le décompte des
traces s’est donc réalisée en divisant les ossements en 5 zones transversales1 : épiphyse
(proximale et distale), diaphyse proximale, médiale et distale (voir figure 15).
Figure 15 : Division des ossements en 5 zones.
Pour repérer les stries, nous nous sommes basés d’une part sur les critères morphologiques
définis par plusieurs auteurs (Binford, 1981 ; Fisher, 1995 ; Lyman, 1994, 2005), puis d’autres
part sur des collections archéologiques ou des pièces expérimentales. Chaque trace ou groupe
de trace est noté à son emplacement exact sur la face et la zone qui lui correspond sur la
planche anatomique (annexe 2 planches 1 à 15).
Pour noter ces traces, nous avons fait la distinction entre traces individuelles et groupes de
traces (tableau 4), traces profondes et traces superficielles.
Repérer avec précision le nombre exact dont se composent certains ensembles de traces
est très fastidieux et ce détail serait d’ailleurs inutile. En effet, un même mouvement de la
pièce lithique sur l’os, caractérisé par une direction, un sens et une force, peut aussi bien avoir
pour résultat une trace ou bien plusieurs caractérisées par toutes ces composantes. Des stries
groupées peuvent être parallèles et il y a alors de fortes chances qu’elles aient crées en même
temps par un même objet. De plus, la profondeur et la distance entre les stries d’un même
groupe peut aussi fournir des indices sur leur formations.
Nous avons donc décidé de représenter sur les schémas, les traces groupées ou isolées de
la même manière : un trait .
1 Il est impropre d’employer les termes de diaphyse et épiphyse pour les os d’oiseaux. Pour plus de facilité, nous utiliserons le terme épiphyse pour désigner l’extrémité et diaphyse pour la partie allongée des os de pigeon.

29
Groupe de
traces
Ensemble de traces produites par le
même «mouvement» et donc de même
direction, sens et profondeur similaire
Trace isolée
Trace de découpe issu d’un seul
passage en un endroit précis de l’outil
sur la surface de l’os
Tableau 4 : Définition et schéma d’une trace isolée et d’un groupe de trace.
D’autre part, l’examen de la surface osseuse a permis de mettre en évidence une
différence réelle concernant la profondeur des traces. Il apparaît donc, sur les schémas, les
traces superficielles et les traces non superficielles, ou «profondes». Pour certains auteurs, le
degré de profondeur des traces est un indice permettant d’identifier leur origine. Les pseudo-
traces de découpe issues du piétinement seraient en majorité superficielles (Olsen et Shipman,
1988).
Les planches anatomiques forment donc une représentation et une interprétation mettant
en évidence les marques créées par le piétinement ainsi que leurs caractéristiques propres :
nombre, longueur, orientation, profondeur et emplacement. Enfin, des photographies
numériques des stries et autres traces ont été réalisé en complément.
Enfin, il est important de souligner que tous les os, même fracturés au cours de l’expérience,
ont pu être remontés entièrement pour l’analyse macroscopique.
3.3. Création de la base de données
L’étude des séries osseuses issues de l’expérimentation a nécessité la création d’une base de
données. Pour la créer, nous avons utilisé le logiciel Excel de Microsoft ; elle prend donc la
forme d’un tableau Excel assez simple contenant les informations quantitatives et qualitatives
qui reprennent et viennent compléter les données fournies par les planches anatomiques.
Outre les informations propres à la partie du squelette, l’espèce et aux marques comme citées
à la fin du paragraphe précédent (emplacement, orientation, profondeur, etc.), la base de

30
données a permis d’ajouter des données relatives aux autres altérations subies par les
ossements. Ainsi, le piétinement a causé, par exemple, des fracturations ou des fissurations.
Ces altérations ont été simplement enregistrées suivant leur présence et leur qualité. En effet,
ce ne sont pas des critères sur lesquels nous nous focalisons mais qui donnent des indices sur
la nature de la force et des mouvements créés par l’action du piétinement. Nous verrons plus
loin les applications issues de la base de données consistant surtout en des statistiques très
simples.
31
III. RESULTATS
Il est important de souligner que la quantité disponible d’ossements n’a pas été équivalente
pour chacune des espèces (voir 1.1. de ce chapitre). Ainsi, on ne peut tirer que des résultats
préliminaires de l’expérience sur les différences inter et intra spécifiques, sur la morphologie
des stries et leur répartition.
1. Représentativité des espèces et des parties anatomiques
On peut tout d’abord mettre en évidence que l’effet du piétinement touchent de manière
variable les espèces mais on peut aussi relever des différences entre les portions squelettiques
elles-mêmes.
1.1. Une espèce présente-t-elle plus de stries ?
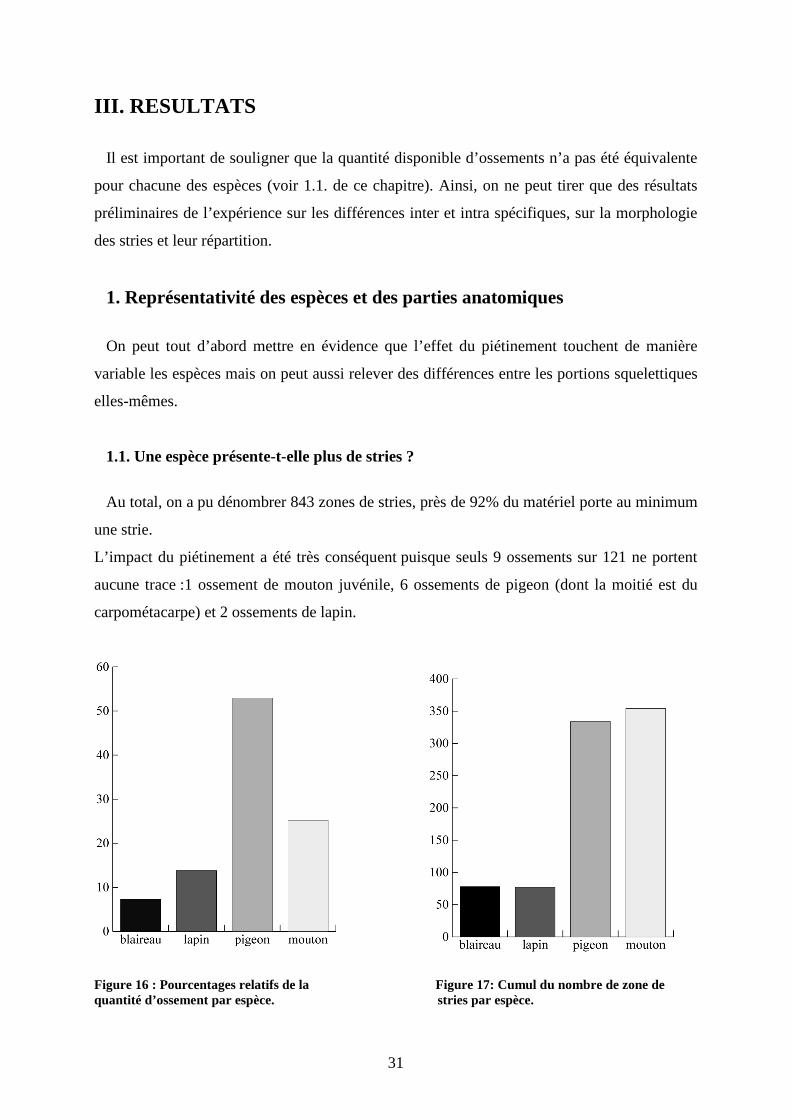
Au total, on a pu dénombrer 843 zones de stries, près de 92% du matériel porte au minimum
une strie.
L’impact du piétinement a été très conséquent puisque seuls 9 ossements sur 121 ne portent
aucune trace :1 ossement de mouton juvénile, 6 ossements de pigeon (dont la moitié est du
carpométacarpe) et 2 ossements de lapin.
Figure 16 : Pourcentages relatifs de la Figure 17: Cumul du nombre de zone de quantité d’ossement par espèce. stries par espèce.

32
On peut constater deux classes dont le nombre de stries est du même ordre (figure 17). Les
résultats pour le blaireau et le lapin sont similaires 78 stries pour le premier, 77 pour le
second. Pourtant, l’effectif du lapin est deux fois plus important que celui du blaireau. Bien
que l’impact du piétinement ait été fort pour ces deux espèces, le blaireau a subi plus d’effet
du piétinement : les os de blaireau portent jusqu’à 22 zones de stries. Cette différence peut
être due à la différence de taille, le blaireau étant plus grand que le lapin ou à la surface des os
de lapin. Les ossements de lapin sont issus d’individus juvéniles, la surface de ces ossements
est plus poreuse que celle d’individus adultes ce qui rend plus difficile la lecture des os et la
mise en évidence de stries.
Le mouton et le pigeon sont les espèces portant le plus de stries. En effet, le pigeon présente
un total de 354 stries pour 64 ossements et le mouton 334 stries pour 30 ossements.
Ainsi, il ne faut pas négliger la différence de volume et donc de surface entre ces deux
espèces (Lyman, 2005). La différence de gabarit va entraîner une différence concernant les
possibilités de mouvements et l’importance de la surface osseuse, et donc plus ou moins de
chance d’entrer en contact avec les objets lithiques (Fiorillo, 1989).
1.2. Y’a-t-il plus de stries sur un ossement ?
La morphologie propre de chaque type d’ossement peut être un paramètre jouant sur
l’impact du piétinement. Ainsi est-il possible que certains os aient été plus touchés que
d’autres ?
Comme on va le voir ci-après, l’impact a été différent non seulement d’une portion
squelettique à l’autre mais aussi entre les os au sein d’une même espèce. Chaque ossement
présente un schéma de répartition de stries unique (voir annexe 2), pourtant on peut remarquer
des similarités pour certaines portions squelettiques d’une même espèce.
Tout d’abord, certaines portions squelettiques sont plus représentées que d’autres quand on
prend les effectifs globaux.
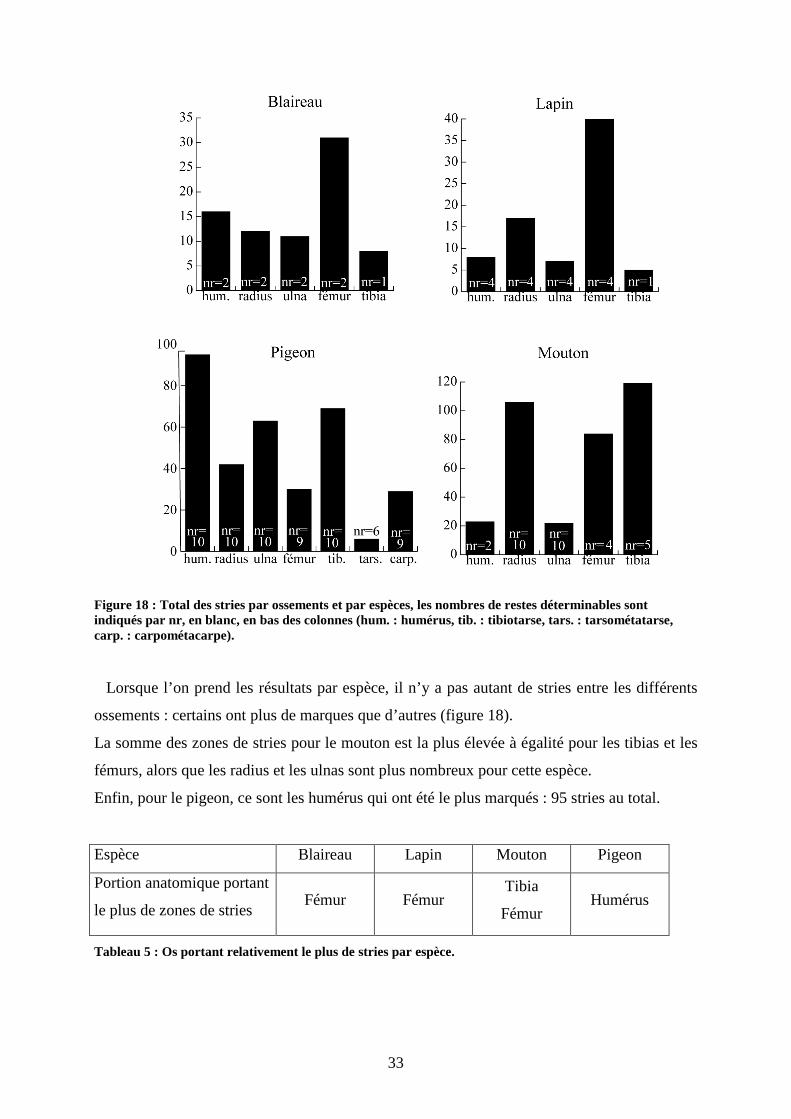
33
Figure 18 : Total des stries par ossements et par espèces, les nombres de restes déterminables sont indiqués par nr, en blanc, en bas des colonnes (hum. : humérus, tib. : tibiotarse, tars. : tarsométatarse, carp. : carpométacarpe).
Lorsque l’on prend les résultats par espèce, il n’y a pas autant de stries entre les différents
ossements : certains ont plus de marques que d’autres (figure 18).
La somme des zones de stries pour le mouton est la plus élevée à égalité pour les tibias et les
fémurs, alors que les radius et les ulnas sont plus nombreux pour cette espèce.
Enfin, pour le pigeon, ce sont les humérus qui ont été le plus marqués : 95 stries au total.
Tableau 5 : Os portant relativement le plus de stries par espèce.
Espèce Blaireau Lapin Mouton Pigeon
Portion anatomique portant
le plus de zones de stries Fémur Fémur
Tibia
Fémur Humérus

34
On peut remarquer la différence entre le pigeon et les autres espèces (Tableau 5). Chez le
pigeon, l’humérus est l’os le plus volumineux, le moins allongé et il présente des surfaces
planes. Etant donné que l’effectif osseux du pigeon est plus important que pour les autres
espèces et que chaque portion anatomique est représentée à part égale, il est plus intéressant
de faire une comparaison portion par portion.
.
Figure 19 : Deux humérus de pigeon,
l’un (à gauche) portant de nombreuses
stries, l’autre (à droite) très peu
2. Schéma de répartition
Bien que le contact entre l’os et l’objet lithique mis en mouvement par le piétinement relève
du hasard, l’impact et la création des marques sont influencés en partie par la morphologie de
l’os (Fiorillo, 1989). Nous allons essayer de dresser une carte de la répartition des zones de
stries en fonction des segments de l’os, de sa morphologie propre (ses reliefs) et du
regroupement des stries. Ceci nous amènera à voir si certaines zones de l’os subissent plus de
contacts avec le matériel lithique.
2.1. Répartition en fonction des segments osseux
De part leur morphologie, les différentes portions d’un os vont subir différemment les
impacts des objets lithiques. Tout d’abord, les extrémités osseuses ont plus de reliefs que les
diaphyses. Puis les portions proximale, médiale et distale de la diaphyse peuvent présenter des
crêtes osseuses, des surfaces plus plates ou plus bombées.
Lorsque l’on regarde la totalité des zones de stries pour toutes les espèces confondues, on peut
constater que la portion osseuse portant le plus de stries est la diaphyse. Les ossements de
pigeon, de mouton et de lapin présentent plus de stries sur la portion médiale de la diaphyse
Bien que l’on puisse constater un nombre important
de stries sur les humérus; seulement, 3 sur 10 sont
massivement marqués avec plus de 20 zones de
stries, les autres ont moins de 10 stries. La
répartition des stries sur des os similaires est donc
très hétérogène comme le montre la figure 19.
L’humérus de gauche porte de multiples zones de
stries tandis que celui de droite presque pas.
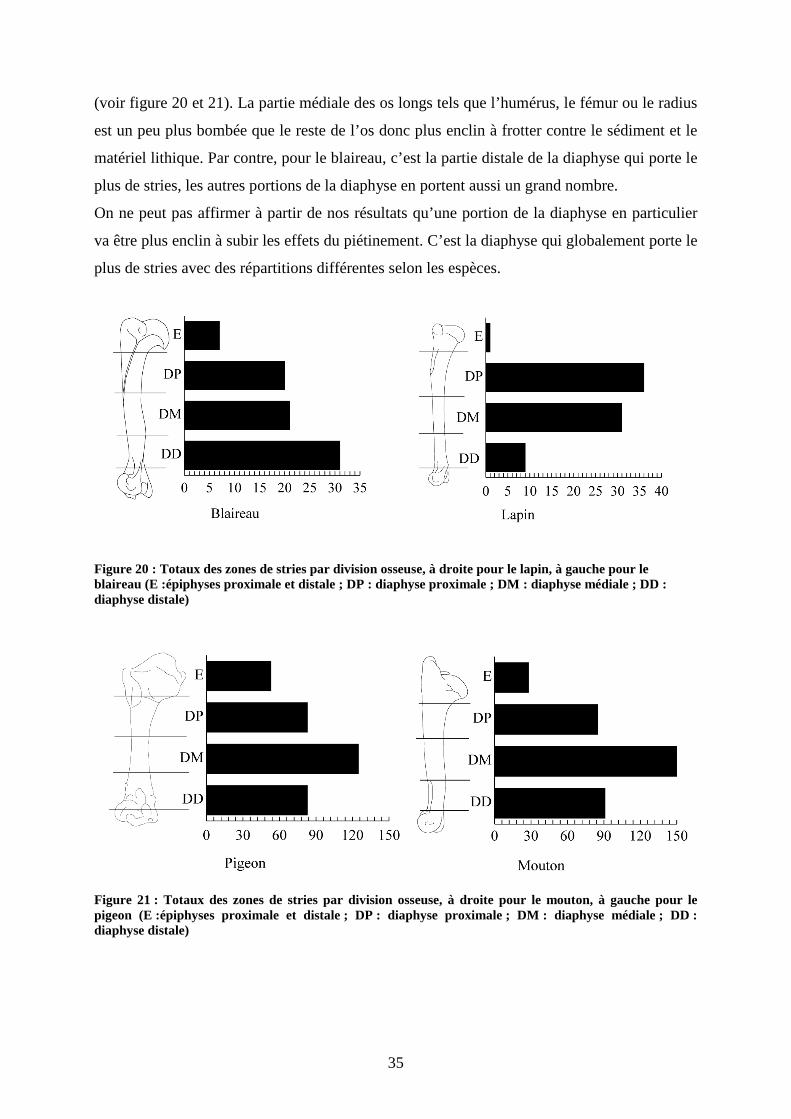
35
(voir figure 20 et 21). La partie médiale des os longs tels que l’humérus, le fémur ou le radius
est un peu plus bombée que le reste de l’os donc plus enclin à frotter contre le sédiment et le
matériel lithique. Par contre, pour le blaireau, c’est la partie distale de la diaphyse qui porte le
plus de stries, les autres portions de la diaphyse en portent aussi un grand nombre.
On ne peut pas affirmer à partir de nos résultats qu’une portion de la diaphyse en particulier
va être plus enclin à subir les effets du piétinement. C’est la diaphyse qui globalement porte le
plus de stries avec des répartitions différentes selon les espèces.
Figure 20 : Totaux des zones de stries par division osseuse, à droite pour le lapin, à gauche pour le blaireau (E :épiphyses proximale et distale ; DP : diaphyse proximale ; DM : diaphyse médiale ; DD : diaphyse distale)
Figure 21 : Totaux des zones de stries par division osseuse, à droite pour le mouton, à gauche pour le pigeon (E :épiphyses proximale et distale ; DP : diaphyse proximale ; DM : diaphyse médiale ; DD : diaphyse distale)

36
2.2. Rôle du relief osseux ?
On vient de le voir, les ossements ne sont pas marqués uniformément, les diaphyses portent
plus de marques mais elles ont des reliefs plus ou moins marqués qui ne vont pas subir les
impacts à parts égales. Ainsi, on pourrait penser que les crêtes osseuses se trouvant sur les
diaphyses auraient plus de stries pourtant ce n’est pas ce que l’on voit. Les crêtes, si elles sont
très développées comme la crête tibiale, n’ont pas plus de marques qu’une autre zone osseuse
voire pas de marque du tout. Ceci est vrai aussi pour des reliefs osseux moins marqués comme
ceux se trouvant sur la partie distale antérieure du fémur chez le lapin, le blaireau ou le
mouton. Ainsi en général pour ces trois espèces, on ne peut pas définir un rôle particulier des
reliefs osseux ; c’est la diaphyse qui porte le plus de traces sans schéma particulier. Le
nombre restreint d’ossements ne permet pas d’aller plus loin.
Par contre, sur certains ossements du pigeon portant beaucoup de stries comme l’humérus,
la superposition des schémas de tous les humérus permet de déterminer des zones
préférentiellement touchées.
Sur ce schéma (figure 22), on voit apparaître 4
zones plus marquées :
- une zone à relief abrupt : 2
- des zones plutôt planes :1 et 3
- une zone bombée : 4
Trois de ces zones se trouvent sur la diaphyse et
n’ont pas une forte convexité ; une seule zone, la
zone 2, est sur l’extrémité de l’os. Le cas de zones
osseuses préférentiellement touchées au niveau
des parties allongées ne se retrouve pas
uniquement, en ce qui concerne le pigeon, sur
l’humérus mais aussi sur la partie distale du
tibiotarse (voir annexe 2, planche 10).
Hormis ces exemples, le reste des ossements de pigeon ne montrent pas de répartition de
stries particulière en fonction des reliefs anatomiques.
Figure 22 : Diagramme cumulé des 10 humérus de pigeon, côté droits et gauche confondus avec numérotation des zones présentant le plus de marques

37
2.3. Regroupement
Le nombre de stries isolées (429) est quasiment égal à celui des stries groupées (423). Le
regroupement des stries apportent de nombreuses informations sur le contact entre os et objets
lithiques. En effet, les caractéristiques des groupements de stries renseignent d’une part sur la
simultanéité et l’origine de leur création : ont-elles été créées par un seul et même impact avec
un objet lithique ? D’autre part, elles informent aussi sur la surface de l’objet en contact avec
l’os (étendue du groupe de stries) et sur l’intensité de l’impact (profondeur).
Sur la photo de la figure 23, la surface a été raclée créant des traces parallèles, courtes, plus
profondes dans la partie supérieure. Ce groupement de stries a donc peut-être été créé par le
même mouvement d’un objet sur la surface osseuse. Par contre, sur la figure 24, on peut voir
des stries de morphologies très diverses. Ce type de schéma a pu se produire au cours du
frottement prolongé par un ou plusieurs objets lithiques contre une même zone de l’os avec
des mouvements dans toutes les directions.
Etendue et force de l’impact
Une zone de stries peut contenir une ou plusieurs stries reflétant une différence dans la
surface de l’objet étant rentré en contact avec la surface de l’os. Un strie unique a plutôt été
créée par un relief plus saillant d’un objet lithique ; un groupe de strie avec les mêmes
caractéristiques que celui de la Figure 23 est engendré par un contact avec un surface saillante
Figure 23 : Groupement de stries sur un ulna de blaireau (référence Bl7, annexe 2 planche 3)
Figure 24 : Zone avec stries diverses sur un humérus de pigeon (référence Pi 7, annexe 2 planche 6)

38
plus large. Enfin, la profondeur des stries donne un indice sur la force de l’impact de la pièce
lithique sur l’os (voir ci-après sous partie 3.1.).
3. Morphologie
3.1. Profondeur
Ce critère, selon Olsen et Shipman (1988), est important pour définir des traces créées par
piétinement ; selon ces auteurs, les stries de piétinement seraient plus superficielles que des
stries de découpe. La profondeur d’une strie donne une idée sur la puissance de l’impact qui
l’a créé. Bien que définir la profondeur d’une strie soit subjectif, on a ici essayé de bien
différencier une strie superficielle (figure 26), d’une strie plus profonde (figure 25). Une strie
profonde entaille largement la surface osseuse, tandis qu’une strie superficielle marque très
peu la surface créant un sillon assez fin.
Figure 26 : Traces superficielles sur un radius de mouton (référence A 9, annexe 2, planche 14).
Figure 25 : Trace profonde sur un fémur de lapin (référence L1, annexe 2, planche 4).

39
Globalement, le nombre de stries profondes est de 201 sur 843 ossements, dont 28 localisées
sur les épiphyses. Les stries profondes sont donc plus nombreuses sur les diaphyses (peut-être
à cause d’une plus grande superficie) mais minoritaires sur la totalité de l’effectif
expérimental. C’est sur les ossements de mouton que les traces profondes sont le plus
nombreuses (110 au total). Enfin, d’une manière générale, les stries profondes n’ont pas de
répartition ou de direction particulières. Elles peuvent néanmoins faire partie d’un groupe de
stries (figure 27).
3.2. Orientation
Trois orientations par rapport à l’axe d’allongement de l’os ont été déterminées :
l’horizontale, l’oblique et la verticale. Dans le graphique ci-dessous, la totalité des zones de
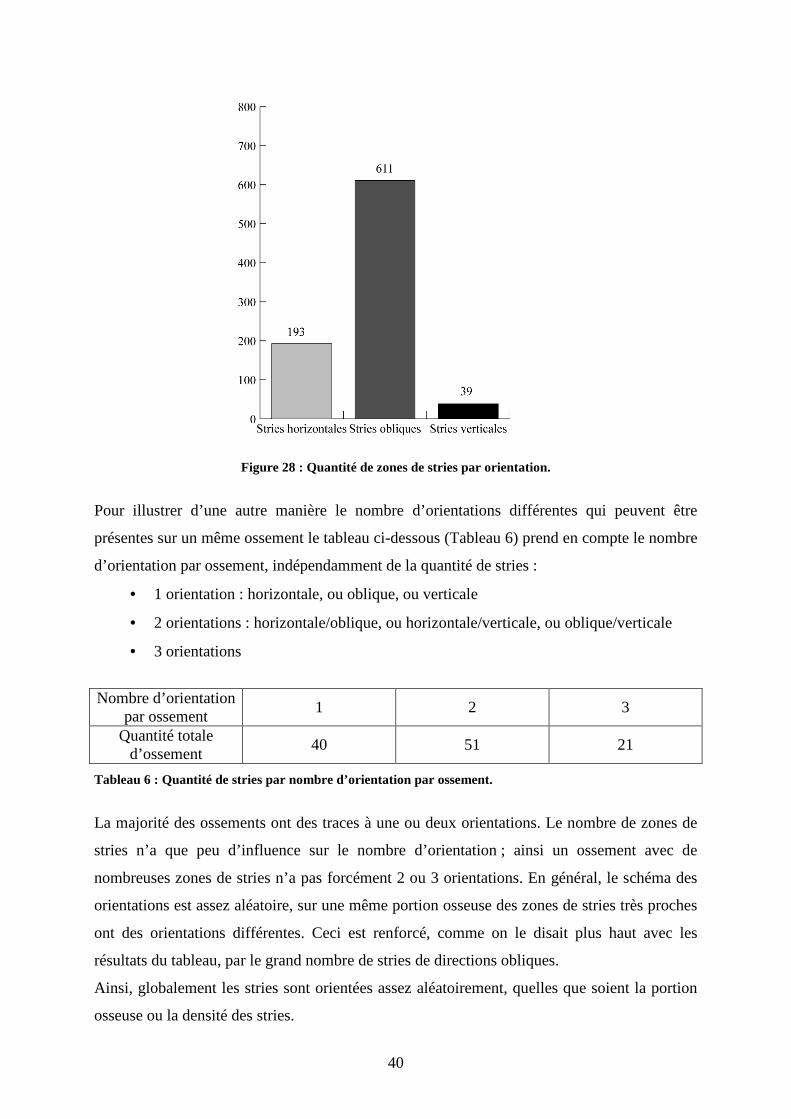
stries a été décomptée suivant ces trois orientations. On constate que sur les 843 zones de
stries totales plus de 600 sont obliques (figure 28). Les stries verticales sont très minoritaires,
seulement 39 ont été comptabilisées. L’obliquité des stries peut avoir des directions et des
angles très différents. L’hétérogénéité dans la direction des stries reflète le caractère aléatoire
des mouvements lors des contacts entre ossements et matériel lithiques.
Figure 27 : Groupement de traces sur un ulna de blaireau (référence Bl 6, annexe 2, planche 3).
40
Figure 28 : Quantité de zones de stries par orientation.
Pour illustrer d’une autre manière le nombre d’orientations différentes qui peuvent être
présentes sur un même ossement le tableau ci-dessous (Tableau 6) prend en compte le nombre
d’orientation par ossement, indépendamment de la quantité de stries :
• 1 orientation : horizontale, ou oblique, ou verticale
• 2 orientations : horizontale/oblique, ou horizontale/verticale, ou oblique/verticale
• 3 orientations
Tableau 6 : Quantité de stries par nombre d’orientation par ossement.
La majorité des ossements ont des traces à une ou deux orientations. Le nombre de zones de
stries n’a que peu d’influence sur le nombre d’orientation ; ainsi un ossement avec de
nombreuses zones de stries n’a pas forcément 2 ou 3 orientations. En général, le schéma des
orientations est assez aléatoire, sur une même portion osseuse des zones de stries très proches
ont des orientations différentes. Ceci est renforcé, comme on le disait plus haut avec les
résultats du tableau, par le grand nombre de stries de directions obliques.
Ainsi, globalement les stries sont orientées assez aléatoirement, quelles que soient la portion
osseuse ou la densité des stries.
Nombre d’orientation par ossement
1 2 3
Quantité totale d’ossement
40 51 21

41
3.3. Longueur
Sur l’effectif total des traces, la proportion de traces allongées c’est-à-dire de longueur
supérieure à 1cm, est très réduite : 17, toutes espèces confondues. Ainsi la majorité des traces
sont courtes, résultant d’un contact bref entre surface osseuse et objet lithique. De plus, les
traces allongées n’ont pas d’orientation particulière, elles sont horizontales, longitudinales ou
souvent oblique à l’axe d’allongement de l’os.
3.4. Particularités morphologiques
Ce sont des figures rencontrées de manière ponctuelle dans le corpus des ossements. Ces
stries sont particulières car leur morphologie reflète un impact particulier ou une particularité
du relief de la pièce lithique.
Figures révélant un impact particulier
La strie en forme de vague suit les
reliefs de la surface de l’os (figure 29).
Figure 29 : Strie en forme de vague sur
un radius de pigeon (Pi 16, annexe 2,
planche 7).
Le raclage provoqué par un contact
fort et prolongé entre l’os et la pièce
lithique (figure 30).
Figure 30 :Raclage sur un radius de lapin (L15, annexe 2 planche 5).

42
Figures révélant un ou des reliefs de la pièce lithique
L’effet d’épaulement : on peut voir
une strie plus fine en parallèle de la
strie principale créée par une
irrégularité de l’angle de l’objet
lithique entré en contact avec l’os
(figure 31).
Figure 31 : Strie avec effet d’épaulement
sur un tibiotarse de pigeon (Pi 60, annexe
2, planche 10)
Cette strie est caractérisée par
de nombreuses stries
superficielles parallèles (figure
32).
Figure 32 : Strie avec multiples stries parallèles sur un tibia de blaireau (Bl 3, annexe 2, planche 2).
4. Tableau récapitulatif des résultats
Le tableau ci-après présente les principaux résultats par espèces, ainsi que les principales
particularités morphologiques. La partie distribution particulière concerne une répartition non
aléatoire des stries sur la surface osseuse qui peut être par exemple une redondance sur
certaines zones d’une portion squelettique.

43
Espèce
Total
zones
de
stries
Portion
squelettique
avec le plus
de stries
Portion
osseuse
avec le
plus de
stries
Orientation
majoritaire
Longueur
des stries
Profondeur
majoritaire
des stries
Distribution
particulière
Blaireau 78 Fémur Diaphyse Oblique Courte Superficielle Non
Lapin 77 Fémur Diaphyse Oblique Courte Superficielle Non
Mouton 354 Fémur
Tibia Diaphyse Oblique Courte Superficielle Non
Pigeon 334 Humérus Diaphyse Oblique Courte Superficielle Oui pour 2
types d’os
Tableau 7 : Récapitulatif des résultats.
5. Autres effets du piétinement
Nous avons décompté les autres modifications qu’ont subi les ossements lors de
l’expérience. Ainsi, 41 os ont été fracturés, 9 montrent des fissurations et 3, un enfoncement.
On a aussi pu relevé des altérations de surfaces telles que des traces d’abrasion et des
comètes (25), et des traces de desquamation : 10.
Figure 33 : Enfoncement (tâche grise sur le schéma) sur un humérus de pigeon (Pi 2, annexe 2, planche 6).

44
IV. DISCUSSION : APPORTS ET BIAIS
Les résultats que nous présentons sont conséquents (beaucoup de stries) mais il reste à
mettre en avant des caractéristiques propres aux stries de piétinement pour les différencier par
rapport à un schéma de découpe. Il est important de mettre en place des critères de
reconnaissance clairs et globaux bien qu’au niveau isolé, les traces soient très similaires à des
traces de découpe. Enfin, les paramètres de notre expérience sont très précis et l’application
des résultats à l’archéologie est donc restreinte.
1. Implications archéozoologiques
L’objectif de notre expérience est de formuler des critères de reconnaissance pour les stries
de piétinement, nos résultats permettent de mettre en avant un schéma de répartition et une
morphologie particulière des stries.
1.1. Similarité entre espèces : schéma de répartition correspondant au piétinement
Aucune expérience n’ayant été réalisée sur des ossements de petite faune comme du lapin,
du blaireau ou du pigeon, on ne peut comparer nos résultats qu’avec des critères issus
d’études expérimentales réalisées sur de la grande faune : cheval, vache et cochon (Andrews
and Cook, 1985 ; Behrensmeyer et al., 1986 ; Fiorillo, 1984,1989 ; Olsen et Shipman, 1988).
Le nombre de stries que portent les ossements après l’expérience est très élevé (843), 112
des 121 ossements portent au moins une strie. L’impact du piétinement a été très important.
Pourtant, il faut relativiser ces résultats (voir partie 2. de ce chapitre) : la durée du piétinement
a été longue et la densité de matériel lithique a été très importante par rapport à celle du
matériel osseux.
Les observations que nous avons pu réaliser sur les différentes espèces nous mènent aux
même conclusions pour les petits et les grands gabarits Les traces diffèrent en général des
critères de E. D. Guilday et al. (1962) définissant un schéma de découpe par la récurrence de
la distribution des stries et leur placement en fonction des zones anatomiques (tendons,
muscles,...). Le mouton est la seule espèce de notre corpus à être proche des espèces utilisées
pour les expériences précédentes. Il rentre très bien dans les schémas définis par les auteurs
les ayant réalisées : orientation des stries aléatoires, diaphyses préférentiellement touchées,
pas de distribution en fonction de l’anatomie, stries courtes en grande majorité. En ce qui
concerne la morphologie même des stries, les résultats se rapprochent de ceux de J. S. Oliver
45
(1989), A. R. Fiorillo (1989) et A. K. Behrensmeyer et al. (1986) car elles sont tout à fait
similaires à des traces de découpe.
En ce qui concerne le lapin et le blaireau, on retrouve aussi ce type de schéma qui met en
avant le caractère aléatoire de l’occurrence des stries.
Le pigeon présente, comme le mouton, un nombre important de stries et globalement un
schéma présentant les caractéristiques énoncées ci-dessus.
1.2. Différences entre espèces
Il est possible de distinguer des différences sur l’effet du piétinement entre les espèces et
entre certaines portions squelettiques.
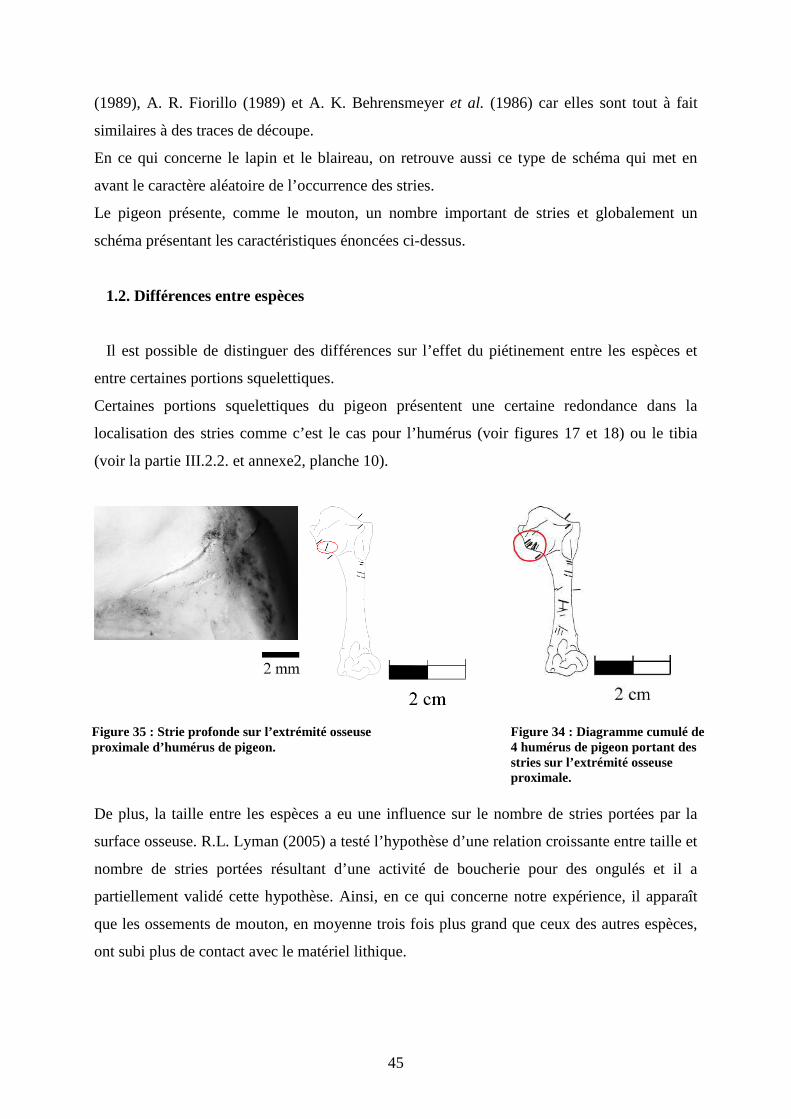
Certaines portions squelettiques du pigeon présentent une certaine redondance dans la
localisation des stries comme c’est le cas pour l’humérus (voir figures 17 et 18) ou le tibia
(voir la partie III.2.2. et annexe2, planche 10).
De plus, la taille entre les espèces a eu une influence sur le nombre de stries portées par la
surface osseuse. R.L. Lyman (2005) a testé l’hypothèse d’une relation croissante entre taille et
nombre de stries portées résultant d’une activité de boucherie pour des ongulés et il a
partiellement validé cette hypothèse. Ainsi, en ce qui concerne notre expérience, il apparaît
que les ossements de mouton, en moyenne trois fois plus grand que ceux des autres espèces,
ont subi plus de contact avec le matériel lithique.
Figure 35 : Strie profonde sur l’extrémité osseuse proximale d’humérus de pigeon.
Figure 34 : Diagramme cumulé de 4 humérus de pigeon portant des stries sur l’extrémité osseuse proximale.

46
1.3. Modèle de reconnaissance pour l’archéozoologie
Certaines stries de piétinement, prises isolément peuvent être confondues avec des traces de
découpe, de part leur morphologie (groupement, profondeur, effet d’épaulement ou autre) ou
leur emplacement (sur une surface articulaire par exemple). Elles peuvent être interprétées,
suivant les critères de L. R. Binford (1981), comme résultant d’une activité de boucherie
spécifique par rapport à leurs caractéristiques.
J. S. Oliver (1989) a mis en évidence un critère important : la morphologie aléatoire des stries
créées par du piétinement. En effet, sur un même ossement ou sur un même type de portion
squelettique la profondeur, l’orientation, la longueur et en général la morphologie des stries
sont très diverses ; ceci vient renforcer les autres critères cités précédemment et repris ci-
dessous.
Critères de reconnaissance de stries causées par le contact d’objets lithiques avec des
ossements sous l’effet du piétinement (Fiorillo, 1989 ; Oliver, 1989 ; Olsen et Shipman,
1988)
• Schéma de répartition des stries aléatoire
• Diaphyses, ou parties allongées, préférentiellement touchées
• Orientation aléatoire des stries
• Stries majoritairement courtes
• Morphologie de stries très diverse
• Stries majoritairement superficielles
2. Biais et restrictions à l’application archéologique
Les résultats que nous mettons en évidence sont le reflet des critères que nous avons choisis
et mis en place. Cela veut dire d’une part, qu’au niveau archéologique ils ne peuvent être
appliqués qu’à un site se trouvant dans le même contexte d’accumulation et dans un substrat
de même type ; d’autre part, certains paramètres de l’expérience ont pu créer des biais.
2.1. Le terreau
Ce sédiment est très meuble, au cours de l’expérience les pièces s’enfonçaient beaucoup et
le mouvement des os comme celui du matériel lithique était très diminué voire nul. Les

47
expérimentateurs ont du augmenter artificiellement le mouvement des objets par une marche
plus appuyée et un remaniement de la cellule (voir II. 2.3.).
En outre, l’analyse du terreau a révélé la présence de sable et de gravier (voir annexe 1). Ces
éléments lithiques peuvent causer des traces très similaires à des traces des découpes
(Behrensmeyer, 1986 ; Fiorillo, 1989). Il est donc impossible de distinguer les stries dues au
sable, de celle dues aux objets lithiques taillés. Ceci s’intègre dans notre problématique
puisque notre but est de voir les « pseudotraces » causées par le piétinement.
2.2. L’intensité
L’intensité du piétinement a été très forte : 3 personnes qui marchent d’un pas appuyé sur
moins de 4 m carrés pendant 3h15. Puis, la quantité de matériel lithique est de 12,4 kg pour
3,1 kg d’ossements seulement. Tous ces paramètres ont d’abord contribué à la quantité
importante de traces produites lors de l’expérience, mais ils doivent aussi être pris comme des
critères limitant la comparaison des résultats de l’expérience seulement à des cas de sites
archéologiques de même ordre.
2.3. Le corpus expérimental
Le matériel lithique a pu nous être fourni en grande quantité, par contre, la quantité de
matériel osseux est moindre. Bien que les ossements de pigeon soient nombreux, la quantité
globale d’os est trop réduite pour tirer des résultats généralisables à chaque espèces.
L’expérience pourrait être complétée en utilisant plus d’ossements de lapin et de blaireau.

48
CONCLUSION
Notre expérience est inédite, c’est la première expérience testant les effets du piétinement
sur ossements de petite faune mélangés à des éléments lithiques. L’effet du piétinement a été
très intense. Il a entraîné la création en grand nombre de traces identiques à des traces de
découpe causées par le contact d’objets lithiques (éclats, lames, nucléus...) contre la surface
des ossements.
Cette ressemblance est importante à première vue. Pourtant une observation plus poussée du
corpus permet de déterminer un schéma de répartition et des caractéristiques spécifiques
quelle que soit l’espèce : profondeur superficielle, orientation plutôt oblique, localisation sur
la diaphyse. Pour les quatre espèces étudiées, on retrouve ce même schéma aléatoire qui est
identique au schéma de piétinement décrit par les expériences de piétinement réalisées
précédemment (voir partie I).
On peut souligner la différence entre la taille d’un ossement et le nombre des stries portées
qu’il porte : un ossement plus volumineux va porter plus de stries.
Le corpus expérimental est encore trop petit pour tirer des généralités. Il serait utile de réaliser
le même type d’expérience mais avec plus d’ossements ou d’établir la fréquence des stries en
fonction de la surface des ossement. En outre, on pourrait aussi réaliser en parallèle un corpus
d’ossements issus de boucherie expérimentale pour tester la corrélation entre les deux
schémas piétinement et découpe, puis d’utiliser un plus fort grossissement pour déterminer
des critères de distinctions entre stries faites par piétinement et stries de découpe.

49
BIBLIOGRAPHIE
ABE Y.; MAREAN C. W.; NILSSEN P. J.; ASSEFA .Z. ; STONE E. C. 2002 - The analysis
of cutmarks on archaeofauna: A review and critique of quantification procedures, and a new
image-analysis gis approach. American antiquity, 67 (4), p. 643-663
ANDREWS P. ; COOK J. 1985 – Natural modifications to bones in a temperate setting. Man,
4, p. 675-691, 2 tabl., 8 fig.
BARONE R. 1966 – Anatomie comparée des mammifères domestiques, Tome I : Ostéologie.
Lyon : Laboratoire d’anatomie et Ecole vétérinaire de Lyon
BEHRENSMEYER A. K.; GORDON K. D.; YANAGI G. T. 1986 - Trampling as a cause of
bone surface damage and pseudo-cutmarks. Nature, 319, p. 768-771, 4 fig.
BINFORD L. R. 1981 – Bones, ancient men and modern myths. London : Academic press,
320 p. ill.
BINFORD L. R. 1984 – Faunal remains from Klasies River Mouth. Orlando : Academic
press
BROMAGE T. G. ; BOYDE A. 1984 – Microscopic criteria for the determination of
directionality of cutmarks on bone. American Journal of Physical Anthropology, 65, p. 359-
366, fig. 9
BUNN H. T. 1989 – Diagnosing Plio-Pleistocene hominid activity with bone fracture
evidence. In : R. Bonnichsen et M. H. Sorg (Ed.), Bone modification. Orono : Center for the
study of first Americans, p. 299-315
COHEN A. ; SERJEANTSON D. 1996 – A manual for the identification of birds from
archaeological sites (revised edition). London : Archetype Edition
COSTAMAGNO S. 1999 – Stratégie de chasse et fonction des sites au Magdalénien dans le
Sud de la France. Bordeaux : Université de Bordeaux I, 1999. 2 t., 495 p., 329 tabl., 132
fig., 128 pl. et ill. Thèse N.D. : Sc. Bordeaux I
COURTIN J. ; VILLA P. 1982 - Une expérience de piétinement. Bulletin de la Société
Préhistorique Française. 79 (4), p. 117-123, 3 fig.
DENYS C. 2002 – Taphonomy and experimentation. Archaeometry, 44 (3), p. 469-484, 3 fig.
DOMINGUEZ-RODRIGO M. 1997 – Testing meat-eating in early hominids : an analysis of
butchery marks on defleshed carcasses. Human Evolution, 12 (3), p. 169-182, tabl., fig.

50
EGELAND C. P. 2003 - Carcass processing intensity and cutmark creation: An experimental
approach. Plains Anthropologist, 48 (184), p. 39-51, tabl., fig.
FIORE I. ; PINO URIA B. ; TAGLIACOZZO A. 2004 – L’exploitation des petits animaux au
Paléolithique supérieur-Mésolithique en Italie : l’exemple de la Grotta del Santuario della
Madonna de Praia a Mare. In : APDCA (Ed.), Petits animaux et Sociétés humaines : du
complément alimentaire aux ressources utilitaires. Actes des XXIVe rencontres
internationales d’Archéologie et d’Histoire d’Antibes (23-25 Octobre 2003), Antibes, p.
417-430
FIORILLO A. R. 1984 - An introduction to the identification of trample marks. Current
Research in the Pleistocene (Orono, Me.), 1, p. 47-48
FIORILLO A. R. 1989 – An experimental study of trampling : implication for the fossil
record. In : R. Bonnichsen et M. H. Sorg (Ed.), Bone modification. Orono : Center for the
study of first Americans, p. 61-72
FIORILLO A. R. 1991 – Pattern and process in bone modification. Anthropologie, 29 (3), p.
157-161, 2 fig.
FISHER J. W. Jr 1995 – Bone suface modifications in zooarchaeology. Journal of
Archaeological Method and Theory, 2 (1), p. 7-67, tabl., fig.
GIFFORD-GONZALEZ D. P.; DAMROSCH D. B.; DAMROSCH D. R.; PRYOR J.;
THUNEN R .L. 1985 - The Third Dimension in Site Structure : An Experiment in
Trampling and Vertical Dispersal. American Antiquity, 50 (4), p. 803-818, 9 fig., 3 tabl.
GUILDAY J. E. ; PARMALEE P. W. ; TANNER D. P. 1962 – Aboriginal butchering
techniques at the Eschelman site (36 La 12), Lancaster County, Pennsylvania. Pennsylvania
Archaeologist, vol. 32, p.59-83, tabl., fig.
HILL A. 1986 – Tools, teeth and trampling. Nature, 319, p. 719- 720
LAROULANDIE V. 2004 - Exploitation des ressources aviaires durant le Paléolithique en
France : bilan critique et perspective. In : APDCA (Ed.), Petits animaux et Sociétés
humaines : du complément alimentaire aux ressources utilitaires. Actes des XXIVe
rencontres internationales d’Archéologie et d’Histoire d’Antibes (23-25 Octobre 2003),
Antibes, p. 163-172
LENOBLE A. ; BORDES J.-G. 2001 – Une expérience de piétinement et de résidualisation
par ruissellement. In : M. Mergoil (Ed.), Préhistoire et Approche expérimentale. sous la

51
direction de M. C. Frère-Sautot, p 295-311, 3 tabl., 15 fig.
LYMAN R. L. 1994 – Vertebrate taphonomy. Cambridge : Cambridge Press University, 524
p. ill.
LYMAN R. L. 2005 – Analyzing cut marks : lessons from artiodactyl remains in the
northwestern United States. Journal of Achaeological Science, 32, p. 1722-1732, tab. ,fig.
MALLYE J. B. ; LAROULANDIE V. 2004 - L’exploitation des petits animaux et les pièges
de l’os : exemple chez les oiseaux et les mustélidés. In : APDCA (Ed.), Petits animaux et
Sociétés humaines : du complément alimentaire aux ressources utilitaires. Actes des XXIVe
rencontres internationales d’archéologie et d’histoire d’Antibes (23-25 Octobre 2003),
Antibes, p.185-190
MC BREARTHY S. ; BISHOP L. ; PLUMMER T.; DEWAR R.; CONARD N. 1998 – Tools
underfoot : human trampling as an agent of lithic artifact edge modification. American
Antiquity, 63 (1), p. 108-129, 5 tabl., 11 fig.
MORLAN E. R. 1984 – Toward the definition of criteria for the recognition of artificial bone
alterations. Quaternary Research, 22, p. 160-171, 1 tabl., 3 fig.
NICHOLSON R. A. 2001 – Taphonomic ivestigations. In : D. R. Brothwell et Pollard A. M.
(Ed.). Handbook of Archaeological Sciences, Ch. 15, p. 179-190, 2 fig.
NIELSEN A .E. 1991 - Trampling the Archaeological Record : An Experimental Study.
American Antiquity, 56 (3), p. 483-503, 9 fig., 11 tabl.
OLIVER J. S. 1989 - Analogous and site context : bone damage from Shield Trap Cave
(24CB91), Carbon county, Montana, U.S.A. In : R. Bonnichsen et M. H. Sorg (Ed.), Bone
modification. Orono : Center for the study of first Americans, p. 73-98
OLSEN S. L. ; SHIPMAN P. 1988 – Surface modification on bone : trampling versus
butchery. Journal of Archaeological Science, 15, p. 535-553, 10 fig.
PEREZ RIPOLL M. 2004 - La consommation humaine des lapins pendant le Paléolithique
dans la région de Valencia (Espagne) et l’étude des niveaux gravétiens de la Cova de les
Cendres (Alicante). In : APDCA (Ed.), Petits animaux et Sociétés humaines : du
complément alimentaire aux ressources utilitaires. Actes des XXIVe rencontres
internationales d’Archéologie et d’Histoire d’Antibes (23-25 Octobre 2003), Antibes, p.191-
206

52
ROBERTS S. J. ; SMITH C. I. ; MILLARD A. ; COLLINS M. J. 2002 – The taphonomy of
cooked bone : characterizing and its physico-chemical effects. Archeometry, 44 (3), p. 485-
494
SELVAGGIO M. M. 1994 – Carnivore tooth marks and stone tool butchery marks on
scavenged bones : archaeological implications. Journal of Human Evolution, 27, p. 215-
228, 4 tabl., 4 fig.
SHIPMAN P. ; ROSE J. 1983 – Early hominid hunting, butchering, and carcass-processing
behaviors : approaches to the fossil record. Journal of Anthropological Archaeology, 2, p.
57-98, tabl., fig.
SHIPMAN P. ; ROSE J. 1984 – Cutmarks mimics on modern and fossil bovid bones. Current
Anthropology, 25 (1), p. 116-117, 3 fig.
STOCKTON E. D. 1973 – Shaw’s Creek Shelter : human displacement of arteacts and its
significance. Mankind, 9, p. 112-117, 2 tabl., 2 fig.
WALKER P. L. ; LONG J. C. 1977 – An experimental study of the morphological
characteristics of tool marks. American Antiquity, 42 (4), p. 605-616, 3 tabl., 7 fig.

53
ANNEXES

54
Annexe 1
Protocole pour l’analyse granulométrique du terreau
Cette analyse complète l’expérience, il est très important de connaître la part d’éléments
sableux ou de graviers que peut contenir le terreau. En effet, ces derniers par l’action du
piétinement peuvent créer des traces sur les os. Le but de cette analyse est de connaître la part
de fraction sableuse, supérieure à 0,25 mm.
Masse initiale de l’échantillon humide : 54,5 g
Masse de l’échantillon sec : 44,7 g
1. Destruction de la matière organique
Utilisation de peroxyde d’hydrogène à 25%, et pour accélérer la réaction chauffage à
environ 90°
2. Séparation de fraction supérieure/inférieure à 50 µm
3. Séchage et évaporation à l’étuve à une température de 40°
4. Tamisage de 1,25 mm à 50 µm
5. Pesée :
Poids total fraction minérale : 21,07 g
Fraction inférieure à 50 µm (limon et argile) : 5,08 g
Fraction supérieure à 50 µm (caillou, gravier, sable) : 15,86 g
Classe de
taille >1,25 mm 1,25 mm>x>0,25 mm 0,25 mm>x>0,1 mm 0,1 mm >x> 50 µm
Poids de la
fraction (g) 7,35 6,72 1,47 0,32
55
Annexe 2 :
Légende :
strie profonde trace d’abrasion ou en comète
strie superficielle desquamation
fracturation enfoncement
Planche 1 : Stries relevées sur les fémurs de Blaireau.
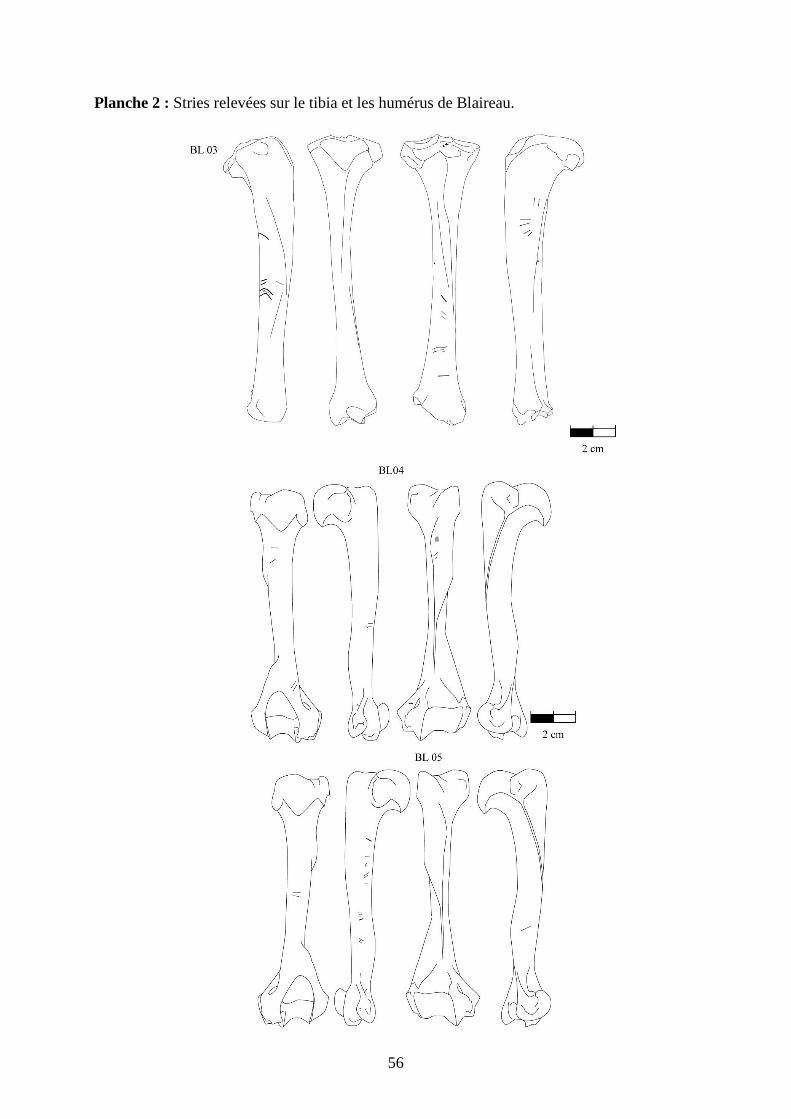
56
Planche 2 : Stries relevées sur le tibia et les humérus de Blaireau.
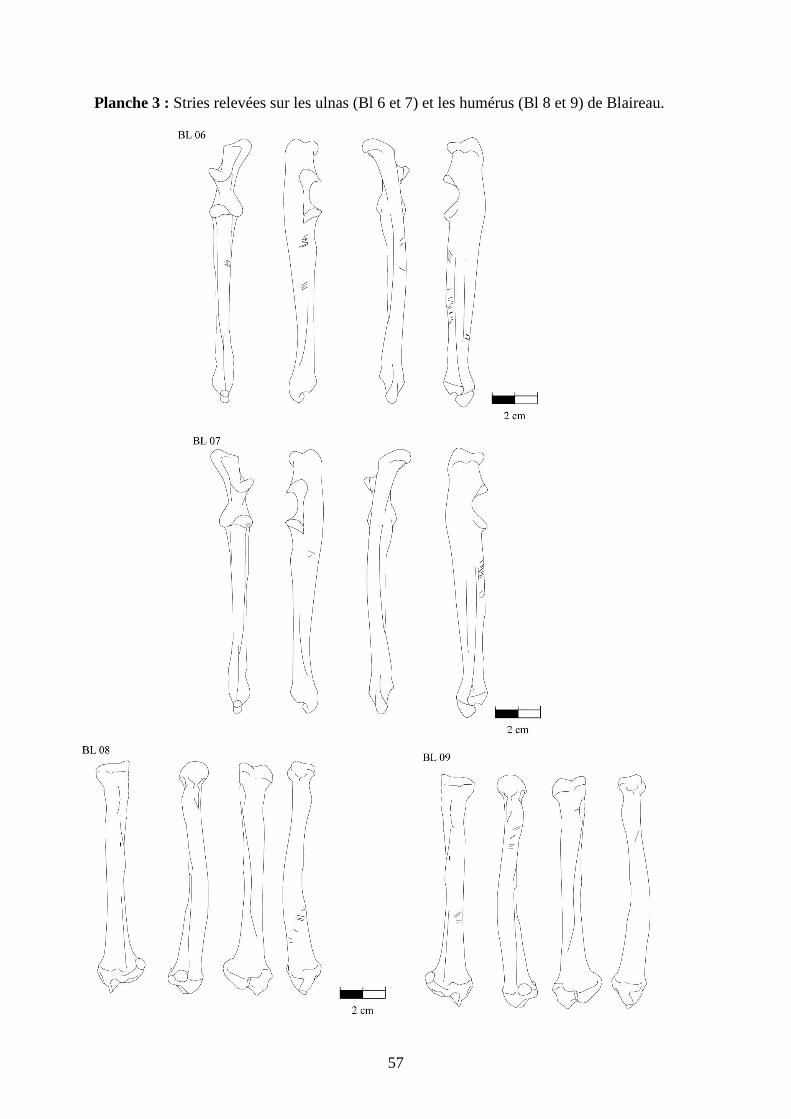
57
Planche 3 : Stries relevées sur les ulnas (Bl 6 et 7) et les humérus (Bl 8 et 9) de Blaireau.
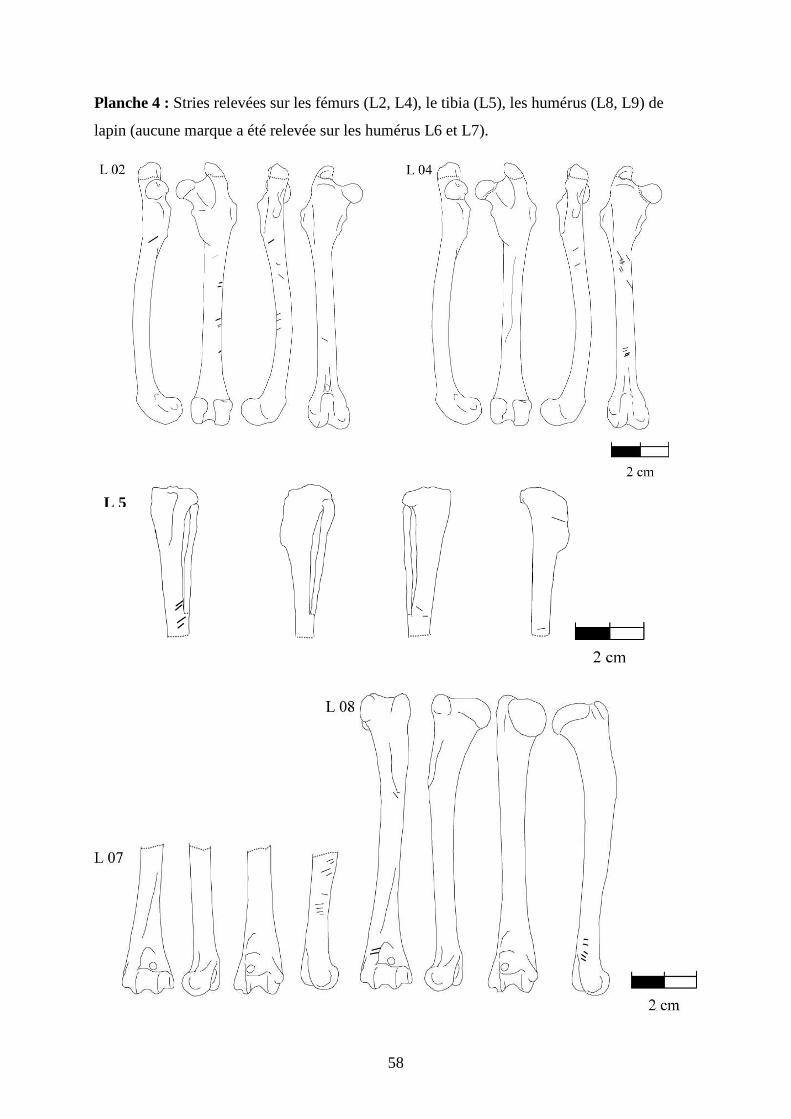
58
Planche 4 : Stries relevées sur les fémurs (L2, L4), le tibia (L5), les humérus (L8, L9) de
lapin (aucune marque a été relevée sur les humérus L6 et L7).
L 5
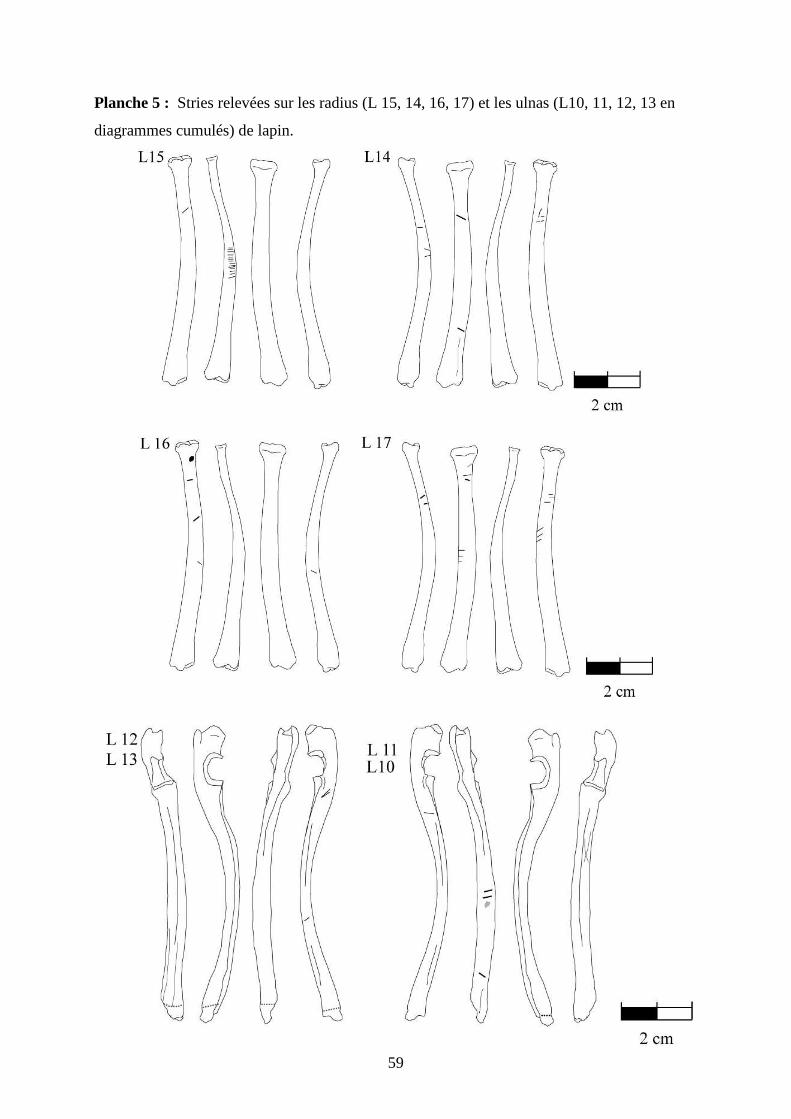
59
Planche 5 : Stries relevées sur les radius (L 15, 14, 16, 17) et les ulnas (L10, 11, 12, 13 en
diagrammes cumulés) de lapin.
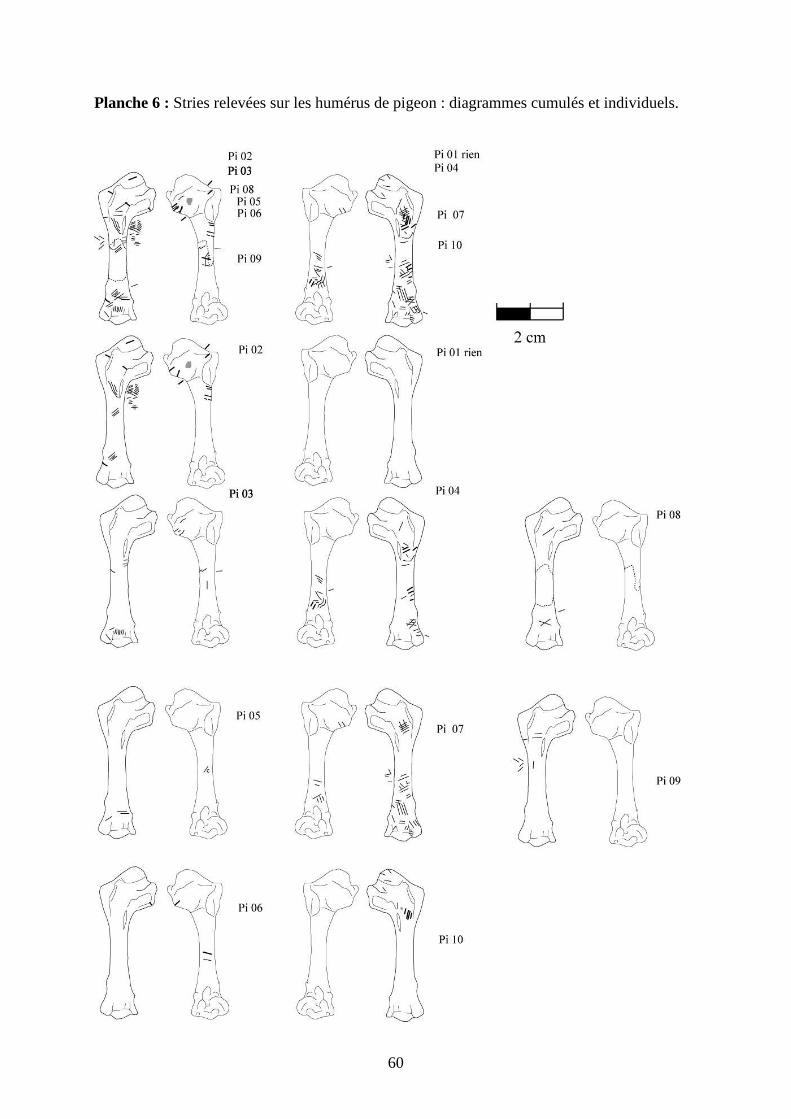
60
Planche 6 : Stries relevées sur les humérus de pigeon : diagrammes cumulés et individuels.

61
Planche 7 : Stries relevées sur les radius de pigeon : diagrammes cumulés et individuels.
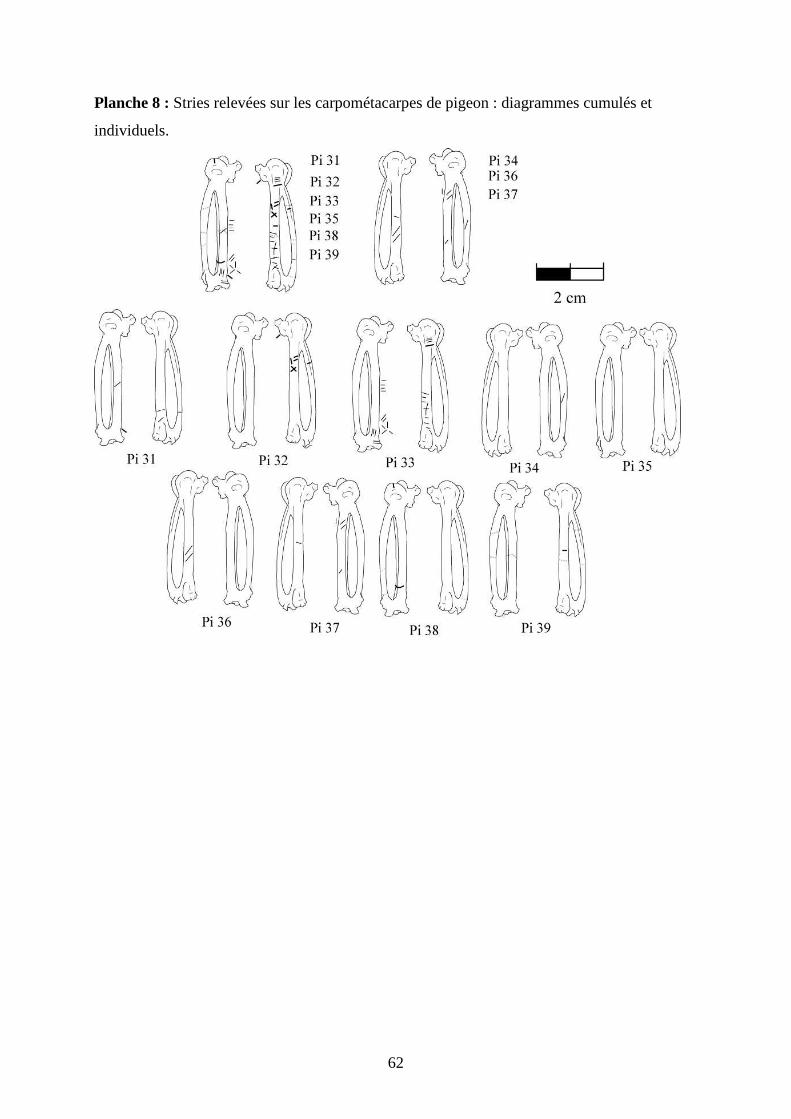
62
Planche 8 : Stries relevées sur les carpométacarpes de pigeon : diagrammes cumulés et
individuels.
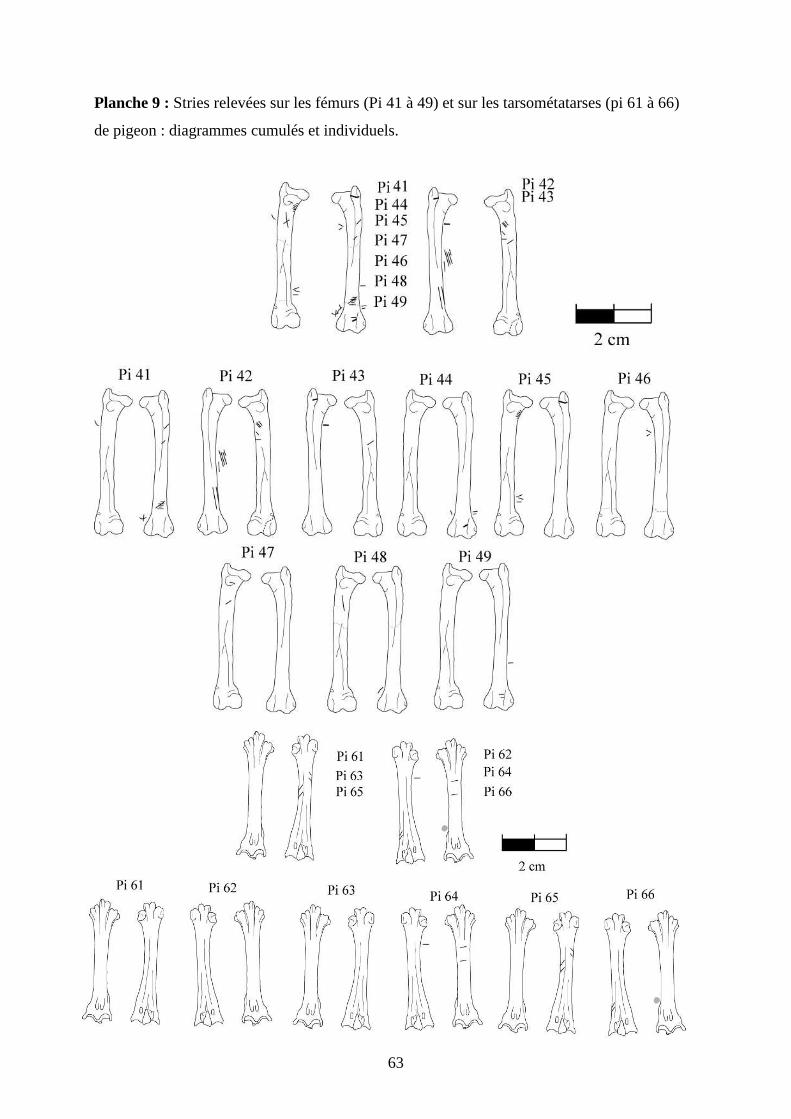
63
Planche 9 : Stries relevées sur les fémurs (Pi 41 à 49) et sur les tarsométatarses (pi 61 à 66)
de pigeon : diagrammes cumulés et individuels.
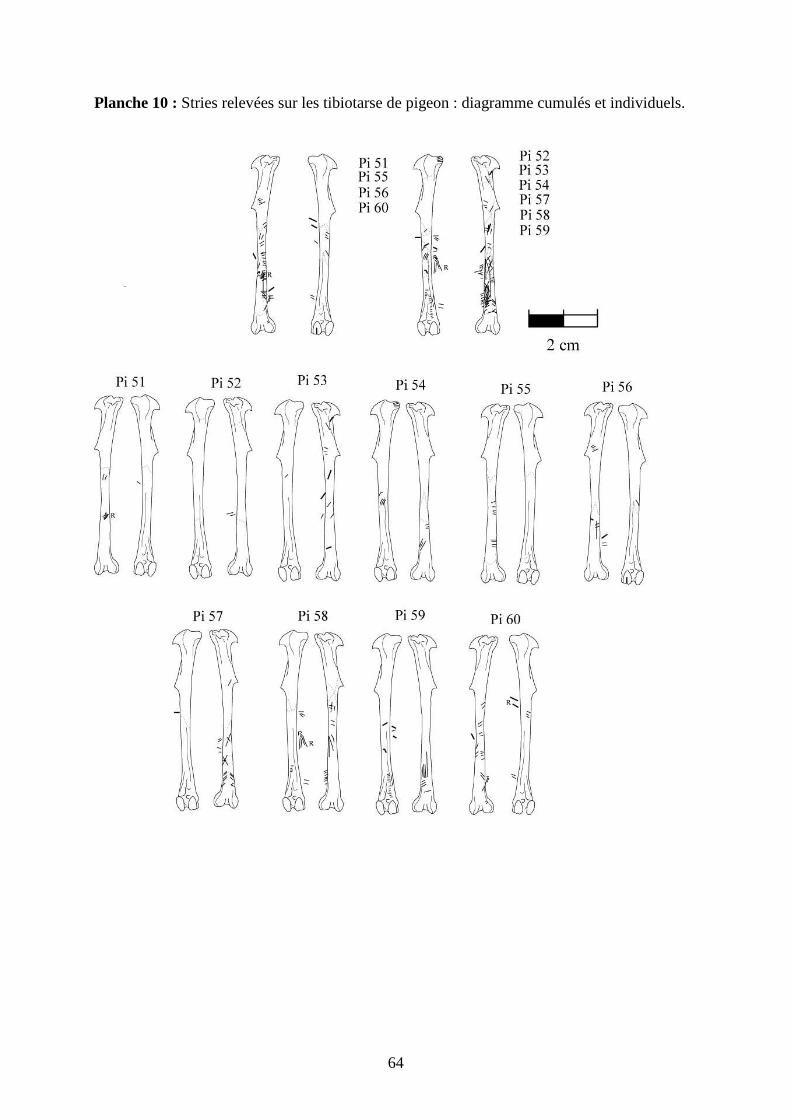
64
Planche 10 : Stries relevées sur les tibiotarse de pigeon : diagramme cumulés et individuels.
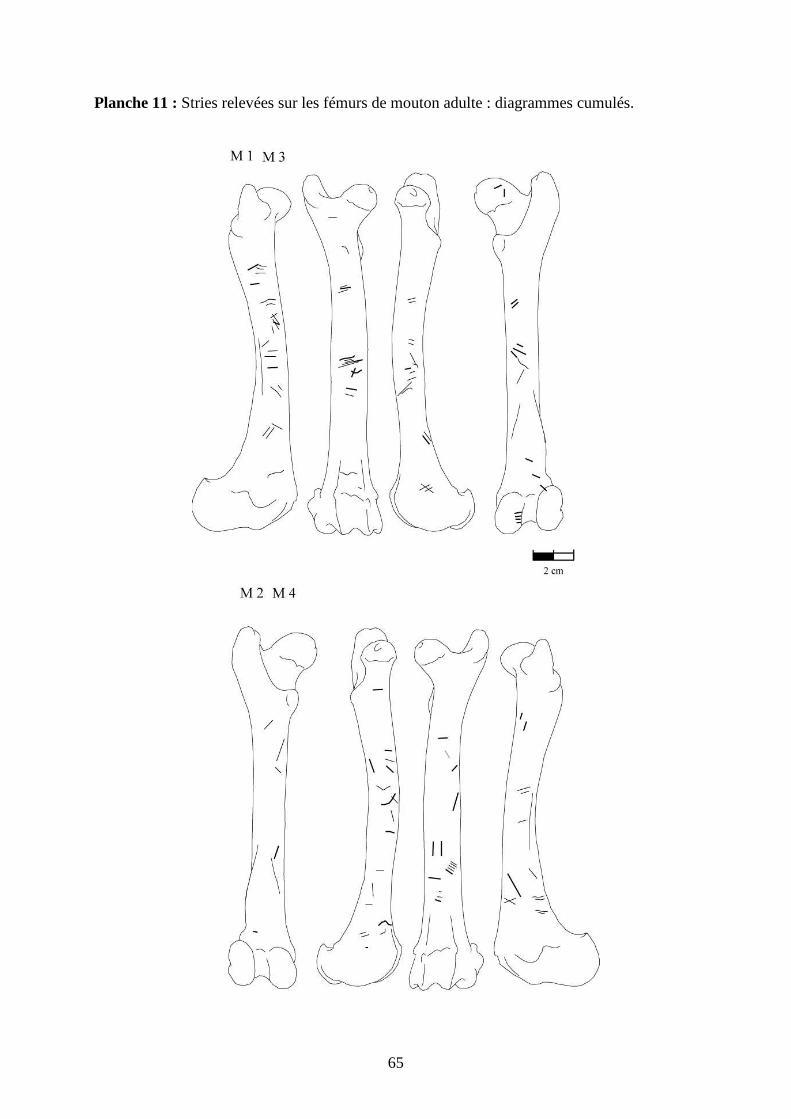
65
Planche 11 : Stries relevées sur les fémurs de mouton adulte : diagrammes cumulés.
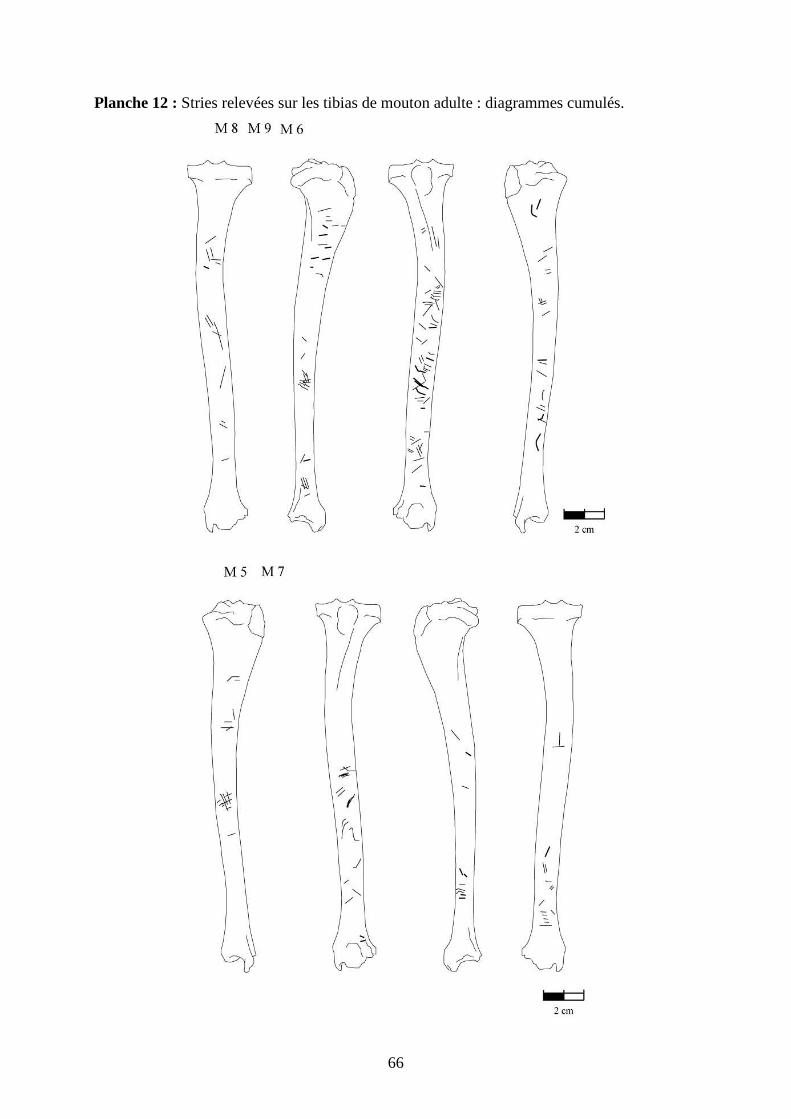
66
Planche 12 : Stries relevées sur les tibias de mouton adulte : diagrammes cumulés.
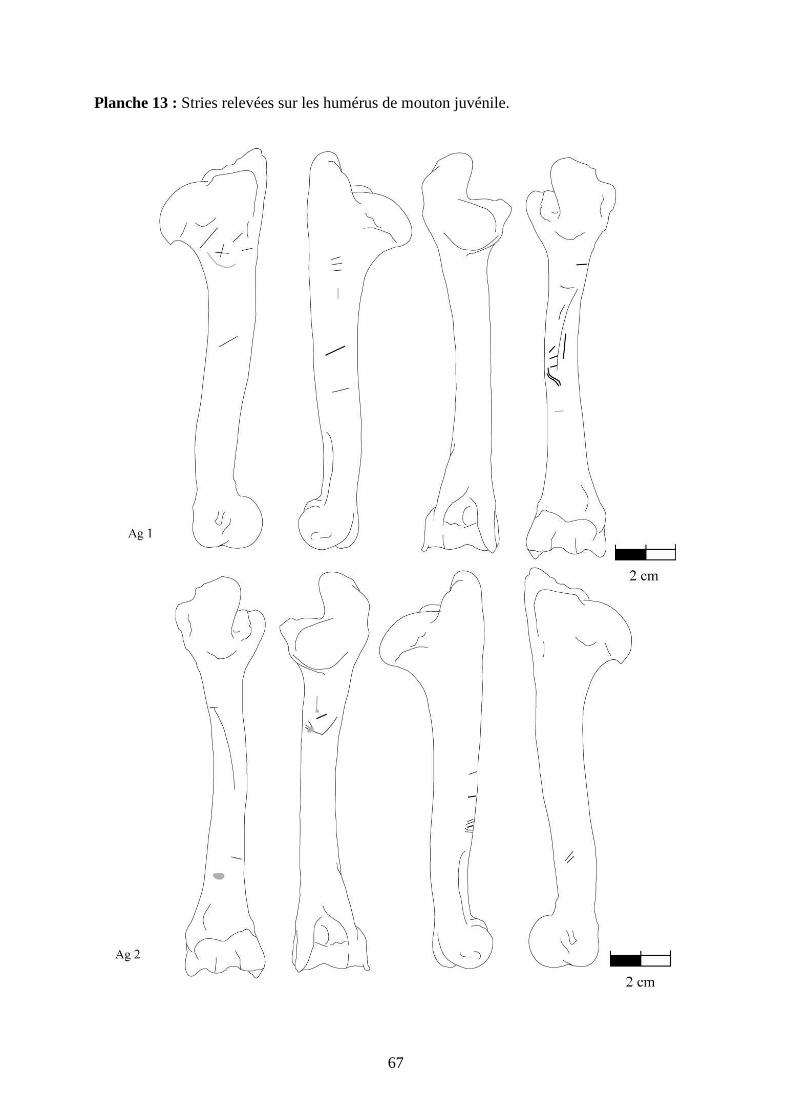
67
Planche 13 : Stries relevées sur les humérus de mouton juvénile.
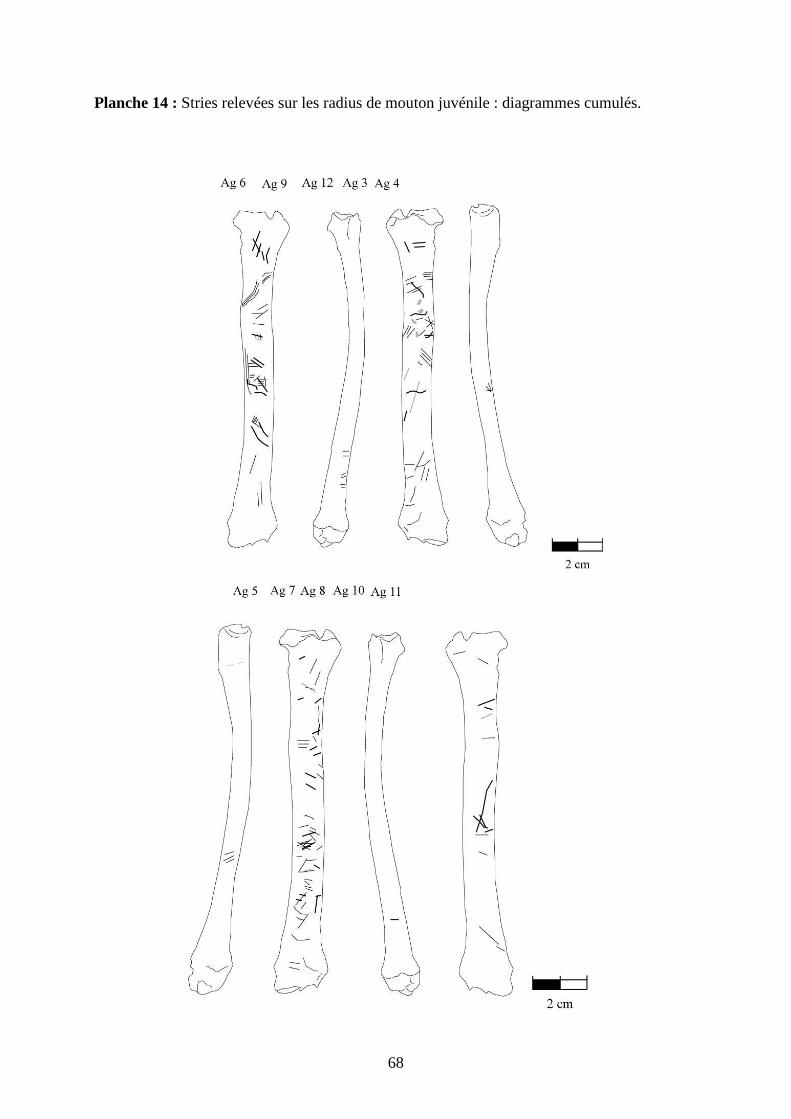
68
Planche 14 : Stries relevées sur les radius de mouton juvénile : diagrammes cumulés.
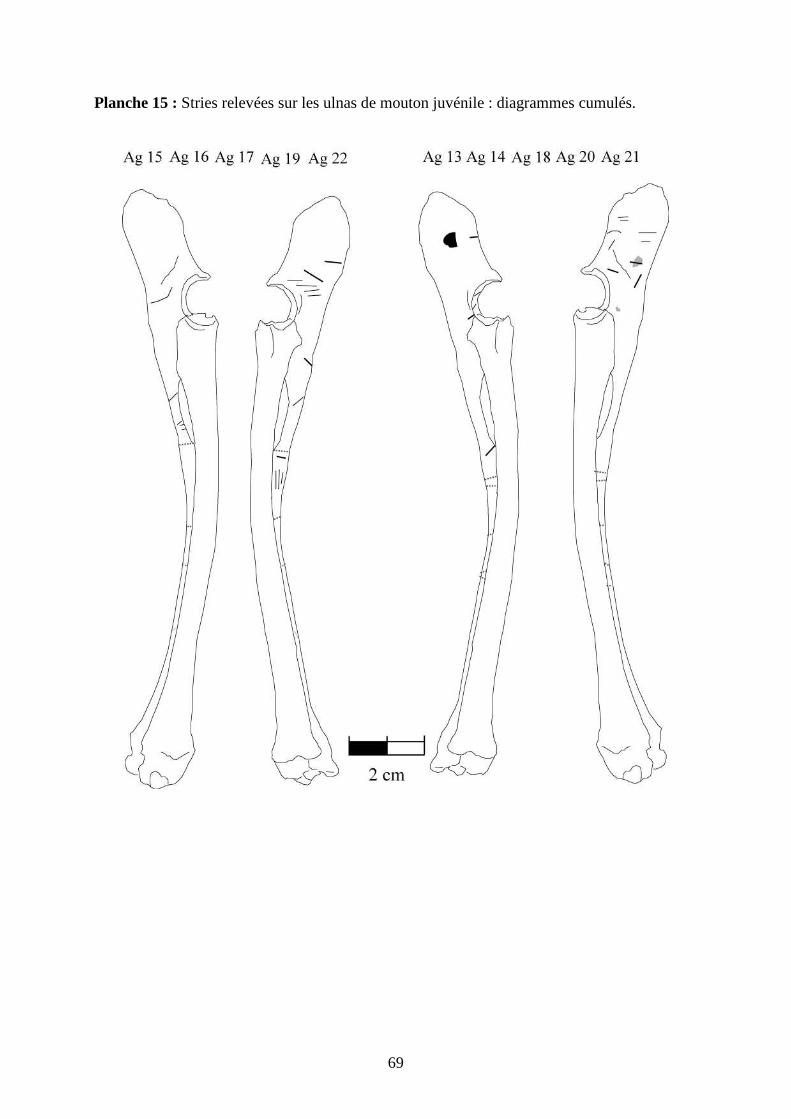
69
Planche 15 : Stries relevées sur les ulnas de mouton juvénile : diagrammes cumulés.
Copyright © 2022 FDOKUMEN


















![Spécificateurs engendrés par les traits [±ANIMÉ], [±HUMAIN], [±CONCRET] et structures d’arguments en arabe et en français](https://static.fdokumen.com/doc/165x107/63196bfebc8291e22e0f184b/specificateurs-engendres-par-les-traits-anime-humain-concret-et.jpg)

