
Ethogram of the free ranging Nasalis larvatus in Bako National Park, Sarawak. 2013
21
1 Ethogram of the free ranging Nasalis larvatus in Bako National Park, Sarawak MOHAMAD BIN KOMBI 1* and MOHD TAJUDDIN ABDULLAH 2 Abstract: An ethogram on the behaviour and daily activities of the Nasalis larvatus (Proboscis Monkey) provided, based on the field observation conducted in Bako National Park, Sarawak, from December 2011 until December 2012. There were seven categories of behavior successfully described, comprising various activities and movement patterns. They are feeding, resting, agonistic, grooming, solicitation, copulation and locomotion. These categories are then break down in several characteristic activities and actions, such as, jumping, leaping, quadrupedal walking, chasing, slapping, biting, facial threats, postural threats, grooming, solicitation, copulation and others miscellaneous behavior. The most frequently observed behavior recorded during the observation are feeding, travelling and resting. The ethogram and behavioural data provided have improved ourstanding on the behavioural patterns of the N. larvatus in Bako National Park. Keywords: Proboscis Monkey, Nasalis lavartus, ethogram, Bako Natioanal Park. 1 Forest Department Sarawak, Wisma Sumber Alam Jalan Stadium, 93060 Kuching, Sarawak, Malaysia. * Corresponding authors: [email protected] 2 Department of Zoology, Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia Malayan Nature Journal 2013, 65(2&3), 1-21
-
Upload
terengganu -
Category
Documents
-
view
3 -
download
0
Transcript of Ethogram of the free ranging Nasalis larvatus in Bako National Park, Sarawak. 2013

1
Ethogram of the free ranging Nasalis larvatus in Bako National Park, Sarawak
MOHAMAD BIN KOMBI1* and MOHD TAJUDDIN ABDULLAH2
Abstract: An ethogram on the behaviour and daily activities of the Nasalis larvatus (Proboscis Monkey) provided, based on the field observation conducted in Bako National Park, Sarawak, from December 2011 until December 2012. There were seven categories of behavior successfully described, comprising various activities and movement patterns. They are feeding, resting, agonistic, grooming, solicitation, copulation and locomotion. These categories are then break down in several characteristic activities and actions, such as, jumping, leaping, quadrupedal walking, chasing, slapping, biting, facial threats, postural threats, grooming, solicitation, copulation and others miscellaneous behavior. The most frequently observed behavior recorded during the observation are feeding, travelling and resting. The ethogram and behavioural data provided have improved ourstanding on the behavioural patterns of the N. larvatus in Bako National Park.
Keywords: Proboscis Monkey, Nasalis lavartus, ethogram, Bako Natioanal Park.
1 Forest Department Sarawak, Wisma Sumber Alam Jalan Stadium, 93060 Kuching, Sarawak, Malaysia.* Corresponding authors: [email protected] Department of Zoology, Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia
Malayan Nature Journal 2013, 65(2&3), 1-21

2
INTRODUCTION
Proboscis monkeys (Nasalis larvatus) are large and sexually dimorphic Colobines that are endemic to the island of Borneo (Bennett & Gombek 1993; Rowe 1996). The natural habitat of proboscis monkeys includes lowland coastal rainforests that contain mangroves and peat swamps (Salter et al. 1985). These animals are classified by the IUCN (1996) as endangered, and are listed in the CITES Appendix I. The subfamily Colobinae that is distributed in Asia and Africa includes 44 species from seven genera and 15 species of three genera, respectively (Groves 2001; Fashing 2007; Kirkpatrick 2007). This species is endemic to Borneo, occurring in Brunei, Indonesia (Kalimantan) and Malaysia (Sabah and Sarawak). It was originally found over the whole of coastal Borneo, as well as on the satellite islands of Berhala, Sebatik and Pulau Laut (Groves 2001).The species is in greater abundance in Kalimantan (Indonesian Borneo), where Meijaard surveyed the population. The Indonesian populations range in size from over 1,000 to less than 100, depending on past and current threats (Meijaard & Nijman 2000). The Pulau Kaget Nature Reserve population in South Kalimantan has reportedly been extinct since 1997 (Meijaard & Nijman 2000). Also, the population in the Mahakam Delta, East Kalimantan which would have numbered in the thousands up until the early 1990s, has now been decimated due to conversion of the coastal swamps to shrimp farms (IUCN 2012). In Sarawak, less than 1,000 animals are thought to remain in patchily distributed populations (Bennett et al. 1987) and in Sabah it was found to be more widespread and abundant than previously thought, with a minimum population size of c. 5,907 individuals found along major coastal river systems in Sabah (John et al. 2008). It has been suggested that it is restricted to the coastal areas and areas near rivers because the interior has soils that are low in minerals and salts, which are a necessary part of its diet (Bennett & Sebastian 1988). The species is mostly folivorous (about 52% of all feedings) and frugivorous (about 40% of all feedings), and prefers young leaves and unripened non-fleshy fruits (Bennett & Sebastian 1988; Yeager 1989). Monthly diet of this species changes with availability throughout the year (Yeager 1989). The home range size of a focal one-male group was 220.5 ha and never entered agricultural lands, nor areas used intermittently as log dumps for logging operations (Boonratana 2000). They moved inland during the day and returned to the river every night to sleep (Boonrata-na 2000). It avoids areas with heavy deforestation, such as agricultural land (Salter & MacKenzie 1985). Where there is no hunting the species can persist in disturbed forests and secondary habitats. In Malaysia, the proboscis is totally protected animal under Sarawak Wildlife Protection Ordinance 1998 and Sabah Wildlife Conservation Enactment 1997.

3
Living in the harsh environment of coastal areas where forest grows on nutrient-rich alluvial soils such as mangrove or alongside riversprovide large body species like the proboscis need an ample supply ofdigestible food where both mangroves and riverine forests have a much higher productivity than the dipterocarps forest (Bennett & Sabastian 1988). Due to the lack of minerals or other nutrients for inland forest of Borneo, the leaves of trees growing on the soil contain low levels of protein and higher levels fiber than those of trees growing on better soil (Bennett & Sabastian 1988). It seems that the poor soils of Bornean forest limit the habitats available to proboscis monkey either limiting the amounts of specific minerals or by limiting the absolute amount of food available. Kawabe and Mano (1972) stated that the mangrove trees serve as a key feature of habitat, providing leaves as food for the proboscis mon-key and a place to rest or sleep. Davis (1962) and Kern (1964) also stated that the major habitat-type of the proboscis monkey is a nipah mangrove association or a nipah-mangrove-mixed forest in the tidal swamp. Since the mangrove at Telok Asam was depleted and die-off due to natural causes, the foraging site that offered them fresh young leaves of mangrove species had been lost. So the dependency on mangrove species as the main resources of food change to the mixed dipterocarp species and beach forest dwelling species such as sea hibiscus (Hibiscus tiliaceus), tapang (Terminalia catappa), pandan (Pandan us utilis), bintangor (Calophylum inophyllum), bird nest fern (Asplenium nidus), putat (Barringtonia asiatica), fruit and soft part of the fronds of Nibung Palm (Oncosperma tigillaria) and others species that was not identified. Even the absent of mangrove forest at Telok Asam may not affect the diet of the proboscis monkey as tree growing in coastal and riverine had a high contain of protein than fiber. Other sources of nutritional food are available and enough digestible food needed with the ample supply of salt and other minerals that are found in coastal area. Although mangrove leaves are often cited as the primary food (Kay & Hylander, 1978), but other field studies indicate more diet choice, including non-sweet fruits (Bennett and Sabatian 1988), flowers (Salter et al. 1985) and seeds (Yeager 1989). The objective of this study is to construct an ethogram, describe and define the behavioural patterns and specific daily activities of Proboscis monkey populations in their natural habitat in Bako National Park, Malaysia. Hence constructing the ethogram was to better understand the behavior of the proboscis that has a function in the maintenance and ecological significance of the species in the changing or altering of the ecosystem by human or naturally. Unmanaged tourism threatened the proboscis populations where the increasing number of visitors to the park may affect the population behavior. Some activities of the visitors also disturb the proboscis monkey feeding time,

4
as some group of proboscis are not used to the present of human.
MATERIALS AND METHODS
Study area
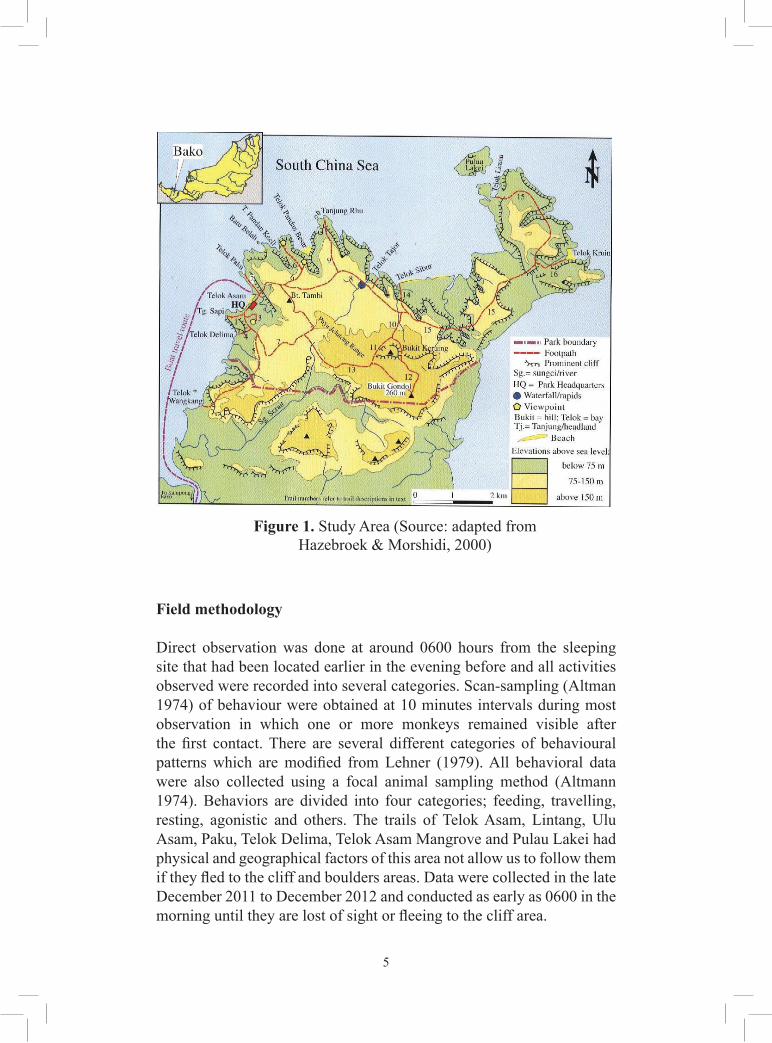
This study was conducted from December 2011 to December 2012 at Bako National Park (Figure 1) located about 30 km northeast of Kuching, at latitude 01º 41’ N and longtitude 110º 26’ E and altitude ranging from sea level to 244 meters (Hazebrock & Abang Kashim 2000). Covering an area of 2,727 hectares and known as the oldest, smallest National Park in Sarawak (Hazebrock & Abang Kashim 2000).The park consist of a diverse type of vegetation consists of seven distinct type of vegetation that includes beach forest, cliff forest, mangrove forest, tropical heath forest (kerangas forest), mixed-dipterocarp forest, padang or grassland vegetation and peat swamp forest (Ashton 1961; 1971; Hazebrock & Abang Kashim 2000) offered a wide range of habitats and ecosystem for plants and animals (Anderson 1961; Rothschild 1971). There is no major river in the vicinity of the park but only streamlet of Asam. The park is inhabited by not only proboscis monkeys but also 23 species of mammals, 150 species of birds and 24 species reptiles such as bearded pig (Sus barbatus), silver-leaf monkey (Presbytis cristata), long-tailed macaques (Macaca fascicularis) and colugo or flying lemur (Galeopterus variegatus). This area is an important tourism spot, and many tourists visited the park regularly in which the popular species is proboscis monkey. The number of visitors increased during week-end and public holidays. Some group of the proboscis monkeys are habituated to the presence of human in their natural habitat.
5
Field methodology
Direct observation was done at around 0600 hours from the sleeping site that had been located earlier in the evening before and all activities observed were recorded into several categories. Scan-sampling (Altman 1974) of behaviour were obtained at 10 minutes intervals during most observation in which one or more monkeys remained visible after the first contact. There are several different categories of behavioural patterns which are modified from Lehner (1979). All behavioral data were also collected using a focal animal sampling method (Altmann 1974). Behaviors are divided into four categories; feeding, travelling, resting, agonistic and others. The trails of Telok Asam, Lintang, Ulu Asam, Paku, Telok Delima, Telok Asam Mangrove and Pulau Lakei had physical and geographical factors of this area not allow us to follow them if they fled to the cliff and boulders areas. Data were collected in the late December 2011 to December 2012 and conducted as early as 0600 in the morning until they are lost of sight or fleeing to the cliff area.
Figure 1. Study Area (Source: adapted from Hazebroek & Morshidi, 2000)

6
ETHOGRAM CONSTRUCTION
An ethogram is a repertoire of all functional and non-functional behavioural patterns for the daily maintenance of an organism (Lehner, 1979). According to Torr and Shine (1994) a comprehensive knowledge of the basic behavioural patterns is needed before detailed studies can be conducted on social interactions and a basic definition and description are needed in order to classify animal behavior into categories (Dzulhelmi & Abdullah 2009). Behavioral studies in non-human primates have become ideal models for further investigations into advanced cognitive function in humans (Fan Xu et al. 2012). Therefore, to support future behavioral investigations and concomitant quantitative studies, it is imperative to establish systematic ethogram of non-human primates on a species-specific basis. However, to date, no systematic ethogram of the proboscis monkey (N. larvatus) has been constructed in Sarawak, Malaysian Borneo. In studying the behaviour of the proboscis monkeys in Bako National Park, major challenges are locating the harem group or the one-male group where most of their ranging areas are located inland with cliff, boulders and inaccessible. The behaviour the individual within the group is very curious to the present of human and will be quickly fleeing when they know the present of observer. Since this hook tail group (all-male) of proboscis monkey was habituated, we can make detailed observations not only the behaviour but also the feeding behavior for a longer observation time than the harem or one-male group. Therefore, we always choose to observe the all-male group that can easily be located and the behavior of adaptable to human present can be tolerated among the group members. The group that we call the hook-tail group where one the individual can be identified that having hook-like at the end of the tail. The observation was based on the individual from several groups of one-male and all male groups that was observed for 9,645 minutes among 63 individuals in the 27 groups that had been followed and observed. The study of behavior encompasses all of the movements and sensations by which animals and men mediate their relationships with their external environment of physical, biotic and social (Lehner 1979). So a comprehensive knowledge of the basic behavioural patterns is needed before detailed studies can be conducted on social interactions, hence basic definition and description are needed in order to classify animal behavior into categories (Torr & Shine 1994).

7
Feeding
Groups usually sleep in one or several nearby trees (Yeager 1990b) and feeding peaks were in the morning and at dusk (Boonratana 1993). Depending on the weather, if it was raining and cold they were normally not actively foraging but stayed in stationary position, resting or sleeping. For this activity they selected the young leaves, fruits and the soft part of the leaves by plucking, pulling, picking even breaking the small branches of tree. In this process, biting and chewing was done until all the food had been consume but it also happened that part of the leave are thrown away not been consumed. Sometime taking of food was done direct by the mouth, holding the branch without the activities of pulling, plucking or breaking. During the active feeding that was in the evening at about 1700 before sleeping and sometime early morning at 0630 hrs, they moved frequently and fast. Searching for another young leaf was usually done while eating and chewing (Figure 2), so the next move would be directly to the leaf to be consumed. In both the male and females, the ratio of feeding time was the highest at 1500 to 1700 hrs, i.e. shortly before sleeping, as they required about 50 hours for processing leaves so they may feed on lots of foods in late afternoon to spend the night time not only for sleeping but also for digestion (Matsuda 2008). For the fruits to be consumed, they did not eat the whole fruit but peeled the fruit to get the seed. Through our observation, bintagor fruit (Calophyllum inophyllum) was their favorite so as the leave, where they would stay for hours foraging in that tree. The distribution of food-bearing trees change seasonally (Paul & Petra 2005) hence influenced the foraging route. Compared to mobile prey such as flying insects and vertebrates, many species of primates spend the majority of their daily activity budget searching for and consuming plant resources such as fruits, flowers, leaves, and seeds (Garber 1987). Through observation period of 9,645 minutes, feeding activities get the most recorded with 1,513 times. This showed that feeding behaviour is frequently done activity compared to other activity. For the male they would spend more time on feeding than female to maintain his larger body (Matsuda 2008).

8
Figure 2. Feeding activity
The amount of food resources depleted from the environment will definitely affect the present and feeding pattern of the proboscis monkey. As what had been observed in Telok Asam, the depleted mangrove forest does not only reduce the source of food but also affect the daily foraging ground. The seasonally pattern of food resource tree such as Terminalia catappa, where the entire leaves drop off during the month of December will affect the foraging route so as the present of proboscis monkey in the area. The feeding behaviour that combined with the ability to discriminate between higher and low-quality food items require sophisticated gustatory, digestive and the cellulolytic bacteria in the forestomach to limit the potential toxic effects for certain alkaloid-rich foods (Gaber 1987).
Travelling and Moving
Travelling from one point to another either in the same tree or to another tree involves the activities of jumping, leaping, walking on their hands and legs (Figure 3) or running on two legs. Jumping is usually done when moving to another tree that might have quite a distance from each others that may need this activity to cross or reach the point. Leaping is movements that involve both hands, swinging from branch to branch mostly moving in the same tree with the short distant point. Quadrupedal walking (Figure 4) involved moving on the branch in the same tree or on the ground in the slow manner. But running on two legs happened

9
when crossing an open area on the ground where tree are separated in a manner that there could be danger or threat to them. It leaps with outstretched arms, using the whip-like action of slender branches to provide a springboard effect for take-off. It is known to hang for short periods, and sometimes moves for short distances, using only the arms. They classified its locomotors behaviour into the type quadrupedalism, a sub-type of Old World semi-brachiation. Activity of travelling or moving is actively happened during the feeding hours where they need to move from branches to branches foraging for young leaves especially when young leaves are scarce. But the abundant of food resource will limit the movement so as foraging distance. Jumping and leaping in a fast way usually happened when there is a threat and sign of danger that the adult male might had warn through vocalization.
Figure 3. Moving on the ground

10
Figure 4. Moving on the tree
Since there is no major river in vicinity of the park, swimming would not be seen in this observation and the streamlet of Asam are narrow where the canopy of trees on both side cover the streamlet, so create a passage for not only proboscis but other species of primate to cross. And there is a small bridge that cross Asam that link to other trails in the park main for visitors but species of primate tend to use it as a medium crossing passage. Our assumption of no swimming would happened was wrong on one occasion of my observation in this streamlet area for a groups of all-male proboscis, instead of going inland for big trees to cross, they jump in the water to cross to the side swimming (Figure 5) as Napier 1985; Yeager 1991a; mentioned that the monkeys are adept at swimming due to morphological feature, such as webbing fingers and toes and sufficient body fat.

11
Figure 5. Crossing the river
Resting
Immobile, station in one places either they are sleeping, scratching, yawning and self-grooming. This resting activity usually happened around 1000 to 1700 hours during the hot weather and heavily rain. Sleeping is either sitting with closed eyes or lying with face down on the branch or leaning the body to the tree. But this is not the behaviors of sleeping during the night sleep. During the night sleep, they just sit (squat) with face between the back limb on the branch and not that happened during the day due to the alertness to the danger. The most obvious behavior are their will not facing the observer and normally sitting backing (Figure 6) or sitting sideway and something that might cover or block direct contact such as leaves and branches. During heavily rainfall they would not make any movement, station in one place with head down between hind legs but sometime they do shake the body to clear off the rain water on the fur.

12
Figure 6. Not facing observer
Others activities during resting are sleeping (Figure 7 and 8) while sitting with eyes close and this happened in few minute without moving any part of the body. Scratching only happened sometime and so as self-grooming. Stretching the hind leg do happened when they squat for quiet long just what human do when sitting for too long. Self-directed behavior such as self-scratching, self-grooming and yawning happen in the situation of resting. Sleepy seem to be obvious during resting, where action like leaning the body to the trunk (Figure 7) and even lying with face down on the branch (Figure 8) assumed that the night sleep is more to alertness and anti predator strategy.

13
Figure 7. Resting activity
Figure 8. Asleep during resting

14
Agonistic
Agonistic interaction and action consisting of chasing, slapping, biting, vocalize, facial threats (Figure 9) and postural threats. Facial threats are a posture of the upper lip or both lips are vertically retracted, exposing the teeth and sometimes the gums where the corners of the mouth may be drawn back (Thierry, et al. 2000). In general, agonism is not usually seen between groups and members of the same group, for serious aggression is rare but minor aggression occurred among individual in the group such as facial threats, postural threats and vocalizations (Yeager 1990b, 1992b). Most of this agonistic behavior happened among juvenile and sub-adult where the sub-adult use to disturbed the juvenile with chases and facial threats for that the juvenile would vocalize with feared. It occurred between the younger animals or juveniles playing around among themselves chasing, slapping and biting. The activity continued where the others member of the group might be resting or sleeping. For adult male vocalization is common characteristic as a dominance and leader among the all male group. The different types frequency of high, low, soft vocalizations such as <Ngoh>, function as several meaning to warn, responding, instruction and quieting confusion among group members (Kawabe & Mano 1972). The sounds of <Gwat>, <Gat>, <Nat>, <Kat> in high-pitched is a warning against intruder and warning and threatening (Kawabe & Mano 1972) and this honking sound continue as the male proboscis watched cautiously at the intruder until all members of the group disappear to the cliff than he moved following his group. A loud call or a high-pitched vocalization emitted by adult males either in non-specific situations or in the context of arousal and social tension (e.g., social conflict, presence of a stranger) (Thierry et al. 2000). Agonistic interaction also happened when groups meet while foraging in the same area. There would an action of chasing among the individual and so as postural threats, by shaking, breaking the trees or branch. Before that a support shake with a violent shaking of environmental object (e.g., branch or trunk), producing an audible noise in the conditions of social tension and agonistic context (Thierry et al. 2000). During the chase some individual would accidently drop to the ground when they accidently jumping and landing on the small tree that might not strong enough to stand the body weight. In this situation the action was done by individual from both groups but not the adult male or the leader and it happened very fast and last in few minute. When it stop all of the members of both groups back doing their normal activity as if there is no incident happened among them.

15
The curiosity among the individual in the group on the present of observer is shown by facial threat to the observer. This behaviour usually accompanied vocalization and is done by all members of the group except the dominant adult male, especially when they saw someone is around them while feeding. At the same time they will move away from the tree or high in the tree where they not visible.
Figure 9. Facial threats
Grooming
This behavior is seldom seen in the daily activity of the proboscis, even during resting or position at sleeping site. This behaviour of comforting each others when resting or sleeping doesn’t happened in the proboscis social life. The only age that needs the need comfort and grooming is the infant. Even self-groom is seldom seen in the daily activity of the proboscis only some scratching done during resting. There is no gathering or sitting next to each others in the social life of the proboscis not like other non-human primate such as macaque (Macaca fascicularis), mutual grooming where an individual cleans the skin or fur of a partner while resting, sitting, gathering together and both mutually groom each other (Thierry et al. 2000). Scratching is done while resting or feeding, a repeated rubbing on the part of the body with hand and foot. Self-groom is seldom seen in

16
the activity of the proboscis but social grooming between juvenile and the adult male happened where the juvenile need protection from the adult male. The shake of the body is usually happened when the animals fur is wet during rainy or morning mist to cleared the fur before drying in the sunlight.
Solicitation and Copulation
No direct observation was made on this particular behavior during the study. Copulations are single mount and last on average around half a minute (Yeager 1990a; Boonratana 1993; Murai 2004; 2006). Solicitation happened before copulation between both male and female of proboscis monkey but solicitation does not always result in copulation (Murai 2006). Solicitation behavior between male and female start with pouted face, while male sometimes vocalize, hence female will approach and turn away, presenting her backside and looking backward at the male and sometime shake their heads to solicit copulation ( Hollihn 1973; Yeager 1990a; Boonratana 1993; Rajanthan & Bennet 1990; Murai 2006)
Interaction
On one occasion that was in January 2012 intraspecific interaction happened between a solitary male proboscis with a group of silver-leaf monkey (Presbytis cristata) reported by a group of researchers from University Malaysia Sarawak comprising of four ladies observing the behaviour the silver-leaf monkey. The solitary that had been abandon from the group either harem or all-male is protecting silver-leaf monkey from being attack by a group of proboscis monkey. Early the next morning I locate the solitary male sleeping in the same tree with the silver-leaf monkey in a position where intruder can access to attack the silver-leaf monkey. For that day the proboscis act like a body guard to the group of silver-leaf monkeys following and watching them from a distance, sometime communicate by sound or making sound as if instructing the group to make a move and the group move hence the proboscis follow from behind. This incident is very rare when two different species of primate that can interact and communicate though they are from different species of primate. During the observation the solitary proboscis male tried to avoid from my observation by hiding but still in vicinity of the group of silver-leaf monkey. Immediately after my absent from the area the proboscis monkey appeared. It seems that the present of man is a threat.

17
CONCLUSION
Although a number of studies had been conducted on the species, knowledge and detailed data on the behavior was not really emphasized. Most of the study done back in the mid-1980s and early 1990s and later in 2001- 2002 were more on ecology, population decline and habitat used. So lots of factors that may have influenced and change the behaviors of the proboscis monkey due loss of habitat, environmental changes and human interference. As the first author experienced in the late 1980s in Bako National Park, proboscis was not easily seen or observed due to their behavior that are not used to human present. But the situation has changed and they had to adapt to the present of human around them as they are the most popular species in Bako National Park. The systematic ethogram of N. larvatus in a natural environment precisely describes, defines and classifies the behaviors observed in the daily routine behavioural of this species. Though the habitat is not only inhibiting by the proboscis monkey, an overlapping among the long-tailed macaque and silver-leaf monkey seemed to be peaceful. Sometime proboscis monkey would moved when a group of macaque approaching or foraging in the same area as if to avoid confrontation, especially macaque due to aggressiveness behavior of macaque. The study on the behavior of the proboscis monkey has it own significance value in managing and conserving the habitats in a ways that will benefit both the resources of the park and the need of the proboscis monkey in term of it natural environment. The N. larvatus have been serological tested positive of Leptospira indicating that the species have once been exposed to the potentially pathogenic microbe (Thayaparan et al., 2013). Their habituated behaviour may have potential health risk to the visiting community in the park area that must be periodically monitored by the authorities. Nevertheless, further study on the behavior activities and patterns should be done to understand more and some function still had not been determined. Hence, the ethogram knowledge would help us to improve our understanding on the proboscis monkey conservation and management needs of this endangered species.
ACKNOWLEDGEMENTS
We would like to thank to Director Forest, Forest Department Sarawak, COE of Sarawak Forest Corporation and Bako National Park managers (Siali Aban and Abang Mutalib Abang Zainudin) for their permissionand permits to conduct research; the Universiti Malaysia Sarawak for the administrative and logistic support in conducting this study. Thank

18
you to Anuar Japar and Halik Akop for assisting in collecting of data and searching and tracking of the Proboscis monkeys. A special thank to all members of Proboscis Genome Project (Nur Aida Mohd Tamrin, Mohd Hanif Ridzuan, Wahap Marni, Madinah Arus, Mohd Isham, Ho Licia, Millawati, undergraduate and postgraduate students of Department of Zoology, UNIMAS) for sharing their information and knowledge, review and comments to improve the initial draft of this paper. Mr. Karim Hercus and Dr Lim Boo Liat chance meeting over coffee and roti in 2010, had given us a window opportunity and faith for us to initiate and conceptualise this project. The UNIMAS-MGI-MoSTI-MGRC Proboscis Genome Project lead by Prof. Dr. M.T. Abdullah, Associate Professor Yuzine Esa and Dr. Ramlah Zainuddin had financially supporting this research project is gratefully acknowledged. This work formed part of the MSc thesis by MBK at UNIMAS that was supported by the Proboscis Genome Project.
REFERENCES
Anderson, J.A.R. 1961. Bako National Park, Sarawak. Malayan Nature Journal, Special Issue. pp. 1-30.Alexander, R.D. 1974. The evolution of social behaviour. Annual Review of Ecology and Systematics.5:325–383.Altmann, J. 1974. Observational study of behavior: sampling methods. Behaviour 69:227–267.Altmann S.A., & Altmann. J. 1970. Baboon ecology: African field research. Karger, Basel.Ampeng, A. & B. M. Md-Zain. 2011. Ranging Patterns of Critically Endangered Colobine, Presbytis chrysomelas chrysomelas. The Scientific World Journal. doi:10.1100/2012/594382.Ashton, P.S. 1971. The Plants and Vegetation of Bako National Park. Malayan Nature Journal 24: 151-162.Bennett, E. 1988. Proboscis monkeys and their swamp forests in Sarawak. Oryx 22(1): 69-74.Bennett, E. & Sebastian, A. 1988. Social organization and ecology of proboscis monkeys (Nasalis larvatus) in mixed coastal forest of Sarawak. International Journal of Primatology 9(3): 233 -255.Boonratana, R. 1993. The ecology and behaviour of the proboscis monkey (Nasalis larvatus) in the lower Kinabatangan, Sabah. Asian Primates 4(1): 13-14.Boonratana, R. 1999. Dispersal in proboscis monkeys (Nasalis larvatus) in the Lower Kinabatangan, northern Borneo. Tropical Biodiversity 6: 179-187.Boonratana, R. 2000. Ranging behavior of proboscis monkeys (Nasalis larvatus) in the Lower Kinabatangan, northern Borneo. International Journal of Primatology 21: 497-518.Boonratana, R. 2002. Social organisation of proboscis monkeys (Nasalis larvatus) in

19
the Lower Kinabatangan, Sabah, Malaysia. Malayan Nature Journal 56: 57- 75.Brodie, J.F., & W. Brockleman. 2009. Bed site selection of red muntjac (Muntiacus muntjak) and sambar (Rusa unicolor) in a tropical seasonal forest. Ecol Res. 24 (6) 1251-1256.Cheney, D.L., & Wrangham. R.W. 1987. Predation. In: Smuts BB, Cheney DL, Seyfarth RM, Wrangham RW & Struhsaker TT (eds) Primate societies. University of Chicago Press, Chicago, pp 227–239Clutton-Brock,T.H, & Harvey, P.H. 1977. Primate ecology and social organization.J. Zoo. Lond. 183:1-39Dzulhelmi, M.N. & M.T. Abdullah. 2009. The Foraging Ecology of the Malayan Flying Lemur (Galeopterus variegatus) in Bako National Park, Sarawak, Malaysia. Journal Of Tropical Biology And Conservation 5: 31- 42.Fan, P.F. & Jlang, X.L. 2008. Sleeping sites, sleeping trees, and sleeping-related behaviors of black crested gibbons (Nomascus concolor jingdongensis) at Mt. Wuliang, Central Yunnan, China. American Journal of Primatology 70:153–160.Fashing. P.J. 2007. African Colobine Monkeys: Patterns of Between-Group Interaction. In: Campbell CJ, Fuentes A, Mackinnon KC, Panger M, Bearder SK (eds) Primates in perspective. Oxford University Press, New York, pp. 201-224Garber, P. A. 1987, Foraging strategies among living primates. Ann. Rev. Anthropol. 16:339–364.Groves, C. P. 2001. Primate taxonomy. Smithsonian Institution Press, Washington DC.Hamilton, W.J. 1982. Baboon sleeping site preference and relationship to primates grouping patterns. American Journal of Primatology 3:41–53.Hazebroek, H.P. & A.M. Abang Kashim. 2000. National Parks of Sarawak. Natural History Publication (Borneo), Kota Kinabalu. Ikki Matsuda. 2008. Feeding and Ranging Behaviors of Proboscis Monkey Nasalis larvatus in Sabah, Malaysia. Hokkaido University Collection of Scholarly and Academic Papers : HUSCAPJeffrey, S. 1982. Threats to the proboscis monkey. Oryx 16: 337-339.John, C. M. S., Henry, B. & Senthival, N. 2008. Status and Conservation of Proboscis Monkeys (Nasalis larvatus) in Sabah, East Malaysia, Primate Conservation (23): 107–120.Kirkpatrick, R.C. 2007. The Asian Colobines Diversity Among Leaf-Eating Monkeys. In: Campbell CJ, Fuentes A, Mackinnon KC, Panger M, Bearder SK (eds) Primats in perspective. Oxford University Press, New York, pp 186–200.
Koenig, A. 2000. Competitive regimes in forest-dwelling Hanuman langur females (Semnopithecus entellus). Behav. Ecol. Sociobiol. 48:93–109.Lehner, P.N. 1979. Handbook of Ethological Methods. Garland STPM Press, New York.Makey, D.& Waterman, P.G. 1982. Ranging behaviour of a group of black colobus

20
(Colobus satanas) in the Douala-Edea Reserve, Cameroon. Folia Primatol. 39:264–304.Meijaard, E. & Nijman, V. 1999. The local extinction of the proboscis monkey Nasalis larvatus in Pulau Kaget Nature Reserve, Indonesia. Oryx 34: 66-70.Meijaard, E. & Nijman, V. 2000. Distribution and conservation of the proboscis monkey (Nasalis larvatus) in Kalimantan, Indonesia. Biological Conservation 92: 15-24.Miller, L.E. & Treves, A. 2007. Predation on primates. In: Campbell CJ, Fuentes A, Mackinnon KC, Panger M & Bearder SK (eds) Primates in perspective. Oxford University Press, New York, pp 525–543.Miyoshi, K. & Higashi, S. 2005 Home range and habitat use by the sable Martes zibellina brachyuran in Japanese cool-temperate mixed forest. Ecological Research 20:95–101.B. Thierry, E.L. Bynum, S. Baker, M.F. Kinnaird, S. Matsumura, Y. Muroyama, T.G. O’Brien, O. Petit, & K. Watanabe 2000. The Social Repertoire of Sulawesi Macaques. Primate Research 16: 203-226.Murai, T. 2004. Social structure and mating behavior of proboscis monkey Nasalis larvatus (Primates; Colobinae). PhD Thesis. Graduate School of Environmental Earth Science, Hokkaido University. (Unpublished).Murai, T. 2006 Mating behaviors of the proboscis monkey (Nasalis larvatus). American Journal of Primatology 68:832-837.Murai, T. Mohamed, M., Bernard, H., Mahedi, P.A., Saburi, R., & Higashi, S. 2007 Female transfer between one-male groups of proboscis monkey (Nsalis larvatus). Primates 48:117–121.Napier, J.R. & Napier, P.H. 1967 A Handbook of Living Primates. Academic Press, London.Napier, P.H. 1985. Catalogue of primates in the British Museum (natural history) and elsewhere in the subfamily Colobinae. British Museum (Natural History), London.Oates, J.F. 1987. Food distribution and foraging behavior. In: Smuts BB, Cheney DL, Seyfarth RM, Wrangham RW & Struhsaker TT (eds) Primate societies. University of Chicago Press, Chicago, pp 197–209.Paul, A.G. & Petra, E.J. 2005. Travel Patterns and Spatial Mapping in Nicaraguan Mantled Howler Monkeys (Alouatta palliata). New Perspectives in the Study of Mesoamerican Primates: Distribution, Ecology, Behavior, and Conservation.Payne, J. C. M., Francias, C. M. & Phillipps, K. 1985. A field guide to the mammals of Borneo. The Sabah Society, Kota Kinabalu.Rajanthan R & Bennett EL. 1990. Note on the social behaviour of wild proboscis monkeys (Nasalis larvatus). Malayan Nature Journal 44:35–44.Reichard, U. 1998 Sleeping sites, sleeping places, and pre-sleep behavior of gibbons (Hylobates lar). American Journal of Primatology 46:35–62.Rothschild, G. 1971. Animals in Bako National Park. Malayan Nature Journal 24:

21
163-169.Salter, R.E, Mackenzie, N.A., Aken KM & Chai PPK. 1985. Habitat use, ranging behaviour, and food habits of the proboscis monkey, Nasalis larvatus (van Wurmb), in Sarawak. Primates 26:436-451.Salter, R. & MacKenzie, N. 1985. Conservation status of proboscis monkey in Sarawak. Biological Conservation 33: 119 - 132.Thayaparan, S., Robertson, I., Amraan, F., Su’ut, L. & Abdullah, M.T., 2013. Serological Prevalence of Leptospiral Infection in Wildlife in Sarawak, Malaysia. Borneo J. Resour. Sci. Tech. 2(2), 79-82.Torr, G.A. & R. Shine. 1994. An Ethogram for the Small Scincid Lizard, Lampropholis guichenoti. Amphibia-Reptilia 15: 21-34.Tuen, A.A. & Pandong, J.J.A.G. 2007. Habitat use and population density of proboscis monkeys (Nasalis larvatus) at Samunsam Wildlife Sanctuary, Sarawak. Malayan Nature Journal 59(3):269-79.Van Schaik, C.P.& van Hooff, J. 1983. On the ultimate causes of primate social systems. Behaviour 85:91–117.Van Schaik, C.P., van Amerongen A, & van Noordwijk, M.A. 1996. Riverine refuging by wild Sumatran long-tailed macaques. In: Fa JA, Lindburg DG (eds) Evolution and ecology of macaque societies. Cambridge University Press, Cambridge, pp 160–181.Xu, F., Xie. L., Li, X., Li, Q. & Wang, T., 2012. Construction and Validation of a Systematic Ethogram of Macaca fascicularis in a Free Enclosure. PLoS ONE 7(5): e37486. doi:10.1371/journal.pone.0037486.Yeager, C. P. & S.C.Silver, et al. 1997. Mineral and Phytochemical Influences on Foliage Selection by the Proboscis Monkey (Nasalis larvatus). American Journal of Primatology 41(2): 117–128.Yeager, C. 1989. Feeding ecology of the proboscis monkey (Nasalis larvatus). International Journal of Primatology 10(6): 497-529.Yeager, C. 1990a. Notes on the sexual behavior of the proboscis monkey (Nasalis larvatus). American Journal of Primatology 21(3): 223-227.Yeager, C. 1990b. Proboscis monkey (Nasalis larvatus) social organization: group structure. American Journal of Primatology 20(2): 95-106. Yeager, C. P. 1991a. Proboscis monkey (Nasalis larvatus) social organization: intergroup patterns of association. American Journal of Primatology 23: 73- 86.Yeager, C. P. 1991b. Possible Antipredator Behavior Associated With River Crossings by Proboscis Monkeys (Nasalis larvatus) American Journal of Primatology 24: 61-66.Yeager, C. P. 1992. Proboscis monkey (Nasalis larvatus) social organization: nature and possible functions of intergroup patterns of association. American Journal of Primatology 26: 133-137.Yeager, C. 1996. Conservation status of proboscis monkey groups and the effects of habitat degradation. Asian Primates Newsletter 5: 3-5.




















