
The Appropriate Use of Neurostimulation of the Spinal Cord ...
Upload
independentCategory
view
4download
0
Ž .Brain Research 753 1997 269–282
Research report
Epibatidine binding sites and activity in the spinal cord
Imran M. Khan a, Tony L. Yaksh a,b, Palmer Taylor a,)
a Department of Pharmacology, 0636, UniÕersity of California, San Diego, La Jolla, CA 92093, USAb Department of Anesthesiology, 0636, UniÕersity of California, San Diego, La Jolla, CA 92093, USA
Accepted 17 December 1996
Abstract
Epibatidine has been shown to be the most potent nicotinic agonist in several neuronal nicotinic receptor preparations. Similar to otherŽ .nicotinic agonists, intrathecal y -epibatidine elicits dose-dependent increases in pressor, heart rate and pain responses in rats, as well as
an increase in latency to withdraw from a noxious thermal stimulus. The latter response requires higher doses and is of shorter duration,suggesting interaction with multiple subtypes of spinal nicotinic receptors. In the present study, we relate the binding properties ofŽ . w3 x Ž . w3 x" - H epibatidine in spinal cord membrane preparations to the cardiovascular and behavioral responses. Unlike y - H cytisine orŽ . w3 x Ž . w3 xy - H nicotine, " - H epibatidine reveals two sites; the ratio of high affinity to low affinity sites is 2:1. The rank ordering of
Ž . w3 xpotencies of the nicotinic agonists in displacing " - H epibatidine binding from spinal cord membranes correlates with the potencies ineliciting cardiovascular and behavioral responses upon spinal administration. The nicotinic receptor antagonists, a-lobeline, dihydro-b-
Ž . w3 x Ž . w3 xerythroidine and methyllycaconitine, also displayed similar rank ordering of potencies in displacing " - H epibatidine, y - H cyti-Ž . w3 xsine or y - H nicotine binding to spinal nicotinic receptors. Virtually all the nicotinic analogs exhibited a Hill coefficient of less than
Ž . w3 xone in competing with " - H epibatidine to spinal cord membranes indicating their interaction with at least two classes of binding sites.Ž .Intrathecal y -epibatidine, in addition to eliciting an initial and subsequently a sustained pressor and tachycardic response, also exhibited
a transient intervening bradycardia which coincided temporally with the duration of the analgesia. Repeated administration ofŽ .y -epibatidine desensitized its responses as well as the cardiovascular and behavioral responses to spinal nicotine and cytisine.Intrathecal a-lobeline showed selectivity for blocking the analgesic response, whereas methyllycaconitine exhibited selectivity for thepressor and irritation responses. The NMDA receptor antagonist, AP-5, inhibited the pressor, tachycardic and irritation responses elicited
Ž .by intrathecal y -epibatidine, confirming a role for spinal excitatory amino acids released by the nicotinic agonist. q 1997 ElsevierScience B.V. All rights reserved.
Keywords: Epibatidine; Nicotinic receptor; Spinal cord; Cardiovascular response; Behavioral response
1. Introduction
Molecular cloning of nicotinic receptors from avian andmammalian species suggests a multigene family of central
Ž w x.nicotinic receptors cf. 19,20 . To date, nine subunits ofthe a-subtype and four subunits of the b-subtype havebeen identified in various species. The possibilities ofsubtypes of neuronal nicotinic receptors from differentcombinations of a- and b-subunits are extensive, yet onlycertain combinations appear in the central and peripheral
Abbreviations: IML, intermediolateral cell column; SBP, systolicblood pressure; HR, heart rate; S.E., standard error; i.t., intrathecal; AP-5,2-amino-5-phosphopentanoic acid
) Ž .Corresponding author. Fax q1 619 534-8248.
nervous system and form active receptors in expressionsystems. Thus, assigning a pharmacological signature tospecific combinations of a- and b-subunits requires acorrelation of pharmacology and binding properties inparticular regional areas of the CNS. We have previouslydemonstrated that nicotinic agonists administered intrathe-cally elicit dose-dependent increases in blood pressure,
w xheart rate and irritation responses in rats 15 . Moreover,the rank-ordering of potencies of the nicotinic agonists ineliciting the cardiovascular and behavioral responses corre-late with affinities of the agonists in binding to the spinal
w xnicotinic receptors 16 . The nicotinic antagonists alsoshow specificities in antagonizing the pharmacological re-sponses and binding of nicotinic agonists to spinal nico-
w xtinic receptors 15,16 . The studies collectively suggest thatthe complement of spinal nicotinic receptors are distinct
0006-8993r97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 97 00031-0

( )I.M. Khan et al.rBrain Research 753 1997 269–282270
from the neuronal nicotinic receptors present in the higherbrain centers and the ganglia.
Recently, epibatidine, a neurotoxin obtained from theskin of Epipedobates tricolor, has been shown to be themost potent nicotinic agonist in various neuronal nicotinic
w xreceptor preparations 2,5,8,26 . Interestingly, it has alsobeen shown to be many fold more potent than morphine in
w xeliciting a transient analgesic response 25 . The analgesicresponse elicited by epibatidine is also observed at the
w xspinal level in rats 9 . In addition, similar to other nico-tinic agonists, spinal epibatidine also elicits dose-depen-dent increases in blood pressure, heart rate and nociceptive
w xresponses 13 . However, spinal epibatidine is more potentin eliciting the nociceptive response than the antinocicep-
w xtive response 9 indicating interaction with multiple sub-types of spinal nicotinic receptors. In rat and human brain,w3 xH epibatidine binds to two or multiple classes of binding
w xsites of different affinities 12 . In the present study, weŽ . w3 xcharacterize the binding properties of " - H epibatidine
to spinal cord membranes and correlate binding with thepharmacological responses to spinal epibatidine, desensiti-zation by epibatidine and selectivity of spinal nicotinicantagonists in inhibiting the responses to i.t. epibatidine.
2. Materials and methods
2.1. Experimental animals
Ž .Male Sprague-Dawley rats 300–350 g were purchasedŽ .from Harlan Co. Indianapolis, IN . They received stan-
dard Purina Rat Chow and water ad libitum. All studieswere carried out according to protocol approved by theUCSD Institutional Animal Care Committee.
2.2. Spinal cord membrane preparation
Membranes were prepared according to the procedurew x Ždescribed earlier 16 . Briefly, a 5 cm 1 cm from the
.sacral end segment of the spinal cord was dissected andplaced in a polypropylene tube. Tissue sections were storedat y708C. Spinal cord sections were homogenized inice-cold 50 mM Tris-HCl buffer, pH 7.4. The homogenatewas centrifuged at 48,000=g for 10 min, the pelletresuspended in fresh buffer, centrifuged a second time, andthe final pellet resuspended in fresh buffer.
2.3. Equilibrium binding assays
Equilibrium binding assays were conducted accordingto the procedures described earlier for nicotine and cytisinew x16 . Briefly, the assay mixture consisted of 400–600 mgof membrane protein in a final incubation volume of 125
Ž . w3 xml. Final concentrations of " - H epibatidine varied be-Ž .tween 0.02 to 5 nmolrl 250 to 50,000 cpm , respectively;
Ž . w3 xstock solutions were prepared in assay buffer. " - H Ep-ibatidine incubations were carried out in the cold roomŽ .48C on a gentle shaker for 60 min. Assays were initiatedby the addition of the membrane suspension with rapid
Ž . w 3 xmixing to the " - H epibatidine solutions in apolypropylene tube. The incubations were terminated bydilution with 3 ml of ice-cold assay buffer and immedi-ately followed by rapid filtration under vacuum through
Ž .0.5% polyethylene imine PEI treated Whatman GFrCfilter papers at 48C. Filters were then rinsed three timeswith 3 ml of ice-cold buffer. Specific binding was deter-mined as the difference in binding between samples con-
Ž . Ž .taining excess unlabeled y -nicotine 40 mmolrl andwŽ . w3 xthose containing only " - H epibatidine. Protein was
Ž .assayed by the bicinchoninic acid BCA assay.Total binding sites and dissociation constants were de-
termined by Scatchard analysis of the saturation bindingdata. For the curvilinear Scatchard plots, dissociation con-stants were determined by regression analysis of each
Ž .slope using Sigma plot Jandel Scientific, CA, USA .
2.3.1. Competition studiesŽ .To determine dissociation constants K of the variousi
Ž . w3 xnicotinic agonists in inhibiting " - H epibatidine bind-ing to spinal cord membranes, two concentrations, 0.08and 0.8 nM, of radioligand were used in the competition
Ž . w3 xassay. A concentration of 0.08 nM " - H epibatidinewould occupy approximately 60% of the high affinitysites, whereas less than 20% of the low affinity sites wouldbe occupied. On the other hand, for the higher concentra-
Ž . w3 xtion of " - H epibatidine, virtually all of the high affin-ity sites and more than 60% of the low affinity sites would
Žbe occupied by the radioligand. Membranes 400–600 mg. Ž . w3 xprotein were incubated with " - H epibatidine and at
least 8 concentrations of a competing ligand in a totalvolume of 125 ml at 48C for 60 min. After dilution andfiltration, bound radioactivity was quantified as describedfor the equilibrium binding assays. IC values and Hill50
coefficients for each competing ligand were determinedgraphically by nonlinear regression analysis of log-logit
Ž .plots InPlot, GraphPad, San Diego, CA . K ’s for eachi
competing ligand for the high affinity sites were calculatedŽby InPlot using the equation K s IC r 1 qi 50
w x .epibatidine rK .d
2.4. Implantation of intrathecal catheters
Ž .For spinal drug delivery, intrathecal i.t. catheters werew ximplanted as described previously 15 . Briefly, rats were
Ž .anesthetized with halothane 2–3% , placed in a stereo-taxic frame and the occipital membrane was exposed. A 9
Ž .cm saline-filled polyethylene PE-10 tubing was placedinto the intrathecal space through the cisterna membraneand passed down to the rostral edge of the lumbar enlarge-ment. The catheter was externalized on top of the skull and

( )I.M. Khan et al.rBrain Research 753 1997 269–282 271
sealed with a piece of stainless steel wire. The rats wereallowed to recover for at least 4 days prior to further study.Only animals exhibiting normal motor behavior were em-ployed in the study.
2.5. Arterial blood pressure and heart rate recording
Rats with i.t. catheters were reanesthetized withŽ .halothane 2–3% and their tail arteries were catheterized
Ž .with a polyethylene PE 50 tube filled with heparinŽ .containing saline 1 unitrml . The wound was covered
with gauze-tape and the rat was placed in a restrainer cage.The cage was constructed to enable the animal to maintaina typical crouching posture. The tail artery catheter wasconnected to a blood pressure transducer coupled to a
Ž .Grass polygraph Model 7 . Heart rates were measuredusing a cardiotachometer triggered from pressure pulses.Rats were allowed to recover for at least 30 min afterplacement in the cage for administration of drugs. Bloodpressure and heart rate were monitored continuously forthe duration of the experiment.
2.6. Administration of drugs
The following chemicals were obtained from SigmaŽ . Ž . Ž .Chemical Co. St. Louis, MO : y -nicotine, q -nico-
Ž .tine, y -cytisine, a-lobeline and mecamylamine. Dihy-Ž . Ž .dro-b-erythroidine, q - and y -epibatidine-L-tartrate,
Ž .q -anatoxin-A hydrochloride, methyllycaconitine andŽ .AP-5 2-amino-5-phosphopentanoic acid were obtained
Ž .from Research Biochemicals International Natick, MA .Ž . Ž .y -Epibatidine as the free base or q -epibatidine as thetartrate salt corresponds to the natural enantiomer; our
Ž .designation of chirality refers to the free base. " -w3 xH Epibatidine was obtained from New England NuclearResearch with a specific activity of 56.2 Cirmmol and99% specified purity.
Drugs were dissolved in sterile saline solutions andadministered using a hand-driven syringe pump over aninterval of 15–25 s. Solutions of drugs were prepared toachieve a 10 ml final delivery volume. Drug administra-tion was followed by 10 ml of saline flush within 15 s toclear the catheter.
2.7. Experimental paradigms for drug administration
Ž .1 Agonist dose–response relationships. Dose–re-Ž .sponse curves for y -epibatidine were developed by ad-
ministration of 0.005, 0.05 or 0.5 mg of the agonist inseparate groups of rats, and cardiovascular and behavioralresponses elicited by each dose of the agonist measured.
Ž .2 Agonist desensitization. Desensitization to agonistswas evaluated by repeated administration of the highestdose within the 35–40 min interval and the cardiovascularand behavioral parameters measured.
2.8. Statistics
All values presented are means"S.E. A Student’s t-testfor unpaired data was used to determine differences be-tween two groups of treatment. Differences between multi-ple groups were compared using an ANOVA.
3. Results
( ) [ 3 ]3.1. Equilibrium binding analyses for " - H epibatidine
Ž . w3 xBinding of " - H epibatidine to rat spinal cord mem-Žbranes prepared from a 5-cm segment of thoracic and
. Ž .lumbarosacral regions was found to be saturable Fig. 1a .Non-specific binding was always less than 20% of totalbinding over the entire concentration range between 0.02
Ž . w3 xand 5 nM of " - H epibatidine. Ligand binding to 0.5%
Ž . w3 xFig. 1. A: equilibrium binding of " - H epibatidine in spinal cordŽmembranes of Sprague-Dawley rats. Membranes 400–600 mg of pro-
. Ž . w3 x Žtein were incubated with varying amounts of " - H epibatidine 0.02.to 5 nM in 125 ml for 60 min at 48C. Specific binding was estimated as
the difference between total binding and non-specific binding. B:Scatchard analysis of the saturation binding data. The curves are represen-tative of one of three individual assays which comprised of spinal cordmembrane preparations pooled from 4–5 rats. Equilibrium binding assaysfor the saturation binding analysis were also done in 8-fold greatervolume and corresponding reduction in concentration. Scatchard analysesof these data yielded a K of 0.046"0.001 nM and total number ofd
binding sites of 19.1"1.3 fmolrmg protein for the high affinity sites.

( )I.M. Khan et al.rBrain Research 753 1997 269–282272
w3 xFig. 2. Inhibition of H epibatidine binding to spinal cord membranes byŽ .various nicotinic agonists. Membranes 400–600 mg of protein were
Ž . Ž . w3 xincubated with 0.08 nM A or 0.8 nM B of H epibatidine and 10–12concentrations of competing agents for 60 min at 48C.
PEI pretreated glass-fiber filters in the absence of tissueprotein was less than 2% of the assay values over therange of concentrations.
Scatchard analyses of the binding data revealed twoclasses of binding sites differing in affinity and number of
Ž .binding sites with equilibrium dissociation constants Kd
of 0.36"0.10 nM and 0.07"0.01 nM for the low andŽ .high affinity sites, respectively Fig. 1b . Total saturable
Ž . Ž .sites for the high site 1 and low site 2 affinity bindingsites were 20.5"1.0 fmolrmg protein and 9.2"1.6fmolrmg protein, respectively.
Ž . w3 xFig. 3. Inhibition of " - H epibatidine binding to spinal cord mem-Žbranes by various nicotinic antagonists. Membranes 400–600 mg of
. Ž . Ž . Ž . w3 xprotein were incubated with 0.08 nM A or 0.8 nM B of " - H epi-batidine and 10–12 concentrations of competing agents for 60 min at 48C.
In order to obtain a better estimate of the dissociationconstants and the total number of sites for the high affinitysites, equilibrium binding assays for the saturation bindinganalysis were done in 8-fold greater volume than for theregular assay. Scatchard analyses of these data yielded aK of 0.046"0.001 nM and total number of binding sitesd
of 19.1"1.3 fmolrmg protein for the high affinity sites.Ž . w3 xTwo classes of binding sites for " - H epibatidine
were found in thoracic, and dorsal and ventral sections ofŽ .the lumbarosacral regions of the spinal cord Table 1 .
w3 xSimilar to our previous observations with H cytisine andw3 x Ž .H nicotine, " -epibatidine exhibited greater number ofbinding sites in the dorsal-lumbar than in the ventral-lumbar
Table 1Ž . Ž . Ž . w3 xEquilibrium dissociation constants K and total saturable sites B of " - H epibatidine binding to membrane preparations from dorsal-lumbar,d max
ventral-lumbar and thoracic spinal cord of Sprague-Dawley rats
Ž . Ž .Spinal cord regions K nmolrl B fmolrmg proteind max
Site 1 Site 2 Site 1 Site 2
Dorsal-lumbar 0.13"0.01 0.31"0.03 38.9"2.2 6.7"0.7Ventral-lumbar 0.08"0.01 0.31"0.03 17.9"0.5 5.9"0.9Thoracic 0.10"0.01 0.33"0.05 16.7"2.2 5.5"1.1
Standard errors of the mean represent 3–5 measurements on separate preparations.

( )I.M. Khan et al.rBrain Research 753 1997 269–282 273
or thoracic regions of the spinal cord. By far, the greatestregional differences in binding site numbers were observedfor the high affinity sites.
( ) [ 3 ]3.2. Inhibition of " - H epibatidine binding by nicotinicagonists
To determine the pharmacological specificity of thenicotinic binding sites, the capacity of the various nicotinic
Ž .agonists and antagonists to inhibit the binding of " -w3 xH epibatidine to spinal cord membranes was measuredŽ . Ž . w3 xTable 2 and Figs. 2 and 3 . Displacement of " - H epi-batidine in the presence of various concentrations of sev-eral nicotinic analogs was assayed at ;0.08 nM and 0.8
Ž . w3 x Ž . w3 xnM of " - H epibatidine. Since " - H epibatidinebinds to two different classes of sites in the spinal cord,using the low and high concentrations of the radioligandshould enable one to obtain pharmacological profiles for
Ž .the various nicotinic analogs in competing with " -w3 xH epibatidine for the two classes of sites. A concentra-
Ž . w3 xtion of 0.08 nM " - H epibatidine would occupy ap-proximately 65% of the high affinity sites, whereas lessthan 20% of the low affinity sites would be occupied. On
Ž . w3 xthe other hand, for the higher concentration of " - H ep-Ž .ibatidine 0.8 nM , almost all of the high affinity sites and
more than 60% of the low affinity sites would be occupied.Ž .Thus, the dissociation constants K determined for thei
Ž . w3 xvarious nicotinic analogs with 0.08 nM " - H epibati-dine would be a close approximation for their dissociationconstants for the high affinity site. In contrast, the K ’si
determined for the nicotinic analogs from the competitionŽ . w3 xbinding profiles in the presence of 0.8 nM of " - H epi-
batidine would reflect a composite of the dissociationconstants from the high and low affinity sites.
Ž . Ž . Ž .Fig. 4. Temporal analysis of pressor A , heart rate B and irritation CŽ . Ž .responses following i.t. injection of 0.5 mg of y -epibatidine. In B , the
open and hatched bars denote the tachycardia and bradycardia, respec-Ž .tively observed in the rats. Each value represents the mean"S.E. ns13 .
Table 2w3 xCompetition by ligands for H epibatidine binding sites in rat spinal cord membranes
Competing ligands Competition at the high affinity site Competition at both sites
Ž . Ž . Ž .IC nM K nM n IC nM n50 i H 50 H
Nicotinic agonistsŽ .y -Epibatidine 0.17"0.03 0.05"0.01 1.28"0.30 0.96"0.15 0.83"0.12Ž .q -Epibatidine 0.13"0.01 0.05"0.01 0.91"0.05 1.01"0.06 0.85"0.25Cytisine 1.45"0.19 0.56"0.11 0.57"0.13 18r22 0.7r0.6Ž .y -Nicotine 12.70"5.00 3.11"0.88 0.53"0.03 70r46 0.4r0.7Ž .q -Anatoxin 4.20"0.90 1.27"0.09 0.72"0.01 15.8r1.6 0.7"0.1Ž .q -Nicotine 115"17 43.4"6.20 0.80"0.15 562 NDNicotinic antagonistsa-Lobeline 8.8"1.0 2.2"0.3 0.8"0.03 121r316Dihydro-b-erythroidine 57.7r41.1 14.5r15.5 0.8r0.9 251r537Methyllycaconitine 385r411 ND )1000
Ž . w3 x Ž .Homogenates of spinal cord were incubated with 8–10 concentrations of each drug and 0.08 or 0.8 nM " - H epibatidine. Dissociation constants K ,iŽ . Ž .IC and Hill coefficients n were determined by non-linear regression analysis In-Plot . Dissociation constants for the competing ligands were50 H
Ž Ž . w3 x .determined using 0.08 nM " - H epibatidine only for site 1. Standard errors of the mean represent three to four measurements on separatepreparations. When the standard error is not shown, values for individual measurements are listed.

( )I.M. Khan et al.rBrain Research 753 1997 269–282274
Fig. 2 and Table 2 show the competition of variousŽ . w3 x Ž .nicotinic agonists for " - H epibatidine binding. y -
Epibatidine was the most potent of the agonists followedŽ .by cytisine, anatoxin and y -nicotine, respectively. Bind-
Ž .ing was stereospecific since q -nicotine was 14 timesw3 x Ž .less potent in inhibiting H epibatidine binding than y -
Ž . Ž .nicotine Fig. 2 and Table 2 . However, y -epibatidinedid not reveal a significant difference in potency in inhibit-
Ž . w3 x Ž .ing " - H epibatidine binding compared to q -epi-batidine. Hill coefficients for the various nicotinic agonists
Ž .were close to 1.0 except for q -anatoxin, cytisine andŽ .y -nicotine, even when the competition binding assays
Ž . w3 x Žwere conducted with 0.08 nM of " - H epibatidine Ta-.ble 2 .
( ) [ 3 ]3.3. Inhibition of " - H epibatidine binding by antago-nists
a-Lobeline was the most potent competitor among an-Ž .tagonists followed by dihydro-b-erthyroidine Db E and
Ž . Ž .methyllycaconitine MLA , respectively Table 2 . Al-though Db E acts as an antagonist in various brain regionsw x3,4,18,22,27 , a-lobeline is known to be a partial agonistw x1,7,24 . Methyllycaconitine, a potent antagonist for neu-
Ž . Ž . Ž . Ž . Ž . Ž .Fig. 5. Effect of repeated prior administrations i.t. of y -epibatidine 0.5 mg on i.t. administered cytisine A–C and y -nicotine D,E elicitedŽ . Ž .cardiovascular and behavioral responses. For the treatment sequence 1 open bars , the second dose of y -epibatidine was administered 35–40 min
Ž . Ž . Ž . Žfollowing the first dose. Then cytisine 5 mg or y -nicotine 5 mg was administered after another 35–40 min. Under treatment sequence 2 shaded. Ž . Ž . Ž .bars , a single dose of cytisine or y -nicotine 5 mg, each was administered in the rats without any prior treatment with y -epibatidine. The
cardiovascular and behavioral responses shown indicate the maximal values for each category of response following each dose of agonist administration.Each value represents the mean"S.E.; n indicates the number of animals in the treatment group.

( )I.M. Khan et al.rBrain Research 753 1997 269–282 275
ronal a-bungarotoxin binding receptors but a less potentw xinhibitor of nicotine binding to rat brain membranes 21,28 ,
Ž . w3 xweakly inhibited " - H epibatidine binding to spinalcord membranes.
Typical of neuronal nicotinic receptors, the binding ofŽ . w3 x" - H epibatidine was not blocked by a-bungarotoxinor a-conotoxin M1, suggesting that the spinal nicotinicreceptors were distinct from those found in muscle and the
Ž . Ž .Fig. 6. Effect of repeated prior administrations i.t. of cytisine 5 mg onŽ . Ž . Ž . Ž .i.t. y -Epibatidine 0.5 mg elicited pressor A , heart rate B and
Ž . Ž .irritation C responses. For the treatment sequence a1 open bars , thesecond dose of cytisine was administered 35–40 min following the first
Ž . Ž .dose. Then y -epibatidine 0.5 mg was administered after anotherŽ .35–40 min. Under treatment sequence a2 shaded bars , a single dose of
Ž . Ž .y -epibatidine 0.5 mg was administered in the rats without any priortreatment with cytisine. In B, the hatched bars denote the bradycardiaobserved in the rats. The cardiovascular and behavioral responses shownrepresent the maximal values for each category of response followingeach dose of agonist administration. Each value represents the mean"
S.E.; n indicates the number of animals in the treatment group.
Ža and a subtypes identified in neuronal tissues data not7 8.shown .
( )3.4. Responses to i.t. y -epibatidine
Ž .We have previously demonstrated that i.t. y -epibati-dine elicits dose-dependent increases in blood pressure,heart rate and behavioral pain responses similar to the
Ž . w xspinal nicotinic agonists, cytisine and y -nicotine 13 .Moreover, rank-ordering of potencies for the various spinalnicotinic agonists in manifesting the specific responses
Ž .show y -epibatidine to be the most potent nicotinicw xagonist 13 .
Fig. 4 shows the various cardiovascular and behavioralresponses at intervals of 0–3, 3–4, 6–7, and 14–15 min
Ž .following i.t. administration of 0.5 mg y -epibatidine.The onsets of the pressor, heart rate and irritation re-sponses were very rapid. Within 3–5 min from the initia-tion of the responses, the irritation response disappearedand simultaneously the tachycardia reverted to a bradycar-
Ž .dia Fig. 4b . Yet, in a significant number of rats, onset ofbradycardia appeared within 2–3 min. Maximal bradycar-
Ž .dia was observed around 7 min after y -epibatidineadministration, a time when the irritation response wasvirtually absent. The pressor response diminished consider-ably with time but remained above the base-line for 15–20min. By 15–25 min an intermittent irritation responsereappeared. By this time, the bradycardia began to disap-pear and the pressor response almost returned to base-lineor remained elevated by 10–15 mmHg. These transientincreases in blood pressure corresponded to the time courseof the intermittent irritation response. All the responsesreturned to base-line by 25–35 min.
3.5. Desensitization and cross-desensitization
Ž . Ž .Prior administration of y -epibatidine 0.5 mg, i.t.resulted in significant inhibition of the pressor, tachycardia
Žand the irritation responses to subsequent doses within. Ž . Ž .35–40 min of y -epibatidine Fig. 5a–f . However, the
bradycardia was not affected. The cardiovascular and be-Ž .havioral responses to i.t. cytisine and y -nicotine admin-
istrations following the second dose of epibatidine werealso reduced when compared to the responses to i.t. cyti-
Ž . Žsine and y -nicotine in untreated control rats Fig. 5a–.c,e,f, shaded bars . Interestingly, in the rats previously
Ž .treated with two sequential doses of y -epibatidine fol-Ž .lowed by a dose of cytisine, an additional dose of y -epi-
Ž .batidine 0.5 mg; 105–110 min exhibited responses ofŽ .similar magnitude to those for the second dose of y -epi-
Ž .batidine data not shown . Thus, even though the desensiti-zation of epibatidine’s responses following repeated dosingwas significant compared to the first dose, the residual
Ž .responses to y -epibatidine did not decrease further.Desensitization of the responses induced by repeated ad-
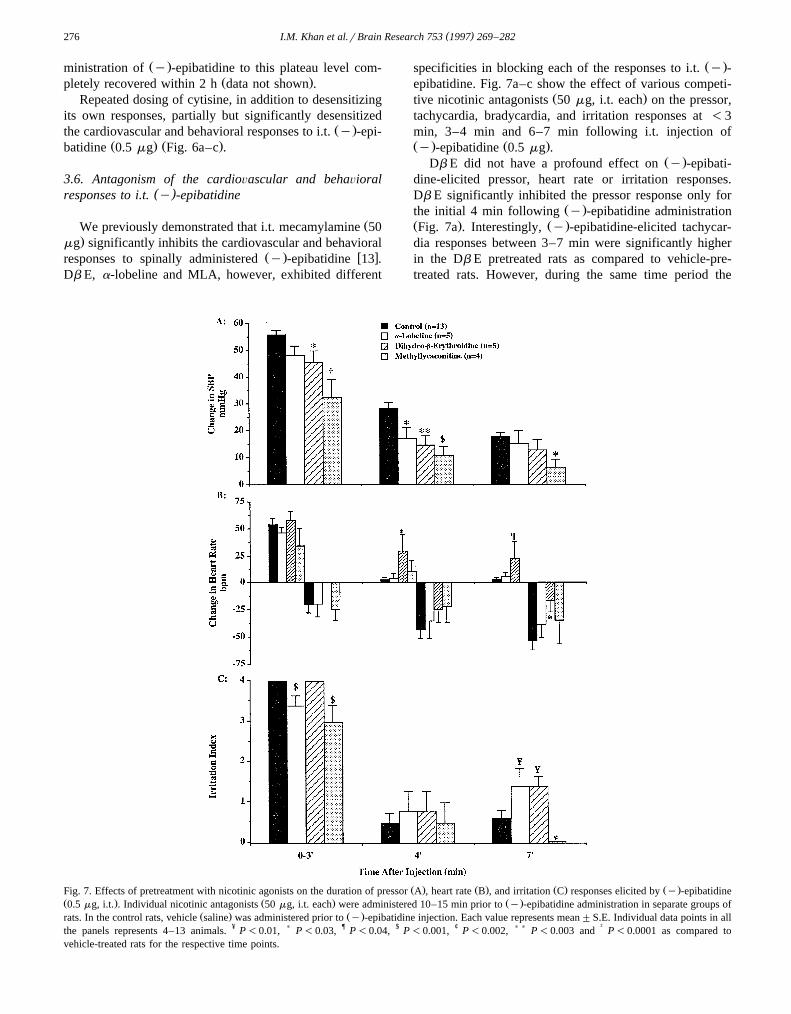
( )I.M. Khan et al.rBrain Research 753 1997 269–282276
Ž .ministration of y -epibatidine to this plateau level com-Ž .pletely recovered within 2 h data not shown .
Repeated dosing of cytisine, in addition to desensitizingits own responses, partially but significantly desensitized
Ž .the cardiovascular and behavioral responses to i.t. y -epi-Ž . Ž .batidine 0.5 mg Fig. 6a–c .
3.6. Antagonism of the cardioÕascular and behaÕioral( )responses to i.t. y -epibatidine
ŽWe previously demonstrated that i.t. mecamylamine 50.mg significantly inhibits the cardiovascular and behavioral
Ž . w xresponses to spinally administered y -epibatidine 13 .Db E, a-lobeline and MLA, however, exhibited different
Ž .specificities in blocking each of the responses to i.t. y -epibatidine. Fig. 7a–c show the effect of various competi-
Ž .tive nicotinic antagonists 50 mg, i.t. each on the pressor,tachycardia, bradycardia, and irritation responses at -3min, 3–4 min and 6–7 min following i.t. injection ofŽ . Ž .y -epibatidine 0.5 mg .
Ž .Db E did not have a profound effect on y -epibati-dine-elicited pressor, heart rate or irritation responses.Db E significantly inhibited the pressor response only for
Ž .the initial 4 min following y -epibatidine administrationŽ . Ž .Fig. 7a . Interestingly, y -epibatidine-elicited tachycar-dia responses between 3–7 min were significantly higherin the Db E pretreated rats as compared to vehicle-pre-treated rats. However, during the same time period the
Ž . Ž . Ž . Ž .Fig. 7. Effects of pretreatment with nicotinic agonists on the duration of pressor A , heart rate B , and irritation C responses elicited by y -epibatidineŽ . Ž . Ž .0.5 mg, i.t. . Individual nicotinic antagonists 50 mg, i.t. each were administered 10–15 min prior to y -epibatidine administration in separate groups of
Ž . Ž .rats. In the control rats, vehicle saline was administered prior to y -epibatidine injection. Each value represents mean"S.E. Individual data points in allthe panels represents 4–13 animals. ¥ P-0.01, ) P-0.03, ¶ P-0.04, $ P-0.001, ¢ P-0.002, ) ) P-0.003 and † P-0.0001 as compared tovehicle-treated rats for the respective time points.
( )I.M. Khan et al.rBrain Research 753 1997 269–282 277
Ž .bradycardia response to y -epibatidine was diminished inthe antagonist pretreated rats.
Ž .Although a-lobeline significantly inhibits spinal y -w xepibatidine-elicited analgesic response 9 , the antagonist
showed no effect on the heart rate responses and onlyminor inhibition of the pressor and irritation responses
Ž .elicited by i.t. y -epibatidine. Of the antagonists onlya-lobeline exerted a perceptible response at the dosesemployed, and its irritation response is likely to arise fromits partial agonist characteristics.
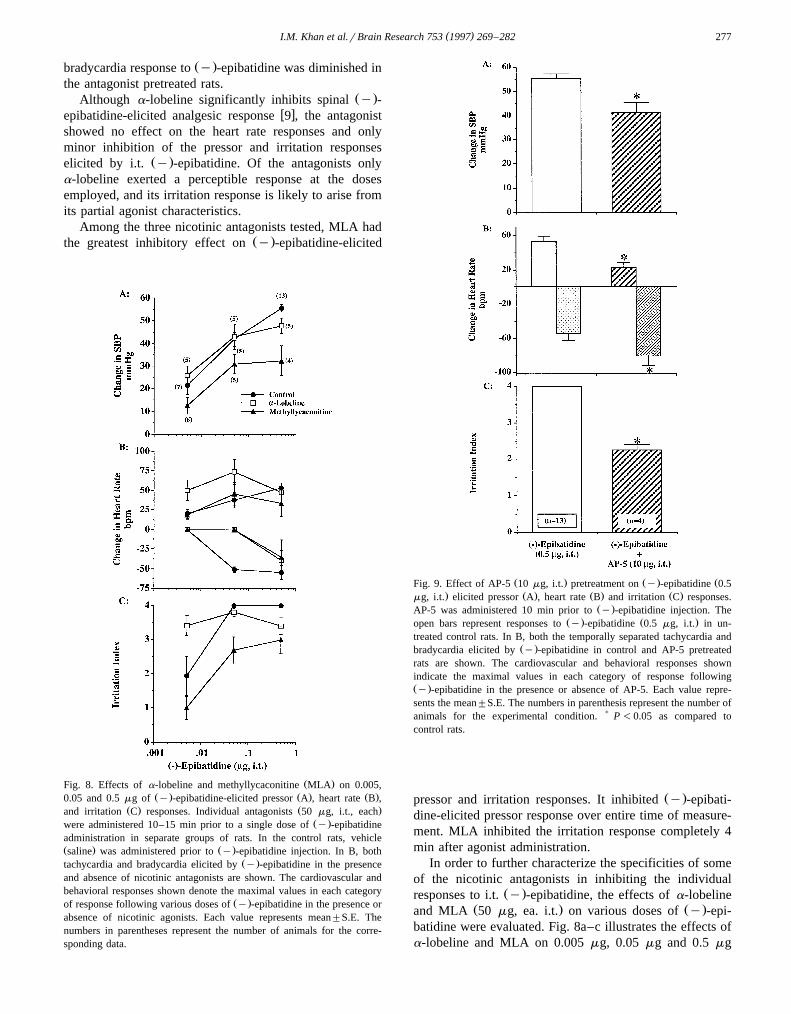
Among the three nicotinic antagonists tested, MLA hadŽ .the greatest inhibitory effect on y -epibatidine-elicited
Ž .Fig. 8. Effects of a-lobeline and methyllycaconitine MLA on 0.005,Ž . Ž . Ž .0.05 and 0.5 mg of y -epibatidine-elicited pressor A , heart rate B ,
Ž . Ž .and irritation C responses. Individual antagonists 50 mg, i.t., eachŽ .were administered 10–15 min prior to a single dose of y -epibatidine
administration in separate groups of rats. In the control rats, vehicleŽ . Ž .saline was administered prior to y -epibatidine injection. In B, both
Ž .tachycardia and bradycardia elicited by y -epibatidine in the presenceand absence of nicotinic antagonists are shown. The cardiovascular andbehavioral responses shown denote the maximal values in each category
Ž .of response following various doses of y -epibatidine in the presence orabsence of nicotinic agonists. Each value represents mean"S.E. Thenumbers in parentheses represent the number of animals for the corre-sponding data.
Ž . Ž . ŽFig. 9. Effect of AP-5 10 mg, i.t. pretreatment on y -epibatidine 0.5. Ž . Ž . Ž .mg, i.t. elicited pressor A , heart rate B and irritation C responses.
Ž .AP-5 was administered 10 min prior to y -epibatidine injection. TheŽ . Ž .open bars represent responses to y -epibatidine 0.5 mg, i.t. in un-
treated control rats. In B, both the temporally separated tachycardia andŽ .bradycardia elicited by y -epibatidine in control and AP-5 pretreated
rats are shown. The cardiovascular and behavioral responses shownindicate the maximal values in each category of response followingŽ .y -epibatidine in the presence or absence of AP-5. Each value repre-sents the mean"S.E. The numbers in parenthesis represent the number ofanimals for the experimental condition. ) P -0.05 as compared tocontrol rats.
Ž .pressor and irritation responses. It inhibited y -epibati-dine-elicited pressor response over entire time of measure-ment. MLA inhibited the irritation response completely 4min after agonist administration.
In order to further characterize the specificities of someof the nicotinic antagonists in inhibiting the individual
Ž .responses to i.t. y -epibatidine, the effects of a-lobelineŽ . Ž .and MLA 50 mg, ea. i.t. on various doses of y -epi-
batidine were evaluated. Fig. 8a–c illustrates the effects ofa-lobeline and MLA on 0.005 mg, 0.05 mg and 0.5 mg

( )I.M. Khan et al.rBrain Research 753 1997 269–282278
Ž .y -epibatidine elicited cardiovascular and behavioral re-sponses.
a-Lobeline did not inhibit the pressor responses to anyŽ .dose of y -epibatidine. a-Lobeline significantly inhibited
Ž .the irritation response to the highest dose of y -epibati-dine, but did not block the irritation response to 0.05 mgŽ .y -epibatidine. Interestingly, irritation response to 0.005
Ž .mg y -epibatidine was significantly higher in the antago-nist pretreated rats as compared to vehicle controls. Similar
Ž . Žto the irritation response, y -epibatidine-elicited 0.005.mg tachycardia was higher in a-lobeline pretreated rats.
Although a-lobeline did not inhibit the bradycardia elicitedŽ .by 0.5 mg of y -epibatidine, it completely abolished the
Ž .bradycardia elicited by 0.05 mg y -epibatidine.In contrast to a-lobeline, MLA significantly inhibited
Ž .the pressor response to all three doses of i.t. y -epibati-Ž .dine Fig. 9a–d . MLA also significantly inhibited the
irritation response to 0.05 mg and 0.5 mg epibatidine.Changes in heart rate elicited by 0.05 and 0.005 mgŽ .y -epibatidine were not inhibited by MLA pretreatment.Analogous to a-lobeline, MLA significantly blocked the
Ž .bradycardia elicited by 0.05 mg of y -epibatidine.
( )3.7. Antagonism of y -epibatidine-elicited responses byi.t. NMDA receptor antagonist
Ž .As previously described, i.t. y -epibatidine elicits adose-dependent release of EAA in spinal microdialysatesw x13 . In addition, the cardiovascular and irritation re-sponses as well as the spinal release of EAA elicited by i.t.Ž . w xy -epibatidine are inhibited by mecamylamine 13 . Thissuggested that a significant portion of the cardiovascularand irritation responses elicited by spinal epibatidine maybe mediated by the released EAA acting via the glutamatereceptors. Accordingly, we determined the effect of AP-5Ž . Ž .NMDA-receptor antagonist on y -epibatidine-elicited
Ž .cardiovascular and behavioral responses. AP-5 10 mg, i.t.significantly inhibited the initial pressor, tachycardia and
Ž .irritation responses to epibatidine 0.5 mg, P-0.05 . In-terestingly, the bradycardia to epibatidine was significantly
Žgreater in AP-5 versus vehicle pretreated control rats Fig..9a–c .
4. Discussion
Two classes of binding sites differing in affinities andtotal number of binding sites can be identified in the spinal
Ž . w3 xcord membranes for " - H epibatidine. To date, fewstudies have been directed to characterizing spinal nico-
w xtinic receptors and delineating their responses 11 . Previ-ously, we demonstrated a single class of high affinity
w3 x w3 xbinding sites for H cytisine and H nicotine in rat spinalcord membranes and characterized the sites with respect to
w xtheir pharmacological specificities 15,16 .
[ 3 ]4.1. H Epibatidine binding in the central nerÕous system
w3 xIn rat brain, H epibatidine also reveals two classes ofbinding sites in equal proportions but differing in their
w x w xaffinities 12 . Perry and Kellar 23 , using in vitro receptorw3 xautoradiographic techniques, demonstrated specific H ep-
ibatidine binding to various regions in rat brain. As com-w3 x w3 xpared to H cytisine, H epibatidine binding is found to
be greater in several brain regions. The authors concludedw3 xthat the differences in labelling between H cytisine and
w3 x w3 xH epibatidine could arise from the ability of H epi-batidine to label more than a single subtype of neuronal
Ž . w3 xnicotinic receptors. In our studies, " - H epibatidine inrat spinal cord membranes exhibits binding to at least twoindependent receptor sites; the ratio of high affinity to lowaffinity binding sites is 2:1. The K for the high affinityd
site in the rat spinal cord is slightly greater than that foundŽ .in the rat brain 0.046 nM vs. 0.015 nM, respectively .
ŽThe total number of binding sites low and high affin-. Ž . w3 xity estimated from " - H epibatidine binding to spinal
cord membranes is greater than the maximum number ofw3 x w3 xbinding sites estimated from H cytisine or H nicotine
w xbinding to spinal cord membranes 16 . The lower non-Ž . w3 x w3 xspecific binding of " - H epibatidine versus H cyti-
w3 x Ž . w3 xsine or H nicotine, and the higher affinity of " - H ep-ibatidine for spinal nicotinic receptors may allow detectionof the lower affinity receptors. Interestingly, the maximumnumber of binding sites for the higher affinity sites forŽ . w3 x" - H epibatidine is roughly similar to the maximum
w3 xnumber of binding sites detected by H cytisine orw3 xH nicotine.
4.2. Distribution of nicotinic binding sites in spinal cord— physiological implications
w3 x w3 x w xSimilar to H cytisine and H nicotine binding 16 ,Ž . w3 x" - H epibatidine binding to the dorsal lumbarosacral
Ž . w3 xregion showed the highest density of " - H epibatidinebinding sites as compared to those in the thoracic orventral lumbarosacral regions. While a ratio of 2:1 for thehigh:low affinity receptor numbers was found in the tho-racic and ventral lumbarosacral regions, in the dorsallumbarosacral region the ratio was 3:1. Autoradiographic
Ž . w3 xstudies demonstrate a high density of " - H epibatidinebinding in the superficial layer of dorsal lumbar and
w x Ž . w3 xthoracic spinal cord 14 . In addition, specific " - H epi-batidine binding can also be observed in the intermediolat-eral cell column region of the thoracic spinal cord. Most ofthe binding in laminae I and II appears to be on smallprimary afferents, since capsaicin pretreatment signifi-
Ž . w3 xcantly inhibits " - H epibatidine binding in this regionŽ .Khan et al. unpublished observations .
w xBased on our previous reports 13–17 , nicotinic recep-tors in the various spinal regions appear to be involved inmediating cardiovascular and behavioral responses to in-

( )I.M. Khan et al.rBrain Research 753 1997 269–282 279
trathecal nicotinic agonists. Accordingly, the potency inbinding to spinal nicotinic receptors for epibatidine corre-lates with its potencies in eliciting the cardiovascular andbehavioral responses following spinal administration. In-
Ž .trathecal y -epibatidine also elicits profound but short-lasting analgesia in rats and appears to be the most potent
w xof the agonists in producing this response 9 . However,the concentration of epibatidine to produce the analgesicresponse is higher than that for eliciting the cardiovascularand nociceptive responses. Thus, we might speculate that asignificant portion of the low affinity binding sites forŽ . w3 x" - H epibatidine represent the analgesic receptor forthis nicotinic agonist.
Ž .An interesting observation with i.t. y -epibatidine isthe reversal of tachycardia to bradycardia accompanyingthe disappearance of the irritation response, 3–10 minpost-injection. This period of bradycardia correlates tem-porally with epibatidine analgesia to thermal stimulation in
w xrats 9 . Tachycardia following nicotinic agonists is mostlyw xassociated with the irritation response 13,15,16 . Thus,
Ž .during the period of analgesia elicited by i.t. y -epibati-dine, when the irritation response is virtually absent, the
Ž .increased sustained pressor response results in activationof the baroreceptor reflex and the corresponding bradycar-dia.
( ) [ 3 ]4.3. Specificity of spinal " - H epibatidine binding sites
4.3.1. Agonist specificitiesŽThe rank order of potencies inverse of the dissociation
.constants, K for the agonists in binding closely correlatesi
with their rank-order in eliciting cardiovascular and behav-ioral responses when administered intrathecally. In studiesin vivo with neuronal nicotinic receptors, epibatidine has
w xbeen shown to be the most potent agonist 2,5,8,26 .Likewise, the rank order of potencies of most other ago-
Ž . w3 xnists in competing with " - H epibatidine binding inspinal cord membranes was roughly comparable to thosefound in other investigations of nicotinic receptors in brainw x12 .
Ž . w3 xAt the higher concentrations of " - H epibatidine,where epibatidine binds to both the high and low affinityspinal nicotinic receptors, analysis of the competition bind-
Ž . w3 xing profiles of " - H epibatidine with various nicotinicanalogs can be complicated by the contribution of the twoindependent sites on the overall binding. Thus, in thepresent study, the K ’s for the various nicotinic analogsi
Ž .were determined at low concentration ;0.08 nM ofŽ . w3 x" - H epibatidine, conditions which should minimizethe influence of low affinity sites. The K ’s determined fori
Ž . w3 xthe various nicotinic agonists in inhibiting " - H epi-batidine binding from the high affinity sites were similar to
w3 xthose for the nicotinic agonists in displacing H cytisinew3 xor H nicotine binding from spinal cord nicotinic recep-
w xtors 16 .Ž .It is noteworthy that q -anatoxin was more potent
Ž .than y -nicotine and almost equipotent as cytisine inw3 x Ž .displacing H epibatidine binding. Spinal q -anatoxin
Ž .appears more potent than y -nicotine in eliciting cardio-vascular and irritation responses and similar in potency to
Ž .i.t. cytisine. q -Anatoxin also elicits desensitization uponŽrepeated i.t. administration Khan et al., unpublished ob-
.servations .Hill slopes for the various nicotinic agonists including
Ž . Ž .q -anatoxin, y -nicotine and cytisine were less thanone revealing competition at both classes of epibatidinebinding sites. Although a Hill slope of less than one forcytisine and nicotine in competition binding assays at 0.8
Ž . w3 xnM " - H epibatidine is expected, Hill slopes of lessthan one at 0.08 nM indicate that these nicotinic agonists
Ž . w3 xmay identify more than one high affinity " - H epi-batidine binding sites. One interpretation for this may bethat the presumed two binding sites in the pentamericnicotinic receptor may have different affinities for nico-tinic agonists other than epibatidine. Alternatively, theremay be more than one class of the higher affinity spinalnicotinic receptors which show selectivity for the nicotinicagonists other than epibatidine.
Ž .Repeated cytisine administrations i.t. elicit near com-plete desensitization to its own responses and that of most
w xother agonists 15 . Yet, cytisine only partially desensitizesŽ .the cardiovascular and behavioral responses to i.t. y -epi-
Ž .batidine. However, repeated y -epibatidine administra-tion desensitizes the responses to cytisine and nicotine, butonly partially desensitizes its own responses. Similar to
Ž .cytisine, q -anatoxin desensitizes its own responses andto cytisine’s responses, and it only partially desensitizesepibatidine’s cardiovascular and behavioral responsesŽ .Khan et al., unpublished observations . Thus, the higher
Ž . w3 xaffinity class of receptors for " - H epibatidine not onlydiffers in selectivity for the nicotinic agonists, they alsohave different desensitization properties. A subset of recep-tors for epibatidine remain refractory to desensitization byall other agonists studied. Recent studies show that toler-ance to the epibatidine enantiomers may differ after chronic
Ž . Ž . w xtreatment with y - and q -epibatidine 10 .It is also apparent from the present study that the
various nicotinic agonists are less potent than epibatidinefor the low affinity epibatidine site. Repeated administra-
Ž .tion of y -epibatidine also desensitizes the analgesicw xresponse 9 . Although a single administration of cytisine
Ž . Ž .5 mg, i.t. desensitizes y -epibatidine’s irritation re-sponse, it does not desensitize the analgesic response toŽ .y -epibatidine. Yet, repeated administrations of cytisine
Ž .do inhibit y -epibatidine’s analgesic response indicatingminimal cytisine interaction with the low affinity nicotinic
w xreceptors presumably mediating analgesia 9 .
4.3.2. Antagonist specificitiesThe rank order of potencies of the three competitive
Ž . w3 xantagonists examined in displacing " - H epibatidinebinding from spinal cord membranes is a-lobeline)Db E

( )I.M. Khan et al.rBrain Research 753 1997 269–282280
4MLA. However, Db E and a-lobeline are equipotent inw3 x w3 x w xdisplacing H cytisine or H nicotine binding 16 . Simi-
lar to several other nicotinic analogs, both a-lobeline andDb E exhibit Hill slopes of less than one in displacement
w3 xat both concentrations of H epibatidine.Table 3 summarizes the antagonistic effect of a-lobe-
Ž .line, Db E and MLA on y -epibatidine, cytisine andŽ .y -nicotine elicited cardiovascular and behavioral re-sponses. Only a-lobeline is selective in blocking the anal-gesic response to spinal epibatidine. In contrast, it isvirtually ineffective in blocking the cardiovascular re-sponses to epibatidine. a-Lobeline is less effective ininhibiting epibatidine-elicited irritation response as com-pared to blocking the irritation response to nicotine. Yet, ithas lower potency in inhibiting the irritation response tonicotine than the pressor response to nicotine. a-Lobelinedoes not antagonize the irritation response to cytisine. Thecomplex pharmacological profile for a-lobeline may beexplained on the basis of a-lobeline having both agonistand antagonistic actions on neuronal nicotinic receptorsw x1,7,15,24 . This dual action complicates the estimation ofits antagonist potency. Thus, the above observations, to-gether with a Hill slope of less than unity for a-lobeline
Ž . w3 xdisplacement of " - H epibatidine binding, indicate thatthe antagonist interacts with a significant portion of thehigh affinity epibatidine binding sites as an agonist ineliciting the irritation response. In addition, it behaves as apartial agonist on the other receptors involved in mediatingcardiovascular and irritation responses and as an antagonist
Ž .for the analgesic response to y -epibatidine. Interest-Ž .ingly, a-lobeline 50 mg, i.t. augments the irritation re-
Ž . Ž .sponse to low doses of y -epibatidine 0.005 mg . This is
consistent with the fact that a-lobeline selectively antago-nizes the analgesic response to epibatidine, and epibatinehas a lower affinity for the analgesic receptors than the
w xirritation receptors 9 .Db E significantly inhibits the pressor, heart rate and
Ž .irritation responses to i.t. y -nicotine, but is largelyineffective in inhibiting these responses to i.t. cytisine orŽ . Ž .y -epibatidine Table 3 . Db E does not inhibit the initial
Ž .irritation response or heart rate increase to y -epibati-dine; however, it inhibits the pressor response for 4 minfollowing epibatidine injection. It also does not inhibit the
Ž .analgesic response to y -epibatidine in the thermal es-w xcape model 9 , yet it has been reported previously that
Ž .Db E can inhibit epibatidine s.c. elicited analgesia in thew xhot-plate antinociceptive assay 6 . Db E is equipotent to
w3 x w3 xa-lobeline in displacing either H cytisine or H nicotinebinding; however, it is less potent than a-lobeline in
Ž . w3 xinhibiting " - H epibatidine binding from spinal cordmembranes. The limited efficacies of Db E in antagonizing
Ž .the various responses to y -epibatidine or cytisine aswell as in competing for the binding of the radiolabelednicotinic agonists is a distinguishing characteristic of thespinal nicotinic receptors.
Among the three antagonists, MLA was most potent ininhibiting the pressor and irritation responses to all thenicotinic agonists examined. However, MLA exhibited the
Ž . w 3 xlowest potency in inhibiting " - H epibatidine,w3 x w3 xH cytisine or H nicotine binding to spinal cord mem-
w xbranes. As described previously 15,16 , the divergence inconcentrations found in the pharmacologic response andbinding experiments might indicate that MLA has channelblocking activity on nicotinic binding sites in the spinal
Table 3Ž .Summary of a-lobeline, dihydro-b-erythroidine and methyllycaconitine 50 mg, i.t., each antagonism of the cardiovascular and behavioral responses
Ž . Ž .elicited by spinal y -epibatidine, cytisine or y -nicotinea aŽ . Ž .y -Epibatidine Cytisine y -Nicotine
a-LobelinePressor 0 q qqq
Ž .Heart rate tachycardia 0 q qqqIrritation q 0 qqAnalgesia qq NT NTDb EPressor q q qqq
Ž .Heart rate tachycardia 0 q qqqIrritation 0 0 qqqAnalgesia 0 NT NTMLAPressor qq qq qq
Ž .Heart rate tachycardia 0 qq qqqIrritation qq qq qqqAnalgesia 0 NT NT
NT, not tested.a w xData from 15 .
Ž . ŽRats were treated with the respective antagonists. Ten min following antagonist administration, near maximal doses for y -epibatidine 0.5 mg for. Ž . Ž . Ž . Ž .cardiovascular and irritation responses, 1.0 mg for analgesia , cytisine 5.0 mg and y -nicotine 5.0 mg were used and initial responses -3 min to the
agonist were measured, except for analgesia where measurements were taken after 5 min. Antagonist scoring: 0, no antagonism; q, 10–20% reduction;qq, 20–50% reduction; qqq, )50% reduction.

( )I.M. Khan et al.rBrain Research 753 1997 269–282 281
cord. Yet, MLA displays selectivity in its antagonisticproperties by not blocking the analgesic response to spinalepibatidine.
4.4. Role of EAA in epibatidine-elicited cardioÕascularand behaÕioral responses
Ž .We have previously demonstrated that i.t. y -epibati-dine is the most potent of the agonists in eliciting dose-de-pendent increases in Asp and Glu release in spinal micro-
w xdialysates in rats 13 . Intrathecal AP-5 significantly inhib-ited epibatidine elicited pressor, tachycardia and irritationresponses. The data indicate that a significant portion ofthe cardiovascular and pain responses elicited by spinalepibatidine is mediated by the EAA release. Interestingly,
Ž .the bradycardia response to y -epibatidine was potenti-ated by the NMDA receptor antagonist. This may indicatethat the irritation and the analgesic responses to spinalepibatidine are mediated by activation of opposing neu-ronal pathways.
In summary, the data indicate that spinal epibatidine,similar to other nicotinic agonists, elicits dose-dependentincreases in blood pressure, heart rate and pain responses.
Ž .In addition, y -epibatidine also produces a subsequentbradycardia which temporally coincides with the period ofanalgesia elicited by the nicotinic agonist. The rank order-ing of the potencies of the the nicotinic agonists in elicit-ing the cardiovascular, behavioral and amino acid releaseend points correlates with the binding characteristics of thenicotinic agonists to spinal nicotinic receptors. The differ-
Ž .ence in potencies of y -epibatidine in eliciting the irrita-tion and analgesic responses and the selective blockade ofthe analgesic and pain responses by a-lobeline and MLAor AP-5, respectively support the contention that epibati-dine activates distinct subtypes of nicotinic receptors inspinal cord. These receptors might be expected to bepresynaptic and control the release of other transmitterssuch as the excitatory amino acids.
Acknowledgements
Supported by USPHS Grant HL 35018 and the Univer-sity of California Tobacco-Related Disease Research Pro-gram to P.T.
References
w x1 Abood, L.G., Salles, K.S. and Maiti, A., Structure activity studies ofcarbamate and other esters: agonists and antagonists to nicotine,
Ž .Pharmacol. Biochem. BehaÕ., 30 1988 403–408.w x2 Alkondon, M. and Albuquerque, E.X., Diversity of nicotinic acetyl-
choline receptors in rat hippocampal neurons. III. Agonist actions ofthe novel alkaloid epibatidine and analysis of type II current, J.
Ž .Pharmacol. Exp. Ther., 274 1995 771–782.w x3 Alkondon, M. and Albuquerque, E.X., Initial characterization of the
nicotinic acetylcholine receptors in rat hippocampal neurons, J.Ž .Recep. Res., 11 1991 1001–1021.
w x4 Araujo, D.M., Lapchak, P.A., Collier, B. and Quirion, R. Characteri-w xzation of N- 3H -methylcarbamylcholine binding sites and effect of
N-methylcarbamylcholine on acetylcholine release in rat brain, J.Ž .Neurochem., 51 1988 292–299.
w x5 Badio, B. and Daly, J.W., Epibatidine, a potent analgesic andŽ .nicotinic agonist, Mol. Pharmacol., 45 1994 563–569.
w x6 Badio, B., Shi, D., Garraffo, M. and Daly, J.W., Antinociceptiveeffects of the alkaloid epibatidine: further studies on involvement of
Ž .nicotinic receptors, Drug DeÕ. Res., 36 1995 46–59.w x7 Barlow, R.B. and Johnson, O., Relations between structure and
Ž .nicotine-like activity: X-ray crystal structure and analysis of y -Ž .cytisine and y -lobeline hydrochloride and a comparison with
Ž .y -nicotine and other nicotine-like compounds, Br. J. Pharmacol.,Ž .98 1989 799–808.
w x8 Bonhaus, D.W., Bley, K.R., Broka, C.A., Fontana, D.J., Leung, E.,Lewis, R., Shieh, A. and Wong, E.H.F., Characterization of theelectrophysiological, biochemical and behavioral actions of epibati-
Ž .dine, J. Pharmacol. Exp. Ther., 272 1995 1199–1203.w x9 Buerkle, H., Khan, I.M., Taylor, P. and Yaksh, T.L., Nociceptive
and antinociceptive responses to intrathecally administered epibati-Ž .dine. Submitted to Neuropharmacology, 1996 .
w x10 Damaj, M.I. and Martin, B.R., Tolerance to the antinociceptiveeffect of epibatidine after acute and chronic administration in mice,
Ž .Eur. J. Pharmacol., 300 1996 51–57.w x11 Gillberg, P.-G. and Askmark, H., Changes in cholinergic and opioid
receptors in the rat spinal cord, dorsal root and sciatic nerve afterŽ .ventral and dorsal root lesion, J. Neural Transm., 85 1991 31–39.
w x12 Houghtling, R.A., Davila-Garcia, M.I. and Kellar, K.J., Characteri-Ž . w xzation of " - 3H -epibatidine binding to nicotinic cholinergic re-
Ž .ceptors in rat and human brain, Mol. Pharmacol., 48 1995 280–287.
w x13 Khan, I.M., Marsala, M., Printz, M.P., Taylor, P. and Yaksh, T.L.,Intrathecal nicotinic agonist-elicited release of excitatory aminoacids as measured by in vivo spinal microdialysis in rats, J. Phar-
Ž .macol. Exp. Ther., 278 1996 97–106.w x14 Khan, I.M., Youngblood, K.L., Yaksh, T.L., Printz, M.P. and Tay-
lor, P., Regulation of spinal nicotinic receptors in spontaneouslyŽ .hypertensive rats, Hypertension, 28 1996 1093–1098.
w x15 Khan, I.M., Taylor, P. and Yaksh, T.L., Cardiovascular and behav-ioral responses to nicotinic agonists administered intrathecally, J.
Ž .Pharmacol. Exp. Ther., 270 1994 150–158.w x16 Khan, I.M., Yaksh, T.L. and Taylor, P., Ligand specificity of
nicotinic acetylcholine receptors in rats spinal cord: studies withŽ .nicotine and cytisine, J. Pharmacol. Exp. Ther., 270 1994 159–
166.w x17 Khan, I.M., Taylor, P. and Yaksh, T.L., Stimulatory pathways and
sites of actions of intrathecally administered nicotinic agonists, J.Ž .Pharmacol. Exp. Ther., 271 1994 1550–1557.
w x18 Lapchak, P.A., Araujo, D.M., Quirion, R. and Collier, B., Presynap-tic cholinergic mechanisms in the rat cerebellum: evidence for
Ž .nicotinic, but not muscarinic autoreceptors, J. Neurochem., 53 19891843–1851.
w x19 Lindstrom, J., Anand, R., Peng, X., Gerzanich, V., Wang, F. and Li,Y., Neuronal nicotinic receptor subtypes, Ann. NY Acad. Sci., 757Ž .1995 100–119.
w x20 Lukas, R.J., Diversity and patterns of regulation of nicotinic receptorŽ .subtypes, Ann. NY Acad. Sci., 757 1995 151–168.
w x21 Macallan, D.R.E., Lunt, G.G., Wonnacott, S., Swanson, K.L., Rap-Ž .paport, H. and Albuquerque, E.X., Methyllycaconitine and q -
anatoxin-a differentiate between nicotinic receptors in vertebrate andŽ .invertebrate nervous system, FEBS Lett., 226 1988 357–363.
w x22 Mulle, C., Vidal, C., Benoit, P. and Changeux, J.P., Existence ofdifferent subtypes of nicotinic acetylcholine receptors in the rat
Ž .habenulo-interpeduncular system, J. Neurosci., 11 1991 2588–2597.

( )I.M. Khan et al.rBrain Research 753 1997 269–282282
w x w3 x23 Perry, D.C. and Kellar, K.J., H Epibatidine labels nicotinic recep-tors in rat brain: an autoradiographic study, J. Pharmacol. Exp.
Ž .Ther., 275 1995 1030–1034.w x24 Reavill, C., Walther, B., Stolerman, I.P. and Testa, B., Behavioral
and pharmacokinetic studies on nicotine, cytisine and lobeline,Ž .Neuropharmacology 29 1990 619–624.
w x25 Spande, T.F., Garrafo, H.M., Edwards, M.W., Yeh, H.J.C., Pannel,Ž .L. and Daly, J.W., Epibatidine: a novel chloropyridyl azabicyclo-
heptane with potent analgesic activity from a Ecuadorian poisonŽ .frog, J. Am. Chem. Soc., 114 1992 3475–3480.
w x26 Sullivan, J.P., Decker, M.W., Brioni, J.D., Donnelly-Roberts, D.,
Anderson, D.J., Bannon, A.W., Kang, C.-H., Adams, P., Piattoni-Kaplan, M., Buckley, M.J., Gopalakrishnan, M, Williams, M. and
Ž .Arneric, S.P., " -Epibatidine elicits a diversity of in vitro and invivo effects mediated by nicotinic acetylcholine receptors, J. Phar-
Ž .macol. Exp. Ther., 271 1994 624–631.w x27 Vidal, C. and Changeux, J.-P., Pharmacological profile of nicotinic
acetylcholine receptors in rat pre-frontal cortex: an electrophysiolog-Ž .ical study in slice preparation, Neuroscience 29 1989 261–270.
w x28 Ward, J.M., Cockcroft, V.B., Lunt, G.G., Smillie, F.S. and Wonna-cott, S., Methyllycaconitine: a selective probe for neuronal a-
Ž .bungarotoxin binding sites, FEBS Lett., 270 1990 45–48.
Copyright © 2022 FDOKUMEN




















