Ephrin B2/EphB4 pathway in hepatic stellate cells stimulates Erk-dependent VEGF production and...
9
doi:10.1152/ajpgi.00510.2009 298:908-915, 2010. First published Mar 25, 2010; Am J Physiol Gastrointest Liver Physiol Debabrata Mukhopadhyay and Vijay H. Shah Amitava Das, Uday Shergill, Lokendra Thakur, Sutapa Sinha, Raul Urrutia, You might find this additional information useful... 41 articles, 19 of which you can access free at: This article cites http://ajpgi.physiology.org/cgi/content/full/298/6/G908#BIBL including high-resolution figures, can be found at: Updated information and services http://ajpgi.physiology.org/cgi/content/full/298/6/G908 can be found at: AJP - Gastrointestinal and Liver Physiology about Additional material and information http://www.the-aps.org/publications/ajpgi This information is current as of May 18, 2010 . http://www.the-aps.org/. Society. ISSN: 0193-1857, ESSN: 1522-1547. Visit our website at American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the American Physiological abnormal function of the gastrointestinal tract, hepatobiliary system, and pancreas. It is published 12 times a year (monthly) by the publishes original articles pertaining to all aspects of research involving normal or AJP - Gastrointestinal and Liver Physiology on May 18, 2010 ajpgi.physiology.org Downloaded from
Transcript of Ephrin B2/EphB4 pathway in hepatic stellate cells stimulates Erk-dependent VEGF production and...

doi:10.1152/ajpgi.00510.2009 298:908-915, 2010. First published Mar 25, 2010;Am J Physiol Gastrointest Liver Physiol
Debabrata Mukhopadhyay and Vijay H. Shah Amitava Das, Uday Shergill, Lokendra Thakur, Sutapa Sinha, Raul Urrutia,
You might find this additional information useful...
41 articles, 19 of which you can access free at: This article cites http://ajpgi.physiology.org/cgi/content/full/298/6/G908#BIBL
including high-resolution figures, can be found at: Updated information and services http://ajpgi.physiology.org/cgi/content/full/298/6/G908
can be found at: AJP - Gastrointestinal and Liver Physiologyabout Additional material and information http://www.the-aps.org/publications/ajpgi
This information is current as of May 18, 2010 .
http://www.the-aps.org/.Society. ISSN: 0193-1857, ESSN: 1522-1547. Visit our website at American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the American Physiologicalabnormal function of the gastrointestinal tract, hepatobiliary system, and pancreas. It is published 12 times a year (monthly) by the
publishes original articles pertaining to all aspects of research involving normal orAJP - Gastrointestinal and Liver Physiology
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

Ephrin B2/EphB4 pathway in hepatic stellate cells stimulates Erk-dependentVEGF production and sinusoidal endothelial cell recruitment
Amitava Das,2 Uday Shergill,1 Lokendra Thakur,1 Sutapa Sinha,3 Raul Urrutia,1
Debabrata Mukhopadhyay,3 and Vijay H. Shah1
1Gastrointestinal Research Unit, Mayo Clinic College of Medicine, Rochester, Minnesota; 2Departments of Basic Sciences,Biochemistry Division and Medicine, Loma Linda University School of Medicine, Loma Linda, California; and 3Departmentof Biochemistry and Molecular Biology, Mayo Clinic Foundation, Rochester, Minnesota
Submitted 15 December 2009; accepted in final form 20 March 2010
Das A, Shergill U, Thakur L, Sinha S, Urrutia R, Mukho-padhyay D, Shah VH. Ephrin B2/EphB4 pathway in hepatic stellatecells stimulates Erk-dependent VEGF production and sinusoidal endothelialcell recruitment. Am J Physiol Gastrointest Liver Physiol 298: G908–G915,2010. First published March 25, 2010; doi:10.1152/ajpgi.00510.2009.—Chemotaxis signals between hepatic stellate cells (HSC) and sinusoi-dal endothelial cells (SEC) maintain hepatic vascular homeostasis andintegrity and also regulate changes in sinusoidal structure in responseto liver injury. Our prior studies have demonstrated that the bidirec-tional chemotactic signaling molecules EphrinB2 and EphB4 areexpressed in HSC. The aim of our present study was to explorewhether and how the EphrinB2/EphB4 system in HSC could promoteSEC recruitment, which is essential for sinusoidal structure andremodeling. Stimulation of human HSC (hHSC) with chimeric ago-nists (2 �g/ml) of either EphrinB2 or EphB4 (EphrinB2 Fc or EphB4Fc, respectively) significantly increased VEGF mRNA levels in hHSCas assessed by quantitative PCR, with respective small interferingRNAs for EphrinB2 and EphB4 inhibiting this increase (P � 0.05,n � 3). EphrinB2 agonist-induced increase in VEGF mRNA levels inhHSC was associated with increased phosphorylation of Erk and wassignificantly blocked by U0126 (20 �M), an inhibitor of MEK, whichis a kinase upstream from Erk (P � 0.05, n � 3). The EphB4 agonistalso significantly increased human VEGF promoter activity (P �0.05, n � 3) as assessed by promoter reporter luciferase assay intransfected LX2-HSC. This was associated with upregulation of thevasculoprotective transcription factor, Kruppel-like factor 2 (KLF2).In Boyden chamber assays, conditioned media from hHSC stimulatedwith agonists of EphrinB2 or EphB4 increased SEC chemotaxis in aVEGF-dependent manner, compared with control groups that in-cluded basal media with agonists of EphrinB2, EphB4, or HSC-conditioned media from HSC in absence of agonist stimulation (P �0.05, n � 3). EphB4 expression was detected in situ within liversinusoidal vessels of rats after carbon tetrachloride-induced liverinjury. In summary, activation of the EphrinB2/EphB4 signalingpathway in HSC promotes chemotaxis of SEC through a pathway thatinvolves Erk, KLF2, and VEGF. These studies identify EphrinB2 orEphB4 as a key intermediary that links HSC signal transductionpathways with angiogenesis and sinusoidal remodeling.
hepatic stellate cell; sinusoidal endothelial cell; VEGF; ephrins
HEPATIC STELLATE CELLS (HSC) are liver-specific pericytes (3,30), which signal with adjacent sinusoidal endothelial cells(SEC) to regulate vascular homeostasis and sinusoidal struc-tural changes that occur in response to liver injury and chronicliver disease (1, 19, 34). HSC embrace SEC with their longcytoplasmic processes and extensions that ideally position
HSC for paracrine signaling with SEC. Indeed, multiple mol-ecules are implicated in HSC-SEC cross talk, especially vas-cular endothelial growth factor A (VEGF), which is a keymolecule released by HSC that promotes SEC migration,recruitment, and angiogenesis (19, 26, 38). Although failure ofsuch signaling results in severe vascular defects (20, 22, 23),the molecular mechanisms that mediate HSC-SEC cross talkmechanisms such as VEGF secretion are not fully defined.
Eph receptor tyrosine kinases constitute a large family oftransmembrane proteins with a single cytoplasmic kinase do-main that is activated in response to binding of ephrin ligandsto the extracellular globular domain of the receptor (11). Basedon their ligand-binding characteristics, Ephs have been subdi-vided into Eph A and Eph B receptors, although significantredundancy and cross talk exists between subclasses (28).Several Eph/ephrin molecules are expressed in blood vessels,and targeted inactivation of genes encoding EphrinB2 andEphB4 in mice has demonstrated their essential role for angio-genesis and vascular remodeling (2, 15, 40). Our prior studies(31) have identified the ephrin family of proteins as keyregulators of HSC motility downstream from platelet derivedgrowth factor (PDGF) stimulation. However, effects of thisclass of molecules on release of angiogenic factors from HSChas not been explored. Therefore, these proteins appear to beinteresting candidates for studying SEC and HSC cross talk.
In this study, we interrogate the role of ephrins in SECrecruitment signaling. Especially since vascular endothelialgrowth factor (VEGF) is a canonical SEC chemotactic agentsecreted by HSC, we hypothesized that ephrin activation inHSC may promote recruitment of SEC through HSC secretionof VEGF. Indeed, we demonstrate, using complementarymechanistic approaches, that ephrin-Eph interactions regulateVEGF gene transcription in HSC and that this in turn promotesSEC recruitment. Additionally, we provide insights into sev-eral key signal intermediaries that determine this process bothin cytosol and in nucleus. We anticipate that these resultsshould expand our understanding of the dynamic interplay ofSEC and HSC within the hepatic sinusoids.
MATERIALS AND METHODS
Cell culture. All tissue culture reagents were obtained from GIBCO(Rockville, MD). Isolated primary human HSC (ScienCell ResearchLaboratories) and primary human SEC (ScienCell Research Labora-tories) were used between passages 2 and 6 (31). HSC and SEC werecultured in defined medium obtained from the vendor and supple-mented with 10% fetal bovine serum, penicillin (100 IU/ml), andstreptomycin (100 �g/ml).
Address for reprint requests and other correspondence: V. H. Shah, Gastro-enterology Research Unit, Mayo Clinic and Foundation, 200 First St. SW,Rochester, MN 55905 (e-mail: [email protected]).
Am J Physiol Gastrointest Liver Physiol 298: G908–G915, 2010.First published March 25, 2010; doi:10.1152/ajpgi.00510.2009.
0193-1857/10 Copyright © 2010 the American Physiological Society http://www.ajpgi.orgG908
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

EphB4/EphrinB2 chimeras. EphB4 and EphrinB2 chimeras werepurchased from R & D Biosystems (Minneapolis, MN). EphrinB2 andEphB4 chimeras consisted of the extracellular domain of the respec-tive ephrin protein fused to the Fc portion of human IgG and wereutilized to activate EphB4 and EphrinB2, respectively (14, 31).
Quantitative real-time PCR. Total RNA was extracted from HSCby using the RNeasy kit according to the manufacturer’s instruction(Qiagen, Valencia, CA). Total RNA (1 �g) was used for the cDNAsynthesis by using random hexamer primer of the SuperScript IIIFirst-Strand Synthesis System (Invitrogen, Carlsbad, CA). For Taq-Man-based real-time PCR analysis, 25 ng of each cDNA was added tothe Taqman Universal PCR Master Mix along with 900 nM of eachprimer and 200 nM of probe according to the manufacturer’s instruc-tion (Applied Biosystems, Carlsbad, CA). Real-time fluorescencemonitoring was performed with the Applied Biosystems 7500 Real-time PCR System instrument. Amplification of human GAPDH andeukaryotic 18S rRNA was used in the same reaction of all samples asan internal control. Gene-specific mRNA was subsequently normal-ized to GAPDH mRNA. Levels of VEGF A mRNA were expressed asfold difference of EphrinB2 Fc- or EphB4 Fc-treated cells comparedwith vehicle-treated cells. For KLF2 mRNA levels, SYBRgreen wasused for quantitative real-time PCR with GAPDH mRNA serving asnormalization control.
Gene silencing. Small interfering RNA (siRNA) targeting humanEphrinB2, EphB4, and a scrambled control were obtained fromQiagen (Valencia, CA). Cells were transfected with siRNA by use ofoligofectamine (Invitrogen). Conditions and concentrations requiredfor specificity of knockdown with high transfection efficiency wereindividually established. In some experiments, reconstitution of func-tional EphrinB2 signaling was achieved by addition of the agonisticrecombinant EphrinB2 Fc chimera.
VEGF ELISA. VEGF-A concentration in supernatants of vehicle,EphrinB2-FC (2 �g/ml), and EphB4-FC (2 �g/ml)-stimulated HSCwere measured by use of a hVEGF Quantitative kit (R & D Biosys-tems) as per the manufacturer’s instructions. Data were calculated aspicograms of VEGF per microgram of cellular protein.
Luciferase promoter assay. LX2 cells, an immortalized humanHSC line (18), were transfected with human VEGF-promoter-lucif-erase reporter constructs (pVEGF) and 0.01 �g Renilla luciferasereporter vector to control for transfection efficiency (pRL-TK) by useof Lipofectamine 2000 (Invitrogen). At 10 h later, culture mediumwas changed and cells were cultured for an additional 12–14 h inpresence or absence of stimulation by Fc chimeras of EphrinB2 orEphB4. In a separate set of experiments, LX2 cells were cotransfectedwith human pVEGF and either EphrinB2/EphB4 siRNA or scrambled(control) siRNA. Luciferase assays were conducted by using a dualluciferase kit (Promega) as described previously (8, 9).
Immunoblotting. Cells were homogenized in a lysis buffer (50 mMTris·HCl, 0.1 mM EGTA, 0.1 mM EDTA, 1% NP-40, 0.1% deoxy-cholic acid, 0.1% SDS, pH 7.5). Protein quantification of samples wasperformed with the Lowry assay. Detergent-soluble protein lysateswere separated by SDS-PAGE on a 10% acrylamide gel, and proteinswere transferred onto nitrocellulose membrane. The membranes werewashed in Tris-buffered saline with 0.05% Tween, blocked in 5%nonfat dry milk, and incubated with p44/42 MAPK, phospho-p44/42MAPK antibody (Ab) (Thr202/Tyr204), Akt, and phospho-Akt Ab(Ser473 and Thr308) (Cell Signaling, Santa Cruz, CA). Membranewas reprobed for actin to confirm equal protein loading and transferbetween lanes.
Boyden chamber assay. Cell migration assays were performed byusing a Boyden chamber (Becton Dickinson, Heidelberg, Germany)with 12-�m-pore-size polycarbonated filters coated with type I col-lagen (50 �g/ml). Cells suspended in serum free medium were seededto the upper wells (20 � 103 cells/well) and lower chambers werefilled with 26 �l of EphrinB2-Fc, EphB4-Fc, or vehicle-treated HSCcell culture supernatant (conditioned media). In another set of exper-iments, cell culture supernatants from vehicle, EphrinB2-Fc or
EphB4-Fc-stimulated HSC, preincubated with either VEGF-A neu-tralizing antibody (10 �g/ml) (R & D Biosystems) or isotype IgGcontrol (10 �g/ml), were used to assess VEGF dependency of theeffect. After 4 h of incubation at 37°C the polycarbonated filter wasremoved and migrated cells on the lower surface were stained withHEMA-3. Cells passed through the filter were quantified from randommicroscopic fields. Each assay was carried out in triplicate with sixreplicates of each group per assay. Results are expressed as themean number of migrated cells � SD.
Confocal immunofluorescence microscopy. Liver tissues fromCCl4-treated rats were harvested and embedded in optimal cuttingtemperature compound. Then 5-�m sections were cut and mounted onslides. Samples were blocked with 10% goat serum in PBS for 1 h andthen incubated with goat anti-mouse EphB4 (1:50), rabbit monoclonalPDGFR-� antibody (1:80), and von Willebrand factor (vWF) anti-body [1:400; vWF is detected in situ on SEC after liver injury (33)]overnight at 4°C degrees, followed by incubation for 1 h with AlexaFluor 488-conjugated donkey anti-goat (1:250) and Alexa Fluor546-conjugated goat anti-rabbit (1:250) secondary antibodies, re-spectively. The slides were then counterstained with Toto3, andconfocal microscopy was performed by LSM 5 Pascal (Carl Zeiss,Thornwood, NY).
Statistical analysis. Experiments were performed in triplicate witha minimum of three independent experiments. Data are depicted asmeans � SE. Comparisons were performed via Student’s t-test orone-way analysis of variance when comparing more than two samplegroups, with statistical significance set at P � 0.05.
RESULTS
EphrinB2 and EphB4 activation stimulates VEGF secretionby HSC. HSC interact with SEC through bidirectional para-crine signaling to form angiogenic vascular networks in liver.We had previously shown that PDGF signaling through EphrinB2regulates HSC motility and vascular coverage (31); however,effects on SEC were not explored. Since VEGF signaling iscritical for endothelial cell sprouting, proliferation, migration,permeability, and survival signaling (10), we looked at theability of HSC to produce VEGF in response to EphrinB2/EphB4 stimulation. VEGF mRNA levels were increased afterstimulation with EphrinB2/EphB4 Fc chimera as measured byquantitative real-time PCR (Fig. 1A). The stimulation ofVEGF-A mRNA induced by EphrinB2/EphB4 Fc occurred ina time-dependent manner (Fig. 1A). Additionally, cyclohexi-mide, an inhibitor of protein translational, blocked EphB4Fc-induced increases in VEGF mRNA, indicating that thepathway of regulation involved de novo protein translation(Fig. 1B). To further corroborate these results, we silencedEphrinB2 and EphB4 with their respective siRNA, both re-agents that we have previously validated with regard to theirspecificity and magnitude of knockdown (31). Interestingly,VEGF mRNA levels were significantly decreased by 40% inpresence of either EphrinB2 siRNA or EphB4 siRNA, indicat-ing that inhibition of endogenous cell-to-cell activation ofEphrinB2/EphB4 signaling is important for maintaining basalVEGF mRNA production (Fig. 1C). Furthermore, this siRNA-induced decrease in VEGF A mRNA expression could bereconstituted and amplified by adding back the relevant Fcchimeras in an exogenous manner (Fig. 1C). A similar andstatistically significant effect, albeit quantitatively smaller, wasalso observed by VEGF ELISA performed from supernatantsof EphrinB2-Fc-stimulated HSC (vehicle: 373.2 pg/�g � 2.5pg/�g vs. EphrinB2 FC: 413.6 pg/�g � 6.7 pg/�g, n � 3separate experiments; P � 0.05 between the groups). Thus
G909EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

these studies demonstrate that EphrinB2/EphB4 activation in-creases VEGF mRNA levels in HSC.
Ephrins activate VEGF transcription by regulating its pro-moter activity. To explore the potential mechanisms of VEGFtranscription regulation by EphrinB2 and EphB4, we studiedthe activity of a human VEGF reporter luciferase construct(pVEGF) in presence or absence of ephrin Fc chimeras in LX2cells. Interestingly, the EphB4 Fc chimeric agonist was more
Fig. 1. EphrinB2 and EphB4 increase VEGF mRNA levels in human hepaticstellate cells (HSC). HSC were treated with EphrinB2 chimera (2 �g/ml),EphB4 chimera (2 �g/ml), and/or respective small interfering RNA (siRNA)for 3, 12, and 48 h, respectively, and the total RNA was extracted for VEGFmRNA estimation by quantitative RT-PCR analysis. A: there was a significantincrease in VEGF mRNA levels in both EphrinB2- and EphB4-treated groupscompared with their respective time controls at 12 and 48 h (n � 3 separateexperiments, each in triplicate, *P � 0.05). B: cyclohexamide (10 �g/ml)blocked EphB4 Fc-induced VEGF A mRNA production (n � 3 separateexperiments, each in triplicate, *P � 0.05). C: basal VEGF mRNA levels wereattenuated by incubating HSC with EphrinB2 or EphB4 siRNA (n � 3 separateexperiment, each in triplicate, *P � 0.05). Stimulation of siRNA-transfectedcells with EphrinB2 and EphB4 chimeras reversed the siRNA-mediatedinhibitory effect on VEGF mRNA levels.
Fig. 2. EphB4 activates VEGF promoter. A: LX2 cells cotransfected withRenilla control and full-length human VEGF reporter luciferase construct(pVEGF) showed an increased luciferase activity upon stimulation with bothEphrinB2 (2 �g/ml) and EphB4 (2 �g/ml) chimeras, with EphB4 stimulationachieving a statistically significant effect (n � 3 separate experiments, each intriplicate, *P � 0.05). B: knockdown of EphrinB2 or EphB4 with theirrespective siRNA resulted in a 40–50% reduction in luciferase activity com-pared with control siRNA, whereas stimulation of siRNA-knockdown cellswith activating chimera reconstituted basal promoter activity (n � 3 separateexperiments, each in triplicate, *P � 0.05). C: stimulation of LX2 cells byEphB4 or EphrinB2 chimeras significantly increased KLF2 mRNA levels asassessed by quantitative real-time PCR (n � 3 separate experiments, each intriplicate, *P � 0.05).
G910 EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

potent than the EphrinB2 Fc and significantly increased VEGFpromoter activity by 2.3-fold (Fig. 2A). To substantiate the roleof EphB4 on VEGF gene transcription, we performed reporterassays in cells cotransfected with human VEGF promoter andeither EphrinB2 siRNA or EphB4 siRNA or scrambled (con-trol) siRNA. Relative luciferase activity in cells cotransfectedwith either EphrinB2 or EphB4 siRNA was significantly de-creased by 40–50% compared with cells cotransfected withscrambled siRNA, consistent with the effects of these siRNAVEGF mRNA levels shown in Fig. 1 and reflective of theeffects of endogenous cell-cell EphrinB2/EphB4 interactionson VEGF transcription (Fig. 2B). This decrease in VEGFpromoter activity could again be rescued by the respective Fcchimeras, which increased the relative luciferase activity withEphB4 Fc, again showing a more prominent effect than Eph-rinB2 Fc (Fig. 2B). Lastly, EphrinB2/EphB4-stimulated HSC
showed an increase in mRNA levels of the vasculoprotec-tive transcription factor KLF2 (27) compared with thevehicle-treated group as assessed by quantitative real-timePCR (Fig. 2C). In summary, these results indicate a signif-
Fig. 3. Ephrin-induced VEGF-A upregulation requires Erk phosphorylation.A: EphrinB2 (EFNB2) and EphB4 stimulation of HSC increased Erk phos-phorylation compared with vehicle, whereas Akt phosphorylation (pAKT-Ser473 and pAKT-Thr308) was unchanged (0–4 �g/ml; n � 3 separateexperiments; pThr308-to-total Akt ratio from densitometric analysis is shownbelow the Western blot). B: U0126 (10 �m) attenuated the increase in Erkphosphorylation as assessed by Western blot. Total Erk served as a proteinloading control. PDGF stimulation was used as a positive control for Erk andAkt activation. C: U0126 (10 �m) decreased VEGF mRNA levels in Eph-rinB2- and EphB4-stimulated cells compared with vehicle (*P � 0.05).
Fig. 4. Conditioned media from EphrinB2/EphB4-stimulated HSC increasessinusoidal endothelial cell (SEC) migration through VEGF-A production.A: conditioned medium (CM) from HSC stimulated with EphrinB2-Fc, EphB4-Fc, or vehicle for 48 h was used to assess the migratory effect on SEC viaBoyden chamber. Ave No., average number; HHSEC, human hepatic sinusoi-dal endothelial cells; Ab, antibody. Media from both EphrinB2- and EphB4-treated groups (4th and 6th bars) increased SEC migration compared with themedia from the vehicle-treated HSC (2nd bar) and compared with groupswhere basal medium (BM) containing EphrinB2 or EphB4 was used as achemokine (3rd and 5th bars) (n � 3 separate experiment, *P � 0.05).B: conditioned media from vehicle or EphrinB2-Fc-treated HSC when prein-cubated with VEGF-A neutralizing antibody (10 �g/ml) significantly de-creased (�50% reduction) the ability of SEC to migrate compared with therespective groups in which the conditioned medium was preincubated withisotype IgG control (n � 3 separate experiments, *P � 0.05).
G911EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from
icant effect of EphrinB2 and EphB4 on VEGF gene tran-scription.
EphrinB2/EphB4 regulate VEGF mRNA levels via Erksignaling. We next sought to identify key cytosolic signalingintermediates that may regulate EphrinB2/EphB4 stimulationof the VEGF promoter. We focused our attention on Erk andAkt (Ser473 and Thr308), both key signaling intermediariesimplicated in VEGF production (12, 17, 24, 36). Western blotanalysis showed that EphrinB2/EphB4 stimulation in HSCsignals intracellularly via Erk phosphorylation (although in-creasing doses of chimera did not have an incremental intra-cellular response) (Fig. 3A). Interestingly, the positive controlPDGF stimulated both Erk and Akt, whereas EphrinB2/EphB4activation specifically activated Erk but not Akt (Fig. 3B). Thespecificity of Erk phosphorylation by EphrinB2/EphB4 wasfurther examined by using the MEK inhibitor U0126. Eph-rinB2 induced Erk phosphorylation was significantly reducedin cells stimulated with U1026 (Fig. 3B). Lastly, VEGF mRNAlevels were analyzed in EphrinB2/EphB4-stimulated HSC pre-treated with U0126. VEGF mRNA levels were significantlyreduced after pretreatment of cells with U0126, indicating thatEphrinB2/EphB4 stimulates VEGF release from HSC throughan Erk-dependent but Akt-independent pathway (Fig. 3C).
EphrinB2/EphB4 stimulation of HSC promotes SEC migra-tion through a VEGF-dependent mechanism. SEC migration isa key step in angiogenesis and therefore to ascertain whether
EphrinB2/EphB4 stimulation of HSC was sufficient to mediateHSC release of chemotactic molecules that regulate SEC mi-gration, HSC were stimulated with EphrinB2 or EphB4 Fcchimera and conditioned medium was collected to ascertainchemotactic effects of the conditioned media on SEC by use ofa modified Boyden chamber. Indeed, conditioned media fromEphrinB2 Fc-stimulated HSC enhanced SEC chemotaxis com-pared with its relevant control groups, including non-chimera-stimulated conditioned media and nonconditioned chimera-containing media (Fig. 4A; *P � 0.05 for EphrinB2-Fc vs.conditioned media). Preincubation of the condition media witheither VEGF-A-neutralizing antibody (10 �g/ml) significantlyreduced (�50%) the ability of SEC to migrate in response toEphrinB2 Fc compared with an isotype IgG control-treatedgroup (Fig. 4B). The neutralizing antibody also significantlyreduced migration by �50% in response to EphB4 Fc as well(data not shown). Thus these studies highlight the role ofEphrinB2/EphB4 stimulation of HSC as a mechanism to pro-mote SEC chemotaxis through VEGF-A production.
Lastly, since EphrinB2/EphB4 activation and signaling re-quire cell-to-cell contact because both the ligand and its cog-nate receptor are cell membrane bound and the ligand andreceptor both signal bidirectionally, we looked at the expres-sion of EphB4 in rats with CCl4-induced liver injury, since theprocess of liver fibrosis involves robust angiogenic responses(13) requiring cross talk between EC and HSC for sinusoidal
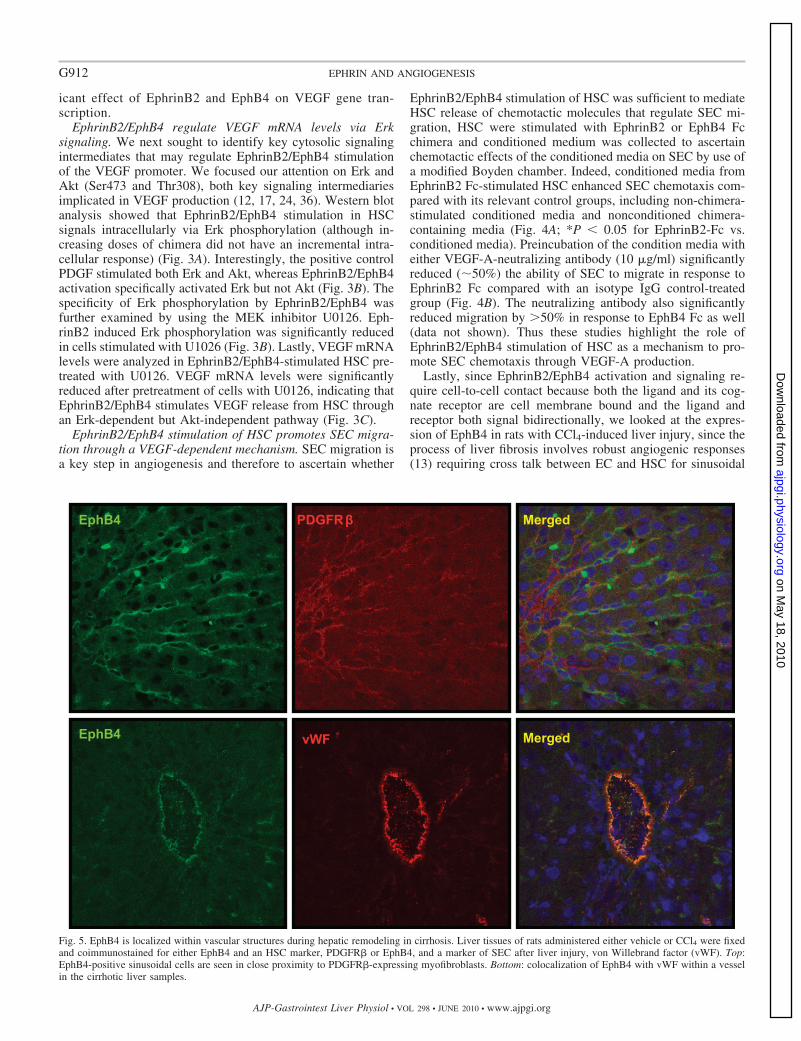
Fig. 5. EphB4 is localized within vascular structures during hepatic remodeling in cirrhosis. Liver tissues of rats administered either vehicle or CCl4 were fixedand coimmunostained for either EphB4 and an HSC marker, PDGFR� or EphB4, and a marker of SEC after liver injury, von Willebrand factor (vWF). Top:EphB4-positive sinusoidal cells are seen in close proximity to PDGFR�-expressing myofibroblasts. Bottom: colocalization of EphB4 with vWF within a vesselin the cirrhotic liver samples.
G912 EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

remodeling. Although the immunofluorescent signal was verylow in vehicle-treated rats (data not shown), CCl4-treated ratsevidenced prominent expression of EphB4 within the sinusoidswith close proximity of both EC and HSC (Fig. 5), therebyproviding in situ positioning of ephrins within the hepaticsinusoids.
DISCUSSION
Hepatic microcirculation, by virtue of its sinusoidal config-uration, modulates hepatic blood flow and facilitates transportof nutrients to parenchymal liver cells. The structural proxim-ity and communication between HSC and SEC within thesinusoids is essential for maintaining sinusoidal integrity andfunction. Paracrine signaling between these two predominantcell populations in the sinusoids also plays a significant role insettings of chronic liver diseases (10, 31) wherein remodelingand distortion of the sinusoidal architecture occurs in coordi-nation with derangements of vascular growth factor productionsuch as VEGF (7, 25, 29). Although the ability of HSC tosecrete VEGF (4) and their angiogenic effects (26) are wellknown, the pathways that modulate VEGF production fromHSC are not as well defined. In this regard, the present studiesidentify a novel observation, expanded with mechanistic de-tails, whereby the ephrin signaling pathway regulates HSC-derived VEGF production and ensuing SEC recruitment.
Ephrins have been shown to play an important role inregulation of vascular assembly and homeostasis (6). Indeed,they provide molecular distinction to endothelium when it isdeveloping into artery or vein and also play an important rolein angiogenic interaction between embryonic vessels (35, 40).Our prior studies (31) showed that the ephrin pathway couldregulate HSC whereby HSC acquire an angiogenic phenotypein response to PDGF through an ephrin-dependent pathway.The present studies tie together these two lines of work in HSCand SEC by demonstrating that the ephrin pathway in HSC can
promote SEC recruitment via release of substances that arechemoattractive for SEC such as VEGF. Indeed, HSC havebeen implicated in the secretion of angiogenic factors such asangiopoietin and VEGF (37). We observed that EphrinB2/EphB4 chimeras increased VEGF mRNA production fromHSC along with an increase in VEGF promoter activity andthat this effect could be specifically blocked by silencing withEphB4 and EphrinB2 siRNA. Interestingly, members of theephrin family have been observed to be upregulated in hepatitisC virus-associated cirrhosis (32) and cancer (16), both condi-tions that may require increased vascularity to maintain aconducive environment for the disease process to progress andconsistent with the immunofluorescent analysis performed inthis study using rats exposed to CCl4.
Our study also highlights some interesting transcriptionalmechanisms by which EphB4/EphrinB2 stimulation leads toVEGF secretion. For example, EphB4/EphrinB2 stimulationalso led to the upregulation of KLF2, a member of the zincfinger-containing family of transcription factor (5). A previousstudy by Wu et al. (41) has shown that KLF2/ mouseembryos show a defect in vascular maturation due to the failureof the mural cells to migrate to the arterial wall. Other work hasalso implicated KLF2 as a key modulator of vascular integrity(21, 27), making it a logical transcriptional molecule to regu-late HSC and SEC cross talk requisite for homeostasis as wellas remodeling.
An important signaling pathway by which ephrins can stim-ulate VEGF secretion from HSC might involve Erk phosphor-ylation as we have observed in our study. Phosphorylation ofErk, a key cell survival and proliferation protein, has beenshown previously by other to regulate VEGF-A expressionthrough HIF-1 phosphorylation (39). We observed that Eph-rinB2 stimulated phosphorylation of Erk. Furthermore, theMEK inhibitor U0126 blocked the phosphorylation secondaryto EphrinB2 treatment, showing the specificity of the signaling
Fig. 6. Proposed mechanism. Stimulation ofeither EphB4 receptor by EphrinB2 or Eph-rinB2 by EphB4 results in increased Erkphosphorylation (circled P) that leads to in-creased KLF2 and VEGF promoter activity.HSC-derived VEGF, in turn, provides a che-motactic stimulus for SEC migration andrecruitment. In an in vivo setting EphrinB2/EphB4 stimulation may occur by cell to cellcontact between HSC or between HSC andSEC.
G913EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

pathway. Since VEGF mRNA expression was also attenuatedby U0126, these studies indicate that Erk phosphorylation maybe an important step in ephrin-dependent VEGF productionand highlight the important role of Erk in vascular integrity andfunction.
Given that SEC motility was enhanced by supernatant fromHSC treated with EphB4 or EphrinB2 chimera, it can bespeculated that HSC might play an important role in recruit-ment of SEC through VEGF production upon stimulation withephrins. This mutual interaction between HSC and SECthrough VEGF and PDGF (31) production, respectively, wouldbe essential for juxtaposing these cells in the hepatic sinusoids,thereby maintaining vascular homeostasis in the liver and inturn for the changes involved in sinusoidal vascular remodel-ing. One question that emerges from these data is the autocrineas opposed to paracrine stimulation of ephrins in HSC. Indeed,EphrinB2 and EphB4 bind and signal bidirectionally, a processthat may provide redundancy of function. Indeed, in our studiessome of the mechanistic readouts were more prominent inresponse to stimulation of EphrinB2-Fc whereas others weremore prominent in response to EphB4-Fc. For example, Eph-rinB2-FC revealed a greater stimulatory effect in studies fo-cused on the protein level such as chemotaxis and ELISA,whereas EphB4-FC revealed quantitatively greater effects onstudies focused on transcription studies. This observation high-lights that although signaling through these receptors is bidi-rectional, there may be quantitative and qualitative differencesin the way that signals are transduced between the two recep-tors. In vivo, we anticipate that ephrin stimulation may occurthrough cell-cell interactions between adjacent stellate cells aswell as between adjacent EC and HSC (Fig. 6). Indeed, theimmunofluorescence analysis of liver from animals with ex-perimental cirrhosis depicts close proximity of EphB4 withboth SEC and HSC.
In summary, EphrinB2 and EphB4 regulate VEGF produc-tion from HSC via Erk phosphorylation and is associated withKLF2 upregulation. We anticipate that this pathway impor-tantly regulates HSC and SEC cross talk that regulates sinu-soidal function in health and disease.
GRANTS
This work was supported by National Institutes of Health Grants R01 DK59615 and R01 HL 86990 (V. H. Shah), P30 DK 084567, Clinical Core.Amitava Das was supported through the Pilot Feasibility Award by MayoClinic Centre on Cell Signaling in Gastroenterology.
DISCLOSURES
No conflicts of interest are declared by the author(s).
REFERENCES
1. Adams DH, Burra P, Hubscher SG, Elias E, Newman W. Endothelialactivation and circulating vascular adhesion molecules in alcoholic liverdisease. Hepatology 19: 588–594, 1994.
2. Adams RH, Wilkinson GA, Weiss C, Diella F, Gale NW, Deutsch U,Risau W, Klein R. Roles of ephrinB ligands and EphB receptors incardiovascular development: demarcation of arterial/venous domains, vas-cular morphogenesis, and sprouting angiogenesis. Genes Dev 13: 295–306, 1999.
3. Allt G, Lawrenson JG. Pericytes: cell biology and pathology. CellsTissues Organs 169: 1–11, 2001.
4. Ankoma-Sey V, Wang Y, Dai Z. Hypoxic stimulation of vascularendothelial growth factor expression in activated rat hepatic stellate cells.Hepatology 31: 141–148, 2000.
5. Atkins GB, Jain MK. Role of Kruppel-like transcription factors inendothelial biology. Circ Res 100: 1686–1695, 2007.
6. Augustin HG, Reiss Y. EphB receptors and ephrinB ligands: regulators ofvascular assembly and homeostasis. Cell Tissue Res 314: 25–31, 2003.
7. Corpechot C, Barbu V, Wendum D, Kinnman N, Rey C, Poupon R,Housset C, Rosmorduc O. Hypoxia-induced VEGF and collagen Iexpressions are associated with angiogenesis and fibrogenesis in experi-mental cirrhosis. Hepatology 35: 1010–1021, 2002.
8. Das A, Fernandez-Zapico ME, Cao S, Yao J, Fiorucci S, Hebbel RP,Urrutia R, Shah VH. Disruption of an SP2/KLF6 repression complex bySHP is required for farnesoid X receptor-induced endothelial cell migra-tion. J Biol Chem 281: 39105–39113, 2006.
9. Das A, Yaqoob U, Mehta D, Shah VH. FXR promotes endothelial cellmotility through coordinated regulation of FAK and MMP-9. ArteriosclerThromb Vasc Biol 29: 562–570, 2009.
10. Deleve LD, Wang X, Guo Y. Sinusoidal endothelial cells prevent ratstellate cell activation and promote reversion to quiescence. Hepatology48: 920–930, 2008.
11. Dodelet VC, Pasquale EB. Eph receptors and ephrin ligands: embryo-genesis to tumorigenesis. Oncogene 19: 5614–5619, 2000.
12. Dong G, Chen Z, Li ZY, Yeh NT, Bancroft CC, Van Waes C.Hepatocyte growth factor/scatter factor-induced activation of MEK andPI3K signal pathways contributes to expression of proangiogenic cyto-kines interleukin-8 and vascular endothelial growth factor in head andneck squamous cell carcinoma. Cancer Res 61: 5911–5918, 2001.
13. El-Assal ON, Yamanoi A, Soda Y, Yamaguchi M, Igarashi M,Yamamoto A, Nabika T, Nagasue N. Clinical significance of microves-sel density and vascular endothelial growth factor expression in hepato-cellular carcinoma and surrounding liver: possible involvement of vascularendothelial growth factor in the angiogenesis of cirrhotic liver. Hepatology27: 1554–1562, 1998.
14. Foo SS, Turner CJ, Adams S, Compagni A, Aubyn D, Kogata N,Lindblom P, Shani M, Zicha D, Adams RH. Ephrin-B2 controls cellmotility and adhesion during blood-vessel-wall assembly. Cell 124: 161–173, 2006.
15. Gerety SS, Wang HU, Chen ZF, Anderson DJ. Symmetrical mutantphenotypes of the receptor EphB4 and its specific transmembrane ligandephrin-B2 in cardiovascular development. Mol Cell 4: 403–414, 1999.
16. Hirai H, Maru Y, Hagiwara K, Nishida J, Takaku F. A novel putativetyrosine kinase receptor encoded by the eph gene. Science 238: 1717–1720, 1987.
17. Kosmidou I, Xagorari A, Roussos C, Papapetropoulos A. Reactiveoxygen species stimulate VEGF production from C2C12 skeletal myotubesthrough a PI3K/Akt pathway. Am J Physiol Lung Cell Mol Physiol 280:L585–L592, 2001.
18. Lee JS, Kang Decker N, Chatterjee S, Yao J, Friedman S, Shah V.Mechanisms of nitric oxide interplay with Rho GTPase family members inmodulation of actin membrane dynamics in pericytes and fibroblasts. AmJ Pathol 166: 1861–1870, 2005.
19. Lee JS, Semela D, Iredale J, Shah VH. Sinusoidal remodeling andangiogenesis: a new function for the liver-specific pericyte? Hepatology45: 817–825, 2007.
20. Leveen P, Pekny M, Gebre-Medhin S, Swolin B, Larsson E, BetsholtzC. Mice deficient for PDGF B show renal, cardiovascular, and hemato-logical abnormalities. Genes Dev 8: 1875–1887, 1994.
21. Lin Z, Kumar A, SenBanerjee S, Staniszewski K, Parmar K, VaughanDE, Gimbrone MA Jr, Balasubramanian V, Garcia-Cardena G, JainMK. Kruppel-like factor 2 (KLF2) regulates endothelial thrombotic func-tion. Circ Res 96: e48–57, 2005.
22. Lindahl P, Johansson BR, Leveen P, Betsholtz C. Pericyte loss andmicroaneurysm formation in PDGF-B-deficient mice. Science 277: 242–245, 1997.
23. Liu Y, Wada R, Yamashita T, Mi Y, Deng CX, Hobson JP, RosenfeldtHM, Nava VE, Chae SS, Lee MJ, Liu CH, Hla T, Spiegel S, Proia RL.Edg-1, the G protein-coupled receptor for sphingosine-1-phosphate, isessential for vascular maturation. J Clin Invest 106: 951–961, 2000.
24. Mazure NM, Chen EY, Laderoute KR, Giaccia AJ. Induction ofvascular endothelial growth factor by hypoxia is modulated by a phos-phatidylinositol 3-kinase/Akt signaling pathway in Ha-ras-transformedcells through a hypoxia inducible factor-1 transcriptional element. Blood90: 3322–3331, 1997.
25. Medina J, Arroyo AG, Sanchez-Madrid F, Moreno-Otero R. Angio-genesis in chronic inflammatory liver disease. Hepatology 39: 1185–1195,2004.
G914 EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from

26. Olaso E, Salado C, Egilegor E, Gutierrez V, Santisteban A, Sancho-Bru P, Friedman SL, Vidal-Vanaclocha F. Proangiogenic role of tumor-activated hepatic stellate cells in experimental melanoma metastasis Hepa-tology 37: 674–685, 2003.
27. Parmar KM, Larman HB, Dai G, Zhang Y, Wang ET, Moorthy SN,Kratz JR, Lin Z, Jain MK, Gimbrone MA Jr, Garcia-Cardena G.Integration of flow-dependent endothelial phenotypes by Kruppel-likefactor 2. J Clin Invest 116: 49–58, 2006.
28. Pasquale EB. The Eph family of receptors. Curr Opin Cell Biol 9:608–615, 1997.
29. Rosmorduc O, Wendum D, Corpechot C, Galy B, Sebbagh N, RaleighJ, Housset C, Poupon R. Hepatocellular hypoxia-induced vascular endo-thelial growth factor expression and angiogenesis in experimental biliarycirrhosis. Am J Pathol 155: 1065–1073, 1999.
30. Sato M, Suzuki S, Senoo H. Hepatic stellate cells: unique characteristicsin cell biology and phenotype. Cell Struct Funct 28: 105–112, 2003.
31. Semela D, Das A, Langer D, Kang N, Leof E, Shah V. Platelet-derivedgrowth factor signaling through ephrin-b2 regulates hepatic vascularstructure and function. Gastroenterology 135: 671–679, 2008.
32. Shackel NA, McGuinness PH, Abbott CA, Gorrell MD, McCaughanGW. Insights into the pathobiology of hepatitis C virus-associated cirrho-sis: analysis of intrahepatic differential gene expression. Am J Pathol 160:641–654, 2002.
33. Shah V, Haddad FG, Garcia-Cardena G, Frangos JA, Mennone A,Groszmann RJ, Sessa WC. Liver sinusoidal endothelial cells are respon-sible for nitric oxide modulation of resistance in the hepatic sinusoids. JClin Invest 100: 2923–2930, 1997.
34. Soriano P. Abnormal kidney development and hematological disorders inPDGF beta-receptor mutant mice. Genes Dev 8: 1888–1896, 1994.
35. Steinle JJ, Meininger CJ, Forough R, Wu G, Wu MH, Granger HJ.Eph B4 receptor signaling mediates endothelial cell migration and prolif-eration via the phosphatidylinositol 3-kinase pathway. J Biol Chem 277:43830–43835, 2002.
36. Takahashi A, Kureishi Y, Yang J, Luo Z, Guo K, Mukhopadhyay D,Ivashchenko Y, Branellec D, Walsh K. Myogenic Akt signaling regu-lates blood vessel recruitment during myofiber growth. Mol Cell Biol 22:4803–4814, 2002.
37. Taura K, De Minicis S, Seki E, Hatano E, Iwaisako K, OsterreicherCH, Kodama Y, Miura K, Ikai I, Uemoto S, Brenner DA. Hepaticstellate cells secrete angiopoietin 1 that induces angiogenesis in liverfibrosis. Gastroenterology 135: 1729–1738, 2008.
38. Tugues S, Fernandez-Varo G, Munoz-Luque J, Ros J, Arroyo V,Rodes J, Friedman SL, Carmeliet P, Jimenez W, Morales-Ruiz M.Antiangiogenic treatment with sunitinib ameliorates inflammatory infil-trate, fibrosis, and portal pressure in cirrhotic rats. Hepatology 46: 1919–1926, 2007.
39. Wang FS, Wang CJ, Chen YJ, Chang PR, Huang YT, Sun YC, HuangHC, Yang YJ, Yang KD. Ras induction of superoxide activates Erk-dependent angiogenic transcription factor HIF-1alpha and VEGF-A ex-pression in shock wave-stimulated osteoblasts. J Biol Chem 279: 10331–10337, 2004.
40. Wang HU, Chen ZF, Anderson DJ. Molecular distinction and angio-genic interaction between embryonic arteries and veins revealed by eph-rin-B2 and its receptor Eph-B4. Cell 93: 741–753, 1998.
41. Wu J, Bohanan CS, Neumann JC, Lingrel JB. KLF2 transcriptionfactor modulates blood vessel maturation through smooth muscle cellmigration. J Biol Chem 283: 3942–3950, 2008.
G915EPHRIN AND ANGIOGENESIS
AJP-Gastrointest Liver Physiol • VOL 298 • JUNE 2010 • www.ajpgi.org
on May 18, 2010
ajpgi.physiology.orgD
ownloaded from