![Molecular Imaging of Murine Intestinal Inflammation With 2-Deoxy-2-[ 18F]Fluoro- d-Glucose and Positron Emission Tomography](https://static.fdokumen.com/doc/165x107/6344fff26cfb3d4064097a1a/molecular-imaging-of-murine-intestinal-inflammation-with-2-deoxy-2-18ffluoro-.jpg)
Ependyma as a Possible Morphological Basis of Ischemic Preconditioning Tolerance in Rat Spinal Cord...
13
Cellular and Molecular Neurobiology, Vol. 24, No. 3, June 2004 ( C 2004) Ependyma as a Possible Morphological Basis of Ischemic Preconditioning Tolerance in Rat Spinal Cord Ischemia Model: Nestin and Fluoro-Jade B Observations Judita Orend ´ aˇ cov ´ a, 1,3 Enik ¨ o Raˇ cekov ´ a, 1 Karol´ ına Kuch ´ arov ´ a, 1 Barbora Pouˇ sov ´ a, 1 Tom ´ aˇ s Ondrejˇ c´ ak, 1 Marcela Martonˇ c´ ıkov ´ a, 1 Zuzana Daxnerov ´ a, 2 and Jozef Marˇ sala 1 Received September 30, 2003; accepted November 13, 2003 SUMMARY 1. To test our hypothesis that a transient nonlethal ischemic insult benefits the lum- bosacral spinal cord ischemic injury, nestin, the marker of proliferating cells, and Fluoro-Jade B, the marker of degenerating cells, were used in rats. Morphological outcome was evalu- ated after 12-min ischemia versus 12-min ischemia preconditioned by 3-min ischemic period and 30-min recirculation (IPC), in each group followed by 2, 3, and 4 days of posttreatment survival. 2. Twelve-minute ischemia, inducing nestin-positivity in ependyma and reactive astro- cytes at the L 1-3 spinal cord segments, shows this region as the viable region of spinal cord in all postischemic survival periods. On the other hand, abundance of Fluoro-Jade B-positive cells, distributed throughout the dorsal horn and intermediate zone of L 4 –S 2 segments, points out the most injured spinal cord region by ischemia. 3. After the same ischemic insult in IPC rats only a few nestin-positive ependymal cell and reactive astrocytes appeared beside the nestin-positive vessels in the lower lumbar and sacral spinal cord segments of all survival periods. The appearance of nestin-positive cells in the spinal cord segments, which “should have been affected” by ischemia indicates protection of this region by the IPC treatment. 4. The number and density evaluation of Fluoro-Jade B fluorescent cells of L 4 –S 2 seg- ments after ischemia and IPC confirmed that degenerating cells were significantly reduced in the IPC rats in all survival periods. 5. Our results showing the immunohistochemical response of epemdyma, committed to the presence of viable tissue, indicate that the ependymal cells may contribute to the ischemic resistance in the IPC rats. KEY WORDS: ischemia; ischemic preconditioning; ependyma; nestin; Fluoro-Jade B; quantitative evaluation. 1 Institute of Neurobiology, Slovak Academy of Sciences, Koˇ sice, Slovak Republic. 2 Institute of Biology and Ecology, Faculty of Sciences, P.J. ˇ Saf ´ arik University, Koˇ sice, Slovak Republic. 3 To whom correspondence should be addressed at Institute of Neurobiology, Slovak Academy of Sciences, ˇ Solt ´ esovej 4, Koˇ sice 040 01, Slovak Republic; e-mail: [email protected]. 477 0272-4340/04/0600-0477/0 C 2004 Plenum Publishing Corporation
Transcript of Ependyma as a Possible Morphological Basis of Ischemic Preconditioning Tolerance in Rat Spinal Cord...

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Cellular and Molecular Neurobiology, Vol. 24, No. 3, June 2004 ( C© 2004)
Ependyma as a Possible Morphological Basis of IschemicPreconditioning Tolerance in Rat Spinal Cord IschemiaModel: Nestin and Fluoro-Jade B Observations
Judita Orendacova,1,3 Eniko Racekova,1 Karolına Kucharova,1
Barbora Pousova,1 Tomas Ondrejcak,1 Marcela Martoncıkova,1
Zuzana Daxnerova,2 and Jozef Marsala1
Received September 30, 2003; accepted November 13, 2003
SUMMARY
1. To test our hypothesis that a transient nonlethal ischemic insult benefits the lum-bosacral spinal cord ischemic injury, nestin, the marker of proliferating cells, and Fluoro-JadeB, the marker of degenerating cells, were used in rats. Morphological outcome was evalu-ated after 12-min ischemia versus 12-min ischemia preconditioned by 3-min ischemic periodand 30-min recirculation (IPC), in each group followed by 2, 3, and 4 days of posttreatmentsurvival.
2. Twelve-minute ischemia, inducing nestin-positivity in ependyma and reactive astro-cytes at the L1−3 spinal cord segments, shows this region as the viable region of spinal cord inall postischemic survival periods. On the other hand, abundance of Fluoro-Jade B-positivecells, distributed throughout the dorsal horn and intermediate zone of L4–S2 segments,points out the most injured spinal cord region by ischemia.
3. After the same ischemic insult in IPC rats only a few nestin-positive ependymalcell and reactive astrocytes appeared beside the nestin-positive vessels in the lower lumbarand sacral spinal cord segments of all survival periods. The appearance of nestin-positivecells in the spinal cord segments, which “should have been affected” by ischemia indicatesprotection of this region by the IPC treatment.
4. The number and density evaluation of Fluoro-Jade B fluorescent cells of L4–S2 seg-ments after ischemia and IPC confirmed that degenerating cells were significantly reducedin the IPC rats in all survival periods.
5. Our results showing the immunohistochemical response of epemdyma, committedto the presence of viable tissue, indicate that the ependymal cells may contribute to theischemic resistance in the IPC rats.
KEY WORDS: ischemia; ischemic preconditioning; ependyma; nestin; Fluoro-Jade B;quantitative evaluation.
1 Institute of Neurobiology, Slovak Academy of Sciences, Kosice, Slovak Republic.2 Institute of Biology and Ecology, Faculty of Sciences, P.J. Safarik University, Kosice, Slovak Republic.3 To whom correspondence should be addressed at Institute of Neurobiology, Slovak Academy of Sciences,
Soltesovej 4, Kosice 040 01, Slovak Republic; e-mail: [email protected].
477
0272-4340/04/0600-0477/0 C© 2004 Plenum Publishing Corporation

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
478 Orendacova et al.
INTRODUCTION
In lower vertebrates, the spinal cord ependyma plays a significant role in neuronalregeneration. In these animals the ependymal cells rapidly proliferate, migrate, anddifferentiate to regenerate the injured spinal cord. They extend long processes thatare believed to provide longitudinal channels to guide regenerating central axonscaudally (Simpson and Duffy, 1994). Ependymal proliferation in response to spinalcord injury has been also described in morphological studies using adult mammals(Beattie et al., 1997; Vaquero et al., 1981; Wallace et al., 1987). However, it is notknown if these proliferating ependymal cells have similar beneficial regenerativeeffect as those in the lower vertebrates. In the adult rat brain, nestin, a neuralstem cell protein, can be found only in endothelial and selected subventricular cells(Morshead et al., 1994; Tohyama et al., 1992). Some evidence suggests that stem-likecells can be localized in or near the ependymal layer of the brain and spinal cord(Chiasson et al., 1999; Doetsch et al., 1999; Garcia-Verdugo et al., 1998; Horner et al.,2000; Johansson et al., 1999). Although Weiss et al. (1996) demonstrated multipo-tent stem cells, which can be isolated, expanded, and differentiated in vitro in theadult mice lumbosacral spinal cord (Shihabuddin et al., 1997) the normal ependymashows no immunoreactivity to nestin, a marker for neural precursor cells (Mokry,1999).
On the basis of experimental interventions mimicking spinal cord injuries, thefindings on nestin-positivity were found to be different from those mentioned above.Frisen et al. (1995) proposed an interesting hypothesis that a precursor cell popula-tion, which can be identified by nestin expression, exists around the central canal,and can proliferate and migrate to the injury site and contribute to the glial scar.The spinal cord, injured by clip compression, demonstrated a dramatic increase inependymal proliferation 3 days after injury and concomitant strong nestin expres-sion in ependyma (Namiki and Tator, 1999). Nestin expression was present in someneurons localized in the outer boundary zone of the ischemic lesion in the cortexand striatum, and in most ependymal cells in the ventricular and subventricular zonefrom the second day after middle cerebral artery occlusion (Li and Chopp, 1999).After a stroke, nestin immunoreactivity in glial, neuronal, and ependymal cells issuggestive of protein expression pattern found in the developing brain.
Ischemic preconditioning (IPC) has been defined as an endogenous cellularprotective mechanism, evoked by a brief ischemic period. IPC renders the CNStissue more resistant to the subsequent lethal ischemic insult (Burda et al., 2003;Danielisova et al., 2003; Kitagawa et al., 1991; Lukacova, 1999; Nemethova et al.,2003; Orendacova et al., 2003; Radonak et al., 2003; Zvara et al., 1999). A similarprotective effect of IPC has been observed after experimental traumatic brain injury(Perez-Pinzon et al., 1999). Contrary to the cerebral trauma, IPC does not improvethe outcome after spinal cord contusion injury, and even worsens the damage to thespinal cord white matter in rat (Ondrejcak et al., 2003).
A novel fluorochrome Fluoro-Jade B, like its predecessor Fluoro-Jade, is ananionic fluorescein derivative useful for histological staining of neurons undergoingdegeneration (Schmued et al., 1997; Schmued and Hopkins, 2000). It is a reliabletracer of high affinity, which makes it easier to detect labeled fine neuronal processes

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Ischemic Preconditioning Benefits Ischemic Injury 479
including dendrites and axons. To date, we know of no published studies using thisfluorochrome for detection of degenerating cells after ischemic injury.
In the present work we used nestin and Fluoro-Jade B labeling to study theregional differences between ischemic and IPC-induced changes of ependyma andthe gray matter of rat lumbosacral spinal cord.
MATERIALS AND METHODS
The experimental procedure was performed with approval from the AnimalCare Committee of the Institute of Neurobiology.
Surgical Procedure
Wistar albino rats were anesthetized with 1.5% halothane in a 45% oxygenair mixture. The tail artery was cannulated with a PE-50 polyethylene catheter formonitoring the mean distal aortic blood pressure (MDAP). The left femoral arterywas isolated and a Fogarty 2F balloon-tipped catheter was introduced for later ad-vancement into the descending thoracic aorta. The left internal carotid artery wascannulated with a 20-gauge catheter for measurement of the mean proximal aorticblood pressure (MPAP); this cannula was connected to a head blood-collection cir-cuit (37.5◦C) that included an 88-cm vertical column of heparinized (1 U/ml) salinesolution. When the aorta was occluded, proximal aortic blood was allowed to bleedinto the heparinized column, maintaining MPAP at 65 ± 3 mmHg. To induce spinalcord ischemia, the Fogarty 2F catheter (situated in the left femoral artery) was in-serted retrogradely into the descending thoracic aorta 10.5 cm from the femoralarteriotomy. During the occlusion the catheter was inflated with 0.05 ml of salinesolution, and aortic occlusion was confirmed by reduction in MDAP. (1) Rats of theischemic group were subjected to the 12-min occlusion of thoracic aorta. (2) Rats ofthe ischemic preconditioned group (IPC) were first subjected to the 3-min occlusionfollowed by 30-min reperfusion, and then again to the 12-min occlusion of thoracicaorta. Then, the vented blood from the carotid artery cannula and the heat exchangerwas returned, the catheters removed, the surgical wounds closed, and the rats of bothexperimental groups put to recovery.
Tissue Sampling for Immunohistochemical and Fluorescent Analysis
The animals were subjected to neurological testing (ambulation and stepping)after two, 24 hr and at the end of each survival period, i.e. 2, 3, and 4 days, for eval-uation of the hind limb motor function. The animals without loss of motor functionafter ischemia or with loss of motor function after IPC were discarded. The rats wereperfused with 4% paraformaldehyde in a deep anesthesia. The lumbosacral spinalcord was removed from the backbone, postfixed, cryoprotected in 30% sucrose andfrozen 40µm thick coronal sections were cut and collected in phosphate buffer (PB).
After blocking in normal serum, the free-floating sections were treated withthe primary monoclonal antibody (mAb) anti-nestin Rat-401 (Developmental

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
480 Orendacova et al.
Hybridoma Studies Bank) diluted 1:400. After thorough washing, the sections wereincubated with antimouse IgG (Dako) secondary antibody. Diaminobenzidine(DAB) was then used as a sensitive chromogen for light microscopic visualization ofnestin-positive cells.
Fluoro-Jade B (Histo-Chem Inc.) was used as a marker of neurons undergoingdegeneration. The sections mounted on 2% gelatin coated slides and air-dried wereheated at 50◦C for at least half an hour before staining. The slides were immersed inabsolute alcohol for 3 min, for 1 min in 70% alcohol, and for 1 min in distilled water.Then the slides were transferred to the solution of 0.06% potassium permanganatefor 15 min and rinsed in distilled water for 2 min. After 30 min in the staining solution,prepared according to the Schmued and Hopkins (2000), 3× 1 min rinses in distilledwater followed. The slides were dried at room temperature, cleared by xylene, andcover slipped with Fluoromont. The sections were observed by Olympus BX51 lightmicroscope.
Some alternate sections were processed according to the Gallyas reduced silvermethod for degenerating neurons (Gallyas et al., 1980).
Quantitative Assessment
The sections were transformed into digital images by the Olympus ReflectedFluorescence system U-RFL-T, the Olympus BX51, the digital camera DP50, andthe visual communication suite Olympus DP-soft, version 3.0. To determine thenumber and optical density of Fluoro-Jade B-exhibiting neurons in the L4–S2 spinalcord the UTHSCSA Image Tool and Microsoft Excel were used in the rats surviving4 days after ischemia (n = 3) and after IPC (n = 3). The gray scale values relativeto the highest concentration of Fluoro-Jade B in degenerating neurons and the lackof the fluorescent marker in the section was defined as the 100 and 0% density values.The results were statistically evaluated by Dunn’s Method and Tukey Test and havebeen given as means ± SD.
RESULTS
Ischemia
In the most injured L4–S2 spinal cord segments ischemia caused widespreadneuronal necrosis, which appeared as distinct demarcated area throughout the dorsalhorn and intermediate zone on Gallyas impregnated sections (Fig. 1(A)). Pathomor-phological signs of degeneration detected along the rostro-caudal extent of the mostinjured segments increased from the second postischemic day and were fully devel-oped after 4-day survival. Nestin immunohistochemistry did not show characteristicexpression pattern either of vessel walls or of ependymal or glial cells (results notshown).
The L3 spinal cord segments, comprising the immediate rostral border of thelesioned region, revealed distinct nestin immunoreactvity of the ependymal cellsand reactive astrocytes in all postinjury survival periods. The ependymal wall was

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Ischemic Preconditioning Benefits Ischemic Injury 481
Fig. 1. Representative micrographs of Gallyas silver impregnation in L4–S2 segments after 4-daysurvival. (A) Pale circumscribed area of degenerated cells in the dorsal horn (DH) and the inter-mediate zone (IZ) after 12-min ischemia. (B) The same region without any signs of degeneration,either in the dorsal horn (DH) or in the intermediate zone (IZ), after IPC. Central canal (CC), bar =400 µm.
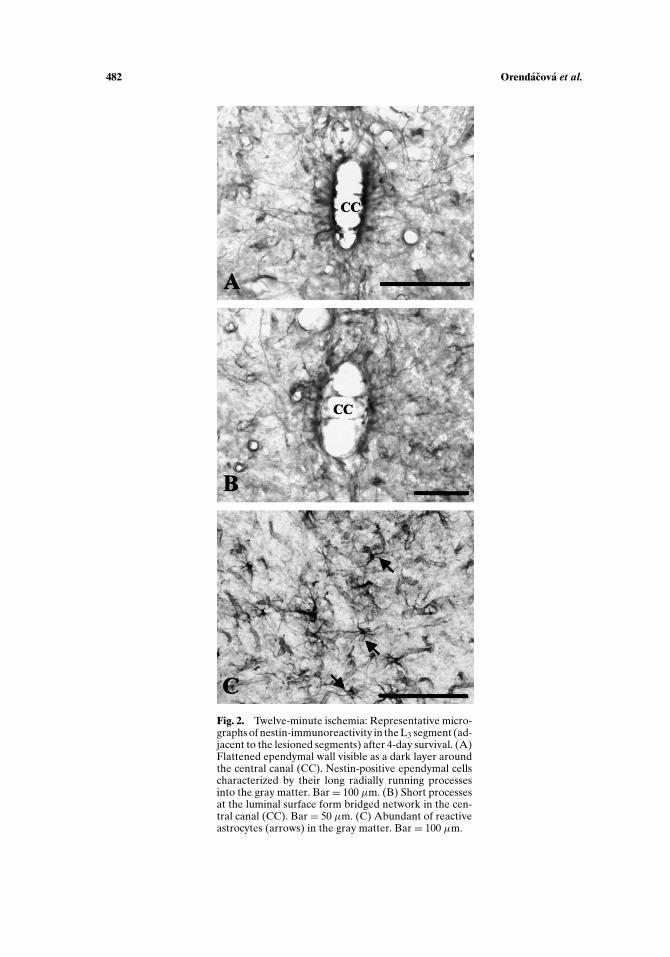
visible as a dark layer of varying thickness around the spinal cord central canal. Theapical cytoplasm of the ependymal cell was reduced and the nuclei appeared to belocated closer to the lumen of the central canal (Fig. 2(A)). The ependymal cells werecharacteristic by their long nestin-positive processes extending radially from the cellbasal pole and traceable for a long distance from the cell bodies. At the luminal sur-face of ependyma short processes progressed to the opposite side forming bridgednetwork in the central canal lumen (Fig. 2(B)). The reactive astrocytes, character-ized by their starry shape body due to multiple running processes, expressed nestin(Fig. 2(C)). The vessels in the immediate border to the necrotic zone were alsonestin-positive. In general, the immunohistochemical observation of lumbosacralspinal cord in rats exposed to 12-min ischemia showed that injured spinal cordsegments were rostrally delineated by a region of characteristic nestin immunore-activity.
Fluoro-Jade B fluorescence exhibited a different distribution pattern in compar-ison to the distribution pattern of nestin-positivity mentioned above. DegeneratedFluoro-Jade B-positive cells were observed at L4–S2 spinal cord segments, i.e. in seg-ments injured by ischemia and were characterized by a conspicuous green fluorescentlabeling distributed throughout the dorsal horns and intermediate zone except mo-toneurons in the ventral horns (Fig. 4(A)).
P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
482 Orendacova et al.
Fig. 2. Twelve-minute ischemia: Representative micro-graphs of nestin-immunoreactivity in the L3 segment (ad-jacent to the lesioned segments) after 4-day survival. (A)Flattened ependymal wall visible as a dark layer aroundthe central canal (CC). Nestin-positive ependymal cellscharacterized by their long radially running processesinto the gray matter. Bar= 100 µm. (B) Short processesat the luminal surface form bridged network in the cen-tral canal (CC). Bar = 50 µm. (C) Abundant of reactiveastrocytes (arrows) in the gray matter. Bar = 100 µm.

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Ischemic Preconditioning Benefits Ischemic Injury 483
Ischemic Preconditioning
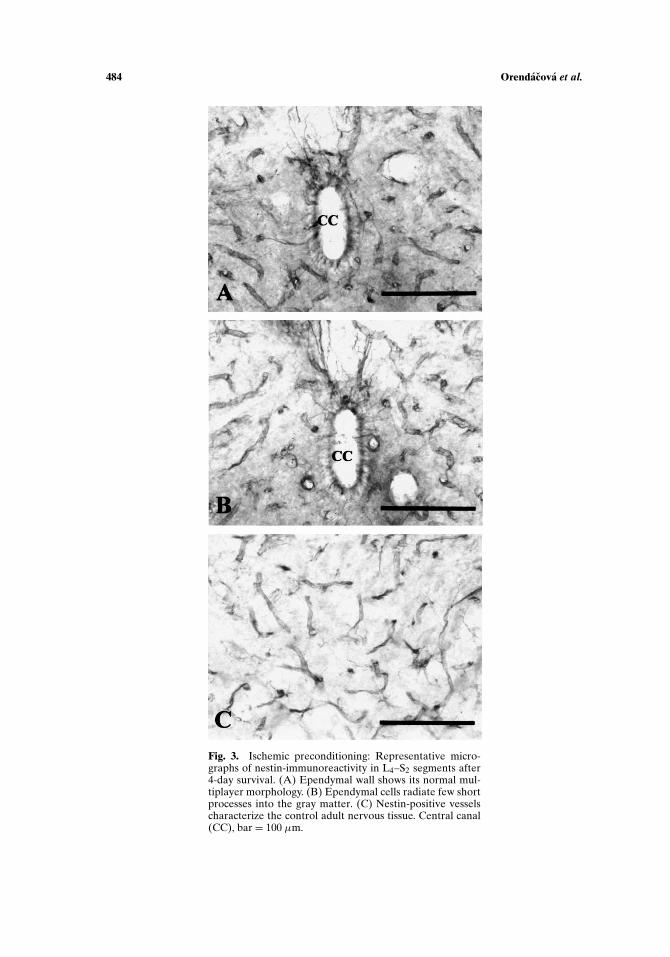
In the L4–S2 spinal cord segments there were no signs of degeneration afterGallyas silver impregnation (Fig. 1(B)). Nestin-immunohistochemistry showed char-acteristic labeling of vessels. Nestin-expressing ependymal cells appeared randomlyin L4–S2 segments in all survival periods. The ependymal wall clearly displayed its nor-mal multilayered morphology, and no irregularities in thickness could be detected asdescribed after ischemia (Fig. 3(A)). Nuclei of ependymal cells were located basallyand the apical cytoplasm surrounded continually the lumen of the central canal.The labeled radially running processes of ependymal cells were fewer and shorter incomparison with that visible after 12-min ischemia (Fig. 3(B)). Nestin-positivity ofreactive astrocytes occurred very rarely. Nestin-positive vessels were seen throughtthe lumbosacral spinal cord segments in all survival periods (Fig. 3(C)).
Fluoro-Jade B fluorescence showed only mild presence of the degenerating cellssituated preferentially in the intermediate zone. The degenerating cells were rarewithin the L4–S2 spinal cord segments (Fig. 4(B)). The Flouro-Jade B findings ofdegenerated cells are in accordance with demonstration of postischemic spinal corddegeneration by the silver impregnation (Fig. 1(B)).
Quantitative Assesment
Quantitative analysis of Fluoro-Jade B sections showed a significant increaseof fluorescent cells throughout the gray matter of L4–S2 spinal cord segments afterischemia in comparison to the IPC rats (Table I). In the rats submitted to the 12-min ischemia the highest number of Fluoro-Jade B-exhibiting cells were present inthe dorsal horn and intermediate zone. In the rats preconditioned by ischemia onlynegligible number of Fluoro-Jade B-expressing cells appeared and they were mostlydistributed in the intermediate zone. Distribution of Fluoro-Jade B fluorescent cellsin the upper lumbar spinal cord segments was of no importance, therefore it wasnot evaluated quantitatively. The optical density values, reflecting concentration ofFluoro-Jade B in degenerating cells, were 63.15 ± 1.17. These values were the samein the rats after ischemia and IPC. There were no significant differences between 2-,3-, and 4-day survival.
DISCUSSION
Ischemia and Ischemic Preconditioning
Although the recent research in the ischemic myocardium, brain, and spinal corddemonstrates different degree of the tissue protection with IPC involving longerreperfusion intervals ranging from 1 to 5 days (for review, see Lukacova, 1999),shorter reperfusion intervals ranging from 10 min to 6 hr also offers some degreeof neuroprotection, thus suggesting that several mechanisms may play a role in IPCprotection of the nervous tissue. The present work investigated whether 30-min reper-fusion after 3-min ischemia is effective against ischemic injury. To address this ques-tion, we used a reliable rat model of spinal cord ischemia (Taira and Marsala, 1996).
P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
484 Orendacova et al.
Fig. 3. Ischemic preconditioning: Representative micro-graphs of nestin-immunoreactivity in L4–S2 segments after4-day survival. (A) Ependymal wall shows its normal mul-tiplayer morphology. (B) Ependymal cells radiate few shortprocesses into the gray matter. (C) Nestin-positive vesselscharacterize the control adult nervous tissue. Central canal(CC), bar = 100 µm.

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Ischemic Preconditioning Benefits Ischemic Injury 485
Fig. 4. Representative micrographs of Fluoro-Jade B-labeling in the L4–S2 segments after 4-day sur-vival. (A) Twelve-minute ischemia induces fluorescence throughout the dorsal horn and intermediatezone. (B) IPC-induced scant fluorescence in the intermediate zone. Central canal (CC), bar = 200 µm.
With this model it was showed that ischemic intervals longer than 8-min trigger arapid degeneration of spinal cord interneurons and detectable neurohistopathologi-cal (Marsala et al., 1997) and histochemical (Yang et al., 2000) changes in lumbosacralspinal cord segments. It was shown that 12-min ischemia is sufficient to induce spasticparaplegia in all experimental animals and 2-day survival postoperation was crucialfor development of either the neurological injury in ischemic rats (Ondrejcak, 2001),or the immunohistochemically detectable protective effect of IPC. Silver impreg-nation analysis of neuronal damage and concurrent histochemical characterizationof NADPH-diaphorase positive neuronal pools in the rabbit lumbosacral segmentsshowed extensive neuronal degeneration in the central gray matter between 1 and4 days after ischemia (Marsala et al., 1997).
Distribution of Nestin-Positive Cells
Hockfield and McKay (1985), who first produced the Rat-401 anti-nestin mon-oclonal antibody, identified cell types immunoreactive for nestin. Nestin, an inter-mediate filament protein, found in the neuroepithelial precursor cells can also beexpressed in vitro in the multipotent precursor cells in the adult CNS (Gritti et al.,1996; Reynolds and Weiss, 1992; Weiss et al., 1996). Although the normal ependyma
Table I. Number of Fluoro-Jade B-Exhibiting Neurons FollowingIschemia (I) and Ischemia With Ischemic Preconditioning (IPC)Distributed Throughout the Gray Matter of L4 Segment after 4-Day
Survival
Dorsal horn Zona intermedia Ventral horn
I 160.13 ± 5.73 123.07 ± 3.37 76.14 ± 8.76IPC 7.79 ± 2.09∗ 17.48 ± 8.81∗ 5.33 ± 1.52∗
Note. Values are expressed as means ± SD in the section of 10 rats.The results were evaluated by Dunn’s Method and Tukey Test.∗P < 0.001.

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
486 Orendacova et al.
of the central canal maintained very limited cell proliferation (1.5% of the cells areproliferating), it did not exhibit nestin immunoreactivity, which would indicate pre-cursor cell qualities (Namiki and Tator, 1999). Since it has been reported that allof the known multipotent neural stem cells express nestin (Dahlstrand et al., 1995;Stemple and Mahanthappa, 1997), we used nestin immunoreactivity to identify cellswith precursor cell qualities in the spinal cord ependyma of adult rats after ischemicinjury.
Expression of the stem cell marker, nestin, by the ependymal cells and by reactiveastrocytes indicates that these differentiated cells have potential of dedifferentiation.This attribute was considered to be characteristic for viable tissue distributed mainlyin the vicinity of injury. Therefore, our findings that after ischemic injury, nestin-positivity was found only in the areas adjacent to the lesion characterize this regionas viable noninjured. Similarly, the occurrence of nestin-positivity in the ependymalcells and reactive astrocytes after IPC in those spinal cord segments (L4–S2), which“should have been affected” by ischemia, confirms the viability of this region, i.e.indicate protective effect of short ischemic preconditioning. Our data, showing theperilesion distribution of nestin expression after ischemia, are consistent with similarobservations after a clip-compression spinal cord injury (Liu et al., 2002; Namiki andTator, 1999). Even though the mechanism of nestin reexpression is unclear, the classicstatic view of the glial scar is challenged by the observation of reactive astrocytesexpressing nestin. Kaya et al. (1999) consider the possibility that nestin expressionmay contribute to the dynamic feature of glial scar formation, and Duggal et al.(1997) have suggested that nestin may also facilitate the formation of new synapticconnections in neurons in the immediate neighborhood of the glial scar.
Distibution of Fluoro-Jade B Degeneration
Fluoro-Jade B, a novel fluorochrome, is demonstrated to be high-affinity trac-ers detecting neuronal degeneration (Schmued et al., 1997; Schmued and Hopkins,2000). It allows a rapid, reliable, and sensitive labeling of fine neuronal processesincluding distal dendrites, axons, and terminals resulting in a yellow-green appearingfluorescence in ultraviolet excitation. Although kainic acid exposure was the onlyneurotoxic insult systematically examined by this fluorochrome, it is believed thatthis fluorescent tracer will detect dead neurons regardless of the cause of death. Pu-tative chemistry and mode of Fluoro-Jade B action remains to be confirmed, butregardless of the mechanism of action, the authors discuss that the tracer can be usedto resolve a number of questions involving neuronal cell death, including studies onbrain pathology, toxicology, development, dementia, and neuroprotection.
Recently we have successfully used Fluoro-Jade B to label the dying cells withinrostral migratory stream (RMS) at various postnatal periods (Martoncıkova et al.,2003) as well as in adult rats after olfactory bulbectomy, i.e. in the absence of themigrating cells target (Pousova et al., 2003). Use of Fluoro-Jade B after 12-min is-chemia resulted in labeling numerous degenerating cells throughout the gray matterof lumbosacral spinal cord segments. The presented distribution of ischemia-induceddegeneration correlates with the ischemic neuronal damage detected by some re-cently developed silver impregnation procedures after Gallyas (Gallyas et al., 1980)

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Ischemic Preconditioning Benefits Ischemic Injury 487
and Nauta (Marsala et al., 1989). Although the qualitative observation of the Fluoro-Jade B-labeling was plausible, we evaluated the nuber and density of degeneratedcells by the methods used in some previous papers (Kucharova et al., 2001; Marsalaet al., 2003; Maslej et al., 2003). This approaching put an evidence of significant de-crease of degenerating cells number after IPC. The published quantitative assessmentwas provided in animals of both experimental groups after 4-day survival, becausethe differences between 2, 3, and 4 days were negligible.
CONCLUSIONS
Nestin-immunohistochemistry indicates significance of the spinal cord region inthe vicinity of damaged tissue.
Fluoro-Jade B appears to be a reliable neuropathological technique comparablewith the silver impregnation allowing demonstration of the IPC benefit effect.
ACKNOWLEDGMENTS
This work was supported by the VEGA grants: 2/2082/22, 2/3216/23, 2/3217/23,and 2/3218/23; the APVT grant: 51-013002. The mAb Rat-401, developed by S.Hockfield, was obtained from the Developmental Studies Hybridoma Bank NICHD,Department of Biological Sciences, University of Iowa, Iowa City, IA 52242.
REFERENCES
Beattie, M. S., Bresnahan, J. C., Koman, J., Tovar, C. A., Van Meter, M., Anderson, D. K., Faden, A. I., Hsu,C. Y., Noble, L. J., Salzman, S., and Young, W. (1997). Endogenous repair after spinal cord contusioninjuries in the rat. Exp. Neurol. 148:453–463.
Burda, J., Hrehorovska, M., Garcia Bonilla, L., Danielisova, V., Cızkova, D., Burda, R., Nemethova, M.,Fando, J. L., and Salinas, M. (2003). Two different mechanisms are essential for the acquisition ofischemic tolerance in CA1 field of hippocampus: Role for protein synthesis. Physiol. Res. 52:1P.
Chiasson, B. J., Tropepe, V., Morshead, C. M., and Van Der Kooy, D. (1999). Adult mammalian forebrainependymal and subependymal cells demonstrate proliferative potential, but only subependymal cellshave neural stem cell characteristics. J. Neurosci. 19:4462–4471.
Dahlstrand, J., Lardelli, M., and Lendahl, U. (1995). Nestin mRNA expression correlates with the centralnervous system progenitor cell state in many, but not all, regions of developing central nervous system.Dev. Brain Res. 84:109–129.
Danielisova, V., Gottlieb, M., Nemethova, M., and Burda, J. (2003). Ischemic preconditioning defends theexcessive iron deposition of post-ischemic rat brain. Physiol. Res. 52:3P.
Doetsch, F., Caille, I., Lim, D. A., Garcia-Verdugo, J. M., and Alvarez-Buylla, A. (1999). Subventricularzone astrocytes are neural stem cells in the adult mammalian brain. Cell 97:703–716.
Duggal, N., Schmidt-Kastner, R., and Hakim, A. M. (1997). Nestin expression in reactive astrocytesfollowing focal cerebral ischemia in rats. Brain Res. 768:1–9.
Frisen, J., Johansson, C. B., Torok, C., Risling, M., and Lendahl, U. (1995). Rapid, widespread, and lon-glasting induction of nestin contributes to the generation of glial scar tissue after CNS injury. J. CellBiol. 131:453–464.
Gallyas, F., Wolff, J. R., Bottcher, H., and Zaborszky, L. (1980). A reliable and sensitive method to localizeterminal degeneration and lysosomes in the central nervous system. Stain Technol. 55:299–306.
Garcia-Verdugo, J. M., Doetsch, F., Wichterle, H., Lim, D. A., and Alvarez-Buylla, A. (1998). Architectureand cell types of the adult subventricular zone: In search of the stem cells. J. Neurobiol. 36:234–248.

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
488 Orendacova et al.
Gritti, A., Parati, E. A., Cova, L., Frolichstall, P., Galli, R., Wanke, E., Faravelli, L., Morassutti, D. J.,Roisen, F., Nickel, D. D., and Vescovi, A. L. (1996). Multipotential stem cells from the adult mousebrain proliferate and self-renew in response to basic fibroblast growth factor. J. Neurosci. 16:1091–1100.
Hockfield, S., and Mckay, R. D. G. (1985). Identification of major cell classes in the developing mammaliannervous system. J. Neurosci. 5:3310–3328.
Horner, P. J., Power, A. E., Kempermann, G., Kuhn, H. G., Palmer, T. D., Winkler, J., Thal, L. J., andGage, F. H. (2000). Proliferation and differentiation of progenitor cells throughout the intact adultrat spinal cord. J. Neurosci. 20:2218–2228.
Johansson, C. B., Momma, S., Clarke, D. L., Risling, M., Lendahl, U., and Frisen, J. (1999). Identificationof a neural stem cell in the adult mammalian central nervous system. Cell 96:25–34.
Kaya, S. S., Mahmood, A., Li, Y., Yavuz, E., and Chopp, M. (1999). Expression of nestin after traumaticbrain injury in rat brain. Brain Res. 840:153–157.
Kitagawa, K., Matsumoto, M., Kuwabara, K., Tagaya, M., Ohtsuki, T., Hata, R., Ueda, H., Handa, N.,Kimura, K., and Kamada, T. (1991). “Ischemic tolerance” phenomenon detected in various brainregions. Brain Res. 561:203–211.
Kucharova, K., Fercakova, A., Jalc, P., and Marsala, J. (2001). NADPH diaphorase activity in the primarysensory neurons following peripheral axotomy. Biologia 56:321–328.
Li, Y., and Chopp, M. (1999). Temporal profile of nestin expression after focal cerebral ischemia in adultrat. Brain Res. 838:1–10.
Liu, K., Wang, Z. C., Wang, H. Y., and Zhang, Y. Z. (2002). Nestin expression and proliferation ofependymal cells in the adult rats spinal cord after injury. Chin. Med. J. 11593:339–341.
Lukacova, N. (1999). The relevance of ischemic preconditioning and tolerance in the neuroprotectivity ofischemia-induced neuronal damage: Up-to-date review. Biologia 54:29–34.
Marsala, J., Kluchova, D., and Marsala, M. (1997). Spinal cord gray matter layers rich in NADPHdiaphorase-positive neurons are refractory to ischemia-reperfusion-induced injury: A histochemi-cal and silver impregnation study in rabbit. Exp. Neurol. 145:165–179.
Marsala, J., Marsala, M., Lukacova, N., Ishikawa, T., and Cızkova, D. (2003). Localization and distributionpattern of nicotinamide adenine dinucleotide phosphate diaphorase exhibiting axons in the whitematter of the spinal cord of the rabbit. Cell. Mol. Neurobiol. 23:57–92.
Marsala, J., Sulla, I., Santa, M., Marsala, M., Mechırova, E., and Jalc, P. (1989). Early neurohistopatholog-ical changes of canine lumbosacral spinal cord segments in ischemia-reperfusion-induced paraplegia.Neurosci. Lett. 106:83–88.
Martoncıkova, M., Racekova, E., Orendacova, J., and Pousova, B. (2003). Changes in rostral migratorystream and subventricular zone during postnatal development in rat: Anatomy and immunohisto-chemistry. Physiol. Res. 52:10P.
Maslej, P., Pomfy, M., and Kucharova, K. (2003). Morphometry and densitometry of NADPH-d positiveneurons in the dog forebrain: Ontogenic study. Biologia 58:409–412.
Mokry, J. (1999). Immunohistochemicka detekce bunık exprimujıcich nestin. In Houdek, L. (ed.), Neuralnıprekurzorove buoky a jejich kultivace, Galen, Praha, pp. 44–49.
Morshead, C. M., Reynolds, B. A., Craig, C. G., Mcburney, M. W., Staines, W. A., Morassutti, D., Weiss,S., and Van Der Kooy, D. (1994). Neural stem cells in the adult mammalian forebrain: A relativelyquiescent subpopulation of subependymal cells. Neuron 13:1071–1082.
Namiki, J., and Tator, Ch. H. (1999). Cell proliferation and nestin expression in the ependyma of the adultrat spinal cord after injury. J. Neuropathol. Exp. Neurol. 58:489–497.
Nemethova, M., Danielisova, V., and Burda, J. (2003). Ischemic preconditioning and c-fos immunoreac-tivity in the CA1 pyramidal layer of the postischemic rat brain. Physiol. Res. 52:13P.
Ondrejcak, T. (2001). Sledovanie funkcnych zmien po poskodenı miechy potkana. Tezy diplomovej prace.Prırodovedecka fakulta, UPJS Kosice, pp. 1–65.
Ondrejcak, T., Vanicky, I., and Galik, J. (2003). Ischemic preconditioning does not improve the outcomeafter spinal cord contusion injury in the rat. Physiol. Res. 52:13P.
Orendacova, J., Racekova, E., Ondrejcak, T., Pousova, B., and Martoncıkova, M. (2003). Ependymalregion of rat spinal cord after ischemia and ischemic preconditioning: An immunohistochemicalstudy. Physiol. Res. 52:14P.
Perez-Pinzon, M. A., Alonso, O., Kraydieh, S., and Dietrich, W. D. (1999). Induction of tolerance againsttraumatic brain injury by ischemic preconditioning. NeuroReport 10:2951–2954.
Pousova, B., Racekova, E., Orendacova, J., and Martoncıkova, M. (2003). Neurogenesis and apoptosis inthe SVZ and RMS of the adult rats after bilateral olfactory bulbectomy. Physiol. Res. 52:15P.
Radonak, J., Vajo, J., Racekova, E., Lukacova, N., and Marsala, J. (2003). The use of ischemic precondi-tioning against ischemic damage in rats. Physiol. Res. 52:16P.

P1: JLS
Cellular and Molecular Neurobiology [cemn] pp1172-cemn-484794 March 25, 2004 21:20 Style file version Oct 23, 2000
Ischemic Preconditioning Benefits Ischemic Injury 489
Reynolds, B. A., and Weiss, S. (1992). Generation of neurons and astrocytes from isolated cells of theadult mammalian central nervous system. Science 255:1707–1710.
Schmued, L. C., Albertson, C., and Slikker, W., Jr. (1997). Fluoro-Jade: A novel fluorochrome for thesensitive and reliable histochemical localization of neuronal degeneration. Brain Res.751:37–46.
Schmued, L. C., and Hopkins, K. J. (2000). Fluoro-Jade B: A high affinity fluorescent marker for thelocalization of neuronal degeneration. Brain Res. 874:123–130.
Shihabuddin, L. S., Ray, J., and Gage, F. H. (1997). FGF-2 is sufficient to isolate progenitors found in theadult mammalian spinal cord. Exp. Neurol. 148:577–586.
Simpson, S. B., Jr., and Duffy, M. T. (1994). The lizard spinal cord: A model system for study of spinalcord injury and repair. Prog. Brain Res. 103:229–241.
Stemple, D. L., and Mahanthappa, N. K. (1997). Neural stem cells are blasting off. Neuron 18:1–4.Taira, Y., and Marsala, M. (1996). Effect of proximal arterial pressure on function, spinal cord blood flow,
and histopathologic changes after increasing intervals of aortic occlusion in the rat. Stroke 27:1850–1858.
Tohyama, T., Lee, V. M.-Y., Rorke, L. B., Marvin, M., Mckay, R. D. G., and Trojanowski, J. Q. (1992).Nestin expression in embryonic human neuroepithelium and in human neuroepithelial tumor cells.Lab. Invest. 66:303–313.
Vaquero, J., Ramiro, M. J., Oya, S., and Cabezudo, J. M. (1981). Ependymal reaction after experimentalspinal cord injury. Acta Neurochir. 55:295–302.
Wallace, M. C., Tator, C. H., and Lewis, A. J. (1987). Chronic regenerative changes in the spinal cord aftercompression injury in rats. Surg. Neurol. 27:209–219.
Weiss, S., Dunne, Ch., Hewson, J., Wohl, Ch., Wheatley, M., Peterson, A. C., and Reynolds, B. A. (1996).Multipotent CNS stem cells are present in the adult mammalian spinal cord and ventricular neuroaxis.J. Neurosci. 16:7599–7609.
Yang, L. Ch., Orendacova, J., Wang, V., Ishikawa, T., Zaksh, T. L., and Marsala, M. (2000). Transient spinalcord ischemia in rat: The time course of spinal FOS protein expression and the effect of intraischemichzpothermia (27◦C). Cell. Mol. Neurobiol. 20:351–365.
Zvara, D. A., Colonna, D. M., Deal, D. D., Vernon, J. C., Gowda, M., and Lundell, J. C. (1999). Ischemicpreconditioning reduces neurologic injury in arat model of spinal cord ischemia. Ann. Thorac. Surg.68:874–880.

![Molecular Imaging of Murine Intestinal Inflammation With 2-Deoxy-2-[18F]Fluoro-d-Glucose and Positron Emission Tomography](https://static.fdokumen.com/doc/165x107/6344fffc596bdb97a908b96f/molecular-imaging-of-murine-intestinal-inflammation-with-2-deoxy-2-18ffluoro-d-glucose.jpg)


















