
Enhancement of in vitro fungicidal activity of fuberidazole to Botrytis cinerea by cucurbiturils
11
1 23 Journal of Inclusion Phenomena and Macrocyclic Chemistry and Macrocyclic Chemistry ISSN 1388-3127 J Incl Phenom Macrocycl Chem DOI 10.1007/s10847-013-0352-8 Enhancement of in vitro fungicidal activity of fuberidazole to Botrytis cinerea by cucurbiturils Na’il Saleh, Suad M. Ajeb, Arjun Sham & Synan F. AbuQamar
-
Upload
dubaimonsters -
Category
Documents
-
view
0 -
download
0
Transcript of Enhancement of in vitro fungicidal activity of fuberidazole to Botrytis cinerea by cucurbiturils

1 23
Journal of Inclusion Phenomena andMacrocyclic Chemistryand Macrocyclic Chemistry ISSN 1388-3127 J Incl Phenom Macrocycl ChemDOI 10.1007/s10847-013-0352-8
Enhancement of in vitro fungicidal activityof fuberidazole to Botrytis cinerea bycucurbiturils
Na’il Saleh, Suad M. Ajeb, Arjun Sham &Synan F. AbuQamar

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL ARTICLE
Enhancement of in vitro fungicidal activity of fuberidazoleto Botrytis cinerea by cucurbiturils
Na’il Saleh • Suad M. Ajeb • Arjun Sham •
Synan F. AbuQamar
Received: 19 March 2013 / Accepted: 1 July 2013
� Springer Science+Business Media Dordrecht 2013
Abstract The host–guest interaction of fuberidazole
(FBZ) fungicide with cucurbituril (CB) macromolecules
was characterized in pure water using UV–vis spectro-
photometric and NMR techniques. The in vitro applications
(at pH 5.5 in pure water) of host–guest complexes were
conducted against Botrytis cinerea. While addition of CB5
to FBZ had no significant effect in vitro, mixing CB7 or
CB8 with FBZ in a 1:2 ratio improved fungal growth
inhibition at least threefold, when compared to the corre-
sponding concentration of the unbound fungicide mole-
cules. Empty CB hosts were completely inactive.
Furthermore, the inhibitory activity to B. cinerea was rel-
atively maintained down to a concentration of 5:10 lM of
the CB7/8@FBZ complexes, relative to any of controls.
Complexation by CB7/8 further improved the photosta-
bility of the fungicides with photostabilization factors of 7
and 3, respectively. CB7/8 bound the protonated forms of
these guests very strongly but their neutral forms were
significantly weaker, which reflects a complexation-
induced increase of their pKa values by 3.8 units with CB7
and 1.4 units with CB8. The present investigation
constitutes an innovative, nonclassical, approach to
enhance fungicides efficacy utilizing macromolecules with
a potential application in crop protection technology.
Keywords Botrytis cinerea � Cucurbiturils � Fuberidazole
Introduction
Non-covalent/reversible interactions are the basis of the
most impressive functions of living systems and life sci-
ences. The preparation of host–guest complexes based on
such recognitions has been the focus of many supramolec-
ular chemists ever since the pioneer work of Cram and Cram
[1]. Such host–guest approaches have been utilized in many
biological and medicinal applications, such as drug delivery
[2, 3], pharmaceutical (pre)formulations [4], sensing/detec-
tions of bioactive analytes [5–7], diagnostic tools [8], and in
agricultural uses [9]. The microcapsule formulation tech-
nology, i.e., Warrior II insecticide with zeon, is a lambda-
cyhalothrin insecticide that has been widely used against a
broad spectrum of primary and secondary insects on crops
such as corn, cotton, soybean and tomato (Syngenta Crop
Protection Inc.). Encapsulation has been used in the for-
mulation of pesticides including fungicides. A recent report
demonstrated, for instance, the effect of CB8 on carboxin’s
activity against Rhizoctonia solani in aqueous solution [9].
The antifungal activity of this systemic anilide fungicide
towards Rhizoctonia solani significantly improved upon
complex formation with CB8 in vitro.
Recent examples of such macromolecules, cucur-
bit[n]uril (CB) macrocycles in Fig. 1 [10], are readily
synthesized on a gram-to-kilogram scale by the conden-
sation reaction of glycouril with formaldehyde under
acidic conditions. CB hosts possess negligible in vitro
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10847-013-0352-8) contains supplementarymaterial, which is available to authorized users.
N. Saleh (&)
Department of Chemistry, College of Science,
United Arab Emirates University, P.O. Box 15551, Al-Ain,
United Arab Emirates
e-mail: [email protected]
S. M. Ajeb � A. Sham � S. F. AbuQamar (&)
Department of Biology, College of Science,
United Arab Emirates University, P.O. Box 15551, Al-Ain,
United Arab Emirates
e-mail: [email protected]
123
J Incl Phenom Macrocycl Chem
DOI 10.1007/s10847-013-0352-8
Author's personal copy
cytotoxicity and in vivo toxicity and can be isolated in
different sizes (n = 5, 6, 7, 8, and 10) [11, 12]. Among
these containers, CB5 and CB7 have excellent solubility in
water [13].
Recently, the use of CB macrocycles in biomedical fields
has attracted attention due to their distinct properties when
compared to other classes of delivery vehicles such as den-
drimers, nanoparticles, or carbon nanotubes [2]. For example,
CB vehicles have the ability to embed the drug within their
cavity, maintaining the drug stability and in a manner where
the drug retention and release can be tuned. While liposomes
and micelles are known to sequester drugs within their
structure, they are unable to control drug release and retention.
Furthermore, CB stand out among other macromole-
cules such as cyclodextrins (CD), calixarenes, cryptands,
crown-ethers, etc., particularly in the protection and
transport of therapeutic agents, due to their high guest-
loading capacity (K values up to 1015; [14] as well as their
ability to significantly increase the pKa value of the
encapsulated ionized drug molecules (from 2 to *5 units)
[5, 15–18]. While the induced pKa shifts were utilized in
enhancing the solubility, chemical stability, and penetra-
tion effects of some commercial drugs [15, 17, 18], the
high binding affinities motivated others to functionalize CB
with the goal of improving its selectivity towards the bio-
logical targets in living systems [19, 20].
In the United States of America, Botrytis cinerea
(Pers.:Fr) is considered among the most important disease
of tomato, after Alternaria solani, Phytophthora infestans,
and Pseudomonas sp. This fungal pathogen causes the grey
mold disease over a broad host range [21–23]. Applications
of chemical fungicides are common control strategies to
control B. cinerea. Previous reports have shown that this
fungus has developed resistance against benzimidazole
(BZ) and dicarboximide fungicides [24, 25]. New chemical
classes of fungicides, such as fluopyram, fenhexamid, bo-
scalid, fludioxonil, and strobilurins, have been labeled for
control against B. cinerea [26]. Therefore, the principal
means of disease management is often the use of
fungicides.
In the present work, host–guest approaches using CB
were implemented to enhance the activity of aqueous
solution of the fungicide fuberidazole, FBZ (see chemical
structures in Fig. 1) [27] towards the model fungus
B. cinerea at submicromolar levels. The test on FBZ,
which belongs to BZ family of compounds, was performed
in vitro using B. cinerea as a pathogen in combination with
cucurbituril (CB) containers CB5, CB7 and CB8. Inter-
estingly, the observed enhancement on FBZ activity
against B. cinerea in the present study was achieved using
a non-conventional approach that is based on supramo-
lecular interactions.
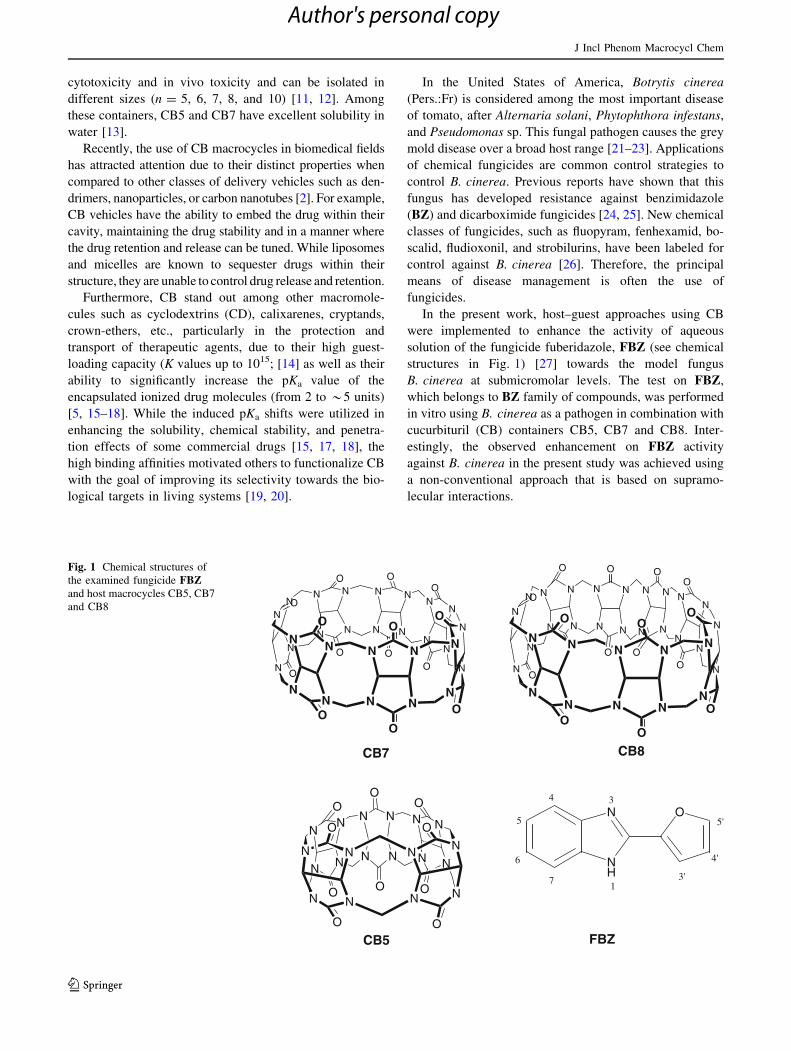
N
N
NNO
N
O
N NN
O
NO
O
N N
N
N
N N
O
N
N
N
NO
N
NN
O
N N
N
N
O
N
O
N
O
N
N
N
O
O
N
O O
CB8
N
N
NN
O
O
O
N
O
N NNO
NO
O
N N
N
N
N N
ON
N
N
NON
NN
O
N N
N
N
O
N
O
O
N
O
CB7
NH
N O
FBZ
4'
5'
1
34
5
6
7 3'
N
N
N N
O
O
O
O
NO
N
N
O
N N
O
N
N
O
O
NNNNN
N
NN
O
N
CB5
Fig. 1 Chemical structures of
the examined fungicide FBZand host macrocycles CB5, CB7
and CB8
J Incl Phenom Macrocycl Chem
123
Author's personal copy
Results
Absorption spectroscopy and determination
of host–guest binding affinities
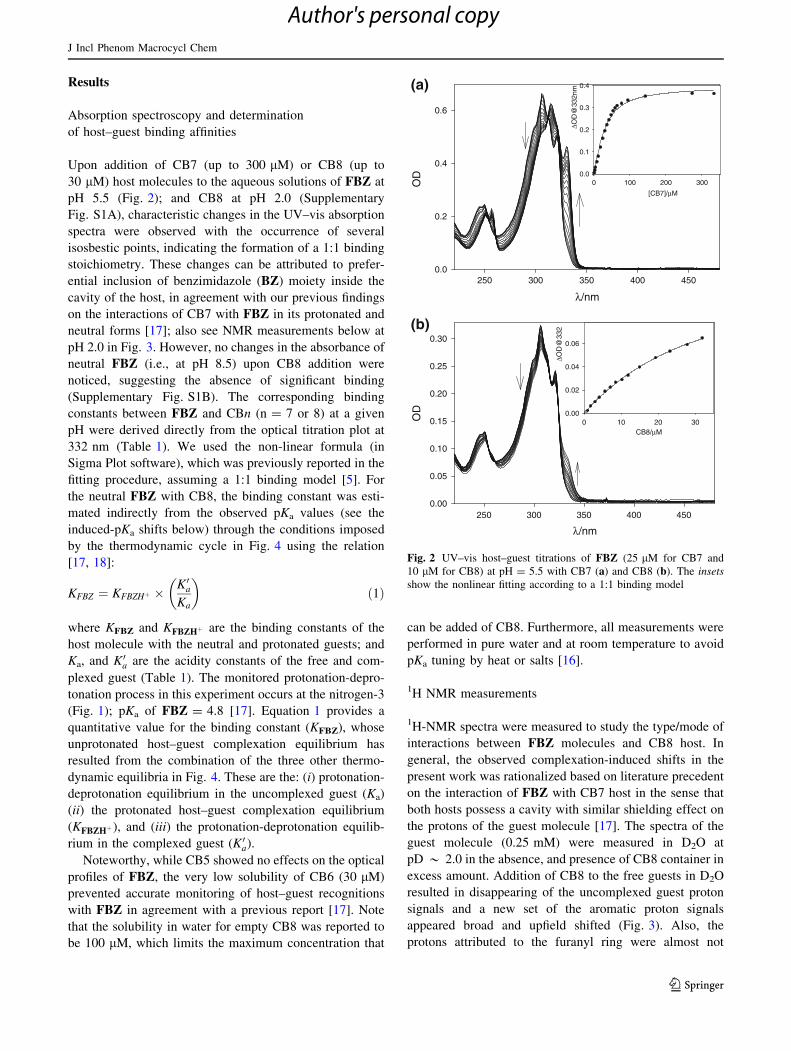
Upon addition of CB7 (up to 300 lM) or CB8 (up to
30 lM) host molecules to the aqueous solutions of FBZ at
pH 5.5 (Fig. 2); and CB8 at pH 2.0 (Supplementary
Fig. S1A), characteristic changes in the UV–vis absorption
spectra were observed with the occurrence of several
isosbestic points, indicating the formation of a 1:1 binding
stoichiometry. These changes can be attributed to prefer-
ential inclusion of benzimidazole (BZ) moiety inside the
cavity of the host, in agreement with our previous findings
on the interactions of CB7 with FBZ in its protonated and
neutral forms [17]; also see NMR measurements below at
pH 2.0 in Fig. 3. However, no changes in the absorbance of
neutral FBZ (i.e., at pH 8.5) upon CB8 addition were
noticed, suggesting the absence of significant binding
(Supplementary Fig. S1B). The corresponding binding
constants between FBZ and CBn (n = 7 or 8) at a given
pH were derived directly from the optical titration plot at
332 nm (Table 1). We used the non-linear formula (in
Sigma Plot software), which was previously reported in the
fitting procedure, assuming a 1:1 binding model [5]. For
the neutral FBZ with CB8, the binding constant was esti-
mated indirectly from the observed pKa values (see the
induced-pKa shifts below) through the conditions imposed
by the thermodynamic cycle in Fig. 4 using the relation
[17, 18]:
KFBZ ¼ KFBZHþ �K 0aKa
� �ð1Þ
where KFBZ and KFBZHþ are the binding constants of the
host molecule with the neutral and protonated guests; and
Ka, and K 0a are the acidity constants of the free and com-
plexed guest (Table 1). The monitored protonation-depro-
tonation process in this experiment occurs at the nitrogen-3
(Fig. 1); pKa of FBZ = 4.8 [17]. Equation 1 provides a
quantitative value for the binding constant (KFBZ), whose
unprotonated host–guest complexation equilibrium has
resulted from the combination of the three other thermo-
dynamic equilibria in Fig. 4. These are the: (i) protonation-
deprotonation equilibrium in the uncomplexed guest (Ka)
(ii) the protonated host–guest complexation equilibrium
(KFBZHþ ), and (iii) the protonation-deprotonation equilib-
rium in the complexed guest (K 0a).
Noteworthy, while CB5 showed no effects on the optical
profiles of FBZ, the very low solubility of CB6 (30 lM)
prevented accurate monitoring of host–guest recognitions
with FBZ in agreement with a previous report [17]. Note
that the solubility in water for empty CB8 was reported to
be 100 lM, which limits the maximum concentration that
can be added of CB8. Furthermore, all measurements were
performed in pure water and at room temperature to avoid
pKa tuning by heat or salts [16].
1H NMR measurements
1H-NMR spectra were measured to study the type/mode of
interactions between FBZ molecules and CB8 host. In
general, the observed complexation-induced shifts in the
present work was rationalized based on literature precedent
on the interaction of FBZ with CB7 host in the sense that
both hosts possess a cavity with similar shielding effect on
the protons of the guest molecule [17]. The spectra of the
guest molecule (0.25 mM) were measured in D2O at
pD * 2.0 in the absence, and presence of CB8 container in
excess amount. Addition of CB8 to the free guests in D2O
resulted in disappearing of the uncomplexed guest proton
signals and a new set of the aromatic proton signals
appeared broad and upfield shifted (Fig. 3). Also, the
protons attributed to the furanyl ring were almost not
λ/nm
λ/nm
250 300 350 400 450
OD
0.0
0.2
0.4
0.6
[CB7]/μM
0 100 200 300
Δ OD
@33
2nm
0.0
0.1
0.2
0.3
0.4(a)
250 300 350 400 450
OD
0.00
0.05
0.10
0.15
0.20
0.25
0.30
CB8/μM0 10 20 30
ΔOD
@33
2
0.00
0.02
0.04
0.06
(b)
Fig. 2 UV–vis host–guest titrations of FBZ (25 lM for CB7 and
10 lM for CB8) at pH = 5.5 with CB7 (a) and CB8 (b). The insets
show the nonlinear fitting according to a 1:1 binding model
J Incl Phenom Macrocycl Chem
123
Author's personal copy
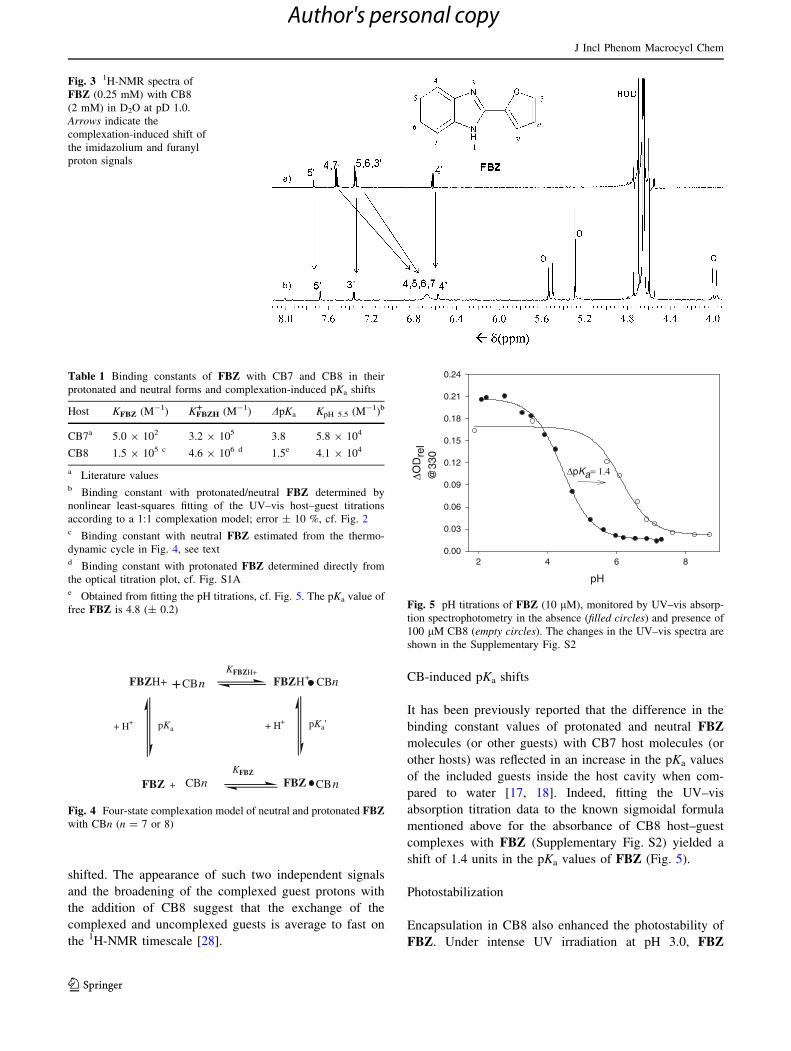
shifted. The appearance of such two independent signals
and the broadening of the complexed guest protons with
the addition of CB8 suggest that the exchange of the
complexed and uncomplexed guests is average to fast on
the 1H-NMR timescale [28].
CB-induced pKa shifts
It has been previously reported that the difference in the
binding constant values of protonated and neutral FBZ
molecules (or other guests) with CB7 host molecules (or
other hosts) was reflected in an increase in the pKa values
of the included guests inside the host cavity when com-
pared to water [17, 18]. Indeed, fitting the UV–vis
absorption titration data to the known sigmoidal formula
mentioned above for the absorbance of CB8 host–guest
complexes with FBZ (Supplementary Fig. S2) yielded a
shift of 1.4 units in the pKa values of FBZ (Fig. 5).
Photostabilization
Encapsulation in CB8 also enhanced the photostability of
FBZ. Under intense UV irradiation at pH 3.0, FBZ
Fig. 3 1H-NMR spectra of
FBZ (0.25 mM) with CB8
(2 mM) in D2O at pD 1.0.
Arrows indicate the
complexation-induced shift of
the imidazolium and furanyl
proton signals
FBZH+ CBnKFBZH+
FBZ CBn
+ H+ + H+
KFBZ
FBZH+
+
pKa pKa'
FBZ
+ CBn
CBn
Fig. 4 Four-state complexation model of neutral and protonated FBZwith CBn (n = 7 or 8)
pH
2 4 6 8
ΔOD
rel
@33
0
0.00
0.03
0.06
0.09
0.12
0.15
0.18
0.21
0.24
ΔpKa= 1.4
Fig. 5 pH titrations of FBZ (10 lM), monitored by UV–vis absorp-
tion spectrophotometry in the absence (filled circles) and presence of
100 lM CB8 (empty circles). The changes in the UV–vis spectra are
shown in the Supplementary Fig. S2
Table 1 Binding constants of FBZ with CB7 and CB8 in their
protonated and neutral forms and complexation-induced pKa shifts
Host KFBZ (M-1) KFBZH1 (M-1) DpKa KpH 5.5 (M-1)b
CB7a 5.0 9 102 3.2 9 105 3.8 5.8 9 104
CB8 1.5 9 105 c 4.6 9 106 d 1.5e 4.1 9 104
a Literature valuesb Binding constant with protonated/neutral FBZ determined by
nonlinear least-squares fitting of the UV–vis host–guest titrations
according to a 1:1 complexation model; error ± 10 %, cf. Fig. 2c Binding constant with neutral FBZ estimated from the thermo-
dynamic cycle in Fig. 4, see textd Binding constant with protonated FBZ determined directly from
the optical titration plot, cf. Fig. S1Ae Obtained from fitting the pH titrations, cf. Fig. 5. The pKa value of
free FBZ is 4.8 (± 0.2)
J Incl Phenom Macrocycl Chem
123
Author's personal copy

decomposed three times more slowly in their complexes
with CB8 than in their free forms (Fig. 6). In comparison,
the photostability of FBZ by CB7 was increased by a factor
of 7 in a precedent report.
In vitro assays
The inhibitory effect of FBZ and its inclusion complex
solutions with CB5, CB7 and CB8 on the mycelia growth
of B. cinerea after 5 days of incubation was illustrated in
Fig. 7 and Supplementary Fig. S3. At pH 5.5, distilled
water (H2O) was used as a negative control treatment (no
FBZ, no CB), the upper panel (right) was the host CB5/7/8
control at 100 lM (Fig. 7a, c; Supplementary Fig. S3a),
and the lower panel (left) was the guest positive control,
FBZ, at 200 lM. The lower panel (right) represents those
treated with CB5/7/8-guest inclusion complex solutions
with micromolar ratios of CB5/7/8@FBZ of 100:200
(Supplementary Fig. S3a; Fig. 7a, c). The inclusion com-
plex of CB5/7/8-guest solutions with micromolar ratios of
CB5/7@FBZ of 5:10, 100:50, 100:100 and 100:200 or
CB8@FBZ of 5:10, 25:50, 50:100, 100:200, 100:100 and
100:50 were tested after 72 h of incubation with B. cinerea
250 300 350 400 450 500
OD
0.00
0.05
0.10
0.15
0.20
0.25
0.30
Time/secs
0 200 400 600 800
f
0.0
0.1
0.2
0.3
0.4
0.5
0.6
X 3
λ /nm
Fig. 6 Photodecomposition of FBZ in the absence (dashed lines)
and presence of CB8 (100 lM, solid lines), followed through the
decrease of the UV absorption band with increasing time of
photoirradiation by UV lamps (300 nm) in a photoreactor (40 min).
In the inset, the characteristic photodecomposition decay function,
f = log [(10Ao - 1)/(10A - 1)], is plotted versus time in seconds
and from the slope the relative photostability was determined by
taking into consideration the molar absorptivity of the CB8-bound
and free FBZ at the excited wavelength before irradiations
Fig. 7 Inhibitory growth effect of inclusion complexes on Botrytis
cinerea. B. cinerea inhibition growth using the inclusion complex
CB7@FBZ (a) and CB8@FBZ (c). Growth inhibition (Y %) on B.
cinerea using inclusion complex solutions CB7@FBZ (b) and
CB8@FBZ (d). In a and c, photos were taken 5 days after
inoculation. In b and d, data were collected 72 h after inoculation.
CB7/8, 100 lM CB7/8; FBZ, 200 lM FBZ; CB7/8:FBZ, 100:200
lM CB7/8:FBZ
J Incl Phenom Macrocycl Chem
123
Author's personal copy

(Supplementary Fig. S3b; Fig. 7b, d). The fungus clearly
grew covering almost the entire surface of the 2xV8
medium in the negative control and any of the containers
(Fig. 7; Supplementary Fig. S3). In the positive control,
although fungal growth could be observed, changes were
minimal and dose-dependent, where a clear fungal semi-
circle was observed on the plates as the concentration of
the FBZ increased from 10 to 200 lM, and the inhibition
rate increased as the FBZ concentration increased. Treat-
ments of the mixtures of CB7 or CB8 containers, but not
CB5, with FBZ at any of the micromolar ratios increased
growth inhibition rate of the fungus (Fig. 7; Supplementary
Fig. S3). The growth inhibition for each of the adminis-
tered treatments in pure water at pH 5.5 was tested. While
no enhancement in inhibition activity of FBZ of B. cinerea
was observed upon CB5 addition (Supplementary Fig. S3),
growth inhibition was increased up to 16–18 % with the
addition of other hosts, CB7 and CB8 (Fig. 7b, d). The
CB7/CB8-bound FBZ inhibited the growth of B. cinerea at
least three times when compared with that of the same
concentration of the unbound fungicides (Fig. 7). Not
surprisingly, the empty CB containers themselves were
completely inactive. It was also observed that the inhibition
activity was dependent on the administrated micromolar
ratio of the host:guest molecules with the highest inhibition
rate at 1:2 ratio compared to that at the 2:1 and 1:1 ratio in
both CB8@FBZ and CB7@FBZ systems, as compared
with their corresponding empty containers or fungicide
controls alone (Fig. 7). In addition, lowering the concen-
tration of either hosts, CB7 or CB8 container, and the guest
fungicide (FBZ) by 20 times from 200:100 to 10:5 lM
lowered the activity of the fungicide composite by a factor
of 0.75 only. Altogether, this suggests that the growth
inhibitory effect of FBZ increased in a dose-dependent
manner when FBZ is coupled with either CB7 or CB8
container, but not CB5.
To determine the pH effect of FBZ on B. cinerea
growth, we tested the maximum dose of the fungicide
(200 lM) used in previous experiments (Methods) at pH
3.0 and 5.5 (Table 2). The negative controls were also
tested at the same pH values towards B. cinerea growth. As
expected, the negative control at any of the pH values
showed a complete mycelium-covered area to the greatest
extent compared with the positive control samples
(Table 2). Also, there was no significant inhibition effect of
FBZ treatments at either of the pH tested. Altogether, this
suggests that while the inhibitory effect of FBZ on
B. cinerea is dose-dependent, it is not pH-dependent.
Noteworthy, the change of pH from 3.0 to 5.5 means that
the FBZ fungicide has changed its structure from proton-
ated to neutral forms (pKa of FBZ = 4.8, Table 1 and
Fig. 5). Consequently, the similar inhibition effect at the
two selected pH also means that changing the acidic-basic
properties has no influence on the fungicide activity against
B. cinerea at least in the meaningful pH range.
Discussion
In the present study, we have employed the CB host–guest
approach in vitro. Our in vitro findings demonstrated that
the antifungal activity toward B. cinerea of CB7@FBZ
and CB8@FBZ were quite similar and enhanced up to
three times with the formation of CB7/CB8 host–guest
complexes at pH 5.5 (and pH 3.0) when compared to the
free FBZ. Furthermore, the inhibition activity was found to
depend on the administrated molar ratio of the guest and
host molecules. Noteworthy, lowering the concentration by
20 times from 200:100 to 10:5 micromolar ratio of the
CB7/CB8@FBZ complexes had negligible effects on the
activity of the composite.
From the measured UV–vis titration data (see ‘‘Results’’
section), the CB8@FBZ system has a 1:1 stoichiometry
and not other ratios. This could be attributed to the size
complementary of the guests to the cavity volume of CB8
[27]. Other things being equal, the binding constants
become highest when the ratios of the volumes to the
guests with the inner cavity volume of CB are close to
0.5529 [27]. In addition, it has also been previously
reported [27, 28] that the release of (high energy) water
molecules from the cavity of CB plays a particularly
important role for the binding of guest molecule. The
results in Table 2 also demonstrated that the protonated
and neutral forms of FBZ have similar activity towards
B. cinerea. Thus, the induced pKa shifts do not seem to
rationalize the trends in the observed data (Fig. 7 and
Supplemetary Fig. S3). For example, the population of the
protonated FBZ is manifested in the degree of ionization,
which is calculated at pH 5.5 to be 99 % and 86 % for FBZ
in its complexed forms with CB7 and CB8, respectively,
taking into consideration the pKa values for the free and
complexed FBZ (Table 1) and using the relation:
a ¼ 1
1þ 10pH�pKað2Þ
Although the precise mechanism of action of the
FBZ@CB7/8 complexes needs further experiments, one
Table 2 pH effect of FBZ on growth Botrytis cinerea. Growth
inhibition (Y %) of FBZ solution (200 lM)
pH Solution (lM) Y (%)
3.0 H2O 0
FBZ 6.36
5.5 H2O 0
FBZ 6.01
J Incl Phenom Macrocycl Chem
123
Author's personal copy

possibility is that the macrocyclic complex actually binds
more tightly to the biological targets (particularly proteins)
[2]. Nau and co-workers [7], has shown that the binding
affinities of a drug molecule with the target site can be
enhanced (with a concomitant increase in the target
specificity) in the presence of a CB macrocycle. The
binding events were monitored through changes in the
fluorescence intensity and lifetime of the drug, providing a
proof-of-principle for the formation of suprabiomolecular
ternary complexes that might potentially be useful in the
context of targeted drug delivery and drug specifity. Note that,
CB7 and CB8 bind FBZ with comparable affinities under the
in vitro condition (pH 5.5; Table 1), which explain their
similar activity against B. cinerea. Yet, in vivo experiments
on this or other systems are needed to prove the formation of a
stable ternary complex under the in vivo condition (pH
7.0–7.5), which are ongoing in our laboratories.
We have used B. cinerea as a model pathogen in our
study, which is a fungus with a broad host range. This
‘‘economic’’ pathogen reduces crop quality and produc-
tivity under diverse production conditions. Application of
chemical fungicides is a common control strategy against
B. cinerea. Noteworthy, B. cinerea develops resistance to
some fungicides, which severely limits chemical control
options [24, 25]. Nowadays, fluopyram, iprodione, fen-
hexamid, boscalid, fludioxonil, various triazoles and stro-
bilurins, are widely used fungicides to control B. cinerea
[26]. The selected BZ derivative in this work (e.g. FBZ) is
broadly used as efficient fungicides particularly as pre- or
post-harvest fungicides to control plant diseases on a wide
variety of fruits, vegetables or other field crops in Asia
[29]. We have demonstrated that the CB-complexation of
FBZ enhances the solubility (e.g., three times with CB7)
and increases the photostability by seven and three times in
the current results with CB7 and CB8, respectively [17].
The present results are, thus, important to BZ antifungal
action as the active concentration (10:5 lM) should be less
harmful to human health than the current uses in crop
treatment. Moreover, the enhanced photostability furbishes
another evidence for the host–guest complex formation and
confirms that the new non-conventional methods have
additional benefits to crop protections.
CB hosts bind the FBZ guest molecules via hydrophobic
and ion–dipole interactions. The glycoluril core at the CB
equator provides a hydrophobic environment for the bind-
ing of neutral forms, while the ion–dipole interactions
occur between the ionizable hydrogen atoms on the nitro-
gen-3 of the FBZ guests and the oxygens at the portals of
the CB [17]. Recently, it has been reported that the release
of water molecules from the cavity of CB plays a signifi-
cant role in the binding of guest molecules [30]. In addi-
tion, the size complementary of the guests with the cavity
volume of CB is manifested in the absolute binding con-
stants of the CB@guest complexes [31]. Such noncovalent
encapsulation inside the nonpolar cavity of CB was
reported to modify the physical and chemical properties of
the guest molecules due to the isolation and confinement of
the guest molecules from the surrounding water medium.
For example, it was reported that CB7 can induce deag-
gregation and photostabilization of some fluorescent dyes
[32]. In addition to the modification of the physical prop-
erties of the guest molecules, CB was demonstrated to
modify the chemical reactivities of the guest molecules for
different applications. As an example, CB can modify the
protonation-deprotonation equilibria of the guest mole-
cules. The carbonyl at the ureidyl portals of CB biases its
negative dipoles towards the positive charges of the guest
over neutral species through the ion–dipole interactions.
Thus, CB hosts, preferentially, bind the protonated species
of a guest molecule with respect to its neutral form and,
consequently, shift the protonation–deprotonation equilib-
ria towards higher pH, the so-called ‘CBs-induced pKa
shifts’. While the induced pKa shifts were utilized in
enhancing the solubility, chemical stability, and penetration
effects of some commercial drugs [2], the present studies
demonstrated no influence of the induced pKa shifts on the
FBZ activity against the fungus in vitro. Utilization the
CB-induced pKa shifts was planned from the beginning, as
it may be desirable to trigger the release of drugs by
shifting the equilibrium towards the uncomplexed drug in
dependence on a certain stimulus in a spatially and/or
temporally controlled manner. As the neutral forms of the
FBZ bind much more weakly with CB than their corre-
sponding protonated forms, a change in the pH of the
medium from below pK 0a (the pKa value of the complex) to
above pK 0a effectively decreases the binding constants of
the drugs and the subsequent rapid release of the encap-
sulated drugs shifts the chemical equilibrium toward the
uncomplexed guest (and host). Note that this concept can
be advantageously employed at different stages of FBZ
drug delivery to plant cells. For example, FBZ@CB
complexes which have pK 0a values between 6 and 9 could
be manufactured below pH 4 (because of the higher bind-
ing constants of the protonated forms and related higher
degree of complexation) while the physiological pH itself
(e.g., pH 7.0–7.5 for the cytoplasm of higher plant cells)
could subsequently favor the efficient release of the drug
molecules into the biological targets. On the contrary, the
CB-induced pKa shifts and the accompanied tuning in the
CB7/8 affinity towards FBZ in its neutral state at alkaline
pH (Supplementary Fig. S1 lower panel) might become
disadvantageous in the context of the need for highly stable
host–guest CB@FBZ complexes that could sustain com-
petitive displacements by the cytoplasmic compartments.
J Incl Phenom Macrocycl Chem
123
Author's personal copy

Therefore, research is undergoing to understand the
relationship between tomato-B. cinerea and the inclusion
complex in vivo, yet this new suggested crop treatment using
a novel host–guest approach in our in vitro findings should
have a lot of potential in the agrochemical fungicide busi-
ness. In particular, CB hosts have unique supramolecular
recognition properties, which offer numerous advantages
over the use of conventional macrocycles. These include
ultrahigh-affinity binding, slow and tunable complexation
kinetics, pKa shifts in favor of the binding of the protonated
forms of guests, and an increased photochemical and
chemical stability of encapsulated guests. While these
properties are well recognized, their implementation in drug
delivery systems has rapidly unfolded during the past
10 years. Among the different supramolecular host mole-
cules, CD [33] have been most widely used for the formu-
lation of therapeutic and diagnostic agents in pharmacology.
However, their use in clinical conditions is generally limited
to oral and topical drug delivery forms, since they are
nephrotoxic if they enter the body in a nonmetabolized state
[34]. In addition, the poor selectivity and low binding
affinities (Kbinding \ 104 M-1) [35] of nonderivatized (nat-
ural) CD towards guest molecules in general, and drug
molecules in particular, requires excess concentrations of
CD to form quantitative CD@guest complexes. Several
alternative macrocyclic host molecules have been suggested
in the literature to have potential as drug delivery vehicles,
for example derivatized calixarenes [36], but not has yet
found actual applications. Thus, CB hosts hold already now
more prospects for drug delivery than calixarenes, and they
are expected to rival CD in many areas.
Conclusions
This study presents an innovative way to enhance the
activity of fungicides against microbes, utilizing the host–
guest non-covalent interactions. Inclusion complexes of
CB7@FBZ and CB8@FBZ enhanced the inhibitory effect
of FBZ on B. cinerea. Although this antifungal effect of
FBZ was improved when mixed with the CB containers
in vitro, additional research is required to elucidate the
specific mode of action and to understand the conditions
that facilitate the performance of these inclusion complexes
and their efficacy.
Experimental section
Chemicals and equipment
CB5, CB7 and CB8 (purity [99.9 %) were purchased and
used as recommended (Sigma-Aldrich). The FBZ was also
purchased from Sigma-Aldrich and used without further
purification. Millipore water was used for dilution. The
UV–vis spectra were measured on Cary-50 instrument
(Varian). All NMR spectra were performed on a Varian
400 MHz spectrometer in D2O. All 1H-NMR spectra are
referenced in ppm with respect to a TMS standard. The pH
values of the solutions were adjusted (±0.2 units) by
adding adequate amounts of HCl (DCl) or NaOH (NaOD)
and recorded using a pH meter (WTW 330i equipped with
a WTW SenTix Mic glass electrode). The photostabiliza-
tion of FBZ was monitored using the change in the
absorbance of the free and complexed fungicides at pH 3.0
upon irradiation in a Luzchem LZC-4V photoreactor
equipped with UVB lamps (kex = 300 nm). Also, the
average power intensity was kept at 12 mW cm-2 during
irradiation.
Fungal culture and antifungal activity determination
Botrytis cinerea strain BO5-10 was grown on 2xV8 agar
(36 % V8 juice, 0.2 % CaCO3, 2 % Bacto-agar, Becton,
Dickinson and Company, Sparks, MD USA). Fungal cul-
tures were initiated and subcultured by transferring pieces
of agar containing mycelium to fresh 2xV8 agar and
incubated at 23 ± 2 �C as described [37].
The in vitro fungicidal activities against B. cinerea were
tested according to [9] with some modifications. An
aqueous solution of FBZ was prepared with a concentra-
tion of 10, 50, 100, or 200 lM. The aqueous solution of
CB5, CB7, or CB8 was prepared with a concentration of 5,
25, 50 or 100 lM. Solutions of CB mixed with FBZ in a
ratio of 1:2, 1:1 and 2:1 were prepared in pure water at pH
of 5.5. Inocula of a diameter of 8-mm were removed from
the margins of actively growing colonies of mycelium, and
placed on one side of a Petri dish on 2xV8 medium as
described above. Filter papers (6 mm in diameter) were
placed on the opposite side of the dish, and soaked onto the
solutions of CB5, CB7, CB8, FBZ (15 lL) or each of the
previously prepared inclusion complex (sterilized water
was used as a negative control). Plates were then covered,
sealed with parafilm, and incubated for 72 h in a 25 �C
incubator. Each treatment was prepared in four replicates.
The radius of the mycelium was measured at 72 h. The
growth inhibition rate was calculated according to the
growth inhibition (Y) formula developed previously [38].
Acknowledgments The authors would like to acknowledge the
Office of Research Support and Sponsored Projects at UAEU for their
support under the grant numbers 31S075 to S.AQ. and 31S074 to
N.S., within the framework of National Research Foundation (NRF)
funding program, the Interdisciplinary fund number 31S035 to S.AQ.
We would also like to thank Prof. Dr. Werner Nau and his research
group for their significant contributions in previous joint accom-
plishments on the recognitions of benzimidazole fungicides by CB in
J Incl Phenom Macrocycl Chem
123
Author's personal copy

aqueous solutions. We thank Ms. Zeinab A. Abdalla for her contri-
bution in the initiation of this study.
References
1. Cram, D.J., Cram, J.M.: Container Molecules and Their Guests.
Royal Society of Chemistry, Cambridge (1997)
2. Ghosh, I., Nau, W.M.: The strategic use of supramolecular pKa
shifts to enhance the bioavailability of drugs. Adv. Drug Deliv.
Rev. 64(9), 764–783 (2012)
3. Day, A.I., Collins, J.G.: Cucurbituril receptors and drug delivery.
In: Gale, P.A., Steed, J.W. (eds.) Supramolecular Chemistry:
From Molecules to Nanomaterials, vol. 3, pp. 983–1000. Wiley,
New York (2012)
4. Saleh, N., Khaleel, A., Al-Dmour, H., al-Hindawi, B., Yak-
ushenko, E.: Host-guest complexes of cucurbit[7]uril with
albendazole in solid state—thermal and structural properties.
J. Therm. Anal. Calorim. 111(1), 385–392 (2013)
5. Saleh, N., Al-Soud, Y.A., Al-Kaabi, L., Ghosh, I., Nau, W.M.: A
coumarin-based fluorescent PET sensor utilizing supramolecular
pKa shifts. Tetrahedron Lett. 52(41), 5249–5254 (2011)
6. Saleh, N., Al-Rawashdeh, N.A.F.: Fluorescence enhancement of
carbendazim fungicide in cucurbit[6]uril. J. Fluoresc. 16(4),
487–493 (2006)
7. Bhasikuttan, A.C., Mohanty, J., Nau, W.M., Pal, H.: Efficient
fluorescence enhancement and cooperative binding of an organic
dye in a supra-biomolecular host-protein assembly. Angew.
Chem. Int. Ed. 46(22), 4120–4122 (2007)
8. Barooah, N., Mohanty, J., Pal, H., Bhasikuttan, A.C.: Supramo-
lecular assembly of hoechst-33258 with cucurbit[7]uril macro-
cycle. Phys. Chem. Chem. Phys. 13(28), 13117–13126 (2011)
9. Liu, H., Wu, X., Huang, Y., He, J., Xue, S.F., Tao, Z., Zhu, Q.J.,
Wei, G.: Improvement of antifungal activity of carboxin by
inclusion complexation with cucurbit[8]uril. J. Incl. Phenom.
Macro. Chem. 71(3–4), 583–587 (2011)
10. Masson, E., Ling, X., Joseph, R., Kyeremeh-Mensah, L., Lu, X.:
Cucurbituril chemistry: a tale of supramolecular success. RSC
Adv. 2(4), 1213–1247 (2012)
11. Hettiarachchi, G., Nguyen, D., Wu, J., Lucas, D., Ma, D., Isaacs,
L., Briken, V.: Toxicology and drug delivery by cucurbit[n]uril
type molecular containers. PLoS ONE 5(5), e10514 (2010)
12. Uzunova, V.D., Cullinane, C., Brix, K., Nau, W.M., Day, A.I.: Tox-
icity of cucurbit[7]uril and cucurbit[8]uril: an exploratory in vitro and
in vivo study. Org. Biomol. Chem. 8(9), 2037–2042 (2010)
13. Bardelang, D., Udachin, K.A., Leek, D.M., Margeson, J.C., Chan,
G., Ratcliffe, C.I., Ripmeester, J.A.: Cucurbit[n]urils (n = 5–8): a
comprehensive solid state study. Cryst. Growth Des. 11(12),
5598–5614 (2011)
14. Moghaddam, S., Yang, C., Rekharsky, M., Ko, Y.H., Kim, K.,
Inoue, Y., Gilson, M.K.: New ultrahigh affinity host-guest com-
plexes of cucurbit[7]uril with bicyclo[2.2.2]octane and adaman-
tane guests: thermodynamic analysis and evaluation of M2 affinity
calculations. J. Am. Chem. Soc. 133(10), 3570–3581 (2011)
15. Saleh, N., Koner, A.L., Nau, W.M.: Activation and stabilization
of drugs by supramolecular pKa shifts: drug-delivery applications
tailored for cucurbiturils. Angew. Chem. Int. Ed. 47(29),
5398–5401 (2008)
16. Barooah, N., Mohanty, J., Pal, H., Bhasikuttan, A.C.: Stimulus-
responsive supramolecular pKa tuning of cucurbit[7]uril encapsu-
lated coumarin 6 dye. J Phys Chem B 116(12), 3683–3689 (2012)
17. Koner, A.L., Ghosh, I., Saleh, N., Nau, W.M.: Supramolecular
encapsulation of benzimidazole-derived drugs by cucurbit[7]uril.
Can. J. Chem. 89(2), 139–147 (2011)
18. Saleh, N., Meetani, M., Al-Kaabi, L., Ghosh, I., Nau, W.M.:
Effects of cucurbiturils on tropicamide and potential applications
in ocular drug delivery. Supramol. Chem. 23(9), 654–661 (2011)
19. Zhao, N., Lloyd, G.O., Scherman, O.A.: Monofunctionalised
cucurbit[6]uril synthesis using imidazolium host-guest complex-
ation. Chem. Commun. 48(25), 3070–3072 (2012)
20. Ma, D., Hettiarachchi, G., Duc, N., Zhang, B., Wittenberg, J.B.,
Zavalij, P.Y., Briken, V., Isaacs, L.: Acyclic cucurbit[n]uril
molecular containers enhance the solubility and bioactivity of
poorly soluble pharmaceuticals. Nat. Chem. 4(6), 503–510 (2012)
21. Javris, W.R.: Botryotinia and Botrytis species: taxonomy, phys-
iology, and pathology. Can. Dep. Agric. Monogr. No. 15 (1977)
22. Williamson, B., Duncan, G.H., Harrison, J.G., Harding, L.A.,
Elad, Y., Zimand, G.: Effect of humidity on infection of rose
petals by dry-inoculated conidia of Botrytis cinerea. Mycol. Res.
99, 1303–1310 (1995)
23. Elad, Y.: Responses of plants to infection by Botrytis cinerea and
novel means involved in reducing their susceptibility to infection.
Biol. Rev. 72(3), 381–422 (1997)
24. Yarden, O., Katan, T.: Mutations leading to substitutions at
amino-acids 198 and 200 of b-tubulin that correlate with beno-
myl-resistance phenotypes of field strains of Botrytis cinerea.
Phytopathology 83(12), 1478–1483 (1993)
25. Yourman, L.F., Jeffers, S.N.: Resistance to benzimidazole and
dicarboximide fungicides in greenhouse isolates of Botrytis
cinerea. Plant Dis. 83(6), 569–575 (1999)
26. Kanetis, L., Forster, H., Adaskaveg, J.E.: Determination of natural
resistance frequencies in penicillium digitatum using a new air-
sampling method and characterization of fludioxonil- and pyri-
methanil-resistant isolates. Phytopathology 100(8), 738–746 (2010)
27. Selms, R.C.D.: Benzimidazoles. I. 2-(Heterocyclic Substi-
tuted)benzimidazoles. J. Org. Chem. 27(6), 2163–2165 (1962)
28. Tang, H., Fuentealba, D., Ko, Y.H., Selvapalam, N., Kim, K., Bohne,
C.: Guest binding dynamics with cucurbit[7]uril in the presence of
cations. J. Am. Chem. Soc. 133(50), 20623–20633 (2011)
29. Jenkyn, J.F., Prew, R.D.: Activity of 6 fungicides against cereal
foliage and root diseases. Ann. Appl. Biol. 75(2), 241–252 (1973)
30. Biedermann, F., Uzunova, V.D., Scherman, O.A., Nau, W.M., De
Simone, A.: Release of high-energy water as an essential driving
force for the high-affinity binding of cucurbit[n]urils. J. Am.
Chem. Soc. 134(37), 15318–15323 (2012)
31. Nau, W.M., Florea, M., Assaf, K.I.: Deep inside cucurbiturils:
physical properties and volume of their inner cavity determine the
hydrophobic driving forces for host-guest complexation. Israel J.
Chem. 51, 559 (2011)
32. Mohanty, J., Nau, W.M.: Ultrastable rhodamine with cucurbituril.
Angew. Chem. Int. Ed. 44(24), 3750–3754 (2005)
33. Li, J., Loh, X.J.: Cyclodextrin-based supramolecular architec-
tures: syntheses, structures, and applications for drug and gene
delivery. Adv. Drug Delivery Rev. 60(9), 1000–1017 (2008)
34. Shchepotina, E., Pashkina, E., Yakushenko, E., Kozlov, V.: Cu-
curbiturils as containers for medicinal compounds. Nanotechnol.
Russ. 6(11), 773–779 (2011)
35. Rekharsky, M.V., Inoue, Y.: Complexation thermodynamics of
cyclodextrins. Chem. Rev. 98(5), 1875–1917 (1998)
36. Da Silva, E., Lazar, A.N., Coleman, A.W.: Bioharmaceuticalapplications of calixarenes. J. Drug Deliv. Sci. Technol. 14(1),
3–20 (2004)
37. AbuQamar, S., Luo, H., Laluk, K., Mickelbart, M.V., Mengiste,
T.: Crosstalk between biotic and abiotic stress responses in
tomato is mediated by the AIM1 transcription factor. Plant J.
58(2), 347–360 (2009)
38. Ware, G.W.: Fundementals of pesticides: a self-instruction guide.
Thomson publication, Frensno (1986)
J Incl Phenom Macrocycl Chem
123
Author's personal copy




















