
Enhanced Osteoclastogenesis in 4-1BB?Deficient Mice Caused by Reduced Interleukin10
6
Enhanced Osteoclastogenesis in 4-1BB–Deficient Mice Caused by Reduced Interleukin-10 Hyun-Hee Shin, Ji-Eun Lee, Eun A Lee, Byoung Se Kwon, and Hye-Seon Choi ABSTRACT: Enhanced osteoclastogenesis was observed in bone marrow–derived macrophage cells from 4-1BB–deficient mice than in those from wildtype mice. 4-1BB and 4-1BB ligand interaction may play a role at a certain stage of osteoclast formation through increased level of IL-10, a negative regulator of osteoclas- togenesis. Introduction: 4-1BB is an inducible T-cell costimulatory molecule and a member of the TNF receptor family. The expression pattern of 4-1BB and 4-1BB ligand (4-1BBL) has suggested that 4-1BB plays a role not only in various responses related to innate immunity but also in bone metabolism. Materials and Methods: Osteoclast formation was evaluated in bone marrow–derived macrophage cells (BMMs) from wildtype and 4-1BB–deficient (4-1BB -/- ) mice. Expression of interleukin-10 (IL-10) during osteoclast formation was analyzed at the mRNA and protein levels. Results: Expression of IL-10 was higher in RANKL-stimulated wildtype BMMs than 4-1BB -/- BMMs. When 4-1BBL was stimulated with 4-1BB–Fc fusion protein, the expression of IL-10 in BMMs increased. Neutral- ization of IL-10 was not as effective in preventing inhibition by IL-10 of osteoclast differentiation in 4-1BB -/- BMMs as in wildtype BMMs. When IL-10 was added to the culture medium, osteoclast formation was inhibited more efficiently in the 4-1BB -/- BMMs than in the wildtype BMMs. Conclusions: Interaction of 4-1BB and 4-1BBL stimulates IL-10 production through 4-1BBL signaling. 4-1BBL plays a role at a certain stage of osteoclast formation, and IL-10 may mediate this effect. The elevated level of osteoclastogenesis in 4-1BB -/- BMMs may thus be caused, in part, by a lower level of IL-10. J Bone Miner Res 2006;21:1907–1912. Published online on August 21, 2006; doi: 10.1359/JBMR.060813 Key words: osteoclastogenesis, 4-1BB, 4-1BB ligand, interleukin-10, bone marrow–derived macrophage INTRODUCTION O STEOCLASTS, THE CELLS responsible for bone resorp- tion, differentiate from hematopoietic stem cells. Os- teoclast precursors have properties in common with precur- sors of the monocyte and macrophage cell lineage. They not only have an important role in physiological bone remod- eling, but also participate in the bone destruction associated with chronic inflammatory disease. (1) Osteoclastogenesis requires two molecules generated from bone marrow mes- enchymal cells: macrophage-colony stimulating factor (M-CSF) and RANKL, a member of the TNF family. (2) Cooperation between the M-CSF and RANKL systems generates an essential signal for osteoclast differentiation produced by an interaction between stromal cells and cells of the osteoclast lineage. (3) Osteoclastogenesis is regulated by positive and negative osteotropic factors. Most osteo- tropic factors regulating osteoclast differentiation also af- fect the immune system, indicating that the two systems are closely related. The positive factors are stimulatory cyto- kines such as interleukin (IL)-1, IL-6, BMP-7, IL-11, and TNF-, and the negative factors include IL-4, IL-10, IL-12, and interferon (IFN)-. (4) The anti-inflammatory cytokine IL-10 generated by a variety of cells, including T cells and macrophages, functions in a negative feedback loop that suppresses the generation of inflammatory cytokines and dampens the acute inflammatory response. (5) IL-10 also suppresses osteoclast formation, suggesting that it plays a role in modulating bone loss in inflammatory disor- ders. (6,7) 4-1BB is an inducible T-cell costimulatory molecule and a member of the TNF receptor (TNFR) family that includes TNFR1, TNFR2, CD40, OX40, and RANK. (8) 4-1BB is expressed mainly on activated T cells, but recent studies have shown that it is present on a variety of other cells including myeloid cells. (9) 4-1BB is also known to be a den- dritic cell–activating molecule like RANK. 4-1BBL is ex- pressed on activated antigen-presenting cells and on several leukemia cell lines. (9) The expression pattern of 4-1BB and 4-1BBL has suggested that 4-1BB plays a role in various responses related to innate immunity and bone metabolism. Therefore, we studied whether 4-1BB and 4-1BBL inter- action plays a role in osteoclastogenesis, using 4-1BB– deficient mice. The authors state that they have no conflicts of interest. Department of Biological Sciences and the Immunomodulation Research Center, University of Ulsan, Ulsan, Korea. JOURNAL OF BONE AND MINERAL RESEARCH Volume 21, Number 12, 2006 Published online on August 21, 2006; doi: 10.1359/JBMR.060813 © 2006 American Society for Bone and Mineral Research 1907
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Enhanced Osteoclastogenesis in 4-1BB?Deficient Mice Caused by Reduced Interleukin10

Enhanced Osteoclastogenesis in 4-1BB–Deficient Mice Caused byReduced Interleukin-10
Hyun-Hee Shin, Ji-Eun Lee, Eun A Lee, Byoung Se Kwon, and Hye-Seon Choi
ABSTRACT: Enhanced osteoclastogenesis was observed in bone marrow–derived macrophage cells from4-1BB–deficient mice than in those from wildtype mice. 4-1BB and 4-1BB ligand interaction may play a roleat a certain stage of osteoclast formation through increased level of IL-10, a negative regulator of osteoclas-togenesis.
Introduction: 4-1BB is an inducible T-cell costimulatory molecule and a member of the TNF receptor family.The expression pattern of 4-1BB and 4-1BB ligand (4-1BBL) has suggested that 4-1BB plays a role not onlyin various responses related to innate immunity but also in bone metabolism.Materials and Methods: Osteoclast formation was evaluated in bone marrow–derived macrophage cells(BMMs) from wildtype and 4-1BB–deficient (4-1BB−/−) mice. Expression of interleukin-10 (IL-10) duringosteoclast formation was analyzed at the mRNA and protein levels.Results: Expression of IL-10 was higher in RANKL-stimulated wildtype BMMs than 4-1BB−/− BMMs. When4-1BBL was stimulated with 4-1BB–Fc fusion protein, the expression of IL-10 in BMMs increased. Neutral-ization of IL-10 was not as effective in preventing inhibition by IL-10 of osteoclast differentiation in 4-1BB−/−
BMMs as in wildtype BMMs. When IL-10 was added to the culture medium, osteoclast formation wasinhibited more efficiently in the 4-1BB−/− BMMs than in the wildtype BMMs.Conclusions: Interaction of 4-1BB and 4-1BBL stimulates IL-10 production through 4-1BBL signaling.4-1BBL plays a role at a certain stage of osteoclast formation, and IL-10 may mediate this effect. The elevatedlevel of osteoclastogenesis in 4-1BB−/− BMMs may thus be caused, in part, by a lower level of IL-10.J Bone Miner Res 2006;21:1907–1912. Published online on August 21, 2006; doi: 10.1359/JBMR.060813
Key words: osteoclastogenesis, 4-1BB, 4-1BB ligand, interleukin-10, bone marrow–derived macrophage
INTRODUCTION
OSTEOCLASTS, THE CELLS responsible for bone resorp-tion, differentiate from hematopoietic stem cells. Os-
teoclast precursors have properties in common with precur-sors of the monocyte and macrophage cell lineage. They notonly have an important role in physiological bone remod-eling, but also participate in the bone destruction associatedwith chronic inflammatory disease.(1) Osteoclastogenesisrequires two molecules generated from bone marrow mes-enchymal cells: macrophage-colony stimulating factor(M-CSF) and RANKL, a member of the TNF family.(2)
Cooperation between the M-CSF and RANKL systemsgenerates an essential signal for osteoclast differentiationproduced by an interaction between stromal cells and cellsof the osteoclast lineage.(3) Osteoclastogenesis is regulatedby positive and negative osteotropic factors. Most osteo-tropic factors regulating osteoclast differentiation also af-fect the immune system, indicating that the two systems areclosely related. The positive factors are stimulatory cyto-kines such as interleukin (IL)-1, IL-6, BMP-7, IL-11, and
TNF-�, and the negative factors include IL-4, IL-10, IL-12,and interferon (IFN)-�.(4) The anti-inflammatory cytokineIL-10 generated by a variety of cells, including T cells andmacrophages, functions in a negative feedback loop thatsuppresses the generation of inflammatory cytokines anddampens the acute inflammatory response.(5) IL-10 alsosuppresses osteoclast formation, suggesting that it playsa role in modulating bone loss in inflammatory disor-ders.(6,7)
4-1BB is an inducible T-cell costimulatory molecule anda member of the TNF receptor (TNFR) family that includesTNFR1, TNFR2, CD40, OX40, and RANK.(8) 4-1BB isexpressed mainly on activated T cells, but recent studieshave shown that it is present on a variety of other cellsincluding myeloid cells.(9) 4-1BB is also known to be a den-dritic cell–activating molecule like RANK. 4-1BBL is ex-pressed on activated antigen-presenting cells and on severalleukemia cell lines.(9) The expression pattern of 4-1BB and4-1BBL has suggested that 4-1BB plays a role in variousresponses related to innate immunity and bone metabolism.
Therefore, we studied whether 4-1BB and 4-1BBL inter-action plays a role in osteoclastogenesis, using 4-1BB–deficient mice.The authors state that they have no conflicts of interest.
Department of Biological Sciences and the Immunomodulation Research Center, University of Ulsan, Ulsan, Korea.
JOURNAL OF BONE AND MINERAL RESEARCHVolume 21, Number 12, 2006Published online on August 21, 2006; doi: 10.1359/JBMR.060813© 2006 American Society for Bone and Mineral Research
1907
JO606431 1907 1912 December

MATERIALS AND METHODS
Reagents
Recombinant mouse M-CSF, RANKL, recombinantmouse IL-10, neutralizing anti-mouse IL-10 Ab, and biotin-labeled anti-mouse IL-10 Ab were obtained from R&Dsystems(Minneapolis, MN, USA). Antibodies againstCD11b, F4/80, CD3, and CD45R were obtained from eBio-science (San Diego, CA, USA). Recombinant mouse IFN-�was purchased from PBL Biomedical Laboratory (Piscata-way, NJ, USA). �-MEM and FBS were purchased fromLife Technologies (Grand island, NY, USA).
Osteoclast formation
Bone marrow cells (typically five mice per preparation)were isolated from 4- to 5-week-old 4-1BB+/+ (WT),4-1BB+/− (hybrid), and 4-1BB−/− (KO) Balb/c mice pro-vided by the University of Ulsan, Immunomodulation Re-search Center. The genotypes of offspring were determinedby Southern blot analysis of DNA from tail biopsies. Allmice were housed in the specific pathogen-free animal fa-cility of the Immunomodulation Research Center. All ani-mals were handled in accordance with the Institutional Ani-mal Care and Use Committee of the ImmunomodulationResearch Center. Standards were approved by the Institu-tional Animal Care and Use Committee of the Immuno-modulation Research Center. Femora and tibias were asep-tically removed and dissected free of adherent soft tissue.The bone ends were cut, and the marrow cavity was flushedout with �-MEM from one end of the bone using a sterile21-gauge needle. The bone marrow suspension was care-fully agitated with a plastic Pasteur pipette to obtain singlecells; these were washed twice and resuspended in �-MEMcontaining 10% FBS, and the suspension was incubated onplates with M-CSF (20 ng/ml). After 16 h, nonadherent cellswere harvested and cultured for 2 more days, at which timelarge populations of adherent monocyte/macrophage-likecells had formed on the bottom of the culture plates. Thesmall number of nonadherent cells and adherent stromalcells were removed by washing the dishes with PBS, fol-lowed by incubation for 5 minutes in 0.25% trypsin/0.05%EDTA. The remaining adherent bone marrow–derivedmacrophages (BMMs) were harvested by vigorous pipet-ting and confirmed by fluorescence-activated cell sorter(FACS) as positive for CD11b and F4/80 and negative forCD3 and CD45R (data not shown). The absence of con-taminating stromal cells was confirmed by lack of growthwhen M-CSF was omitted (data not shown). The isolatedBMMs were seeded at a density of 3 × 104 cells/well in48-well plates, additional medium containing M-CSF (20ng/ml) and RANKL (40 ng/ml) was added, and the mediumwas replaced on day 3. After incubation for the indicatedtimes, the cells were fixed in 10% formalin for 10 minutesand prepared for TRACP staining as described.(10) Thenumber of TRACP+ multinucleated cells (MNCs) contain-ing three or more nuclei was scored.
RNA isolation and RT-PCR
IL-10 and GAPDH mRNA was measured by RT-PCRanalysis. Total RNA was isolated from M-CSF– and
RANKL-treated BMM cells with TRI reagent (SigmaChemical, St Louis, MO, USA), and cDNA was synthesizedusing the reverse transcriptase supplied with the cDNAsynthesis kit (Invitrogen, San Diego, CA, USA). ThecDNA was amplified by 32 cycles (IL-10) and 25 cycles(GAPDH) of PCR with the following specific PCR prim-ers: mouse IL-10 5�-CCCTGGGTGAGAAGCTGAAG-3�(forward) and 5�-GGAAGAACCCCTCCCATCAT-3�(reverse); mouse GAPDH, 5�-ACCACAGTCCATGC-CATCAC-3� (forward) and 5�-TCCACCACCCTGTTGC-TGTA-3� (reverse). Each cycle consisted of 30 s of dena-turation at 94°C, 30 s of annealing at 60°C, and 30 s ofextension at 72°C. GAPDH was used as internal control.The sizes of the PCR products for mouse IL-10 andGAPDH were 526, and 452 bp, respectively.
Statistical analysis
All values are expressed as means ± SE. Student’s t-testwas used to evaluate differences between samples of inter-est and the corresponding controls. A p value of <0.05 wasconsidered statistically significant.
RESULTS
Osteoclastogenesis is stimulated in 4-1BB KO mice
Whole bone marrow cells were obtained from 4-1BB+/+
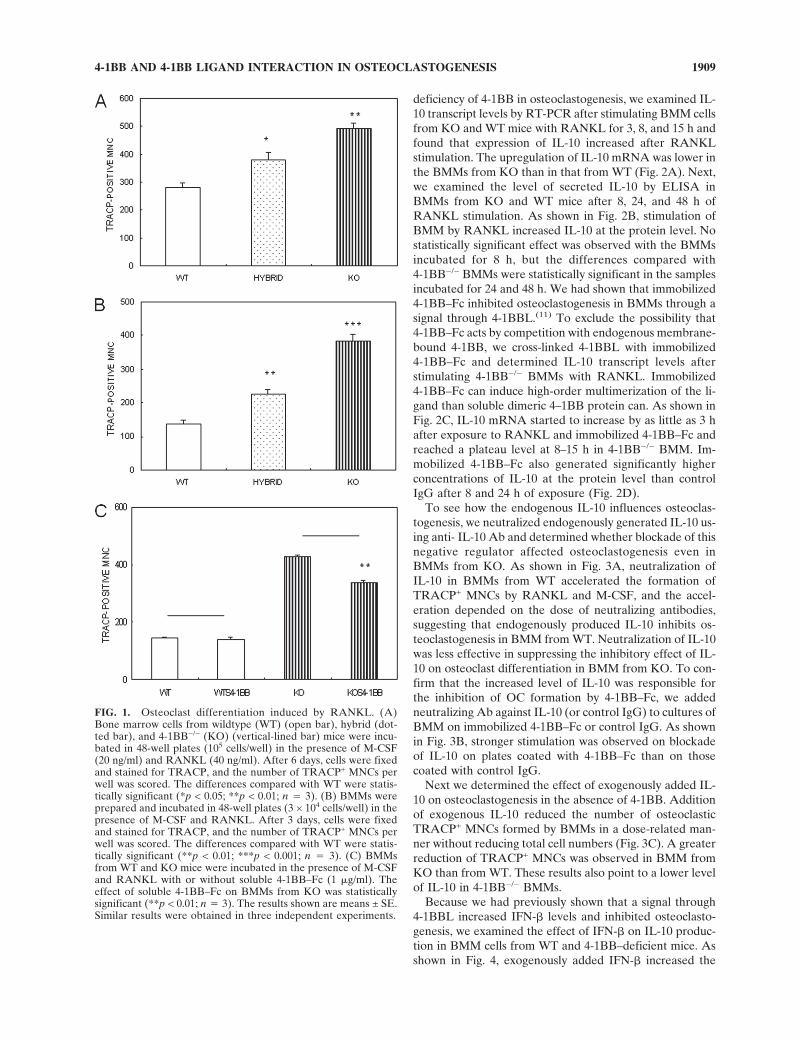
(WT), 4-1BB+/− (hybrid), and 4-1BB−/− (KO) littermates ona Balb/c background and incubated for 6 days with M-CSFand RANKL to generate osteoclasts. RANKL induced theformation of a large number of mononuclear and multi-nuclear osteoclasts that were TRACP+. The numbers ofTRACP+ MNCs formed by the bone marrow cells from KOand hybrid mice were 1.8- and 1.4-fold higher, respectively,than the number formed by those from WT mice (Fig. 1A).
Because the bone marrow cells used were a heteroge-neous population, we determined the effect of 4-1BB di-rectly on cultures of BMM cells free of stromal cells andlymphocytes (Fig. 1B). In the presence of M-CSF andRANKL, the maximum level of osteoclast formation oc-curred at 3 days. The stimulation of osteoclast formation inthe KO and hybrid mice was 2.8- and 1.7-fold, respectively,higher than in the WT mice. When we stimulated BMMswith RANKL and M-CSF in the presence of soluble 4-1BB-Fc fusion protein (1 �g/ml), osteoclastogenesis in theBMMs from KO decreased significantly, whereas there wasno effect on those from WT (Fig. 1C). These results suggestthat absence of 4-1BB stimulates osteoclast formation inBMM cells, indicating that 4-1BB and 4-1BBL interactionreduces osteoclastogenesis. All additional experimentswere performed using BMMs.
Stimulation of osteoclastogenesis results from lowerlevel of IL-10 in 4-1BB-KO mice
We showed previously that the 4-1BB and 4-1BBL inter-action inhibits osteoclastogenesis in BMMs by increasingthe level of IFN-�.(11) However, neutralization of IFN-� didnot completely suppress this effect, indicating that otherfactors were involved in the inhibition of osteoclastogen-esis. To see whether IL-10 levels were also related to the
SHIN ET AL.1908
deficiency of 4-1BB in osteoclastogenesis, we examined IL-10 transcript levels by RT-PCR after stimulating BMM cellsfrom KO and WT mice with RANKL for 3, 8, and 15 h andfound that expression of IL-10 increased after RANKLstimulation. The upregulation of IL-10 mRNA was lower inthe BMMs from KO than in that from WT (Fig. 2A). Next,we examined the level of secreted IL-10 by ELISA inBMMs from KO and WT mice after 8, 24, and 48 h ofRANKL stimulation. As shown in Fig. 2B, stimulation ofBMM by RANKL increased IL-10 at the protein level. Nostatistically significant effect was observed with the BMMsincubated for 8 h, but the differences compared with4-1BB−/− BMMs were statistically significant in the samplesincubated for 24 and 48 h. We had shown that immobilized4-1BB–Fc inhibited osteoclastogenesis in BMMs through asignal through 4-1BBL.(11) To exclude the possibility that4-1BB–Fc acts by competition with endogenous membrane-bound 4-1BB, we cross-linked 4-1BBL with immobilized4-1BB–Fc and determined IL-10 transcript levels afterstimulating 4-1BB−/− BMMs with RANKL. Immobilized4-1BB–Fc can induce high-order multimerization of the li-gand than soluble dimeric 4–1BB protein can. As shown inFig. 2C, IL-10 mRNA started to increase by as little as 3 hafter exposure to RANKL and immobilized 4-1BB–Fc andreached a plateau level at 8–15 h in 4-1BB−/− BMM. Im-mobilized 4-1BB–Fc also generated significantly higherconcentrations of IL-10 at the protein level than controlIgG after 8 and 24 h of exposure (Fig. 2D).
To see how the endogenous IL-10 influences osteoclas-togenesis, we neutralized endogenously generated IL-10 us-ing anti- IL-10 Ab and determined whether blockade of thisnegative regulator affected osteoclastogenesis even inBMMs from KO. As shown in Fig. 3A, neutralization ofIL-10 in BMMs from WT accelerated the formation ofTRACP+ MNCs by RANKL and M-CSF, and the accel-eration depended on the dose of neutralizing antibodies,suggesting that endogenously produced IL-10 inhibits os-teoclastogenesis in BMM from WT. Neutralization of IL-10was less effective in suppressing the inhibitory effect of IL-10 on osteoclast differentiation in BMM from KO. To con-firm that the increased level of IL-10 was responsible forthe inhibition of OC formation by 4-1BB–Fc, we addedneutralizing Ab against IL-10 (or control IgG) to cultures ofBMM on immobilized 4-1BB–Fc or control IgG. As shownin Fig. 3B, stronger stimulation was observed on blockadeof IL-10 on plates coated with 4-1BB–Fc than on thosecoated with control IgG.
Next we determined the effect of exogenously added IL-10 on osteoclastogenesis in the absence of 4-1BB. Additionof exogenous IL-10 reduced the number of osteoclasticTRACP+ MNCs formed by BMMs in a dose-related man-ner without reducing total cell numbers (Fig. 3C). A greaterreduction of TRACP+ MNCs was observed in BMM fromKO than from WT. These results also point to a lower levelof IL-10 in 4-1BB−/− BMMs.
Because we had previously shown that a signal through4-1BBL increased IFN-� levels and inhibited osteoclasto-genesis, we examined the effect of IFN-� on IL-10 produc-tion in BMM cells from WT and 4-1BB–deficient mice. Asshown in Fig. 4, exogenously added IFN-� increased the
FIG. 1. Osteoclast differentiation induced by RANKL. (A)Bone marrow cells from wildtype (WT) (open bar), hybrid (dot-ted bar), and 4-1BB−/− (KO) (vertical-lined bar) mice were incu-bated in 48-well plates (105 cells/well) in the presence of M-CSF(20 ng/ml) and RANKL (40 ng/ml). After 6 days, cells were fixedand stained for TRACP, and the number of TRACP+ MNCs perwell was scored. The differences compared with WT were statis-tically significant (*p < 0.05; **p < 0.01; n � 3). (B) BMMs wereprepared and incubated in 48-well plates (3 × 104 cells/well) in thepresence of M-CSF and RANKL. After 3 days, cells were fixedand stained for TRACP, and the number of TRACP+ MNCs perwell was scored. The differences compared with WT were statis-tically significant (**p < 0.01; ***p < 0.001; n � 3). (C) BMMsfrom WT and KO mice were incubated in the presence of M-CSFand RANKL with or without soluble 4-1BB–Fc (1 �g/ml). Theeffect of soluble 4-1BB–Fc on BMMs from KO was statisticallysignificant (**p < 0.01; n � 3). The results shown are means ± SE.Similar results were obtained in three independent experiments.
4-1BB AND 4-1BB LIGAND INTERACTION IN OSTEOCLASTOGENESIS 1909
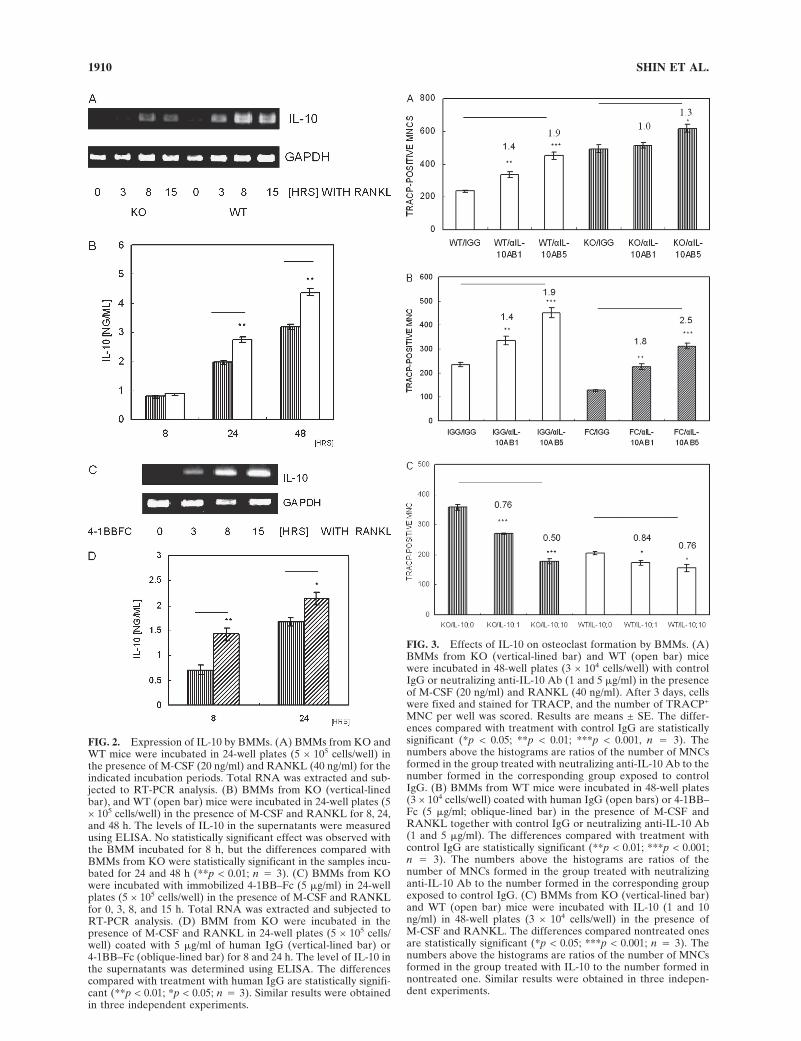
FIG. 2. Expression of IL-10 by BMMs. (A) BMMs from KO andWT mice were incubated in 24-well plates (5 × 105 cells/well) inthe presence of M-CSF (20 ng/ml) and RANKL (40 ng/ml) for theindicated incubation periods. Total RNA was extracted and sub-jected to RT-PCR analysis. (B) BMMs from KO (vertical-linedbar), and WT (open bar) mice were incubated in 24-well plates (5× 105 cells/well) in the presence of M-CSF and RANKL for 8, 24,and 48 h. The levels of IL-10 in the supernatants were measuredusing ELISA. No statistically significant effect was observed withthe BMM incubated for 8 h, but the differences compared withBMMs from KO were statistically significant in the samples incu-bated for 24 and 48 h (**p < 0.01; n � 3). (C) BMMs from KOwere incubated with immobilized 4-1BB–Fc (5 �g/ml) in 24-wellplates (5 × 105 cells/well) in the presence of M-CSF and RANKLfor 0, 3, 8, and 15 h. Total RNA was extracted and subjected toRT-PCR analysis. (D) BMM from KO were incubated in thepresence of M-CSF and RANKL in 24-well plates (5 × 105 cells/well) coated with 5 �g/ml of human IgG (vertical-lined bar) or4-1BB–Fc (oblique-lined bar) for 8 and 24 h. The level of IL-10 inthe supernatants was determined using ELISA. The differencescompared with treatment with human IgG are statistically signifi-cant (**p < 0.01; *p < 0.05; n � 3). Similar results were obtainedin three independent experiments.
FIG. 3. Effects of IL-10 on osteoclast formation by BMMs. (A)BMMs from KO (vertical-lined bar) and WT (open bar) micewere incubated in 48-well plates (3 × 104 cells/well) with controlIgG or neutralizing anti-IL-10 Ab (1 and 5 �g/ml) in the presenceof M-CSF (20 ng/ml) and RANKL (40 ng/ml). After 3 days, cellswere fixed and stained for TRACP, and the number of TRACP+
MNC per well was scored. Results are means ± SE. The differ-ences compared with treatment with control IgG are statisticallysignificant (*p < 0.05; **p < 0.01; ***p < 0.001, n � 3). Thenumbers above the histograms are ratios of the number of MNCsformed in the group treated with neutralizing anti-IL-10 Ab to thenumber formed in the corresponding group exposed to controlIgG. (B) BMMs from WT mice were incubated in 48-well plates(3 × 104 cells/well) coated with human IgG (open bars) or 4-1BB–Fc (5 �g/ml; oblique-lined bar) in the presence of M-CSF andRANKL together with control IgG or neutralizing anti-IL-10 Ab(1 and 5 �g/ml). The differences compared with treatment withcontrol IgG are statistically significant (**p < 0.01; ***p < 0.001;n � 3). The numbers above the histograms are ratios of thenumber of MNCs formed in the group treated with neutralizinganti-IL-10 Ab to the number formed in the corresponding groupexposed to control IgG. (C) BMMs from KO (vertical-lined bar)and WT (open bar) mice were incubated with IL-10 (1 and 10ng/ml) in 48-well plates (3 × 104 cells/well) in the presence ofM-CSF and RANKL. The differences compared nontreated onesare statistically significant (*p < 0.05; ***p < 0.001; n � 3). Thenumbers above the histograms are ratios of the number of MNCsformed in the group treated with IL-10 to the number formed innontreated one. Similar results were obtained in three indepen-dent experiments.
SHIN ET AL.1910
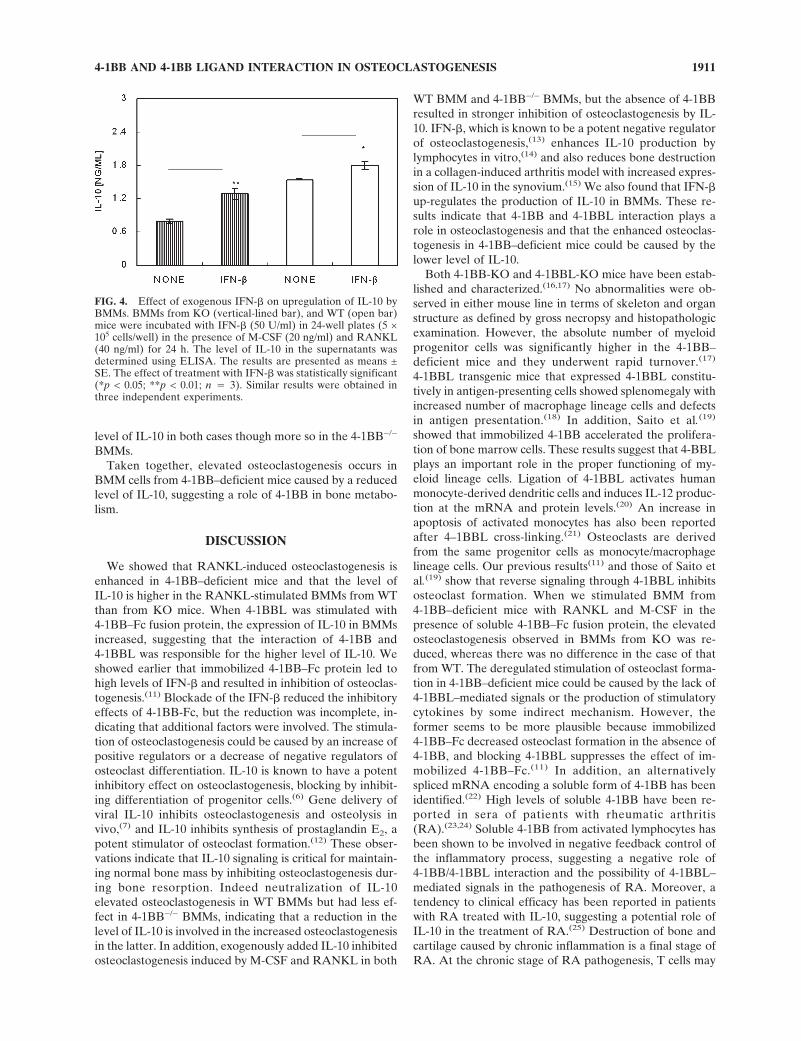
level of IL-10 in both cases though more so in the 4-1BB−/−
BMMs.Taken together, elevated osteoclastogenesis occurs in
BMM cells from 4-1BB–deficient mice caused by a reducedlevel of IL-10, suggesting a role of 4-1BB in bone metabo-lism.
DISCUSSION
We showed that RANKL-induced osteoclastogenesis isenhanced in 4-1BB–deficient mice and that the level ofIL-10 is higher in the RANKL-stimulated BMMs from WTthan from KO mice. When 4-1BBL was stimulated with4-1BB–Fc fusion protein, the expression of IL-10 in BMMsincreased, suggesting that the interaction of 4-1BB and4-1BBL was responsible for the higher level of IL-10. Weshowed earlier that immobilized 4-1BB–Fc protein led tohigh levels of IFN-� and resulted in inhibition of osteoclas-togenesis.(11) Blockade of the IFN-� reduced the inhibitoryeffects of 4-1BB-Fc, but the reduction was incomplete, in-dicating that additional factors were involved. The stimula-tion of osteoclastogenesis could be caused by an increase ofpositive regulators or a decrease of negative regulators ofosteoclast differentiation. IL-10 is known to have a potentinhibitory effect on osteoclastogenesis, blocking by inhibit-ing differentiation of progenitor cells.(6) Gene delivery ofviral IL-10 inhibits osteoclastogenesis and osteolysis invivo,(7) and IL-10 inhibits synthesis of prostaglandin E2, apotent stimulator of osteoclast formation.(12) These obser-vations indicate that IL-10 signaling is critical for maintain-ing normal bone mass by inhibiting osteoclastogenesis dur-ing bone resorption. Indeed neutralization of IL-10elevated osteoclastogenesis in WT BMMs but had less ef-fect in 4-1BB−/− BMMs, indicating that a reduction in thelevel of IL-10 is involved in the increased osteoclastogenesisin the latter. In addition, exogenously added IL-10 inhibitedosteoclastogenesis induced by M-CSF and RANKL in both
WT BMM and 4-1BB−/− BMMs, but the absence of 4-1BBresulted in stronger inhibition of osteoclastogenesis by IL-10. IFN-�, which is known to be a potent negative regulatorof osteoclastogenesis,(13) enhances IL-10 production bylymphocytes in vitro,(14) and also reduces bone destructionin a collagen-induced arthritis model with increased expres-sion of IL-10 in the synovium.(15) We also found that IFN-�up-regulates the production of IL-10 in BMMs. These re-sults indicate that 4-1BB and 4-1BBL interaction plays arole in osteoclastogenesis and that the enhanced osteoclas-togenesis in 4-1BB–deficient mice could be caused by thelower level of IL-10.
Both 4-1BB-KO and 4-1BBL-KO mice have been estab-lished and characterized.(16,17) No abnormalities were ob-served in either mouse line in terms of skeleton and organstructure as defined by gross necropsy and histopathologicexamination. However, the absolute number of myeloidprogenitor cells was significantly higher in the 4-1BB–deficient mice and they underwent rapid turnover.(17)
4-1BBL transgenic mice that expressed 4-1BBL constitu-tively in antigen-presenting cells showed splenomegaly withincreased number of macrophage lineage cells and defectsin antigen presentation.(18) In addition, Saito et al.(19)
showed that immobilized 4-1BB accelerated the prolifera-tion of bone marrow cells. These results suggest that 4-BBLplays an important role in the proper functioning of my-eloid lineage cells. Ligation of 4-1BBL activates humanmonocyte-derived dendritic cells and induces IL-12 produc-tion at the mRNA and protein levels.(20) An increase inapoptosis of activated monocytes has also been reportedafter 4–1BBL cross-linking.(21) Osteoclasts are derivedfrom the same progenitor cells as monocyte/macrophagelineage cells. Our previous results(11) and those of Saito etal.(19) show that reverse signaling through 4-1BBL inhibitsosteoclast formation. When we stimulated BMM from4-1BB–deficient mice with RANKL and M-CSF in thepresence of soluble 4-1BB–Fc fusion protein, the elevatedosteoclastogenesis observed in BMMs from KO was re-duced, whereas there was no difference in the case of thatfrom WT. The deregulated stimulation of osteoclast forma-tion in 4-1BB–deficient mice could be caused by the lack of4-1BBL–mediated signals or the production of stimulatorycytokines by some indirect mechanism. However, theformer seems to be more plausible because immobilized4-1BB–Fc decreased osteoclast formation in the absence of4-1BB, and blocking 4-1BBL suppresses the effect of im-mobilized 4-1BB–Fc.(11) In addition, an alternativelyspliced mRNA encoding a soluble form of 4-1BB has beenidentified.(22) High levels of soluble 4-1BB have been re-ported in sera of patients with rheumatic arthritis(RA).(23,24) Soluble 4-1BB from activated lymphocytes hasbeen shown to be involved in negative feedback control ofthe inflammatory process, suggesting a negative role of4-1BB/4-1BBL interaction and the possibility of 4-1BBL–mediated signals in the pathogenesis of RA. Moreover, atendency to clinical efficacy has been reported in patientswith RA treated with IL-10, suggesting a potential role ofIL-10 in the treatment of RA.(25) Destruction of bone andcartilage caused by chronic inflammation is a final stage ofRA. At the chronic stage of RA pathogenesis, T cells may
FIG. 4. Effect of exogenous IFN-� on upregulation of IL-10 byBMMs. BMMs from KO (vertical-lined bar), and WT (open bar)mice were incubated with IFN-� (50 U/ml) in 24-well plates (5 ×105 cells/well) in the presence of M-CSF (20 ng/ml) and RANKL(40 ng/ml) for 24 h. The level of IL-10 in the supernatants wasdetermined using ELISA. The results are presented as means ±SE. The effect of treatment with IFN-� was statistically significant(*p < 0.05; **p < 0.01; n � 3). Similar results were obtained inthree independent experiments.
4-1BB AND 4-1BB LIGAND INTERACTION IN OSTEOCLASTOGENESIS 1911

directly or indirectly modulate osteoclastogenesis. 4-1BB isconsidered a costimulatory molecule of T cells, and T cellsalso secrete RANKL,(3) which is not only involved in boneresorption but also in several immune responses. 4-1BBexpressed on activated T cells could stimulate 4-1BBL–bearing osteoclast cells to generate IL-10, a negative regu-lator of osteoclastogenesis.
Taken together, our findings show that osteoclast differ-entiation by whole bone marrow cells and BMM is in-creased in the absence of 4-1BB, a member of the TNFRfamily. In this study, we presented evidence that 4-1BBaffects osteoclastogenesis induced by RANKL, anothermember of the TNFR family, by modulating the level ofIL-10. The fact that the absence of 4-1BB stimulates osteo-clastogenesis may point to cross-talk between the immunesystem and bone metabolism. Further study of the 4-1BBand 4-1BBL interaction may clarify the role of 4-1BB incross-talk between the bone and immune system and pro-vide a rational for its use as a therapeutic agent in RA.
ACKNOWLEDGMENTS
This work was supported by the SRC fund to the IRC,University of Ulsan from KOSEF, and the Ministry of Ko-rean Sciences and Technology. JEL is supported by 2ndProject of BK21.
REFERENCES
1. Roodman GD 1996 Advances in bone biology: The osteoclast.Endocrinol Rev 17:308–332.
2. Suda T, Takahashi N, Udagawa N, Jimi E, Gillespie MT, Mar-tin TJ 1999 Modulation of osteoclast differentiation and func-tion by the new members of the tumor necrosis factor receptorand ligand families. Endocrinol Rev 20:345–357.
3. Kong YY, Yoshida H, Sarosi I, Tan HL, Timms E, CapparelliC, Morony S, Olivereira-dos-Santos AJ, Van G, Itie A, KhooW, Wakeham A, Dunstan CR, Lacey DL, Mak TW, Boyle WJ,Penninger JM 1999 OPGL is a key regulator of osteoclasto-genesis, lymphocyte development and lymph-node organogen-esis. Nature 397:315–323.
4. Horowitz MC, Xi Y, Wilson K, Kacena MA 2001 Control ofosteoclastogenesis and bone resorption by members of theTNF family of receptors and ligands. Cytokine Growth FactorRev 12:9–18.
5. De Vries JE 1995 Immunosuppressive and anti-inflammatoryproperties of interleukin 10. Ann Med 27:537–541.
6. Xu LX, Kukita T, Kukita A, Otsuka T, Niho Y, Iijima T 1995IL-10 inhibits cytokine selectively inhibits osteoclastogenesisby inhibiting differentiation of osteoclst progenitors into pre-osteoclast-like cells in rat bone marrow culture system. J CellPhysiol 165:624–629.
7. Carmody EE, Schwarz EM, Puzas E, Roiser RN, O’Keefe RJ2002 Viral interleukin-10 gene inhibition of inflammation, os-teoclastogenesis, and bone resorption in response to titaniumparticles. Arthritis Rheum 46:1298–1308.
8. Kwon BS, Weismann SM 1989 cDNA sequences of two induc-ible T-cell genes. Proc Natl Acad Sci USA 86:1963–1967.
9. Kwon B, Lee HW, Kwon BS 2002 New insights into the role of4-1BB in immune responses: Beyond CD8+T cells. Trends Im-munol 23:378–380.
10. Kobayashi K, Takahashi N, Jimi E, Udagawa N, Takami M,Kotake S, Nakagawa N, Kinosaki M, Yamaguchi K, Shima N,Yasuda H, Morinaga T, Higashio K, Martin TJ, Suda T 2000Tumor necrosis factor stimulates osteoclast differentiation by amechanism independent of the ODF/RANKL-RANK interac-tion. J Exp Med 191:275–285.
11. Shin HH, Lee EA, Kim SJ, Kwon BS, Choi HS 2006 A signal
through 4-1BB ligand inhibits receptor for activation of nuclearfactor-kB ligand-induced osteoclastogenesis by increasing in-terferon-beta production. FEBS Lett 580:1601–1606.
12. Shibata Y, Nishiyama A, Ohata H, Gabbard J, Myrvik QN,Henriksen RA 2005 Differential effects of IL-10 on prostaglan-din H synthase-2 expression and prostaglandin E2 biosynthesisbetween spleen and bone marrow macrophages. J Leukoc Biol77:544–551.
13. Takayanagi H, Kim S, Matsuo K, Suzuki T, Sato K, Yokochi Y,Oda H, Nakamura K, Ida N, Wagner EF, Taniguchi T 2002RANKL maintains bone homeostasis through c-Fos-dependent induction of interferon-�. Nature 416:744–749.
14. Rep MH, Hintzen RQ, Polman CH, van Lier RA 1996 Re-combinant interferon-beta blocks proliferation but enhancesinterleukin-10 secretion by activated human T-cells. J Neuro-immunol 67:111–118.
15. van Holten J, Reedquist K, Sattonet-Roche P, Smeets TJM,Plater-Zyberk C, Vervoordeldonk MJ, Tak PP 2003 Treatmentwith recombinant interferon-� reduces inflammation and slowscartilage destruction in the collagen-induced arthritis model ofrheumatoid arthritis. Arthritis Res Ther 6:R239–R249.
16. DeBenedette MA, Wen T, Bachmann MF, Ohashi PS, BarberBH, Stockling KL, Peschon JJ, Watts TH 1999 Analysis of4-1BB ligand-deficient mice and of mice lacking both 4-1BBLand CD28 reveals a role for 4-1BB in skin allograft rejectionand in the cytotoxic T cell response to influenza virus. J Im-munol 163:4833–4841.
17. Kwon BS, Hurtado JC, Lee ZH, Kwack KB, Seo SK, Choi BK,Koller BH, Wolisi G, Broxmeyer HE, Vinay DS 2002 Immuneresponse in 4-1BB-deficient mice. J Immunol 168:5483–5490.
18. Zhu G, Flies D, Tamada K, Sun Y, Rodriguez M, Fu Y-X,Chen L 2001 Progressive depletion of peripheral B lympho-cytes in 4-1BB ligand/I-E�-Transgenic mice. J Immunol167:2671–2676.
19. Saito K, Ohara N, Hotokezaka H, Fukumoto S, Yuhasa K,Naito M, Fujiwara T, Nakayama K 2004 Infection-induced up-regulation of the costimulatory molecule 4-1BB in osteoblasticcells and its inhibitory effect on M-CSF/RANKL-induced invitro osteoclastogenesis. J Biol Chem 279:13555–13563.
20. Laderach D, Wesa A, Galy A 2003 4-1BB-ligand is regulatedon human dendritic cells and induces the production of IL-12.Cell Immunol 226:37–44.
21. Langstein J, Michel J, Schwarz H 1999 CD137 induces prolif-eration and endomitosis in monocytes. Blood 94:3161–3168.
22. Setareh M, Schwarz H, Lotz M 1995 A mRNA variant encod-ing a soluble form of 4-1BB, a member of the murine NGF/TNF receptor family. Gene 27:311–315.
23. Michel J, Langstein J, Hofstadter F, Schwarz H 1998 A solubleform of CD137 (ILA/4-1BB), a member of the TNF receptorfamily, is released by activated lymphocytes and is detectablein sera of patients with rheumatoid arthritis. Eur J Immunol28:290–295.
24. Jung HW, Choi SW, Choi JI, Kwon BS 2004 Serum concen-trations of soluble 4-1BB and 4-1BB ligand correlated with thedisease severity in rheumatoid arthritis. Exp Mol Med 36:13–22.
25. Haraoui B, Strand V, Keystone E 2000 Biologic agents in thetreatment of rheumatoid arthritis. Curr Pharmaceut Biotech-nol 1:217–233.
Address reprint requests to:Hye-Seon Choi, PhD
Department of Biological Sciences and theImmunomodulation Research Center
University of UlsanUlsan 680-749, Korea
E-mail: [email protected]
Received in original form June 29, 2006; revised form August 1,2006; accepted August 16, 2006.
SHIN ET AL.1912




















