
Cyclin D1 Is a Selective Modifier of Androgen-dependent Signaling and Androgen Receptor Function

Enhanced latent inhibition in dopamine receptor-deficient miceis sex-specific for the D1 but not D2 receptor subtype:implications for antipsychotic drug action
Cecilie Bay-Richter1, Colm M. P. O’Tuathaigh2, Gerard O’Sullivan2, David M. Heery3, JohnL. Waddington2, and Paula M. Moran1
1School of Psychology, University of Nottingham, Nottingham, UK 2Molecular and CellularTherapeutics, Royal College of Surgeons in Ireland, St Stephen’s Green, Dublin, Ireland 3Schoolof Pharmacy, Centre for Biomolecular Sciences, University of Nottingham, Nottingham, UK
AbstractLatent inhibition (LI) is reduced learning to a stimulus that has previously been experiencedwithout consequence. It is an important model of abnormal allocation of salience to irrelevantinformation in patients with schizophrenia. In rodents LI is abolished by psychotomimetic drugsand in experimental conditions where LI is low in controls, its expression is enhanced byantipsychotic drugs with activity at dopamine (DA) receptors. It is however unclear what theindependent contributions of DA receptor subtypes are to these effects. This study thereforeexamined LI in congenic DA D1 and D2 receptor knockout (D1 KO and D2 KO) mice.Conditioned suppression of drinking was used as the measure of learning in the LI procedure.Both male and female DA D2 KO mice showed clear enhancement of LI reproducingantipsychotic drug effects in the model. Unexpectedly, enhancement was also seen in D1 KOfemale mice but not in D1 KO male mice. This sex-specific pattern was not replicated inlocomotor or motor coordination tasks nor in the effect of DA KOs on baseline learning in controlgroups indicating some specificity of the effect to LI. These data suggest that the dopaminergicmechanism underlying LI potentiation and possibly antipsychotic action may differ between thesexes, being mediated by D2 receptors in males but by both D1 and D2 receptors in females. Thesedata suggest that the DA D1 receptor may prove an important target for understanding sexdifferences in the mechanisms of action of antipsychotic drugs and in the aetiology of aberrantsalience allocation in schizophrenia.
KeywordsAntipsychotic; dopamine D1 receptor; dopamine D2 receptor; dopamine knockout mice; latentinhibition; sex difference
Copyright © 2008 CINP
Address for correspondence: P. M. Moran, Ph.D., School of Psychology, University of Nottingham, Nottingham NG7 2RD, UK. Tel.:+44 115 9515312 Fax : +44 115 9515324, [email protected].
Statement of InterestNone.
Europe PMC Funders GroupAuthor ManuscriptInt J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Published in final edited form as:Int J Neuropsychopharmacol. 2009 April ; 12(3): 403–414. doi:10.1017/S1461145708009656.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

IntroductionLatent inhibition (LI) is a process of learning to ignore irrelevant stimuli and is manifestedas slower learning about a stimulus that has been pre-exposed without consequence (PE),compared to learning about the same stimulus without pre-exposure (NPE) (Lubow andMoore, 1959). LI is used to model abnormalities in information filtering in schizophrenia,where patients have difficulty selecting between relevant and irrelevant information (Barakand Weiner, 2007; Gray et al., 1997; Kapur et al., 2005; Lubow, 2005; Weiner, 2003). LI isdisrupted in acute schizophrenia patients, their relatives and high schizotypal individuals,and normalized by antipsychotic drug treatment in patients (Baruch et al., 1988; Cohen etal., 2004; Gray et al., 1991; Lubow, 2005; Rascle et al., 2001). LI shows parallelpharmacological profiles in animals and humans (Moser et al., 2000; Weiner, 2003). Inrodents, psychotomimetic drugs such as amphetamine impair LI, while antipsychotic drugsenhance low levels of LI produced by decreasing the amount of pre-exposure (Moran et al.,1996; Moser et al., 2000; Palsson et al., 2005; Weiner, 2003; Weiner et al., 2003). Thesefeatures make it a powerful preclinical animal model of relevance to schizophrenia.
Pharmacology, microdialysis and lesion studies in rats have implicated the importance of theneurotransmitter dopamine (DA) in the regulation of LI (Gray et al., 1997; Joseph et al.,2000; Weiner, 2003; Young et al., 2005). In the brain the effects of DA are mediatedthrough activation of differently coupled receptor subtypes referred to as either DA D1-like(D1, D5) or DA D2-like (D2, D3, D4) (Missale et al., 1998). There is increasing evidence fora differential contribution of DA D1 and DA D2 receptors (hereafter designated D1 and D2respectively) to a variety of individual behaviours such as locomotor activity, memory andreward as well as responses to drugs of abuse and neuroprotection (Bozzi and Borrelli, 2006;Clifford et al., 2000; Doherty et al., 2008; Drago et al., 1994; Fowler et al., 2002; Kruzichand Grandy, 2004; Welter et al., 2007). The assignment of specific roles for DA receptorsubtypes in behaviour has been facilitated by the availability of knockout (KO) mice foreach DA receptor subtype (El-Ghundi et al., 2007; Holmes et al., 2004; Tan et al., 2003;Waddington et al., 2005). Given the importance of the DA hypothesis of schizophreniawhich suggests an overactivity in mesolimbic DA (Carlsson, 1988; Guilin et al., 2007;Kapur and Mamo, 2003; Kapur et al., 2005; Toda and Abi-Dargham, 2007) and the affinityof antipsychotic drugs for both D1 and D2 receptors (Missale et al., 1998; Miyamoto et al.,2005), it is of particular importance to establish the role of these receptor subtypes inbehaviours relevant to schizophrenia. Furthermore, understanding the potential separation ofside-effect and antipsychotic consequences of D1 and D2 blockade is important for thedevelopment of novel antipsychotic drugs with fewer extra-pyramidal side-effects (Farde etal., 1992; Horacek et al., 2006; Reynolds, 2007; Strange, 2001; Zai et al., 2007).
A number of studies have shown that D2 but not D1 antagonists enhance low levels of LIinduced by reducing the amount of stimulus pre-exposure, suggesting an action of DAthrough D2 but not D1 receptors in LI. D2 antagonists also reverse LI disruption byamphetamine while D1 antagonists do not (Moran et al., 1996; Moser et al., 2000; Trimbleet al., 2002; Warburton et al., 1994). However, nicotine-induced disruption of LI(considered to be mediated by nucleus accumbens DA release), is reversed by the D1antagonist SCH 23390 (Young et al., 2005). None of the drugs used in these studies is fullyselective for either the D1 or D2 receptor with many of the atypical antipsychotic drugshaving similar affinity for both receptors as well as for D3 and D4 (Missale et al., 1998;Miyamoto et al., 2005).
In this study congenic D1 or D2 receptor KO mice were used to determine whether theenhancing effects of D2 antagonists and the lack of effect of D1 antagonists on low LI arereproduced in mice that have genes for D1 or D2 receptors deleted independently and
Bay-Richter et al. Page 2
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

whether D1 or D2 receptors are independently essential for the normal expression of (high)LI. LI was investigated using two levels of pre-exposure, one producing LI in wild-type(WT) animals (60 PE) and one not (40 PE). This facilitates the observation of potentialenhancement or reduction of LI. To assess consistency with prior behavioural studies andrelevance to motor side-effects of antipsychotic drugs, locomotor activity and motorcoordination on the rotarod were also assessed.
Materials and methodsAnimals
The original F2 hybrid strain (129/Sv×C57BL/6J) containing the mutated D1 or D2 receptoralleles were generated as reported previously (Drago et al., 1994; Kelly et al., 1997). Thetargeted gene deletion was constructed in 129/Sv embryonic stem cells and male chimaerasmated with C57BL/6 females to produce heterozygous mutants (D1
+/- or D2+/-). Incipient
congenic D1 and D2 lines were established by back-crossing heterozygous mutants to WTC57BL/6 for 14 generations. Homozygous KO mice (D1
-/- and D2-/-) and WT (D1
+/+ andD2
+/+) littermates were bred by heterozygous intermatings of the incipient congenicheterozygote mutants imported from Royal College of Surgeons, Dublin, Ireland. Congenicstrains have significant advantages over mixed background strains in reducing inter-animalvariability and maximizing the specificity of the phenotype to the targeted gene (Gerlai,1996; Silva et al., 1997). D1 KO mice had low body weight and poor growth and wereweaned later than D2 KO mice to facilitate growth and survival. They were given a wetmash diet in the home cages after weaning, this has been reported previously in these mice(Doherty et al., 2008). D1 KO mice were bred with the kind permission of Professor JohnDrago, University of Melbourne. D2 KO mice were bred with the kind permission ofProfessor Malcolm Low, Oregon Health and Science University, USA.
Male and female KO mice and WT littermates aged at least 10 wk were used in theexperiments. Animals were housed 1-4 per cage under a 12-h light-dark cycle (lights on07:00 hours), at constant temperature and humidity with food available ad libitum.Experiments were performed during the light period. Mice used for the suppression oflicking model were subjected to water restriction periods of 23 h. All experiments wereperformed in accordance with local and national rules on animal experimentation, and withappropriate personal and project licence authority under the Animals (Scientific Procedures)Act, UK 1986 (PPL no: 40/2883).
GenotypingGenotyping was performed by PCR using genomic DNA extracted from ear biopsies by the‘hot sodium hydroxide and tris’ (hotSHOT) method (Truett et al., 2000). The reactionsdetermined the presence of both the neomycin phosphotransferase (NEO) gene and presenceof either the D1 gene or the D2 gene. The following primers were used for the D1 reaction:
NEOf: 5′-CTG AAT GAA CTG CAG GAC GA-3′,
NEOr: 5′-ATA CTT TCT CGG CAG GAG CA-3′,
D1f: 5′-CAA TCA GAA AGT TCC TTT AAG ATG TCC-3′,
D1r: ATG GTG GCT GGA AAA CAT CAC AGC CCC-3′,
and for the D2 reaction:
NEOf: 5′-CTT GGG TGG AGA GGC TAT TC-3′,
NEOr: 5′-AGG TGA GAT GAC AGG AGA TC-3′,
Bay-Richter et al. Page 3
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

D2f: 5′-TGT GAC TGC AAC ATC CCA CC-3′,
D2r: 5′-GCG GAA CTC AAT GTT GAA GG -3′.
PCR products were separated on a 1.5 % agarose gel. For D1 gene, amplification of a 172-bpPCR product indicated the presence of at least one transgenic allele and amplification of a353-bp PCR product indicated the presence of at least one WT allele. For the D2 gene,amplification of a 280-bp band indicated the presence of at least one transgenic allele and a105-bp band indicated the presence of at least one WT allele.
Instruments and proceduresLatent inhibition—Training and testing occurred in six identical conditioning chambers(21.6 cm×17.8 cm×12.7 cm, ENV-307W; Med Associates Inc., St Albans, VT, USA)housed in light and sound-attenuating boxes (ENV-022V, Med Associates Inc.). Theconditioning chambers were composed of Plexiglas walls in the front and back, stainlesssteel sides, and a metal grid floor connected to a shock scrambler and generator. Each boxcontained a ventilation fan, mounted at the back to provide air exchange and backgroundnoise (69 dB white noise), and a sonalert mounted on the right wall for delivering theconditioned stimulus (CS) (85 dB) (ENV-323AW, Med Associates Inc.). Each chamber wasequipped with a removable drink spout located in the left wall of the chamber. The lickspout was connected to a lickometer (ENV-250, Med Associates Inc.) which recorded thenumber of licks. The chambers were interfaced with a PC computer running MED-PCsoftware (SOF-735, Med Associates Inc.) to control stimulus presentation and record data.
The LI protocol consisted of six stages: water restriction, pre-training, pre-exposure/no pre-exposure, conditioning, re-establishment of drinking, and testing.
Water restriction (days 1-7): Mice were placed on a 23-h water restriction schedule 7 dbefore pre-training began. Water in the test apparatus was given in addition to the dailyration of 1 h delivered in the home cages. This regimen was maintained throughout theexperiment.
Pre-training (days 8-13): Mice were placed in the conditioning chambers and allowed todrink freely from a water sipper for 15 min. The number of licks made and latency to lickwas recorded.
Pre-exposure (day 14): Mice were placed in the conditioning chambers without access tothe water sipper. They were given 40 (expts 1 and 3), 60 (expt 2), 5 (expt 4) or 10 (expt 4)presentations of an 85-dB 5-s tone with a 15-s interstimulus interval. NPE control mice wereplaced in the chambers for the same amount of time but received no pre-exposures to thetone.
Conditioning (day 15): Mice were placed in the experimental chambers without access tothe water sipper. After 5 min, two tone-footshock pairings were presented. Each tone (CS)was of 5-s duration and was followed by a 1-s 0.38-mA US footshock. There was a 2.5-mininterval between pairings and mice remained in the chamber for 5 min following the secondtone-shock presentation.
Lick training (days 16-17): Mice were placed in the conditioning chambers for 15 min andgiven free access to the water sipper to re-establish licking in the chamber prior to testing.Mice that did not lick consistently were omitted from the experiment at this stage (n=2 forexpts 1-4).
Bay-Richter et al. Page 4
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Testing (day 18): Mice were placed in the conditioning chambers with access to the watersipper. The number of licks and the time to complete licks 80-90 (A) and 90-100 (B) wererecorded electronically. After the first 90 licks, the tone CS was presented until the mousereached lick 100. The standard measure of conditioned suppression was the time taken tocomplete licks 90-100 in the presence of the CS. A suppression ratio (SR) was calculatedaccording to the formula A/(A+B) yielding a scale of 0-0.5. Low SR indicates good learningwhile high SR indicates poor learning of the association between the tone and footshock. LIis demonstrated as higher SR in the PE group compared to the NPE group.
Spontaneous locomotor activity—Spontaneous locomotor activity was recorded usingthe open-field test. The open-field apparatus consisted of six Plexiglas boxes measuring 30cm×17 cm with 25 cm walls. All walls were dark except for one transparent wall in eachbox allowing light to enter. Animals were placed in the boxes to habituate for 30 min.Twenty-four hours after the habituation session the animals were again placed into the open-field boxes for 30 min and total distance moved was recorded and analysed using acomputerized video-tracking program EthoVision (version 3.1, Noldus, The Netherlands).Six animals were tested simultaneously in six separate open-field arenas. Between mice, allarenas were thoroughly cleaned with water and odour-free soap.
Motor coordination—The rotarod test was used as a measure of motor coordination.Animals were placed on a plastic rod rotating at 3 rpm (Rota-Rod/RS, LSI, Letica,Barcelona, Spain). Latency (s) to fall from the rod up to a maximum of 300 s was manuallyrecorded. If a mouse fell within the first 5 s, the trial was restarted.
Experimental procedureExperiments were run in the following order:
Expt 1—LI at 40 PE in D2 KO mice (n=19 WT, n=24 D2 KO). Mice were tested first onrotarod activity, 4 d later on locomotor activity and 14 d later began water deprivationschedule for LI. The LI procedure was run using 40 PE to produce low levels of LI in WTmice, this number was determined by pilot experiments in our laboratory.
Expt 2—LI at 60 PE in D2 KO mice (n=33 WT, n=25 D2 KO). The LI procedure was runusing 60 PE to produce LI in WT mice.
Expt 3—LI at 40 PE in D1 KO mice (n=24 WT, n=21 D1 KO). Mice were tested first onrotarod activity, 2 d later on locomotor activity and 18 d later began water deprivationschedule for LI. The LI procedure was run using 40 PE.
Expt 4—This experiment was performed to further investigate LI in D1 KO males as expt 3showed that LI was still high at 40 PE in male D1 KO WT mice. LI was tested at 5 PE and10 PE in male D1 KO mice (n=36 male WT, n=15 male D1 KO) to produce low LI in WTmice.
StatisticsStatistics were performed using SPSS software, version 15 (SPSS Inc., Chicago, IL, USA).For LI experiments independent analysis of variance (ANOVA) was used with SR asdependent variable and exposure (NPE/PE) genotype (WT/KO) and sex (male/female) asfactors. For post-hoc comparisons and other behavioural experiments Student’s independentt tests were used. Five extreme values (>3 standard deviations of the mean) occurred in expt3 and were removed from analysis: one WT PE female, one WT NPE female, one KO NPEmale, two WT NPE males. No extreme values were found in any other experiment. Unless
Bay-Richter et al. Page 5
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

otherwise indicated, data were collapsed across sex when there was no effect of sex and nointeraction between sex and exposure or genotype.
ResultsLI in D2 receptor KO mice
There was a clear enhancement of low LI (40 PE) in D2 KO mice compared to WTlittermates (Figure 1a). A two-way ANOVA with exposure, genotype and sex as factorsdemonstrated a significant effect of exposure [F(1, 34)=6.113, p=0.01], genotype [F(1,34)=6.812, p=0.01] and a significant exposure×genotype interaction [F(1, 34)=5.667,p<0.05]. No sex differences were found [F(1, 34)=0.001, n.s.]. Post-hoc t tests confirmedthat D2 KO animals showed a significant difference between NPE and PE groups (denotingLI) whereas WT animals did not [D2 KO NPE vs. D2 KO PE: t(21)=−2.853, p<0.01; WTNPE vs. WT PE: t(14)=−1.139, n.s.]. Moreover, PE groups were significantly differentbetween genotypes whereas NPE groups were not [WT PE vs. D2 KO PE: t(17)=2.448,p<0.05; WT NPE vs. D2 KO NPE: t(18)=1.856, n.s.]. Figure 1(b, c) shows the same animalsfrom expt 1 (40 PE) divided by sex.
Figure 2 shows that when WT mice show robust LI (at 60 PE) there is no difference betweengenotypes, with both groups showing clear LI. There was a significant effect of exposure[F(1, 46)=9.481, p<0.01] indicating the presence of LI but no effect of genotype [F(1,46)=0.948, n.s.] and no exposure×genotype interaction [F(1, 47)=0.561, n.s.]. No sexdifferences were found [F(1, 46)=0.080, n.s.] and therefore, in post-hoc analysis the sexeswere pooled. Post-hoc t tests confirmed an exposure effect (i.e. the presence of LI) in bothgenotypes [WT NPE vs. WT PE: t(28)=−2.545, p=0.01; D2 KO NPE vs. D2 KO PE: t(19)=−2.409, p<0.05]. Post-hoc t tests showed no differences between NPE groups or PE groups[WT NPE vs. D2 KO NPE: t(23)=1.230, n.s.; WT PE vs. D2 KO PE: t(27)=0.897, n.s.].
Baseline differences in drinking rates during pre-training between genotypes were not foundin any experiment. Repeated-measures ANOVA on drinking rates in expt 1 with pre-trainingday and genotype as factors showed a significant effect of pre-training day [F(5, 35)=4.840,p<0.01], no effect of genotype [F(5, 39)=0.123, n.s.] and no pre-training day×genotypeinteraction [F(1, 35)=1.911, n.s.]. The same was found for expt 2: a significant effect of pre-training day was found [F(5, 52)=8.446, p<0.001], no significant effect of genotype [F(1,56)=0.136, n.s.] and no pre-training day×genotype interaction [F(5, 52)=1.111, n.s.]. Similarresults were found for expts 3 and 4 (data not shown). No baseline changes in drinkingbehaviour were seen in the test phase of the experiment either as measured by the time Ascores. Expt 1: no effect of group [F(1, 34)=1.580 n.s.], genotype [F(1, 34)=0.710, n.s.], orsex [F(1, 34)=0.470, n.s.]. Expt 2: no effect of group [F(1, 46)=0.394, n.s.], genotype [F(1,46)=0.313, n.s.] or sex [F(1, 46)=0.434, n.s.]. Expt 3: no effect of group [F(1, 33)=0.641,n.s.], genotype [F(1, 33)=1.742, n.s.] or sex [F(1, 33)=1.373, n.s.]. Expt 4: no effect of group[F(1, 50)=1.01, n.s.] or genotype [F(1, 50)=1.05, n.s.].
No effect of sex or genotype was found for NPE control learning groups in any of the fourexperiments (all F values <1).
Spontaneous locomotor activity and motor coordination in D2 receptor KO miceFigure 3a shows that D2 KO animals are hypoactive compared to WT littermates [F(1,40)=90.840, p<0.001] in the open-field test where spontaneous locomotor activity over 30min was measured. A sex difference was also found [F(1, 40)=10.859, p<0.01], where malestended to be hyperactive compared to females. Post-hoc t tests showed significant sexdifferences between D2 KO animals [male D2 KO vs. female D2 KO: t(23)=2.923, p<0.01]but not between WT animals [male WT vs. female WT: t(17)=1.927, n.s.]. The significant
Bay-Richter et al. Page 6
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

genotype effect was found in both males and females [male WT vs. male D2 KO: t(19)=−5.895, p<0.001; female WT vs. female D2 KO: t(21)=−7.973, p<0.001].
Figure 3b shows that D2 KO mice demonstrate impaired motor coordination as measured bythe rotarod test when compared to WT littermates [F(1, 42)=12.328, p<0.001]. No sexdifferences were found [F(1, 42)=0.969, n.s.]. Post-hoc t tests confirmed the genotype effect[male WT vs. male D2 KO: t(21)=−2.905, p<0.01; female WT vs. female D2 KO: t(21)=−2.005, p=0.05].
D1 receptor KO mice and LIFigure 4a indicates slightly enhanced LI in D1 KO mice. There was a significant effect ofexposure [F(1, 67)=7.223, p<0.01] and a significant effect of genotype [F(1, 67)=4.716,p<0.05], but no exposure×genotype interaction suggesting the effect is not specific to the PEgroup. There was a significant effect of sex [F(1, 52)=7.159, p<0.01] in WT mice.Subsequent ANOVAs for separate sexes with exposure and genotype as factors showed asignificant effect of exposure [F(1, 34)=9.045, p<0.01] and importantly no effect ofgenotype [F(1, 34)=0.036, n.s.] in males (Figure 4b) suggesting no potentiation in male KOmice. Females showed a significant effect of exposure [F(1, 29)=5.472, p<0.05] but incontrast with data in male mice, significant enhancement of LI was found in female KOmice (Figure 4c). In females there was a significant effect of genotype [F(1, 29)=15.649,p<0.001] and a significant exposure×genotype interaction [F(1, 29)=5.754, p<0.05]. Post-hoc t tests confirmed a genotype difference in females in the PE group [female WT PE vs.female D1 KO PE: t(14)=3.574, p<0.01] but not the NPE group [female WT NPE vs. femaleD1 KO NPE: t(15)=1.768, n.s.]. Post-hoc tests revealed no effect of exposure [female WTNPE vs. female WT PE: t(22)=0.317, n.s.; female D1 KO NPE vs. female D1 KO PE: t(7)=−1.379, n.s.]. In male animals no genotype differences were found [male WT PE vs. maleD1 KO PE: t(18)=0.274, n.s.; male WT NPE vs. male D1 KO NPE: t(15)=−0.789, n.s.] but asignificant effect of exposure was found in WT but not D1 KO animals [male WT NPE vs.WT PE: t(26)=−3.099, p<0.01; male D1 KO NPE vs. male D1 KO PE: t(7)=−1.515, n.s.].This suggests potentiation of LI in female but not male D1 KO mice.
There was no significant effect of D1 genotype on baseline drinking rate. Repeated-measuresANOVA showed a significant effect of pre-training day [F(5, 36)=6.047, p<0.001], no effectof genotype [F(2, 40)=2.236, n.s.] and no genotype×pre-training day interaction [F(10,74)=1.852, n.s.].
To investigate further the suggestion from the above data that male D1 KO mice do notshow enhanced LI a further experiment was performed investigating LI at very low numbersof exposures such that WT mice do not show LI. Figure 6 shows that at 5 PE where WTmale mice do not show LI, there is similarly no LI in D1 KO male mice. Furthermore, at 10PE there is a trend towards disruption in D1 KO mice. There was a significant effect ofexposure [F(1, 47)=11.76, p<0.0001], no effect of genotype (F<1), nor a genotype×exposureinteraction [F(2, 47)=1.573, n.s.] t test-confirmed presence of LI in WT at 10 PE [NPE vs.10 PE (t=7.9, p<0.001)].
Spontaneous locomotor activity and coordination in D1 receptor KO miceFigure 5a shows that in the open-field test of spontaneous locomotor activity, D1 KOanimals are hyperactive compared to WT littermates with a significant effect of genotype[F(1, 41)=40.596, p<0.001] and no effect of sex [F(1, 41)=0.471, n.s.].
Figure 5b shows no effect on motor coordination in D1 KO mice of either sex. No effect ofgenotype [F(1, 41)=0.097, n.s.] or sex [F(1, 41)=0.685, n.s.] was found.
Bay-Richter et al. Page 7
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

DiscussionThese data demonstrate that D2 KO mice show markedly enhanced LI in low PE conditionswhere WT mice do not show LI. This profile reproduces the established profile of a range ofantipsychotic drugs such as haloperidol, clozapine, remoxipride in LI in rats and mice(Mongeau et al., 2007; Moran et al., 1996; Moser et al., 2000; Trimble et al., 1997;Warburton et al., 1994; Weiner et al., 2003). These data for the first time confirm using agenetic approach, a specific role for the D2 receptor in the modulation of LI. Furthermore,our results suggest that the enhancement of LI characteristic of antipsychotics can be elicitedby deletion of the D2 receptor subtype alone, and is therefore probably not dependent uponD3 and D4 receptors for which antipsychotic drugs also have affinity. In conditions where LIis present in controls, i.e. high number of preexposures (60 PE), D2 receptor gene deletion iswithout effect confirming that LI expression does not require the D2 receptor in conditionswhere WT mice show robust LI. This reproduces well-established effects of D2 receptorantagonists on LI (Moran et al., 1996; Ruob et al., 1998; Shadach et al., 2000).
It has been suggested that D2 KO mouse models possess limited value in modellingantipsychotic drug effects as, with the notable exception of side-effect-relevant behaviourssuch as motor deficits, they have not straightforwardly reproduced the behavioural effects ofD2 antagonist antipsychotic drugs (Chen et al., 2006; Gainetdinov et al., 2001; Holmes et al.,2004; Waddington et al., 2005). For example D2 KO mice do not show similar effects toantipsychotic drugs in sensory gating in the pre-pulse inhibition (PPI) model, althoughamphetamine abolition of PPI is prevented in these mice (Ralph et al., 1999). The finding ofa similar profile of D2 receptor knockout and antipsychotic drugs in LI suggest that, for theconstituent KO model, such a conclusion is premature. It is noteworthy that in theconditioned avoidance response model which is also sensitive to the effects of antipsychoticdrugs, D2 KO mice similarly reproduce the effects of antipsychotic drugs, although sexdifferences in this model have not been addressed (Smith et al., 2002).
The present study also confirms that D2 gene deletion produces hypoactivity and impairedmotor coordination as has been shown previously (Aoyama et al., 2000; Baik et al., 1995;Chausmer et al., 2002; Clifford et al., 2000; Fowler et al., 2002). This confirms that in thiscongenic line the established locomotor activity and motor coordination-impairing profile ofD2 antagonists is also reproduced (Aoyama et al., 2000).
In D1 KO mice we unexpectedly found significant enhancement of low LI in female but notmale mice. However, at 40 PE in WT males there was a higher baseline LI than in thefemale WT cohort perhaps allowing enhancement to be seen more readily in D1 KOfemales. We have subsequently replicated this higher LI in male WT D1 mice at 20, 40 and60 PE suggesting it is a robust effect (data not shown) and expt 4 has shown that we still seeLI at 10 PE in male WT animals although this is not seen at 5 PE. We cannot fully explainthis difference at present, however, it is possible that longer weaning in D1 WT mice mayhave exerted a beneficial cognitive effect on males as it is known that curtailment ofweaning in male mice has negative behavioural consequences and induces corticosteroneabnormalities (Wurbel and Stauffacher, 1997), this may also explain the baseline differencesin WT activity and rotarod that we observed. It is clear that the amount of pre-exposurerequired to produce LI in WT animals differs between the sexes in these mice, with malesshowing no LI at 5 PE and females no LI at 40 PE. These differences are, however,consistent with studies of LI in humans where females show reduced LI compared to males(Lubow and De la Casa, 2002). Despite this higher baseline in males there is still aconsiderable margin to detect enhancement of LI if it existed and we see no evidence forthis. At 5 PE we see no LI in either WT males or D1 KO males and a trend towards reducedLI in D1 KO males at 10 PE. This supports the suggestion that D1 KO males do not show
Bay-Richter et al. Page 8
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

enhanced LI compared to WT males. Further studies using larger cohorts than used in thepresent study would be desirable before concluding definitively that D1 KO males do notshow enhanced LI under any experimental conditions. At 10 PE there is some suggestionthat D1 KO males may show a deficit in LI produced by 10 PE, although this is notsignificant in the present study. This possibility will need to be replicated and extended tolarger cohorts in follow-up studies. These data nonetheless suggest that D1 receptor deletionmay enhance LI exclusively in females. This is contrary to prior studies in rats showing noeffect of D1 antagonists on low LI (Trimble et al., 2002). It is notable that these studies incommon with the majority of behavioural studies in rodents were carried out exclusively inmales (Hughes, 2007).
D1 KO mice showed hyperactivity and no deficits in rotarod motor coordination that wassimilar between the sexes confirming previous studies in these mice (Holmes et al., 2004).The lack of a similar profile of sex difference in activity and motor coordination additionallysuggests that the LI sex difference we have identified is highly specific to LI and notattributable to generalized behavioral/motor compromise in these mice. This is furthersupported by the fact that there is no effect of genotype or sex on the NPE learning groups.
Clinical trials reporting no benefit of D1 antagonists in schizophrenia patients have beenprimarily carried out with male patients (de Beaurepaire et al., 1995; Den Boer et al., 1995).The present data might help to explain this lack of efficacy. These data indicate thatenhancement of LI may involve differential regulation by D1 and D2 receptor subtypes inmales and females.
The existence of sex differences in phenomenology, epidemiology, neuroanatomy andresponse to antipsychotic drugs in schizophrenia has been well described (Goldstein et al.,2002; Lewine et al., 1996; Usall et al., 2007). Recently, Rubin et al. (2008) found sexdifferences in cognitive responses to antipsychotic drugs in patients with schizophrenia. Thepresent results may help to explain sex differences in response to antipsychotic drugs interms of dissociable dopaminergic substrates; one possible mechanism is an interaction withsex hormones. It is well established that the oestrous cycle influences variation in basalextracellular concentration of striatal DA, both in drug-stimulated DA release and in DA-mediated behaviours (Becker, 1999). Oestrogen directly acts on the striatum and nucleusaccumbens to enhance DA release and DA-mediated behaviours (Le Saux et al., 2006).
High levels of oestrogen have been shown to increase mesolimbic DA transmission (knownto disrupt LI) (Gray et al., 1995, 1997, 1999; Joseph et al., 1993; Van Hartesveldt and Joyce,1986) while ovariectomy reduces basal extracellular DA, amphetamine-induced striatal DArelease, and behaviours mediated by the striatal DA system (Becker, 1999).
Sex differences have also been reported in rodents in behavioural response to drugs of abusesuch as cocaine, ethanol and MDMA; these differences have been shown to be mediated inpart by the D1 receptor (Ferris et al., 2007; Festa et al., 2006; Melnick and DowEdwards,2001). Behavioural responses to D1 agonists and antagonists have also been demonstrated todiffer between the sexes (Nazarian et al., 2004).
Sexually dimorphic behavioural phenotypes in mice with mutations for genes mediatingdiverse aspects of DA neurotransmission have also been reported including DARPP-32 andcatechol-O-methyltransferase (COMT) function (Babovic et al., 2007; Harrison andTunbridge, 2007; Waddington et al., 2005).
In all KO mouse studies where the genetic deletion is present from birth there is thepotential for compensatory developmental changes to occur. While this cannot be excludedhere, this is currently the only approach to differentiate the impact of the absence of specific
Bay-Richter et al. Page 9
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

DA receptor subtypes in isolation. The recent development of inducible D1 mutants will beof value in future exploration of sexually dimorphic contributions of different DA receptorsubtypes to salience allocation processes and their aberrant states in psychosis (e.g. Gantoiset al., 2007). The present results are, however, consistent with predictions frompharmacological studies in LI for the D2 receptor, while the novel sex differences for D1suggested will generate further investigation using pharmacological approaches and a widerrange of behavioural models known to be sensitive to antipsychotic drug effects.
In summary we have shown that the antipsychoticlike profile of enhanced LI can be elicitedby D2 but not D1 receptor KO in males but by both D1 and D2 receptor KO in females. Thissuggests the possibility that the mechanism of action of antipsychotic drugs may differbetween the sexes and that the D1 receptor may be an important factor in dissociable maleand female responses to antipsychotic drugs.
AcknowledgmentsWe acknowledge the kind permission of Professor John Drago University of Melbourne, Australia and ProfessorMalcolm Low, Oregon Health and Science University, USA to use the KO mice. We are grateful for the support ofthe University of Nottingham New Researchers Fund, Science Foundation Ireland, and the Wellcome Trust.
ReferencesAoyama S, Kase H, Borrelli E. Rescue of locomotor impairment in dopamine D2 receptor-deficient
mice by an adenosine A2A receptor antagonist. Journal of Neuroscience. 2000; 20:5848–5852.[PubMed: 10908627]
Babovic D, O’Tuathaigh CM, O’Sullivan GJ, Clifford JJ, Tighe O, Croke DT, Karayiorgou M, GogosJA, Cotter D, Waddington JL. Exploratory and habituation phenotype of heterozygous andhomozygous COMT knockout mice. Behavioural Brain Research. 2007; 183:236–239. [PubMed:17707921]
Baik JH, Picetti R, Saiardi A, Thiriet G, Dierich A, Depaulis A, De Paulis A, Le Meur M, Borrelli E.Parkinsonian-like locomotor impairment in mice lacking dopamine D2 receptors. Nature. 1995;377:424–428. [PubMed: 7566118]
Baruch I, Hemsley DR, Gray JA. Differential performance of acute and chronic-schizophrenics in alatent inhibition task. Journal of Nervous and Mental Disease. 1988; 176:598–606. [PubMed:2903219]
Barak S, Weiner I. Scopolamine induces disruption of latent inhibition which is prevented byantipsychotic drugs and an acetylcholinesterase inhibitior. Neuropsychopharmacology. 2007;32:989–999. [PubMed: 16971898]
Becker JB. Gender differences in dopaminergic function in striatum and nucleus accumbens.Behavioural Pharmacology. 1999; 18:583–589.
Bozzi Y, Borrelli E. Dopamine in neurotoxicity and neuroprotection: what do D2 receptors have to dowith it? Trends in Neuroscience. 2006; 29:167–174.
Carlsson A. The current status of the dopamine hypothesis of schizophrenia.Neuropsychopharmacology. 1988; 1:179–186. [PubMed: 3075131]
Chausmer AL, Elmer GI, Rubinstein M, Low MJ, Grandy DK, Katz JL. Cocaine-induced locomotoractivity and cocaine discrimination in dopamine D2 receptor mutant mice. Psychopharmacology.2002; 163:54–61. [PubMed: 12185400]
Chen JS, Lipska BK, Weinberger DR. Genetic mouse models of schizophrenia, from hypothesis-basedto susceptibility gene-based models. Biological Psychiatry. 2006; 59:1180–1188. [PubMed:16631133]
Clifford JJ, Usiello A, Vallone D, Kinsella A, Borrelli E, Waddington JL. Topographical evaluation ofbehavioural phenotype in a line of mice with targeted gene deletion of the D2 dopamine receptor.Neuropharmacology. 2000; 39:382–390. [PubMed: 10698004]
Bay-Richter et al. Page 10
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Cohen E, Sereni N, Kaplan O, Weizman A, Kikinzon L, Weiner I, Lubow RE. The relation betweenlatent inhibition and symptom-types in young schizophrenics. Behavioural Brain Research. 2004;149:113–122. [PubMed: 15129775]
de Beaurepaire R, Labelle A, Naber D, Jones BD, Barnes TR. An open trial of the D1 antagonist SCH39166 in six cases of acute psychotic states. Psychopharmacology (Berlin). 1995; 121:323–327.[PubMed: 8584613]
Den Boer JA, van Megen HJ, Fleischhacker WW, Louwerens JW, Slaap BR, Westenberg HG,Westenberg HG, Burrows GD, Srivastava ON. Differential effects of the D1-DA receptorantagonist SCH39166 on positive and negative symptoms of schizophrenia. Psychopharmacology(Berlin). 1995; 121:317–322. [PubMed: 8584612]
Doherty JM, Masten VL, Powell SB, Ralph RJ, Klamer D, Low MJ, Geyer MA. Contributions ofdopamine D1, D2, and D3 receptor subtypes to the disruptive effects of cocaine on prepulseinhibition in mice. Neuropsychopharmacology. 2008; 33:2648–2656. [PubMed: 18075489]
Drago J, Gerfen CR, Lachowicz JE, Steiner H, Hollon TR, Love PE, Ooi GT, Grinberg A, Lee EJ,Huang SP, et al. Altered Striatal Function in a Mutant Mouse Lacking D1A Dopamine Receptors.Proceedings of the National Academy of Sciences USA. 1994; 91:12564–12568.
El-Ghundi M, O’Dowd BF, George SR. Insights into the role of dopamine receptor systems in learningand memory. Reviews in the Neurosciences. 2007; 18:37–66. [PubMed: 17405450]
Farde L, Nordstrom AL, Weisel FA, Pauli S, Halldin C, Sedvall G. Positron emittion tomographicanalysis of central D1 and D2 dopamine receptor occupancy in patients treated with classicalneuroleptics and clozapine. Relation to extrapyramidal side effects. Archives of GeneralPsychiatry. 1992; 49:538–544. [PubMed: 1352677]
Festa ED, Jenab S, Weiner J, Nazarian A, Niyomchai T, Russo SJ, Kemen LM, Akhavan A, Wu HBK,Quinones-Jenab V. Cocaine-induced sex differences in D1 receptor activation and binding levelsafter acute cocaine administration. Brain Research Bulletin. 2006; 68:277–284. [PubMed:16377433]
Ferris MJ, Mactutus FC, Silvers JM, Hasselrot U, Beaudin SA, Struppb BJ, Booze RM. Sex mediatesdopamine and adrenergic receptor expression in adult rats exposed prenatally to cocaine.International Journal of Developmental Neuroscience. 2007; 25:445–454. [PubMed: 17933484]
Fowler SC, Zarcone TJ, Vorontsova E, Chen R. Motor and associative deficits in D2 dopaminereceptor knockout mice. International Journal of Developmental Neuroscience. 2002; 20:309–321.[PubMed: 12175868]
Gainetdinov RR, Mohn AR, Caron MG. Genetic animal models: focus on schizophrenia. Trends inNeuroscience. 2001; 24:527–533.
Gantois I, Fang K, Jiang LN, Babovic D, Lawrence AJ, Ferreri V, Teper Y, Jupp B, Ziebell J,Morganti-Kossmann CM, et al. Ablation of D1 dopamine receptor-expressing cells generates micewith seizures, dystonia, hyperactivity, and impaired oral behaviour. Proceedings of the NationalAcademy of Sciences USA. 2007; 104:4182–4187.
Gerlai R. Gene-targeting studies of mammalian behavior: is it the mutation or the backgroundgenotype? Trends in Neuroscience. 1996; 19:177–181.
Goldstein JM, Seidman CJ, O’Brien LM, Horton NJ, Kennedy DN, Makris N, Caviness VS Jr.Faraone SV, Tsuang MT. Impact of normal sexual dimorphisms on sex differences in structuralbrain abnormalities in schizophrenia assessed by magnetic resonance imaging. Archives ofGeneral Psychiatry. 2002; 59:154–164. [PubMed: 11825137]
Gray JA, Feldon J, Rawlins JNP, Smith AD, Hemsley DR. The neuropsychology of schizophrenia.Behavioral and Brain Sciences. 1991; 14:1–19.
Gray JA, Joseph MH, Hemsley DR, Young AMJ, Warburton EC, Boulenguez P, Grigoryan GA, PetersSL, Rawlins JN, Tai BCT, et al. The role of mesolimbic dopaminergic and retrohippocampalafferents to the nucleus accumbens in latent inhibition: implications for schizophrenia.Behavioural Brain Research. 1995; 71:19–31. [PubMed: 8747172]
Gray JA, Kumari V, Lawrence N, Young AMJ. Functions of the dopaminergic innervation of thenucleus accumbens. Psychobiology. 1999; 27:225–235.
Bay-Richter et al. Page 11
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Gray JA, Moran PM, Grigoryan G, Peters SL, Young AMJ, Joseph MH. Latent inhibition: the nucleusaccumbens connection revisited. Behavioural Brain Research. 1997; 88:27–34. [PubMed:9401705]
Guilin O, Abi-Dargham A, Laruelle M. Neurobiology of dopamine in schizophrenia. InternationalReview of Neurobiology. 2007; 78:1–39. [PubMed: 17349856]
Harrison PJ, Tunbridge EM. Catechol-O-methyltransferase (COMT): a gene contributing to sexdifferences in brain function, and to sexual dimorphism in the predisposition to psychiatricdisorders. Neuropsychopharmacology. 2007 Published online: 5 September 2007. doi: 10.1038/sj.npp.1301543.
Holmes A, Lachowicz JE, Sibley DR. Phenotypic analysis of dopamine receptor knockout mice; recentinsights into the functional specificity of dopamine receptor subtypes. Neuropharmacology. 2004;47:1117–1134. [PubMed: 15567422]
Horacek J, Bubenikova-Valesova V, Kopecek M, Palenicek T, Dockery C, Mohr P, Hoschl C.Mechanisms of action of atypical antipsychotic drugs and the neurobiology of schizophrenia. CNSDrugs. 2006; 20:389–409. [PubMed: 16696579]
Hughes RN. Sex does matter: comments on the prevalence of male-only investigations of drug effectson rodent behaviour. Behavioral Pharmacology. 2007; 18:583–589.
Joseph MH, Peters SL, Gray JA. Nicotine blocks latent inhibition in rats - evidence for a critical roleof increased functional-activity of dopamine in the mesolimbic system at conditioning rather thanpreexposure. Psychopharmacology. 1993; 110:187–192. [PubMed: 7870882]
Joseph MH, Young AMJ, Moran P. Animal models of schizophrenia: the interface betweenneuropharmacology and cognitive deficit. European Journal of Neuroscience. 2000; 12:412.
Kapur S, Mamo D. Half a century of antipsychotics and still a central role for dopamine D-2 receptors.Progress in Neuro-Psychopharmacology & Biological Psychiatry. 2003; 27:1081–1090. [PubMed:14642968]
Kapur S, Mizrahi R, Li M. From dopamine to salience to psychosis - linking biology, pharmacologyand phenomenology of psychosis. Schizophrenia Research. 2005; 79:59–68. [PubMed: 16005191]
Kelly MA, Rubinstein M, Asa SL, Zhang G, Saez C, Bunzow JR, Allen RG, Hnasko R, Ben-JonathanN, Grandy DK, Low MJ. Pituitary lactotroph hyperplasia and chronic hyperprolactinemia indopamine D2 receptor-deficient mice. Neuron. 1997; 19:103–113. [PubMed: 9247267]
Kruzich PJ, Grandy DK. Dopamine D-2 receptors mediate two-odor discrimination and reversallearning in C57BL/6 mice. BMC Neuroscience. 2004; 5:12. [PubMed: 15061865]
Le Saux M, Morissette M, Di Paolo T. Erbeta mediates the estradiol increase of D2 receptors in ratstriatum and nucleus accumbens. Neuropharmacology. 2006; 50:451–457. [PubMed: 16309717]
Lewine RR, Walkert EF, Shurett R, Caudle J, Haden C. Sex differences in neuropsychologicalfunctioning among schizophrenic patients. American Journal of Psychiatry. 1996; 153:1178–1184.[PubMed: 8780422]
Lubow RE. Construct validity of the animal latent inhibition model of selective attention deficits inschizophrenia. Schizophrenia Bulletin. 2005; 31:139–153. [PubMed: 15888432]
Lubow RE, De la Casa G. Latent inhibition as a function of schizotypality and gender: implications forschizophrenia. Biological Psychology. 2002; 59:69–86. [PubMed: 11790444]
Lubow RE, Moore AU. Latent inhibition: the effect of nonreinforced pre-exposure to the conditionalstimulus. Journal of Comparative Physiology and Psychology. 1959; 52:415–419.
Melnick SM, Dow-Edwards DL. Differential behavioural responses to chronic amphetamine in adultmale and female rats exposed to postnatal cocaine treatment. Pharmacology Biochemistry andBehavior. 2001; 69:219–224.
Missale C, Nash SR, Robinson SW, Jaber M, Caron MG. Dopamine receptors: from structure tofunction. Physiological Reviews. 1998; 78:189–225. [PubMed: 9457173]
Miyamoto S, Duncan GE, Marx CE, Lieberman JA. Treatments for schizophrenia: a critical review ofpharmacology and mechanisms of action of antipsychotic drugs. Molecular Psychiatry. 2005;10:79–104. [PubMed: 15289815]
Mongeau R, Marcello S, Andersen JS, Pani L. Contrasting effects of diazepam and repeated restraintstress on latent inhibition in mice. Behavioural Brain Research. 2007; 183:147–155. [PubMed:17629580]
Bay-Richter et al. Page 12
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Moran PM, Fischer TR, Hitchcock JM, Moser PC. Effects of clozapine on latent inhibition in the rat.Behavioral Pharmacology. 1996; 7:42–48.
Moser PC, Hitchcock JM, Lister S, Moran PM. The pharmacology of latent inhibition as an animalmodel of schizophrenia. Brain Research Reviews. 2000; 33:275–307. [PubMed: 11011070]
Nazarian A, Russo SJ, Festa ED, Kraish M, Quinones-Jenab V. The role of D1 and D2 receptors in thecocaine conditioned place preference of male and female rats. Brain Research Bulletin. 2004;63:295–299. [PubMed: 15196654]
Palsson E, Klamer D, Wass C, Archer T, Engel JA, Svensson L. The effects of phencyclidine on latentinhibition in taste aversion conditioning: differential effects of preexposure and conditioning.Behavioural Brain Research. 2005; 157:139–146. [PubMed: 15617780]
Ralph RJ, Varty GB, Kelly MA, Wang YM, Caron MG, Rubinstein M, Grandy DK, Low MJ, GeyerMA. The dopamine D-2, but not D-3 or D-4, receptor subtype is essential for the disruption ofprepulse inhibition produced by amphetamine in mice. Journal of Neuroscience. 1999; 19:4627–4633. [PubMed: 10341260]
Rascle C, Mazas O, Vaiva G, Tournant M, Raybois O, Goudemand M, Thomas P. Clinical features oflatent inhibition in schizophrenia. Schizophrenia Research. 2001; 51:149–161. [PubMed:11518635]
Reynolds GP. The impact of pharmacogenetics on the development and use of antipsychotic drugs.Drug Discovery Today. 2007; 12:953–959. [PubMed: 17993414]
Rubin LH, Haas GL, Keshavan MS, Sweeney JA, Maki PM. Sex difference in cognitive response toantipsychotic treatment in first episode schizophrenia. Neuropsychopharmacology. 2008; 33:290–297. [PubMed: 17392734]
Ruob C, Weiner I, Feldon J. Haloperidol-induced potentiation of latent inhibition: interaction withparameters of conditioning. Behavioral Pharmacology. 1998; 9:245–253.
Shadach E, Gaisler I, Schiller D, Weiner I. The latent inhibition model dissociates between clozapine,haloperidol, and ritanserin. Neuropsychopharmacology. 2000; 23:151–161. [PubMed: 10882841]
Silva AJ, Simpson EM, Takahashi JS, Lipp HP, Nakanishi S, Wehner JM, Giese KO, Tully T, Abel T,Chapman PF, et al. Mutant mice and neuroscience: recommendations concerning geneticbackground. Neuron. 1997; 19:755–759. [PubMed: 9354323]
Smith JW, Fetsko LA, Xu R, Wang Y. Dopamine D2L receptor knockout mice display deficits inpositive and negative reinforcing properties of morphine and in avoidance learning. Neuroscience.2002; 113:755–765. [PubMed: 12182883]
Strange PG. Antipsychotic drugs: importance of dopamine receptors for mechanisms of therapeuticactions and side effects. Pharmacological Reviews. 2001; 53:119–133. [PubMed: 11171942]
Tan S, Hermann B, Borrelli E. Dopaminergic mouse mutants: investigating the roles of the differentdopamine receptor subtypes and the dopamine transporter. International Review of Neurobiology.2003; 54:145–197. [PubMed: 12785287]
Toda M, Abi-Dargham A. Dopamine hypothesis of schizophrenia: making sense of it all. CurrentPsychiatry Reports. 2007; 9:329–336. [PubMed: 17880866]
Trimble KM, Bell R, King DJ. Enhancement of latent inhibition in the rat by the atypical antipsychoticagent remoxipride. Pharmacology Biochemistry and Behavior. 1997; 56:809–816.
Trimble KM, Bell R, King DJ. Effects of the selective dopamine D(1) antagonists NNC 01-0112 andSCH 39166 on latent inhibition in the rat. Physiology & Behavior. 2002; 77:115–123. [PubMed:12213509]
Truett GE, Heeger P, Mynatt RL, Truett AA, Walker JA, Warman ML. Preparation of PCR-qualitymouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). Biotechniques. 2000;29:52, 54. [PubMed: 10907076]
Usall J, Suarez D, Haro JM. Gender differences in response to antipsychotic treatment in outpatientswith schizophrenia. Psychiatry Research. 2007; 153:225–231. [PubMed: 17681611]
Van Hartesveldt C, Joyce JN. Effects of estrogen on the basal ganglia. Neuroscience & BiobehavioralReviews. 1986; 10:1–14. [PubMed: 2871534]
Waddington JL, O’Tuathaigh C, O’Sullivan G, Tomiyama K, Koshikawa N, Croke DT. Phenotypicstudies on dopamine receptor subtype and associated signal transduction mutants: insights and
Bay-Richter et al. Page 13
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

challenges from 10 years at the psychopharmacology-molecular biology interface.Psychopharmacology (Berlin). 2005; 181:611–638. [PubMed: 16041535]
Warburton EC, Joseph MH, Feldon J, Weiner I, Gray JA. Antagonism of amphetamine-induceddisruption of latent inhibition in rats by haloperidol and ondansetron - implications for a possibleantipsychotic action of ondansetron. Psychopharmacology. 1994; 114:657–664. [PubMed:7855229]
Weiner I. The ‘two-headed’ latent inhibition model of schizophrenia: modeling positive and negativesymptoms and their treatment. Psychopharmacology (Berlin). 2003; 169:257–297. [PubMed:12601500]
Weiner I, Schiller D, Gaisler-Salomon I. Disruption and potentiation of latent inhibition byrisperidone: the latent inhibition model of atypical antipsychotic action.Neuropsychopharmacology. 2003; 28:499–509. [PubMed: 12629529]
Welter M, Vallone D, Samad TA, Meziane H, Usiello A, Borrelli E. Absence of dopamine D2receptors unmasks an inhibitory control over the brain circuitries activated by cocaine.Proceedings of the National Academy of Sciences USA. 2007; 16:6840–6845.
Wurbel H, Stauffacher G. Age and weight at weaning affect corticosterone level and development ofstereotypies in ICR - mice. Animal Behavior. 1997; 53:891–900.
Young AM, Moran PM, Joseph MH. The role of dopamine in conditioning and latent inhibition: what,when, where and how? Neuroscience & Biobehavioral Reviews. 2005; 29:963–976. [PubMed:16045987]
Zai CC, Hwanga RW, De Luca V, Müller DJ, King N, Zai GC, Remington G, Meltzer HY, LiebermanJA, Potkin SG, Kennedy JL. Association study of tardive dyskinesia and twelve DRD2polymorphisms in schizophrenia patients. American Journal of Psychiatry. 2007; 163:1747–1753.
Bay-Richter et al. Page 14
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 1.Latent inhibition (LI) in D2 KO mice with 40 pre-exposures. Columns represent mean±S.E.M.(a) LI in males and females (n=9-12 per group). Panels (b) and (c) show the data from panel(a) divided into (b) males and (c) females. ** p<0.01 compared to no pre-exposure (NPE)for same genotype; § p<0.05 compared to pre-exposed (PE) for wild-type (WT) mice.Enhanced LI is found in D2 KO mice at 40 PE where LI is not present in WT littermatecontrols.
Bay-Richter et al. Page 15
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
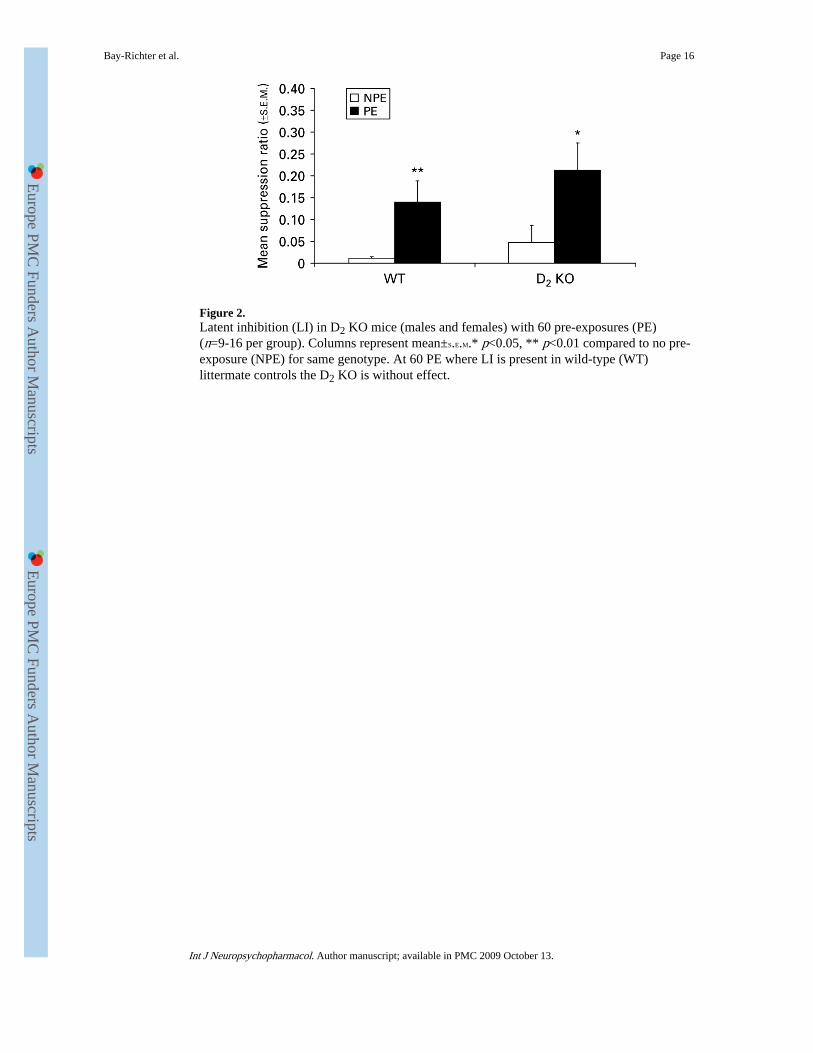
Figure 2.Latent inhibition (LI) in D2 KO mice (males and females) with 60 pre-exposures (PE)(n=9-16 per group). Columns represent mean±S.E.M.* p<0.05, ** p<0.01 compared to no pre-exposure (NPE) for same genotype. At 60 PE where LI is present in wild-type (WT)littermate controls the D2 KO is without effect.
Bay-Richter et al. Page 16
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
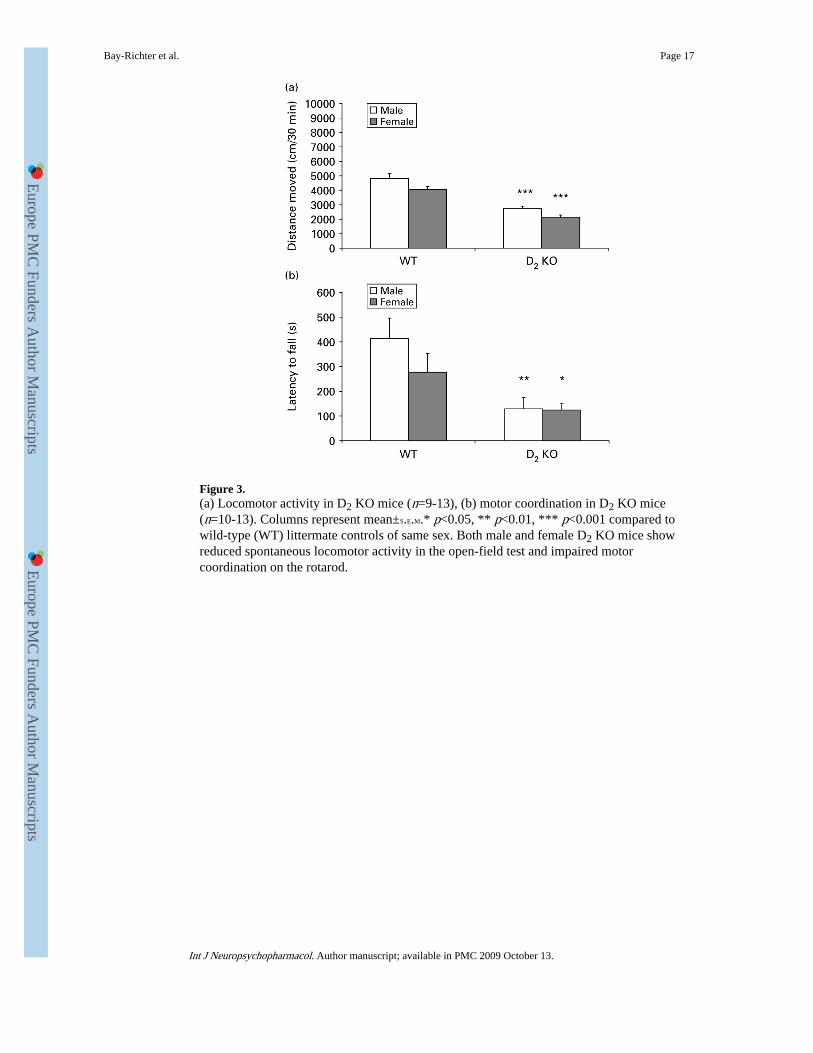
Figure 3.(a) Locomotor activity in D2 KO mice (n=9-13), (b) motor coordination in D2 KO mice(n=10-13). Columns represent mean±S.E.M.* p<0.05, ** p<0.01, *** p<0.001 compared towild-type (WT) littermate controls of same sex. Both male and female D2 KO mice showreduced spontaneous locomotor activity in the open-field test and impaired motorcoordination on the rotarod.
Bay-Richter et al. Page 17
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
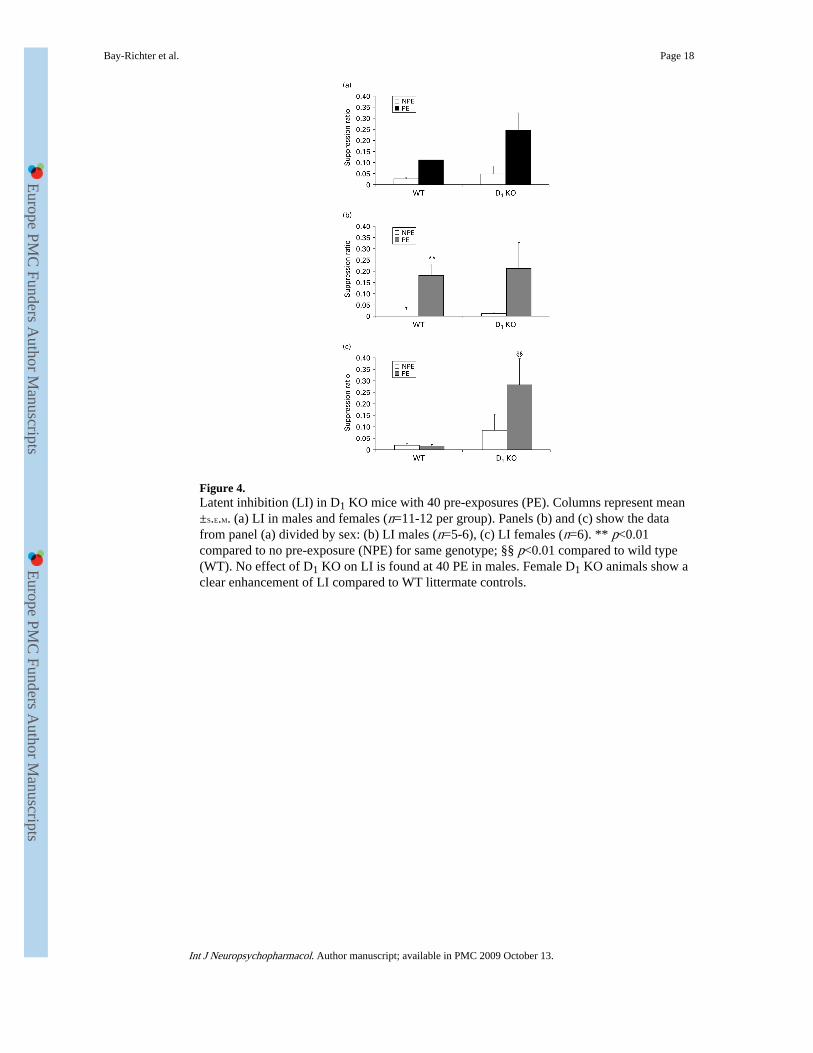
Figure 4.Latent inhibition (LI) in D1 KO mice with 40 pre-exposures (PE). Columns represent mean±S.E.M. (a) LI in males and females (n=11-12 per group). Panels (b) and (c) show the datafrom panel (a) divided by sex: (b) LI males (n=5-6), (c) LI females (n=6). ** p<0.01compared to no pre-exposure (NPE) for same genotype; §§ p<0.01 compared to wild type(WT). No effect of D1 KO on LI is found at 40 PE in males. Female D1 KO animals show aclear enhancement of LI compared to WT littermate controls.
Bay-Richter et al. Page 18
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 5.(a) Locomotor activity in D1 KO mice (n=10-12), (b) motor coordination in D1 KO mice(n=10-12). Columns represent mean±S.E.M. *** p<0.01 compared to wild-type (WT)littermate controls of same sex. D1 KO mice of both sexes are hyperactive compared to WTlittermate controls as measured in the open-field test. The D1 KO had no effect on motorcoordination in either sex measured on the rotarod.
Bay-Richter et al. Page 19
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
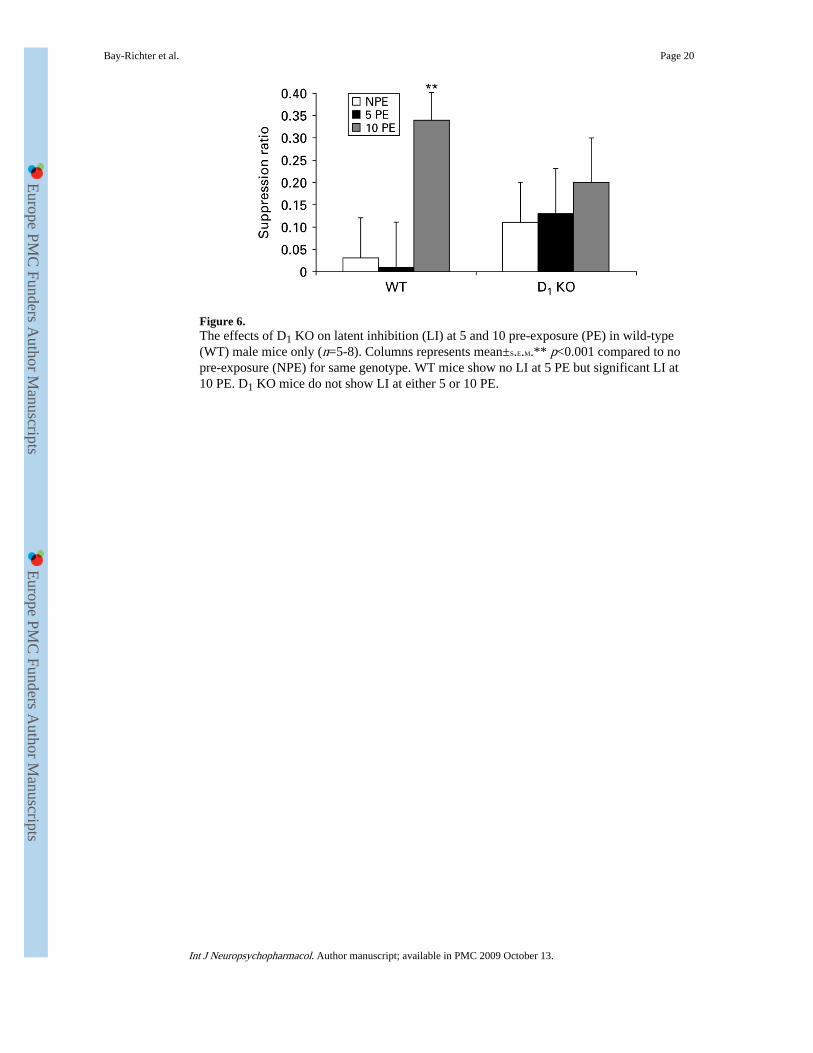
Figure 6.The effects of D1 KO on latent inhibition (LI) at 5 and 10 pre-exposure (PE) in wild-type(WT) male mice only (n=5-8). Columns represents mean±S.E.M.** p<0.001 compared to nopre-exposure (NPE) for same genotype. WT mice show no LI at 5 PE but significant LI at10 PE. D1 KO mice do not show LI at either 5 or 10 PE.
Bay-Richter et al. Page 20
Int J Neuropsychopharmacol. Author manuscript; available in PMC 2009 October 13.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
Copyright © 2022 FDOKUMEN




















