
Sclairpipe - Marine Pipeline Installation - Engineered Pipe ...
Upload
independentCategory
view
3download
0
Engineered antibody intervention strategies forAlzheimer’s disease and related dementias bytargeting amyloid and toxic oligomers
Remy Robert1,4, Olan Dolezal1, Lynne Waddington1,Meghan K. Hattarki1, Roberto Cappai2,3,Colin L. Masters2, Peter J. Hudson1 andKim L. Wark1
1CSIRO Molecular and Health Technologies, 343 Royal Parade,2The Mental Health Research Institute, University of Melbourne,Parkville, Victoria 3052 and 3Department of Pathology,Bio21 Molecular Science and Biotechnology Institute, University ofMelbourne, Melbourne, Victoria 3010, Australia
4To whom correspondence should be addressed. E-mail: [email protected]
Most neurodegenerative disorders, such as Alzheimer’s(AD), Parkinson’s, Huntington’s and Creutzfeldt–Jakobdisease, are characterised by the accumulation of insolublefilamentous aggregates known as amyloid. These pathol-ogies share common pathways involving protein aggrega-tion which can lead to fibril formation and amyloidplaques. The 4 kDa Ab peptide (39–43 amino acids)derived from the proteolysis of the amyloid precursorprotein is currently a validated target for therapy in AD.Both active and passive immunisation studies against Abare being trialled as potential AD therapeutic approaches.In this study, we have characterised engineered antibodyfragments derived from the monoclonal antibody, WO-2which recognises an epitope in the N-terminal region of Ab(amino acids 2–8 of Ab). A chimeric recombinant Fab(rFab) and single chain fragments (scFvs) of WO-2 wereconstructed and expressed in Escherichia coli. Rationallydesigned mutants to improve the stability of antibody frag-ments were also constructed. All antibody formats retainedhigh affinity (KD �8 3 1029 M) for the Ab peptide,comparable with the intact parental IgG as measuredby surface plasmon resonance. Likewise, all engineeredfragments were able to: (i) prevent amyloid fibrillisation,(ii) disaggregate preformed Ab1 –42 fibrils and (iii) inhibitAb1– 42 oligomer-mediated neurotoxicity in vitro as effi-ciently as the whole IgG molecule. These data indicatethat the WO-2 antibody and its fragments have immu-notherapeutic potential. The perceived advantages of usingsmall Fab and scFv engineered antibody formats whichlack the effector function include more efficient passageacross the blood-brain barrier and minimising the riskof triggering inflammatory side reactions. Hence, theserecombinant antibody fragments represent attractive can-didates and safer formulations of passive immunotherapyfor AD.Keywords: Alzheimer’s disease (AD)/amyloid b modulation/engineered antibody/SPR/stability
Introduction
Alzheimer’s disease (AD) is the most prevalent neurode-generative form of dementia (Selkoe, 2002; Masters and
Beyreuther, 2006) and results in memory loss and beha-vioural problems including mood swings, confusion andbreakdown of language in affected individuals. It is esti-mated that 26.6 million people worldwide were afflictedby AD in 2006 (Suh and Checler, 2002), which couldpotentially quadruple by 2050 (Brookmeyer et al., 2007).Currently there is no cure for AD, only treatments which canslow the progression of the disease and manage some of thesymptoms. For example, if diagnosed and treated early withcholinesterase inhibitors, patients can retain cognitive andfunctional abilities for longer as well as experience beha-vioural improvements (McGleenon et al., 1999).
The Ab protein has been identified as playing a major rolein the pathogenesis of AD. This 4 kDa peptide (39–43amino acids in length) is derived from the proteolysis of theamyloid precursor protein (APP) (Kang et al., 1987) and isone of the main constituents of the amyloid plaques associ-ated with the disease (Selkoe, 1999). Although the Abpeptide fibrillogenesis remains the subject of many studies,the precise mechanisms are not yet fully understood (Arimonet al., 2005). Nevertheless, it is known that the Ab peptidehas a tendency to self-associate and can adopt several formsincluding soluble oligomers (also referred to as Ab-deriveddiffusible ligands or ADDLs), protofibrils and insolubleamyloid fibrils. Importantly, all these forms are neurotoxicin vitro (Lorenzo and Yankner, 1994) with soluble oligomersdisplaying the highest level of toxicity (Lambert et al., 1998;Walsh et al., 2002).
A number of approaches have been used to design thera-peutic agents that can prevent or reverse the formation ofthese toxic Ab species (Weiner and Frenkel, 2006). Many ofthese strategies involve small drug molecules which inhibitor interfere with Alzheimer’s amyloidogenesis (Chernyet al., 2001; Yang et al., 2005; Zhao et al., 2008). Some ofthese compounds are now in clinical trials.
An alternative approach has focused on antibodies aspotential therapeutics. Studies have now demonstrated thatactive immunisation with the Ab peptide triggers the for-mation of antibodies that can clear and prevent the develop-ment of amyloid plaques in transgenic AD mice (Schenket al., 1999; Janus et al., 2000). These data prompted humanclinical trials to evaluate the efficiency of the AN1792vaccine (aggregated Ab). However, the trial was abandoneddue to 6% of the participants developing sterile meningoen-cephalitis (Orgogozo et al., 2003). Consequently, researchefforts were redirected towards developing a passive vaccine.The murine IgG monoclonal antibody 3D6, raised againstthe N-terminal 1–5 amino acids of Ab, was tested for itsability to reduce pathology in AD transgenic mice. Althougha reduction in diffuse amyloid deposits was observed (Bardet al., 2000), side effects such as cerebral amyloidangiopathy-related microhemorrhages still occurred (Rackeet al., 2005). To reduce immunogenicity, the 3D6 mAb was
# The Author 2008. Published by Oxford University Press. All rights reserved.
For Permissions, please e-mail: [email protected]
199
Protein Engineering, Design & Selection vol. 22 no. 3 pp. 199–208, 2009Published online October 16, 2008 doi:10.1093/protein/gzn052
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from

humanised (Bapineuzumab or AAB-001) and is now inphase III clinical trials.
Side effects observed after administration of some thera-peutic antibodies can be attenuated by deglycosylating wholeIgG antibody (Wilcock et al., 2006) or removing the Fcportion by proteolysis to produce Fab’2 (Tamura et al.,2005). An alternative strategy, and the subject of this study,is to design recombinant Fab (rFab) or scFv (Holliger andHudson, 2005; Levites et al., 2006). We constructed recombi-nant antibody fragments (scFv and chimeric Fab) derivedfrom the well-characterised WO-2 hybridoma cell line (Idaet al., 1996) and demonstrated their immunotherapeuticpotential compared with the intact IgG in targeting the Abpeptide. The WO-2 derived Fab and scFv fragments retainedthe same high affinity as the intact IgG and can efficiently:(i) prevent amyloid fibrillisation, (ii) disaggregate preformedAb1 – 42 fibrils and (iii) inhibit soluble oligomer-mediatedneurotoxicity.
Material and methods
Purification of WO-2 IgG and preparationof papain-digested WO-2 FabThe WO-2 hybridoma (Ida et al., 1996) was grown andmaintained in serum-free medium (Invitrogen, USA) forantibody production. WO-2 immunoglobulin was purifiedfrom culture supernatant by affinity chromatography on aProsep-vA protein A column (Millipore, Billerica, USA).The bound WO-2 IgG was eluted with 0.2 M glycine/1 MNaCl, pH 2.0 into a neutralising solution of 1 M Tris,pH 8.0, then dialysed against PBS overnight at 48C. WO-2Fab fragment was generated by digestion with papain(Sigma, St Louis, MO, USA) (Gruen et al., 1993).Undigested IgG and the Fc portion were removed by affinitychromatography using protein A.
Generation of recombinant scFv and rFabTotal RNA, extracted from the WO-2 hybridoma cell line(Ida et al., 1996) using the Trizol Reagent (Invitrogen,Mount Waverley, VIC, Australia), was utilised for cDNAsynthesis. The V genes were isolated from cDNA by RT–PCR using primers annealing to mouse light (mIgCk-1:50-CTTCCACTTGACATTGATGTCTTTG-30) and heavyconstant region (mIgG2a-1: 50-CAGGTCAAGGTCACTGGCTCAGG-30). The VH and VL genes were sequenced andanalysed by the IMGT databases.
To construct the WO-2 scFv, the VH (GenBank accessionno. AM950178) and VL (GenBank accession no.AM950179) genes were amplified with primers WO-2VHFOR (50-GCCATTAACCATGGCCCAGGTTACTCTGAAAGAGTCT-30), WO-2 VH BACK (50-TGAACCGCCACCGCCACTACCGCCACCGCCTGAGGAGACTGTGAGAGTGGTGCCTTG-30), WO-2 VLK FOR (50-GGTAGTGGCGGTGGCGGTTCAGGCGGTGGCGGTAGCGATGTTTTGATGACCCAAACT-30) and WO-2 VLK BACK (50-GCCATTAAGGATCCAGCGGCCGCCCGCTTCAGCTCCAGCTTGGT-30).The PCR products were assembled and cloned into the pGCexpression vector (Fig. 1a) (Coia et al., 1997) as describedby Gilliland et al. (1996). The chimeric rFab was generatedusing the same approach with primers WO-2 VH FOR/WO-2VH Fab BACK (50-TGAGGAGACGGTGACCGTGGTG
CCTTGGCCCCA-30) and WO-2 Fab K FOR(50-CAGGCCGATATCGTGATGACCCAAACTCCA-30)/WO-2Fab K BACK (50-TTTCAGCTCGAGCTTGGTCCCAGCACCGAA-30). Amplified VH and VL fragments were digestedwith restriction enzymes (NcoI/BstEII for VH and EcoRV/XhoI for VL) and ligated sequentially into the pGC-Fabexpression vector (Fig. 1b).
Three-dimensional structure modellingof the antibody fragmentThe molecular model of the WO-2 scFv was obtained byusing the web antibody modelling (WAM) algorithm(Whitelegg and Rees, 2000) (http://antibody.bath.ac.uk/).Images of the model were generated using PyMOL (PyMOLversion 0.82, http://pymol.sourceforge.net/) (DeLanoScientific LLC).
Mutagenesis of the antibody fragmentsSite-directed mutagenesis of the WO-2 scFv was performedusing the Quick Changew Site-directed Mutagenesis Kit II(Stratagene, La Jolla, CA, USA). Mutations were verified bysequencing the coding region of the antibody fragments onboth strands.
Production, purification and biochemical characterisationof the recombinant antibody fragmentsRecombinant proteins were expressed in the bacterial peri-plasm as described by Dolezal et al. (2000). Expression andisolation were monitored by gel filtration chromatographythrough a Superdexw 200 HR 10/30 column (AmershamPharmacia Biotech, Uppsala, Sweden) and Coomassie stain-ing. The presence of individual antibody fragments wasconfirmed by western blotting using a mouse anti-FLAGM2 antibody (Sigma) as previously described (Kortt et al.,1994).
Enzyme linked immunosorbent assaysWells of 96-well Maxisorb Immuno plates (Nunc, Roskilde,Denmark) were coated with 5 mg/ml of purified MBP-Ab1 – 42
(Caine et al., 2007) in carbonate-coating buffer. Control wellswere coated with 50 mg/ml BSA or 50 mg/ml MBP overnightat 48C. Wells were blocked with 200 ml of 2% MPBS for 2 hat 378C. WO-2 scFv and rFab antibody fragments from crudeperiplasmic extracts and purified preparations (5 mg/ml) wereincubated in wells with MBP-Ab1 – 42. Control wells wereincubated with the anti-FLAG M2 antibody for 1 h at RT.After washing, a 1/2000 dilution of anti-mouse IgGHRP-conjugate (GEB-Pierce, Rockford, IL, USA) was addedto each well and incubated for a further 1 h at 208C. Thesubstrate 2,20-Azino-bis(3-ethylbenzthiazoline-6-sulfonicacid) (ABTS, Roche, Indianapolis, USA) was added andabsorbance was read at 405 nm.
Biosensor binding analysisSurface plasmon resonance (SPR) measurements wereperformed using a Biacore T100 biosensor instrument(GE Healthcare, Uppsala, Sweden). All immobilisation andbinding experiments were performed at 258C in 1�HBS-EPþ buffer (10 mM HEPES, 150 mM NaCl, 3 mMEDTA, 0.05% surfactant P-20, pH 7.4). Analysis of the inter-action between Ab and WO-2 IgG was carried out using anindirect assay whereby WO-2 IgG was captured onto a
R.Robert et al.
200
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from
sensor chip (CM5) surface via rabbit anti-mouse (RAM) IgGantibody. A standard amine coupling protocol was employedto immobilise the RAM IgG antibody onto a surface viaexposed primary amines (Johnsson et al., 1991) in two separ-ate flow cells (1 and 2). This procedure resulted in immobil-isation of �12 000 RU (resonance units) of RAM IgG inboth flow cells. Two different versions of Ab were tested forbinding to captured WO-2 IgG: (i) synthetic Ab1 – 40 pep-tide and (ii) recombinant Im7-Ab1 – 18 fusion protein
(O.D., unpublished data) consisting of Ab residues 1–18fused to the C-terminus of Immunity protein 7—Im7(Kleanthous et al., 1998). Ab samples (2–64 nM)were injected over captured WO-2 and reference surfaces at30 ml/min. Association and dissociation phases were eachmonitored for 5 min (300 s). At least one buffer onlyinjection identical to Ab injections was included for thepurpose of double-referencing. RAM IgG surfaces wereregenerated with a single 45 s injection (at 30 ml/min) of
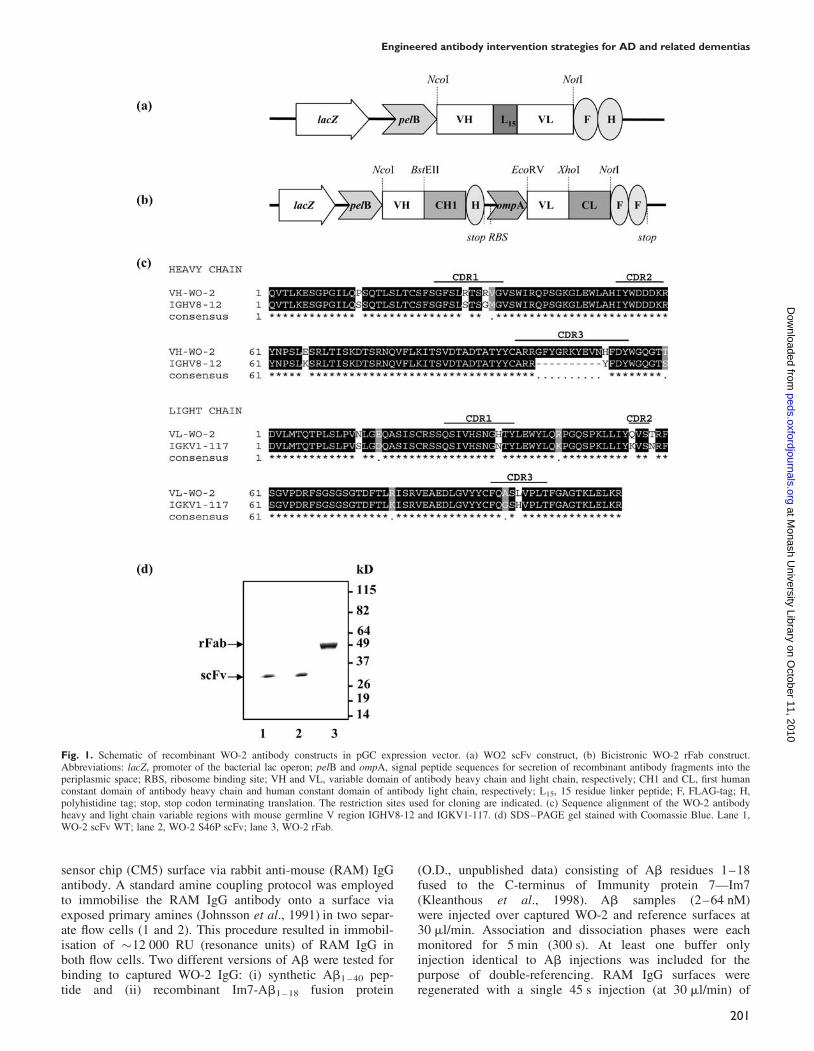
Fig. 1. Schematic of recombinant WO-2 antibody constructs in pGC expression vector. (a) WO2 scFv construct, (b) Bicistronic WO-2 rFab construct.Abbreviations: lacZ, promoter of the bacterial lac operon; pelB and ompA, signal peptide sequences for secretion of recombinant antibody fragments into theperiplasmic space; RBS, ribosome binding site; VH and VL, variable domain of antibody heavy chain and light chain, respectively; CH1 and CL, first humanconstant domain of antibody heavy chain and human constant domain of antibody light chain, respectively; L15, 15 residue linker peptide; F, FLAG-tag; H,polyhistidine tag; stop, stop codon terminating translation. The restriction sites used for cloning are indicated. (c) Sequence alignment of the WO-2 antibodyheavy and light chain variable regions with mouse germline V region IGHV8-12 and IGKV1-117. (d) SDS–PAGE gel stained with Coomassie Blue. Lane 1,WO-2 scFv WT; lane 2, WO-2 S46P scFv; lane 3, WO-2 rFab.
Engineered antibody intervention strategies for AD and related dementias
201
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from

10 mM Glycine, pH 1.5. Binding activities of various WO-2antibody fragments to its Ab antigen were evaluated using arecombinant Im7-Ab1 – 18 fusion protein form. As was thecase for WO-2 IgG, these measurements relied upon anindirect assay format whereby the Im7-Ab1 – 18 fusion proteinwas captured onto a chip surface via the Im7 natural bindingpartner, colicin E7 endonuclease domain of Escherichia coli(Ko et al., 1999), herein referred to as DNaseE7. Basic prin-ciples and concepts of this capture assay will be morethoroughly described elsewhere (Hosse & Dolezal, manu-script in preparation). A standard amine coupling protocolwas employed to immobilise DNaseE7 onto a CM5 chip inflow cells 3 and 4. This procedure resulted in immobilisationof �2500 RU of DNAseE7 protein. Im7-Ab1 – 18 sampleswere diluted (�0.1 mg/ml) and injected for 30 s at a flowrate of 30 ml/min over the DNaseE7 surface in flow cell 4resulting in a capture of �40 RU of fusion protein for scFvmeasurements and 20 RU for Fab measurements. WO-2 anti-body fragment preparations, typically spanning 2–128 nM inconcentration, were injected over the captured Im7-Ab1 – 18 ata flow rate of 30 ml/min. Association and dissociation phaseswere monitored for 3 and 5 min, respectively. At least onebuffer only injection identical to WO-2 fragment injectionswas included for the purpose of double-referencing.DNaseE7 surfaces were regenerated with a single 1 mininjection (at 30 ml/min) of Immuno-PureTM gentle elutionbuffer (GEB-Pierce).
Preparation of Ab1–42 peptideAn Ab1 – 42 synthetic lyophilised peptide (W. M. KeckLaboratory, Yale University, New Haven, CT, USA) was dis-solved in 1,1,1,3,3,3-hexafluoro-2-isopropanol (HFIP)(Sigma) and aliquoted. HFIP was removed by evaporationunder a fume hood and residual traces of HFIP wereremoved by drying under vacuum in a SpeedVac (SavantInstrument). The resulting peptide film was stored at 2808Cuntil required. For an Ab1 – 42 monomeric stock solutionpreparation, the peptide film was dissolved in either DMSO(dimethyl sulfoxide) or 20 mM NaOH for 1 h, sonicated for5 min and centrifuged at high speed for 10 min at 48C,immediately prior to use. To prepare an oligomeric pre-paration, the monomeric Ab1 – 42 stock solution was adjustedto 100 mM in ice-cold PBS, vortexed and incubated for 24 hat 48C with continuous rotation.
Inhibition of Ab1–42 fibrillisationThe monomeric Ab1 – 42 stock solution was diluted to 20 mMin PBS, pH 7.4, and incubated with or without 4 mM purifiedWO-2 IgG, WO-2 rFab, WO-2 scFv or NC10 anti-neuraminidase scFv antibody control (Dolezal et al., 2000)for 5 days at RT. Reactions were monitored by transmissionelectron microscopy (TEM) and thioflavine T (ThT) fluor-escence assay.
Disaggregation of preformed fibrilsAliquots of the monomeric Ab1 – 42 stock solution werediluted to 200 mM in PBS, pH 7.4, and incubated for 5 days atRT. Fibril formation was confirmed by TEM. The fibril sol-ution (200 mM) was next diluted 1:10 with either PBS, NC10scFv, mutant WO-2 S46P scFv, WO-2 rFab or WO-2 IgG(4 mM final concentration for each antibody). Reactions weremonitored after 24 h by TEM and ThT fluorescence assay.
Transmission electron microscopyCarbon-coated gold grids were glow-discharged under nitro-gen. Protein samples (0.1–0.3 mg/ml) were applied to thegrids and negatively stained with 2% potassium phospho-tungstate, pH 6.8. Grids were air-dried and examined usingan FEI Tecnai 12 transmission electron microscope operatingat 120 kV. Images were recorded using an OlympusMegaview III CCD camera.
ThT fluorescence assayAb1 – 42 fibrils were detected by incubating 5 ml of the reac-tions with 200 ml of 5 mM ThT solution (50 mM glycine,pH 9). The fluorescence intensity was measured at an exci-tation wavelength of 450 nm and an emission wavelength of482 nm with a spectrofluorometer LSB-50 (Perkin Elmer,Norwalk, CT, USA) with both excitation and emission band-widths of 5 nm as previously described (LeVine, 1999).Each data point represents the percentage of the maximumThT fluorescence intensity of the Ab1 – 42 fibrils in PBS aftersubtracting out the fluorescence signal from free ThT.
In vitro neuroprotection (MTT) assay againstoligomer-mediated toxicityHuman neuroblastoma cell line M17 was maintained inserum-free OPTI-MEM medium (Invitrogen, Australia)containing 0.1 mM non-essential amino acids, 50 IU/mlpenicillin, 50 mg/ml streptomycin (Gibco, Gaithersburg,MA, USA), 1 mM sodium pyruvate, 10% FCS in 5% CO2 at378C. Cells were seeded in 96-well tissue cultureplates (Nunc) at �104 cells per well and incubated for 24 h.Ab1 – 42 oligomers were incubated either alone or with puri-fied WO-2 IgG, WO-2 rFab, WO-2 S46P scFv or NC10 scFvfor 3 h at RT. These reactions (10 ml) were added to the cells(final concentration of peptide and antibodies of 15 and0.5 mM, respectively). Plates were incubated for anadditional 24 h at 378C.
Cell viability was determined using MTT toxicity assays.Briefly, 10 ml of 5 mg/ml MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] was added to eachwell and incubated for a further 24 h at 378C. Plates werecentrifuged, the medium was removed and 100 ml of 0.1 MHCl in isopropanol was added to each well to dissolve thecrystals. The absorbance was measured at 560 nm.
Results
Generation, expression and purification of theWO-2 rFab and scFvsThe monoclonal antibody WO-2 (IgG2a, k) (Ida et al., 1996)recognises amino acid residues 2–8 of Ab1 – 42 (Miles et al.,2008) in all forms (monomers, oligomers, protofibrils andfibrils) in different biochemical assays (Ida et al., 1996;Jensen et al., 2000). To construct the WO-2 scFv and chi-meric Fab, RNA was extracted from WO-2 hybridoma cellsand used as the VH and VL gene source. The VH (WO-2-H)and VL (WO-2-L) sequences (Fig. 1c) were analysed withIMGT/V-QUEST software (Giudicelli et al., 2004; Lefrancet al., 2005) and the IMGT/JunctionAnalysis tool (YousfiMonod et al., 2004) to identify the immunoglobulin germline V, (D) and J genes from which WO-2 variable regionsare derived. The mouse germ line V, (D) or J alleles most
R.Robert et al.
202
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from
similar to the WO-2-H nucleotide sequence by IMGT/V-QUEST are IGHV8-12*01, IGHD1-1*01 and IGHJ2*01,and those most similar to WO-2-L are IGKV1-117*01 andIGKJ5*01. The WO-2 scFv cassette (VH–VL oriented andseparated by a 15 amino acids linker) was cloned into thepGC expression vector, whereas the WO-2-H and WO-2-Lgenes were cloned sequentially into the pGC Fab expressionvector. Both constructs expressed protein in the periplasmicspace of TOP10 F’ E. coli cells and contained a C-terminalFLAG and 6-HIS tag for identification and affinity purifi-cation. SDS–PAGE profiles of the WO-2 scFv and rFabshowed proteins of expected sizes at �30 and �50 kDa,respectively (Fig. 1d). The activity of these purified frag-ments was assessed by ELISA and western blot against anMBP-Ab1 – 42 purified fusion protein. The fragments revealedhigh binding reactivity, similar to that of the purified parentalIgG in both assays (data not shown).
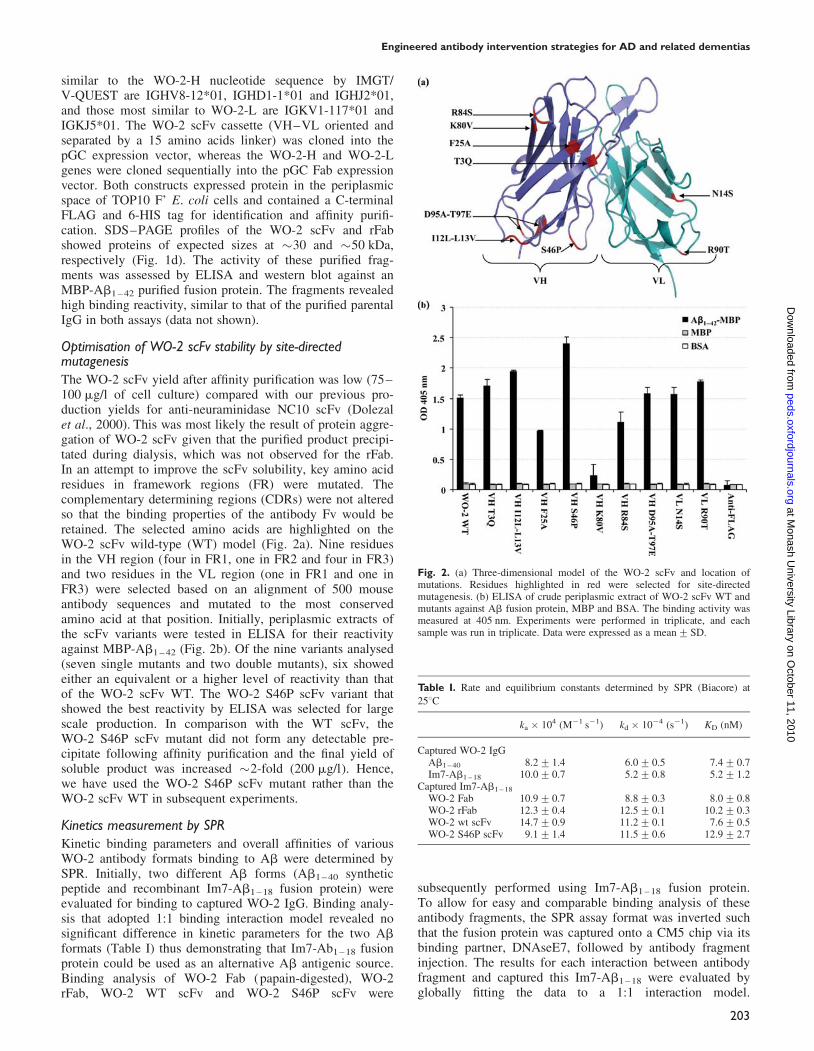
Optimisation of WO-2 scFv stability by site-directedmutagenesisThe WO-2 scFv yield after affinity purification was low (75–100 mg/l of cell culture) compared with our previous pro-duction yields for anti-neuraminidase NC10 scFv (Dolezalet al., 2000). This was most likely the result of protein aggre-gation of WO-2 scFv given that the purified product precipi-tated during dialysis, which was not observed for the rFab.In an attempt to improve the scFv solubility, key amino acidresidues in framework regions (FR) were mutated. Thecomplementary determining regions (CDRs) were not alteredso that the binding properties of the antibody Fv would beretained. The selected amino acids are highlighted on theWO-2 scFv wild-type (WT) model (Fig. 2a). Nine residuesin the VH region (four in FR1, one in FR2 and four in FR3)and two residues in the VL region (one in FR1 and one inFR3) were selected based on an alignment of 500 mouseantibody sequences and mutated to the most conservedamino acid at that position. Initially, periplasmic extracts ofthe scFv variants were tested in ELISA for their reactivityagainst MBP-Ab1 – 42 (Fig. 2b). Of the nine variants analysed(seven single mutants and two double mutants), six showedeither an equivalent or a higher level of reactivity than thatof the WO-2 scFv WT. The WO-2 S46P scFv variant thatshowed the best reactivity by ELISA was selected for largescale production. In comparison with the WT scFv, theWO-2 S46P scFv mutant did not form any detectable pre-cipitate following affinity purification and the final yield ofsoluble product was increased �2-fold (200 mg/l). Hence,we have used the WO-2 S46P scFv mutant rather than theWO-2 scFv WT in subsequent experiments.
Kinetics measurement by SPRKinetic binding parameters and overall affinities of variousWO-2 antibody formats binding to Ab were determined bySPR. Initially, two different Ab forms (Ab1 – 40 syntheticpeptide and recombinant Im7-Ab1 – 18 fusion protein) wereevaluated for binding to captured WO-2 IgG. Binding analy-sis that adopted 1:1 binding interaction model revealed nosignificant difference in kinetic parameters for the two Ab
formats (Table I) thus demonstrating that Im7-Ab1 – 18 fusionprotein could be used as an alternative Ab antigenic source.Binding analysis of WO-2 Fab (papain-digested), WO-2rFab, WO-2 WT scFv and WO-2 S46P scFv were
subsequently performed using Im7-Ab1 – 18 fusion protein.To allow for easy and comparable binding analysis of theseantibody fragments, the SPR assay format was inverted suchthat the fusion protein was captured onto a CM5 chip via itsbinding partner, DNAseE7, followed by antibody fragmentinjection. The results for each interaction between antibodyfragment and captured this Im7-Ab1 – 18 were evaluated byglobally fitting the data to a 1:1 interaction model.
Table I. Rate and equilibrium constants determined by SPR (Biacore) at
258C
ka � 104 (M21 s21) kd � 1024 (s21) KD (nM)
Captured WO-2 IgGAb1 – 40 8.2+1.4 6.0+0.5 7.4+0.7Im7-Ab1 – 18 10.0+0.7 5.2+0.8 5.2+1.2
Captured Im7-Ab1 – 18
WO-2 Fab 10.9+0.7 8.8+0.3 8.0+0.8WO-2 rFab 12.3+0.4 12.5+0.1 10.2+0.3WO-2 wt scFv 14.7+0.9 11.2+0.1 7.6+0.5WO-2 S46P scFv 9.1+1.4 11.5+0.6 12.9+2.7
Fig. 2. (a) Three-dimensional model of the WO-2 scFv and location ofmutations. Residues highlighted in red were selected for site-directedmutagenesis. (b) ELISA of crude periplasmic extract of WO-2 scFv WT andmutants against Ab fusion protein, MBP and BSA. The binding activity wasmeasured at 405 nm. Experiments were performed in triplicate, and eachsample was run in triplicate. Data were expressed as a mean+SD.
Engineered antibody intervention strategies for AD and related dementias
203
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from
Calculated association (ka) and dissociation (kd) rate con-stants as well as the overall KD values are shown in Table I.No significant affinity differences were observed betweenrecombinant and parental monoclonal antibody formats.Thus, the WO-2 rFab, the WO-2 WT scFv and the WO-2S46P scFv bound to Ab antigen with comparable affinity tothat of the parental IgG and the papain-digested Fab.
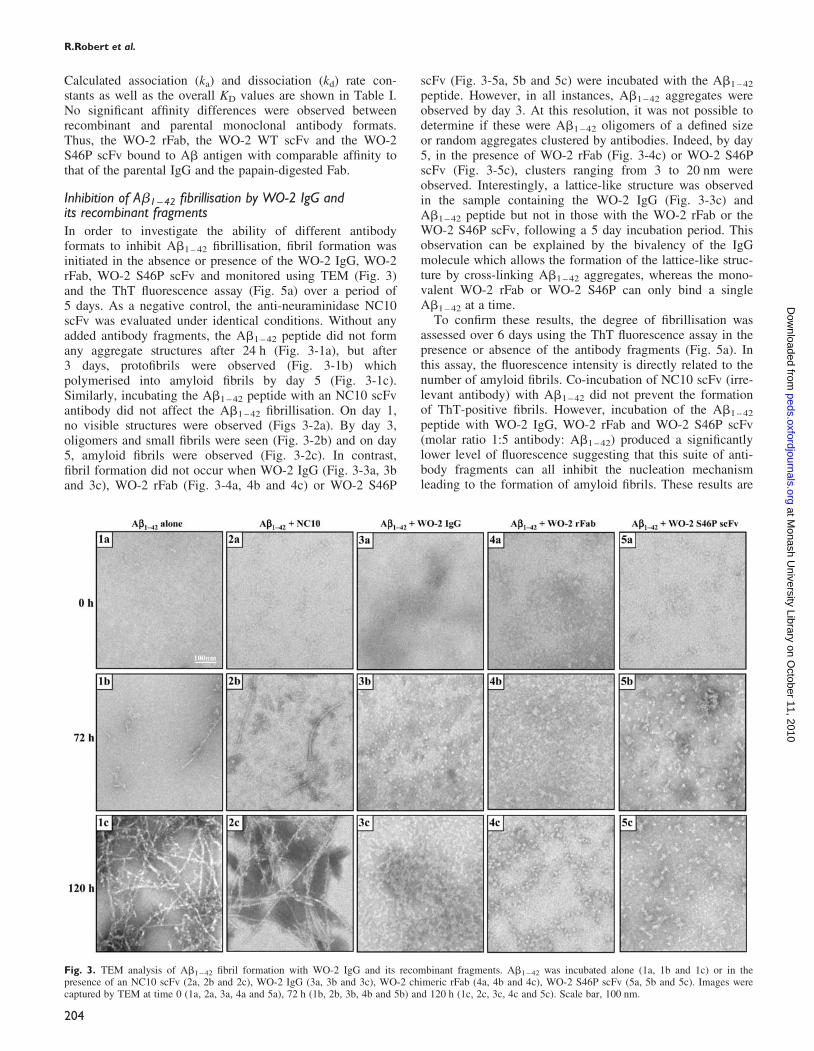
Inhibition of Ab1–42 fibrillisation by WO-2 IgG andits recombinant fragmentsIn order to investigate the ability of different antibodyformats to inhibit Ab1 – 42 fibrillisation, fibril formation wasinitiated in the absence or presence of the WO-2 IgG, WO-2rFab, WO-2 S46P scFv and monitored using TEM (Fig. 3)and the ThT fluorescence assay (Fig. 5a) over a period of5 days. As a negative control, the anti-neuraminidase NC10scFv was evaluated under identical conditions. Without anyadded antibody fragments, the Ab1 – 42 peptide did not formany aggregate structures after 24 h (Fig. 3-1a), but after3 days, protofibrils were observed (Fig. 3-1b) whichpolymerised into amyloid fibrils by day 5 (Fig. 3-1c).Similarly, incubating the Ab1 – 42 peptide with an NC10 scFvantibody did not affect the Ab1 – 42 fibrillisation. On day 1,no visible structures were observed (Figs 3-2a). By day 3,oligomers and small fibrils were seen (Fig. 3-2b) and on day5, amyloid fibrils were observed (Fig. 3-2c). In contrast,fibril formation did not occur when WO-2 IgG (Fig. 3-3a, 3band 3c), WO-2 rFab (Fig. 3-4a, 4b and 4c) or WO-2 S46P
scFv (Fig. 3-5a, 5b and 5c) were incubated with the Ab1 – 42
peptide. However, in all instances, Ab1 – 42 aggregates wereobserved by day 3. At this resolution, it was not possible todetermine if these were Ab1 – 42 oligomers of a defined sizeor random aggregates clustered by antibodies. Indeed, by day5, in the presence of WO-2 rFab (Fig. 3-4c) or WO-2 S46PscFv (Fig. 3-5c), clusters ranging from 3 to 20 nm wereobserved. Interestingly, a lattice-like structure was observedin the sample containing the WO-2 IgG (Fig. 3-3c) andAb1 – 42 peptide but not in those with the WO-2 rFab or theWO-2 S46P scFv, following a 5 day incubation period. Thisobservation can be explained by the bivalency of the IgGmolecule which allows the formation of the lattice-like struc-ture by cross-linking Ab1 – 42 aggregates, whereas the mono-valent WO-2 rFab or WO-2 S46P can only bind a singleAb1 – 42 at a time.
To confirm these results, the degree of fibrillisation wasassessed over 6 days using the ThT fluorescence assay in thepresence or absence of the antibody fragments (Fig. 5a). Inthis assay, the fluorescence intensity is directly related to thenumber of amyloid fibrils. Co-incubation of NC10 scFv (irre-levant antibody) with Ab1 – 42 did not prevent the formationof ThT-positive fibrils. However, incubation of the Ab1 – 42
peptide with WO-2 IgG, WO-2 rFab and WO-2 S46P scFv(molar ratio 1:5 antibody: Ab1 – 42) produced a significantlylower level of fluorescence suggesting that this suite of anti-body fragments can all inhibit the nucleation mechanismleading to the formation of amyloid fibrils. These results are
Fig. 3. TEM analysis of Ab1 – 42 fibril formation with WO-2 IgG and its recombinant fragments. Ab1 – 42 was incubated alone (1a, 1b and 1c) or in thepresence of an NC10 scFv (2a, 2b and 2c), WO-2 IgG (3a, 3b and 3c), WO-2 chimeric rFab (4a, 4b and 4c), WO-2 S46P scFv (5a, 5b and 5c). Images werecaptured by TEM at time 0 (1a, 2a, 3a, 4a and 5a), 72 h (1b, 2b, 3b, 4b and 5b) and 120 h (1c, 2c, 3c, 4c and 5c). Scale bar, 100 nm.
R.Robert et al.
204
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from
consistent with the TEM data, and together indicate thatWO-2 as a whole antibody or as engineered fragments is aneffective inhibitor of fibrillisation.
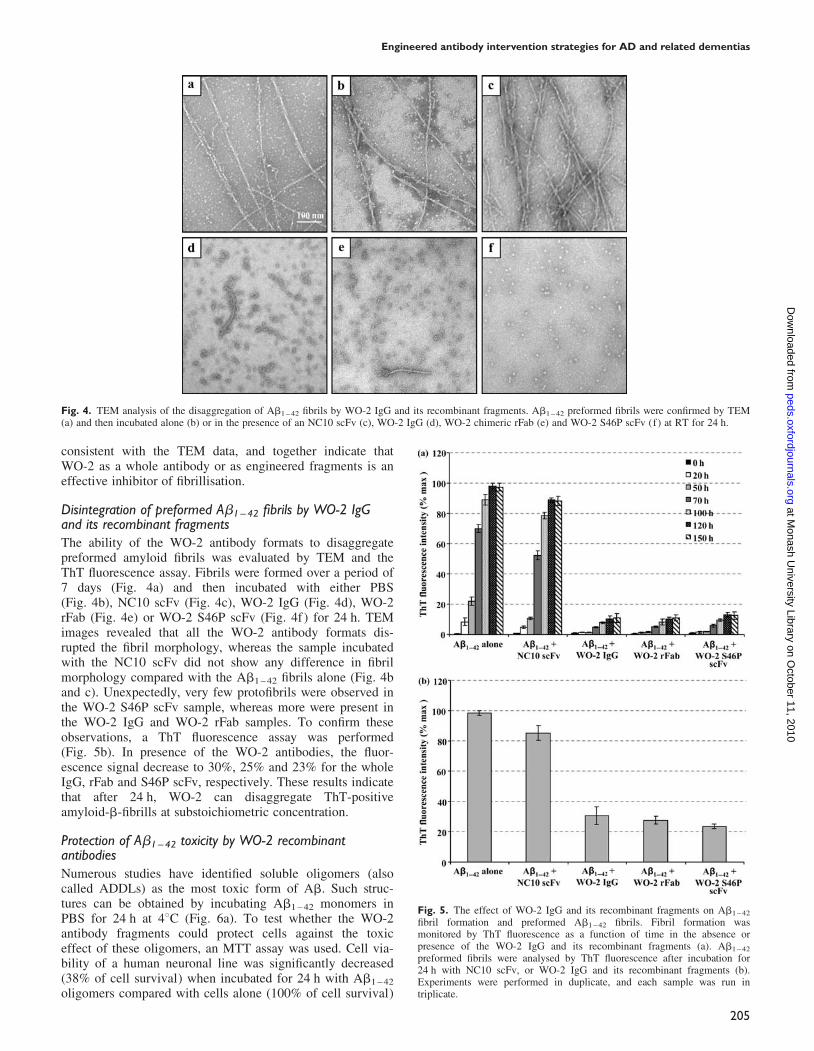
Disintegration of preformed Ab1–42 fibrils by WO-2 IgGand its recombinant fragmentsThe ability of the WO-2 antibody formats to disaggregatepreformed amyloid fibrils was evaluated by TEM and theThT fluorescence assay. Fibrils were formed over a period of7 days (Fig. 4a) and then incubated with either PBS(Fig. 4b), NC10 scFv (Fig. 4c), WO-2 IgG (Fig. 4d), WO-2rFab (Fig. 4e) or WO-2 S46P scFv (Fig. 4f ) for 24 h. TEMimages revealed that all the WO-2 antibody formats dis-rupted the fibril morphology, whereas the sample incubatedwith the NC10 scFv did not show any difference in fibrilmorphology compared with the Ab1 – 42 fibrils alone (Fig. 4band c). Unexpectedly, very few protofibrils were observed inthe WO-2 S46P scFv sample, whereas more were present inthe WO-2 IgG and WO-2 rFab samples. To confirm theseobservations, a ThT fluorescence assay was performed(Fig. 5b). In presence of the WO-2 antibodies, the fluor-escence signal decrease to 30%, 25% and 23% for the wholeIgG, rFab and S46P scFv, respectively. These results indicatethat after 24 h, WO-2 can disaggregate ThT-positiveamyloid-b-fibrills at substoichiometric concentration.
Protection of Ab1–42 toxicity by WO-2 recombinantantibodiesNumerous studies have identified soluble oligomers (alsocalled ADDLs) as the most toxic form of Ab. Such struc-tures can be obtained by incubating Ab1 – 42 monomers inPBS for 24 h at 48C (Fig. 6a). To test whether the WO-2antibody fragments could protect cells against the toxiceffect of these oligomers, an MTT assay was used. Cell via-bility of a human neuronal line was significantly decreased(38% of cell survival) when incubated for 24 h with Ab1 – 42
oligomers compared with cells alone (100% of cell survival)
Fig. 4. TEM analysis of the disaggregation of Ab1 – 42 fibrils by WO-2 IgG and its recombinant fragments. Ab1 – 42 preformed fibrils were confirmed by TEM(a) and then incubated alone (b) or in the presence of an NC10 scFv (c), WO-2 IgG (d), WO-2 chimeric rFab (e) and WO-2 S46P scFv (f) at RT for 24 h.
Fig. 5. The effect of WO-2 IgG and its recombinant fragments on Ab1 –42
fibril formation and preformed Ab1 – 42 fibrils. Fibril formation wasmonitored by ThT fluorescence as a function of time in the absence orpresence of the WO-2 IgG and its recombinant fragments (a). Ab1 – 42
preformed fibrils were analysed by ThT fluorescence after incubation for24 h with NC10 scFv, or WO-2 IgG and its recombinant fragments (b).Experiments were performed in duplicate, and each sample was run intriplicate.
Engineered antibody intervention strategies for AD and related dementias
205
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from
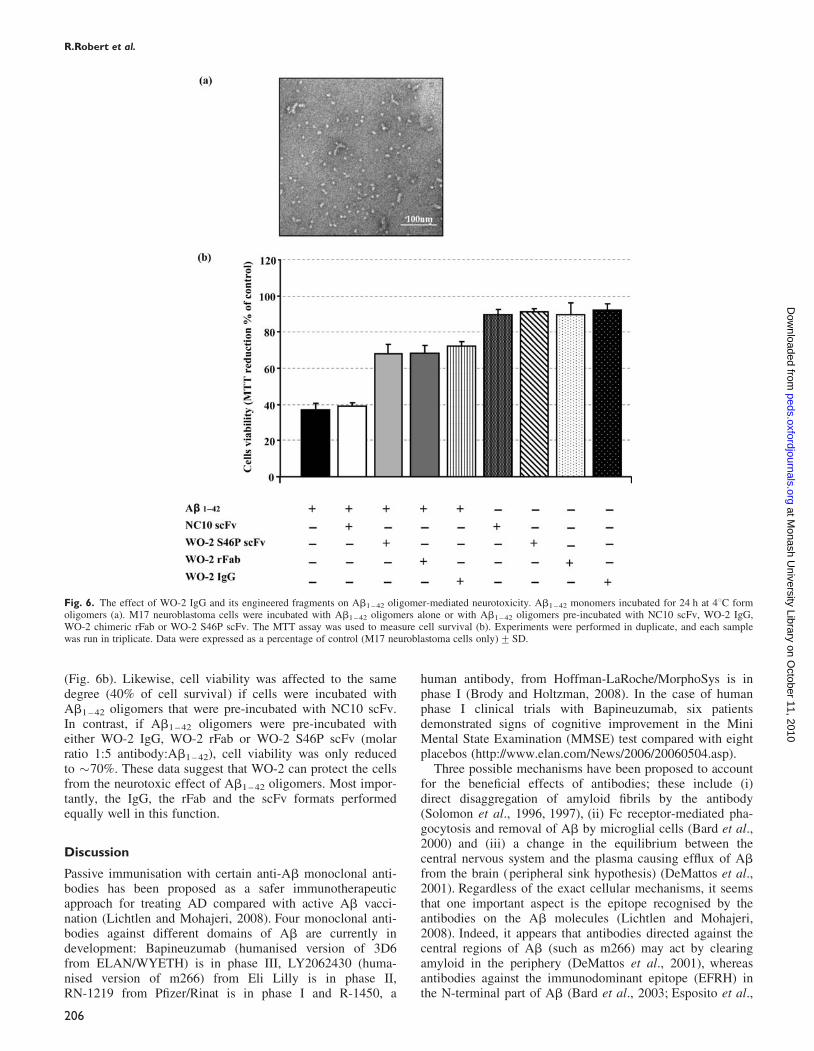
(Fig. 6b). Likewise, cell viability was affected to the samedegree (40% of cell survival) if cells were incubated withAb1 – 42 oligomers that were pre-incubated with NC10 scFv.In contrast, if Ab1 – 42 oligomers were pre-incubated witheither WO-2 IgG, WO-2 rFab or WO-2 S46P scFv (molarratio 1:5 antibody:Ab1 – 42), cell viability was only reducedto �70%. These data suggest that WO-2 can protect the cellsfrom the neurotoxic effect of Ab1 – 42 oligomers. Most impor-tantly, the IgG, the rFab and the scFv formats performedequally well in this function.
Discussion
Passive immunisation with certain anti-Ab monoclonal anti-bodies has been proposed as a safer immunotherapeuticapproach for treating AD compared with active Ab vacci-nation (Lichtlen and Mohajeri, 2008). Four monoclonal anti-bodies against different domains of Ab are currently indevelopment: Bapineuzumab (humanised version of 3D6from ELAN/WYETH) is in phase III, LY2062430 (huma-nised version of m266) from Eli Lilly is in phase II,RN-1219 from Pfizer/Rinat is in phase I and R-1450, a
human antibody, from Hoffman-LaRoche/MorphoSys is inphase I (Brody and Holtzman, 2008). In the case of humanphase I clinical trials with Bapineuzumab, six patientsdemonstrated signs of cognitive improvement in the MiniMental State Examination (MMSE) test compared with eightplacebos (http://www.elan.com/News/2006/20060504.asp).
Three possible mechanisms have been proposed to accountfor the beneficial effects of antibodies; these include (i)direct disaggregation of amyloid fibrils by the antibody(Solomon et al., 1996, 1997), (ii) Fc receptor-mediated pha-gocytosis and removal of Ab by microglial cells (Bard et al.,2000) and (iii) a change in the equilibrium between thecentral nervous system and the plasma causing efflux of Abfrom the brain (peripheral sink hypothesis) (DeMattos et al.,2001). Regardless of the exact cellular mechanisms, it seemsthat one important aspect is the epitope recognised by theantibodies on the Ab molecules (Lichtlen and Mohajeri,2008). Indeed, it appears that antibodies directed against thecentral regions of Ab (such as m266) may act by clearingamyloid in the periphery (DeMattos et al., 2001), whereasantibodies against the immunodominant epitope (EFRH) inthe N-terminal part of Ab (Bard et al., 2003; Esposito et al.,
Fig. 6. The effect of WO-2 IgG and its engineered fragments on Ab1 – 42 oligomer-mediated neurotoxicity. Ab1 – 42 monomers incubated for 24 h at 48C formoligomers (a). M17 neuroblastoma cells were incubated with Ab1 – 42 oligomers alone or with Ab1 – 42 oligomers pre-incubated with NC10 scFv, WO-2 IgG,WO-2 chimeric rFab or WO-2 S46P scFv. The MTT assay was used to measure cell survival (b). Experiments were performed in duplicate, and each samplewas run in triplicate. Data were expressed as a percentage of control (M17 neuroblastoma cells only)+SD.
R.Robert et al.
206
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from

2008) have been reported to cross the blood-brain barrier andact on Ab in brain tissue (Bard et al., 2000). Unfortunately,passive administration of these anti-Ab antibodies directedagainst the N-terminal part of Ab to AD mice models havealso been reported to trigger inflammatory processes such ascerebral microhemorrhages (Pfeifer et al., 2002). This hasprompted the generation of engineered antibody fragmentsincluding scFvs (Frenkel et al., 2000; Liu et al., 2004;Fukuchi et al., 2006; Solorzano-Vargas et al., 2008; Yueet al., 2008), rFabs and V-domains (Habicht et al., 2007)which lack the Fc region. Given that IgG molecules havelimited access to the brain (Banks et al., 2002), it is antici-pated that due to their smaller size, these antibody fragmentsmay enter more efficiently into this area.
In this study, we constructed and characterised antibodyfragments of the monoclonal antibody WO-2 recognising theN-terminal region of the Ab peptide (amino acids 2–8).Although other anti-Ab scFvs have been described for theirimmunotherapeutic potential, this work is the first examplein which three different formats have been compared.Moreover, to our knowledge, this study is the first todescribe a recombinant chimeric Fab fragment displayinganti-aggregating, disaggregating and neuroprotectiveactivities.
Given that the amount of soluble functional WO-2 WTscFv (100 mg/l) produced was not sufficient to carry out bio-chemical assays, our first step was to improve stability bymutating WO-2 WT scFv in the VH and VL FR usingrational design and site-directed mutagenesis. Of the variantsgenerated, the WO-2 S46P scFv with a point mutation inthe VH FR2 region produced the best results including a2-fold increase in final yield of soluble product (100 versus200 mg/l) and no precipitate. This finding was consistentwith other studies which demonstrated not only that the VHFR2 is an important region for scFv stability (Masuda et al.,2006) but similar substitutions (any amino acid to a proline)resulted in increased stability (Matthews et al., 1987).Likewise, reformatting the WO-2 scFv into an rFab furtherenhanced the stability which probably resulted from theadditional binding forces provided by the constant regions(Quintero-Hernandez et al., 2007).
Binding kinetics of the three fragments (WO-2 scFv WT,WO-2 S46P scFv and WO-2 rFab) revealed that the affinitieswere essentially the same as the parental IgG and as thepapain-digested Fab. Furthermore, all were in the nanomolarrange required for therapeutic applications. These datasuggest that the three-dimensional structure of the bindingdomains of the parental IgG is not affected by the insertionof a linker for the scFv constructs or the grafting of humanconstant region for the rFab. Our current work involves gen-erating a humanised recombinant antibody by CDR-graftingand further improvement of the expression yield will be doneon these humanised versions of WO-2.
Next, we determined in biochemical assays in vitro theimmunotherapeutic potential of the three formats. The resultspresented here suggest that this suite of antibodies shows theability to inhibit Ab1 – 42 fibril formation. One possible mech-anism for inhibition involves the binding of antibody toAb1 – 42 molecules preventing the filament nucleation process(Chirita et al., 2004). In our case, WO-2 appears to lead tothe formation of amorphous aggregates suggesting that theantibodies cannot inhibit the early stage of aggregation as
previously described by Mamikonyan et al. (2007) with apolyclonal antibody preparation. Interestingly, theco-incubation of WO-2 IgG with Ab1 – 42 molecules leads toa lattice-like structure which has not been previously reportedin similar assays (Legleiter et al., 2004; Mamikonyan et al.,2007). Presumably, the bivalency of the IgG moleculesallows cross-linking of small amorphous aggregates present-ing multiple epitopes, resulting in the lattice structures thatare not observed with the monovalent binders, WO-2 scFvand WO-2 rFab. Moreover, these antibody fragments canalso disaggregate preformed fibrils. Again, the exact mechan-ism remains unclear. It has been proposed that the high affi-nity antibody binding to soluble Ab1 – 42 species disturbs theequilibrium between fibrillar and non-fibrillar Ab1 – 42 andthus causes the amyloid fibrils to disaggregate (Chirita et al.,2004).
The antibodies were also tested in the context of cellularneurotoxicity in the M17 human neuroblastoma cell line. Inall instances, WO-2 IgG, WO-2 rFab and WO-2 S46P scFvreduced the neurotoxicity caused by Ab1 – 42 oligomers.These results are promising as oligomers have beensuggested to be the most toxic form of Ab and are involvedin the early stage of AD pathogenesis (Lambert et al., 1998;Walsh et al., 2002).
Recently, it has been demonstrated that expression of intra-bodies directed against the EFRH motif of Ab can decreaseand prevent the generation of Ab peptide by shielding theb-secretase cleavage site of APP (Paganetti et al., 2005).Therefore, it will be worth testing the in situ expression ofWO-2 S46P scFv as an intrabody in a similar system. Inaddition, testing current WO-2 antibody formats in an ADmouse model will allow a direct comparison of the efficiencyon amyloid load and cognitive function of such moleculesin vivo.
It is likely that all the advantages of engineered antibodyfragments (smaller size, low cost production in microbialexpression systems, possibility to express as intracellularprotein and so on) will make these molecules attractive toolsfor safer immunotherapy of AD and other neurodegenerativediseases.
Acknowledgements
We thank Pr K. Beyreuther for providing us the WO-2 hybridoma cell line,Dr Qiao-Xin Li for the Ab1 – 40 peptide sample and Dr J. Caine for theAb-MBP fusion protein. We are very grateful to Lesley Pearce for hertechnical assistance. We thank Dr S.D. Nuttall and Dr N.M. McKern forcritical reading of the manuscript.
ReferencesArimon,M., Dıez-Perez,I., Kogan,M.J., Durany,N., Giralt,E., Sanz,F. and
Fernandez-Busquets,X. (2005) FASEB J., 10, 1344–1346.Banks,W.A., Terrell,B., Farr,S.A., Robinson,S.M., Nonaka,N. and
Morley,J.E. (2002) Peptides, 23, 2223–2226.Bard,F., et al. (2000) Nat. Med., 6, 916–919.Bard,F., et al. (2003) Proc. Natl Acad. Sci. USA, 100, 2023–2028.Brody,D.L. and Holtzman,D.M. (2008) Annu. Rev. Neurosci., 31, 175–193.Brookmeyer,R., Johnson,E., Ziegler-Graham,K. and Arrighi,M.H. (2007)
Dementia, 3, 186–191.Caine,J., Volitakis,I., Cherny,R., Varghese,J. and Macreadie,I. (2007)
Protein Pept. Lett., 14, 83–86.Cherny,R.A., et al. (2001) Neuron, 30, 665–676.Chirita,C., Necula,M. and Kuret,J. (2004) Biochemistry, 43, 2879–2887.Coia,G., Ayres,A., Lilley,G.G., Hudson,P.J. and Irving,R.A. (1997) Gene,
201, 203–209.
Engineered antibody intervention strategies for AD and related dementias
207
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from

DeMattos,R.B., Bales,K.R., Cummins,D.J., Dodart,J.C., Paul,S.M. andHoltzman,D.M. (2001) Proc. Natl Acad. Sci. USA, 98, 8850–8855.
Dolezal,O., Pearce,L.A., Lawrence,L.J., McCoy,A.J., Hudson,P.J. andKortt,A.A. (2000) Protein Eng., 13, 565–574.
Esposito,M., Luccarini,I., Cicatiello,V., De Falco,D., Fiorentini,A., Barba,P.,Casamenti,F. and Prisco,A. (2008) Mol. Immunol., 45, 1056–1062.
Frenkel,D., Solomon,B. and Benhar,I. (2000) J. Neuroimmunol., 106, 23–31.Fukuchi,K., Tahara,K., Kim,H.D., Maxwell,J.A., Lewis,T.L.,
Accavitti-Loper,M.A., Kim,H., Ponnazhagan,S. and Lalonde,R. (2006)Neurobiol. Dis., 2, 502–511.
Gilliland,L.K., Norris,N.A., Marquardt,H., Tsu,T.T., Hayden,M.S.,Neubauer,M.G., Yelton,D.E., Mittler,R.S. and Ledbetter,J.A. (1996)Tissue Antigens, 47, 1–20.
Giudicelli,V., Chaume,D. and Lefranc,M.P. (2004) Nucleic Acids Res., 32,W435–W440.
Gruen,L.C., Kort,A.A. and Nice,E. (1993) Eur. J. Biochem., 217, 319–325.Habicht,G., et al. (2007) Proc. Natl Acad. Sci. USA, 104, 19232–19237.Holliger,P. and Hudson,P.J. (2005) Nat. Biotechnol., 23, 1126–1136.Ida,N., Hartmann,T., Pantel,J., Schroder,J., Zerfass,R., Forstl,H.,
Sandbrink,R., Masters,C.L. and Beyreuther,K. (1996) J. Biol. Chem., 271,22908–22914.
Janus,C., et al. (2000) Nature, 408, 979–982.Jensen,M., et al. (2000) Mol. Med., 6, 291–302.Johnsson,B., Lofas,S. and Lindquist,G. (1991) Anal Biochem., 198,
268–277.Kang,J., et al. (1987) Nature, 325, 733–736.Kleanthous,C., Hemmings,A.M., Moore,G.R. and James,R. (1998) Mol.
Microbiol., 2, 227–233.Ko,T.P., Liao,C.C., Ku,W.Y., Chak,K.F. and Yuan,H.S. (1999) Structure, 7,
91–102.Kortt,A.A., Malby,R.L., Caldwell,J.B., Gruen,L.C., Ivancic,N.M.,
Lawrence,M.C., Howlett,G.J., Webster,R.G., Hudson,P.J. andColman,P.M. (1994) Eur. J. Biochem., 221, 151–157.
Lambert,M.P., et al. (1998) Proc. Natl Acad. Sci. USA, 95, 6448–6453.Lefranc,M.P., Giudicelli,V., Kaas,Q., Duprat,E., Jabado-Michaloud,J.,
Scaviner,D., Ginestoux,C., Clement,O., Chaume,D. and Lefranc,G. (2005)Nucleic Acids. Res., 33, D593–D597.
Legleiter,J., Czilli,D.L., Gitter,B., DeMattos,R.B., Holtzman,D.M. andKowalewski,T. (2004) J. Mol. Biol., 335, 997–1006.
LeVine,H., 3rd (1999) Methods Enzymol., 309, 274–284.Levites,Y., Jansen,K., Smithson,L.A., Dakin,R., Holloway,V.M., Das,P. and
Golde,T.E. (2006) J. Neurosci., 26, 11923–11928.Lichtlen,P. and Mohajeri,M.H. (2008) J. Neurochem., 104, 859–874.Liu,R., Yuan,B., Emadi,S., Zameer,A., Schulz,P., McAllister,C.,
Lyubchenko,Y., Goud,G. and Sierks,M.R. (2004) Biochemistry, 43,6959–6967.
Lorenzo,A. and Yankner,B.A. (1994) Proc. Natl Acad. Sci. USA, 91,12243–12247.
Mamikonyan,G., et al. (2007) J. Biol. Chem., 282, 22376–22386.Masters,C.L. and Beyreuther,K. (2006) Brain, 129, 2823–2839.Masuda,K., Sakamoto,K., Kojima,M., Aburatani,T., Ueda,T. and Ueda,H.
(2006) FEBS J., 273, 2184–2194.Matthews,B.W., Nicholson,H. and Becktel,W.J. (1987) Proc. Natl Acad. Sci.
USA, 84, 6663–6667.McGleenon,B.M., Dynan,K.B. and Passmore,A.P. (1999) Br. J. Clin.
Pharmacol., 48, 471–480.Miles,L.A., et al. (2008) J. Mol. Biol., 377, 181–192.Orgogozo,J.M., et al. (2003) Neurology, 61, 46–54.Paganetti,P., Calanca,V., Galli,C., Stefani,M. and Molinari,M. (2005) J. Cell.
Biol., 168, 863–868.Pfeifer,M., Boncristiano,S., Bondolfi,L., Stalder,A., Seller,T., Staufenbiel,M.,
Mathews,P.M. and Jucker,M. (2002) Science, 298, 1379.Quintero-Hernandez,V., Juarez-Gonzalez,V.R., Ortız-Leon,M., Sanchez,R.,
Possani,L.D. and Becerril,B. (2007) Mol. Immunol., 44, 1307–1315.Racke,M.M., et al. (2005) J. Neurosci., 25, 629–636.Schenk,D., et al. (1999) Nature, 400, 173–177.Selkoe,D.J. (1999) Nature, 399, A23–A31.Selkoe,D.J. (2002) Science, 298, 789–791.Solomon,B., Koppel,R., Hanan,E. and Katzav,T. (1996) Proc. Natl Acad.
Sci. USA, 93, 452–455.Solomon,B., Koppel,R., Frankel,D. and Hanan-Aharon,E. (1997) Proc. Natl
Acad. Sci. USA, 94, 4109–4112.Solorzano-Vargas,R.S., et al. (2008) Mol. Immunol., 45, 881–886.Suh,Y.H. and Checler,F. (2002) Pharmacol. Rev., 54, 469–525.Tamura,Y., Hamajima,K., Matsui,K., Yanoma,S., Narita,M., Tajima,N.,
Xin,K.Q., Klinman,D. and Okuda,K. (2005) Neurobiol. Dis., 20,541–549.
Walsh,D.M., Klyubin,I., Fadeeva,J.V., Cullen,W.K., Anwyl,R., Wolfe,M.S.,Rowan,M.J. and Selkoe,D.J. (2002) Nature, 416, 535–539.
Weiner,H.L. and Frenkel,D. (2006) Nat. Rev. Immunol., 6, 404–416.Whitelegg,N.R. and Rees,A.R. (2000) Protein Eng., 13, 819–824.Wilcock,D.M., Alamed,J., Gottschall,P.E., Grimm,J., Rosenthal,A., Pons,J.,
Ronan,V., Symmonds,K., Gordon,M.N. and Morgan,D. (2006)J. Neurosci., 26, 5340–5346.
Yang,F., et al. (2005) J. Biol. Chem., 280, 5892–5901.Yousfi Monod,M., Giudicelli,V., Chaume,D. and Lefranc,M.P. (2004)
Bioinformatics, 20 (Suppl. 1), i379–i385.Yue,S., Li,Y., Wang,X., Bai,H., Xia,J., Jiang,L., Ji,Y., Fan,L., He,Z. and
Chen,Q. (2008) Life. Sci., 82, 1249–1255.Zhao,B., Yu,M., Neitzel,M., Marugg,J., Jagodzinski,J., Lee,M., Hu,K.,
Schenk,D., Yednock,T. and Basi,G. (2008) J. Biol. Chem., 283,2927–2938.
Received September 9, 2008; revised September 9, 2008;accepted September 9, 2008
Edited by Anna Wu
R.Robert et al.
208
at Monash U
niversity Library on October 11, 2010
peds.oxfordjournals.orgD
ownloaded from
Copyright © 2022 FDOKUMEN




















