
The Intrinsic Properties of Reconstituted Soils - ScholarWorks ...
Upload
independentCategory
view
2download
0
ISSN: 1524-4571 Copyright © 2010 American Heart Association. All rights reserved. Print ISSN: 0009-7330. Online
TX 72514Circulation Research is published by the American Heart Association. 7272 Greenville Avenue, Dallas,
DOI: 10.1161/CIRCRESAHA.109.206326 published online Apr 29, 2010; Circ. Res.
Sardone, Giacomo Matteucci, Georg C. Terstappen and Salvatore Oliviero Francesco Neri, Maurizio Orlandini, Felice Petraglia, Stefano Gotta, Gian Luca Federico Galvagni, Susanna Pennacchini, Ahmad Salameh, Marina Rocchigiani,
Kinase ActivityVEGFR-3 Phosphorylation Without the Activation of the Receptor Intrinsic
Endothelial Cell Adhesion to the Extracellular Matrix Induces c-Src�Dependent
http://circres.ahajournals.org/cgi/content/full/CIRCRESAHA.109.206326/DC1Data Supplement (unedited) at:
http://circres.ahajournals.org
located on the World Wide Web at: The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circres.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation Research is online at
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Endothelial Cell Adhesion to the Extracellular MatrixInduces c-Src–Dependent VEGFR-3 Phosphorylation
Without the Activation of the Receptor IntrinsicKinase Activity
Federico Galvagni, Susanna Pennacchini, Ahmad Salameh, Marina Rocchigiani, Francesco Neri,Maurizio Orlandini, Felice Petraglia, Stefano Gotta, Gian Luca Sardone, Giacomo Matteucci,
Georg C. Terstappen, Salvatore Oliviero
Rationale: Integrins cooperate with growth factor receptors to promote downstream signaling for cell proliferationand migration. However, the mechanism of receptor activation is still unknown.
Objective: To analyze the mechanism of phosphorylation of the vascular endothelial growth factor receptor(VEGFR)-3 by cell adhesion.
Methods and Results: We show that VEGFR-3 phosphorylation, induced by cell attachment to the extracellularmatrix, is independent from the intrinsic kinase activity of the receptor, as evidenced from phosphorylation celladhesion experiments with a mutant kinase dead receptor or in the presence of the specific kinase inhibitor MAZ51. Cell adhesion experiments in the presence of the c-Src inhibitor PP2 or in fibroblast triple knockout for c-Src,Yes, and Fyn (SYF) demonstrate that VEGFR-3 phosphorylation, induced by extracellular matrix, is mediatedby c-Src. Kinase assays in vitro with recombinant c-Src show that VEGFR-3 is a direct c-Src target and massspectrometry analysis identified the sites phosphorylated by c-Src as tyrosine 830, 833, 853, 1063, 1333, and 1337,demonstrating that integrin-mediated receptor phosphorylation induces a phosphorylation pattern that isdistinct from that induced by growth factors. Furthermore, pull-down assays show that integrin-mediatedVEGFR-3 phosphorylation activates the recruitment to the receptor of the adaptor proteins CRKI/II and SHCinducing activation of JNK.
Conclusions: These data suggest that cell adhesion to extracellular matrix induces a downstream signaling using thetyrosine kinase receptor VEGFR-3 as scaffold. (Circ Res. 2010;106:00-00.)
Key Words: cell adhesion � endothelial cells � angiogenesis � biochemistry � signaling
Crosstalk between integrins and growth factor receptorsplays an important role in vascular development and its
maintenance. Several examples demonstrated that crosstalkbetween integrins and tyrosine kinase receptors is requiredfor growth factor–induced biological processes to ensure cellgrowth, survival, and differentiation in normal and patholog-ical processes.1 The angiogenic processes require the coordi-nation of signals from the extracellular environment toactivates specific tyrosine kinase receptors and integrins.1,2
Integrins associating with growth factor receptors regulate thecapacity of the integrin/receptor complexes to propagatedownstream signals.3,4 Integrin-dependent activation of re-ceptor tyrosine kinases is a general mechanism to enhancegrowth factor signals, the recruitment of transducing proteinsto membrane cytoskeletal complexes as well as nuclear
responses. This cooperation has been shown for severalsignaling pathways including insulin, epidermal growth fac-tor (EGF), platelet-derived growth factor, fibroblast growthfactor, and vascular endothelial growth factor (VEGF) sig-naling.5–18 Integrins are able to trigger ligand-independentEGF receptor autophosphorylation leading to activation ofthe downstream pathway.16 Integrins, c-Src, p130Cas andEGF receptor associate in a macromolecular complex on thecell membrane and integrin-dependent adhesion inducesphosphorylation of specific tyrosine residues of EGF recep-tor, distinct from those obtained by the soluble ligandEGF.11,19
A large body of evidence demonstrates that the angiogenicprocess is initiated by mitogenic signals induced by growthfactors such as VEGF and the interplay between the tyrosine
Original received January 7, 2009; resubmission received July 31, 2009; revised resubmission received April 14, 2010; accepted April 16, 2010.From Dipartimento di Biologia Molecolare (F.G., S.P., A.S., M.R., F.N., M.O., S.O.), Universita degli Studi di Siena; Dipartimento di Pediatria (F.P.),
Ostetricia e Medicina della riproduzione, Universita degli Studi di Siena; Siena Biotech (S.G., G.L.S., G.C.T.); and Novartis Vaccines (G.M.), Siena, Italy.Correspondence to Salvatore Oliviero, Dipartimento di Biologia Molecolare, Universita degli Studi di Siena via Fiorentina 1-53100 Siena, Italy. E-mail
[email protected]© 2010 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.109.206326
1 by on May 18, 2011 circres.ahajournals.orgDownloaded from

kinase receptors and integrins expressed on endothelialcells plays a critical role in this process.20,21 VEGFstimulation of endothelial cells induces the formation of acomplex between vascular endothelial growth factor recep-tor (VEGFR)-2 and �v�3 that induces a synergistic re-sponse crucial for angiogenesis.8,21–26
VEGFR-3 (Flt-4) plays a role in vasculogenesis andangiogenesis.27–33 VEGFR-3 localizes in caveolae and acti-vates cell cycle progression, survival, and migration by theactivation of MAPK pathways.34–36
VEGFR-3, in contrast to its highly related endothelialreceptor VEGFR-2 (which, after VEGF stimulation, forms acomplex with integrin �3 subunit8,20,21,24,25,37), associatesselectively with integrin �1.10,12 Cell attachment to fibronec-tin or collagen induces the phosphorylation of VEGFR-3 inthe absence of a ligand and significantly enhances thephosphorylation of the receptor induced by its ligand.10,12
However, the mechanism of integrin activation of VEGFR-3has not been clarified.
Here, we demonstrate that collagen I–induced activationof VEGFR-3 is independent from the intrinsic catalyticactivity of the receptor but it is a direct target of c-Src.c-Src phosphorylates VEGFR-3 at specific tyrosine resi-dues with a pattern of phosphorylation that is distinct fromthe pattern induced by the ligand. Cell adhesion inducesthe phosphorylation of the tyrosine residues 1063 and1337, which are known binding sites for CRKI/II and SHC.Pull-down assays demonstrated that integrin-mediated re-ceptor phosphorylation induces the recruitment of CRKI/IIand SHC second messengers to the receptor suggestingthat the integrin/receptor complex can activate growth andsurvival signaling in the absence of activation of theintrinsic receptor kinase by its cognate ligands.
Methods
Cell Lines, Cultures, and AssaysDetails of primary endothelial cells, cell lines, growing conditions,inductions with extracellular matrix (ECM) proteins or VEGF-D,
Figure 1. VEGFR-3 phosphorylation by cell adhesion to the ECM does not require the receptor kinase activity. A, HUVECs wereplated on type I collagen (4 �g/cm2), and VEGFR-3 phosphorylation was analyzed at the times indicated. Total cell extracts were immunopre-cipitated with anti–P-tyrosine and revealed with anti–VEGFR-3. B, HUVECs were plated on type I collagen at increasing concentration for 15minutes and VEGFR-3 phosphorylation was analyzed as described previously. C, HEK 293 cells transfected with pcDNA3 vectors expressingVEGFR-3 wild-type (WT) or VEGFR-3 kinase dead mutant (KD) were treated for 30 minutes with conditioned media from 293-FT expressingVEGF-D. D, HEK 293 cells transfected with pcDNA3 vectors expressing VEGFR-3 WT and KD were plated on collagen I for 15 minutes. Totalcell extracts were immunoprecipitated with anti–P-tyrosine and revealed with anti–VEGFR-3. E, HUVECs transduced with PINCO retroviralvector expressing the chimeric receptor EGFR-VEGFR3 (E-R3) WT or KD as indicated were treated with 1 ng/mL EGF for 10 minutes. Immu-noprecipitates were performed with anti–VEGFR-3 antibody and detected with anti–P-tyrosine or anti–VEGFR-3 antibodies. F, HUVECs trans-duced with chimeric receptor E-R3 WT or KD as indicated were plated on collagen I for 15 minutes. Cell lysates were immunoprecipitatedwith anti–P-tyrosine antibody and revealed with anti–VEGFR-3. G, HUVECs transduced with chimeric receptor E-R3 WT were pretreated for30 minutes with 5 �mol/L MAZ 51 before treatment with 1 ng/mL EGF for 10 minutes. Total cell extracts were immunoprecipitated with anti–VEGFR-3 antibody and detected with anti–P-tyrosine and anti–VEGFR-3 antibodies. H, HUVECs transduced with chimeric receptor E-R3 WTwere pretreated for 30 minutes with 5 �mol/L MAZ51 before plating for 15 minutes on collagen I. Immunoprecipitates were performed withanti–P-tyrosine and revealed with anti–VEGFR-3 antibody.
Non-standard Abbreviations and Acronyms
ECM extracellular matrix
EGF epidermal growth factor
GFP green fluorescent protein
GST glutathione S-transferase
HLMVEC human lymphatic microvascular endothelial cell
HUVEC human umbilical vein endothelial cell
JNK Jun N-terminal kinase
KD kinase dead mutant
PTB phosphotyrosine binding
SH2 Src homology domain 2
shRNA small hairpin RNA
VEGF vascular endothelial growth factor
VEGFR vascular endothelial growth factor receptor
WT wild type
2 Circulation Research June 25, 2010
by on May 18, 2011 circres.ahajournals.orgDownloaded from

[3H]thymidine incorporation, and TUNEL and migration assays areprovided in the Online Data Supplement at http://circres.ahajournals.org.
DNA Construct, Antibodies, andRecombinant ProteinsConstructs expressing human VEGFR-3, E-R3 chimeric recep-tors, GST-SH2 of CRK, and GST-PTB of Shc were previouslydescribed.35 Small hairpin (sh)RNA expressing lentiviral vectorswere described.38 Details of immunoblotting, immunoprecipita-tion procedures, and antibodies are provided in the Online DataSupplement.
c-Src kinase assay, mass spectrometry analysis and GST pull-down assay are described in the Online Data Supplement.
ResultsIntegrin-Dependent VEGFR-3 PhosphorylationDoes Not Require the Intrinsic Kinase Activityof VEGFR-3It has been recently demonstrated that VEGFR-3 is highlyexpressed in angiogenic sprouts and specific inhibition ofVEGFR-3 in vascular endothelial cells inhibits angiogenesisin vivo.31 Because the interplay between ECM and endothe-lial cells plays a key role in angiogenesis, we here analyzedVEGFR-3 activation by type I collagen in human umbilicalvein endothelial cells (HUVECs). We observed that platingHUVECs on type I collagen induces a transient VEGFR-3phosphorylation with peak at 15 minutes (Figure 1A and 1B;Online Figure I). To verify whether the receptor phosphory-lation by cell adhesion is independent from the intrinsickinase activity of the receptor, we generated a kinase dead
VEGFR-3 receptor by site directed mutagenesis of tyrosine1068 (VGFR-3-Y1068F) as this residue substitution inacti-vates the receptor kinase.35 The wild-type and the VEGFR-3kinase dead receptor were transiently transfected in HEK 293cells as these cells express integrin �1 (Online Figure II).Analysis of the receptor phosphorylation revealed, as ex-pected, phosphorylation of the wild-type receptor followingcell treatment with VEGF-D, whereas VEGFR-3-Y1068Fmutant resulted inactive (Figure 1C and Online Figure I). Incontrast, both, the wild-type as well as the kinase inactivereceptor, were phosphorylated by plating cells on type Icollagen (Figure 1D and Online Figure I). To analyze the celladhesion–dependent receptor phosphorylation in endothelialcells we used a chimeric receptor containing the extracellulardomain of the EGF receptor fused to the transmembrane andcytoplasmic domain of VEGFR-3, either wild-type (E-R3) ormutant (E-R3KD), carrying the Y1068F substitution. Inserum-free starved HUVEC treatment with EGF ligand failedto activate E-R3KD (Figure 1E). Conversely, adhesion ofHUVECs on collagen I–coated dishes resulted in an evidentphosphorylation of both E-R3 wild-type and the kinase deadreceptor (Figure 1F).
Furthermore, cell adhesion in the presence of MAZ 51,a drug that specifically inhibits VEGFR-3 kinase activity,inhibited VEGFR-3 autophosphorylation in response togrowth factor activation, whereas it had no effect onadhesion-dependent receptor phosphorylation (Figure 1Gand 1H).
Taken together these results demonstrate that cell adhesioninduces VEGFR-3 phosphorylation independently from thereceptor kinase activity.
Figure 2. Integrin �1 and c-Src form a com-plex with VEGFR-3. A, HUVECs were plated ontype I collagen or gelatin, and total cell extractswere immunoprecipitated with anti–integrin �1antibodies as indicated and revealed with theanti–VEGFR-3 and anti–integrin �1 N20. B, Totalcell extracts of HUVECs plated on type I colla-gen were immunoprecipitated with anti–VEGFR-3 and revealed with anti–integrin �1. C,Immunoprecipitation of total HUVEC extractsplated on fibronectin with anti–integrin �3 andrevealed with anti–VEGFR-2, VEGFR-3, and anti–integrin �3 as indicated. D, Integrin �1 coimmu-noprecipitated with VEGFR-3 and c-Src in cellextracts from HEK 293 plated on type I collagentransiently transfected with a pcDNA3 vectorexpressing VEGFR-3 either wild-type (WT) orKD. Immunoprecipitates were performed withanti–�1-integrin and detected by Western blotwith anti–VEGFR-3, anti– c-Src, and anti–�1-integrin. E, Integrin �1 coimmunoprecipitatedwith E-R3 and c-Src in cell extracts fromHUVECs plated on type I collagen transducedwith chimeric receptor E-R3 either WT and KD.Cells extracts were immunoprecipitated withanti–�1-integrin and detected with anti–VEGFR-3, anti– c-Src, and anti–�1-integrin anti-bodies. F, In vitro kinase assay using recombi-nant c-Src to phosphorylate E-R3 KDimmunoprecipitated from total cell lysates ofHUVECs transduced with E-R3 KD. Sp3 wasused as a control of c-Src specific phosphoryla-
tion. Immunoprecipitates were performed with anti–VEGFR-3 antibody, subjected to kinase assay with active c-Src and detectedwith anti–P-tyrosine and anti–VEGFR-3 antibodies.
Galvagni et al Endothelial Cell Adhesion to the ECM 3
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Integrin-Mediated Adhesion Triggers c-Src toPhosphorylate VEGFR-3The above results establish that collagen I–mediatedVEGFR-3 phosphorylation is dissociated from the intrinsic
catalytic activity of the receptor, suggesting that a specificactivating complex is induced by integrin engagement toECM. As shown in Figure 2A, VEGFR-3 was alwaysassociated with integrin �1 because on type I collagen, it
Figure 3. Cell adhesion induces c-Src to phos-phorylate VEGFR-3. A, Analysis of VEGFR-3phosphorylation in HUVECs pretreated with1 �mol/L PP2 as indicated before induction withconditioned media containing VEGF-D for 30 min-utes. Total cell extracts were immunoprecipitatedwith anti–P-tyrosine antibody and detected withanti–VEGFR-3 antibody. B, Analysis of E-R3 chi-mera phosphorylation in HUVECs transduced withE-R3 wild-type (WT) pretreated with 1 �mol/L PP2as indicated before treatment with 1 ng/mL EGFfor 10 minutes. Cell extracts were immunoprecipi-tated with anti–VEGFR-3 antibody and detectedwith anti–P-tyrosine and anti–VEGFR-3 antibodies.C, Analysis of VEGFR-3 as in A of HUVECs platedon type I collagen for 15 minutes. Total cell lysateswere immunoprecipitated with anti–P-tyrosine anti-body. Immunoprecipitates and inputs wererevealed with anti–VEGFR-3 antibody. D, Analysisof E-R3 phosphorylation as in B of HUVECs trans-duced with E-R3 WT plated on type I collagen for15 minutes. Total cell lysates were immunoprecipi-tated with anti–P-tyrosine antibody and revealedwith anti–VEGFR-3 antibody. E, Western blot anal-ysis of VEGFR-3 and integrin �1 expression inHUVECs and in HLMVEC. F, Analysis of VEGFR-3as in A of HLMVEC plated on type I collagen for15 minutes. Total cell lysates were immunoprecipi-tated with anti–P-tyrosine antibody. Immunopre-cipitates and inputs were revealed with anti–VEGFR-3 antibody. G, Analysis of E-R3 WTphosphorylation transduced in WT fibroblasts(SYF�/�) and fibroblasts triple knockout for c-Src,Yes, and Fyn (SYF�/�)44 plated on type I collagenfor 15 minutes. c-Src rescue by transfection ofSYF�/� previously transduced with E-R3 WT. Afteradhesion, cells were collected and lysates wereimmunoprecipitated with anti–P-tyrosine anddetected with anti–VEGFR-3. Endogenous andtransfected c-Src were detected in whole cellextracts with anti–c-Src antibody. The chimerawas phosphorylated in adhesion dependent-manner on the reintroduction of c-Src in SYF cells.
Table. Phosphotyrosine-Containing Peptides Identified by LC-MS/MS Analysis
Peptide Sequence pYsite SEQUEST XCorr Observed m/z z Delta Mass (ppm)
TGYLSIIMDPGEVPLEEQCEpYLSYDASQWEFPR 830 5.913 1334.92432 3 2.4
TGYLSIIMDPGEVPLEEQCEYLSpYDASQWEFPR 833 5.642 1334.92432 3 2.4
VLGpYGAFGK 853 2.753 496.23642 2 0.7
DIpYKDPDFVR 1063 3.145 674.30225 2 �0.6
GGQVFpYNSEYGELSEPSEEDHCSPSAR 1333 5.888 1037.75122 3 2.6
GGQVFYNSEpYGELSEPSEEDHCSPSAR 1337 7.285 1037.75122 3 2.6
GGQVFpYNSEpYGELSEPSEEDHCSPSAR 1333–1337 7.052 1064.40552 3 1.5
Peptide sequences are indicated along with the position of pYsites, the indicated peptide scores, observed mass-to-charge ratios(m/z), charge state (z), and mass error. The isobaric phosphopeptides GGQVFpYNSEYGELSEPSEEDHCSPSAR and GGQVFYNSEpY-GELSEPSEEDHCSPSAR, which differ only for the position of the phosphoryl moiety, are chromatographically indistinguishable, andhence the resulting MS/MS spectrum is the combination of both contributions. The same can be said for the couple of isomericphosphopeptides TGYLSIIMDPGEVPLEEQCEpYLSYDASQWEFPR and TGYLSIIMDPGEVPLEEQCEYLSpYDASQWEFPR.
4 Circulation Research June 25, 2010
by on May 18, 2011 circres.ahajournals.orgDownloaded from

coimmunoprecipitated with both an antibody that immuno-precipitates total integrin �1 (N20) and an antibody thatspecifically detects the integrin �1 in its active conformation(9EG7),39,40 whereas we could not detect VEGFR-3 associa-tion with integrin �1 on gelatin. In contrast, integrin �3 didnot associate with VEGFR-3, whereas it associates withVEGFR-2 (Figure 2C).
As it was previously demonstrated that integrin �1 coim-munoprecipitates with c-Src,41,42 we investigate whetherVEGFR-3 could be part of a complex with c-Src and integrin�1. As shown in Figure 2D, c-Src and VEGFR-3 coimmu-noprecipitated with integrin �1 suggesting that a VEGFR-3/Src/integrin�1 complex is present on the cell surface. Similarresults were obtained with the chimeric receptor (Figure 2E).
To demonstrate a direct phosphorylation of VEGFR-3 byc-Src we performed by an in vitro kinase. E-R3 was
detergent-extracted from transduced HUVECs. The lysateswere immunoprecipitated with an anti–VEGFR-3 and anti-Sp3 as a control and immunoprecipitates were subjected to akinase assay in the presence of recombinant active Src Y527Fand ATP. As shown in Figure 2F, kinase dead receptor aredirect substrates for Src.
To demonstrate the functional role of Src in VEGFR-3phosphorylation we performed adhesion assays with cellsexposed to PP2, a c-Src kinase inhibitor highly specific ifused at low concentration. Cell treatment with PP2 at a finalconcentration of 1 �mol/L did not alter VEGFR-3 autophos-phorylation by VEGF-D treatment or E-R3 autophosphory-lation by cell treatment with EGF (Figure 3A and 3B). Incontrast, 1 �mol/L PP2 significantly reduced both VEGFR-3and E-R3 phosphorylation induced by plating the cells oncollagen I–coated dishes (Figure 3C and 3D; Online Figure
Figure 4. Mapping c-Src phosphorylation sites of VEGFR-3.A through G, Representative spectra of the phosphopeptidesderived from the cytoplasmic domain of VEGFR-3 mutantkinase dead (KD) demonstrating tyrosine phosphorylation atpositions 830, 833, 853, 1063, 1333, and 1337. The b- andy-ion series are indicated. The sequence of the peptide isshown at the top of each panel. Additional information is listedin the Table.
Galvagni et al Endothelial Cell Adhesion to the ECM 5
by on May 18, 2011 circres.ahajournals.orgDownloaded from

I). As in adults VEGFR-3 is constitutively expressed inlymphatic vessels, we performed the adhesion experimentalso in human lymphatic microvascular endothelial cells(HLMVECs), which express high levels of VEGFR-3 (Figure3E).43 As shown in Figure 3F, plating HLMVEC on type Icollagen induces VEGFR-3 phosphorylation and cell treat-ment with PP2 significantly reduced VEGFR-3 phosphoryla-tion demonstrating that the crosstalk between VEGFR-3 andintegrin �1 is a general event.
To further verify the role of c-Src in VEGFR-3 phosphor-ylation we performed collagen induction in fibroblasts wild-type (SYF�/�) and “knocked out” for c-Src, Yes, Fyn(SYF�/�)44 which express integrin �1 (Online Figure II). InSYF�/� VEGFR-3 receptor was not phosphorylated on celladhesion to type I collagen and its phosphorylation wasrestored by cotransfection of VEGFR-3 with c-Src (Figure3G). Taken together, these results demonstrate that c-Srcphosphorylates VEGFR-3 following cell adhesion on type Icollagen.
Mapping Src-Dependent Phosphorylation Siteson VEGFR-3To identify direct Src-target residues we performed in vitrophosphorylation by incubating the recombinant C-terminaldomain of the kinase dead VEGFR-3 with active c-Src andsearched for tyrosine-phosphorylated residues by massspectrometric analysis of peptides after trypsin digestion.The phosphorylation sites 830 and 833 contained in thepeptide 810 to 842, (m/z 1334.92432) were both detectedin the phosphorylated form (Table and Figure 4A and 4B).The phosphorylation site 853 contained in the peptide 850to 858 (m/z 496.23642) was detected in the phosphorylatedform (Table and Figure 4C). The phosphorylation site 1063contained in the peptide 1061 to 1070 (m/z 674.30225) wasdetected in the phosphorylated form (Table and Figure4D). The phosphorylation sites 1333 and 1337 contained inthe peptide 1328 to 1354 (m/z 1037.75122) were bothdetected in the phosphorylated form (Table and Figure 4Ethrough 4G).
These results show that c-Src-dependent receptor phos-phorylation induces a phosphorylation pattern that is distinctfrom the phosphorylation pattern induced by the treatmentwith the growth factor.35,45
Collagen I–Dependent VEGFR-3 PhosphorylationCan Mediate Downstream SignalingThe above results demonstrated that c-Src phosphorylatesVEGFR-3 C-terminal domain at several tyrosine residuesincluding 1063 and 1337 previously known to recruitCRKI/II and SHC on receptor phosphorylation.35,46 Incu-bation of the in vitro phosphorylated receptor with thefusion adaptor proteins GST-SH2-CRK or GST-PTB-SHCsuggests that c-Src phosphorylation of VEGFR-3 at thesetyrosine residues may induce the recruitment of CRKI/IIand SHC to the receptor (Figure 5A). Pull-down experi-ments in vivo demonstrated that after adhesion to collagenthe chimeric E-R3 KD was able to recruit both CRKI/II
and SHC adaptor proteins (Figure 5B and Online Figure I).In contrast, the wild-type receptor, but not the kinase-inactive mutant, could bind these adaptor proteins in aligand-dependent manner. These experiments indicate thatthe tyrosine residues 1063 and 1337, phosphorylated oncell adhesion to the ECM, independently from activationof the receptor kinase activity, can recruit CRKI/II andSHC adaptor proteins to the receptor and suggest thatVEGFR-3 phosphorylation by c-Src contributes to activatereceptor downstream signaling.
The functional significance of VEGFR-3 transactivation bycell adhesion was analyzed by in vitro sprouting thus theformation of tube-like structures resembling early capillariesin a 3D type I collagen matrix. HUVECs transduced witheither the wild-type or the kinase dead receptor showed asignificant increase of the sprouting activity on the collagenmatrix, whereas the Y1063F or the Y1337F mutant receptorsdid not show an increased sprouting with respect to the mockinfected cells (Figure 6B and 6C).
Analysis of downstream second messengers revealed anincrease of JNK phosphorylation on cell induction by type I
Figure 5. c-Src phosphorylates E-R3 KD at specific andfunctional tyrosines. A, Pull-down assay of E-R3 KD and Sp3immunoprecipitated from HUVECs transduced with E-R3 KD,subjected to in vitro c-Src phosphorylation and incubated withthe fusion proteins GST-SH2-CRK and GST-PTB-SHC. Immuno-precipitates were analyzed by Western blot with anti–P-tyrosineand anti–VEGFR-3 antibodies and the pull-down revealed withanti-GST antibody. B, Pull-down assay with recombinant fusionproteins GST-SH2-CRK and GST-PTB-SHC with HUVECextracts transduced with E-R3 wild-type (WT) or KD eithertreated with EGF or plated on collagen I. Lanes 1 to 4 pull-downfrom 1�105 cells, lanes 5 to 8 from 1�106 cells to obtain com-parable phosphorylation signals. Cell extracts were incubatedwith GST fusion proteins and revealed with anti–VEGFR-3antibody.
6 Circulation Research June 25, 2010
by on May 18, 2011 circres.ahajournals.orgDownloaded from

collagen both with wild-type and kinase dead receptor,whereas JNK phosphorylation was impaired with eitherY1063F or Y1337F mutant (Figure 6D and 6E).
Furthermore, to evaluate which angiogenic functions weremediated by VEGFR-3 induced by type I collagen weperformed cell proliferation, adhesion, migration and apopto-sis tests in control (shGFP) and in VERGFR-3 (shR3-D andshR3-F) silenced HUVECs (Figure 7). VEGFR-3 silencingsignificantly affected endothelial cell migration on type Icollagen, which is a ligand for the integrin �1, whereas it didnot affect HUVEC migration on vitronectin, which is a ligandfor integrin �3 (Figure 7B). VEGFR-3 silencing also inducedincrease of apoptosis only on type I collagen (Figure 7C),whereas it did not affect cell proliferation measured by DNAsynthesis and cell adhesion (Figure 7D and 7E).
DiscussionMacromolecular complexes of integrins and growth factorreceptors have been documented to cooperate for the optimalresponse to cognate growth factors and the propagation of
downstream signaling. This cooperation has been demon-strated to be crucial for the angiogenic process.20,21
We now observed that adhesion-dependent phosphory-lation of the receptor VEGFR-3 triggered by the formationof the complex between VEGFR-3 and integrin �1 inducesthe phosphorylation of the receptor independently from theintrinsic kinase activity of the receptor. These results showa remarkable difference from the behavior of VEGFR-2which is positively regulated by plating cells on vitronec-tin, whereas type I collagen inhibits VEGFR-2 activationby TCPTP-dependent receptor dephosphorylation.26,47,48
It has been recently shown that VEGFR-3 is specificallyexpressed in tip cells during sprouting angiogenesis whereit plays a role in sprouting, vascular density, and vesselbranching.31,49 Our results suggest that during the angio-genic process the interactions of sprouting endothelial cellswith the ECM, modulating VEGFR-3 signaling, contributeto determine the correct shape of the vascular network.
The activation of VEGFR-3 by the formation of thereceptor integrin complex differs from EGF receptor be-cause in the latter case, integrin-dependent adhesion trig-
Figure 6. Wild-type VEGFR-3 but not VEGFR-3 mutants inducessprouting and JNK activation on type I collagen. A, Western blot oftotal cell extract from HUVECs transduced with the empty retroviralvector or with retroviruses for the expression of the chimeric receptorE-R3 wild-type (wt), kinase dead (KD), with the amino acid substitutionY1063F, Y1337F revealed with anti–VEGFR-3 antibody. B, Spheroids oftransduced cells were embedded in collagen matrix without specificgrowth factors and analyzed 24 hours later. C, Quantitative analysis of 3independent sprouting experiments measured by digitized image analy-sis of spheroids. The results are expressed as the means�SD. *P�0.05.D, Transduced HUVECs were starved for 16 hours in M199 mediumcontaining 0.25 FCS% and 0.25% BSA and subsequently induced with8 �g/mL soluble type I collagen. Total cell extracts were analyzed bySDS-PAGE and Western blotting with the antibodies recognizingphospho-JNK and total JNK. E, Western blot bands intensities from 3different experiments were quantified and plotted as phospho-JNK/JNKratio. The results are expressed as the means�SD of 3 independentexperiments. *P�0.05.
Galvagni et al Endothelial Cell Adhesion to the ECM 7
by on May 18, 2011 circres.ahajournals.orgDownloaded from

gers the receptor autophosphorylation,16 whereas we ob-served VEGFR-3 phosphorylation independently ofintrinsic receptor kinase activity and mediated by c-Srcfollowing adhesion. In fact, we observed that VEGFR-3 isa direct target of c-Src that phosphorylates the C-terminaldomain of VEGFR-3 at several residues including thosecritical for the activation of downstream signaling. It waspreviously shown that c-Src plays a role also in thesignaling of VEGFR-2. In this case, on VEGF treatment,c-Src regulates the formation of the complex betweenVEGFR-2 and �v�3 by phosphorylating the cytoplasmictyrosine residues of integrin �3 inducing structuralchanges on the complex that promote capillary formationand chemotaxis.24,25
Because cell adhesion– dependent receptor phosphoryla-tion could be dissociated from the receptor kinase activity,we were able to identify the tyrosine residues specificallydependent from the integrin/Src-mediated activation bymass spectrometric analysis. This analysis revealed 6tyrosine residues on VEGFR-3 C-terminal domain, namelytyrosine 830, 833, 853, 1063, 1333, and 1337 demonstrat-ing that integrin-mediated receptor phosphorylation in-duces a phosphorylation pattern that is distinct from thatinduced by growth factors.
Functional assays in 3D collagen matrix suggest thatadhesion per se could activate a signaling pathway usingVEGFR-3 as a scaffold module to activate a downstreamsignal. c-Src-dependent VEGFR-3 phosphorylation includestyrosine residues 1063 and 1337, which are docking sites forCRKI/II and SHC, respectively. Our experiments demon-strate that on cell adhesion, and in absence of the intrinsickinase activity of the receptor, these 2 sites do recruit CRKI/IIand SHC adaptor proteins suggesting that cell adhesioncontributes to the survival and migration of endothelial cellsin agreement with our previous observations.35 Thus, the
ability of integrins to transactivate VEGFR-3 suggests thatactivation of growth factor receptors in the absence of theirspecific ligands can be a mechanism of adhesion-mediatedsignaling. This could play a role during the sprouting processof angiogenesis because endothelial cells, expressingVEGFR-3, get in contact with the ECM and the engagementof integrin in cell matrix interactions may activate a cellresponse even in the absence or at low doses of the cognategrowth factor. Finally, the integrin-mediated VEGFR-3 acti-vation may have implications in medicine because weshowed that blocking the receptor with a specific drug is notsufficient to switch off the signaling derived from thereceptor.
AcknowledgmentsWe thank all members Novartis vaccines research team for supportand discussions and Andrea Graziani, Federico Bussolino, GuidoSerini, and Guido Tarone for helpful suggestions.
Sources of FundingThis study was supported by grants from Associazione Italianaricerca sul Cancro (AIRC), Ministero Italiano dell’Istruzione,dell’Unversita e della Ricerca (MIUR), Istituto Toscano Tumori(ITT).
DisclosuresNone.
References1. Somanath PR, Ciocea A, Byzova TV. Integrin and growth factor receptor
alliance in angiogenesis. Cell Biochem Biophys. 2009;53:53–64.2. Serini G, Napione L, Arese M, Bussolino F. Besides adhesion: new
perspectives of integrin functions in angiogenesis. Cardiovasc Res. 2008;78:213–222.
Figure 7. VEGFR-3 silencing affects en-dothelial cell migration and survival ontype I collagen. A, Western blot analysisof HUVEC whole cell extracts transducedwith a lentiviral vector for the expressionof a control shRNA (GFP) or 2 indepen-dent shRNA specific for VEGFR-3 (R3-D)and (R3-F). B, Quantitative analysis of 3independent migration experiments inBoyden chamber of HUVECs expressingshRNA as indicated. The results areexpressed as the means�SD. *P�0.05.C, Quantitative analysis of apoptosismeasured by TUNEL assay from 3 inde-pendent experiments expressed as per-centage over total nuclei stained withDAPI of HUVECs expressing shRNA asindicated. The results are expressed asthe means�SD. *P�0.05. D, Quantitativeanalysis of [3H] incorporation of HUVECsexpressing shRNA as indicated. Theresults are expressed as the means�SD.E, Quantitative analysis of cell attachmenton collagen (C), gelatin (G), or vitronectin(V) of HUVECs expressing shRNA as indi-cated. The results are expressed as themeans�SD.
8 Circulation Research June 25, 2010
by on May 18, 2011 circres.ahajournals.orgDownloaded from

3. Schwartz MA, Ginsberg MH. Networks and crosstalk: integrin signallingspreads. Nat Cell Biol. 2002;4:E65–E68.
4. Giancotti FG, Tarone G. Positional control of cell fate through jointintegrin/receptor protein kinase signaling. Annu Rev Cell Dev Biol. 2003;19:173–206.
5. Sundberg C, Rubin K. Stimulation of beta1 integrins on fibroblastsinduces PDGF independent tyrosine phosphorylation of PDGF beta-receptors. J Cell Biol. 1996;132:741–752.
6. Miyamoto S, Teramoto H, Gutkind JS, Yamada KM. Integrins can col-laborate with growth factors for phosphorylation of receptor tyrosinekinases and MAP kinase activation: roles of integrin aggregation andoccupancy of receptors. J Cell Biol. 1996;135:1633–1642.
7. Short SM, Talbott GA, Juliano RL. Integrin-mediated signaling events inhuman endothelial cells. Mol Biol Cell. 1998;9:1969–1980.
8. Soldi R, Mitola S, Strasly S, Defilippi P, Tarone G, Bussolino F. Role ofavb3 integrin in the activation of vascular endothelial growth factorreceptor-2. EMBO J. 1999;18:882–892.
9. Kuwada SK, Li X. Integrin alpha5/beta1 mediates fibronectin-dependentepithelial cell proliferation through epidermal growth factor receptoractivation. Mol Biol Cell. 2000;11:2485–2496.
10. Wang JF, Zhang XF, Groopman JE. Stimulation of beta 1 integrin inducestyrosine phosphorylation of vascular endothelial growth factor receptor-3and modulates cell migration. J Biol Chem. 2001;276:41950–41957.
11. Moro L, Dolce L, Cabodi S, Bergatto E, Erba EB, Smeriglio M, Turco E,Retta SF, Giuffrida MG, Venturino M, Godovac-Zimmermann J, Conti A,Schaefer E, Beguinot L, Tacchetti C, Gaggini P, Silengo L, Tarone G,Defilippi P. Integrin-induced epidermal growth factor (EGF) receptoractivation requires c-Src and p130Cas and leads to phosphorylation ofspecific EGF receptor tyrosines. J Biol Chem. 2002;277:9405–9414.
12. Zhang X, Groopman JE, Wang JF. Extracellular matrix regulates endo-thelial functions through interaction of VEGFR-3 and integrinalpha5beta1. J Cell Physiol. 2005;202:205–214.
13. Schneller M, Vuori K, Ruoslahti E. Alphavbeta3 integrin associates withactivated insulin and PDGFbeta receptors and potentiates the biologicalactivity of PDGF. EMBO J. 1997;16:5600–5607.
14. Clemmons DR, Maile LA. Interaction between insulin-like growth factor-Ireceptor and alphaVbeta3 integrin linked signaling pathways: cellularresponses to changes in multiple signaling inputs. Mol Endocrinol. 2005;19:1–11.
15. Del Pozo MA, Kiosses WB, Alderson NB, Meller N, Hahn KM, SchwartzMA. Integrins regulate GTP-Rac localized effector interactions throughdissociation of Rho-GDI. Nat Cell Biol. 2002;4:232–239.
16. Cabodi S, Moro L, Bergatto E, Boeri Erba E, Di Stefano P, Turco E,Tarone G, Defilippi P. Integrin regulation of epidermal growth factor(EGF) receptor and of EGF-dependent responses. Biochem Soc Trans.2004;32:438–442.
17. Kaneko Y, Kitazato K, Basaki Y. Integrin-linked kinase regulatesvascular morphogenesis induced by vascular endothelial growth factor.J Cell Sci. 2004;117:407–415.
18. Aplin AE, Stewart SA, Assoian RK, Juliano RL. Integrin-mediatedadhesion regulates ERK nuclear translocation and phosphorylation ofElk-1. J Cell Biol. 2001;153:273–282.
19. Boeri Erba E, Bergatto E, Cabodi S, Silengo L, Tarone G, Defilippi P,Jensen ON. Systematic analysis of the epidermal growth factor receptorby mass spectrometry reveals stimulation-dependent multisite phosphor-ylation. Mol Cell Proteomics. 2005;4:1107–1121.
20. Somanath PR, Ciocea A, Byzova TV. Integrin and growth factor receptoralliance in angiogenesis. Cell Biochem Biophys. 2009;53:53–64.
21. Serini G, Napione L, Bussolino F. Integrins team up with tyrosine kinasereceptors and plexins to control angiogenesis. Curr Opin Hematol. 2008;15:235–242.
22. Borges E, Jan Y, Ruoslahti E. Platelet-derived growth factor receptor betaand vascular endothelial growth factor receptor 2 bind to the beta 3integrin through its extracellular domain. J Biol Chem. 2000;275:39867–39873.
23. Masson-Gadais B, Houle F, Laferriere J, Huot J. Integrin alphavbeta3,requirement for VEGFR2-mediated activation of SAPK2/p38 and forHsp90-dependent phosphorylation of focal adhesion kinase in endothelialcells activated by VEGF. Cell Stress Chaperones. 2003;8:37–52.
24. Mahabeleshwar GH, Feng W, Reddy K, Plow EF, Byzova TV. Mech-anisms of integrin-vascular endothelial growth factor receptor cross-activation in angiogenesis. Circ Res. 2007;101:570–580.
25. Mahabeleshwar GH, Chen J, Feng W, Somanath PR, Razorenova OV,Byzova TV. Integrin affinity modulation in angiogenesis. Cell Cycle.2008;7:335–347.
26. Somanath PR, Malinin NL, Byzova TV. Cooperation between integrinalphavbeta3 and VEGFR2 in angiogenesis. Angiogenesis. 2009;12:177–185.
27. Carmeliet P. Angiogenesis in health and disease. Nat Med. 2003;9:653–660.
28. Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and itsreceptors. Nat Med. 2003;9:669–676.
29. Dumont DJ, Jussila L, Taipale J, Lymboussaki A, Mustonen T, PajusolaK, Breitman M, Alitalo K. Cardiovascular failure in mouse embryosdeficient in VEGF receptor-3. Science. 1998;282:946–949.
30. Kubo H, Fujiwara T, Jussila L, Hashi H, Ogawa M, Shimizu K, AwaneM, Sakai Y, Takabayashi A, Alitalo K, Yamaoka Y, Nishikawa SI.Involvement of vascular endothelial growth factor receptor-3 in mainte-nance of integrity of endothelial cell lining during tumor angiogenesis.Blood. 2000;96:546–553.
31. Tammela T, Zarkada G, Wallgard E, Murtomaki A, Suchting S, Wir-zenius M, Waltari M, Hellstrom M, Schomber T, Peltonen R, Freitas C,Duarte A, Isoniemi H, Laakkonen P, Christofori G, Yla-Herttuala S,Shibuya M, Pytowski B, Eichmann A, Betsholtz C, Alitalo K. BlockingVEGFR-3 suppresses angiogenic sprouting and vascular network for-mation. Nature. 2008;454:656–660.
32. Laakkonen P, Waltari M, Holopainen T, Takahashi T, Pytowski B,Steiner P, Hicklin D, Persaud K, Tonra JR, Witte L, Alitalo K. Vascularendothelial growth factor receptor 3 is involved in tumor angiogenesisand growth. Cancer Res. 2007;67:593–599.
33. Matsumura K, Hirashima M, Ogawa M, Kubo H, Hisatsune H, Kondo N,Nishikawa S, Chiba T. Modulation of VEGFR-2-mediated endotheli-al-cell activity by VEGF-C/VEGFR-3. Blood. 2003;101:1367–1374.
34. Marconcini L, Marchio S, Morbidelli L, Cartocci E, Albini A, Ziche M,Bussolino F, Oliviero S. c-fos-induced growth factor/vascular endothelialgrowth factor-D induces angiogenesis in vivo and in vitro. Proc Natl AcadSci U S A. 1999;96:9671–9676.
35. Salameh A, Galvagni F, Bardelli M, Bussolino F, Oliviero S. Directrecruitment of CRK and GRB2 to VEGFR-3 induces proliferation,migration, and survival of endothelial cells through the activation ofERK, AKT, and JNK pathways. Blood. 2005;106:3423–3431.
36. Galvagni F, Anselmi F, Salameh A, Orlandini M, Rocchigiani M,Oliviero S. Vascular endothelial growth factor receptor-3 activity ismodulated by its association with caveolin-1 on endothelial membrane.Biochemistry. 2007;46:3998–4005.
37. Mahabeleshwar GH, Feng W, Phillips DR, Byzova TV. Integrin signalingis critical for pathological angiogenesis. J Exp Med. 2006;203:2495–2507.
38. Orlandini M, Spreafico A, Bardelli M, Rocchigiani M, Salameh A,Nucciotti S, Capperucci C, Frediani B, Oliviero S. Vascular endothelialgrowth factor-D activates VEGFR-3 expressed in osteoblasts inducingtheir differentiation. J Biol Chem. 2006;281:17961–17967.
39. Lenter M, Uhlig H, Hamann A, Jeno P, Imhof B, Vestweber D. Amonoclonal antibody against an activation epitope on mouse integrinchain beta 1 blocks adhesion of lymphocytes to the endothelial integrinalpha 6 beta 1. Proc Natl Acad Sci U S A. 1993;90:9051–9055.
40. Bazzoni G, Shih DT, Buck CA, Hemler ME. Monoclonal antibody 9EG7defines a novel beta 1 integrin epitope induced by soluble ligand andmanganese, but inhibited by calcium. J Biol Chem. 1995;270:25570–25577.
41. Moro L, Venturino M, Bozzo C, Silengo L, Altruda F, Beguinot L,Tarone G, Defilippi P. Integrins induce activation of EGF receptor: rolein MAP kinase induction and adhesion-dependent cell survival. EMBO J.1998;17:6622–6632.
42. Kim E, Lee S, Mian MF, Yun SU, Song M, Yi KS, Ryu SH, Suh PG.Crosstalk between Src and major vault protein in epidermal growthfactor-dependent cell signalling. FEBS J. 2006;273:793–804.
43. Norgall S, Papoutsi M, Rossler J, Schweigerer L, Wilting J, Weich HA.Elevated expression of VEGFR-3 in lymphatic endothelial cells fromlymphangiomas. BMC Cancer. 2007;7:105.
44. Klinghoffer RA, Sachsenmaier C, Cooper JA, Soriano P. Src familykinases are required for integrin but not PDGFR signal transduction.EMBO J. 1999;18:2459–2471.
45. Dixelius J, Makinen T, Wirzenius M, Karkkainen MJ, Wernstedt C,Alitalo K, Claesson-Welsh L. Ligand-induced vascular endothelialgrowth factor receptor-3 (VEGFR-3) heterodimerization with VEGFR-2in primary lymphatic endothelial cells regulates tyrosine phosphorylationsites. J Biol Chem. 2003;278:40973–40979.
46. Fournier E, Rosnet O, Marchetto S, Turck CW, Rottapel R, Pelicci PG,Birnbaum D, Borg JP. Interaction with the phosphotyrosine bindingdomain/phosphotyrosine interacting domain of SHC is required for the
Galvagni et al Endothelial Cell Adhesion to the ECM 9
by on May 18, 2011 circres.ahajournals.orgDownloaded from

transforming activity of the FLT4/VEGFR3 receptor tyrosine kinase.J Biol Chem. 1996;271:12956–12963.
47. Mitola S, Brenchio B, Piccinini M, Tertoolen L, Zammataro L, Breier G,Rinaudo MT, den Hertog J, Arese M, Bussolino F. Type I collagen limitsVEGFR-2 signaling by a SHP2 protein-tyrosine phosphatase-dependentmechanism 1. Circ Res. 2006;98:45–54.
48. Mattila E, Auvinen K, Salmi M, Ivaska J. The protein tyrosine phos-phatase TCPTP controls VEGFR2 signalling. J Cell Sci. 2008;121:3570–3580.
49. Benedito R, Roca C, Sorensen I, Adams S, Gossler A, Fruttiger M,Adams RH. The notch ligands Dll4 and Jagged1 have opposing effects onangiogenesis. Cell. 2009;137:1124–1135.
Novelty and Significance
What Is Known?● Cell adhesion induces the phosphorylation of several receptors.● Integrins mediate receptor activation by extracellular matrix (ECM).● ECM activates the intrinsic kinase activity of the epidermal growth
factor (EGF) receptor.
What New Information Does This Article Contribute?● This article describes the mechanism of activation of the tyrosine
kinase receptor vascular endothelial growth factor receptor(VEGFR)-3 by cell adhesion.
● Cell adhesion–dependent VEGFR-3 phosphorylation does not requirethe intrinsic kinase activity of the receptor, but is mediated byc-Src.
● This study also identifies the tyrosine residues that are phosphory-lated following cell adhesion to the ECM.
Cell adhesion activates surface receptors by promoting theformation of receptor-integrin complexes. During angiogenesis,
endothelial cells expressing VEGFR-3 come in contact with theECM. In this article, we report a novel mechanism of receptoractivation. Our data show that integrin-dependent VEGFR-3phosphorylation is mediated by c-Src, which directly phosphor-ylates VEGFR-3 at critical tyrosine residues. Phosphorylation ofthese residues facilitates the recruitment of second messengers.These observations suggest that Src-mediated receptor phos-phorylation is sufficient to activate cell adhesion–induced intra-cellular signaling. Previous studies showed that, on adhesion,phosphorylation of the EGF receptor at critical residues activatesthe intrinsic kinase activity of the receptor, leading to autophos-phorylation and subsequent stimulation of downstream signal-ing. We demonstrate that blocking the receptor with a specificdrug is not sufficient to switch off the cell signaling triggered bythe receptor. Our results suggest that a therapy that uses aspecific tyrosine kinase receptor inhibitor would not be sufficientto switch off the signaling originating from the receptor.
10 Circulation Research June 25, 2010
by on May 18, 2011 circres.ahajournals.orgDownloaded from

ONLINE DATA SUPPLEMENT
Endothelial cell adhesion to the extra-cellular matrix induces c-Src-dependent VEGFR-3
phosphorylation without the activation of the receptor intrinsic kinase activity.
Federico Galvagni1, Susanna Pennacchini1, Ahmad Salameh1, Marina Rocchigiani1, Francesco
Neri1, Maurizio Orlandini1, Felice Petraglia2, Stefano Gotta3, Gian Luca Sardone3, Giacomo
Matteucci4, Georg C. Terstappen3, and Salvatore Oliviero1
MATERIALS AND METHODS
Cell lines, cultures and assays.
Human Umbilical Vein Endothelial Cells (HUVEC) were derived from umbilical cordons and were
grown on gelatin-coated dishes in M199 medium supplemented with 10% Fetal Bovine Serum
(FBS), 10 U/ml Heparin, 50 U/ml penicillin-streptomycin and 0,1 mg/ml bovine brain extract.
Human embryonic kidney HEK 293, 293 FT, HEK 293 GPG were grown in DMEM high glucose
medium supplemented with 10% Fetal Bovine Serum (FBS) and 50 U/ml penicillin-streptomycin.
Human Lymphatic Microvascular Endothelial Cells (neonatal) (HLMVEC 13517, Lonza) were
grown in EGM-2 MV (Lonza).
Mouse Fibroblasts wild-type and triple knockout for c-Src, Yes and Fyn were grown in DMEM
supplemented with 10% heat inactivated FBS and 50 U/ml penicillin-streptomycin. Transient
transfections of HEK 293 cells were performed using Polyfectamine reagent (Qiagen).
Virus preparations
A total of 7x106 293GPG cells were transfected with 24 µg of retroviral plasmid constructs by using
Lipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions. The collected
supernatant was centrifuged in a Beckman ultracentrifuge into Ultraclear tubes (Beckman, Hialeah,
FL). Infected cells were assayed by flow cytometry for the expression level of the GFP protein
expressed by the PINCO vector and the expression level of chimeric receptors on the cell surface.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

The results were confirmed by Western blot analysis using the whole-cell lysate.
DNA construct, antibodies, and recombinant proteins
Both the full-length human VEGFR-3 and E-R3 chimeric receptors wild type and mutants were
previously described 1. Small hairpin RNA (shRNA) expressing lentiviral vectors were previously
described 2.
The following antibodies were used for immunoblotting and immunoprecipitation: Anti-VEGFR-3
(sc-321), anti-P-Tyrosine (sc-7020), anti-c-Src (sc-18), anti-GST (sc-138 HRP), anti-integrin β1
N20 (sc-6622), and anti-Histidine (sc-8036) (Santa Cruz Biotecnology), anti-active-integrin β1
(9EG7, BD pharmingen), anti-integrin β3 (BV4, Abcam), anti-JNK (#9252, Cell Signaling
Technology, Beverly, MA) and anti-P-JNK (#9251S, Cell Signaling Technology, Beverly, MA).
Secondary antibodies anti-mouse IgG-HRP, anti-rabbit IgG-HRP, and anti-goat IgG-HRP were
obtained from GE Healthcare. Anti-β−actin (A5441) and human epidermal growth factor (EGF)
was purchased from Sigma.
The recombinant VEGF-D was obtained by conditioned media from 293 FT transfected with
VEGF-D expressing vector.
The recombinant GST-SH2 of CRK and GST-PTB of Shc fusion proteins used in protein
interaction assays were previously described 1.
The recombinant VEGFR-3 used for mass spectrometry analysis was obtained by subcloning the C-
Terminal domain of VEGFR-3 kinase dead into pET 15b expressing vector from aminoacis 805 to
1363 downstream to the Histidine tag (SP806). Human VEGFR-3 (805-1363) transformed E.Coli
cells (BL21-DE3) grown for 3h at 37°C after IPTG induction were pelleted by centrifugation at
6000 g for 10 min and solubilized in 6M guanidium chloride, 20 mM Tris-Hcl pH7.9, 2 mM β-
mercaptoethanol and 0.5 M NaCl. After centrifugation (23700 x g, 30 min 4°C) VEGFR-3 was
purified by IMAC (Immobilized Metal Affinity Chromatography) in denaturing conditions (7M
Urea; 20 mM Tris-Hcl pH 7.9; 2 mM β-mercaptoethanol; 10% glycerol and 0.5 M NaCl), using an
AKTA purifier (GE Healthcare). On-column refolding of immobilized VEGFR-3 was achieved by
applying a linear gradient reducing the concentration of denaturant in 200 ml column volumes and
the refolded protein was eluted in a buffer containing 20 mM Tris-Hcl pH 8; 100 mM NaCl; 2 mM
β-mercaptoethanol; 10% glycerol and 300 mM imidazole. Imidazole was removed on a HiTrap
desalting column (GE Healthcare) in 20 mM Tris-Hcl pH 8; 100 mM NaCl; 10% glycerol buffer.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

Protein phosphorylation and Immunoprecipitation
Stimulation of cell by ECM proteins was performed by loading cells onto tissue culture dishes
coated with the indicated amount of type I collagen (collagen rat tail, Roche) or with the
concentration of 4 µg/cm2 for different times as indicated at 37°C. Stimulation of cells by soluble
ligands was obtained by incubating cells with 1 ng/ml of EGF, 8 µg/ml type I collagen or
conditioned media containing VEGF-D at indicated time. When indicated cells were starved in the
corresponding medium 1% BSA for 20 hours and treated with pharmacological inhibitors 1 µM
PP2 (Calbiochem) and 5 µM MAZ 51 (Alexis Biochemicals). After stimulation, cells were washed
and solubilized with RIPA lysis buffer (50 mM TrisHCL pH 7.2, 100 mM NaCl, 1% Triton, 15
DOC, 0.1% SDS, 50 mM NaF, 2 mM Na3VO4 and antiprotease cocktail Sigma).
Immunoprecipitation was performed by using Dynabeads Protein A or G (Invitrogen) complexed
with receptor specific anti-VEGFR-3, anti-integrin β1 and anti-P-Tyrosine antibodies as indicated,
according to manufacturer’s instructions. Samples were separated by SDS-PAGE and analyzed by
Western blotting.
c-Src kinase assay
E-R3 kinase dead receptor (E-R3 KD) was immunoprecipitated from total cell extracts of infected
HUVEC with anti-VEGFR-3 antibody cooupled to Dynabeads protein A. After 3 washes beads
were incubate at 30º C for 10 min in kinase buffer (20 mM Hepes pH 7.5, 10 mM MgCl2, and 2 mM
Na3VO4) containing 100 µM ATP and 50 ng of human active c-Src (#14-326, Upstate-Millipore).
The reaction was analyzed by SDS-PAGE and Western blotting.
Mass spectrometry
Mass spectrometry identification of phosphopepetides was done according to standard protocol.
Briefly, after reduction and alkylation proteins were digested in solution with Trypsin Gold
(Promega). Digestion was carried out overnight at 37º C in a thermomixer. Samples were prepared
by using SpeedVac centrifuge evaporator (Thermo Scientific), redissolved in 0.1% trifluoroacetic
acid and desalted on ZipTip (Millipore) according to manufacturer’s protocol. Samples were
analyzed by nano-LC-MS/MS coupled to LTQ-Orbitrap mass spectrometer. Chromatographic
separation of peptides was performed using a vented column configuration with trapping and
analytical capillary columns from Nano Separations operated at a flow rate of 110 nl/min by
splitting their flow of an Agilent 1100 capillary pump. An 80 min gradient from 3% to 55%
acetonitrile in water, containing 0.2% formic acid, was used.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

The mass spectrometer was operated in data-dependent mode to automatically measure MS and
MS/MS spectra of top 2 most intense precursor ions with a charge state greater than + 2. Multistage
activation was applied on neutral loss of phosphoric acid. Both MS and MS/MS spectra were
acquired in the orbitrap analyser at a resolution of 30,000 and 7,500 respectively.
Data analysis was accomplished with BioWorks rev. 3.3.1 SP1 (Thermo Scientific). Peptide
identifications were obtained by performing database searches with the embedded SEUQEST
search engine on the Ensembl Human protein database to which common laboratory contaminating
proteins were also added.
The search parameters used in SEQUEST were: up to 4 PMTs per peptide allowed; cysteine
carbamodomethylation as a fixed modification; asparagine deamidation, methionine and histidine
oxidation, phosphorylation of serine, threonine and tyrosine as variable modifications. The
maximum error tolerance of MS scans was set to 10 ppm and 0.1 Da for fragments ions.
GST pull down
After stimulation cells were extracted in lysis buffer containing 50 mM Hepes pH 7.0; 150 mM
NaCl; 0.1% Triton X-100; 10% glycerol; 1.5 mM MgCl2; 1 mM EGTA; 10 mM Na pyrophosphate;
100 mM NaF; 1 mM DTT and 2 mM Na3V04. Equal amounts of cell extracts were incubated with
recombinant fusion proteins GST-SH2-CRK and GST-PTB-SHC bound to Glutathione Sepharose
beads (Roche). The beads were subsequently washed with HNTGN buffer (50 mM Hepes pH 7.0;
150 mM NaCl; 0.1% Triton X-100; 10% glycerol and 2 mM Na3VO4) and analyzed by Western
blotting.
DNA synthesis, chemotaxis, and sprouting assays
Mitogenic activity of HUVEC was assayed in 96-well plates. A total of 6 x103 cells per well were
grown for 16 hours in complete M199 medium. The cells were then starved for 24 hours in M199
medium containing 2% fetal bovine serum (FBS) and 0.5% BSA. Cells were then incubated with
3H-thymidine (0,5 µCi per well) and incubated for 8 hours at 37°C. They were trypsinized, and 3H-
thymidine incorporation was measured by using a -counter (Hewlett Packard, Palo Alto, CA).
Chemotaxis activity was performed in Boyden chambers. A total of 1.5 x 06 HUVECs were grown
for 16 hours in complete M199 medium. The cells were then starved for 24 hours in M199 medium
containing 2% FBS and 0.5% BSA. Polyvinylpyrrolidone-free polycarbonate filters (Nucleopore;
Corning Costar, Acton, MA) with a pore size of 8 µm coated with 0.1% gelatin were used. For
sprouting assays, HUVECs resuspended in complete M199 culture medium containing 0.12%
methylcellulose were seeded in non adherent round-bottom 96-well plates. Single spheroids per
by on May 18, 2011 circres.ahajournals.orgDownloaded from

well (800 cells per spheroid) were embedded into rat tail collagen gels (Boehringer Mannheim,
Mannheim, Germany). HUVEC spheroids were resuspended in M199 containing 20% FBS and 2%
methylcellulose and mixed with an equal volume of 0.4% collagen solution (rapidly transferred into
48-well plates pre-warmed at 37°C and allowed to polymerase) and incubated for 18 to 24 hours.
Sprouting was quantified by measuring the length of the capillary sprouts (ocular grid at 100
magnification) that had grown out of each spheroid (5 to 10 spheroids per experimental group and
experiment were measured).
Apoptosis assay
A total of 5000 cells per cm2 were grown in complete M199 medium for 16 hours on a 1% gelatin-
coated 8-well Chamber slide. Cells were starved for 24 hours in M199 medium supplemented with
0.25% BSA and fixed in 2% paraformaldehyde in PBS; apoptosis was assessed by terminal
deoxynucleotidyl transferase deoxyuridine triphosphate (dUTP) nick end labeling (TUNEL)
staining (Promega), following the manufacturer’s instructions. The percentage of apoptotic cells
was calculated from the number of TUNEL-positive cells divided by the total number of counted
DAPI (4,6 diamidino-2-phenylindole)–stained cells.
References
1. Salameh A, Galvagni F, Bardelli M, Bussolino F, Oliviero S. Direct recruitment of CRK andGRB2 to VEGFR-3 induces proliferation, migration, and survival of endothelial cellsthrough the activation of ERK, AKT, and JNK pathways. Blood. 2005;106:3423-31.
2. Orlandini M, Spreafico A, Bardelli M, Rocchigiani M, Salameh A, Nucciotti S, CapperucciC, Frediani B, Oliviero S. Vascular endothelial growth factor-D activates VEGFR-3expressed in osteoblasts inducing their differentiation. J Biol Chem. 2006;281:17961-17967.
by on May 18, 2011 circres.ahajournals.orgDownloaded from

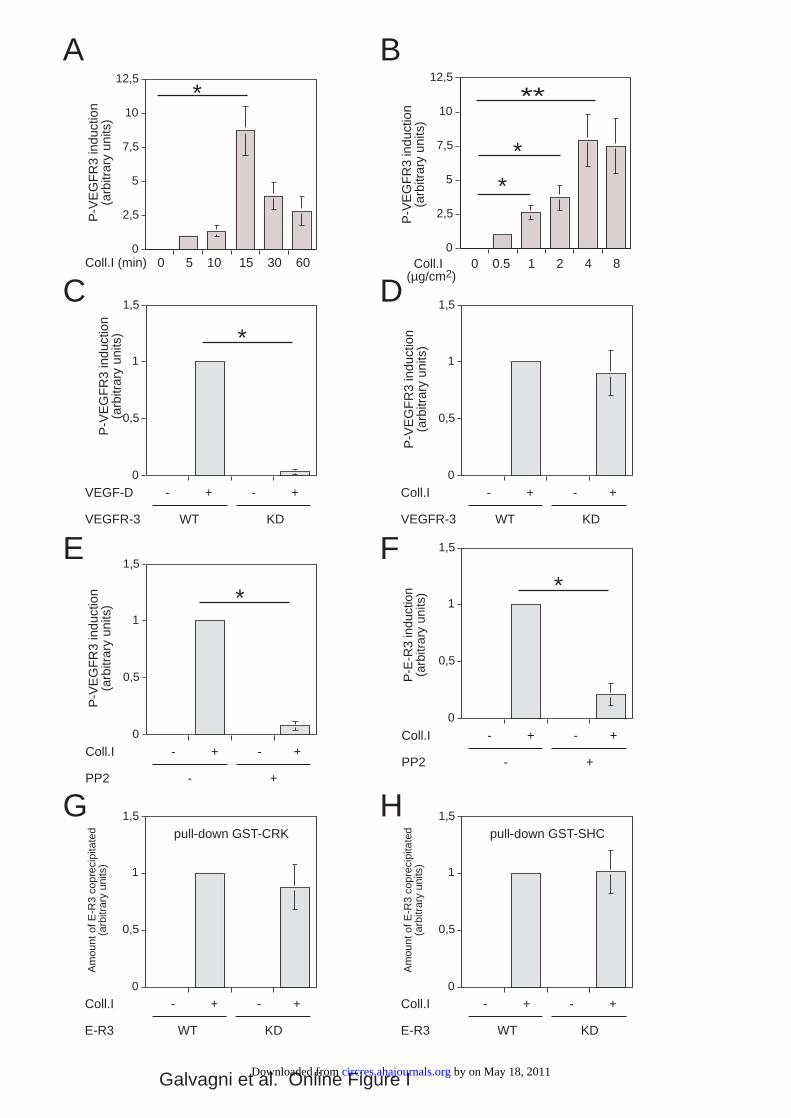
Online Figure I. (A) Quantitative analysis of 3 independent experiments of VEGFR-3
phosphorylation by type I collagen of which one representative experiment is shown Figure 1A. At
the minimum detectable signal was assigned the arbitrary value 1. Statistical significance of
differences were determinated by Student T-test (* p < 0.001). (B) Quantitative analysis of 3
independent experiments of VEGFR-3 phosphorylation by type I collagen of which one
representative experiment is shown Figure 1B. At the minimum detectable signal was assigned the
arbitrary value 1. Statistical significance of differences were determinated by Student T-test (* p <
0.001). (C) Quantitative analysis of 3 independent experiments of VEGF-D-induced VEGFR-3
phosphorylation of which one representative experiment is shown in Figure 1C. The arbitrary value
1 was assigned at the signal obtained after VEGF-D induction in HUVEC over-expressing the wild
type VEGFR-3 (WT). Statistical significance of differences were determinated by Student T-test (*
p < 0.001). (D) Quantitative analysis of 3 independent experiments of VEGFR-3 phosphorylation
by type I collagen of which one representative experiment ris shown in Figure 1D. The arbitrary
value 1 was assigned at the signal obtained after type I collagen induction in HUVEC over-
expressing the wild type VEGFR-3 (WT). (E, F) Quantitative analysis of 3 independent
experiments of VEGFR-3 phosphorylation by type I collagen in cells either treated or untreated
with the c-Src inhibitor PP2 of which one representative experiment is shown in Figure 3C and D.
The arbitrary value 1 was assigned at the signal measured after collagen I induction in untreated
cells. Statistical significance of differences were determinated by Student T-test (* p < 0.001). (G,
H) Quantitative analysis of 3 independent GST-pull-down experiments of which one representative
experiment is shown in Figure 5B. The arbitrary value 1 was assigned at the signal obtained with
extracts of cells over-expressing the wild type chimeric receptor E-R3.
Online Figure II. FACS analysis of the cell types used in the present work for integrin β1, α1, α2
and α3. The antibodies used were: anti human and mouse integrin β1 (Mab1965, Chemicon), anti
human integrin α1 (SM1122FT, Acris Antibodies, GmbH), anti human integrin α2 (SM1125PT,
Acris Antibodies, GmbH), anti human and mouse integrin α3 (SM1544PT, Acris Antibodies,
GmbH), anti mouse integrin α1 (SM2099FT, Acris Antibodies, GmbH) and anti mouse integrin α2
(AM01268PU-T, Acris Antibodies, GmbH).
by on May 18, 2011 circres.ahajournals.orgDownloaded from
0
2,5
5
7,5
10
12,5
Coll.I (min) 0 5 10 15 30 60
P-V
EG
FR
3 in
duct
ion
(arb
itrar
y un
its)
P-V
EG
FR
3 in
duct
ion
(arb
itrar
y un
its)
Coll.I 0 0.5 1 2 4 8 (µg/cm2)
0
0,5
1
1,5
P-V
EG
FR
3 in
duct
ion
(arb
itrar
y un
its)
Coll.I - + - +
VEGFR-3 WT KD
0
0,5
1
1,5
P-V
EG
FR
3 in
duct
ion
(arb
itrar
y un
its)
VEGF-D - + - +
VEGFR-3 WT KD
0
0,5
1
1,5
P-V
EG
FR
3 in
duct
ion
(arb
itrar
y un
its)
Coll.I - + - +
PP2 - +
0
0,5
1
1,5
P-E
-R3
indu
ctio
n(a
rbitr
ary
units
)
0
0,5
1
1,5
Am
ount
of E
-R3
copr
ecip
itate
d(a
rbitr
ary
units
)
Coll.I - + - +
E-R3 WT KD
pull-down GST-CRK
0
0,5
1
1,5
Am
ount
of E
-R3
copr
ecip
itate
d(a
rbitr
ary
units
)
Coll.I - + - +
E-R3 WT KD
pull-down GST-SHC
A B
C D
E F
G H
* **
* *
*
*
0
2,5
5
7,5
10
12,5
*
Coll.I - + - +
PP2 - +
Galvagni et al. Online Figure I by on May 18, 2011 circres.ahajournals.orgDownloaded from

0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102
103
104
105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102
103
104
105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102
103
104
105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102
103
104
105
FITC-A
0
20
40
60
80
100
% o
f M
ax
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
FITC-A
0
20
40
60
80
100
0 102 103 104 105
PE-A
0
20
40
60
80
100
0 102 103 104 105
PE-A
0
20
40
60
80
100
HUVEC
HLMVEC
HEK 293
SYF +/+
SYF -/-
Integrin α1 Integrin α2 Integrin α3 Integrin β1
% o
f M
ax
% o
f M
ax
% o
f M
ax
% o
f M
ax
Galvagni et al. Online Figure II
by on May 18, 2011 circres.ahajournals.orgDownloaded from
Copyright © 2022 FDOKUMEN




















