
gasoline engine intake manifold leakage diagnosis ... - ijicic
Upload
law-rutgersCategory
view
0download
0
Endosomal Leakage and NuclearTranslocation of Multiwalled CarbonNanotubes: Developing a Model for CellUptakeQingxin Mu,†,‡ Dana L. Broughton,† and Bing Yan*,†,§
St. Jude Children’s Research Hospital, Memphis, Tennessee 38105, and School ofPharmaceutical Sciences, School of Chemistry and Chemical Engineering, ShandongUniVersity, Jinan, China, 250100
Received August 13, 2009; Revised Manuscript Received October 14, 2009
ABSTRACT
We report our findings on cellular membrane penetration, endocytosis, endosomal leakage, and nuclear translocation of multiwalled carbonnanotubes (MWCNTs). Our data is consistent with a working model for MWCNTs’ cell uptake and cellular translocations.
Among all nanomaterials, carbon nanotubes (CNTs) possessthe broadest application potential including electrical cir-cuits,1 hydrogen storage,2 drug delivery,3 and tumor imaging.4
In recent years, the perturbation of CNTs on proteins,5
cells,6-8 and live animals9 have also attracted concerns inboth public and scientific communities. Because of theirtubular structure and extremely high aspect-ratio, CNTs canreadily penetrate various biological barriers in mammals,9
plants,10 and microorganisms.11 Furthermore, the largesurface area enables CNTs to bind small molecules,7 pro-teins,5 and nucleic acids.12 They may deliver therapeuticpayloads or DNA and RNA molecules to cells and interferewith biological processes such as signaling transduction.6
Therefore, the behavior of CNTs in living cells includingcell entrance, subcellular locations, and excretion is crucialfor their effects on cellular functions. However, the mech-anism of cell uptake and cellular fate of CNTs is not fullyunderstood and current descriptions are still controversialespecially on cell uptake of CNTs,13-18 their intracellulartranslocation, and subcellular localization.19-23 A popularview is that CNTs are taken up by cells through clathrin-dependent endocytosis.14 Protein- or DNA-coated single-walled carbon nanotubes (SWCNTs) were shown to entercells in an energy-dependent manner. Although the majorityof published data agree with the endocytosis model, theenergy-independent cell uptake was also reported.15 Theauthors argued that the different mechanisms of cell uptake
could be due to the different surface characteristics of CNTs.Another controversy is on subcellular locations of CNTs.Reports described that CNTs go into cells without enteringthe nucleus21,24,25 while other reports found that SWCNTentered the cell nucleus17,19,22,23 and this entrance might bereversible.22 However, there is no knowledge whethermultiwalled carbon nanotubes (MWCNTs) can enter cellnucleus. Furthermore, it is still not clear how CNTs can entercytoplasm to interact with proteins, enzymes, and delivertherapeutic agents.
Understanding of mechanisms of cell entrance, intracellulartranslocation, subcellular locations, and the excretion ofCNTs is of utmost importance in order to elucidate CNTs’possible hazards and their potentials for drug/gene delivery.Herein, we report our ultrastructural observations of celluptakes of CNTs into human embryonic kidney epithelialcells (HEK293). On the basis of our experimental observa-tions and inference from previous reports, a plausible modelfor MWCNTs’ cellular circulation path is proposed. SingleMWCNTs enter cells through direct penetration whileMWCNT bundles enter through endocytosis. MWCNTbundles in endosomes may release single nanotubes that thenpenetrate the endosome membrane and escape into cyto-plasm. Short MWCNTs can also enter the cell nucleus. Allclasses of MWCNTs are recruited into lysosome for excre-tion. This model helps unify previous controversies andprovides novel insights into MWCNTs’ cellular circulations.
Endocytosis is energy-dependent and characterized by itstemperature dependence.26 To compare the effect of surfacecharge on cell uptake, we synthesized both carboxlyated(MWCNT-COOH) and aminated (MWCNT-NH2) CNTs
* To whom correspondence should be addressed. Phone: +9014952797.Fax: +9014955715. E-mail: [email protected].
† St. Jude Children’s Research Hospital.‡ School of Pharmaceutical Sciences, Shandong University.§ School of Chemistry and Chemical Engineering.
NANOLETTERS
2009Vol. 9, No. 12
4370-4375
10.1021/nl902647x CCC: $ 2009 American Chemical SocietyPublished on Web 11/10/2009
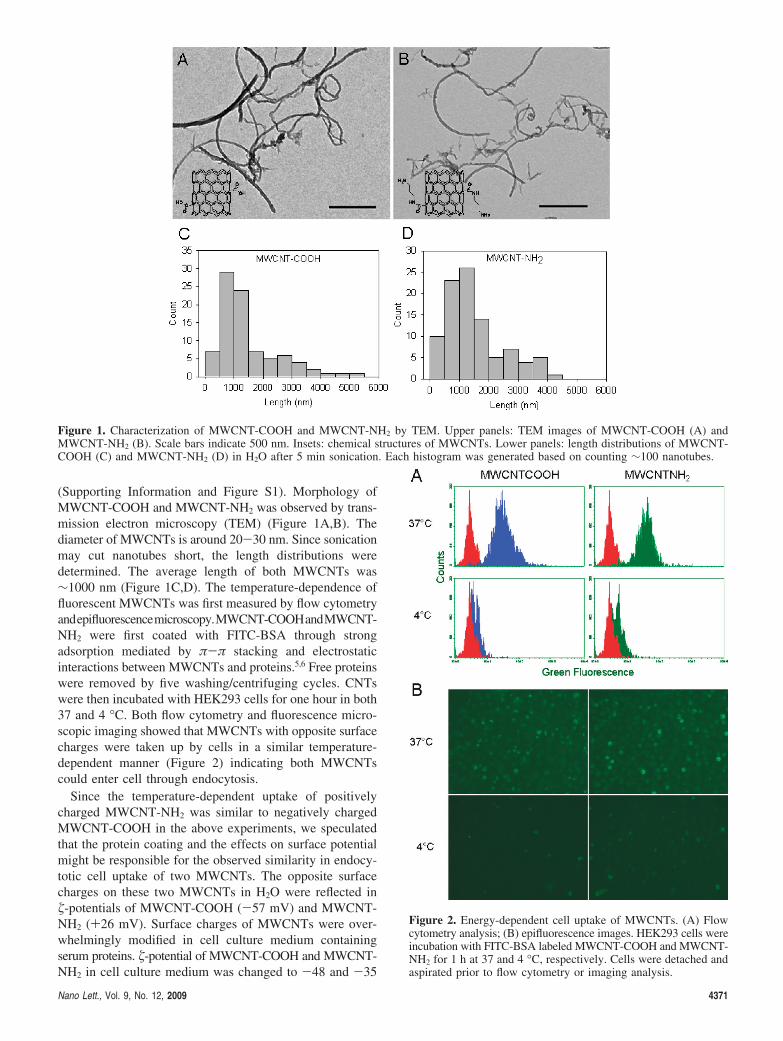
(Supporting Information and Figure S1). Morphology ofMWCNT-COOH and MWCNT-NH2 was observed by trans-mission electron microscopy (TEM) (Figure 1A,B). Thediameter of MWCNTs is around 20-30 nm. Since sonicationmay cut nanotubes short, the length distributions weredetermined. The average length of both MWCNTs was∼1000 nm (Figure 1C,D). The temperature-dependence offluorescent MWCNTs was first measured by flow cytometryandepifluorescencemicroscopy.MWCNT-COOHandMWCNT-NH2 were first coated with FITC-BSA through strongadsorption mediated by π-π stacking and electrostaticinteractions between MWCNTs and proteins.5,6 Free proteinswere removed by five washing/centrifuging cycles. CNTswere then incubated with HEK293 cells for one hour in both37 and 4 °C. Both flow cytometry and fluorescence micro-scopic imaging showed that MWCNTs with opposite surfacecharges were taken up by cells in a similar temperature-dependent manner (Figure 2) indicating both MWCNTscould enter cell through endocytosis.
Since the temperature-dependent uptake of positivelycharged MWCNT-NH2 was similar to negatively chargedMWCNT-COOH in the above experiments, we speculatedthat the protein coating and the effects on surface potentialmight be responsible for the observed similarity in endocy-totic cell uptake of two MWCNTs. The opposite surfacecharges on these two MWCNTs in H2O were reflected in�-potentials of MWCNT-COOH (-57 mV) and MWCNT-NH2 (+26 mV). Surface charges of MWCNTs were over-whelmingly modified in cell culture medium containingserum proteins. �-potential of MWCNT-COOH and MWCNT-NH2 in cell culture medium was changed to -48 and -35
Figure 1. Characterization of MWCNT-COOH and MWCNT-NH2 by TEM. Upper panels: TEM images of MWCNT-COOH (A) andMWCNT-NH2 (B). Scale bars indicate 500 nm. Insets: chemical structures of MWCNTs. Lower panels: length distributions of MWCNT-COOH (C) and MWCNT-NH2 (D) in H2O after 5 min sonication. Each histogram was generated based on counting ∼100 nanotubes.
Figure 2. Energy-dependent cell uptake of MWCNTs. (A) Flowcytometry analysis; (B) epifluorescence images. HEK293 cells wereincubation with FITC-BSA labeled MWCNT-COOH and MWCNT-NH2 for 1 h at 37 and 4 °C, respectively. Cells were detached andaspirated prior to flow cytometry or imaging analysis.
Nano Lett., Vol. 9, No. 12, 2009 4371

mV due to protein binding (Supporting Information andTable S1). As proteins abundantly exist in biological fluids,protein bindings in general may significantly change CNTs’surface characteristics in vivo. Proteins bound to bothMWCNTs were identified by LC-MS/MS analysis showingthat 55 of the most abundant proteins bound to MWCNT-COOH and 37 to MWCNT-NH2 (Supporting Informationand Table S1). Therefore, heavy protein coating on bothMWCNTs made MWCNTs with opposite charges similarnanostructures in term of surface charges. This similarity maydirectly impact their abilities to interact with cells.
Flow cytometry and fluorescence microscope only detectgross fluorescence that emits from cells; highly dispersedCNTs, such as single nanotubes, might not be detectable byeither technique. The cell uptake of single nanotubes mustbe investigated by other methods, such as TEM. TEM wasperformed on cells incubated with MWCNTs for one hourand 48 h. About 20 images were acquired for each sampleand the representative images are shown in Figures 3 and 4
and Supporting Information (Figures S3 and S4). Theentrance of single MWCNTs was repeatedly observed byTEM. For example, Figure 3A shows that a single MWCNT-COOH is penetrating the plasma membrane. Our data suggestthat direct penetration can be designated as one of the routesto deliver MWCNTs into cytoplasm. In contrast to singleMWCNTs, bundled MWCNT-COOH tended to enter cellsthrough endocytosis and in the beginning trapped in intra-cellular endosomes (Figure 3B) consistent with the resultfrom flow cytometry and fluorescence microscopy. MWCNT-COOH bundles within endosomes then released singleMWCNTs that penetrated the endosomal membrane andescaped into cytoplasm (Figure 3C,D, Supporting Informa-tion Figure S3) providing another route for MWCNTs toenter cytoplasm. Because of the direct penetration into cellsand escaping from endosomes, single MWCNTs werefrequently observed in the cytoplasm (Figures 3D,G and 4C,Eand Figure S3 in Supporting Information). At 48 h, cyto-plasmic MWCNT and endosomal escape were also observed
Figure 3. TEM characterization of cell uptake of MWCNT-COOHs and their cellular localizations in HEK293 cells. Cells were incubatedwith nanotubes (100 µg/mL) at 37 °C for 1 and 48 h, respectively. The cells were then fixed, embedded, and sectioned followed by imaginganalysis. Arrows indicate organelles or MWCNT-COOHs: green, plasma membrane; red, endosome; yellow, cytoplasm; blue, lysosome;black, MWCNT-COOH. All scale bars represent 500 nm.
4372 Nano Lett., Vol. 9, No. 12, 2009

(Figure 3F,G). At this time point, much more MWCNT-COOHs were recruited into lysosomes (Figure 3E,H). Theplasma and endosomal membrane penetration mode of cellentrance based on TEM complements the endocytosis modelobserved previously by other researchers14 and our data inprevious sections.
Similar to MWCNT-COOH, MWCNT-NH2s also entercells throughendocytosisanddirectpenetration.TheMWCNT-NH2 bundles in endosomes also released single MWCNTsthat might penetrate endosomal membrane and enter the
cytoplasm (Figure 4A,B). The cytoplasmic accumulation ofsingle MWCNT-NH2s (Figure 4C,E, Supporting InformationFigure S4) was also results of both direct permeation andendosomal escape of single MWCNT-NH2 (Figure 4B). Theendosomal escape of MWCNT-NH2 indicated that the strongmembrane insertion ability of MWCNTs did not depend onsurface charge of MWCNTs after protein coating modifica-tions in biological fluid. The fusion of endosome-like vesicleswith lysosomes was observed (Figure 4D) and the accumula-tion of MWCNTs in lysosome was observed (Figure 4F).As a sorting station, substances in endosome are oftentransported into various compartments such as Golgi ap-paratus, endoplasmic reticulum (ER) or lysosome.27 Throughcarefully examination, we could not find nanotubes in Golgiapparatus or ER in our experiments. Our observationsindicated that recruitment of MWCNT bundles into lysos-omes was the most possible route for their cellular fate.
The accumulation of cytoplasmic MWCNTs increasedtheir chance to enter nucleus. Cell nucleus consists of doublelayered membrane envelop that separates its contents fromthe cytoplasm. It is impermeable to most molecules unlessthose that are able to penetrate through nuclear pores.28 TEMexaminations identified a MWCNT-NH2 (Figure 4G,H) inthe nucleus. This result documented the first direct observa-tion of MWCNT located in a cell nucleus.
Cell uptake of MWCNTs, whether through endocytosisor direct penetration, does not depend on their surface chargeswhen they are modified by protein coating. The effect ofthe dimension or length of MWCNTs on cellular uptake stillneeds to be addressed. We next measured the length ofindividual nanotubes that were taken up by cells after 1 h.About 100 nanotubes with proper orientations were measuredand the histograms were generated (Figure 5). The histogramsshow that the average lengths of MWCNT-COOH andMWCNT-NH2 in cells are around 250 and 175 nm, respec-tively, while the average length of MWCNTs is ∼1000 nmwhen they first are added to the cell culture. Since only tubeslying in the cell section plane could be measured for theirlength, we examined ∼20 TEM micrographs from randomtransverse sections of cells and only counted those tubes withproper orientations. The nanotube length distribution there-fore represents the best approximation on the basis of ourTEM results. The data demonstrate that shorter MWCNTshave a stronger propensity to penetrate plasma membranes.This finding offers us an essential strategy for targetingcytoplasm and nucleus using CNTs as delivery vehicles.Although MWCNTs occupied most subcellular organelles,no cell death was observed except some cell growthinhibition for both MWCNTs (Supporting Information,Figure S2).
Recently processes for cell uptake were described asclathrin-dependent endocytosis14 and endocytosis-indepen-dent cell uptake.15 Subcellular locations of CNTs were alsopinpointed in scattered studies.18,20,22,23,25 CNTs were foundin cytoplasm, endosomes, and lysosomes. Only SWCNTswere found in the cell nucleus. These phenomena wereseparately reported and currently there is no model thatdescribes the cellular circulation of CNTs from cell entrance
Figure 4. TEM characterization of cell uptake of MWCNT-NH2sand their cellular localizations in HEK293 cells. Cells wereincubated with nanotubes (100 µg/mL) at 37 °C for 1 and 48 h,respectively. The cells were then fixed, embedded, and sectionedfollowed by imaging analysis. Arrows indicate organelles orMWCNT-NH2s: green, plasma membrane; red, endosome; yellow,cytoplasm; blue, lysosome; black, MWCNT-NH2. All scale barsrepresent 500 nm, except the scale bar in H, which indicates 20nm.
Nano Lett., Vol. 9, No. 12, 2009 4373

to excretion. Previous reports and our own experimentalresults are consistent with a working model for the celluptake of CNTs (Figure 6). In this model, CNTs are dividedinto two classes, clusters and singles. CNT clusters are takenup by cells through energy-dependent endocytosis process.
The CNT bundles then release single nanotubes that escapeendosomes by penetrating endosome membrane and enteringthe cytoplasm. Alternatively, the highly dispersed singleCNTs cross cell membrane and enter cells directly bypenetrating cellular membranes. All CNTs are recruited intolysosomes for excretion. Cytoplasmic CNTs can also enterthe nucleus suggesting the strong ability of CNTs to crossseveral biological membrane barriers.
In summary, we observed the size dependent and directmembrane penetration by both negatively and positivelycharged single MWCNTs, endocytosis of bundled MWCNTs,the endosomal leakage of single MWCNTs from bundles,and the nuclear translocation of MWCNT. On the basis ofour experimental observations and previous reports, weproposed a working model that describes our currentunderstanding of CNTs’ cell uptake. The model will havemajor impacts on both drug delivery and toxicity researchof CNTs. For example, all cellular CNTs may be exposedto cytoplasm so that the unexpected interactions with cellularfunctional molecules are likely to occur. Therefore, a betterunderstanding of CNTs’ cellular behavior provides bothopportunities and challenges on the way to the developmentof nanomedicine.
Acknowledgment. We thank Ms. Linda Mann (Cell andtissue imaging, St. Jude Children’s Research Hospital) forthe technical assistance on TEM and Hartwell Center forBioinformatics & Biotechnology at St. Jude Children’sResearch Hospital for proteomics analysis. This work wassupported by National Cancer Institute (P30CA027165), theAmerican Lebanese Syrian Associated Charities (ALSAC),and St. Jude Children’s Research Hospital.
Supporting Information Available: Details of the ex-perimental procedure, FT-IR characterization of MWCNTs,identification of MWCNT bound proteins, cell growthmeasurement, and additional TEM images are available. Thismaterial is available free of charge via the Internet at http://pubs.acs.org.
References(1) Collins, P. C.; Arnold, M. S.; Avouris, P. Science 2001, 292, 706–
709.(2) Dillon, A. C.; Jones, K. M.; Bekkedahl, T. A.; Kiang, C. H.; Bethune,
D. S.; Heben, M. J. Nature 1997, 386, 377–379.(3) Bianco, A.; Kostarelos, K.; Prato, M. Curr. Opin. Chem. Biol. 2005,
9, 674–679.(4) Zavaleta, C.; de la Zerda, A.; Liu, Z.; Keren, S.; Cheng, Z.; Schipper,
M.; Chen, X.; Dai, H.; Gambhir, S. S. Nano Lett. 2008, 8, 2800–2805.
(5) Mu, Q.; Liu, W.; Xing, Y.; Zhou, H.; Li, Z.; Zhang, Y.; Ji, L.; Wang,F.; Si, Z.; Zhang, B.; Yan, B. J. Phys. Chem. C 2008, 112, 3300–3307.
(6) Mu, Q.; Du, G.; Chen, T.; Zhang, B.; Yan, B. ACS Nano 2009, 3,1139–1144.
(7) Zhou, H. Y.; Mu, Q. X.; Gao, N. N.; Liu, A. F.; Xing, Y. H.; Gao,S. L.; Zhang, Q.; Qu, G. B.; Chen, Y. Y.; Liu, G.; Zhang, B.; Yan, B.Nano Lett. 2008, 8, 859–865.
(8) Wick, P.; Manser, P.; Limbach, L. K.; Dettlaff-Weglikowska, U.;Krumeich, F.; Roth, S.; Stark, W. J.; Bruinink, A. Toxicol. Lett. 2007,168, 121–131.
(9) Qu, G. B.; Bai, Y. H.; Zhang, Y.; Jia, Q.; Zhang, W. D.; Yan, B.Carbon 2009, 47, 2060–2069.
(10) Liu, Q. L.; Chen, B.; Wang, Q. L.; Shi, X. L.; Xiao, Z. Y.; Lin, J. X.;Fang, X. H. Nano Lett. 2009, 9, 1007–1010.
Figure 5. Length distributions of MWCNT-COOH (A) andMWCNT-NH2 (B) inside cells after 1 h incubation with MWCNTsat 37 °C. Each histogram was generated based on counting 100individual nanotubes with proper orientations.
Figure 6. A working model for cell uptake of MWCNTs. Numbers(1-5) and letters (a-d) indicate different steps in two possiblecellular translocation pathways of MWCNTs. The bundled MWCNTsbind to cell membranes (1) and are subsequently internalized intocells (2) inside endosomes. In the endosomes, bundles release singleMWCNTs that penetrate endosomal membranes and enter cyto-plasm (3). Both residual bundled MWCNTs in endosomes and freeMWCNTs in the cytoplasm are recruited into lysosomes forexcretion (4,5). Single MWCNTs enter cells through directmembrane penetration (a) to enter cytoplasm (b). They are recruitedinto lysosomes for excretion (c,d). Short MWCNTs are also ableto enter the nucleus.
4374 Nano Lett., Vol. 9, No. 12, 2009

(11) Kang, S.; Herzberg, M.; Rodrigues, D. F.; Elimelech, M. Langmuir2008, 24, 6409–6413.
(12) Li, X.; Peng, Y. H.; Ren, J. S.; Qu, X. G. Proc. Natl. Acad. Sci. U.S.A.2006, 103, 19658–19663.
(13) Chin, S. F.; Baughman, R. H.; Dalton, A. B.; Dieckmann, G. R.;Draper, R. K.; Mikoryak, C.; Musselman, I. H.; Poenitzsch, V. Z.;Xie, H.; Pantano, P. Exp. Biol. Med. 2007, 232, 1236–1244.
(14) Kam, N. W. S.; Liu, Z. A.; Dai, H. J. Angew. Chem., Int. Ed. 2006,45, 577–581.
(15) Kostarelos, K.; Lacerda, L.; Pastorin, G.; Wu, W.; Wieckowski, S.;Luangsivilay, J.; Godefroy, S.; Pantarotto, D.; Briand, J. P.; Muller,S.; Prato, M.; Bianco, A. Nat. Nanotechnol. 2007, 2, 108–113.
(16) Lacerda, L.; Raffa, S.; Prato, M.; Bianco, A.; Kostarelos, K. NanoToday 2007, 2, 38–43.
(17) Pantarotto, D.; Briand, J. P.; Prato, M.; Bianco, A. Chem. Commun.2004, 16–17.
(18) Zhu, Y.; Li, W. X.; Li, Q. N.; Li, Y. G.; Li, Y. F.; Zhang, X. Y.;Huang, Q. Carbon 2009, 47, 1351–1358.
(19) Mooney, E.; Dockery, P.; Greiser, U.; Murphy, M.; Barron, V. NanoLett. 2008, 8, 2137–2143.
(20) Porter, A. E.; Gass, M.; Muller, K.; Skepper, J. N.; Midgley, P. A.;Welland, M. Nat. Nanotechnol. 2007, 2, 713–717.
(21) Lacerda, L.; Pastorin, G.; Gathercole, D.; Buddle, J.; Prato, M.; Bianco,A.; Kostarelos, K. AdV. Mater. 2007, 19, 1480–1484.
(22) Cheng, J. P.; Fernando, K. A. S.; Veca, L. M.; Sun, Y. P.; Lamond,A. I.; Lam, Y. W.; Cheng, S. H. ACS Nano 2008, 2, 2085–2094.
(23) Porter, A. E.; Gass, M.; Bendall, J. S.; Muller, K.; Goode, A.; Skepper,J. N.; Midgley, P. A.; Welland, M. ACS Nano 2009, 3, 1485.
(24) Zhang, D. W.; Yi, C. Q.; Zhang, J. C.; Chen, Y.; Yao, X. S.; Yang,M. S. Nanotechnology 2007, 18, 9.
(25) Kang, B.; Yu, D. C.; Chang, S. Q.; Chen, D.; Dai, Y. D.; Ding, Y. T.Nanotechnology 2008, 19, 8.
(26) Weigel, P. H.; Oka, J. A. J. Biol. Chem. 1981, 256, 2615–2617.(27) Mellman, I. Ann. ReV. Cell DeV. Biol. 1996, 12, 575–625.(28) Terry, L. J.; Shows, E. B.; Wente, S. R. Science 2007, 318, 1412–1416.
NL902647X
Nano Lett., Vol. 9, No. 12, 2009 4375
Copyright © 2022 FDOKUMEN



















