
Endogenous humoral autoreactive immune responses to apoptotic cells: Effects on phagocytic uptake,...
14
Endogenous humoral autoreactive immune responses to apoptotic cells: Effects on phagocytic uptake, chemotactic migration and antigenic spread Joy Das, Pooja Arora, Donald Gracias, Alagangula Praveen, Beena P. J. Raj, Elishba Martin and Rahul Pal Immunoendocrinology Laboratory, National Institute of Immunology, New Delhi, India Enhanced cell death and deficient clearance of cellular debris are thought to contribute to increased self-antigen exposure in systemic autoimmune disease. To investigate the characteristics of early humoral autoimmune responses, six monoclonal antibodies were generated from two autoimmune prone strains of mice. All antibodies specifically bound the surface of late-stage apoptotic cells. Similar antibody reactivities were present in the sera of patients with systemic lupus erythematosus. While IgM antibodies significantly reduced the phagocytic uptake of apoptotic thymocytes, IgG antibodies enhanced uptake. Poly-reactivity was demonstrated in the recognition of ribonucleoproteins and lipids. An antibody reactive towards lysophosphatidylcholine reversed lysophosphatidylcholine- mediated inhibition of LPS-induced TNF-a production and adversely affected the trans- migration of phagocytes towards an apoptotic stimulus. In several instances, CDR were characterized by the accumulation of somatic mutations. Anti-idiotypic antibodies generated upon immunization bound distinct cellular moieties and self-antigens. Poly- specific, apoptotic cell-reactive autoantibodies can therefore directly impact upon the course of disease by influencing phagocytic uptake of apoptotic cells, by inducing a pro- inflammatory environment through neutralization of bioactive lipids, by blinding phago- cytes to the presence of dying cells through the negation of lipidic chemotactic signals, and by mediating diversification of the humoral autoimmune response via the idiotypic network. Key words: Apoptosis . Autoimmunity . Systemic lupus erythematosus Introduction Systemic lupus erythematosus (SLE) is the prototypical organ non-specific autoimmune disease. The disease generally mani- fests a lifelong, relapsing–remitting phenotype. Multiple organs are targeted; chronic renal failure is a serious consequence, and patients can also exhibit arthritis, neurological involvement and immune dysfunction [1]. Genetic pre-disposition has been described in both animals and humans, and various susceptibility loci have been identified using genetic studies [2]. Both spontaneous and derived murine models of disease exist and have been extensively utilized to study pathology [3]. Despite significant advances in understanding lupus-related organ dysfunction, drawing correlations of disease manifesta- tions with autoimmune specificity remains a high priority. More than 100 different antibody specificities have been described and, while most may be epiphenomena, some autoantibodies may have destructive potential. Anti-dsDNA antibodies have been associated with kidney pathology [4], anti-Ro and anti-La responses with neonatal lupus and congenital heart block [5], Correspondence: Dr. Rahul Pal e-mail: [email protected] & 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu Eur. J. Immunol. 2008. 38: 3561–3574 DOI 10.1002/eji.200838624 Clinical immunology 3561
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Endogenous humoral autoreactive immune responses to apoptotic cells: Effects on phagocytic uptake,...

Endogenous humoral autoreactive immune responsesto apoptotic cells: Effects on phagocytic uptake,chemotactic migration and antigenic spread
Joy Das, Pooja Arora, Donald Gracias, Alagangula Praveen, Beena P. J. Raj,
Elishba Martin and Rahul Pal
Immunoendocrinology Laboratory, National Institute of Immunology, New Delhi, India
Enhanced cell death and deficient clearance of cellular debris are thought to contribute to
increased self-antigen exposure in systemic autoimmune disease. To investigate the
characteristics of early humoral autoimmune responses, six monoclonal antibodies were
generated from two autoimmune prone strains of mice. All antibodies specifically bound
the surface of late-stage apoptotic cells. Similar antibody reactivities were present in the
sera of patients with systemic lupus erythematosus. While IgM antibodies significantly
reduced the phagocytic uptake of apoptotic thymocytes, IgG antibodies enhanced uptake.
Poly-reactivity was demonstrated in the recognition of ribonucleoproteins and lipids. An
antibody reactive towards lysophosphatidylcholine reversed lysophosphatidylcholine-
mediated inhibition of LPS-induced TNF-a production and adversely affected the trans-
migration of phagocytes towards an apoptotic stimulus. In several instances, CDR were
characterized by the accumulation of somatic mutations. Anti-idiotypic antibodies
generated upon immunization bound distinct cellular moieties and self-antigens. Poly-
specific, apoptotic cell-reactive autoantibodies can therefore directly impact upon the
course of disease by influencing phagocytic uptake of apoptotic cells, by inducing a pro-
inflammatory environment through neutralization of bioactive lipids, by blinding phago-
cytes to the presence of dying cells through the negation of lipidic chemotactic signals, and
by mediating diversification of the humoral autoimmune response via the idiotypic
network.
Key words: Apoptosis . Autoimmunity . Systemic lupus erythematosus
Introduction
Systemic lupus erythematosus (SLE) is the prototypical organ
non-specific autoimmune disease. The disease generally mani-
fests a lifelong, relapsing–remitting phenotype. Multiple organs
are targeted; chronic renal failure is a serious consequence, and
patients can also exhibit arthritis, neurological involvement and
immune dysfunction [1]. Genetic pre-disposition has been
described in both animals and humans, and various susceptibility
loci have been identified using genetic studies [2]. Both
spontaneous and derived murine models of disease exist and
have been extensively utilized to study pathology [3].
Despite significant advances in understanding lupus-related
organ dysfunction, drawing correlations of disease manifesta-
tions with autoimmune specificity remains a high priority. More
than 100 different antibody specificities have been described and,
while most may be epiphenomena, some autoantibodies may
have destructive potential. Anti-dsDNA antibodies have been
associated with kidney pathology [4], anti-Ro and anti-La
responses with neonatal lupus and congenital heart block [5],Correspondence: Dr. Rahul Pale-mail: [email protected]
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Eur. J. Immunol. 2008. 38: 3561–3574 DOI 10.1002/eji.200838624 Clinical immunology 3561

and anti-phospholipid antibodies with recurrent spontaneous
abortion [6]. Additionally, titers of complement-fixing antibodies
to cell-surface moieties upon leucocytes are negatively correlated
with circulating leucocyte numbers [7]. While some studies
indicate a correlation between anti-ribosomal P antibodies
and neuropsychiatric manifestations [8], contradictory data
have also emerged.
Interestingly, in both SLE patients and several disease-prone
animal models, aberrance in apoptotic processes has been observed.
Several studies document excessive spontaneous apoptosis (or
excessive susceptibility to apoptotic stimuli) in lupus patients [9], as
well as an inefficient clearance of apoptotic debris [10]. Though
apoptosis is considered a non-inflammatory event, infusion of
experimental animals with syngeneic apoptotic cells generates a
lupus-like autoimmune response [11], albeit with no associated
long-term pathology. In furtherance of the postulate-linking
apoptosis and systemic immune dysfunction, animals genetically
modified to impair the uptake of apoptotic cells exhibit lupus
pathology [12, 13]. Apoptotic debris may therefore be the source
of antigenic material that initiates autoreactivity in genetically
susceptible individuals.
Upon apoptosis, cells externalize phosphatidylserine (PS) as
an early event [14]. Subsequently, ‘‘blebs’’ containing both
nuclear and cytoplasmic autoantigens extrude from the cell
surface [15], and these particles quite possibly constitute the
initial immune trigger. Autoantibodies known to target
several lupus-specific antigens bind to the surface of apoptotic
cells before true cellular permeability is achieved [16]. It is
conceivable, therefore, that autoantibodies specifically reactive
to the apoptotic cell surface could have disease-modifying
effects.
This report seeks to investigate properties of the apoptotic
cell-specific antibody responses in lupus in the context of disease.
Apoptotic cells, rather than individual molecules known to be
externalized during apoptosis, served as targets in the
development of six murine monoclonal antibodies (2C11,
IgMk; 2H8, IgMk; 1B4, IgMk; 2C3, IgG2bk; 1B3, IgG2ak; 1B1,
IgG2ak) from two autoimmune-prone strains of mice.
Caspase activity was shown to be critical for antibody binding.
Antibodies exhibited varying degrees of cross-reactivity to
commonly targeted ribonucleoprotein (RNP) autoantigens as
well as phospholipids, and impacted upon the phagocytic uptake
of apoptotic cells in an isotype-dependent manner. Antibody
2C11, which bound lysophosphatidylcholine (LPC), influenced
two important biological roles mediated by the lipid: the
suppression of inflammatory responses and the chemotaxis of
phagocytic cells. Antisera, generated upon immunization with
either antibody 2C11 or antibody 1B1, exhibited anti-self-reac-
tivity with a diversified antigenic recognition profile. Variable
region analysis revealed that distinct gene families were
employed, with some antibodies exhibiting preferential accumu-
lation of somatic mutations in the CDR. Germline or somatically
mutated early antibody responses specifically directed against
dying cells could thus conceivably impact upon the course and
severity of disease.
Results
Reactivity towards intra-cellular and cell-surfaceantigens
Table 1 presents nomenclature, cellular binding pattern, Ig
isotype and animal source of the antibodies.
A total of 465 independent hybridomas were established, using
spleen cells derived from aging, lupus-prone mice. Of these, 32
secreted antibodies reactive to the surface of healthy cells, and 58
secreted antibodies reactive towards permeabilized cells and non-
reactive towards apoptotic cells. Figures 1 and 2 show the binding
properties of six antibodies chosen for their specificity towards
permeabilized as well as apoptotic cells, and non-reactivity towards
moieties present on the surface of healthy cells. Antibodies 2C11 and
1B4 exclusively recognized cytoplasmic moieties while antibodies
1B1 and 1B3 bound nuclear antigens. Antibodies 2H8 and 2C3
bound antigens in both regions (Fig. 1A–F). Similar binding patterns
were observed on many cell types and across two species (humans
and mice), indicating evolutionary and lineage antigenic conserva-
tion (data not shown). Relevant isotype control antibodies did
not bind either permeabilized or non-permeabilized cells (Fig. 1G
and H).
Reactivity towards apoptotic cells
Eight hours after the induction of apoptosis, cells that bound
antibody also bound Annexin-V (Fig. 2A, top right quadrants) and
cells not binding Annexin-V were also not bound by antibody
(Fig. 2A, bottom right quadrants). Cells that bound Annexin-V but
not antibody (Fig. 2A, top left quadrants), as well as healthy calls
that bound neither reagent (Fig. 2A, bottom left quadrants), were
also observed. Addition of the pan-caspase inhibitor z-VAD-fmk to
cultures during the induction of apoptosis caused a reduction in both
Annexin-V and antibody reactivity; the few cells recognized by the
antibodies also bound Annexin-V (Fig. 2B).
Upon confocal microscopy, all six antibodies exclusively
bound cells also recognized by Annexin-V (Fig. 2C), reiterating
results obtained by FACS analysis. While Annexin-V reactivity
appeared to be relatively more evenly distributed across the cell
surface, in most instances antibody reactivity appeared confined
to restricted regions of the membrane, possibly upon incipient
Table 1. Description of nomenclature, intra-cellular specificity, isotypeand animal source of the monoclonal antibodies
Antibody Specificity Isotype Strain
2C11 Cytoplasmic IgMk C57BL/6lpr/lpr
2H8 Cytoplasmic/nuclear IgMk C57BL/6lpr/lpr
1B4 Cytoplasmic IgMk NZB/W F1
2C3 Cytoplasmic/nuclear IgG2bk NZB/W F1
1B3 Nuclear IgG2ak NZB/W F1
1B1 Nuclear IgG2ak NZB/W F1
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3562
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

blebs. Regions of overlap between Annexin-V and antibody
reactivity appeared yellow or orange in merged images, as in
images depicting Antibodies 1B4 and 2C3. Healthy cells not
binding Annexin-V or antibody (as in the image depicting
Antibody 1B4 reactivity), as well as cells at the early stages
of apoptosis, which bound Annexin-V but not antibody (as in
images depicting antibodies 1B3 and 1B4 reactivity), were
apparent.
At 12 h post-apoptotic stimulus, preferential antibody binding to
surface protrusions (putative apoptotic blebs) was more significant
(Fig. 2D). Antibody 2C11 bound blebs isolated from apoptotic SP2/0
cells (Fig. 2E). In addition, blebs isolated from apoptotic 2C11
hybridoma cells, as opposed to blebs isolated from apoptotic SP2/0
cells, were demonstrated to contain adhered antibody (Fig. 2E).
Antigenic analysis
IgM antibodies appeared to be reactive towards multiple RNP
(Fig. 3A). Antibody 2C11 predominantly bound Ro52 and the
U1-RNP A Protein, while antibody 2H8 bound Ro60 to the most
significant extent, but also demonstrated recognition of Ro52,
SmB, SmD and the U1-RNP A Protein. Antibody 1B4 recognized
Ro52 and Ro60, while demonstrating minor reactivity towards all
the other proteins. Similar (but distinctive) poly-reactivity was
observed with the IgG antibodies (Fig. 3B). Antibodies 2C3 and
1B1 bound Ro60 and SmD in a dominant fashion, while antibody
1B3 exclusively recognized SmD. Antibody 1D1 (IgG3k), which
was generated as part of another study and recognized the cell
membrane of healthy, non-apoptotic Jurkat cells, was employed
as control; it was essentially non-reactive.
Antibodies to oxidized phospholipids have been shown to bind
to the apoptotic cell surface and induce the release of pro-
inflammatory cytokines [17]. The IgM antibody 2C11 specifically
bound LPC and the IgG antibody 2C3 exhibited a variable degree
of cross-reactivity to different lipids. The other antibodies were
poorly reactive (Fig. 3C and D).
Human SLE sera were evaluated to accord clinical relevance
to the data. The autoantigen Ro60 was chosen for this analysis
since five of the six antibodies exhibited appreciable binding to it.
Eve
nts
FL
A
D E F
HG
B C
Eve
nts
FL
Eve
nts
FL
Eve
nts
FL
Eve
nts
FL
Eve
nts
FL
Eve
nts
FL FL
Eve
nts
Figure 1. (A–F) Recognition of permeabilized SP2/O cells (solid, thick profiles) by the IgM (A: 2C11; B: 2H8; C: 1B4) and IgG (D: 2C3; E: 1B3; F: 1B1)monoclonal antibodies. Non-permeabilized cells (dashed profiles) were not recognized. Solid, thin profiles indicate negative controls, stainingobtained in the presence of only secondary antibodies. Also depicted (at right for each antibody) are respective intra-cellular localization patternsupon permeabilized cells on immunofluorescence analysis (top) and corresponding phase contrast images (bottom). (G and H) Binding of isotypecontrol antibodies (IgMk: red profiles; IgG2ak: green profiles; IgG2bk: blue profiles) to (G) permeabilized and (H) non-permeabilized SP2/O cells. Grayprofiles indicate negative controls, staining obtained in the presence of only secondary antibodies.
Eur. J. Immunol. 2008. 38: 3561–3574 Clinical immunology 3563
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
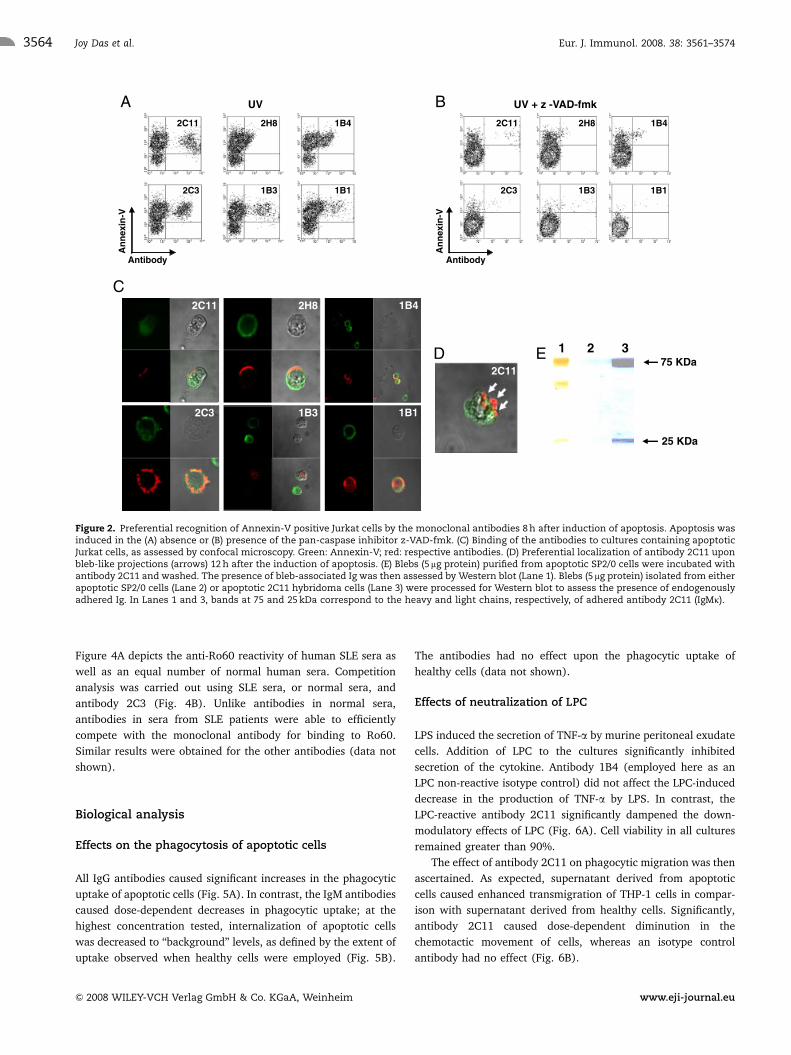
Figure 4A depicts the anti-Ro60 reactivity of human SLE sera as
well as an equal number of normal human sera. Competition
analysis was carried out using SLE sera, or normal sera, and
antibody 2C3 (Fig. 4B). Unlike antibodies in normal sera,
antibodies in sera from SLE patients were able to efficiently
compete with the monoclonal antibody for binding to Ro60.
Similar results were obtained for the other antibodies (data not
shown).
Biological analysis
Effects on the phagocytosis of apoptotic cells
All IgG antibodies caused significant increases in the phagocytic
uptake of apoptotic cells (Fig. 5A). In contrast, the IgM antibodies
caused dose-dependent decreases in phagocytic uptake; at the
highest concentration tested, internalization of apoptotic cells
was decreased to ‘‘background’’ levels, as defined by the extent of
uptake observed when healthy cells were employed (Fig. 5B).
The antibodies had no effect upon the phagocytic uptake of
healthy cells (data not shown).
Effects of neutralization of LPC
LPS induced the secretion of TNF-a by murine peritoneal exudate
cells. Addition of LPC to the cultures significantly inhibited
secretion of the cytokine. Antibody 1B4 (employed here as an
LPC non-reactive isotype control) did not affect the LPC-induced
decrease in the production of TNF-a by LPS. In contrast, the
LPC-reactive antibody 2C11 significantly dampened the down-
modulatory effects of LPC (Fig. 6A). Cell viability in all cultures
remained greater than 90%.
The effect of antibody 2C11 on phagocytic migration was then
ascertained. As expected, supernatant derived from apoptotic
cells caused enhanced transmigration of THP-1 cells in compar-
ison with supernatant derived from healthy cells. Significantly,
antibody 2C11 caused dose-dependent diminution in the
chemotactic movement of cells, whereas an isotype control
antibody had no effect (Fig. 6B).
1 375 KDa
25 KDa
2C11
1B42C11 2H8
C
D
Antibody
An
nex
in-V
UV
2C11 2H8
2C3 1B3
Antibody
An
nex
in-V
UV + z -VAD-fmk
2C11 2H8 1B4
2C3 1B3 1B1
B
E
1B4
1B1
1B32C3 1B1
A
2
Figure 2. Preferential recognition of Annexin-V positive Jurkat cells by the monoclonal antibodies 8 h after induction of apoptosis. Apoptosis wasinduced in the (A) absence or (B) presence of the pan-caspase inhibitor z-VAD-fmk. (C) Binding of the antibodies to cultures containing apoptoticJurkat cells, as assessed by confocal microscopy. Green: Annexin-V; red: respective antibodies. (D) Preferential localization of antibody 2C11 uponbleb-like projections (arrows) 12 h after the induction of apoptosis. (E) Blebs (5 mg protein) purified from apoptotic SP2/0 cells were incubated withantibody 2C11 and washed. The presence of bleb-associated Ig was then assessed by Western blot (Lane 1). Blebs (5mg protein) isolated from eitherapoptotic SP2/0 cells (Lane 2) or apoptotic 2C11 hybridoma cells (Lane 3) were processed for Western blot to assess the presence of endogenouslyadhered Ig. In Lanes 1 and 3, bands at 75 and 25 kDa correspond to the heavy and light chains, respectively, of adhered antibody 2C11 (IgMk).
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3564
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Anti-idiotypic responses
Antibody 1B1 (IgG2ak) was immunized in 6 wk-old NZB/W F1
lupus-prone mice, the mouse strain from which the antibody was
derived. As shown in Fig. 7A (right panel), upon FACS analysis,
sera demonstrated enhanced presence of IgG antibodies reactive
towards self-antigen(s), in comparison with sera generated by
immunization with adjuvant alone; antibodies of the IgM isotype
did not exhibit such anti-self-reactivity (Fig. 7A, left panel). As
previously indicated, antibody 1B1 predominantly demonstrated
reactivity towards nuclear antigen(s), with the cytoplasm
remaining unbound (Fig. 7B, top left panel). Antibodies in the
sera of animals immunized with antibody 1B1, on the other hand,
bound both nuclear and cytoplasmic antigens (Fig. 7B, top right
panel). In contrast, minimal binding was observed when serum
obtained from animals immunized with adjuvant was employed
(Fig. 7B, bottom left panel). Antibodies in the sera of antibody-
1B1-immunized animals demonstrated significantly enhanced
recognition of Ro60, La and the U1-RNP A protein in ELISA
assays (Fig. 7B, bottom right panel). Though Ro60 was
recognized by antibody 1B1 (which also bound SmD; Fig. 3B),
the reactivity towards the latter two proteins indicated the
presence of newer specificities in the sera of animals immunized
with this antibody.
Evidence of such a diversification of anti-self-responses was
also obtained upon immunization of C57BL/6 animals with
antibody 2C11 (IgMk). While such sera demonstrated significant
IgG autoreactive responses, no appreciable IgM responses were
observed. Antisera bound both cytoplasmic and nuclear antigens
in contrast to the cytoplasmic reactivity demonstrated by
antibody 2C11. Sera generated upon immunization with the
adjuvant did not contain significant autoreactive antibody
specificities. Further analysis revealed the preferential recogni-
tion of La and the U1-RNP A Protein in anti-2C11 sera. Though
the latter was an antigen bound by antibody 2C11 (which also
bound Ro52 in a dominant fashion, Fig. 3A), the former repre-
sented a new specificity (data not shown).
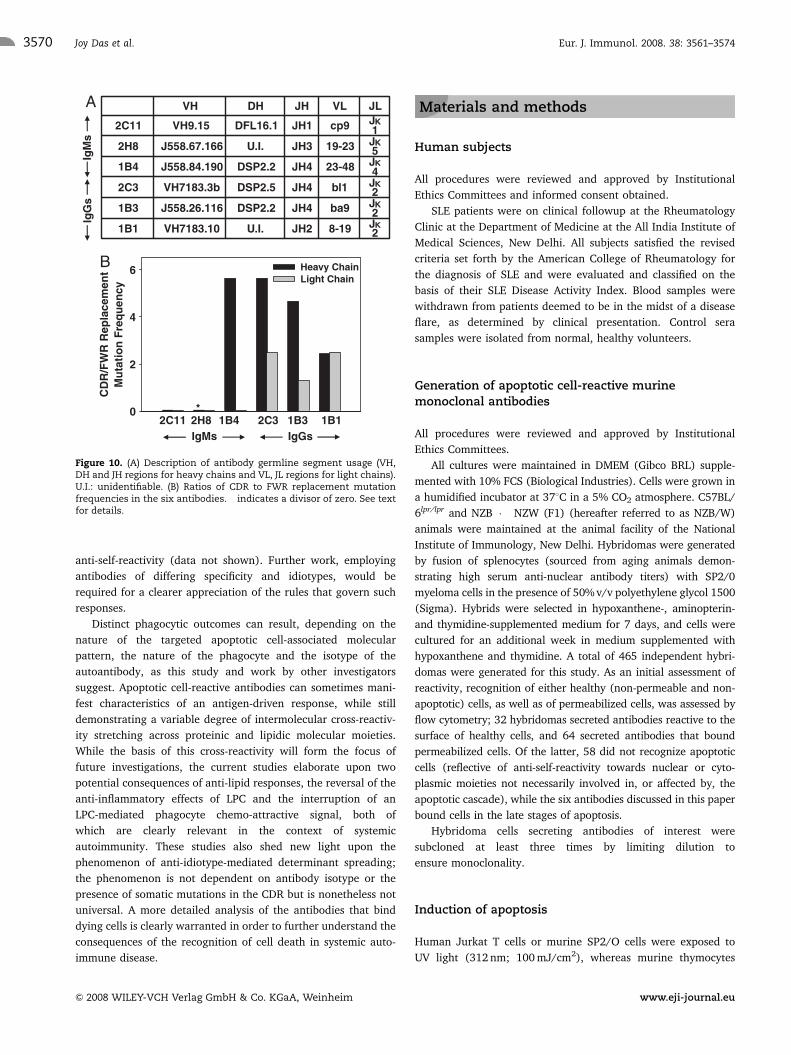
V region analysis
Figure 8 provides nucleotide sequence data for the IgM
antibodies and Fig. 9 for the IgG antibodies. Significantly,
antibodies demonstrated closest homologies to distinct germline
segments (and/or combinations thereof, as shown in Fig. 10A)
for both light and heavy chains, possibly a reflection of the
distinct antigenic specificities observed. For Antibodies 2H8 and
1B1, the heavy chain D regions were unidentifiable; Fig. 10B
shows ratios of the mutation frequencies observed in the CDR and
framework regions (FWR); a value in excess of unity was
obtained for the heavy chain of the IgM antibody 1B4, on account
of amino acid replacements observed in the CDR1 and CDR2
regions. For the heavy chain of antibody 2H8, a numerical value
could not be assigned on account of the divisor being zero. The
CDR of the IgM light chains were not preferentially mutated. Both
the heavy and light chains of the IgG Antibodies 2C3 and 1B1
contained an increased number of somatic mutations in the CDR
compared with FWR. While the heavy chain of antibody 1B3 was
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6Ro52Ro60SmBSmDLaU1-RNP A
0.0
0.5
1.0
1.5
2.0
2.5
3.0Ro52Ro60SmBSmDLaU1-RNP A
0.0
0.1
0.2
0.3
0.4 LPCLPAPSPEPCPACL
0.0
0.2
0.4
0.6
0.8LPCLPAPSPEPCPACL
A B
C D2C11 2H8 1B4
2C11 2H8 1B4
Ab
sorb
ance
(45
0 n
m)
Ab
sorb
ance
(45
0 n
m)
Ab
sorb
ance
(45
0 n
m)
Ab
sorb
ance
(45
0 n
m)
2C3 1B3 1B1 1D1
2C3 1B3 1B1 1D1
Figure 3. Reactivity of the IgM (A and C) and IgG (B and D) monoclonal antibodies towards recombinant RNP autoantigens (A and B) andphospholipids (C and D) by ELISA. LPC, lysophosphatidylcholine; LPA, lysophosphatidic acid; PS, phosphatidylserine; PE, phosphatidylethano-lamine; PC, phosphatidylcholine; PA, phosphatidic acid; CL, cardiolipin. Bars represent arithmetic means of triplicate, plotted along with standarderrors.
Eur. J. Immunol. 2008. 38: 3561–3574 Clinical immunology 3565
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

similarly mutated, its light chain was assigned a value close to
unity, indicating an absence of preferential somatic mutations in
the CDR.
Discussion
In the context of the immune system, the processes of negative
selection in the thymus [18] as well as the killing of virally
infected cells [19] involve activation of the apoptotic cascade.
Normally, apoptotic cells are recognized by professional phago-
cytes and cleared very efficiently. The molecules and processes
involved in the specific recognition by the phagocyte of the
products of cell death have begun to be elucidated [20].
Phagocytosis of apoptotic cells is associated with the regulation
of inflammatory responses [21]. Lupus appears to be character-
ized by excessive apoptosis as well as defective clearance of
apoptotic debris, both of which would be expected to lead to the
accumulation of self-antigens and, possibly as a consequence, an
enhanced susceptibility to autoimmune disease. The specific cell-
surface recognition of ‘‘late-stage’’ apoptotic cells (as defined by
the presence of apoptosis-specific moieties along with demon-
strated retention of plasma membrane integrity) by antibodies
has been previously reported both by us [22, 23] and by
other investigators [16, 24]. In this study, the IgG and IgM
apoptotic cell-specific murine monoclonal antibodies generated
1000.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6A
B
SLE Sera
Normal Sera
1000.0
0.2
0.4
0.6
0.8
1.0SLE Sera
Normal Sera
Ab
sorb
ance
(45
0 n
m)
Sera Dilution
Ab
sorb
ance
(45
0 n
m)
Sera Dilution
1000
1000
Figure 4. (A) Binding of antibodies in sera from SLE patients (dashedlines) and normal subjects (solid lines) to Ro60 by direct-binding ELISA.(B) Competitive ELISA demonstrating the ability of antibodies in serafrom SLE patients (dashed lines), but not from normal subjects (solidlines), to compete with antibody 2C3 for binding to Ro60. The squarerepresents absorbance in the absence of sera.
0
0.4
0.6
0.8
1.0
1.2
1.4A
B
1B3
2C3
1B1
Antibody concentration (µg/ml)
Ph
ago
cyti
c in
dex
00.2
0.4
0.6
0.8
1.0
1.2
1.4 2C11
1B4
2H8
* *
*
**
**
*****
****
*
**
**
****
400300200100
10080604020
Figure 5. Phagocytic uptake by peritoneal macrophages of apoptoticthymocytes in presence of (A) IgG and (B) IgM monoclonal antibodies.The IgG mediated enhanced uptake (Student’s t-test: �po0.05; ��po0.009versus isotype control) while the IgM diminished uptake (Student’s t-test:�po0.05; ��po0.008; ���po0.0009 versus isotype control) of apoptotic cells.The Phagocytic Index, calculated as described in the text, was plotted asfold change over relevant isotype control antibodies, indicated by thehorizontal line with open symbols. In both figures, the diamondrepresents the Phagocytic Index when healthy, non-apoptotic cells wereemployed in the absence of antibodies. Bars represent arithmetic meansof triplicate, plotted along with standard errors.
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3566
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

demonstrated cross-reactivity across several RNP autoantigens.
Extensive cross-reactivity of anti-RNP responses has also
been previously described by other investigators [25]. Cross-
reactivity of autoimmune antibodies across different classes of
molecules has also been reported. For example, antibodies to
dsDNA have been shown to bind phospholipids and peptides
[26, 27]. Antibody 2C11 (IgMk) bound LPC, while antibody 2C3
(IgG2bk) appeared to be more broadly reactive in terms
of anti-lipid specificity. LPC is synthesized and secreted by
apoptotic cells by the action of calcium-independent phosholi-
pase-A2. Lipid extracts obtained from apoptotic thymocytes
contain LPC along with other oxidized phospholipids, and
immunization of mice with such cells generates antibodies that
specifically bind apoptotic cells through recognition of oxidation-
specific epitopes [28]. In certain contexts, LPC has anti-
inflammatory properties [29] and may contribute to the
quiescent milieu of the apoptotic cell-phagocyte environment.
LPC also acts as a chemo-attractant for phagocytes [30], probably
helping them locate dying cells in vivo. Infusion of LPC has
been demonstrated to interfere with the clearance of apoptotic
cells in gld mice [31] and mice deficient in the G2A ‘‘effector’’
exhibit late-onset autoimmunity [32]. Spontaneously arising
autoreactivity against the lipid could thus have far-reaching
consequences.
Sera from lupus-prone mice have been shown to inhibit the
uptake of apoptotic cells [10], a finding of significance, since
apoptotic debris is thought to provide the autoantigenic stimulus
in lupus. IgM antibodies against low-density lipoproteins on
apoptotic cells can inhibit phagocytosis of apoptotic cells [33], as
can an IgM antibody derived from animals immunized with dying
cells [34]. Such studies and others [23, 35, 36] question whether,
in all cases, the uptake of apoptotic cells is strictly PS-dependent
or whether other moieties on dying cells may also contribute.
Antibodies such as IgM 2C11, which binds LPC as well as apop-
totic blebs (also shown to act as chemo-attractants for phago-
cytes) and can also efficiently prevent the phagocytic uptake of
apoptotic cells, could be envisaged to be powerful disease
perpetuators in an environment rendered pro-inflammatory, due
to bacterial infection, for example. The IgG antibodies generated
in this study (2C3, 1B1 and 1B3), in contrast to IgM, enhanced
the internalization of apoptotic cells, possibly by Fc-gR-mediated
mechanisms. IgG anti-phospholipid antibodies that specifically
target apoptotic cells have been shown to increase uptake by
monocytes and induce the release of TNF-a [17]; on the other
hand, IgG antibodies to RNP Ro and La were shown to inhibit
uptake of apoptotic cardiocytes by healthy cardiocytes [37], cells
that do not express Fc receptors. SLE sera as well as small nuclear
RNP-autoantibody and DNA-autoantibody immune complexes
have been shown to stimulate the production of inflammatory
cytokines via the stimulation of Toll-like receptors [38, 39]. In the
present study, in no instances were antibodies found to influence
cytokine production (data not shown). It would be interesting to
attempt correlation between the epitopic specificity of auto-
antibodies directed towards RNP and disease progression and
severity.
Evidence exists implicating the idiotypic network in the
initiation and progression of autoimmune responses. Immuniza-
tion with an anti-dsDNA antibody bearing the 16/6 idiotype led
to the generation of an anti-dsDNA response in non-autoimmune-
prone mice [40]. Immunization with a human monoclonal anti-
body recognizing Ro60 and Ro52 resulted in novel autoantigens
being targeted. Anti-idiotypic antibodies also bound the surface
of healthy cells, unlike the immunizing antibody, which specifi-
cally bound apoptotic cells [23]. We have now addressed this
issue in a system that offers an increased level of stringency;
immunization of syngeneic mice with murine antibodies would
be expected to generate primarily an anti-idiotypic response,
unlike immunization of animals with human antibodies, which
necessitates extensive absorptions to rid the elicited antibodies of
isotypic and allotypic reactivity. Antibodies 1B1 (IgG2ak) and
00
100
200
300
400
500
6002C11Isotype Control
Tran
smig
rati
ng
Cel
ls (
x 10
-2)
***
B
Antibody Concentration (µg/ml)
A
0
200
400
600
80α 0
1000
1200
1400
1600
LPS + + + + LPC --
- -- -
-+ ++
Ab. - 2C11 1B4 - - 2C11 1B4
*
8642
Figure 6. (A) Influence of antibody 2C11 or antibody 1B4 (employed asan isotype control) upon the down-modulatory effects of LPC on LPS-induced TNF-a secretion (Student’s t-test: �po0.001 antibody 2C11versus Antibody 1B4). Antibodies were used at a concentration of20 mg/well. Ab.: antibodies. (B) Effects of varying concentrations ofantibody 2C11 (filled circles) and an isotype control antibody (anti-TNP(IgMk); open circles) on the transmigration of THP-1 cells towardssupernatant derived from apoptotic cells (Student’s t-test: �po0.007;��po0.003 antibody 2C11 versus isotype control). The filled squarerepresents transmigration of THP-1 cells towards supernatant derivedfrom healthy cells. Bars represent arithmetic means of triplicate,plotted along with standard errors.
Eur. J. Immunol. 2008. 38: 3561–3574 Clinical immunology 3567
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

2C11 (IgMk) were both capable of evoking anti-self-responses
and diversification in the repertoire. The ELISA data on select
recombinant proteins are only indicative in nature, and more
comprehensive antigenic analysis using anti-idiotypic sera would
obviously be more revealing. Nevertheless, the results provide
evidence that, in a syngeneic system, the idiotypic network can
mediate epitope spreading in anti-RNP responses. One way in
which this may occur can be postulated. Antibodies reactive
towards one component could conceivably generate ‘‘internal
image’’ anti-idiotypic antibodies that bind to other component
molecule(s) of a macromolecular complex to which the inciting
antigen belongs. While such a scenario is possible, all specificities
that arise in experiments such as these cannot be predicted a
priori, on the basis of knowledge of known interactions.
While the anti-1B1, anti-idiotypic specificity could conceivably be
predicted (based on the known associations of Ro60 with La and
of SmD with the U1-RNP complex), such forecasts for the anti-
2C11, anti-idiotypic specificity would be erroneous. Unexpected
specificities also arise upon immunization with anti-DNA anti-
bodies [40]. It is conceivable that unappreciated (and currently
unpredictable) levels of idiotypic connectivity exist amongst
autoimmune antibody responses. Anti-idiotype-mediated epitope
spreading is clearly not a general phenomenon, however, as
immunization with antibody 2C3 (IgG2bk) does not result in
IgGIgM
Adjuvant
1B1 Anti-IdB
A
0.0
0.2
0.4
0.6
0.8Ro52
Ro60
SmB
SmD
La
U1-RNP A
Ab
sorb
ance
(45
0 n
m)
Eve
nts
FL
1B1
Figure 7. (A) Assessment of IgM (left panel) and IgG (right panel) autoreactivity of sera isolated from NZB/W animals immunized with antibody 1B1(solid, thick profiles) or adjuvant (dashed profiles) by FACS analysis. Solid, thin profiles depict negative controls, staining obtained when just thesecond antibody was employed. FL: log fluorescence. (B) Top left: reactivity of antibody 1B1 upon permeabilized U-87 cells by immunofluorescence.Top right: reactivity of anti-1B1 sera. Bottom left: reactivity of adjuvant-induced sera. Bottom right: differential reactivity of anti-1B1 sera towards apanel of recombinant antigens by ELISA. Results are plotted after respective reactivity obtained upon immunization with adjuvant alone has beensubtracted, and so represent anti-idiotypic responses. Bars represent arithmetic means of triplicate, plotted along with standard errors.
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3568
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Figure 8. Heavy and light chain variable region sequences for the IgM antibodies (A) 2C11, (B) 2H8 and (C) 1B4. FWR and CDR are indicated.Homologies to respective closest germline genes are indicated as dots. Amino acid mutations are indicated in lower case. Underlines indicate non-encoded amino acids.
Figure 9. Heavy and light chain variable region sequences for the IgG antibodies (A) 2C3, (B) 1B3 and (C) 1B1. FWR and CDR are indicated.Homologies to respective closest germline genes are indicated as dots. Amino acid mutations are indicated in lower case. Underlines indicate non-encoded amino acids.
Eur. J. Immunol. 2008. 38: 3561–3574 Clinical immunology 3569
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
anti-self-reactivity (data not shown). Further work, employing
antibodies of differing specificity and idiotypes, would be
required for a clearer appreciation of the rules that govern such
responses.
Distinct phagocytic outcomes can result, depending on the
nature of the targeted apoptotic cell-associated molecular
pattern, the nature of the phagocyte and the isotype of the
autoantibody, as this study and work by other investigators
suggest. Apoptotic cell-reactive antibodies can sometimes mani-
fest characteristics of an antigen-driven response, while still
demonstrating a variable degree of intermolecular cross-reactiv-
ity stretching across proteinic and lipidic molecular moieties.
While the basis of this cross-reactivity will form the focus of
future investigations, the current studies elaborate upon two
potential consequences of anti-lipid responses, the reversal of the
anti-inflammatory effects of LPC and the interruption of an
LPC-mediated phagocyte chemo-attractive signal, both of
which are clearly relevant in the context of systemic
autoimmunity. These studies also shed new light upon the
phenomenon of anti-idiotype-mediated determinant spreading;
the phenomenon is not dependent on antibody isotype or the
presence of somatic mutations in the CDR but is nonetheless not
universal. A more detailed analysis of the antibodies that bind
dying cells is clearly warranted in order to further understand the
consequences of the recognition of cell death in systemic auto-
immune disease.
Materials and methods
Human subjects
All procedures were reviewed and approved by Institutional
Ethics Committees and informed consent obtained.
SLE patients were on clinical followup at the Rheumatology
Clinic at the Department of Medicine at the All India Institute of
Medical Sciences, New Delhi. All subjects satisfied the revised
criteria set forth by the American College of Rheumatology for
the diagnosis of SLE and were evaluated and classified on the
basis of their SLE Disease Activity Index. Blood samples were
withdrawn from patients deemed to be in the midst of a disease
flare, as determined by clinical presentation. Control sera
samples were isolated from normal, healthy volunteers.
Generation of apoptotic cell-reactive murinemonoclonal antibodies
All procedures were reviewed and approved by Institutional
Ethics Committees.
All cultures were maintained in DMEM (Gibco BRL) supple-
mented with 10% FCS (Biological Industries). Cells were grown in
a humidified incubator at 371C in a 5% CO2 atmosphere. C57BL/
6lpr/lpr and NZB � NZW (F1) (hereafter referred to as NZB/W)
animals were maintained at the animal facility of the National
Institute of Immunology, New Delhi. Hybridomas were generated
by fusion of splenocytes (sourced from aging animals demon-
strating high serum anti-nuclear antibody titers) with SP2/0
myeloma cells in the presence of 50% v/v polyethylene glycol 1500
(Sigma). Hybrids were selected in hypoxanthene-, aminopterin-
and thymidine-supplemented medium for 7 days, and cells were
cultured for an additional week in medium supplemented with
hypoxanthene and thymidine. A total of 465 independent hybri-
domas were generated for this study. As an initial assessment of
reactivity, recognition of either healthy (non-permeable and non-
apoptotic) cells, as well as of permeabilized cells, was assessed by
flow cytometry; 32 hybridomas secreted antibodies reactive to the
surface of healthy cells, and 64 secreted antibodies that bound
permeabilized cells. Of the latter, 58 did not recognize apoptotic
cells (reflective of anti-self-reactivity towards nuclear or cyto-
plasmic moieties not necessarily involved in, or affected by, the
apoptotic cascade), while the six antibodies discussed in this paper
bound cells in the late stages of apoptosis.
Hybridoma cells secreting antibodies of interest were
subcloned at least three times by limiting dilution to
ensure monoclonality.
Induction of apoptosis
Human Jurkat T cells or murine SP2/O cells were exposed to
UV light (312 nm; 100 mJ/cm2), whereas murine thymocytes
J28-19JH2U.I.VH7183.101B1
J2ba9JH4DSP2.2J558.26.1161B3
J2bl1JH4DSP2.5VH7183.3b2C3
J423-48JH4DSP2.2J558.84.1901B4
J519-23JH3U.I.J558.67.1662H8
J1cp9JH1DFL16.1VH9.152C11
JLVLJHDHVHA
B
IgM
sIg
Gs
0
2
4
6 Heavy ChainLight Chain
IgMs IgGs2H8 *
CD
R/F
WR
Rep
lace
men
tM
uta
tio
n F
req
uen
cy
1B11B3 2C31B4 2C11
Figure 10. (A) Description of antibody germline segment usage (VH,DH and JH regions for heavy chains and VL, JL regions for light chains).U.I.: unidentifiable. (B) Ratios of CDR to FWR replacement mutationfrequencies in the six antibodies. � indicates a divisor of zero. See textfor details.
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3570
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

were cultured in medium supplemented with 10 mM dexametha-
sone. In both instances, cells were then incubated for
different periods of time. At the time of analysis, apoptotic cells
retained the capacity to exclude the vital dye Trypan
Blue, indicating retention of plasma membrane integrity.
To establish the dependence of antibody binding upon
caspase activity, apoptosis was induced in the presence of
25 mM z-VAD-fmk, a pan-caspase inhibitor.
Antigenic reactivity
Flow cytometry and confocal analysis
All incubations were carried out in FACS buffer (PBS
containing 1% BSA and 0.2% sodium azide) for 1 h at 41C.
Healthy, apoptotic or permeabilized cells (obtained by
brief incubation in chilled methanol containing 0.001% Triton-
X-100) were incubated with antibodies and subsequently
washed to remove unbound antibody by brief centrifugations
at 300g. Fluorochrome-labeled secondary antibodies were
then added at the recommended dilution and another incubation
carried out. After further washes, cells were resuspended
in PBS and analyzed on a Becton Dickinson LSR flowcyto-
meter. To assess exposure of PS, antibody-stained cells
were first resuspended in Annexin-binding buffer (10 mM
Hepes, 140 mM NaCl and 2.5 mM CaCl2) followed by incubation
at room temperature for 20 min with an Annexin-V–fluorochrome
conjugate.
For confocal microscopy, apoptotic Jurkat T were fixed in
freshly prepared ice cold 2% paraformaldehyde in PBS for
10 min, followed by incubation in PBS supplemented with 1%
BSA for 15 min. Cells were then processed as described for flow
cytometry.
Western blot
Apoptotic blebs were prepared by differential centrifugation.
Briefly, apoptosis was induced in SP2/0 cells by UV light
as described under "Induction of apoptosis". Cells were
centrifuged at 400g for 5 min at 41C and the supernatant
re-centrifuged at 1000g for 10 min. The resulting supernatant
was further centrifuged at 17 000g for 30 min. The pellet
contained apoptotic blebs of 6–8 mm, as assessed by scanning
electron microscopy. Blebs (5mg protein) were incubated
with antibody 2C11 for 2 h at 371C and then subjected to
extensive washing to remove unbound antibody. SDS-PAGE
and Western blot were carried out, and the presence of
bleb-bound antibody was revealed by an anti-mouse-Ig-HRP-
secondary antibody conjugate. Additionally, blebs were isolated
from apoptotic 2C11 hybridoma cells and the presence of
endogenous, adhered antibody 2C11 was assessed by SDS-PAGE
and Western blot. In this case, blebs isolated from apoptotic
SP2/0 cells, which were not expected to contain adhered Ig,
served as negative controls.
ELISA
Recombinant autoantigens (Ro52, Ro60, SmB, SmD, La and
U1-RNP A Protein, obtained from Dr. Shu Man Fu, University of
Virginia, USA) were diluted in 0.1 M carbonate buffer, pH 9.2.
Fifty microliters of buffer containing 500 ng antigen
was dispensed in each well of a 96-well ELISA plate (Nunc).
Plates were incubated overnight at 41C. After wells were
washed with 10 mM PBS (pH 7.4) containing 0.05% Tween 20,
un-occupied binding sites were ‘‘blocked’’ by incubation with
200mL of 1% BSA prepared in PBS; this and all subsequent
incubations were at 371C for 2 h; 50 mL of primary antibodies
(murine monoclonal antibodies, SLE sera or normal
sera), appropriately diluted in PBS with Tween 20 containing
1% BSA, were dispensed into the wells and a further incubation
carried out. After further washes, 50 mL of diluted secondary
antibody was added to each well. Following another incubation,
wells were extensively washed. Enzymatic reactivity was
visualized with a tetramethyl benzidine, tetramethyl ammonium
borohydride and N,N-dimethyl acetamide containing substrate
solution. Absorbance was recorded at 450 nm. For competition
assays, diluted SLE sera or normal sera were pre-incubated in
antigen-adsorbed wells for 30 min followed by addition
of the monoclonal antibodies. Incubation was carried on for
another 90 min and reactivity was assessed following incubation
with a goat anti-mouse IgG-HRP conjugate (Jackson Immuno-
Research).
The following phospholipids were obtained from
Avanti Polar Lipids: LPC, lysophosphatidic acid (LPA), PS, phos-
phatidylethanolamine (PE), phosphatidylcholine (PC), phospha-
tidic acid (PA) and cardiolipin (CL). Phospholipids were
resuspended in 1:4 chloroform:methanol at a concentration of
50 mg/mL; 30 mL of this solution was dispensed into wells of
a 96-well ELISA plate. Evaporation of the solvent occurred during
incubation at 16 h at 41C. Un-occupied binding sites were
‘‘blocked’’ by incubation with PBS containing 5% low-fat milk
powder or 0.1% polyvinyl alcohol. Other procedures were as
mentioned for recombinant autoantigens.
Negative controls for all assays included antigen-blank
wells, and wells in which only antigen and second antibodies
were employed. Highest absorbances (which typically
ranged from 0.05 to 0.08) were subtracted.
Phagocytosis
Murine thymocytes were incubated with 10 nM CFSE diacetate
salt mixed isomer (Molecular Probes) for 20 min at 371C. Cells
were washed twice with RPMI-1640 and apoptosis was induced
by the addition of dexamethasone as described under ‘‘Induction
of apoptosis’’.
C57BL/6 mice were injected intraperitoneally with 1 mL of a
4% thioglycolate solution. Peritoneal exudates cells were
‘‘tapped’’ at 72 h. Cells were resuspended in DMEM and allowed
to adhere to tissue culture plates for 2 h. Non-adherent cells were
Eur. J. Immunol. 2008. 38: 3561–3574 Clinical immunology 3571
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

removed by repeated flushing. CFSE-labeled apoptotic thymo-
cytes were added to adhered phagocytes at a ratio of 1:4
(phagocytes:apoptotic cells) in the presence of varying concen-
trations of antibodies. An IgMk antibody monoclonal antibody
recognizing the Vi molecule of Salmonella typhi was employed as
the negative control for Antibodies 2C11, 2H8 and 1B4. Antibody
1D1 (an IgG3k monoclonal antibody generated in the laboratory,
which is reactive to cell-surface moieties upon healthy cells) was
employed as negative control for Antibodies 2C3, 1B3 and 1B1.
Cultures were incubated for 2 h at 371C and then transferred to
41C to stop further uptake. In certain instances, supernatants
were stored for subsequent cytokine analysis. Non-phagocytosed
thymocytes were removed by extensive ‘‘flushing’’. Cells were
stained with an anti-Mac1-phycoerythrin conjugate (PE; BD
Pharmingen) to label the macrophage population. FACS
analysis was then carried out. The Phagocytic Index was assessed
as the number of dually stained cells (macrophages (red) that
had ingested apoptotic cells (green)) over total number of
macrophages (red).
Inflammation
One of the apoptotic cell-specific antibodies (antibody 2C11
(IgMk)) was reactive towards LPC, a bioactive lipid that exhibits
both anti-inflammatory [29] and pro-inflammatory [41] proper-
ties. To delineate the anti-inflammatory effects of LPC in initial
experiments, thioglycolate-elicited peritoneal cells were cultured
in X-Vivo 10 serum-free media and allowed to adhere for 2–3 h at
371C; 100 ng/mL of S. typhi LPS (Sigma) was added to the cells,
along with varying concentrations of LPC or PC. An incubation
was then carried out for 4 h at 371C. LPC, but not PC, caused a
dose-dependent decrease in the secretion of TNF-a (data not
shown), estimated by ELISA (BD OptEIA). To assess the potential
of antibody 2C11 (as opposed to the apoptotic cell-specific but
LPC non-reactive antibody 1B4 (IgMk)) to revert the anti-
inflammatory effects of LPC, 20 mg of the respective antibodies
were added to cultures containing cells, LPS (100 ng/mL) and
LPC (50 mM), and TNF-a was estimated by ELISA.
Transmigration
Apoptotic cells are known to secrete LPC, which, in addition to its
other biological activities, also acts as a chemo-attractant for
phagocytic cells [30]. It is also believed that small membranous
vesicles released by apoptotic cells may have similar properties.
Since the antibody 2C11 bound both LPC and apoptotic
blebs, the ability of the antibody to inhibit the transmigration
of THP-1 cells towards supernatant obtained from apoptotic cell
cultures was assessed. An anti-TNP antibody (IgMk; BD
Pharmingen) was employed as the isotype control. Apoptosis
was induced in Jurkat cells as described under ‘‘Induction of
apoptosis’’. Supernatants were harvested by centrifugation at
1200g for 10 min; 200mL of apoptotic cell-culture supernatant
(either non-supplemented or supplemented with varying concen-
tration of antibody 2C11 or the isotype control antibody) was
added to the lower chamber of transmigration wells. THP-1 cells
(50 mL of a suspension of 2�106 cells/mL) were added to
transwell inserts (8 mm pore size; BD Falcon) and an incubation
carried out for 90 min under standard culture conditions.
Transmigration was assessed by counting the cells harvested
from the lower chamber.
Anti-idiotypic responses
Antibody 2C11 (IgMk) was purified by affinity chromatography
on UltraLink Immobilized Mannan Binding Protein (Pierce).
antibody 1B1 (IgG2ak) and antibody 2C3 (IgG2bk) were purified
using Protein-G Sepharose affinity chromatography (GE Health-
care). Six-wk-old animals (C57BL/6 for Antibodies 2C11, and
NZB/W for Antibodies 1B1 and 2C3) received subcutaneous
injections of 50 mg antibody emulsified in Incomplete Freund’s
Adjuvant; each animal received 100mL at two sites. Two booster
injections were administered at weekly intervals and blood
samples were collected 1 wk after each booster. Anti-self-
reactivity in the sera was assessed by FACS analysis on
permeabilized and non-permeabilized Jurkat cells, by indirect
immunofluorescence assays on HeLa cells and by ELISA upon a
panel of recombinant autoantigens.
Antibody variable region genes
RNA was isolated from extensively subcloned hybridoma cultures
by the TRIzol (Invitrogen) method. cDNA was prepared using
oligo dT primers and avian myeloblastosis virus reverse tran-
scriptase (Promega). RNA and the oligo dT primer (Promega)
were incubated at 701C for 10 min followed by instantaneous
cooling on ice. Additional pre-mixed components were added and
the mixture incubated on a thermal cycler (Perkin Elmer) for
60 min at 481C. Reverse transcriptase was inactivated by heating
to 701C for 5 min. The cDNA preparation was treated with RNase
H (Promega) at 371C for 30 min to degrade residual RNA. A
mouse Ig kit (Novagen) containing primers for the murine light
and heavy chain genes was employed. Forward and reverse
primers (5 pM) were added in a total reaction volume
of 25 mL. PCR amplification was then carried out (30 cycles;
5 min hotstart, 45 s denaturation at 941C, 45 s annealing at 601C).
Extension was carried out for 45 s at 721C; a final extension for
7 min was carried out at 721C. After electrophoresis on 1%
agarose, the PCR product was purified using the Qiagen gel
extraction kit and ligated to the pGEMT vector (Promega). The
DH5a strain of Escherichia coli was transformed with the ligated
plasmid using standard protocols. The transformed cells were
selected on LB agar supplemented with ampicillin (100 mg/mL),
X-gal and iso-propyl thio-galactopyranose. Plasmids were isolated
and cloned antibody gene inserts were sequenced by using M13
(forward and reverse) primers, corresponding to sequences
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3572
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

adjacent to the multiple cloning site of the pGEMT vector.
Automated sequencing was carried out at the Centralized
Sequencing Facility of the National Institute of Immunology.
Analysis of antibody variable region gene sequences
was carried out using the NCBI and IMGT germline databases.
Sequences were submitted to GenBank (Accession Numbers
EF063577–EF063588; http://www.ncbi.nlm.nih.gov/Genbank).
Mutation frequencies (cumulative replacement mutations per
unit amino acid length) were calculated for the CDR and FRW
regions; in order to obtain a conservative estimate, amino acids
not unequivocally assignable to designated gene segments were
not included in the analysis.
Acknowledgements: This study was supported by core and
extramural grants from the Department of Biotechnology,
Government of India to R. P.
Conflict of interest: The authors have declared no financial or
commercial conflict of interest.
References
1 Kotzin, B. L., Systemic lupus erythematosus. Cell 1996. 85: 303–306.
2 Wakeland, E. K., Liu, K., Graham, R. R. and Behrens, T. W., Delineating
the genetic basis of systemic lupus erythematosus. Immunity 2001. 15:
397–408.
3 Singh, R. R., SLE: translating lessons from model systems to human
disease. Trends Immunol. 2005. 26: 572–579.
4 Suzuki, N., Harada, T., Mizushima, Y. and Sakane, T., Possible pathogenic
role of cationic anti-DNA autoantibodies in the development of nephritis
in patients with systemic lupus erythematosus. J. Immunol. 1993. 151:
1128–1136.
5 Tran, H. B., Macardle, P. J., Hiscock, J., Cavill, D., Bradley, J., Buyon, J. P. and
Gordon, T. P., Anti-La/SSB antibodies transported across the placenta
bind apoptotic cells in fetal organs targeted in neonatal lupus. Arthritis
Rheum. 2002. 46: 1572–1579.
6 Shoenfeld, Y., Etiology and pathogenetic mechanisms of the anti-
phospholipid syndrome unraveled. Trends Immunol. 2003. 24: 2–4.
7 Edwards, B. S., Searles, R. P., Brozek, C. M., Richards, R., Savage, S. M.,
Nolla, H. and Hoffman, C. L., Isotype and cytotoxicity spectra of anti-
lymphocyte antibodies in patients with systemic lupus erythematosus.
Clin. Immunol. Immunopathol. 1987. 45: 333–347.
8 Yoshio, T., Hirata, D., Onda, K., Nara, H. and Minota, S., Antiribosomal
P-protein antibodies in cerebrospinal fluid are associated with neurop-
sychiatric systemic lupus erythematosus. J. Rheumatol. 2005. 32: 34–39.
9 Jin, O., Sun, L. Y., Zhou, K. X., Zhang, X. S., Feng, X. B., Mok, M. Y. and
Lau, C. S., Lymphocyte apoptosis and macrophage function: correlation
with disease activity in systemic lupus erythematosus. Clin. Rheumatol.
2005. 24: 107–110.
10 Licht, R., Dieker, J. W., Jacobs, C. W., Tax, W. J. and Berden, J. H.,
Decreased phagocytosis of apoptotic cells in diseased SLE mice. J.
Autoimmun. 2004. 22: 139–145.
11 Mevorach, D., Zhou, J. L., Song, X. and Elkon, K. B., Systemic exposure to
irradiated apoptotic cells induces autoantibody production. J. Exp. Med.
1998. 188: 387–392.
12 Cohen, P. L., Caricchio, R., Abraham, V., Camenisch, T. D., Jennette, J. C.,
Roubey, R. A., Earp, H. S. et al., Delayed apoptotic cell clearance and
lupus-like autoimmunity in mice lacking the c-mer membrane tyrosine
kinase. J. Exp. Med. 2002. 196: 135–140.
13 Hanayama, R., Tanaka, M., Miyasaka, K., Aozasa, K., Koike, M.,
Uchiyama, Y. and Nagata, S., Autoimmune disease and impaired uptake
of apoptotic cells in MFG-E8-deficient mice. Science 2004. 304: 1147–1150.
14 Fadok, V. A., Voelker, D. R., Campbell, P. A., Cohen, J. J., Bratton, D. L. and
Henson, P. M., Exposure of phosphatidylserine on the surface of apoptotic
lymphocytes triggers specific recognition and removal by macrophages.
J. Immunol. 1992. 148: 2207–2216.
15 Casciola-Rosen, L. A., Anhalt, G. and Rosen, A., Autoantigens targeted in
systemic lupus erythematosus are clustered in two populations of
surface structures on apoptotic keratinocytes. J. Exp. Med. 1994. 179:
1317–1330.
16 Radic, M., Marion, T. and Monestier, M., Nucleosomes are exposed at the
cell surface in apoptosis. J. Immunol. 2004. 172: 6692–6700.
17 Manfredi, A. A., Rovere, P., Galati, G., Heltai, S., Bozzolo, E., Soldini, L.,
Davoust, J. et al., Apoptotic cell clearance in systemic lupus erythema-
tosus. I. Opsonization by antiphospholipid antibodies. Arthritis Rheum.
1998. 41: 205–214.
18 Kyewski, B. and Klein, L., A central role for central tolerance. Annu. Rev.
Immunol. 2006. 24: 571–606.
19 Russell, J. H. and Ley, T. J., Lymphocyte-mediated cytotoxicity. Annu. Rev.
Immunol. 2002. 20: 323–370.
20 Henson, P. M. and Hume, D. A., Apoptotic cell removal in development
and tissue homeostasis. Trends Immunol. 2006. 27: 244–250.
21 Cvetanovic, M. and Ucker, D. S., Innate immune discrimination
of apoptotic cells: repression of proinflammatory macrophage transcrip-
tion is coupled directly to specific recognition. J. Immunol. 2004. 172:
880–889.
22 Pal, R., Deshmukh, U. S., Ohyama, Y., Fang, Q., Kannapell, C. C.,
Gaskin, F. and Fu, S. M., Evidence for multiple shared antigenic
determinants within Ro60 and other lupus-related ribonucleoprotein
autoantigens in human autoimmune responses. J. Immunol. 2005. 175:
7669–7677.
23 Gandhi, R., Hussain, E., Das, J., Handa, R. and Pal, R., Anti-idiotype-
mediated epitope spreading and diminished phagocytosis by a human
monoclonal antibody recognizing late-stage apoptotic cells. Cell Death
Differ. 2006. 13: 1715–1726.
24 Dransfield, I., Rossi, A. G., Brown, S. B. and Hart, S. P., Neutrophils:
dead or effete? Cell surface phenotype and implications for phagocytic
clearance. Cell Death Differ. 2005. 12: 1363–1367.
25 Deshmukh, U. S., Kannapell, C. C. and Fu, S. M., Immune responses to
small nuclear ribonucleoproteins: antigen-dependent distinct B cell
epitope spreading patterns in mice immunized with recombinant
polypeptides of small nuclear ribonucleoproteins. J. Immunol. 2002. 168:
5326–5332.
26 Lafer, E. M., Rauch, J., Andrzejewski, C., Jr., Mudd, D., Furie, B., Furie, B.,
Schwartz, R. S. et al., Polyspecific monoclonal lupus autoantibodies
reactive with both polynucleotides and phospholipids. J. Exp. Med. 1981.
153: 897–909.
27 Putterman, C. and Diamond, B., Immunization with a peptide surrogate
for double-stranded DNA (dsDNA) induces autoantibody production and
renal immunoglobulin deposition. J. Exp. Med. 1998. 188: 29–38.
Eur. J. Immunol. 2008. 38: 3561–3574 Clinical immunology 3573
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

28 Chang, M. K., Binder, C. J., Miller, Y. I., Subbanagounder, G., Silverman, G.
J., Berliner, J. A. and Witztum, J. L., Apoptotic cells with oxidation-specific
epitopes are immunogenic and proinflammatory. J. Exp. Med. 2004. 200:
1359–1370.
29 Yan, J. J., Jung, J. S., Lee, J. E., Lee, J., Huh, S. O., Kim, H. S., Jung, K. C. et al.,
Therapeutic effects of lysophosphatidylcholine in experimental sepsis.
Nat. Med. 2004. 10: 161–167.
30 Lauber, K., Bohn, E., Krober, S. M., Xiao, Y. J., Blumenthal, S. G.,
Lindemann, R. K., Marini, P. et al., Apoptotic cells induce migration of
phagocytes via caspase-3-mediated release of a lipid attraction signal.
Cell 2003, 3113: 717–730.
31 Aprahamian, T., Rifkin, I., Bonegio, R., Hugel, B., Freyssinet, J. M., Sato, K.,
Castellot, J. J. Jr. and Walsh, K., Impaired clearance of apoptotic cells
promotes synergy between atherogenesis and autoimmune disease. J.
Exp. Med. 2004. 199: 1121–1131.
32 Le, L. Q., Kabarowski, J. H., Weng, Z., Satterthwaite, A. B., Harvill, E. T.,
Jensen, E. R., Miller, J. F. and Witte, O. N., Mice lacking the orphan G
protein-coupled receptor G2A develop a late-onset autoimmune
syndrome. Immunity 2001. 14: 561–571.
33 Chang, M. K., Bergmark, C., Laurila, A., Horkko, S., Han, K. H., Friedman,
P., Dennis, E. A. and Witztum, J. L., Monoclonal antibodies against
oxidized low-density lipoprotein bind to apoptotic cells and inhibit their
phagocytosis by elicited macrophages: evidence that oxidation-specific
epitopes mediate macrophage recognition. Proc. Natl. Acad. Sci. USA 1999.
96: 6353–6358.
34 Fujii, C., Shiratsuchi, A., Manaka, J., Yonehara, S. and Nakanishi, Y.,
Difference in the way of macrophage recognition of target cells
depending on their apoptotic states. Cell Death Differ. 2001. 8: 1113–1122.
35 Gardai, S. J., McPhillips, K. A., Frasch, S. C., Janssen, W. J., Starefeldt, A.,
Murphy-Ullrich, J. E., Bratton, D. L. et al., Cell-surface calreticulin
initiates clearance of viable or apoptotic cells through trans-activation
of LRP on the phagocyte. Cell 2005. 123: 321–334.
36 Guzik, K., Bzowska, M., Smagur, J., Krupa, O., Sieprawska, M., Travis, J.
and Potempa, J., A new insight into phagocytosis of apoptotic cells:
proteolytic enzymes divert the recognition and clearance of polymorpho-
nuclear leukocytes by macrophages. Cell Death Differ. 2007. 14: 171–182.
37 Clancy, R. M., Neufing, P. J., Zheng, P., O’mahony, M., Nimmerjahn, F.,
Gordon, T. P. and Buyon, J. P., Impaired clearance of apoptotic cardiocytes
is linked to anti-SSA/Ro and SSB/La antibodies in the pathogenesis of
congenital heart block. J. Clin. Invest. 2006. 116: 2413–2422.
38 Means, T. K., Latz, E., Hayashi, F., Murali, M. R., Golenbock, D. T. and
Luster, A. D., Human lupus autoantibody-DNA complexes activate DCs
through cooperation of CD32 and TLR9. J. Clin. Invest. 2005. 115:
407–417.
39 Savarese, E., Chae, O. W., Trowitzsch, S., Weber, G., Kastner, B., Akira, S.,
Wagner, H. et al., U1 small nuclear ribonucleoprotein immune complexes
induce type I interferon in plasmacytoid dendritic cells through TLR7.
Blood 2006. 107: 3229–3234.
40 Mendlovic, S., Brocke, S., Shoenfeld, Y., Ben-Bassat, M., Meshorer, A.,
Bakimer, R. and Mozes, E., Induction of a systemic lupus erythematosus-
like disease in mice by a common human anti-DNA idiotype. Proc. Natl.
Acad. Sci. USA 1988. 85: 2260–2264.
41 Aiyar, N., Disa, J., Ao, Z., Ju, H., Nerurkar, S., Willette, R. N., Macphee, C.
H. et al., Lysophosphatidylcholine induces inflammatory activation of
human coronary artery smooth muscle cells. Mol. Cell. Biochem. 2007. 295:
113–120.
Abbreviations: LPC: lysophosphatidylcholine � PC: phosphatidylcholine
� PE: phosphatidylethanolamine � PS: phosphatidylserine � RNP:
ribonucleoprotein � SLE: systemic lupus erythematosus
Full correspondence: Dr. Rahul Pal, Immunoendocrinology Laboratory,
National Institute of Immunology, Aruna Asaf Ali Marg, JNU Complex,
New Delhi, India
Fax: 191-11-26742125
e-mail: [email protected]
Received: 18/6/2008
Revised: 27/8/2008
Accepted: 29/9/2008
Eur. J. Immunol. 2008. 38: 3561–3574Joy Das et al.3574
& 2008 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu













![NK Cells Mediate Increase of Phagocytic Activity but Not of Proinflammatory Cytokine (Interleukin6 [IL6], Tumor Necrosis Factor Alpha, and IL12) Production Elicited in Splenic Macrophages](https://static.fdokumen.com/doc/165x107/632147240c12e1161503b7d3/nk-cells-mediate-increase-of-phagocytic-activity-but-not-of-proinflammatory-cytokine.jpg)






