
Therapeutic B cell depletion impairs adaptive and autoreactive CD4+ T cell activation in mice
Upload
independentCategory
view
5download
0
doi:10.1182/blood-2009-09-243071Prepublished online March 15, 2010;
Jonathan Samuels, Jessica Berman, Jane H. Buckner, Charlotte Cunningham-Rundles and Eric MeffreIsabelle Isnardi, Yen-Shing Ng, Laurence Menard, Greta Meyers, David Saadoun, Iva Srdanovic, autoreactive unresponsive clonesComplement receptor 2/CD21-negative human naive B cells mostly contain
(4607 articles)Immunobiology �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

1
Complement receptor 2/CD21-negative human naive B cells mostly contain
autoreactive unresponsive clones
Isabelle Isnardi1, Yen-Shing Ng1,2, Laurence Menard1,2, Greta Meyers1,2, David
Saadoun1, Iva Srdanovic1, Jonathan Samuels1,3, Jessica Berman4, Jane H. Buckner5,
Charlotte Cunningham-Rundles6, and Eric Meffre1,2*
1Laboratory of Molecular Immunology, Hospital for Special Surgery, New York, NY
10021, USA, 2Current address: Department of Immunobiology, Yale University School of
Medicine, New Haven, CT 06509, USA, 3Current address: Division of Rheumatology,
Hospital for Joint Diseases, NYU Langone Medical Center, New York, NY 10003, USA;
4Division of Rheumatology, Hospital for Special Surgery, New York, NY 10021, USA,
5Translational Research Program, Benaroya Research Institute, Seattle WA 98105,
USA, 6Department of Medicine and Pediatrics, Mount Sinai Medical Center, New York,
NY 10029, USA.
* Correspondence : [email protected]
Short Title: Characterization of human anergic B cells
Blood First Edition Paper, prepublished online March 15, 2010; DOI 10.1182/blood-2009-09-243071
Copyright © 2010 American Society of Hematology
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

2
Abstract
Complement receptor 2 (CR2/CD21)-negative B cells have been found enriched in
patients with autoimmune diseases and in common variable immunodeficiency (CVID)
patients who are prone to autoimmunity. However, the physiology of CD21-/lo B cells
remains poorly characterized. We found that some rheumatoid arthritis (RA) patients
also display an increased frequency of CD21-/lo B cells in their blood. A majority of CD21-
/lo B cells from RA and CVID patients expressed germline autoreactive antibodies, which
recognized nuclear and cytoplasmic structures. In addition, these B cells were unable to
induce calcium flux, become activated or proliferate in response to B cell receptor and/or
CD40 triggering, suggesting that these autoreactive B cells may be anergic. Moreover,
gene array analyses of CD21-/lo B cells revealed molecules specifically expressed in
these B cells and which are likely to induce their unresponsive stage. Thus, CD21-/lo B
cells mostly contain autoreactive unresponsive clones, which express a specific set of
molecules that may represent new biomarkers to identify anergic B cells in humans.
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

3
Introduction
Random V(D)J recombination generates a large number of autoreactive B cells, which
can be silenced in the bone marrow by three main tolerance mechanisms: deletion,
receptor editing, and anergy 1-3. Deletion results in the removal of autoreactive clones by
apoptosis whereas receptor editing allows autoreactive B cells to alter their self-reactive
B cell receptor (BCR). This process may rescue immature B cell clones from deletion
and allow their differentiation to resume. In contrast to deletion and receptor editing,
anergy does not remove autoreactive B cell clones from the total B cell population but
renders them irresponsive to antigenic stimulation 4-7. Anergic autoreactive B cells
remain in the periphery but they have a short life span, which ultimately result in their
elimination 8,9.
Initial reports have demonstrated that deletion is mainly used to eliminate B cells, which
express highly autoreactive BCRs against membrane-bound antigens 10,11. However,
receptor editing has since been shown to be the major B cell tolerance mechanism
against these antigens, and clonal deletion appears to be a default mechanism when
receptor editing fails to silence autoreactive B cells 12. Alternatively, anergy seems to be
preferentially induced in B cells that express moderately autoreactive BCRs toward
soluble antigens 11. Using transgenic mouse models, anergic B cells have been
described as being unable to become activated, proliferate or secrete antibodies upon
BCR triggering (reviewed in 7). Indeed, BCR signaling is abnormal in these cells and
BCR aggregation fails to induce an increased concentration of intracellular calcium
(Ca2+)i or tyrosine phosphorylation cascades. It is believed that this irresponsive state
results from chronic BCR exposure to self-antigens, which desensitizes BCR signaling
abilities 13,14. The characterization of unresponsive B cells in unmanipulated mice and in
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

4
humans showed that anergic B cells represent a small percentage of circulating B cells
15,16.
We report here that an unusual B cell population, which downregulate the complement
receptor CR2/CD21 and was previously reported in SLE and common variable
immunodeficiency disease (CVID) patients, develops in some RA patients 17,18 ,19-21,22 ,23.
These CD21-/lo B cells are enriched in autoreactive clones that are refractory to most
stimulation, suggesting that human CD21-/lo B cells use an anergic mechanism to be
tolerized.
Methods
Patients and healthy donor controls
CVID and RA patients are described in Table S1-S2. Healthy donors include a 36-year-
old Caucasian male (HD10) and 24-year-old Caucasian female (HD11). Additional
blood leukocyte preparations from control donors were obtained from the New York
Blood Center. Samples were collected after individuals signed informed consent in
accordance with Hospital for Special Surgery institutional review board-approved
protocols and the Declaration of Helsinki.
B cell purification, single cell sorting, cDNA and RT-PCR
Peripheral B cells were purified from the blood of patients and control donors by
negative selection using RosetteSepTM procedure (StemCell Technologies).
Alternatively, mature naïve B cells were enriched from peripheral blood mononuclear
cells using the Naïve B Cell Isolation Kit II (Miltenyi). B cells were stained with
fluorescein isothiocyanate (FITC) anti-human CD27, phycoerythrin (PE) anti-human
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

5
CD10, and either anti-human IgM biotin and allophycocyanin (APC) anti-human CD19,
or PE-Cy7 anti-human CD19 and APC anti-human CD21 (Pharmingen, Beckton
Dickinson). Biotinylated antibodies were revealed using streptavidin-PECy7 (Caltag
Laboratories). Single CD21loCD10+IgMhiCD27- new emigrant, CD19+CD10-CD21+CD27-
conventional mature naive and CD19+CD10-CD21-/loCD27- B cells from patients and
control donors were sorted on a FACSVantage (Becton Deckinson) into 96-well PCR
plates containing 4 μl of lysis solution (0.5X PBS containing 10 mM DTT, 8 U RNAsin
(Promega), and 0.4 U 5’-3’ RNase Inhibitor (Eppendorf)) and immediately frozen on dry
ice. All samples were stored at -70°C. RNA from single cells was reverse-transcribed in
the original 96 well plate in 12.5μl reactions containing 100U of Superscript II RT (Gibco
BRL) for 45 min at 37°C. RT-PCR reactions and primer sequences were as described
24. Immunoglobulin sequences and mutation status were analyzed by Ig BLAST
comparison with GenBank using the National Center for Biotechnology Information
IgBlast server (http://www.ncbi.nlm.nih.gov/igblast/). Heavy chain CDR3 was defined as
the interval between the conserved cysteine at position 92 in the VH framework 3 and the
conserved tryptophan at position 103 in JH segments.
Antibodies
Additional flow cytometry analyses were performed using anti-BAFF-R-FITC, CD86-
FITC, MHC class II/HLA-DR-FITC, CD44-FITC, CD62L-FITC, CD25-FITC, Fas/CD95-
FITC, ICOSL-PE, TACI-PE, CD80-PE, MHC class I/HLA-A,B,C-PE, CD11c-PE, CD32-
PE, CD93-PE, CD40-Alexa Fluor 647, CXCR4-APC (Biolegend), CD22-FITC, CD72-
FITC, CD21-PE, CD38-PE, CD69-PE, CD85j-PE, IgD-PE, CD19-PECy7, CD21-APC
(Pharmingen, Beckton Dickinson), CCR7-FITC, IL-4R-PE (R&D Systems), and CD23-
FITC (Serotec). Unconjugated anti-CD58 (Pharmingen, Beckton Dickinson) and CD85d
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

6
(Abcam) mouse monoclonal antibodies were coupled using the Alexa Fluor 488
Monoclonal Antibody Labeling Kit (Molecular Probes).
Microarray gene expression profile analysis
RNA was extracted from 105-3.105 batch sorted CD19+CD10-CD21+CD27- conventional
mature naive and CD19+CD10-CD21-/loCD27- B cells using the Absolutely RNA
microprep kit (Stratagene). 100-200 ng of RNA was obtained per sample, and the
quality of the purified RNA was assessed by the Bioanalyzer from Agilent. Using the
Ovation biotin system kit from Nugen, 30-50ng of RNA was amplified and labeled to
produce cDNA. Labeled cDNA was hybridized on chips containing the whole human
genome (Human Genome U133 2.0 from Affymetrix). The data discussed in this
manuscript have been deposited in National Center for Biotechnology Information’s
Gene Expression Omnibus and are accessible through GEO Series accession number
GSE13917
(http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?token=hzgjvycoecikyrc&acc=GSE13917)
.
B cell activation, proliferation and survival
Naïve B cells, which contained CD21+ and CD21-/lo B cells from control donors and
patients were plated at 150,000-200,000 cells per well in a 96 well plate in RPMI 10%
serum and 20 μg/mL polyclonal F(ab)’2 rabbit anti-human IgM (Jackson
Immunoresearch), 1 μg/mL multimeric soluble recombinant human CD40L (Alexis
Biochemicals), and/or 1 μg/mL CpG (Invivogen) for 48 hours. To assess B cell
proliferation, naïve B cells were labeled with CFSE (Molecular Probes-Invitrogen), plated
and stimulated as described above. The cells were harvested on day 3 and labeled for
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

7
CD19 and CD21 expression before determining the division number by decreasing
CFSE fluorescence in flow cytometry experiments. The proportions of apoptotic and
dead cells were assessed by flow cytometry to measure binding with annexin V and
propidium iodide using the Annexin V-FITC Apoptosis Detection Kit I (BD Pharmingen).
Calcium Mobilization
To measure free intracellular calcium concentration [Ca2+]i, naïve B cells were first
labeled with anti-CD19 and anti-CD21 antibodies, and loaded with 5 mM Fluo-3 AM
(Molecular Probes-Invitrogen) in the presence of 0.2% Pluronic F-127 (Sigma) for 30 min
at room temperature. Cells were washed, resuspended at 106 cells/mL and [Ca2+]i was
monitored over time by flow cytometry on gated CD19+CD21+ and CD19+CD21-/lo B cells.
After the baseline was established, cells were stimulated with 20 μg/ml F(ab')2 mouse
anti-human IgM. Mean [Ca2+]i was analyzed using the FCS Assistant 1.2.9 software (BD
Biosciences).
Antibody production, ELISAs and IFAs
Cloning strategy, expression vectors, and antibody reactivity against specific antigens
were as described 24. Highly polyreactive ED38 was used as positive control in HEp-2-
reactivity and polyreactivity ELISAs 24. Antibodies were considered polyreactive when
they recognized at least 2 and usually all of the 4 analyzed antigens that include single-
stranded DNA (ssDNA), double-stranded DNA (dsDNA), insulin and lipopolysaccharide
(LPS). For indirect immunofluorescence assays, HEp-2 cell coated slides (Bion
Enterprises, LTD) were incubated in a moist chamber at room temperature with purified
recombinant antibodies at 50-100 μg/mL. FITC-conjugated goat anti-human IgG was
used as detection reagent. Pictures were taken with an Axioskop (Zeiss) using a Plan-
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

8
Neofluar 40x/.75 objective and Axiovision 3.1 acquisition software.
Statistics
Differences between healthy donor and patients populations were analyzed for statistical
significance with unpaired Student’s t-tests, using SigmaPlot software (Systat). A P
value of less than 0.05 was considered significant.
Supporting Information
Supplemental material includes eight figures and nineteen tables.
Results
CVID and RA patients may display unusual peripheral CD21-/lo B cells
CVID comprises a heterogeneous group of patients suffering from reduced serum levels
of several Ig isotypes resulting in an increased susceptibility to bacterial infections (Table
S1) 25. Indeed, CVID patients frequently show severely decreased frequencies of CD27+
memory B cells, especially those expressing switched isotypes 21,26,27. In addition, some
CVID patients display an unusual population of B cells that lack complement receptor 2
(CR2/CD21) 21-23. The presence of 20% or more CD21-/lo B cells in the blood of CVID
patients was proposed to be a criteria for the classification of these patients as CVID
group Ia who are prone to develop autoimmunity (Figure 1A-B) 21,22. Since CD21-/lo B
cells were also reported in the blood of patients with autoimmune diseases including
SLE 17,18 ,19,20, we analyzed the frequency of CD19+CD27-CD21-/lo B cells in healthy
donors and untreated active RA patients (Table S2). We found that the proportion of
CD21-/lo B cells was low in peripheral CD27-depleted B cells from healthy donors (Figure
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

9
1C-D) 17. In contrast, a subgroup of RA patients displayed a high frequency of CD21-/lo B
cells (Figure 1C-D). Similarly to CVID patients, CD19+CD27-CD21-/lo B cells in RA
patients were not new emigrant/transitional B cells because they expressed neither
CD10 nor CD38 (data not shown). Moreover, the frequency of CD19+CD10-CD27-CD21-
/lo (from now on referred to as CD21-/lo) B cells was stable in the same RA patients over a
4 year-period. Thus, both CVID and RA patients may display an increased proportion of
CD21-/lo B cells in their blood.
CD21-/lo B cells express highly autoreactive antibodies including ANAs
The physiology of CD21-/lo B cells has not been thoroughly investigated and the
involvement of these B cells in the development of autoimmunity is unknown. BCR
expression in CD21-/lo B cells was assessed by flow cytometry after depletion of CD27+
memory B cells. In agreement with previous reports, we found that the majority of CD21-
/loCD27- B cells from healthy donors expressed isotype-switched B cells (Figure 2A) 28-30.
In contrast, CD21-/loCD27- B cells from RA and CVID patients preferentially expressed
IgM and/or IgD (Figure 2A). Indeed, patients’ CD21-/lo B cells were enriched in IgMloIgD+
B cells, which represented about 30% of their CD21-/lo B cells whereas this population
only accounted for 10% of CD21-/lo B cells in healthy donors (Figure 2A).
To determine whether unswitched CD21-/lo B cells from RA and CVID patients, as well as
from healthy donors, expressed autoreactive BCRs, we compared the reactivity of
antibodies expressed by single CD21-/lo and single conventional CD19+CD10-CD27-
CD21+ (referred to as CD21+) mature naïve B cells from the same individuals. Most Ig
genes expressed by CD21-/lo B cells from RA and CVID patients were devoid of somatic
hypermutations, thereby demonstrating that these B cells belonged to the naïve
compartment. IgM genes amplified from CD21-/lo B cells from two healthy donors
displayed unmutated and mutated sequences, revealing that these cells contain both
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

10
naïve and memory B cells (Tables S3-S18 and data not shown). Therefore, we only
expressed BCRs encoded by germline Ig genes to compare naïve B cell populations.
We found that the frequency of polyreactive and HEp-2-reactive clones was increased in
CD21-/lo compared to CD21+ B cells in all individuals (Figure 2B and S1). Differences in
HEp-2 reactivity between CD21-/lo and CD21+ B cells reached significance for both
healthy donors and patients (Figure S2). In addition, CD21-/lo B cells often expressed
highly HEp-2 reactive antibodies (arbitrarily defined by ELISA OD405>2) compared to
their CD21+ B cell counterparts (Figure 2C). Highly HEp-2 reactive antibodies often
displayed heavy chain complementarity determining regions containing many positive
charges favoring autoreactivity and were mostly kappa clones, which suggested that
receptor editing using lambda light chains was not induced to silence these B cells
(Figure 2D and Tables S3-S18) 24,31,32. Indeed, CD21-/lo B cells showed a statistically
significant decrease in upstream Vκ gene usage combined to an increase in Jκ1 gene
usage compare to conventional CD21+ B cells, further suggesting decrease secondary
recombination events in CD21-/lo B cells despite their autoreactive features (Figure S3).
Thus, CD21-/lo B cells are enriched in polyreactive and strongly autoreactive clones,
which are normally silenced during B cell development in the bone marrow.
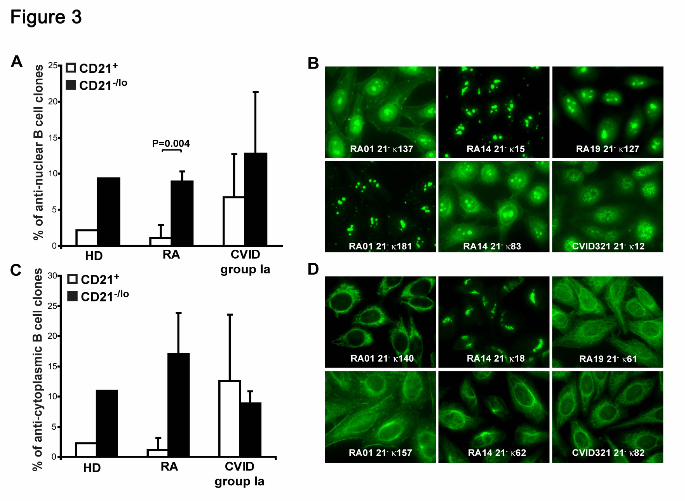
Immunofluorescence assays revealed that 5.6%-22.2% of naïve CD21-/lo B cells
expressed anti-nuclear antibodies (ANAs) (Figure 3A-B). ANAs expressed by CD21-/lo B
cells showed a diversity of nucleolar and speckled anti-nuclear staining patterns (Figure
3B). In contrast, ANAs were rarely detected in CD21+ B cells from healthy donors and
RA patients but could be fairly frequent in CD21-/lo B cells from CVID patients (Figure 3A
and data not shown). In addition, CD21-/lo B cells expressed antibodies that often
recognized specific cytoplasmic structures, which included mitochondria, the golgi
apparatus, and cytoskeleton components (Figure 3C-D, and Tables S3-S18). These
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

11
antibodies mostly corresponded to the highly HEp-2 reactive clones identified by ELISA.
In contrast, antibodies expressed by CD21+ B cells rarely recognized well-defined
structures in HEp-2 cells. We conclude that CD21-/lo B cells differ from CD21+
conventional B cells in that they often express autoreactive antibodies that may
recognize nuclear or cytoplasmic antigens.
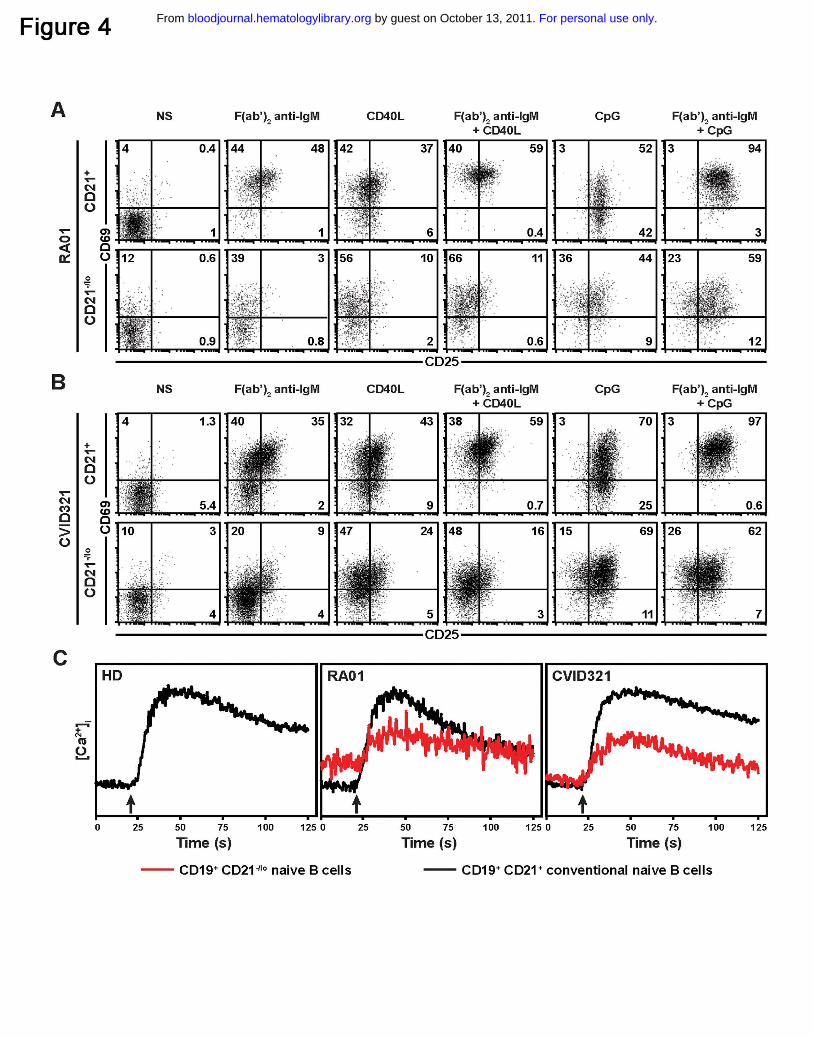
CD21-/lo B cells fail to get activated through BCR and CD40 triggering
We analyzed the physiology of CD21-/lo B cells by studying the induction of CD25, CD69,
CD23, TACI, ICOS-L and Fas by flow cytometry on CD21+ and CD21-/lo gated B cells
after BCR, CD40 and TLR9 stimulations for 2 days. CD21+ B cells from RA01, RA19,
CVID218 and CVID321 patients upregulated CD69 and CD25 after BCR, CD40 or TLR9
triggering (Figure 4A-B and data not shown). In contrast, CD21-/lo B cells from the same
patients failed to properly induce cell surface expression of CD25 and CD69 after BCR
and CD40 triggering (Figure 4A-B). The induction of CD25 and CD69 in CD21-/lo B cells
by TLR9 stimulation with CpG, although decreased compared to conventional B cells,
were less affected than those induced by BCR and/or CD40 triggering (Figure 4A-B).
Defects in the induction of CD23 on CD21-/lo compared to CD21+ B cells were also
observed after BCR, TLR9, or especially CD40 stimulations (Figure S4-S5). These
defects were not the result of a global inability of CD21-/lo B cells to get activated,
because CD21-/lo B cells upregulated cell surface expression of ICOS-L, TACI and Fas
after BCR, CD40 or TLR9 stimulation (Figure S4-S5). In addition, similar results were
obtained when CD21+ and CD21-/lo B cells were initially fractionated and then stimulated
in vitro for 2 days (data not shown). Thus, CD21+ and CD21-/lo B cells display distinct B
cell responses after BCR, CD40 or TLR9 stimulation.
BCR triggering does not induce calcium flux in CD21-/lo B cells
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

12
The inability of CD21-/lo B cells to upregulate some activation markers upon BCR
triggering could result from an inability to properly induce intracellular calcium flux. We
analyzed [Ca2+]i after BCR stimulations in these B cells compared to conventional CD21+
B cells. BCR triggering induced a strong [Ca2+]i increase in CD21+ conventional B cells
from healthy donors as well as RA and CVID patients (Figure 4C and data not shown).
In contrast, CD21-/lo B cells from RA01, RA19, RA26, CVID321, and CVID218 patients
only induced a weak elevation of [Ca2+]i in response to BCR stimulation (Figure 4C and
data not shown). We conclude that CD21-/lo B cells do not exhibit a proper calcium
response following BCR stimulation.
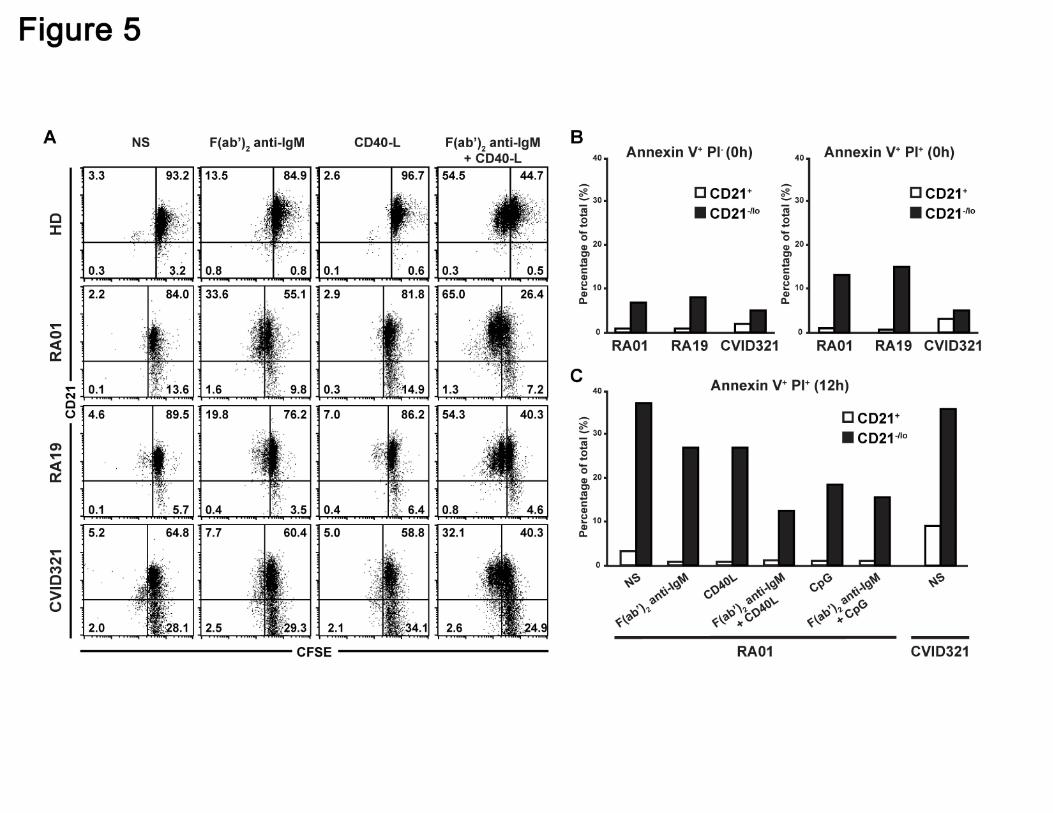
CD21-/lo B cells do not proliferate after BCR and CD40 triggering
Since CD21-/lo B cells showed an impaired activation compared to CD21+ B cells in
response to several stimulations, we investigated their ability to proliferate after antigenic
stimulation. Carboxyfluorescein succinimidyl ester (CFSE) labeled CD21+ and CD21-/lo B
cells from 5 RA and 4 CVID patients and healthy donor controls were stimulated through
their BCR or CD40 for 3 days and CFSE dilution, which reflects B cell proliferation, was
then assessed by flow cytometry (Figure 5A and data not shown). Anti-IgM F(ab’)2
stimulation induced the proliferation of a few CD21+ B cells in all individuals whereas
CD40L did not (Figure 5A). BCR and CD40 co-triggering resulted in the proliferation of a
majority of CD21+ B cells from both healthy donors and patients and some of these B
cells underwent up to two divisions (Figure 5A). In contrast, RA and CVID CD21-/lo B
cells failed to proliferate after BCR and CD40 costimulation. We conclude that unlike
CD21+ conventional B cells, CD21-/lo B cells are unable to proliferate after antigenic
stimulation, further suggesting their unresponsive stage.
CD21-/lo B cells are prone to die by apoptosis
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

13
We assessed the survival potential of CD21-/lo B cells by analyzing the binding of
Annexin V to their surface, which along with propidium iodide (PI) staining can define
Annexin V+PI- early apoptotic cells and Annexin V+PI+ late apoptotic and dead cells. We
found that freshly isolated CD21-/lo B cells from RA and CVID patients contained a higher
frequency of both Annexin V+PI- early apoptotic and Annexin V+PI+ late apoptotic and
dead B cells compared to conventional CD21+ B cells (Figure 5B). When naïve B cells
from RA and CVID patients were left unstimulated for 12 hours, CD21-/lo B cells
contained about 35% of Annexin V+PI+ dying cells whereas there were less than 10% in
CD21+ B cells (Figure 5C). Moreover, triggering through BCR, CD40 and/or TLR9 only
partially rescued these B cells from cell death. In contrast, stimulated CD21+ B cells
contained virtually no Annexin V+PI+ B cells (Figure 5C). In agreement with their
increased susceptibility to die, CD21-/lo B cells also contained a higher proportion of
Annexin V+PI- early apoptotic cells than CD21+ B cells, regardless of the stimulation
(Figure S6). Hence, CD21-/lo B cells are more susceptible to apoptosis and cell death
than CD21+ B cells and their ability to be eliminated is poorly rescued by BCR
stimulation in combination with either CD40 or TLR9 triggering.
The transcriptome of CD21-/lo B cells shows an inhibitory gene signature
We intended to further characterize CD21-/lo autoreactive unresponsive B cells by
performing gene array profiling and compare transcripts expressed in CD21-/lo versus
CD21+ mature naïve B cells isolated from 2 RA and 3 CVID patients (Figure 6). 1,079
transcripts were found expressed with a greater than twofold statistically significant
difference by the 2 B cell subpopulations and included 656 upregulated and 423
downregulated transcripts in CD21-/lo B cells (Figure 6A). Some of those differentially
expressed genes, and a few others of specific interest, are shown on Figure 6B and
Table S19. Correlating with a decreased expression of CD21 on their cell surface, we
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

14
found that CD21-/lo B cells down-regulated the transcription of CR2/CD21, as well as
other complement binding molecules CR1/CD35, CD55 and CD59 (Figure 6B). Many
genes encoding survival and activation molecules like CD40, OX40L and BCMA, as well
as the cytokine receptors IL-4R and IL-13R that stimulate B cell proliferation, were all
downregulated in CD21-/lo B cells (Figure 6B). In contrast, CD21-/lo B cells upregulated
the transcription of a whole set of immunoreceptor tyrosine-based inhibition motif (ITIM)
receptor genes likely to inhibit B cell activation and proliferation. They included CD72,
Fc receptors (FcR) CD32/FCGR2B and FCRL5/IRTA2, as well as LILRB and SIGLEC
genes (Figure 6B). Genes encoding phosphatases potentially mediating the inhibitory
effect of these ITIM-receptors where also found upregulated in the CD21-/lo B cells
(Figure 6B). In addition, these B cells downregulated transcripts encoding the chemokine
receptors CCR7, CXCR4 and CXCR5 as well as lymphotoxin A which regulates B cell
trafficking (Figure 6B). Lymphocyte homing capacities are also likely to differ between
CD21-/lo and CD21+ B cells because transcripts for many integrins (ITG), including
ITGAX, which encodes CD11c molecules, were upregulated in CD21-/lo B cells, while the
transcripts for selectin CD62L were downregulated (Figure 6B). The increased amounts
of growth arrest-specific 7 (GAS7) and CDKN1A/p21 combined to decreased cyclin-
dependent kinase 6 (CDK6) and MYC transcripts in CD21-/lo B cells are likely to result in
the observed defective proliferation of these cells. In addition, the upregulation of
Fas/CD95 and caspase genes in CD21-/lo B cells could be responsible for their increased
susceptibility to cell death. Finally, we found that some transcription factor genes were
upregulated in CD21-/lo B cells compared to CD21+ B cells, such as BCL11B, EGR2,
HOXA3, HOXB7, HOXC4, NFATC2, SOX4, TBET, and TOX, while a few transcription
factor genes were found downregulated, such as FOXP1 (Figure 6B). In conclusion, the
transcriptome of CD21-/lo B cells is characterized by the upregulation of many genes
encoding molecules likely to inhibit B cell activation, proliferation and survival and the
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

15
downregulation of B cell activating genes, supporting the observation that CD21-/lo B
cells are refractory to antigenic stimulation.
CD21-/lo B cells display a specific B cell phenotype
The expression of many genes found either upregulated or downregulated in gene array
experiments was validated by flow cytometry in RA01, RA19, CVID214, CVID218 and
CVID321 after B cell surface staining with specific monoclonal antibodies.
Representative results for RA and CVID patients, as well as for healthy controls, are
shown in Figure 7 and S7 and S8 respectively. In agreement with gene array analysis,
CD21-/lo B cells were CD19hiCD40loCD86+CD58hiIL-4RloCD11c+CCR7-
/loCXCR4loCD44loCD62L-/loCD72hiCD32hiCD85j+CD85d+Fas+ (Figure 7 and S7-S8).
Moreover, we also found that BAFF-R was downregulated and MHC class I and ITIM
receptor CD22 was upregulated in CD21-/lo B cells, whereas changes in their gene
expression were not detected in gene array experiments (Figure 7 and S7-S8). Human
CD21+ and CD21-/lo B cells expressed TACI, ICOS-L, CD80 and MHC class II at similar
levels and did not expressed CD93, which was found on the cell surface of mouse
anergic transitional 3 (T3) B cells (Figure 7 and S7-S8, and data not shown). Hence,
CD21-/lo B cells display a unique phenotype, which is characterized by the
downregulation of receptors that normally favor B cell survival and/or activation, and the
upregulation of molecules inhibiting B cell functions.
Discussion
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

16
We report herein the identification in humans of a unique autoreactive B cell population,
which lacks CD21 expression and is refractory to B cell stimulation. Indeed, naïve
CD21-/lo B cells express a diverse germline BCR repertoire, which is enriched in
autoreactive clones. In addition, CD21-/lo B cells showed impaired calcium mediated
signaling, did not upregulate some activation markers, and did not proliferate in
response to BCR triggering. CD21-/lo B cells were also prone to die faster than CD21+
conventional B cells, suggesting they have a shorter half-life. Because most of these
features are characteristic of mouse anergic B cells, autoreactive CD21-/lo B cells, which
escaped central B cell tolerance deletion and remain in the periphery in an irresponsive
stage, therefore represent a population of anergic B cells in humans 4,14,15,33-37.
B cells which express intrinsically autoreactive BCRs encoded by the VH4-34 gene have
been suggested to be anergic because they fail to induce normal calcium flux after BCR
triggering 38. In addition, VH4-34 B cells are enriched in mature naïve B cells and
remain excluded from germinal centers (GCs) in healthy donors 38,39. However, we did
not find any increase in VH4-34 gene usage in CD21-/lo B cells, which expressed a very
diverse immunoglobulin repertoire. The abundance of self-antigens expressed on
erythrocytes and recognized by VH4-34 BCRs may induce a unique anergic program in
these cells that differs from that of CD21-/lo B cells, which may recognize a broader set of
self-antigens. Another population of irresponsive B cells which downregulated IgM
expression was recently reported in humans 16. These B cells also express autoreactive
antibodies and fail to induce calcium flux and tyrosine phosphorylation cascade after IgD
triggering but could get activated and proliferated in vitro 16. By analogy to anergic
mouse models, which express different autoreactive transgenic antibodies and develop
different anergic B cell programs, we would like to propose that VH4-34-expressing, IgM-
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

17
negative and CD21-/lo B cells represent different populations of anergic B cells in
humans.
The existence of some CD21-/lo B cells have been previously reported in CVID and SLE
patients, in individuals infected with either human immunodeficiency virus (HIV) or
hepatitis C virus combined with mixed cryoglobulinemia, as well as in healthy donors 17-
23,28-30,40. All these CD21-/lo B cells, with the exception of those in CVID patients, belong
to the memory B cell compartment because they frequently express mutated isotype
switched antibodies 28-30. In agreement with these reports, we found that memory B cells
represented the majority of CD21-/lo B cells in healthy donors. In contrast, CD21-/lo B
cells from RA and CVID patients were often naïve B cells because they produced
germline encoded IgM and IgD antibodies. However, both naïve and memory CD21-/lo B
cells displayed a similar gene expression profile and expressed common cell surface
molecules, which differed from conventional CD21+ B cells 28,30. It is unknown whether
memory CD21-/lo B cells from healthy donors are anergic but CD21-/lo B cells from HIV-
infected viremic individuals and CVID patients poorly respond to antigenic stimulation
23,28. Chronic antigen exposure, such as in HIV-infected viremic individuals, may lead
antigen-reactive B cells to an anergic phenotype similar to CD21-/lo naïve B cells and
disable them to potentially secrete antibodies.
The phenotype of CD21-/lo B cells revealed the modulation of the expression of many
molecules that may prevent the activation of these B cells. The downregulation of the
complement receptor CR2/CD21 previously reported on mouse anergic B cells is likely
to result in an increased BCR signaling threshold and compromise the ability of CD21-/lo
B cells to be activated 34,37. In addition, CD21 is able to break anergy in mouse B cells,
further suggesting that its downregulation contribute to an anergic stage in mice and
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

18
humans 34,37,41. The downregulation of CD40, OX40L, IL-4R and IL-13R, which play an
important role in B cell activation and proliferation, may also be responsible for the
anergic responses of CD21-/lo B cells. Moreover, increased amounts of many ITIM
receptors on CD21-/lo B cells, including CD22, FcγR2B/CD32, and CD72 that was
already reported in mouse anergic B cells, can further dampen the activation of these B
cells 42-45. The overexpression of many phosphatases in CD21-/lo B cells is also likely to
decrease their antigenic responses. Chemokine receptors including CCR7, CXCR4, and
CXCR5 as well as selectin CD62L have been shown to play a major role in B cell
trafficking in secondary lymphoid organs and GCs 46,47. The downregulation of these
receptors on CD21-/lo B cells is likely to exclude them from B cell follicules and being
recruited in germinal centers 9,48. In addition, the upregulation of many integrins also
involved in B cell homing may direct these autoreactive B cells to specific compartments
where they may be eventually eliminated. Moreover, the increased expression of
molecules favoring cell death and blocking cell cycle entry may further prevent the
expansion of CD21-/lo B cells and favor their removal. Altogether, CD21-/lo B cells
express a specific set of molecules that may prevent the activation of these B cells.
Some of these molecules may also serve as biomarkers to identify human unresponsive
autoreactive naïve B cells and distinguish them from the other CD21-/lo B cells belonging
to the memory compartment.
Unresponsive autoreactive CD21-/lo B cells express a specific B cell phenotype, which
may be regulated by transcription factors. However, transgenic mouse models did not
identify a specific transcription factor whose expression would be responsible for the
establishment of an anergic B cell program 42. Indeed, it has been postulated that
because the maintenance of an anergic stage requires constant BCR triggering by self-
antigens, the anergic phenotype may not depend on a genetic program ensured by
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

19
transcription factors 13,14. Gene array comparisons between CD21-/lo and CD21+ naïve
B cells revealed the induction of a set of transcription factors in human unresponsive
autoreactive CD21-/lo B cells. It is unknown whether this transcription factor signature is
dependent on constant BCR triggering by self-antigens, but it unveils new leads in the
identification of factors that may control the expression of the molecules resulting in the
inability of CD21-/lo B cells to respond to antigenic stimulations.
Three mechanisms normally ensure the silencing of developing autoreactive B cells.
Receptor editing mediated by secondary recombination has been reported to be very
efficient at silencing autoreactive B cells reacting against membrane bound antigens and
deletion seems to be used when receptor editing failed to silence autoreactive B cells 12.
Anergy may be preferentially induced in B cells that express moderately autoreactive
BCRs toward soluble antigens 11. In agreement with this hypothesis, we found that
CD21-/lo B cells that express unmutated IgM in healthy donors display weakly
autoreactive BCRs and may represent anergic B cells. Healthy donors also showed low
frequencies of CD21-/lo B cells, suggesting that anergy is rarely used to silence
developing autoreactive B cells in physiological conditions. Factors that favor the
development or the maintenance of CD21-/lo B cells in the blood of some RA and CVID
patients remain to be determined. The serum of CVID group Ia but not RA patients
contains elevated concentrations of B-cell activating factor (BAFF), which is known to
inhibit the counterselection of anergic B cells in mice and likely to contribute to the large
accumulation of CD21-/lo B cells in these CVID patients (49 and data not shown).
The induction of an unresponsive program in highly autoreactive B cells may represent
risks to develop autoimmunity. Indeed, elevated numbers of highly autoreactive
unresponsive CD21-/lo B cells remain in the blood of RA and CVID patients instead of
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

20
being eliminated and immune reactions may create a favorable environment to break
tolerance and eventually activate these CD21-/lo B cells. In RA, all patients develop
autoimmunity, whether or not they display CD21-/lo B cells, and it is unclear how these
B cells contribute to the initiation and/or progression of the disease. However, CVID
group Ia patients who display high frequencies of CD21-/lo B cells are more susceptible
to develop autoimmune syndromes, suggesting that CD21-/lo B cells may contribute to
the development of autoimmunity in humans.
Acknowledgments
We are very much indebted to the patients and their families. We thank Dr. S.
Rudchenko and S. Semova for help with the single cell sorter, Dr. D. Orange, D.
Goldenberg and K. Warnatz for providing RA and CVID patient samples and Dr G.
Charvin for microscopy analysis. This publication was made possible by Grant Number
AI061093 and AI071087 from NIH-NIAID.
Authorship Contributions
I. I performed the experiments, with help from E. M. for cell isolation and sorting, and
from Y.-S. N., L. M., G. M., D. S. and I. S. for antibody cloning and reactivity testing. E.
M. initiated the collaboration with J. S., J. B., J. H. B. and C. C.-R. who provided patient
blood samples. I. I. and E. M. designed the research and wrote the manuscript.
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

21
Disclosure of Conflicts of Interest
The authors declare no competing financial interests.
References
1. Nemazee D. Does immunological tolerance explain the waste in the B-
lymphocyte immune system? Experiment and theory. Ann. N. Y. Acad. Sci.
1995;764:397-401
2. Radic MZ, Weigert M. Origins of anti-DNA antibodies and their implications for
B-cell tolerance. Ann. N. Y. Acad. Sci. 1995;764:384-396
3. Goodnow CC, Sprent J, Fazekas de St Groth B, Vinuesa CG. Cellular and
genetic mechanisms of self tolerance and autoimmunity. Nature. 2005;435 (7042):590-
597
4. Goodnow CC, Crosbie J, Adelstein S, Lavoie TB, Smith-Gill SJ, Brink RA,
Pritchard-Briscoe H, Wotherspoon JS, Loblay RH, Raphael K, et al. Altered
immunoglobulin expression and functional silencing of self- reactive B lymphocytes in
transgenic mice. Nature. 1988; 334 (6184):676-682
5. Erikson J, Radic MZ, Camper SA, Hardy RR, Carmack C, Weigert M.
Expression of anti-DNA immunoglobulin transgenes in non-autoimmune mice. Nature.
1991;349 (6307):331-334
6. Tsao BP, Chow A, Cheroutre H, Song YW, McGrath ME, Kronenberg M. B
cells are anergic in transgenic mice that express IgM anti-DNA antibodies. Eur. J.
Immunol. 1993;23 (9):2332-2339
7. Cambier JC, Gauld SB, Merrell KT, Vilen BJ. B-cell anergy: from transgenic
models to naturally occurring anergic B cells? Nat. Rev. Immunol. 2007;7 (8):633-643
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

22
8. Fulcher DA, Basten A. Reduced life span of anergic self-reactive B cells in a
double- transgenic model. J. Exp. Med. 1994;179 (1):125-134
9. Cyster JG, Hartley SB, Goodnow CC. Competition for follicular niches
excludes self-reactive cells from the recirculating B-cell repertoire [see comments].
Nature. 1994;371 (6496):389-395
10. Nemazee DA, Bürki K. Clonal deletion of B lymphocytes in a transgenic
mouse bearing anti-MHC class I antibody genes. Nature. 1989;337 (6207):562-566
11. Hartley SB, Crosbie J, Brink R, Kantor AB, Basten A, Goodnow CC.
Elimination from peripheral lymphoid tissues of self-reactive B lymphocytes recognizing
membrane-bound antigens. Nature. 1991;353 (6346):765-769
12. Halverson R, Torres RM, Pelanda R. Receptor editing is the main
mechanism of B cell tolerance toward membrane antigens. Nat. Immunol. 2004;
5(6):645-650
13. Healy JI, Dolmetsch RE, Timmerman LA, Cyster JG, Thomas ML, Crabtree
GR, Lewis RS, Goodnow CC. Different nuclear signals are activated by the B cell
receptor during positive versus negative signaling. Immunity. 1997;6 (4):419-428
14. Gauld SB, Benschop RJ, Merrell KT, Cambier JC. Maintenance of B cell
anergy requires constant antigen receptor occupancy and signaling. Nat. Immunol.
2005;6 (11):1160-1167
15. Merrell KT, Benschop RJ, Gauld SB, Aviszus K, Decote-Ricardo D, Wysocki
LJ, Cambier JC. Identification of anergic B cells within a wild-type repertoire. Immunity.
2006;25 (6):953-962
16. Duty JA, Szodoray P, Zheng NY, Koelsch KA, Zhang Q, Swiatkowski M,
Mathias M, Garman L, Helms C, Nakken B, Smith K, Farris AD, Wilson PC. Functional
anergy in a subpopulation of naive B cells from healthy humans that express
autoreactive immunoglobulin receptors. J. Exp. Med. 2009;206 (1):139-151
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

23
17. Wehr C, Eibel H, Masilamani M, Illges H, Schlesier M, Peter HH, Warnatz K.
A new CD21low B cell population in the peripheral blood of patients with SLE. Clin.
Immunol. 2004;113 (2):161-171
18. Wei C, Anolik J, Cappione A, Zheng B, Pugh-Bernard A, Brooks J, Lee EH,
Milner EC, Sanz I. A new population of cells lacking expression of CD27 represents a
notable component of the B cell memory compartment in systemic lupus erythematosus.
J. Immunol. 2007;178 (10):6624-6633
19. Culton DA, Nicholas MW, Bunch DO, Zhen QL, Kepler TB, Dooley MA,
Mohan C, Nachman PH, Clarke SH. Similar CD19 dysregulation in two autoantibody-
associated autoimmune diseases suggests a shared mechanism of B-cell tolerance loss.
J. Clin. Immunol. 2007;27 (1):53-68
20. Nicholas MW, Dooley MA, Hogan SL, Anolik J, Looney J, Sanz I, Clarke SH.
A novel subset of memory B cells is enriched in autoreactivity and correlates with
adverse outcomes in SLE. Clin. Immunol. 2008;126 (2):189-201
21. Warnatz K, Denz A, Drager R, Braun M, Groth C, Wolff-Vorbeck G, Eibel H,
Schlesier M, Peter HH. Severe deficiency of switched memory B cells (CD27(+)IgM(-
)IgD(-)) in subgroups of patients with common variable immunodeficiency: a new
approach to classify a heterogeneous disease. Blood. 2002;99 (5):1544-1551
22. Warnatz K, Wehr C, Drager R, Schmidt S, Eibel H, Schlesier M, Peter HH.
Expansion of CD19(hi)CD21(lo/neg) B cells in common variable immunodeficiency
(CVID) patients with autoimmune cytopenia. Immunobiology. 2002;206 (5):502-513
23. Rakhmanov M, Keller B, Gutenberger S, Foerster C, Hoenig M, Driessen G,
van der Burg M, van Dongen JJ, Wiech E, Visentini M, Quinti I, Prasse A, Voelxen N,
Salzer U, Goldacker S, Fisch P, Eibel H, Schwarz K, Peter HH, Warnatz K. Circulating
CD21low B cells in common variable immunodeficiency resemble tissue homing, innate-
like B cells. Proc. Natl. Acad. Sci. USA. 2009;106 (32):13451-13456
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

24
24. Wardemann H, Yurasov S, Schaefer A, Young JW, Meffre E, Nussenzweig
MC. Predominant autoantibody production by early human B cell precursors. Science.
2003;301 (5638):1374-1377
25. Cunningham-Rundles C, Bodian C. Common variable immunodeficiency:
clinical and immunological features of 248 patients. Clin. Immunol. 1999;92 (1):34-48
26. Piqueras B, Lavenu-Bombled C, Galicier L, Bergeron-van der Cruyssen F,
Mouthon L, Chevret S, Debre P, Schmitt C, Oksenhendler E. Common variable
immunodeficiency patient classification based on impaired B cell memory differentiation
correlates with clinical aspects. J. Clin. Immunol. 2003;23 (5):385-400
27. Wehr C, Kivioja T, Schmitt C, Ferry B, Witte T, Eren E, Vlkova M, Hernandez
M, Detkova D, Bos PR, Poerksen G, von Bernuth H, Baumann U, Goldacker S,
Gutenberger S, Schlesier M, Bergeron-van der Cruyssen F, Le Garff M, Debre P,
Jacobs R, Jones J, Bateman E, Litzman J, van Hagen PM, Plebani A, Schmidt RE, Thon
V, Quinti I, Espanol T, Webster AD, Chapel H, Vihinen M, Oksenhendler E, Peter HH,
Warnatz K. The EUROclass trial: defining subgroups in common variable
immunodeficiency. Blood. 2008;111 (1):77-85
28. Moir S, Ho J, Malaspina A, Wang W, DiPoto AC, O'Shea MA, Roby G, Kottilil
S, Arthos J, Proschan MA, Chun TW, Fauci AS. Evidence for HIV-associated B cell
exhaustion in a dysfunctional memory B cell compartment in HIV-infected viremic
individuals. J. Exp. Med. 2008;205 (8):1797-1805
29. Ehrhardt GR, Hsu JT, Gartland L, Leu CM, Zhang S, Davis RS, Cooper MD.
Expression of the immunoregulatory molecule FcRH4 defines a distinctive tissue-based
population of memory B cells. J. Exp. Med. 2005;202 (6):783-791
30. Ehrhardt GR, Hijikata A, Kitamura H, Ohara O, Wang JY, Cooper MD.
Discriminating gene expression profiles of memory B cell subpopulations. J. Exp. Med.
2008;205 (8):1807-1817
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

25
31. Radic MZ, Mackle J, Erikson J, Mol C, Anderson WF, Weigert M. Residues
that mediate DNA binding of autoimmune antibodies. J. Immunol. 1993;150 (11):4966-
4977.
32. Isnardi I, Ng YS, Srdanovic I, Motaghedi R, Rudchenko S, von Bernuth H,
Zhang SY, Puel A, Jouanguy E, Picard C, Garty BZ, Camcioglu Y, Doffinger R,
Kumararatne D, Davies G, Gallin JI, Haraguchi S, Day NK, Casanova JL, Meffre E.
IRAK-4- and MyD88-dependent pathways are essential for the removal of developing
autoreactive B cells in humans. Immunity. 2008;29 (5):746-757
33. Goodnow CC, Brink R, Adams E. Breakdown of self-tolerance in anergic B
lymphocytes. Nature. 1991;352 (6335):532-536
34. Hartley SB, Cooke MP, Fulcher DA, W. HA, Cory S, Basten A, Goodnow
CC. Elimination of Self-Reactive B Lymphocytes Proceeds in Two Stages: Arrested
Development and Cell Death. Cell. 1993;72 (3):325-335
35. Cooke MP, Heath AW, Shokat KM, Zeng Y, Finkelman FD, Linsley PS,
Howard M, Goodnow CC. Immunoglobulin signal transduction guides the specificity of B
cell-T cell interactions and is blocked in tolerant self-reactive B cells. J. Exp. Med.
1994;179 (2):425-438
36. Nguyen KA, Mandik L, Bui A, Kavaler J, Norvell A, Monroe JG, Roark JH,
Erikson J. Characterization of anti-single-stranded DNA B cells in a non-autoimmune
background. J. Immunol. 1997;159 (6):2633-2644
37. Mandik-Nayak L, Bui A, Noorchashm H, Eaton A, Erikson J. Regulation of
anti-double-stranded DNA B cells in nonautoimmune mice: localization to the T-B
interface of the splenic follicle. J. Exp. Med. 1997;186 (8):1257-1267
38. Cappione A, 3rd, Anolik JH, Pugh-Bernard A, Barnard J, Dutcher P,
Silverman G, Sanz I. Germinal center exclusion of autoreactive B cells is defective in
human systemic lupus erythematosus. J. Clin. Invest. 2005;115 (11):3205-3216
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

26
39. Pugh-Bernard AE, Silverman GJ, Cappione AJ, Villano ME, Ryan DH, Insel
RA, Sanz I. Regulation of inherently autoreactive VH4-34 B cells in the maintenance of
human B cell tolerance. J. Clin. Invest. 2001;108 (7):1061-1070
40. Charles ED, Green RM, Marukian S, Talal AH, Lake-Bakaar GV, Jacobson
IM, Rice CM, Dustin LB. Clonal expansion of immunoglobulin M+CD27+ B cells in HCV-
associated mixed cryoglobulinemia. Blood. 2008;111 (3):1344-1356
41. Lyubchenko T, Dal Porto JM, Holers VM, Cambier JC. Cutting edge:
Complement (C3d)-linked antigens break B cell anergy. J. Immunol. 2007;179 (5):2695-
2699
42. Glynne R, Ghandour G, Rayner J, Mack DH, Goodnow CC. B-lymphocyte
quiescence, tolerance and activation as viewed by global gene expression profiling on
microarrays. Immunol. Rev. 2000;176:216-246
43. Fujimoto M, Bradney AP, Poe JC, Steeber DA, Tedder TF. Modulation of B
lymphocyte antigen receptor signal transduction by a CD19/CD22 regulatory loop.
Immunity. 1999;11 (2):191-200
44. Pan C, Baumgarth N, Parnes JR. CD72-deficient mice reveal nonredundant
roles of CD72 in B cell development and activation. Immunity. 1999;11 (4):495-506
45. Nimmerjahn F, Ravetch JV. Fcgamma receptors as regulators of immune
responses. Nat. Rev. Immunol. 2008;8 (1):34-47
46. Allen CD, Ansel KM, Low C, Lesley R, Tamamura H, Fujii N, Cyster JG.
Germinal center dark and light zone organization is mediated by CXCR4 and CXCR5.
Nat. Immunol. 2004;5 (9):943-952
47. Forster R, Davalos-Misslitz AC, Rot A. CCR7 and its ligands: balancing
immunity and tolerance. Nat. Rev. Immunol. 2008;8 (5):362-371
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

27
48. Cyster JG, Goodnow CC. Antigen-induced exclusion from follicles and
anergy are separate and complementary processes that influence peripheral B cell fate.
Immunity. 1995;3 (6):691-701
49. Lesley R, Xu Y, Kalled SL, Hess DM, Schwab SR, Shu H-B, Cyster JG.
Reduced competitiveness of autoantigen-engaged B cells due to increased dependence
on BAFF. Immunity. 2004;20 (4):441-453
Figure legends
Figure 1. CVID and RA patients may display an increased frequency of CD21-/lo B
cells in their blood. (A) CVID group Ia patients were previously defined as patients
displaying a frequency of CD21-/lo naïve B cells > 20% of total B cells 21. Each diamond
represents an individual and (B) dot plots show CD19 and CD21 expression on CD27-
gated peripheral B cells from representative CVID group Ia patients. (C) Increased
CD21-/lo B cell frequency in RA patients. (C) Dot plots show CD19 and CD21 expression
on CD27-depleted peripheral B cells from a representative healthy donor (HD10) and RA
patients. (D) The frequencies of CD21-/lo naïve B cells are compared between healthy
donors and RA patients. Each diamond represents an individual and statistically
significant differences are indicated.
Figure 2. CD21-/lo naïve B cells express autoreactive BCRs. (A) A majority of CD21-/lo
B cells from RA and CVID patients express IgM and IgD. Dot plots show IgM and IgD
expression gated on CD19+CD21+(left) and CD19+CD21-/lo (right) CD27-depleted B cells.
(B) Most CD21-/lo naïve B cells express autoreactive antibodies. Antibodies cloned from
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

28
single CD21+CD27-CD10- and CD21-/loCD27-CD10- B cells from two healthy donors
(HD10 and 11), 3 RA and 3 CVID patients were tested by ELISA for reactivity against
HEp-2 lysates. Dotted lines show ED38-positive control 24. Horizontal lines show cut-off
OD405 for positive reactivity. For each B cell population, the frequency of HEp-2-reactive
and non HEp-2-reactive clones is summarized in pie charts with the number of
antibodies tested indicated in the centers. The frequency of HEp-2 reactive B cells was
higher in CD21-/lo than in CD21+ B cells in all individuals. (C) CD21-/lo B cells express
highly autoreactive antibodies. The frequency of highly HEp-2-reactive antibodies,
arbitrarily defined by ELISA OD405>2, expressed by CD21+ and CD21-/lo B cells from HD,
RA and CVID patients is represented. A large fraction of CD21-/lo B cells from most
individuals were highly HEp-2-reactive. P values calculated using a paired T-test are
indicated when significant. (D) A majority of highly autoreactive antibodies from CD21-/lo
B cells express kappa light chains. The frequency of highly HEp-2-reactive CD21-/lo B
cells from HD, RA and CVID patients is represented subdivided among kappa and
lambda clones. Many highly HEp-2-reactive CD21-/lo B cells express kappa light chains
suggesting that receptor editing using lambda light chains was not induced to silence
these clones.
Figure 3. CD21-/lo B cells contain anti-nuclear and anti-cytoplasmic clones. The
frequencies of anti-nuclear (A) and anti-cytoplasmic (C) clones in CD21+ and CD21-/lo B
cells from HD, RA and CVID patients are shown. P values calculated using a paired T-
test are indicated when significant. (B) ANAs expressed by CD21-/lo B cells showed a
diversity of nucleolar (RA01 21- κ181, RA01 21- κ137, RA14 21- κ15, and RA14 21- κ83),
nucleolar and speckled (RA19 21- κ127) and speckled (CVID 21- κ12) anti-nuclear
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

29
staining patterns. (D) Anti-cytoplasmic CD21-/lo B cells can react with well-defined
intracellular structures which include mitochondria (RA01 21- κ140, and CVID321 21-
κ82), the golgi (RA14 21- κ18), and cytoplasmic fibers (RA19 21- κ61, RA14 21- κ62, and
RA01 21- κ157 that is likely to recognize actin).
Figure 4. CD21-/lo naïve B cells fail to properly respond to BCR triggering. CD21-/lo
naïve B cells from (A) RA01 and (B) CVID321 patients fail to upregulate CD69 and
CD25 expression after stimulation with F(ab’2) anti-IgM, recombinant human CD40L,
and TLR9 agonist CpG. Total naive B cells containing both CD21-/lo and CD21+ B cells
were plated and dot plots show CD69 and CD25 expression after 2 days according to
gated CD21 expression. (C) BCR triggering in CD21-/lo naïve B cells does not properly
increase [Ca2+]i. [Ca2+]i was assessed by flow cytometry in CD21-/lo and CD21+ naive B
cells from a healthy donor (HD), RA01 and CVID321 patients. Both B cell populations
were loaded with Fluo-3 and expressed similar levels of surface IgM expression when
experiments were performed. Arrow indicates when F(ab’)2 anti-IgM was added. Data
from each group of individuals are representative of two-three independent experiments.
Figure 5. CD21-/lo naïve B cells do not proliferate after BCR and CD40 co-
triggering and are prone to cell death. (A) Total naive B cells from healthy donors
(HD), RA and CVID patients containing both CD21-/lo and CD21+ B cells were labeled
with CFSE and stimulated with F(ab’2) anti-IgM and/or recombinant human CD40L. Dot
plots show CD21 expression according to CFSE dilution after 3 days. CFSE dilution in
CD21+ but not in CD21-/lo B cells stimulated with F(ab’2) anti-IgM and recombinant
human CD40L reveals that some CD21+ B cells underwent up to two divisions whereas
CD21-/lo B cells did not divide. (B) Increased frequency of Annexin V+ propidium iodide
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

30
(PI)- early apoptotic, and Annexin V+ PI+ late apoptotic and dead cells in freshly isolated
CD21-/lo naïve B cells from RA and CVID patients compared to their CD21+ B cell
counterparts. (C) BCR, CD40 and/or TLR9 triggering do not prevent the apoptosis and
cell death of CD21-/lo naïve B cells. Total naive B cells containing both CD21-/lo and
CD21+ B cells from RA01 or CVID321 patients were plated and Annexin V and PI
staining was assessed after 12 hours. BCR, CD40 and TLR9 stimulation prevent late
apoptosis and cell death in CD21+ but not in CD21-/lo B cells.
Figure 6. Gene array comparisons of CD21-/lo and CD21+ naive B cells from RA
and CVID patients using the Affimetrix Human Genome U133 Plus 2.0 Array. (A)
Transcripts differentially expressed by CD21-/lo and CD21+ naive B cells from 2 RA and 3
CVID group Ia patients are shown. (B) Selected transcripts well known to B cell biology
or of potential interest are presented. Up- and down-regulated transcripts are indicated
in red and green, respectively. The magnitude of expression is depicted by the color
bar.
Figure 7. CD21-/lo B cells from RA and CVID patients display a phenotype
reflecting gene array profiling data. Overlays for the indicated markers were obtained
when CD27-depleted B cells isolated from the blood of patient RA01 were gated on
either CD19+CD21-/lo (bold line) or CD19+CD21+ (dashed line) B cells. The similar
phenotypes of CD21-/lo B cells from CVID patients and healthy donors are shown in
supplemental Figure S7-S8.
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom
For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom

For personal use only. by guest on October 13, 2011. bloodjournal.hematologylibrary.orgFrom
Copyright © 2022 FDOKUMEN



















