
Emel’yanova N. G., Pavlov D. A., Luong Thi Bich Thuan 2006. Hormonal stimulation of maturation and...
11
768 ISSN 0032-9452, Journal of Ichthyology, 2006, Vol. 46, No. 9, pp. 768–778. © Pleiades Publishing, Inc., 2006. Original Russian Text © N.G. Emel’yanova, D.A. Pavlov, Luong Thi Bich Thuan, 2006, published in Voprosy Ikhtiologii, 2006, Vol. 46, No. 6, pp. 807–817. For more than 15 years, the Russian–Vietnamese Tropical Center has carried out the works connected with the study of the ecology and taxonomic diversity of marine tropical fishes on the basis of the Russian– Vietnamese Intergovernmental Agreement. The devel- opment of the methods for regulating the processes of oocyte ripening in fishes is one of the directions of these researches. Methodical procedures of induction maturation will be apparently different in fishes with different types of oogenesis. Development of these pro- cedures in combination with the study of the specificity of development of sexual cells at finishing stages of oogenesis will make it possible to approach an under- standing of the features of the hormonal regulation of maturation processes. The methods for hormonal stimulation of matura- tion are described mostly for the freshwater and some marine fishes, which are the objects of aquaculture, for example, giltheard seabream Sparus auratus and Euro- pean seabass Dicentrarchus labrax (Moretti et al., 1999). Induction of the maturation of sexual products in many species of tropical fishes, which inhabit the coral reefs and are the objects of aquarium fish culture, can also be a genuine problem. This is connected with the fact that the market of marine ornamental fishes is based almost completely on the barbarous gathering of animals from nature, undermining the natural balance formed in populations, and artificial rearing can signif- icantly decrease the strain on fish communities (McAl- lister, 1999; Holt and Riley, 2000; Wabnitz et al., 2003). In recent years, the experiments have been carried out on the prolonged keeping of marine ornamental fishes under controlled conditions to stimulate natural spawn- ing. First results were obtained on breeding the larvae of some species under artificial conditions (Danilowicz and Brown, 1992; Holt and Riley, 2000; Olivotto et al., 2003). Hormonal stimulation of maturation is, in many cases, the only method for obtaining mature sexual products (Wabnitz et al., 2003). The species from the genus Zebrasoma, family of surgeon fishes (Acanthuridae) are common inhabitants of sea aquariums. The information about the biology of fishes from this genus is little and fragmentary. They are known to be widely distributed in seas with low lat- itudes and inhabit lagoons and coral reefs. They feed on algae and detritus. They spawn pelagic eggs (Randall and Lim, 2000). The spawning in the Indian and Pacific oceans occurs at a depth about 10 m into the reef zone at daytime (Robertson, 1983). In Vietnam, Z. scopas is a widely distributed species, which inhabits coral reefs in the South China Sea and is the object of uncontrolled fishing by the native population. This work was aimed at the development of the methods of hormonal stimulation of Z. scopas matura- tion, the study of the specificity of oocyte development at late stages of oogenesis, and the evaluation of the quality of ovulated oocytes. MATERIAL AND METHODS The work was carried out at the Base of the Coastal Department, Russian–Vietnamese Tropical Research Hormonal Stimulation of Maturation and Ovulation of Oocytes in Zebrasoma scopas (Acanthuridae) N. G. Emel’yanova a , D. A. Pavlov a , and Luong Thi Bich Thuan b a Moscow State University, Vorob’evy gory, Moscow, 119899 Russia b Coastal Department, Russian–Vietnamese Tropical Research and Technological Center, Nha Trang, Vietnam E-mail: [email protected] Received December 29, 2005 Abstract—The experiments were carried out on hormonal stimulation of oocyte maturation in Zebrasoma sco- pas from the South China Sea, Vietnam. Three variants of surfagon injections were studied: 1—double injec- tions (5 + 20 µ g/kg of fish body weight); 2—double injections (2 + 8 µ g/kg); and 3—single injections (20 µ g/kg). The time interval between two injections comprised 15–24 h. Ovulation of oocytes in variants 1 and 2 was observed in most (67%) females 33–47 h after the first injection. The increase of the time interval between injections I and II was followed by the decrease of the interval between injection II and ovulation. In variant 3, oocytes ripened but ovulation was absent. The oocytes possessed with the competence for maturation are always present in the ovaries because of a continuous type of oogenesis. The morphological changes in oocytes in the process of maturation were observed. Ovulated oocytes could be stored in the ovary cavity no more than 4 h; the number of embryos with normal cleavage decreased during this time from 90 to 53%. DOI: 10.1134/S0032945206090098
-
Upload
moscowstate -
Category
Documents
-
view
0 -
download
0
Transcript of Emel’yanova N. G., Pavlov D. A., Luong Thi Bich Thuan 2006. Hormonal stimulation of maturation and...

768
ISSN 0032-9452, Journal of Ichthyology, 2006, Vol. 46, No. 9, pp. 768–778. © Pleiades Publishing, Inc., 2006.Original Russian Text © N.G. Emel’yanova, D.A. Pavlov, Luong Thi Bich Thuan, 2006, published in Voprosy Ikhtiologii, 2006, Vol. 46, No. 6, pp. 807–817.
For more than 15 years, the Russian–VietnameseTropical Center has carried out the works connectedwith the study of the ecology and taxonomic diversityof marine tropical fishes on the basis of the Russian–Vietnamese Intergovernmental Agreement. The devel-opment of the methods for regulating the processes ofoocyte ripening in fishes is one of the directions ofthese researches. Methodical procedures of inductionmaturation will be apparently different in fishes withdifferent types of oogenesis. Development of these pro-cedures in combination with the study of the specificityof development of sexual cells at finishing stages ofoogenesis will make it possible to approach an under-standing of the features of the hormonal regulation ofmaturation processes.
The methods for hormonal stimulation of matura-tion are described mostly for the freshwater and somemarine fishes, which are the objects of aquaculture, forexample, giltheard seabream
Sparus auratus
and Euro-pean seabass
Dicentrarchus labrax
(Moretti et al.,1999). Induction of the maturation of sexual products inmany species of tropical fishes, which inhabit the coralreefs and are the objects of aquarium fish culture, canalso be a genuine problem. This is connected with thefact that the market of marine ornamental fishes isbased almost completely on the barbarous gathering ofanimals from nature, undermining the natural balanceformed in populations, and artificial rearing can signif-icantly decrease the strain on fish communities (McAl-lister, 1999; Holt and Riley, 2000; Wabnitz et al., 2003).In recent years, the experiments have been carried out
on the prolonged keeping of marine ornamental fishesunder controlled conditions to stimulate natural spawn-ing. First results were obtained on breeding the larvaeof some species under artificial conditions (Danilowiczand Brown, 1992; Holt and Riley, 2000; Olivotto et al.,2003). Hormonal stimulation of maturation is, in manycases, the only method for obtaining mature sexualproducts (Wabnitz et al., 2003).
The species from the genus
Zebrasoma
, family ofsurgeon fishes (Acanthuridae) are common inhabitantsof sea aquariums. The information about the biology offishes from this genus is little and fragmentary. Theyare known to be widely distributed in seas with low lat-itudes and inhabit lagoons and coral reefs. They feed onalgae and detritus. They spawn pelagic eggs (Randalland Lim, 2000). The spawning in the Indian and Pacificoceans occurs at a depth about 10 m into the reef zoneat daytime (Robertson, 1983). In Vietnam,
Z. scopas
isa widely distributed species, which inhabits coral reefsin the South China Sea and is the object of uncontrolledfishing by the native population.
This work was aimed at the development of themethods of hormonal stimulation of
Z. scopas
matura-tion, the study of the specificity of oocyte developmentat late stages of oogenesis, and the evaluation of thequality of ovulated oocytes.
MATERIAL AND METHODS
The work was carried out at the Base of the CoastalDepartment, Russian–Vietnamese Tropical Research
Hormonal Stimulation of Maturation and Ovulation of Oocytes in
Zebrasoma scopas
(Acanthuridae)
N. G. Emel’yanova
a
, D. A. Pavlov
a
, and Luong Thi Bich Thuan
b
a
Moscow State University, Vorob’evy gory, Moscow, 119899 Russia
b
Coastal Department, Russian–Vietnamese Tropical Research and Technological Center, Nha Trang, VietnamE-mail: [email protected]
Received December 29, 2005
Abstract
—The experiments were carried out on hormonal stimulation of oocyte maturation in
Zebrasoma sco-pas
from the South China Sea, Vietnam. Three variants of surfagon injections were studied: 1—double injec-tions (5 + 20
µ
g/kg of fish body weight); 2—double injections (2 + 8
µ
g/kg); and 3—single injections(20
µ
g/kg). The time interval between two injections comprised 15–24 h. Ovulation of oocytes in variants 1 and2 was observed in most (67%) females 33–47 h after the first injection. The increase of the time interval betweeninjections I and II was followed by the decrease of the interval between injection II and ovulation. In variant 3,oocytes ripened but ovulation was absent. The oocytes possessed with the competence for maturation arealways present in the ovaries because of a continuous type of oogenesis. The morphological changes in oocytesin the process of maturation were observed. Ovulated oocytes could be stored in the ovary cavity no more than4 h; the number of embryos with normal cleavage decreased during this time from 90 to 53%.
DOI:
10.1134/S0032945206090098

JOURNAL OF ICHTHYOLOGY
Vol. 46
No. 9
2006
HORMONAL STIMULATION OF MATURATION AND OVULATION 769
and Technological Center (City of Nha Trang, Vietnam)from February to April 2005. Preliminary data on hor-monal stimulation of
Z. scopas
were obtained in 2002and 2004.
Fish for the study were captured with nets in NhaTrang Bay of the South China Sea. The time from thecapture to fish delivery to the laboratory averaged sev-eral hours. The fishes were kept in an aquaria room innetting cages 80
×
80
×
100 cm established in concretetanks up to 3 m
3
in volume. Water circulated in everytank in a closed cycle (water replacement comprised 4–8 m
3
/h), passing through the filter and representing apile of perforated trays with a coral crumb. The watertemperature in the tanks was 25–27
°
C and the salinitywas 33–35‰.
The sum total of 168 specimens was used in theexperiments. Some females were used for biologicalanalysis. Total length to the end of a caudal fin (TL),body length to the end of scale covering (
l
), total bodyweight (
G
), body weight of the gutted fish (
G
1
), andgonad weight (
g
) were determined (table).
Preliminary experiments carried out in 2002 and2004 demonstrated that spawners of
Z. scopas
did notmature without hormonal stimulation, being kept underartificial conditions; ovulated oocytes could beobtained using different doses of surfagon (30–80
µ
g/kg) injected twice with the interval between theinjections at about 15 h. Surfagon was sometimes usedtogether with neuroleptic eglonil. Because the applica-tion of eglonil had no effect on the results, it was notused in 2005.
Taking into consideration our preliminary results,we tested the scheme of the hormonal stimulation of
Z. scopas
maturation and ovulation in three directions:1—determining the lowest possible efficient doses ofsurfagon; 2—studying the influence of the intervalbetween the preliminary (I) and booster (II) injectionson the time of oocyte maturation and ovulation; 3—assessing the probability of single hormonal stimula-tion.
The summary dose of surfagon varied from 10 to25
µ
g/kg of fish weight. Three variants of hormonalstimulation of oocyte maturation were used. Variant 1(“common” dose of surfagon): 5
µ
g/kg as a preliminaryinjection and 20
µ
g/kg as a booster injection. Variant 2(“small” dose of surfagon): 2
µ
g/kg and 8
µ
g/kg,respectively, as injections I and II. Variant 3: a single
injection with surfagon dose 20
µ
g/kg. The intervalbetween repeated injections varied from 15 to 24 h.
Injections were made several hours after the deliv-ery of the fishes to the aquaria room. Surfagon wasinjected to the body cavity under the pectoral fin.Because there was no clear sexual dimorphism, theindividuals of both sexes were kept together in the pre-liminary experiments in 2002 and 2004 and at thebeginning of the work in 2005, and the injections werecarried out according to the same scheme. Then, wefound some differences in the morphology of the uro-genital region in males and females (see sectionResults), and fishes were examined immediately afterdelivery to the laboratory and sorted by sex as was pos-sible. Females were placed in cages, separate from themales, and some of them were labeled through remov-ing one or several rays in the dorsal or anal fin.
The ovulation in the result of oocyte maturationunder natural conditions (“natural” ovulation) wasobserved in many females immediately after theirdelivery to the aquaria room. In such cases, the eggswere stripped, and females were used for subsequenthormonal injections.
Injection of males in preliminary experiments (2002and 2004) demonstrated that spermiation rarelyoccurred, usually after injection II. Because spermiaobtained from the testes activated well, the males werenot injected in 2005.
To determine the moment of egg ovulation after hor-monal stimulation, all females were examined every1.5–2.0 h within 7–10 h after injection II (in the case ofdouble injections) or 20 h after a single injection. Theyellow-pinkish convexity covered with a thin mem-brane appeared immediately before the ovulation in theregion of genital opening in a female. When pressing afemale abdomen, the membrane prevented, in somecases, the output of ovulated eggs. Because of this, itwas carefully punctured with a sharpened item, andthen the eggs were stripped. This apparently operationhad no effect on the process of egg maturation: themembrane was formed again several hours later.
To study the size composition of oocytes and theirmorphological features in the process of maturationafter injections, the intravital samples of oocytes weretaken in labeled individuals before injections I and IIand in the interval between injection II and ovulation.The analysis of the size composition of oocytes wascarried out with the help of a system for image analysis
Biological characters of
Zebrasoma scopas
females (
n
= 17)
Characters TL, cm
l
, cm
G
, g
G
1
, g
g
, g GSI, %
M
+
σ
13.0
±
1.1 11.2
±
0.9 56.1
±
11.4 49.0
±
10.6 1.47
±
0.77 3.0
±
1.7
Lim
10.6–14.5 9.3–12.5 35.0–78.0 32.0–70.0 0.25–3.56 0.8–7.6
Note: M +
σ
, average value and standard deviation;
Lim
, limits; GSI (gonadosomatic index):
g
/
G
1
×
100, where
g
is gonad weight and
G
1
is body weight without intestines.

770
JOURNAL OF ICHTHYOLOGY
Vol. 46
No. 9
2006
EMEL’YANOVA et al.
that included the microscope Nikon Eclipse E-200 withan analog black and white camera and a computer.Oocyte diameters were measured using the computerprogram ImageJ.
The oocytes at stages before the beginning of thefusion of yolk granules were immersed in Serra solu-tion to study their morphology (Makeeva andEmel’yanova, 1990). The morphology of oocytes atlater stages of maturation (those obtaining the transpar-ency) was studied after drawing out of the gonad andplacement into the physiological solution.
The quality of ovulated oocytes stripped from thefemale body was evaluated by the results of their artifi-cial insemination with male sperm obtained after draw-ing out the testes. The testes were disintegrated, pressedthrough the gauze to the weighing bottle, and seawaterwas added to activate the spermia. The eggs wereinseminated with the “semidry” method. The mobilityof spermatozoa in the smear of sperm activated in thedrop of seawater was preliminarily recorded under the
microscope. The quality of ovulated eggs was evaluated3–5 h after insemination at the morula stage (at 25
°
C),examining 100 eggs from every female under the bin-ocular microscope and calculating the part of the eggswith normal cleavage.
To obtain eggs of high quality, we must know duringwhat time after ovulation the eggs retain their viabilitybeing in the female body. An experiment with strippingand insemination of small portions of oocytes immedi-ately after ovulation and 4 and 8 h after it was carriedout in 11 females of
Z. scopas.
RESULTS
Sexual Dimorphism and External Structure of Gonads
The genital opening of the female of
Z. scopas
(Fig. 1a) was situated between the ventral and anal fins.Before the completion of oocyte maturation, it had asimilar size with the urogenital opening of males or was
(a) (b)
(c) (d)
(e) (f)
Fig. 1.
Zebrasoma scopas
: a, mature female; b, ventral part of the male body; c, ventral part of the female body having gonads atmaturity stage IV; d, ventral part of the female body with ovulated eggs; e, ovaries; f, testes.

JOURNAL OF ICHTHYOLOGY
Vol. 46
No. 9
2006
HORMONAL STIMULATION OF MATURATION AND OVULATION 771
only insignificantly greater (Figs. 1b and 1c). Becauseother signs of sexual dimorphism were absent, it wasrelatively difficult to differentiate fishes according theirsex. The sexual opening of the female increased beforethe ovulation, and the urogenital region obtained a
slightly convex character (Fig. 1d). Such females dif-fered easily relative to males. Before the ovulation, thesexual opening was closed with a thin membrane,which was torn immediately before the spawning andwas formed again soon after it. The ovaries are short-
0 10 20 30 40 50 60 70
Time, h
n
= 743%III43%
(e)
0 10 20 30 40 50 60 70
n
= 1258%III100%
(d)
0 10 20 30 40 50 60 70
n
= 4100%III
(c)
0 10 20 30 40 50 60 70
n
= 650%III100%
(b)
0 10 20 30 40 50 60 70
n
= 786%III86%
(a)
Ovulation
Prolonged ovulation
706050403020100
n
= 6
Ovulation
67
%
III100%
706050403020100
n
= 967
%
III100%
706050403020100
n
= 4100
%
III
(a)
(b)
(c)
Time, h
Fig. 2.
The schemes of hormonal stimulation of
Zebrasoma scopas
through double injections with the common dose (5
µ
g/kg +20
µ
g/kg) of surfagon. a–e—fish groups. Here and hereinafter: Roman numerals designate injections I and II. Total number offemales (
n
) and the part of females (% from their total number for particular group) with natural ovulation and induced ovulationare presented. Time is calculated from the moment of fish delivery to the aquaria room.
Fig. 3.
The schemes of hormonal stimulation of
Zebrasoma scopas
through double injections with a small dose (2
µ
g/kg + 8
µ
g/kg)of surfagon.

772
JOURNAL OF ICHTHYOLOGY
Vol. 46
No. 9
2006
EMEL’YANOVA et al.
ened, paired, and join together in the lower part approx-imately for one-third of their length, usually yellow incolor (Fig. 1e). The testes are also paired and growntogether in the lower part, similar to the ovaries(Fig. 1f).
Hormonal Stimulationof the Maturation of Sexual Products
Variant 1
(Fig. 2). The fishes in different groups dif-fered in the interval between the delivery to the aquariaroom and the preliminary injection, which varied from2 (Fig. 2b) to 9 h (Fig. 2e). Additionally, “natural” ovu-lation was observed in a great number of individuals(43–100%) in 4 out of 5 groups.
Ovulation induced with hormonal injections wasobserved in 64% of the individuals. When the intervalbetween injections I and II increased, the intervalbetween injection II and ovulation decreased. Particu-larly, the ovulation began in different individuals 23–32 hafter injection II, if the interval between the injectionswas 15 h (Fig. 2a), and after 11–18 h, if the intervalbetween the injections was 24 h (Fig. 2e).
It should be noted that the time of the beginning ofovulation, counted from the moment of preliminaryinjection, varied much less and ranged in most fishesfrom 33 (Fig. 2) to 39 h (Fig. 2b). The time before theinduced ovulation varied even less, if we calculate thetime from natural ovulation: induced ovulation wasobserved in most fish after 41–42 h.
Variant 2
(Fig. 3). The fishes in different groups dif-fered in the time between the delivery to the aquariaroom and injection I and in the interval between injec-tions I and II. Natural ovulation was observed in allfishes from the 1st and 2nd groups (Figs. 3a, 3b),whereas it was not observed in the fishes from the 3rdgroup (Fig. 3c).
Despite a more than twofold decrease in the surf-agon dose, oocyte ovulation, caused by hormonal stim-ulation, occurred in most fishes (67–100%, averaged
74%) from all groups. It was observed at approximatelythe same dates as in fishes from variant 1. The increaseof the interval between injections I and II was accom-panied with the decrease of the interval between injec-tion II and ovulation. The total duration of intervalsbetween injection I and ovulation and between naturaland induced ovulation was approximately the same asin variant 1. Fractional (net) fecundity varied from1160 to 19 042 eggs and averaged 7584 (
n
= 9).
Variant 3
(Fig. 4). The completion of oocyte matu-ration and the appearance (only in 2 females) of a fewovulated oocytes that were recorded in both groups dif-fered in the interval between the delivery to the aquariaroom and injection I and in the part of fishes with natu-ral ovulation. These phenomena were observed atapproximately the same time as the individuals withdouble injections.
The Diameter Distribution of Oocytes
The diameter distribution of oocytes in separatefemales demonstrated the domination of smaller cells,i.e., of junior age groups. The oocytes of intermediateclasses between the minimum and maximum ones werepresented (Fig. 5). The oocytes of the older age grouphad a diameter about 350 µm in the intact females(Fig. 5a). Such oocytes had the competence for matura-tion and could ovulate after hormonal injections. Someincrease of diameter occurred in oocytes of the olderage group before injection II (Fig. 5b). Size composi-tion of oocytes in the ovary after ovulation was mostlysimilar to that of the intact females. Some number ofresidual oocytes, which did not ovulate for some rea-son, was usually presented in the gonad. The size ofsuch cells was usually much greater than the size ofmost oocytes (Fig. 5c). The diameter of ovulatedoocytes varied from 569 to 846 µm and averaged632 µm (n = 322).
10 20 30 40 50 60 700
n = 10I100%
45% I
10 20 30 40 50 60 700
n = 11
Time, h
(a)
(b)
Completion of oocyte maturation
Prolonged completion of oocyte maturation
Fig. 4. The schemes of hormonal stimulation of Zebrasoma scopas through single injections (20 µg/kg) of surfagon.

JOURNAL OF ICHTHYOLOGY Vol. 46 No. 9 2006
HORMONAL STIMULATION OF MATURATION AND OVULATION 773
Morphological Transformationof Oocytes in the Process of Maturation
Oocytes of the older size group were usually repre-sented in the intact females by the cells with a centralnucleus. Numerous small oil globules in the cytoplasmwere distributed in the form of a wide ring surroundingthe nucleus (Fig. 6a). The processes of nucleus migra-tion to an animal pole and fusion of oil globules beganafter the first injection. Just before injection II, thenucleus was situated, as a rule, some distance awayfrom the animal pole, and oil globules became enlargedand localized in the form of a narrower ring around thenucleus (Figs. 6b, 6c). After the booster injection, theprocesses of further migration of the nucleus to the ani-mal pole continued. The localization of the nucleusdirectly on the animal pole was observed only in sepa-rate cases; this state was apparently very short. Then thenuclear membrane disintegrated, the nucleus contoursof this result became less clear, and karyoplasm andcytoplasm mixed. The fusion and enlargement of oilglobules continued in parallel (Fig. 6d, 6e) and finishedwith the formation of one large globule (Fig. 6f). As arule, the process of the homogenization (fusion) of yolkgranules began at the same time, though most yolkremained discrete, and oocytes looked dark in this rela-tion (Figs. 6f, 6g). Oocyte size increased in the hydra-tion result, as these stages passed. Oocytes hydratedeven more by the moment of ovulation and becametransparent in the result of the complete fusion of yolkgranules (Fig. 6h).
Effect of Delay on the Quality of Ovulated Oocytes in the Female Body
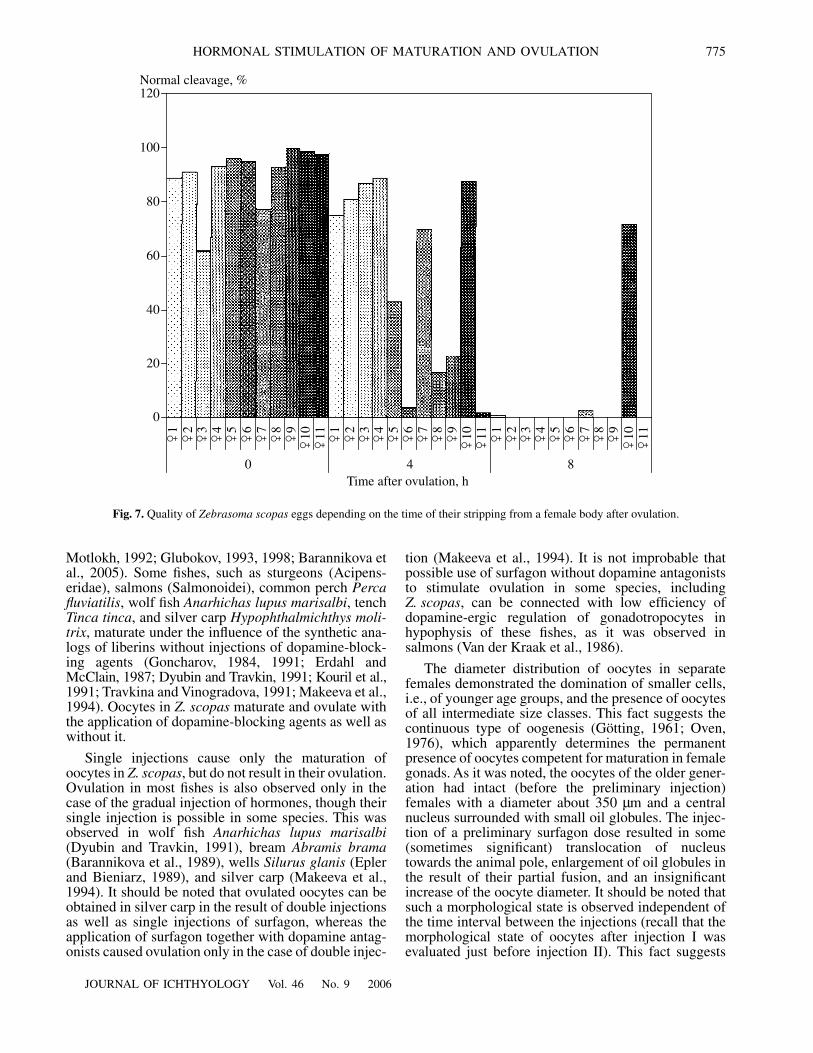
Repeated stripping and insemination of the part ofovulated oocytes from the same females demonstratedthat the number of eggs with normal cleavage averaged90% (σ = 11.2) just after ovulation and 53% (σ = 35.4)and 7% (σ = 20.7), respectively, 4 and 8 h after ovula-tion (Fig. 7). It is interesting to note that the quality ofoocytes in one individual remained high even 8 h afterthe ovulation. These data suggest significant differencein the quality of ovulated oocytes in separate females.
DISCUSSION
The results of the experiments demonstrated that thescheme with the double injection of surfagon should beused to stimulate the maturation and ovulation ofoocytes in Z. scopas. The summary dose of hormonecan vary significantly, but it would be more efficient touse minimal doses: 2 µg/kg as a preliminary injectionand 8 µg/kg as a booster dose (summary dose10 µg/kg). The interval between the injections can bedecreased or increased in certain limits. It is good tobear in mind that ovulation occurred after a longertime interval after injection II, if we decrease thistime, whereas total time from injection I to ovulationvaries insignificantly.
Usually, about 37 (33–39) h pass from the prelimi-nary injection to ovulation, but this interval couldincrease to 47 h in some females (it varied from 22 to48 h in preliminary experiments in 2002 and 2004). Theindividuals, in which oocytes maturated under naturalconditions and the eggs were obtained soon after theirdelivery to the aquaria room (for “natural” ovulation),could produce ovulated oocytes again after hormonalstimulation. The presence of natural ovulation did notaffect the time of ovulation induced with hormonalstimulation. Repeated ovulation occurred, as a rule, 41–
60
50
40
30
20
10
0
n = 104
(a)
Number of oocytes, %
50
40
30
20
10
0
n = 88
(b)
50
40
30
20
10
0
n = 70
(c)
100 200 300 400 500Diameter of oocytes, µm
Fig. 5. The diameter of oocytes in the sample from gonadsof female Zebrasoma scopas: a, before injection I; b, beforeinjection II; c, after ovulation.

774
JOURNAL OF ICHTHYOLOGY Vol. 46 No. 9 2006
EMEL’YANOVA et al.
42 h after the natural ovulation in most females inde-pendent of the injection scheme (excluding singleinjection) and used doses of surfagon. On this basis wecan assume that the interval between the spawning ofthe following egg portions is about 2 days in a naturalenvironment. Analogous data on the spawning with aninterval of 1–2 days are presented for the fishes with acontinuous type of oogenesis in Oven’s monograph(1976).
The synthetic analogs of gonadotropin-releasinghormones (Gn-Rh), which regulate synthesis and theremoval of gonadotropins from the hypophysis, areused more and more often in the last decade of aquac-ulture to stimulate maturation of sexual cells (Lam,
1982; Goncharov, 1984, 1991; Barannikova et al.,1989, 2005; Mikodina et al., 1989; Godukhin and Mot-lokh, 1992; Glubokov et al., 1994; Nocillado et al.,1999; Poliakova et al., 2002). The Gn–Rh are subdi-vided to liberins, stimulating the activity of gonadotropo-cytes in the hypophysis and statins and inhibiting thisactivity (Rozen, 1994). It is known that dopamine is a sta-tin or gonadotropin-releasing-inhibiting factor (Changand Peter, 1983). In this relation, the use of syntheticanalogs of releasing hormones in the practice of artifi-cial fish breeding is accompanied by the injection tofishes of substances blocking the dopamine receptors inthe hypophysis (De Lee et al., 1988; Epler and Bien-iarz, 1989; Barannikova et al., 1991; Godukhin and
0.1 mm
21
3
1
12
2
22
2
2
3
3
4
2
6
5
(a) (b) (c)
(d) (e) (f)
(g) (h)
Fig. 6. The state of Zebrasoma scopas oocytes: a, before injection I; b, c, before injection II; d–g, after injection II, h, ovulatedoocyte. Oocytes (a−e) are clarified in Serra’s solution. 1, nucleus; 2, oil globule; 3, yolk granule; 4, fused yolk granules; 5, micro-pyle; 6, cortical alveole.
JOURNAL OF ICHTHYOLOGY Vol. 46 No. 9 2006
HORMONAL STIMULATION OF MATURATION AND OVULATION 775
Motlokh, 1992; Glubokov, 1993, 1998; Barannikova etal., 2005). Some fishes, such as sturgeons (Acipens-eridae), salmons (Salmonoidei), common perch Percafluviatilis, wolf fish Anarhichas lupus marisalbi, tenchTinca tinca, and silver carp Hypophthalmichthys moli-trix, maturate under the influence of the synthetic ana-logs of liberins without injections of dopamine-block-ing agents (Goncharov, 1984, 1991; Erdahl andMcClain, 1987; Dyubin and Travkin, 1991; Kouril et al.,1991; Travkina and Vinogradova, 1991; Makeeva et al.,1994). Oocytes in Z. scopas maturate and ovulate withthe application of dopamine-blocking agents as well aswithout it.
Single injections cause only the maturation ofoocytes in Z. scopas, but do not result in their ovulation.Ovulation in most fishes is also observed only in thecase of the gradual injection of hormones, though theirsingle injection is possible in some species. This wasobserved in wolf fish Anarhichas lupus marisalbi(Dyubin and Travkin, 1991), bream Abramis brama(Barannikova et al., 1989), wells Silurus glanis (Eplerand Bieniarz, 1989), and silver carp (Makeeva et al.,1994). It should be noted that ovulated oocytes can beobtained in silver carp in the result of double injectionsas well as single injections of surfagon, whereas theapplication of surfagon together with dopamine antag-onists caused ovulation only in the case of double injec-
tion (Makeeva et al., 1994). It is not improbable thatpossible use of surfagon without dopamine antagoniststo stimulate ovulation in some species, includingZ. scopas, can be connected with low efficiency ofdopamine-ergic regulation of gonadotropocytes inhypophysis of these fishes, as it was observed insalmons (Van der Kraak et al., 1986).
The diameter distribution of oocytes in separatefemales demonstrated the domination of smaller cells,i.e., of younger age groups, and the presence of oocytesof all intermediate size classes. This fact suggests thecontinuous type of oogenesis (Götting, 1961; Oven,1976), which apparently determines the permanentpresence of oocytes competent for maturation in femalegonads. As it was noted, the oocytes of the older gener-ation had intact (before the preliminary injection)females with a diameter about 350 µm and a centralnucleus surrounded with small oil globules. The injec-tion of a preliminary surfagon dose resulted in some(sometimes significant) translocation of nucleustowards the animal pole, enlargement of oil globules inthe result of their partial fusion, and an insignificantincrease of the oocyte diameter. It should be noted thatsuch a morphological state is observed independent ofthe time interval between the injections (recall that themorphological state of oocytes after injection I wasevaluated just before injection II). This fact suggests
20
Normal cleavage, %
0
40
60
80
100
120
�1
�2
�3
�4
�5
�6
�7
�8
�9
�10
�11 �1
�2
�3
�4
�5
�6
�7
�8
�9
�10
�11 �1
�2
�3
�4
�5
�6
�7
�8
�9
�10
�11
0 4 8Time after ovulation, h
Fig. 7. Quality of Zebrasoma scopas eggs depending on the time of their stripping from a female body after ovulation.

776
JOURNAL OF ICHTHYOLOGY Vol. 46 No. 9 2006
EMEL’YANOVA et al.
that a preliminary dose of hormone only initiates theprocesses of maturation of sexual cells. It is known forsome objects of aquaculture that polarization of oocytesis observed after a preliminary injection of hormone, andthe nucleus migrates to the animal pole to the region ofmicropyle close to the cell membrane (Lemanova andSakun, 1975; Kazanskii et al., 1978; Duvarova, 1981;Val’ter, 1984; Makeeva and Emel’yanova, 1990). Thishappens in Z. scopas after injection II.
Judging by gradually increased sizes of oocytes inZ. scopas, the process of further fusion of oil globulesand cell hydration run in parallel. Some data (Alekseevand Alekseeva, 1996) suggest that oocyte diameter insome species of marine fishes does not change in theprocess of the fusion of oil globules and increases onlyin the result of the fusion (homogenization) of yolkgranules accompanied with hydration. After formationof a single oil granule, further hydration is observed inZ. scopas, yolk granules fuse completely, and oocytesbecome transparent in this result.
The quality of ovulated oocytes is important for theartificial reproduction of fishes. The quality of eggs candecrease quickly, if the moment of ovulation is deter-mined incorrectly. It is known that time, during whichovulated oocytes retain the capacity for fertilization andnormal development, can vary significantly in differentspecies (Ginzburg, 1968; Korovina, 1986; Pavlov et al.,2004). If ovulated eggs can be stored in vivo in salmo-nids over several days without damage, for example,more than 20 days in coho salmon Oncorhynchuskisutch (Fitzpatrick et al., 1987), and during 10–12 h incold-water marine fishes, turbot Psetta maxima andhalibut Hippoglossus hippoglossus (McEvoy, 1984;Bromage et al., 1994). The decrease of egg quality wasrecorded in carp Cyprinus carpio within 1.5–2.0 h(Korovina, 1986). Oocyte quality decreased in silvercarp during the first hour after ovulation (Makeeva etal., 1987). A short period of viability of the ovulatedeggs (about one hour) was also recorded in striped bassMorone saxatilis (Stevens, 1966) and Prochilodusmarggravii (Rizzo et al., 2003). It is considered thatdeterioration of the quality of ovulated oocytes in thecase of their delay in the ovarian cavity is caused byasphyxia in the result of the absence of a connectionwith the cells of the connective-tissue theca, in whichthe blood vessels run (Korovina, 1986; Makeeva et al.,1987). The decrease of the quality of ovulated oocytesin Z. scopas (by 38%) was observed 4 h after theirkeeping in a female body, though their quality remainedrelatively high in more than half of the individuals. Thetime of delay of ovulated oocytes in the female bodywithout damage was relatively high in Z. scopas ascompared with the other warmwater species.
Hence, maturation and ovulation can be induced inmature females Z. scopas by a double surfagon injec-tion. Because this species is characterized by a contin-uous type of oogenesis, the oocytes possessing the
competence for maturation are constantly presented ingonads.
ACKNOWLEDGMENTS
We are grateful to academician D.S. Pavlov, Scien-tific supervisor of the Tropical Center, Institute of Ecol-ogy and Evolution, Russian Academy of Sciences; Pro-fessor G.G. Novikov, the supervisor of project EcolanE.3.2.2 “Taxonomic Diversity and Ecology of MarineHydrobionts;” codirectors of the Primorskii Depart-ment, the Russian–Vietnamese Tropical Research andTechnological Center V.K. Nezdolii; and the following:Nguyen Van Doan; head of the Ecology DepartmentNgo Chi Thien; researcher of the Ecology DepartmentD.T.H. Ien; researcher of the Tropical Center NguyenDang Thanh; and the researchers of the IchthyologyDepartment, Biological Faculty, Moscow State Univer-sity A.M. Shadrin and S.A. Ponomarev for assistance inarrangement of the work.
This work was supported in part by the RF PresidentCouncil for Grants and State Support of the LeadingScientific Schools (state contract with Rosnaukano. 02.445.11.7064).
REFERENCES1. F. E. Alekseev and E. I. Alekseeva, Determination of
Gonad Maturity Stages and the Study of Sexual Cycles,Fecundity, Egg Production, and Rate of Sexual Matura-tion in Marine Commercial Fish. Methodical Guide(AtlantNIRO, Kaliningrad, 1996).
2. I. A. Barannikova, L. V. Bayunova, N. N. Kolmakov, andT. B. Semenkova, “The Dynamics of Steroid Hormonesin Blood Under Hormonal Stimulation of Maturation inthe Northern Dvina Sterlet Acipenser ruthenus,” Vopr.Ikhtiol. 45 (1), 131–138 (2005) [J. Ichthyol. 45 (1), 124–131 (2005)].
3. I. A. Barannikova, A. A. Boev, V. P. Dyubin, andB. G. Travkin, “The Use of Hormonal Impacts for theInduction of Maturation in Cyprinid Fish,” in Proceed-ings of VII All-Union Conference on the EcologicalPhysiology and Biochemistry in Fish (Yaroslavl, 1989),pp. 30–31.
4. I. A. Barannikova, V. P. Dyubin, S. G. Kiseleva, andL. V. Bayunova, “On the Hypothalamic Regulation ofthe Function of Gonadotropocytes of the Hypophysis inPacific Salmon,” in Reproductive Physiology of Fish:Abstracts of Papers (AN BSSR, Mezhved. Ikhtiol.Komis., Minsk, 1991).
5. N. Bromage, M. Bruce, N. Basavaraja, et al., “Egg Qual-ity Determinants in Finfish: The Role of Overripeningwith Special Reference to the Timing of Stripping in theAtlantic Halibut Hippoglossus hippoglossus,” J. WorldAquac. Soc. 25 (1), 13–21 (1994).
6. J. P. Chang and K. E. Peter, “Effects of Pimiside and desGly10,(D-Ala6)Luteinizing Hormone-Releasing Hor-mone Ethylamide on Serum Gonadotropin Concentra-tions Germinal Vesicle Migration and Ovulation inFemale Gold Fish, Carassius auratus,” Gen. Comp.Endocrinol. 52 (1), 30–37 (1983).

JOURNAL OF ICHTHYOLOGY Vol. 46 No. 9 2006
HORMONAL STIMULATION OF MATURATION AND OVULATION 777
7. B. S. Danilowicz and C. L. Brown, Rearing Methods forTwo Damselfish Species: Dascyllus albisella (Gill) andD. aruanus (L.), Aquaculture 106, 141–149 (1992).
8. U. W. R. De Lee, Van’t C. Veer, et al., “The Dopaminer-gic Regulation of Gonadotropin-Releasing HormoneReceptor Binding in the Pituitary of the African Catfish,Clarias gariepinus,” Gen. Comp. Endocrinol. 2 (2),408–415 (1988).
9. A. S. Duvarova, “The Use of the Biopsy Method for theDetermination of Maturity of Oocytes and the Modifica-tion of the Schemes of Hypophyseal Injections in the Sil-ver Carp,” Tr. Nauchno-Issled. Inst. Ozern. Rechn.Rybn. Khoz., No. 160, 79–86 (1981).
10. V. P. Dyubin and B. G. Travkin, “Hormonal Stimulationof the Wolffish,” in The Reproductive Physiology ofFish: Abstracts of Papers (AN BSSR, Mezhved. Ikhtiol.Komis., Minsk, 1991).
11. P. Epler and K. Bieniarz, Gonad Maturation and Hor-monal Stimulation of Spawning in Wells (Silurus glanis(L.), Pol. Arch. Hydrobiol. 36 (36), 417–429 (1989).
12. D. Erdahl and J. McClain, Effect of LH-RH AnalogueTreatment on Egg Maturation (Ovulation) in Lake TroutBrood Stock, Prog. Fish-Cult. 49 (4), 276–279 (1987).
13. M. S. Fitzpatrick, C. B. Schreck, F. Ratti, and R. Chit-wood, “Viabilities of Eggs Stripped from Coho Salmonat Various Times After Ovulation,” Prog. Fish-Cult. 49,177–180 (1987).
14. A. S. Ginzburg, Fecundation in Fish and the Problem ofPolyspermy (Nauka, Moscow, 1968) [in Russian].
15. A. I. Glubokov, “Some Neurophysiological Prerequi-sites for Using Biologically Active Substances and Fac-tors for the Stimulation of Maturation in Fish,” in Bio-logically Active Substances and Factors in the Aquacul-ture (VNIRO, Moscow, 1993), pp. 3–28.
16. A. I. Glubokov, J. Kouril, E. V. Mikodina, and T. Barth,“Effect of Synthetic GhRH Analogues and DopamineAntagonists on the Maturation of Pacific Mullet, Mugilso-iuy Bas,” Aquacult. Fish. Manag. 25, 419–425(1994).
17. A. I. Glubokov, “Regulation of the Reproductive Func-tion Using Biologically Active Substances and Factors,”in Biological Foundations of Mariculture, Ed. byL.A. Dushkina (VNIRO, Moscow, 1998) pp. 205–215.
18. O. V. Godukhin and N. N. Motlokh, “Regulation of theGonadotrophic Function of Hypophysis in Bony Fish,”Usp. Sovrem. Biol. 112 (1), 115–129 (1992).
19. B. F. Goncharov, “Synthetic Analog of Luliberin—ANew Perspective Stimulator of Maturation of GenitalProducts in Acipenserids,” Dokl. Akad. Nauk SSSR 276(4), 1002–1006 (1984).
20. B. F. Goncharov, “Results and Perspectives of UsingSynthetic Analogs of Gonadotropin-Releasing Hor-mones in Fish Husbandry,” in The Reproductive Physiol-ogy of Fish: Abstracts of Papers (AN BSSR, Mezhved.Ikhtiol. Komis., Minsk, 1991).
21. K. S. Gotting, “Beitrage zur Kenntnis der Grundlagender Fortpflanzung und zur Fruchtböarkeitbestimmungbei marinen Teleosteern,” Helgoland. Wiss. Meersunter-such. 8 (1), 1–41 (1961).
22. G. J. Holt and C. M. Riley, “Laboratory Spawning ofCoral Reef Fishes: Effects of Temperature and Photope-riod,” UJNR Techn. Rep., No. 28, 33–38 (2000).
23. B. N. Kazanskii, Yu. A. Feklov, S. B. Podushka, andA. N. Molodtsov, “Express-Method for Determining theExtent of Gonad Maturity in Acipenserid Spawners,”Rybn. Khoz., No. 2, 24–27 (1978).
24. V. M. Korovina, “Aftereffects of “Overripening” of Eggsin Embryogenesis of Bony Fish,” Tr. Zool. Inst. Akad.Nauk SSSR 154, 115–123 (1986).
25. J. Kouril, T. Barth, J. Hamackova, et al., “The Compari-son of Effects of Two Superactive Gn-RH Analogues byMeans of Hormonal Induction of Ovulation in Tench(Tinca tinca (L.) in Artificial Propagation,” in Interna-tional Symposium on Reproductive Biology in Aquacul-ture (China, 1991), p. 54.
26. T. J. Lam, “Application of Endocrinology to Fish Cul-ture,” Can. J. Fish. Aquat. Sci. 133, 111–137 (1982).
27. N. A. Lemanova and O. F. Sakun, Methodical Guide onthe Hormonal Stimulation of Carp Spawners at EarlyObtaining of Eggs (GosNIORKh, Leningrad, 1975) [inRussian].
28. A. P. Makeeva and N. G. Emel’yanova, “A CytologicalStudy of Preovulation Oocytes of the Silver Carp and theUse of the Biopsy Method for Revealing Their State,”Vopr. Ikhtiol. 30 (3), 469–477 (1990).
29. A. P. Makeeva, N. G. Emel’yanova, N. V. Belova, andA. S. Ekutich, “The Use of the Synthetic Releasing Hor-mone for the Stimulation of Maturation and Spawning ofthe Silver Carp Hypophthalmichthys molitrix,” Vopr.Ikhtiol. 34 (1), 108–114 (1994).
30. A. P. Makeeva, N. G. Emel’yanova, and B. V. Verigin,“On the Quality of Eggs Produced by Far Eastern Her-bivorous Fish Under Conditions of Their Hatchery Pro-duction,” Vopr. Ikhtiol. 27 (5), 809–822 (1987).
31. D. E. McAllister, “Is Mariculture the Remedy to Prob-lems of Coral Reefs of Coastal Communities,” SPC LiveReef Inform. Bull. 5, 47–48 (1999).
32. L.-A. McEvoy, “Ovulatory Rhythms and Over-Ripeningof Eggs in Cultivated Turbot, Scophthalmus maximus,” J.Fish. Biol. 24, 437–448 (1984).
33. E. V. Mikodina, Ya. Kourzhil, A. I. Glubokov, et al.,“Synchronization of Maturation and Ovulation in Rain-bow Trout Using a Synthetic Analog of Salmon Gona-dotropin-Releasing Hormone GnRH (D-Tle6) -GnRH-Rro-Nhet and Inhibitor Dopamine - Metoclopramide,” inProceedings of the International Symposium on ModernProblems of Mariculture (VNIRO, Moscow, 1989),pp. 131–132.
34. A. Moretti, M. Pedini Fernandez-Criado, G. Cittolin,and R. Guidastri, Manual on Hatchery Production ofSeabass and Gilthead Seabream (FAO, Rome, 1999),Vol. 1.
35. J. N. Nocillado, J. P. van Eenennaam, and S. I. Doroshev,“Final Ovarian Maturation and Egg Quality in Hormon-ally-Induced White Sturgeon,” J. Appl. Ichthyol. 15, 319(1999).
36. I. Olivotto, M. Cardinali, L. Barbaresi, et al., “Coral ReefFish Breeding: The Secrets of Each Species,” Aquacul-ture 224 (1-4), 69–78 (2003).
37. L. S. Oven, Specific Features of Oogenesis and the Pat-tern of Spawning in Marine Fish (Naukova Dumka,Kiev, 1976) [in Russian].

778
JOURNAL OF ICHTHYOLOGY Vol. 46 No. 9 2006
EMEL’YANOVA et al.
38. D. Pavlov, E. Kjørsvik, T. Refsti, and Ø. Andersen,Brood Stock and Egg Production, in Culture of Cold-Water Marine Fish, Ed. by E. Moksness, E. Kjørsvik,and Y. Olsen (Blackwell Publ., Oxford, 2004), Ch. 5,pp. 129–203.
39. L. Poliakova, G. Artaly, and J. Giovannini, “The Use ofthe Synthetic Hormone LH- RH (d-ala6) for Reproduc-tive Control of Wild Caught and Captivity Raised Stur-geons, in “Proceedings of the 4th International Sympo-sium on Sturgeon, 2001,” Ed. by H. Rosenthal,R.M. Bruch, and F.P. Binkowski (Oshkosh, Wisconsin,USA, 2002), p. 160.
40. J. E. Randall and K. K. P. Lim, “A Checklist of the Fishesof the South China Sea,” Ruffles Bull. Zool. Suppl.,No. 8, 569–667 (2000).
41. E. Rizzo, H. P. Godinho, and Y. Sato, “Short-Term Stor-age of Oocytes from the Neotropical Teleost FishProchilodus marggravii,” Theriogenology 60 (6), 1059–1070 (2003).
42. D. R. Robertson, “On the Spawning Behavior andSpawning Cycles of Eight Surgeonfishes (Acanturidae)from Indo-Pacific,” Environ. Biol. Fishes 5, 193–223(1983).
43. V. B. Rozen, Essentials of Endocrinology (Mosk. Gos.Univ., Moscow, 1994) [in Russian].
44. R. E. Stevens, “Hormone-Induced Spawning of StripedBass for Reservoir Stocking,” Prog. Fish-Cult. 28, 19–28(1966).
45. G. L. Travkina and O. V. Vinogradova, “Comparison ofthe Action of Synthetic Analogs of Luliberin and Ace-tonized Hypopheses of the Carp on the State of Femalesof River Perch,” in The Reproductive Physiology of Fish:Abstracts of Papers (AN BSSR, Mezhved. Ikhtiol.Komis., Minsk, 1991).
46. G. A. Val’ter, Candidate’s Dissertation in Biology(VNIRO, Moscow, 1984).
47. G. Van der Kraak, E. M. Donaldson, and J. P. Chang,“Dopamine Involvement in the Regulation of Secretionin Coho Salmon,” Can. J. Zool. 64 (6), 1245–1248(1986).
48. C. Wabnitz, M. Taylor, E. Green, and T. Razak, FromOcean to Aquarium: The Global Trade in Marine Orna-mentals (UNEP World Conservation Monitoring Centre,Cambridge, UK, 2003).
Translated by T. B. Chicheva




















