
Bahram Gur's Spectacular Marksmanship and the Art of Illustration in Qajar Lithographed Books

lable at ScienceDirect
Quaternary International 252 (2012) 122e141
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
The scene of spectacular feasts: Animal remains from Pavlov I south-east,the Czech Republic
Piotr Wojtal a,*, Jaros1aw Wilczy�nski a, Zbigniew M. Boche�nski a, Jiri A. Svoboda b,c
a Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31-016 Kraków, PolandbDepartment of Anthropology, Faculty of Science, Masaryk University, Kotlá�rská 2, 602 00 Brno, Czech Republicc Institute of Archaeology at Brno, ASCR, Královopolská 147, 612 00 Brno, Czech Republic
a r t i c l e i n f o
Article history:Available online 29 June 2011
* Corresponding author.E-mail addresses: [email protected] (P. W
krakow.pl (J. Wilczy�nski), [email protected]@iabrno.cz (J.A. Svoboda).
1040-6182/$ e see front matter � 2011 Elsevier Ltd adoi:10.1016/j.quaint.2011.06.033
a b s t r a c t
Gravettian culture arose about 30 thousand years ago and expanded into nearly the whole of Europeduring the next millennia. Gravettian cultural levels are known from caves as well as open-air sites.Pavlov I belongs to the latter category, which is typical in Central and Eastern Europe. Excavations atthis site were conducted from 1952 to 1971. We were able to study materials collected during exca-vations at the south-east part of the site in the years 1952e56. They yielded enormous numbers ofarchaeological artefacts (e.g., stone and bone tools, personal ornaments, clay figurines) and more than47,000 animal bones. Remains of small (birds, hares and foxes) and medium sized animals (reindeer)dominate the bone assemblage. In contrast to other Gravettian sites, mammoth remains are not sonumerous at Pavlov I. A characteristic feature of the site is a very large number of carnivore remains,especially from wolves and foxes. All stages of processing (skinning, dismembering and filleting) areevidenced for different mammal taxa. For the Pavlovian people, herbivores (reindeer, woollymammoth, and horse), carnivores (fox, wolf, wolverine, bears, and cave lion), and small animals (haresand birds) were an important source of food as well as raw material for tool production and orna-ments. All skeletal parts of small, medium, and large animals were found, which indicates that hunterstransported whole bodies of killed animals to the site. More than 300 cut marks were found on theanimal bones. Other signs of human activity are not so numerous. Because the site was occupied forrelatively long periods of time, carnivores had limited access to the remains and only 23 bones havetooth marks.
� 2011 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
The Mid-Upper Palaeolithic, or later Interpleniglacial andearly Upper Pleniglacial (terminal Marine Isotope Stage 3 andearly Stage 2), was a period of global climatic instability (Guthrieand Van Kolfschoten, 2000). In the period between 30 and 20thousand years ago a considerable cultural unification took placein Europe, which resulted in the so called Gravettian tech-nocomplex (Roebroeks et al., 2000). It arose about 30 thousandyears BP and, by the next millennia, extended over various partsof Europe, from the Pyrenees to the Ural mountains (Soffer, 1985;Koz1owski, 1986; Svoboda et al., 1996; Villaverde, 1998). Inwestern Europe, Gravettian is mostly known from cave sites in
ojtal), [email protected] (Z.M. Boche�nski),
nd INQUA. All rights reserved.
France and Spain, whereas in Central and Eastern Europe largeopen-air sites prevail. The Gravettian technocomplex in CentralEurope includes among others the earlier Pavlovian stage (sitessuch a Pavlov I, Dolní Vĕstonice I, Krems Wachtberg), and thelater Willendorf-Kostienkian stage (sites such as Wilendorf II,Petrkovice, Milovice, Moravany, Kraków Spadzista Street)(Koz1owski, 1998; Fladerer, 2001; Wojtal and Sobczyk, 2005;Svoboda, 2007, 2008; Oliva, 2009). The most distinctivefeatures of the individual stages of Gravettian technocomplexcultures were the presence of characteristic stone tools (backedbladelets and shouldered points) and figurative art, includingzoomorphic and anthropomorphic objects. Gravettian sites,especially those in Central Europe, often yielded large mammothbone deposits. These accumulations are interpreted as remains ofspecialized mammoth hunting or mammoth scavenging, result-ing from bone gathering or secondary exploitation of naturaldeath sites (West, 2001; Svoboda et al., 2005; Wojtal andSobczyk, 2005; Brugère et al., 2009).

P. Wojtal et al. / Quaternary International 252 (2012) 122e141 123
The Danube valley and adjacent South Moravia are regarded asa Gravettian region of particular importance. The site of Wil-lendorf II, layer 5, provided the earliest Gravettian dates, reachingup to 30,000 BP, while south Moravia provided a complex systemof famous early Gravettian sites, dated to about 27e25,000 BP.These sites are all located at the foot of Pavlovské (Palava) Hillsand include such large and well known sites as Pavlov I, DolníVĕstonice I, and Dolní Vĕstonice II (Absolon and Klima, 1977;Svoboda, 1991, 1994a,b, 2005; Klima, 2005). Additional discov-eries confirm intensive human occupation at new sites such asPavlov VI or Milovice IV (Svoboda et al., 2009; Svoboda et al. inpreparation).
2. The site of Pavlov I
Pavlov I is one of several sites extending about 2.5 km along thenorthern foot of the Pavlovské Hills, from Dolní V�estonice II in thewest, through Dolní V�estonice I, Pavlov I and II to Pavlov VI in theeast (Fig. 1). It is an example of a large and long-term settlement,excavated by Bohuslav Klíma between 1952e1971, using
Fig. 1. Location of Pavlov I on the roots of Pavlovsk
methodological standards of that time (wet-sieving and recordingprovenience from squares). It is clear that the original modelordering the individual settlement units into a kind of a “village”needs to by tested by an internal analysis. In an ideal case,a settlement unit would be composed of a hearth, associatedstructures of pits and other features, and one or more clusters ofartifacts and faunal remains which may be interpreted as “huts”.However sites with such coarse-grained assemblages, wheremicrolayers and spatial units are not clearly separated, presenta long termmethodological problem.When interpreting these sitestoday, several site-formation factors are involved: the intensity ofoccupation, its repetition, variability of activities, and finally thepostdepositional processes. The analysis includes spatial relation-ships (or overlapping) among the individual settlement units,connections by refittings, 14C dating, and typological and othercomparative study of their content.
The Gravettian cultural layer complex of Pavlov I is anthropo-genic in origin. It is thick and sometimes structured into micro-layers, with soil components, charcoal, burnt bone fragments andvarious osteological materials, ochre and artifacts. In some areas
e Hills, and a plan of the trenches at the site.

Table 1NISP (Number of Identified Specimens), %NISP, MNI (Minimum Number of Indi-viduals) and %MNI of birds and mammals from Pavlov I south-east. Ia – domiantemammal taxa; IIb – complimentary mammal taxa; IIIc – incidental mammal taxa.
Taxon NISP %NISP MNI %MNI
Cygnus cygnus/Cygnus olor 2 0.004 1 0.19Cygnus columbianus 3 0.006 2 0.37Anas querquedula 1 0.002 1 0.19Asio flammeus 1 0.002 1 0.19Haliaetus albicilla 2 0.004 1 0.19Gyps fulvus 2 0.004 1 0.19Accipitridae indet. 1 0.002 e e
Falco cf. tinnunculus 1 0.002 1 0.19Falco cf. peregrinus 2 0.004 1 0.19Lagopus lagopus 269 0.57 27 5.03Lagopus muta 17 0.035 3 0.56Lagopus sp. 47 0.1 e e
Tetrao tetrix 52 0.11 5 0.93Tetraonidae indet (T. tetrix/
Lagopus sp.)52 0.11 e e
Perdix perdix 1 0.002 1 0.19Charadriiformes indet (size
Larus)1 0.002 1 0.19
Turdus sp. 1 0.002 1 0.19Passeriformes indet. (size
Emberiza)1 0.002 1 0.19
cf. Pica pica 1 0.002 e e
Pica/Garrulus 1 0.002 1 0.19Pyrrhocorax pyrrhocorax 4 0.008 1 0.19Pyrrhocorax/Corvus monedula 8 0.016 1 0.19Corvus monedula 11 0.023 2 0.37small Corvidae 2 0.004 e e
Corvus corax 352 0.7 12 2.23Aves indet. 22 0.046 e e
IIIc Castor fibre 9 0.018 2 0.37Rodentia indet. 45 0.095 e e
IIIc Ochotona pussilla 6 0.012 3 0.56IIIc Martes/Mustela 18 0.038 e e
Ia Lepus sp. 6773 14.31 192 35.82IIb Gulo gulo 781 1.65 10 1.86IIb Panthera spelaea 81 0.171 3 0.56Ia Alopex lagopus/
Vulpes vulpes5460 11.54 123 22.94
Ia Canis lupus 6190 13.1 57 10.63IIb Ursus arctos 6 0.012 1 0.19IIb Ursus spelaeus 3 0.006 1 0.19IIb Ursus sp. 41 0.086 2 0.37
Carnivora 165 0.35 e e
IIb Equus sp. 589 1.24 10 1.86IIb Mammuthus
primigenius2264 4.8 7 1.3
IIIc Colodontaantiquitatis
2 0.004 1 0.19
IIIc Cervus elaphus 4 0.008 1 0.19Ia Rangifer tarandus 4026 8.51 56 10.44
Cervidae 491 1.04 e e
IIIc Bison/Bos 3 0.006 1 0.19IIIc Capra ibex 5 0.01 1 0.19
Small mammal(fox-hare sized)
4632 9.8 e e
Medium mammal(reindeer-wolf sized)
1954 4.13 e e
Large mammal(horse-bear sized)
555 1.18 e e
Very large mammal(mammoth sized)
616 1.3 e e
Mammaliaindeterminate
11747 24.82 e e
TOTAL NISP/MNI 47323 536
P. Wojtal et al. / Quaternary International 252 (2012) 122e141124
where artifacts are concentrated, and especially in the depressionsand around hearths, two or more cultural layers are separated bya thin layer of loess. However these superpositions were not datedby 14C during the excavation. In terms of 14C chronology, the 11
dates available belong to a time-span of twomillenia 25e27,000 BP,or 27e29,000 cal BC, which corresponds to the Evolved Pavlovianstage. Given this time-span and intensity of occupation, we expectthat the formation of the whole complex was a relatively rapidprocess, and the typological analysis of artifacts from the individuallayers, wherever the material was separated during the excavation(as in 1953), confirms this expectation.
Excavations at Pavlov I provided many sometimes spectacularfinds. One male burial and numerous isolated human remainswere recovered (Trinkaus and Svoboda, 2006; Trinkaus et al.,2010). A wide range of stone tool classes were found andvariety of activities had been performed, such as processinganimal and plant food and the maintenance of hunting tools,indicating that Pavlov I may have been a long term base camp(Verpoorte, 2005). At the site were also discovered artefactsshowing extensive human actions: decorated bones, carvedsculptures, and antler and ivory tools. Remains of many hearthswere noted at Pavlov I. At the southeast part of the site, theremains of 56 hearths were found. But the most typicalphenomena of Pavlov I and II and Dolní V�estonice I and II arebaked clay figurines, which were probably used in rituals (Sofferand Vandiver, 1994; Svoboda, 1994a,b). At Pavlov I and DolníV�estonice I and II were found negative clay impressions of fibre-based constructions. They are the world’s oldest evidence for theuse of plant fibres in the manufacture of textiles, basketry, andnetting (Adovasio et al., 1996, 1997, 2005).
The picture derived from site-plans and microstratigraphiesmay be supplemented by analysing distribution patterns of thevarious find categories, artifacts and naturefacts. As shown byVerpoorte (2000) who analysed the lithic industry, the settled areahas the character of a “palimpsest”. Following Novák (2005) whostudied distribution patterns of a variety of artifacts (lithics andbone industries, decorative objects, ceramic fragments, ochre) theoriginal scatter patterns of the individual find categories weredisturbed during multiple repeated occupations. No clear distri-bution pattern is visible in central areas having dense accumula-tions of finds, nor is a separation of residential and activity zonespossible. More favourable areas for this type of analysis are siteperipheries, where the various distribution patterns are moreevident. Nevertheless, the site is not a simple mixture of autono-mous discrete units, but rather a representation of a site wheremultiple activities were performed in the same place during a longtime period. Finally, the locations already disturbed by repeatedhuman activities were slightly modified by geological post-depositional processes.
At other large sites such as Dolní V�estonice I and II or Milovice I,mammoth bone deposits were recovered. Such an assemblage wasnot recorded at Pavlov I and the effects of erosion by an active creekon the eastern boundary of the site may be a possible explanationfor this absence (Svoboda, 2005). Compared to Dolní V�estonice orMilovice, this absence has reduced mammoth representation in thetotal faunal spectrum of Pavlov I.
Previous analyses of the faunal remains have focused on thetaxonomic composition and metrical analyses (e.g. Musil, 1955,1959, 1994, 2005a). Our studies supplement and continue thepresentation of results of archaeological and zoological studiesfrom this area of the site (Svoboda, 1994a,b, 2005). The animalbones collected during field work from 1957 until 1971 from otherareas have been not studied yet.
Our studies were carried out on animal remains collected duringexcavations in the southeast part of Pavlov I, done from 1952 to1956 and covering 875 sq.m (25 � 35 m). This area was an inten-sively and repeatedly settled part of the site, providing the richestarchaeological and palaeontological finds. The excavations atPavlov I - Southeast yielded 47,323 remains of mammals, birds and

%NISP
2%
36%
27%
3%
6%
26%
BirdsSmall mammal (fox-hare sized)Medium mammal (reindeer-wolf sized)Large mammal (horse-bear sized)MammothIndeterminate
Fig. 2. %NISP (Number of Identified Specimens) of birds, small mammals (fox-hare sized), medium mammals (reindeer-wolf sized), large mammals (horse-bear sized) and verylarge mammals (mammoth sized).
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 125
a single fish. It is worth mentioning that isolated fossils probablycollected by the Gravettian people were also discovered, includinga Miocene shark tooth and shells of Tertiary molluscs (Hladilováand Mikulá�s, 2005; Hladilová, 2005).
3. Material and methods
The animal bones and teeth collected during all seasons ofexcavations at Pavlov I are stored in the osteological collections ofBudi�sov Castle of the Moravian Museum (Moravské zemskémuzeum). Human remains and stone and bone artefacts from the
%MNI
12%
61%
23%
3% 1%
Fig. 3. %MNI (Minimum Number of Individuals) of birds, small mammals (fox-hare sized),large mammals (mammoth sized).
site are stored in the Institute of Archaeology, Czech Academy ofSciences, in Dolní V�estonice.
Frequencies of skeletal elements and animal individuals fromPavlov I southeast were measured in terms of the Number ofIdentified Specimens (NISP) and Minimum Number of Individ-uals (MNI). NISP is defined and used as the number of identifiedspecimens in a collection, where identified means ascribed totaxon. MNI was estimated by sorting e but not matching e leftand right elements. There were also calculated MNE and MAUvalues. MNE (Minimum Number of skeletal Elements) is anestimation of the number of skeletal elements represented byspecimens in the assemblage, based on the most common
BirdsSmall mammal (fox-hare sized)Medium mammal (reindeer-wolf sized)Large mammal (horse-bear sized)Mammoth
medium mammals (reindeer-wolf sized), large mammals (horse-bear sized) and very
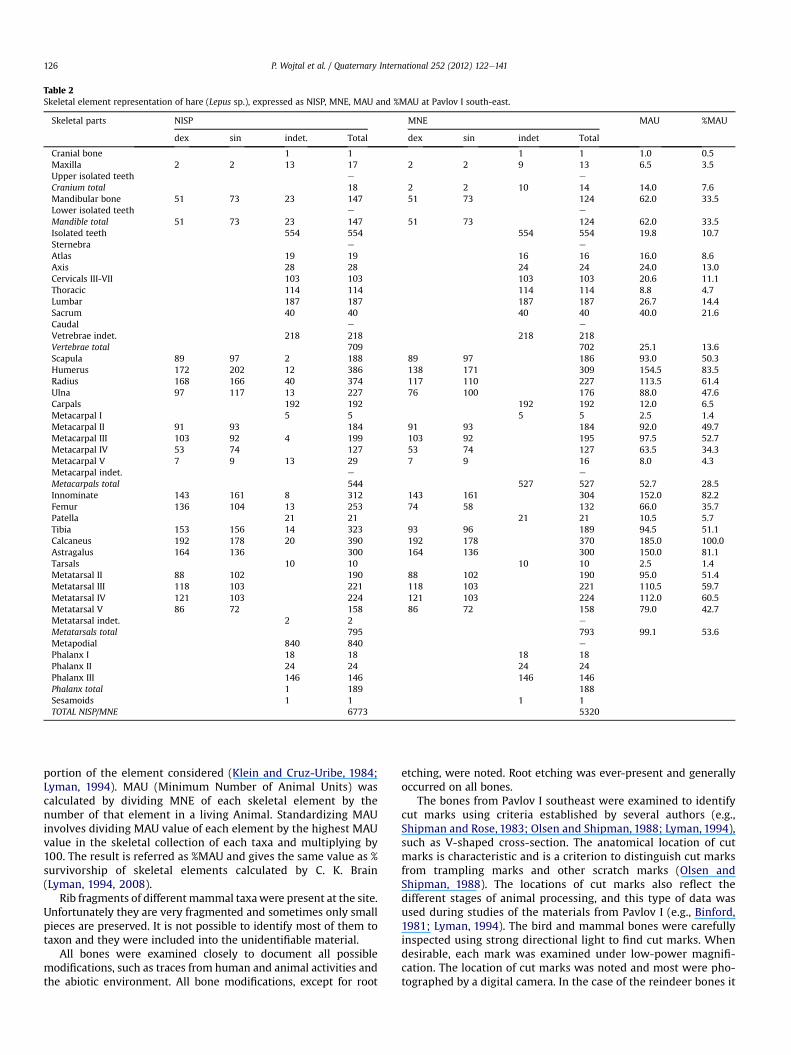
Table 2Skeletal element representation of hare (Lepus sp.), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone 1 1 1 1 1.0 0.5Maxilla 2 2 13 17 2 2 9 13 6.5 3.5Upper isolated teeth e e
Cranium total 18 2 2 10 14 14.0 7.6Mandibular bone 51 73 23 147 51 73 124 62.0 33.5Lower isolated teeth e e
Mandible total 51 73 23 147 51 73 124 62.0 33.5Isolated teeth 554 554 554 554 19.8 10.7Sternebra e e
Atlas 19 19 16 16 16.0 8.6Axis 28 28 24 24 24.0 13.0Cervicals III-VII 103 103 103 103 20.6 11.1Thoracic 114 114 114 114 8.8 4.7Lumbar 187 187 187 187 26.7 14.4Sacrum 40 40 40 40 40.0 21.6Caudal e e
Vetrebrae indet. 218 218 218 218Vertebrae total 709 702 25.1 13.6Scapula 89 97 2 188 89 97 186 93.0 50.3Humerus 172 202 12 386 138 171 309 154.5 83.5Radius 168 166 40 374 117 110 227 113.5 61.4Ulna 97 117 13 227 76 100 176 88.0 47.6Carpals 192 192 192 192 12.0 6.5Metacarpal I 5 5 5 5 2.5 1.4Metacarpal II 91 93 184 91 93 184 92.0 49.7Metacarpal III 103 92 4 199 103 92 195 97.5 52.7Metacarpal IV 53 74 127 53 74 127 63.5 34.3Metacarpal V 7 9 13 29 7 9 16 8.0 4.3Metacarpal indet. e e
Metacarpals total 544 527 527 52.7 28.5Innominate 143 161 8 312 143 161 304 152.0 82.2Femur 136 104 13 253 74 58 132 66.0 35.7Patella 21 21 21 21 10.5 5.7Tibia 153 156 14 323 93 96 189 94.5 51.1Calcaneus 192 178 20 390 192 178 370 185.0 100.0Astragalus 164 136 300 164 136 300 150.0 81.1Tarsals 10 10 10 10 2.5 1.4Metatarsal II 88 102 190 88 102 190 95.0 51.4Metatarsal III 118 103 221 118 103 221 110.5 59.7Metatarsal IV 121 103 224 121 103 224 112.0 60.5Metatarsal V 86 72 158 86 72 158 79.0 42.7Metatarsal indet. 2 2 e
Metatarsals total 795 793 99.1 53.6Metapodial 840 840 e
Phalanx I 18 18 18 18Phalanx II 24 24 24 24Phalanx III 146 146 146 146Phalanx total 1 189 188Sesamoids 1 1 1 1TOTAL NISP/MNE 6773 5320
P. Wojtal et al. / Quaternary International 252 (2012) 122e141126
portion of the element considered (Klein and Cruz-Uribe, 1984;Lyman, 1994). MAU (Minimum Number of Animal Units) wascalculated by dividing MNE of each skeletal element by thenumber of that element in a living Animal. Standardizing MAUinvolves dividing MAU value of each element by the highest MAUvalue in the skeletal collection of each taxa and multiplying by100. The result is referred as %MAU and gives the same value as %survivorship of skeletal elements calculated by C. K. Brain(Lyman, 1994, 2008).
Rib fragments of different mammal taxawere present at the site.Unfortunately they are very fragmented and sometimes only smallpieces are preserved. It is not possible to identify most of them totaxon and they were included into the unidentifiable material.
All bones were examined closely to document all possiblemodifications, such as traces from human and animal activities andthe abiotic environment. All bone modifications, except for root
etching, were noted. Root etching was ever-present and generallyoccurred on all bones.
The bones from Pavlov I southeast were examined to identifycut marks using criteria established by several authors (e.g.,Shipman and Rose, 1983; Olsen and Shipman, 1988; Lyman, 1994),such as V-shaped cross-section. The anatomical location of cutmarks is characteristic and is a criterion to distinguish cut marksfrom trampling marks and other scratch marks (Olsen andShipman, 1988). The locations of cut marks also reflect thedifferent stages of animal processing, and this type of data wasused during studies of the materials from Pavlov I (e.g., Binford,1981; Lyman, 1994). The bird and mammal bones were carefullyinspected using strong directional light to find cut marks. Whendesirable, each mark was examined under low-power magnifi-cation. The location of cut marks was noted and most were pho-tographed by a digital camera. In the case of the reindeer bones it

0
10
20
30
40
50
60
70
80
90
100
llukS
elbidnaM
saltA
sixA
IIV-III slacivreC
cicarohT
rabmuL
murcaS
alupacS
suremu
H
suidaR
anlU
lapracateM
etanimonnI
rumeF
aibiT
alubiF
muenaclaC
sulagartsA
lasratateM
I xnalahP
II xnalahP
III xnalahP
UAM
%
Lepus sp. Alopex/Vulpes
Fig. 4. Relative skeletal element abundance of hare and foxes, expressed as standardized minimal animal units (%MAU) at Pavlov I south-east.
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 127
was possible to distinguish marks made during skinning,dismembering and filleting using Binford’s (1981) criteria. Someof the above-mentioned criteria were also used for the interpre-tation of cutmarks on the bones of other taxa, namely carnivoresand lagomorphs.
Another modification made by humans which was studied onthe bone material from Pavlov I are traces of fire. Humans canburn bones in intentional actions such as using bones as a fuel,cooking and roasting, refuse removal or through unintentionalexposure (Stiner et al., 1995; Bennet, 1999; Théry-Parisot, 2002;Villa et al., 2002; Schiegl et al., 2003). A colour-based scheme(ranging from natural hue to black to white) is the mostcommonly used method for the identification of burned bone (e.g.,Buikstra and Swegle, 1989; Lyman, 1994; Shahack-Gross et al.,1997). Other criteria have been applied to confirm burning ofremains (Shipman et al., 1984; Stiner et al., 1995), but the colour ofthe bones was the only feature we used to distinguish burnedbones at Pavlov I south-east.
Bones from Pavlov I were examined to find carnivore modifi-cations. Carnivores are important taphonomic agents affectingbone preservation, potentially influencing bone survival andrepresentation in the assemblage. Characteristic damages causedby carnivores have been described by Binford (1981), Haynes (1980,1983), Lyman (1994), and Sutclife (1970), among others, who havenoted different types of modification (punctures, pits, scores andfurrows) caused by carnivore teeth. The dimension and shape of themarks sometimes allow identification of the taxon responsible forgnawing (Haynes, 1983).
The bones fromPavlov I were also examined to find rodent gnawmarks, which are closely-spaced and flat bottomed parallel grooves(Lyman, 1994). At the site only one fragment of Cervidae antler hadsigns of rodent gnawing.
4. Results
4.1. Animal taxa and skeletal representation
The mammals are represented by 46,466 bones and teeth.Species and element identification was possible for 26,962
specimens belonging to 16 taxa; the remaining bone and teethfragments (w42% of all remains) are not identifiable (Table 1). Thefollowing size groups of mammals can be distinguished: small(hare-fox sized), medium (reindeer-wolf sized), large (horse-bearsized), and very large (mammoth) (Figs. 2 and 3).
The most numerous remains at Pavlov I southeast are those ofthe hare (Lepus sp.), consisting of nearly 6800 bones belonging toa minimum of 192 individuals (Tables 1 and 2). Noteworthy isthe extreme fragmentation of the hare remains. Most boneswere broken and only some parts of them have been preserved,which made species identification impossible. Appendicularelements are clearly more abundant than axial elements(Table 2, Fig. 4), possibly a result either of greater fragility of theskull and vertebra, or the easier identification of leg bones. Sevenbones of the steppe pika (Ochotona pusilla) were also found.
Surprisingly, carnivore bones and teeth make up more than47% of all identified mammallian remains. The most numerousamong them are foxes (red and polar fox) and wolves. Foxes arerepresented by more than 5400 bones and teeth from a minimumof 123 individuals (Tables 1 and 3). Wolves (Canis lupus) weremore numerous in terms of the number of specimens(NISP ¼ 6190) but they represent only 57 individuals (Tables 1 and4). During the excavations 781 remains were found froma minimum of 10 wolverine (Gulo gulo) individuals (Tables 1 and5). All mentioned carnivores are represented by all skeletalelements. Relative skeletal element abundances for them noted as%MAU show that both axial elements (skull and mandibles) andappendicular elements are well represented. Only vertebrae werefound in smaller numbers. However, both atlas and axis, which areeasier to identify on the basis of fragments, are also well repre-sented (Figs. 4 and 5). Also found were a few dozen bones andteeth of large carnivores e specifically cave lion (Panthera spelaea)and bears (Ursus arctos, Ursus spelaeus, U. sp.), which belong toonly one or two individuals. The axial skeleton is representedmainly by teeth and mandibular fragments, and foot bones(carpals and tarsals, metapodials and phalanges) are very wellpreserved in the material (Tables 6 and 7).
Remains of three taxa of larger herbivores, reindeer (Rangifertarandus), woolly mammoth (Mammuthus primigenius) and horse
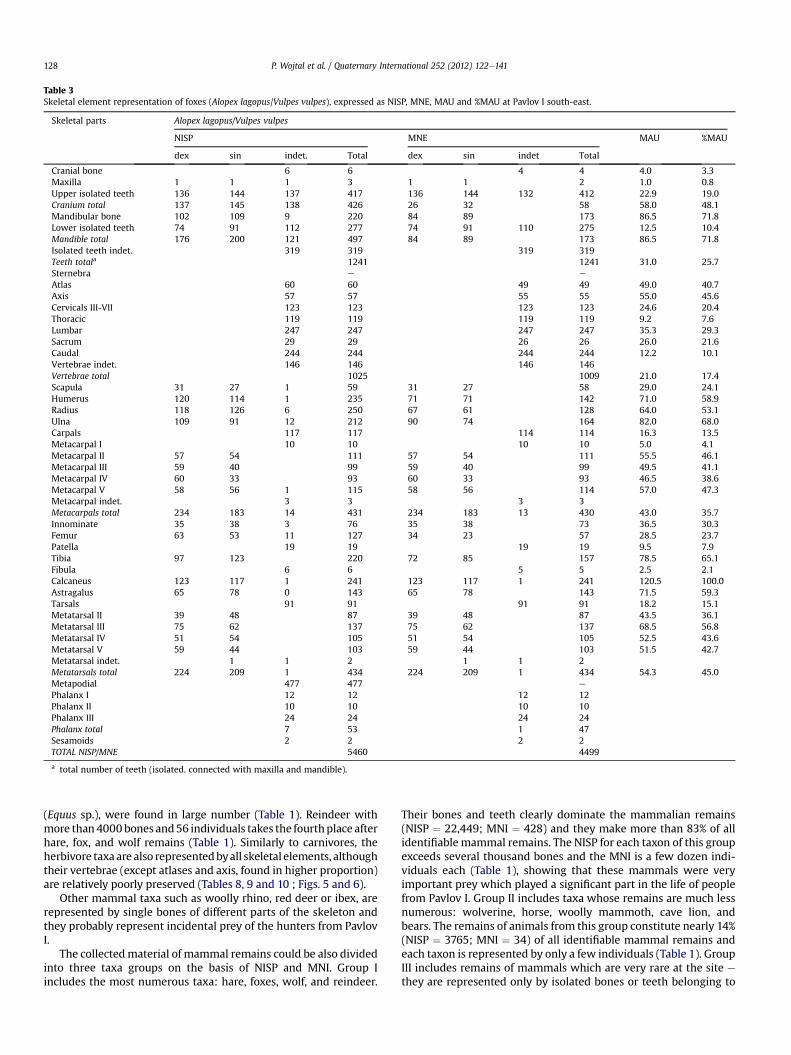
Table 3Skeletal element representation of foxes (Alopex lagopus/Vulpes vulpes), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts Alopex lagopus/Vulpes vulpes
NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone 6 6 4 4 4.0 3.3Maxilla 1 1 1 3 1 1 2 1.0 0.8Upper isolated teeth 136 144 137 417 136 144 132 412 22.9 19.0Cranium total 137 145 138 426 26 32 58 58.0 48.1Mandibular bone 102 109 9 220 84 89 173 86.5 71.8Lower isolated teeth 74 91 112 277 74 91 110 275 12.5 10.4Mandible total 176 200 121 497 84 89 173 86.5 71.8Isolated teeth indet. 319 319 319 319Teeth totala 1241 1241 31.0 25.7Sternebra e e
Atlas 60 60 49 49 49.0 40.7Axis 57 57 55 55 55.0 45.6Cervicals III-VII 123 123 123 123 24.6 20.4Thoracic 119 119 119 119 9.2 7.6Lumbar 247 247 247 247 35.3 29.3Sacrum 29 29 26 26 26.0 21.6Caudal 244 244 244 244 12.2 10.1Vertebrae indet. 146 146 146 146Vertebrae total 1025 1009 21.0 17.4Scapula 31 27 1 59 31 27 58 29.0 24.1Humerus 120 114 1 235 71 71 142 71.0 58.9Radius 118 126 6 250 67 61 128 64.0 53.1Ulna 109 91 12 212 90 74 164 82.0 68.0Carpals 117 117 114 114 16.3 13.5Metacarpal I 10 10 10 10 5.0 4.1Metacarpal II 57 54 111 57 54 111 55.5 46.1Metacarpal III 59 40 99 59 40 99 49.5 41.1Metacarpal IV 60 33 93 60 33 93 46.5 38.6Metacarpal V 58 56 1 115 58 56 114 57.0 47.3Metacarpal indet. 3 3 3 3Metacarpals total 234 183 14 431 234 183 13 430 43.0 35.7Innominate 35 38 3 76 35 38 73 36.5 30.3Femur 63 53 11 127 34 23 57 28.5 23.7Patella 19 19 19 19 9.5 7.9Tibia 97 123 220 72 85 157 78.5 65.1Fibula 6 6 5 5 2.5 2.1Calcaneus 123 117 1 241 123 117 1 241 120.5 100.0Astragalus 65 78 0 143 65 78 143 71.5 59.3Tarsals 91 91 91 91 18.2 15.1Metatarsal II 39 48 87 39 48 87 43.5 36.1Metatarsal III 75 62 137 75 62 137 68.5 56.8Metatarsal IV 51 54 105 51 54 105 52.5 43.6Metatarsal V 59 44 103 59 44 103 51.5 42.7Metatarsal indet. 1 1 2 1 1 2Metatarsals total 224 209 1 434 224 209 1 434 54.3 45.0Metapodial 477 477 e
Phalanx I 12 12 12 12Phalanx II 10 10 10 10Phalanx III 24 24 24 24Phalanx total 7 53 1 47Sesamoids 2 2 2 2TOTAL NISP/MNE 5460 4499
a total number of teeth (isolated. connected with maxilla and mandible).
P. Wojtal et al. / Quaternary International 252 (2012) 122e141128
(Equus sp.), were found in large number (Table 1). Reindeer withmore than 4000 bones and56 individuals takes the fourthplace afterhare, fox, and wolf remains (Table 1). Similarly to carnivores, theherbivore taxa are also representedbyall skeletal elements, althoughtheir vertebrae (except atlases and axis, found in higher proportion)are relatively poorly preserved (Tables 8, 9 and 10 ; Figs. 5 and 6).
Other mammal taxa such as woolly rhino, red deer or ibex, arerepresented by single bones of different parts of the skeleton andthey probably represent incidental prey of the hunters from PavlovI.
The collectedmaterial of mammal remains could be also dividedinto three taxa groups on the basis of NISP and MNI. Group Iincludes the most numerous taxa: hare, foxes, wolf, and reindeer.
Their bones and teeth clearly dominate the mammalian remains(NISP ¼ 22,449; MNI ¼ 428) and they make more than 83% of allidentifiable mammal remains. The NISP for each taxon of this groupexceeds several thousand bones and the MNI is a few dozen indi-viduals each (Table 1), showing that these mammals were veryimportant prey which played a significant part in the life of peoplefrom Pavlov I. Group II includes taxa whose remains are much lessnumerous: wolverine, horse, woolly mammoth, cave lion, andbears. The remains of animals from this group constitute nearly 14%(NISP ¼ 3765; MNI ¼ 34) of all identifiable mammal remains andeach taxon is represented by only a few individuals (Table 1). GroupIII includes remains of mammals which are very rare at the site e
they are represented only by isolated bones or teeth belonging to
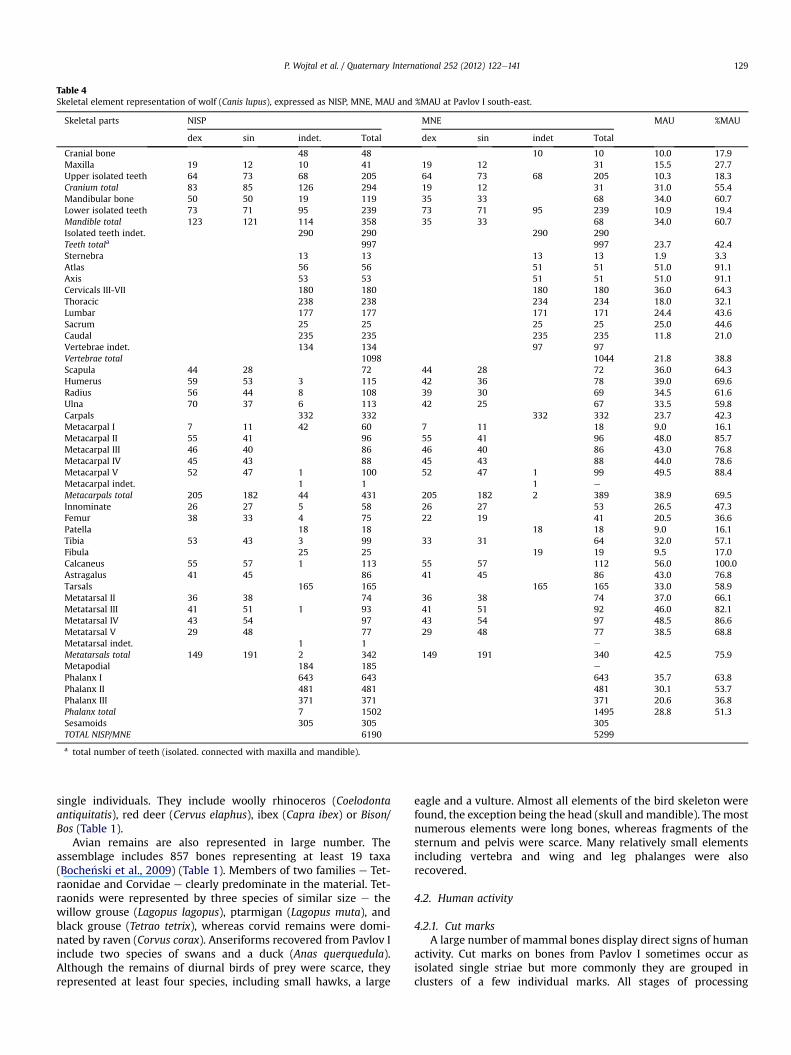
Table 4Skeletal element representation of wolf (Canis lupus), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone 48 48 10 10 10.0 17.9Maxilla 19 12 10 41 19 12 31 15.5 27.7Upper isolated teeth 64 73 68 205 64 73 68 205 10.3 18.3Cranium total 83 85 126 294 19 12 31 31.0 55.4Mandibular bone 50 50 19 119 35 33 68 34.0 60.7Lower isolated teeth 73 71 95 239 73 71 95 239 10.9 19.4Mandible total 123 121 114 358 35 33 68 34.0 60.7Isolated teeth indet. 290 290 290 290Teeth totala 997 997 23.7 42.4Sternebra 13 13 13 13 1.9 3.3Atlas 56 56 51 51 51.0 91.1Axis 53 53 51 51 51.0 91.1Cervicals III-VII 180 180 180 180 36.0 64.3Thoracic 238 238 234 234 18.0 32.1Lumbar 177 177 171 171 24.4 43.6Sacrum 25 25 25 25 25.0 44.6Caudal 235 235 235 235 11.8 21.0Vertebrae indet. 134 134 97 97Vertebrae total 1098 1044 21.8 38.8Scapula 44 28 72 44 28 72 36.0 64.3Humerus 59 53 3 115 42 36 78 39.0 69.6Radius 56 44 8 108 39 30 69 34.5 61.6Ulna 70 37 6 113 42 25 67 33.5 59.8Carpals 332 332 332 332 23.7 42.3Metacarpal I 7 11 42 60 7 11 18 9.0 16.1Metacarpal II 55 41 96 55 41 96 48.0 85.7Metacarpal III 46 40 86 46 40 86 43.0 76.8Metacarpal IV 45 43 88 45 43 88 44.0 78.6Metacarpal V 52 47 1 100 52 47 1 99 49.5 88.4Metacarpal indet. 1 1 1 e
Metacarpals total 205 182 44 431 205 182 2 389 38.9 69.5Innominate 26 27 5 58 26 27 53 26.5 47.3Femur 38 33 4 75 22 19 41 20.5 36.6Patella 18 18 18 18 9.0 16.1Tibia 53 43 3 99 33 31 64 32.0 57.1Fibula 25 25 19 19 9.5 17.0Calcaneus 55 57 1 113 55 57 112 56.0 100.0Astragalus 41 45 86 41 45 86 43.0 76.8Tarsals 165 165 165 165 33.0 58.9Metatarsal II 36 38 74 36 38 74 37.0 66.1Metatarsal III 41 51 1 93 41 51 92 46.0 82.1Metatarsal IV 43 54 97 43 54 97 48.5 86.6Metatarsal V 29 48 77 29 48 77 38.5 68.8Metatarsal indet. 1 1 e
Metatarsals total 149 191 2 342 149 191 340 42.5 75.9Metapodial 184 185 e
Phalanx I 643 643 643 35.7 63.8Phalanx II 481 481 481 30.1 53.7Phalanx III 371 371 371 20.6 36.8Phalanx total 7 1502 1495 28.8 51.3Sesamoids 305 305 305TOTAL NISP/MNE 6190 5299
a total number of teeth (isolated. connected with maxilla and mandible).
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 129
single individuals. They include woolly rhinoceros (Coelodontaantiquitatis), red deer (Cervus elaphus), ibex (Capra ibex) or Bison/Bos (Table 1).
Avian remains are also represented in large number. Theassemblage includes 857 bones representing at least 19 taxa(Boche�nski et al., 2009) (Table 1). Members of two families e Tet-raonidae and Corvidae e clearly predominate in the material. Tet-raonids were represented by three species of similar size e thewillow grouse (Lagopus lagopus), ptarmigan (Lagopus muta), andblack grouse (Tetrao tetrix), whereas corvid remains were domi-nated by raven (Corvus corax). Anseriforms recovered from Pavlov Iinclude two species of swans and a duck (Anas querquedula).Although the remains of diurnal birds of prey were scarce, theyrepresented at least four species, including small hawks, a large
eagle and a vulture. Almost all elements of the bird skeleton werefound, the exception being the head (skull andmandible). Themostnumerous elements were long bones, whereas fragments of thesternum and pelvis were scarce. Many relatively small elementsincluding vertebra and wing and leg phalanges were alsorecovered.
4.2. Human activity
4.2.1. Cut marksA large number of mammal bones display direct signs of human
activity. Cut marks on bones from Pavlov I sometimes occur asisolated single striae but more commonly they are grouped inclusters of a few individual marks. All stages of processing
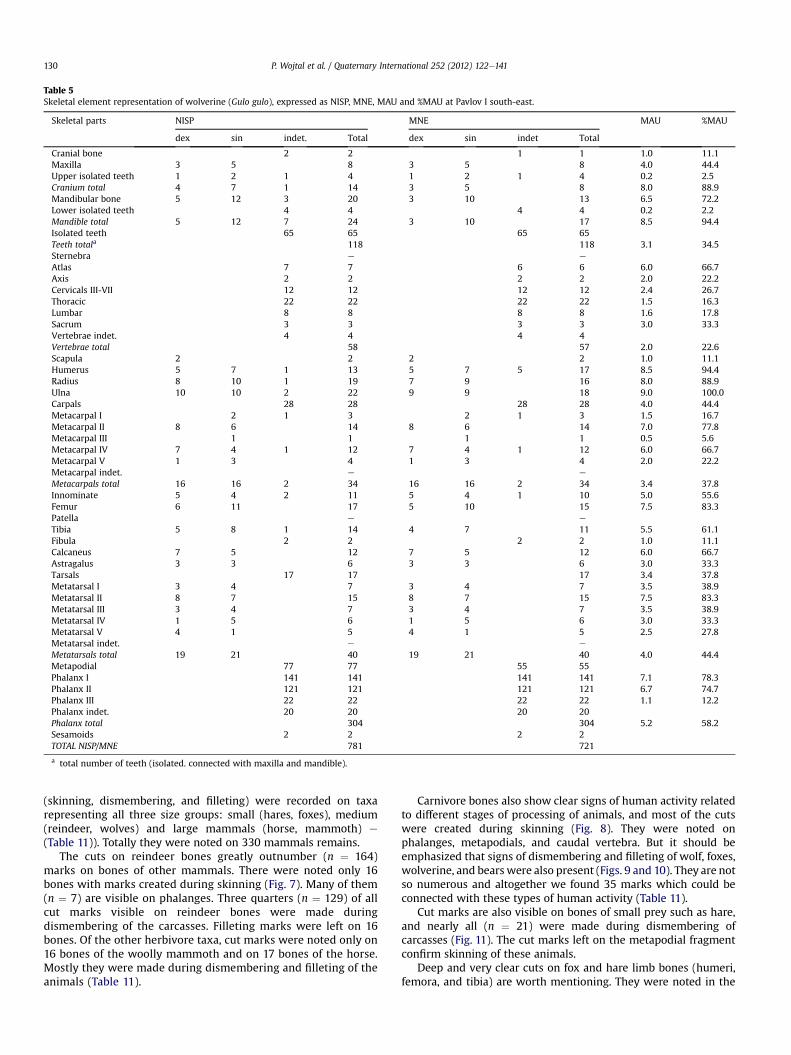
Table 5Skeletal element representation of wolverine (Gulo gulo), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone 2 2 1 1 1.0 11.1Maxilla 3 5 8 3 5 8 4.0 44.4Upper isolated teeth 1 2 1 4 1 2 1 4 0.2 2.5Cranium total 4 7 1 14 3 5 8 8.0 88.9Mandibular bone 5 12 3 20 3 10 13 6.5 72.2Lower isolated teeth 4 4 4 4 0.2 2.2Mandible total 5 12 7 24 3 10 17 8.5 94.4Isolated teeth 65 65 65 65Teeth totala 118 118 3.1 34.5Sternebra e e
Atlas 7 7 6 6 6.0 66.7Axis 2 2 2 2 2.0 22.2Cervicals III-VII 12 12 12 12 2.4 26.7Thoracic 22 22 22 22 1.5 16.3Lumbar 8 8 8 8 1.6 17.8Sacrum 3 3 3 3 3.0 33.3Vertebrae indet. 4 4 4 4Vertebrae total 58 57 2.0 22.6Scapula 2 2 2 2 1.0 11.1Humerus 5 7 1 13 5 7 5 17 8.5 94.4Radius 8 10 1 19 7 9 16 8.0 88.9Ulna 10 10 2 22 9 9 18 9.0 100.0Carpals 28 28 28 28 4.0 44.4Metacarpal I 2 1 3 2 1 3 1.5 16.7Metacarpal II 8 6 14 8 6 14 7.0 77.8Metacarpal III 1 1 1 1 0.5 5.6Metacarpal IV 7 4 1 12 7 4 1 12 6.0 66.7Metacarpal V 1 3 4 1 3 4 2.0 22.2Metacarpal indet. e e
Metacarpals total 16 16 2 34 16 16 2 34 3.4 37.8Innominate 5 4 2 11 5 4 1 10 5.0 55.6Femur 6 11 17 5 10 15 7.5 83.3Patella e e
Tibia 5 8 1 14 4 7 11 5.5 61.1Fibula 2 2 2 2 1.0 11.1Calcaneus 7 5 12 7 5 12 6.0 66.7Astragalus 3 3 6 3 3 6 3.0 33.3Tarsals 17 17 17 3.4 37.8Metatarsal I 3 4 7 3 4 7 3.5 38.9Metatarsal II 8 7 15 8 7 15 7.5 83.3Metatarsal III 3 4 7 3 4 7 3.5 38.9Metatarsal IV 1 5 6 1 5 6 3.0 33.3Metatarsal V 4 1 5 4 1 5 2.5 27.8Metatarsal indet. e e
Metatarsals total 19 21 40 19 21 40 4.0 44.4Metapodial 77 77 55 55Phalanx I 141 141 141 141 7.1 78.3Phalanx II 121 121 121 121 6.7 74.7Phalanx III 22 22 22 22 1.1 12.2Phalanx indet. 20 20 20 20Phalanx total 304 304 5.2 58.2Sesamoids 2 2 2 2TOTAL NISP/MNE 781 721
a total number of teeth (isolated. connected with maxilla and mandible).
P. Wojtal et al. / Quaternary International 252 (2012) 122e141130
(skinning, dismembering, and filleting) were recorded on taxarepresenting all three size groups: small (hares, foxes), medium(reindeer, wolves) and large mammals (horse, mammoth) e
(Table 11)). Totally they were noted on 330 mammals remains.The cuts on reindeer bones greatly outnumber (n ¼ 164)
marks on bones of other mammals. There were noted only 16bones with marks created during skinning (Fig. 7). Many of them(n ¼ 7) are visible on phalanges. Three quarters (n ¼ 129) of allcut marks visible on reindeer bones were made duringdismembering of the carcasses. Filleting marks were left on 16bones. Of the other herbivore taxa, cut marks were noted only on16 bones of the woolly mammoth and on 17 bones of the horse.Mostly they were made during dismembering and filleting of theanimals (Table 11).
Carnivore bones also show clear signs of human activity relatedto different stages of processing of animals, and most of the cutswere created during skinning (Fig. 8). They were noted onphalanges, metapodials, and caudal vertebra. But it should beemphasized that signs of dismembering and filleting of wolf, foxes,wolverine, and bears were also present (Figs. 9 and 10). They are notso numerous and altogether we found 35 marks which could beconnected with these types of human activity (Table 11).
Cut marks are also visible on bones of small prey such as hare,and nearly all (n ¼ 21) were made during dismembering ofcarcasses (Fig. 11). The cut marks left on the metapodial fragmentconfirm skinning of these animals.
Deep and very clear cuts on fox and hare limb bones (humeri,femora, and tibia) are worth mentioning. They were noted in the

Table 6Skeletal element representation of cave lion (Panthera spelaea), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet Total dex sin indet Total
Cranial bone e e
Maxilla e e
Upper isolated teeth 1 1 1 1 0.1 3.1Cranium total 1 1 1 1.0 50.0Mandibular bone 1 1 1 3 1 1 2 1.0 50.0Lower isolated teeth 1 3 4 1 2 3 0.2 10.7Mandible total 7 1 1 2 1.0 50.0Sternebra e e
Atlas 1 1 1 1 1.0 50.0Axis 1 1 1 1 1.0 50.0Cervicals III-VII 1 1 1 1 0.2 10.0Thoracic e e
Lumbar e
Sacrum e e
Caudal 1 1 1 1Vertebrae indet. e e
Vertebrae total 4 3 0.1 5.4Scapula 1 1 1 1 0.5 25.0Humerus 1 1 1 1 0.5 25.0Radius e e
Ulna 1 1 1 1 0.5 25.0Carpals 7 7 7 7 0.5 25.0Metacarpal I 1 1 1 1 0.5 25.0Metacarpal II 1 1 1 1 0.5 25.0Metacarpal III 1 1 2 1 1 2 1.0 50.0Metacarpal IV 1 1 2 1 1 2 1.0 50.0Metacarpal V 1 1 2 1 1 2 1.0 50.0Metacarpal indet. e e
Metacarpals total 8 8 0.8 40.0Innominate e e
Femur e e
Patella e e
Tibia 1 1 1 1 0.5 25.0Fibula 1 1 1 1 0.5 25.0Calcaneus 1 1 2 1 1 2 1.0 50.0Astragalus 2 2 4 2 2 4 2.0 100.0Tarsals 1 1 1 1 0.2 10.0Metatarsal I 1 1 1 1 0.5 25.0Metatarsal II 1 1 1 1 0.5 25.0Metatarsal III 1 1 2 1 1 2 1.0 50.0Metatarsal IV e e
Metatarsal V 3 3 3 3Metatarsal indet. e e
Metatarsals total 7 7 0.7 35.0Metapodial 3 3 3Phalanx I 10 10 10 0.5 25.0Phalanx II 18 18 18 1.1 56.3Phalanx III 4 4 4 0.2 10.0Phalanx total 32 32 0.6 28.6Sesamoids e e
TOTAL NISP/MNE 81 79
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 131
middle of nine bone shafts, and they certainly could not have beenrelated to the processing (skinning, dismembering or filleting) ofthese animals (Fig. 12). However, the aim of these actions is notclear.
Careful examination of bird bones collected from Pavlov I south-east yielded only two bones with cut marks made with stone toolse a distal humerus of a black grouse (T. tetrix) and a proximalhumerus of a whooper swan (Cygnus cygnus) with cut marks on theanterior side of its proximal end. The position of the marks suggeststhat they were made during dismembering of the carcass(Boche�nski et al., 2009).
4.2.2. Burning damageIn the studied material only 382 mammalian remains with signs
of fire activity were found (Table 11). Of them 108 specimens wereidentified to particular taxon, whereas other fragments (n ¼ 274)
were not identifiable, being usually small pieces a few centimetresin length. Among avian remains, only one partly burned distalcarpometacarpus of Lagopus sp. was found (Boche�nski et al., 2009).
4.2.3. Percussion marksOther direct signs of human activity such as percussion marks
are rare and visible only on 41 bones of different mammal taxa.Most of the bones which have the marks are not identifiable.Despite the scarcity of percussion marks it should be noted that allreindeer long limb bones were broken. Long bones of other taxa(hare, wolverine, wolf, foxes,) were not so fragmented and somewere intact.
Despite a large number of broken bones, only isolated boneflakes produced during breaking or hitting bones were discovered(n ¼ 15). We can not exclude the possibility that smaller andunidentifiable bone fragments were not collected during

Table 7Skeletal element representation of bears (Ursus arctos, Ursus spealeus, Ursus sp.), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet. Total
Cranial bone e e
Maxilla e e
Upper isolated teeth 2 4 4 10 2 4 4 10 0.6 55.6Cranium total 10 1 1 1 1.0 100.0Mandibular bone e
Lower isolated teeth 1 1 2 1 1 2 0.1 11.1Mandible total 1 1 2 1 1 2 1.0 100.0Isolated teeth 5 5 4 4 0.1 11.1Sternebra e e
Atlas e e
Axis e e
Cervicals III-VII e e
Thoracic 1 1 1 0.1 7.7Lumbar e e
Sacrum e e
Vertebrae indet. e e
Total vertebrae 1 1 0.04 3.6Scapula e e
Humerus e e
Radius e e
Ulna 1 1 1 1 0.5 50.0Carpals e e
Metacarpal I e e
Metacarpal II 1 1 1 1 0.5 50.0Metacarpal III 1 1 2 1 1 2 1.0 100.0Metacarpal IV e e
Metacarpal V 1 1 1 0.5 50.0Metacarpal indet. e e
Total metacarpals 4 4 0.4 40.0Innominate e e
Femur e e
Patella e e
Tibia 1 1 1 1 0.5 50.0Fibula 1 1 1 1 0.5 50.0Calcaneus e e
Astragalus 1 1 1 1 0.5 50.0Tarsals 2 2 2 0.2 20.0Metatarsal II 1 1 1 1 0.5 50.0Metatarsal III e e
Metatarsal IV e e
Metatarsal V 1 1 2 1 1 2 1.0 100.0Metatarsal indet. e e
Total metatarsals 3 3 0.4 37.5Metapodial 4 4 2 e
Phalanx I 5 5 5 5 0.3 25.0Phalanx II 7 7 7 7 0.4 43.8Phalanx III 2 2 2 2 0.1 10.0Phalanx 1 15 1 15 0.3 26.8Sesamoids e e
TOTAL NISP/MNE 50 46
P. Wojtal et al. / Quaternary International 252 (2012) 122e141132
excavations in the 1950’s. It should be mentioned that tusk flakesare represented in the assemblage (n¼ 27) and they probably comefrom broken tusks which were prepared for tool production.
4.2.4. Bone tools and ornamentsPavlov I south-east yielded 858 art objects and tools made from
mammal teeth, bones, antlers, and tusks. Large number of artefacts(n ¼ 197) were made from mammoth bones and tusks. Some toolswere made from mammoth bones which had been heavily modi-fied, such as shovel- (spoon) like tools (Fig. 13). However, the mostnumerous are the tools in the form of points or spatulas made fromtusk. Most are small fragments, several centimetres in length. Verynumerous in this material are objects, or more often semi-finishedproducts made from reindeer antlers. These are mostly smallfragments of antlers bearing traces of notching, cutting and chop-ping. These traces are the result of split antlers in the production of
“hoes” or other tools. Reindeer bones were rarely used for themanufacture of tools. In the studied material there are only a fewperforators and awls (Fig. 14). These type of tools were also madefrom horse reduced metapodia, and these bones were the onlyremains of this taxon used in tool production.
Pendants were made mainly from carnivore teeth. Gravettianhunters used mainly teeth of foxes (n ¼ 284) and wolves (n ¼ 65),mostly third upper incisors and canines. This type of personal orna-ment was made by gently drilling a hole in the tooth root, or bypercussion. Only two pendants were made from reindeer teeth(incisors). Thebonesof carnivores, especially longbones fromwolfandfoxes, were used for themanufacture of tools. Humanmodification ofthe two ulnae of large birds e raven and Bewick’s swan e are notconnected with processing but may be considered objects of art(Boche�nski et al., 2009). One of the bones was cut and polished frombothendsand theotheroneshowsan interestingpatternof cutmarks.

0
10
20
30
40
50
60
70
80
90
100
llukS
elbidnaM
saltA
sixA
IIV-IIIslacivre
C
cicarohT
rabmuL
murcaS
alupacS
suremu
H
suidaR
anlU
lapracateM
etanimonnI
rumeF
aibiT
alubiF
muenaclaC
sulagartsA
lasratateM
IxnalahP
IIxnalahP
IIIxnalahP
UAM
%
Canis lupus Rangifer tarandus Gulo gulo
Fig. 5. Relative skeletal element abundance of wolf, reindeer, wolverine, expressed as standardized minimal animal units (%MAU) at Pavlov I south-east.
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 133
4.3. Carnivore damage
Only 23 bones with gnawing marks from large carnivores werefound. Most of the gnawed bones belong to woolly mammoth(n ¼ 8) and reindeer (n ¼ 5). Dimensions and shapes of the marksindicate that the marks were probably made by wolves. Only onmammoth bones was noted more intensive gnawing by carnivores.Single bones of the hare, wolf, and fox were also gnawed, and hadpuncture marks.
5. Discussion
The archaeological content, thickness, and multilayer characterof the cultural complex suggest that large sites such as Pavlov I are,in fact, accumulations or palimpsests of smaller ones. The small siteof Pavlov VI has recently provided a good case of such a smaller (anelemental unit) site (Svoboda et al., 2009; Wojtal, NývltováFi�sáková, Wilczy�nski in preparation). However the question to askis whether this is all they are. Pavlov I provides evidence of a longerstay in terms of months and years, and the most importantevidence of specific activities such as symbolism and rituals. Itseems that these locations are not just archaeological summaries ofthe individual episodes, but that there was a pattern of humanaggregation over longer time-spans and centralized activities (bothutilitarian and symbolic).
Without trying to detail the individual stages (or seasons) ofoccupation, we do think it is possible to use the large amount ofthe zooarchaeological and anthropological facts to reconstructvarious aspects of the life of the Gravettian hunters. We were ableto study more than 47,000 animal remains from the south-easternpart of the site. Important roles were played in the life of Pavlovhunters’ not only by hare, fox, wolf, and reindeer, but also bywoolly mammoth. The taxa representation, number of remains,and individuals of different animals found at Pavlov I show thatthe diet of its inhabitants was diversified, with the presence oflarger herbivores e reindeer, woolly mammoths, and horses.Smaller animals (hares, foxes, birds) were also significant for the
inhabitants of the site and they were an important diet compo-nent (Figs. 2 and 3). At other Gravettian sites of Central Europethat are younger than Pavlov I (Milovice I, Kraków Spadzista, orMoravany Lopata), animal taxa representation is not so diversi-fied, with clear domination by woolly mammoth and reindeer(West, 1996; Lipecki and Wojtal, 1998; Wojtal and Sobczyk, 2005;Brugère et al., 2009).
A very characteristic feature of Pavlov I is a very large number ofcarnivore remains, especially wolf and foxes. They certainly had animportantmeaning in the lives of people from this site. It is possiblethat the hides were the main reason for hunting carnivores. Otherbody parts of carnivores were used in the production of tools (awlsmade from bones) and personal ornaments (pendants made fromwolf and fox teeth). The importance of carnivores for people of theGravettian culture is also shown by findings from other sites. Thebones and teeth of carnivores (wolf, foxes, and wolverine), some-times in larger proportion, were found at open-air sites of Moravia,Austria, and Poland. These sites include Krems-Wachtberg (Austria)(Fladerer, 2001), Dolní V�estonice I and II (West, 2001; Musil, 2003;Wojtal and Wilczy�nski, in preparation), Jaro�sov-Podvr�st’a (CzechRepublic) (Musil, 2005b), and Kraków Spadzista Street (E) and (F)(West, 1996; Wilczy�nski and Wojtal in preparation). Carnivoreteeth were used as pendants not only at Pavlov I but also at otherGravettian sites (Valde-Nowak, 2003).
Bones of single individuals of bears and cave lions were found inthe above-mentioned Moravian sites and also at other open-airsites, where the paleontological material was not as rich. Thesites include Milovice I, Dolní Ve�stonice II, Krems-Wachtberg(Austria) (Fladerer, 2001; Fladerer and Salcher, 2004; Brugèreet al., 2009; Wojtal and Wilczy�nski in preparation), showing thatduring the Gravettian hunting large and dangerous carnivores suchas cave lions and cave bears was not accidental but intentional. Thehunting of large carnivores, specifically bears (cave bears or brownbears) or cave lions (P. spelaea), is also documented not only atanother Gravettian site Hohle Fels cave in Germany, but also atolder sites (Auguste, 1992; Tillet and Binford, 2002; Münzel andConard, 2004; Wojtal, 2007; Blasco et al., 2010).

Table 8Skeletal element representation of reindeer (Rangifer tarandus), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone 18 18 8 8 8.0 14.4Maxilla e e
Antler 6 13 152 171 6 13 19Upper isolated teeth 135 132 22 289 135 132 22 289 24.1 43.4Cranium total 478 21 24 45 45.0 81.1Mandibular bone 21 13 12 46 15 8 23 11.5 20.7Lower isolated teeth 196 206 88 490 196 206 88 490 27.2 49.0Mandible total 217 219 100 536 24 39 63 31.5 56.8Isolated teeth 78 78 78 78Teeth totala 893 893 24.8 44.7Sternabra 3 3 3 3 0.4 0.8Atlas 16 16 14 14 14.0 25.2Axis 27 27 21 21 21.0 37.8Cervicals III-VII 53 53 53 53 10.6 19.1Thoracic 76 76 76 76 5.8 10.5Lumbar 39 39 39 39 6.5 11.7Sacrum 9 9 9 9 9.0 16.2Vertebrae indet. 40 40 35 35Vertebrae total 260 247 9.1 16.5Scapula 16 30 1 47 16 30 46 23.0 41.4Humerus 44 29 2 75 39 24 63 31.5 56.8Radius 97 68 14 179 51 35 86 43.0 77.5Ulna 38 46 9 93 36 42 78 39.0 70.3Carpals 333 333 333 27.8 50.0Metacarpal 19 27 99 145 19 27 46 23.0 41.4Metacarpal reduced 45 45 40 40 10.0 18.0Metacarpal indet. e e
Metacarpals total 19 27 144 190 19 27 40 86 14.3 25.8Innominate 6 6 4 16 6 6 12 6.0 10.8Femur 14 18 10 42 7 10 17 8.5 15.3Patella 12 19 3 34 12 19 31 15.5 27.9Tibia 71 63 6 140 56 55 111 55.5 100.0Os malleolare 23 23 46 23 23 46 23.0 41.4Calcaneus 44 37 7 88 44 37 81 40.5 73.0Astragalus 54 51 2 107 54 51 105 52.5 94.6Tarsals 125 125 125 31.3 56.3Metatarsal 20 24 87 131 20 24 44 22.0 39.6Metatarsal indet. e e
Total metatarsals 20 24 87 131 20 24 44 22.0 39.6Metapodial 101 101 e
Phalanx I 298 298 246 246 15.4 27.7Phalanx II 186 186 172 172 10.8 19.4Phalanx III 109 109 109 109 6.8 12.3Phalanx indet. 3 3 3 3Phalanx total 596 527 11.0 19.8Reduced phalanx I 63 63 61 61 3.8 6.9Reduced phalanx II 66 66 63 63 3.9 7.1Reduced phalanx III 53 53 53 53 3.3 6.0Sesamoids 149 149 149 149TOTAL NISP/MNE 4206 3255
a total number of teeth (isolated. connected with maxilla and mandible).
P. Wojtal et al. / Quaternary International 252 (2012) 122e141134
The Pavlov I site can be described as a long term base camp andwe can not exclude that in some periods the site was year-roundinhabited. The large proportion of fur-bearing mammals at PavlovI support winter occupation of the site. A year-round occupationwas confirmed at Dolní Ve�stonice II, another Pavlovian site locateda few hundred metres away (Nývltová Fi�sáková, 2007).
We have no direct evidence of Pavlovian people’s huntingmethods and we can only make assumption about how animalswere killed. The finds from this site provide a clue as to how theswift, fast-moving small game animals, such as hares or ravens,were hunted. Knotted nets made from plant fibres, whose oldestevidence in the world was found at Pavlov I, may have been usedas efficient entrapment devices for catching small mammals andbirds (Adovasio et al., 1997, 2005; Soffer and Adovasio, 2004).Fowling methods known from ethnographic and zooarchaeo-logical studies include nets, snares, sling-stones, bows, spears,
and others, depending on local customs (Lupo and Schmitt, 2002;Serjeantson, 2009; Pangau-Adam and Noske, 2010; Purnama andIndrawan, 2010). Larger mammals (reindeer, horse, wolf) couldbe killed using spears or the bow and arrow, a longer distanceweapon. The presence of large number of raven remains andtheir wing bones suggest that feathers were used in arrow orspear fletching (Boche�nski et al., 2009). Brühl (2005) suggeststhat also shovel- (spoon) like tools (Fig. 13) could be used asa sabre-like slashing weapon during the hunting of smallmammals.
Pavlov I is not a killing site of woolly mammoths. However, thepresence of their bones and teeth show that people transported tothe site at least some parts of their carcasses. The site is also nota killing site of other animals. It was occupied by people for a longertime than a kill or processing camp would have been, and it is veryhard to imagine that a place where people more or less

Table 9Skeletal element representation of woolly mammoth (Mammuthus primigenius), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone 152 152 7 7 7.0 100.0Maxilla e e
Milk tusk 3 3 3 1.5 21.4Tusk fragments 458 458 e
Upper isolated cheek teeth 10 9 4 23 10 9 4 23Hyoid bone 7 7 7 7 3.5 50.0Cranium total 484 7 7 7.0 100.0Mandibular bone 9 9 1 1 1.0 14.3Lower isolated cheek teeth 3 4 4 11 3 4 4 11Mandible total 3 4 4 20 2 3 5 5.0 71.4Teeth unidentifiable 10 10 10 10Teeth fragments (lamellae) 260 260 e
Teeth total 13 13 18 44 13 13 18 44 5.5 78.6Sternebra 2 2 2 2 0.3 4.1Atlas 9 9 4 4 4.0 57.1Axis 3 3 3 3 3.0 42.9Cervicals II-VII 17 e 14 14 2.8 40.0Thoracic 15 e 12 12 0.6 9.0Lumbar 4 4 4 4 1.0 14.3Sacrum 2 2 2 2 2.0 28.6Caudal 46 46 41 41 2.0 27.9Vertebrae indet. 42 42 23 23Vertebrae total 106 103 2.0 28.3Ribs 688 688 44 44 1.2 16.5Scapula 2 5 7 2 2 1.0 14.3Humerus 5 5 10 2 4 6 3.0 42.9Radius 7 1 14 22 6 1 8 15 7.5 107.1Ulna 2 5 8 15 2 5 3 10 5.0 71.4Carpals 44 e 40 40 3.3 47.6Metacarpal I 1 1 1 1 0.5 7.1Metacarpal II 1 1 2 1 1 2 1.0 14.3Metacarpal III e e
Metacarpal IV e e
Metacarpal V 2 1 3 2 1 3 1.5 21.4Metacarpal indet. 3 3 2 2Metacarpals Total 9 9 9 0.9 12.9Innominate 1 2 14 17 1 2 5 8 4.0 57.1Femur 1 1 14 16 1 1 4 6 3.0 42.9Patella 5 5 5 5 2.5 35.7Tibia 3 7 6 16 3 6 3 12 6.0 85.7Fibula 3 4 18 25 2 2 5 9 4.5 64.3Calcaneus 4 2 6 4 2 6 3.0 42.9Astragalus 1 1 4 6 1 1 4 6 3.0 42.9Tarsals 29 29 29 29 2.9 41.4Metatarsal I 1 1 2 1 1 2 1.0 14.3Metatarsal II 3 4 1 8 3 4 1 8 4.0 57.1Metatarsal III 4 1 1 6 4 1 1 6 3.0 42.9Metatarsal IV 1 2 3 1 2 3 1.5 21.4Metatarsal V 1 1 1 1 0.5 7.1Metatarsal indet. 3 3 3 3Metatarsals total 21 18 1.8 25.7Metapodial indet. 60 60 38 38Metapodial total 13 9 64 86 13 9 42 64 3.2 45.7Phalanx I 19 19 19 19 1.0 13.6Phalanx II 30 30 30 30 1.9 26.8Phalanx III 16 16 16 16 0.8 11.4Phalanx indet 56 56 29 29Phalanx total 121 94 1.7 24.0Sesamoids 69 69 69 69TOTAL NISP/MNE 2402 600
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 135
permanently lived was also a killing site for so many animals ofdifferent groups, including mammoths, reindeer, wolves, and foxes.
The large amount of different signs of human activity allows usto reconstruct some behaviours of the inhabitants of Pavlov I. Thesmall number of cutmarks created during skinning practices visibleon reindeer bones, in comparison with large number of marksmade during dismembering and filleting, shows that the hides ofreindeers were not so important as raw material. On the otherhand, cut marks on reindeer phalanges suggest that the main
purpose of obtaining reindeer hides was to make socks and shoes(Binford, 1981). The large number of carnivore remains andnumerous signs of skinning show that they provided hides forclothing preparation. Skinning cut marks were found on 22 carni-vore bones, mostly metapodials, phalanges, and caudal vertebra.They show that hunters tried to get hides of the animals as large aspossible. We have to point out the presence of the cut marks madeduring dismembering and filleting of wolves, foxes, wolverines, andbears, indicating that carnivores were also a source of food for the

Table 10Skeletal element representation of horse (Equus sp.), expressed as NISP, MNE, MAU and %MAU at Pavlov I south-east.
Skeletal parts NISP MNE MAU %MAU
dex sin indet. Total dex sin indet Total
Cranial bone e e
Maxilla 1 1 1 1 0.5 5.3Upper isolated teeth 51 51 51 51 2.6 26.8Cranium total 52 52 4 4 4.0 42.1Mandibular bone 4 2 14 20 4 2 3 9 4.5 47.4Lower isolated teeth 69 69 69 69 3.5 36.3Mandible total 4 2 83 89 4 2 6 6.0 63.2Isolated teeth 64 64 64 64Teeth total 184 184 4.6 48.4Sternebra e e e
Atlas 3 3 3 3 3.0 31.6Axis 1 1 1 1 1.0 10.5Cervicals II-VII 5 5 5 5 1.0 10.5Thoracic 7 7 6 6 0.3 3.5Lumbar 3 3 3 3 0.5 5.3Sacrum e e e
Vertebrae indet. 2 2 1 1Vertebrae total 21 19 0.6 6.3Scapula 4 5 1 10 4 5 9 4.5 47.4Humerus 7 6 2 15 6 5 11 5.5 57.9Radius 6 12 9 27 4 6 10 5.0 52.6Ulna 3 9 2 14 3 6 9 4.5 47.4Carpals 62 62 62 62 8.9 93.2Metacarpal 2 1 3 2 1 3 1.5 15.8Innominate 1 1 2 1 1 2 1.0 10.5Femur 4 7 29 40 2 5 7 14 7.0 73.7Patella 10 7 3 20 10 7 2 19 9.5 100.0Tibia 6 9 13 28 6 8 14 7.0 73.7Calcaneus 1 1 2 1 1 2 1.0 10.5Astragalus 2 2 4 2 2 4 2.0 21.1Tarsals 26 26 26 26 5.2 54.7Metatarsal 2 2 2 1.0 10.5Metapodial 23 23 13 13Splint bone(mc II and IV; mt II and IV)
31 31 31 31 3.9 40.8
Phalanx I 14 14 14 14 1.2 12.3Phalanx II 12 12 12 12 1.0 10.5Phalanx III 6 6 6 6 0.5 5.3Phalanx total 32 32 0.9 9.4Sesamoids 22 22 22 22TOTAL NISP/MNE 730 476
Table 11Summary of burned bones and cut marks related to skinning, dismembering and filleting by taxon.
Taxon Cut marks total Skinning Dismembering Filleting Notidentifiable process Bone/teeth tools and ornaments Burned bones
Cygnus cygnus 1 1Cygnus columbianus 1Lagopus sp. 1Tetrao tetrix 1 1Corvus corax 1Lepus sp. 24 1 21 2 25Gulo gulo 10 3 5 1 1 1 11Panthera spelaea 2 1 1 3Alopex/Vulpes 18 2 5 4 7 293 17Canis lupus 36 16 16 3 1 74 21Ursus sp 1 1 3Carnivora 2Equus sp. 17 2 8 5 2 4 4Mammuthus primigenius 16 1 4 5 6 197 7Rangifer tarandus 164 16 129 16 3 71 20Cervidae 1 1 77 1Medium mammal (reindeer-wolf sized) 11 11Large mammal (horse-bear sized) 10 10Mammalia indeterminate 20 20 133 274TOTAL 332 42 192 34 64 860 383
P. Wojtal et al. / Quaternary International 252 (2012) 122e141136
0
10
20
30
40
50
60
70
80
90
100
llukS
elbidnaM
saltA
sixA
IIV-IIIslacivre
C
cicarohT
rabmuL
murcaS
alupacS
suremu
H
suidaR
anlU
lapracateM
etanimonnI
rumeF
aibiT
alubiF
muenaclaC
sulagartsA
lasratateM
IxnalahP
IIxnalahP
IIIxnalahP
UAM
%
Mammuthus primigenius Equus sp.
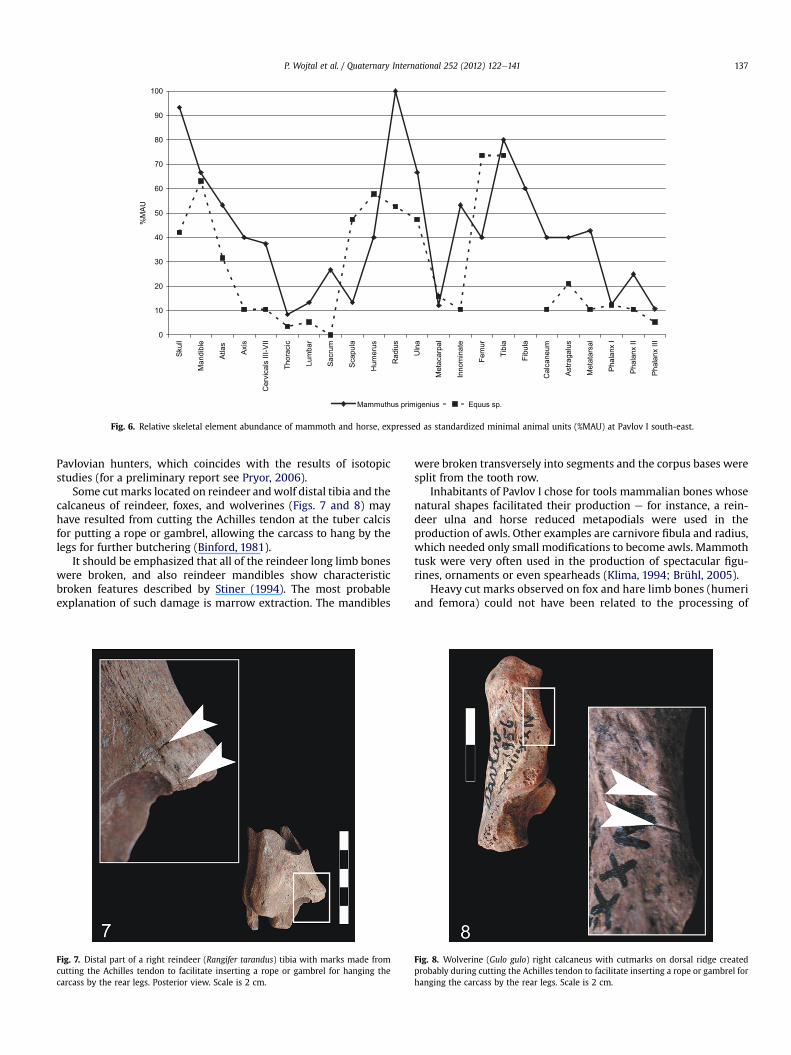
Fig. 6. Relative skeletal element abundance of mammoth and horse, expressed as standardized minimal animal units (%MAU) at Pavlov I south-east.
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 137
Pavlovian hunters, which coincides with the results of isotopicstudies (for a preliminary report see Pryor, 2006).
Some cut marks located on reindeer andwolf distal tibia and thecalcaneus of reindeer, foxes, and wolverines (Figs. 7 and 8) mayhave resulted from cutting the Achilles tendon at the tuber calcisfor putting a rope or gambrel, allowing the carcass to hang by thelegs for further butchering (Binford, 1981).
It should be emphasized that all of the reindeer long limb boneswere broken, and also reindeer mandibles show characteristicbroken features described by Stiner (1994). The most probableexplanation of such damage is marrow extraction. The mandibles
Fig. 7. Distal part of a right reindeer (Rangifer tarandus) tibia with marks made fromcutting the Achilles tendon to facilitate inserting a rope or gambrel for hanging thecarcass by the rear legs. Posterior view. Scale is 2 cm.
were broken transversely into segments and the corpus bases weresplit from the tooth row.
Inhabitants of Pavlov I chose for tools mammalian bones whosenatural shapes facilitated their production e for instance, a rein-deer ulna and horse reduced metapodials were used in theproduction of awls. Other examples are carnivore fibula and radius,which needed only small modifications to become awls. Mammothtusk were very often used in the production of spectacular figu-rines, ornaments or even spearheads (Klima, 1994; Brühl, 2005).
Heavy cut marks observed on fox and hare limb bones (humeriand femora) could not have been related to the processing of
Fig. 8. Wolverine (Gulo gulo) right calcaneus with cutmarks on dorsal ridge createdprobably during cutting the Achilles tendon to facilitate inserting a rope or gambrel forhanging the carcass by the rear legs. Scale is 2 cm.
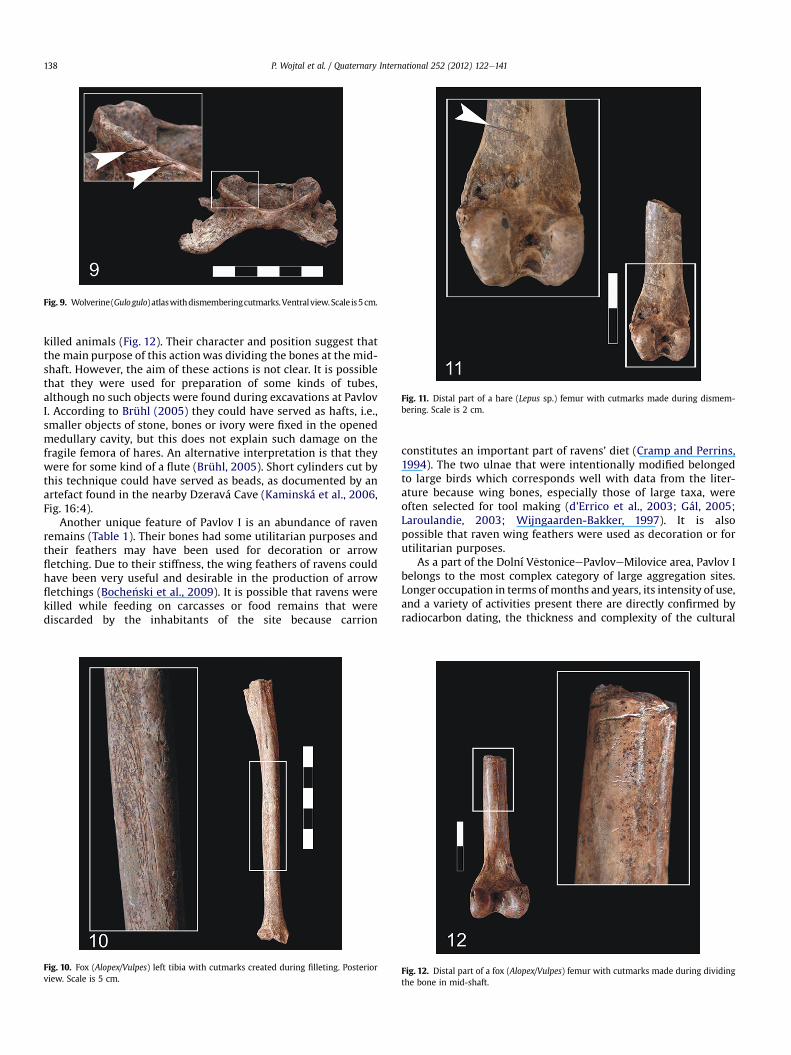
Fig. 9. Wolverine(Gulogulo) atlaswithdismemberingcutmarks.Ventralview.Scale is5cm.
Fig. 11. Distal part of a hare (Lepus sp.) femur with cutmarks made during dismem-bering. Scale is 2 cm.
P. Wojtal et al. / Quaternary International 252 (2012) 122e141138
killed animals (Fig. 12). Their character and position suggest thatthe main purpose of this actionwas dividing the bones at the mid-shaft. However, the aim of these actions is not clear. It is possiblethat they were used for preparation of some kinds of tubes,although no such objects were found during excavations at PavlovI. According to Brühl (2005) they could have served as hafts, i.e.,smaller objects of stone, bones or ivory were fixed in the openedmedullary cavity, but this does not explain such damage on thefragile femora of hares. An alternative interpretation is that theywere for some kind of a flute (Brühl, 2005). Short cylinders cut bythis technique could have served as beads, as documented by anartefact found in the nearby Dzeravá Cave (Kaminská et al., 2006,Fig. 16:4).
Another unique feature of Pavlov I is an abundance of ravenremains (Table 1). Their bones had some utilitarian purposes andtheir feathers may have been used for decoration or arrowfletching. Due to their stiffness, the wing feathers of ravens couldhave been very useful and desirable in the production of arrowfletchings (Boche�nski et al., 2009). It is possible that ravens werekilled while feeding on carcasses or food remains that werediscarded by the inhabitants of the site because carrion
Fig. 10. Fox (Alopex/Vulpes) left tibia with cutmarks created during filleting. Posteriorview. Scale is 5 cm.
constitutes an important part of ravens’ diet (Cramp and Perrins,1994). The two ulnae that were intentionally modified belongedto large birds which corresponds well with data from the liter-ature because wing bones, especially those of large taxa, wereoften selected for tool making (d’Errico et al., 2003; Gál, 2005;Laroulandie, 2003; Wijngaarden-Bakker, 1997). It is alsopossible that raven wing feathers were used as decoration or forutilitarian purposes.
As a part of the Dolní V�estoniceePavloveMilovice area, Pavlov Ibelongs to the most complex category of large aggregation sites.Longer occupation in terms ofmonths and years, its intensity of use,and a variety of activities present there are directly confirmed byradiocarbon dating, the thickness and complexity of the cultural
Fig. 12. Distal part of a fox (Alopex/Vulpes) femur with cutmarks made during dividingthe bone in mid-shaft.
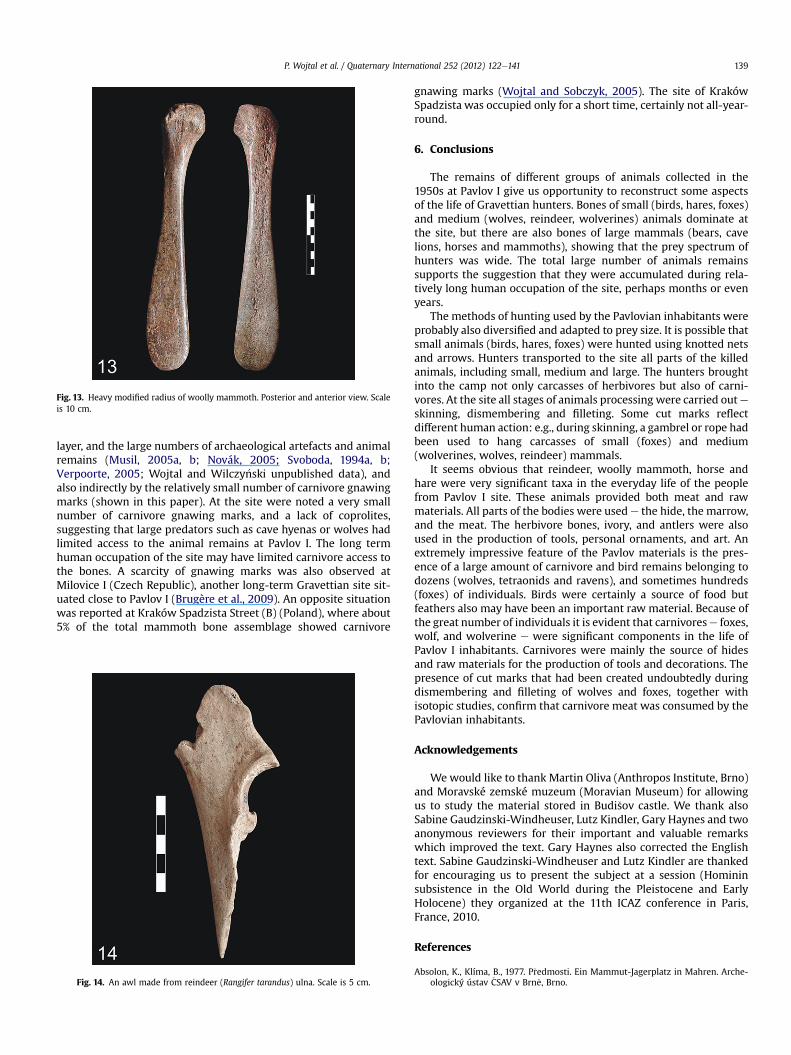
Fig. 13. Heavy modified radius of woolly mammoth. Posterior and anterior view. Scaleis 10 cm.
P. Wojtal et al. / Quaternary International 252 (2012) 122e141 139
layer, and the large numbers of archaeological artefacts and animalremains (Musil, 2005a, b; Novák, 2005; Svoboda, 1994a, b;Verpoorte, 2005; Wojtal and Wilczy�nski unpublished data), andalso indirectly by the relatively small number of carnivore gnawingmarks (shown in this paper). At the site were noted a very smallnumber of carnivore gnawing marks, and a lack of coprolites,suggesting that large predators such as cave hyenas or wolves hadlimited access to the animal remains at Pavlov I. The long termhuman occupation of the site may have limited carnivore access tothe bones. A scarcity of gnawing marks was also observed atMilovice I (Czech Republic), another long-term Gravettian site sit-uated close to Pavlov I (Brugère et al., 2009). An opposite situationwas reported at Kraków Spadzista Street (B) (Poland), where about5% of the total mammoth bone assemblage showed carnivore
Fig. 14. An awl made from reindeer (Rangifer tarandus) ulna. Scale is 5 cm.
gnawing marks (Wojtal and Sobczyk, 2005). The site of KrakówSpadzista was occupied only for a short time, certainly not all-year-round.
6. Conclusions
The remains of different groups of animals collected in the1950s at Pavlov I give us opportunity to reconstruct some aspectsof the life of Gravettian hunters. Bones of small (birds, hares, foxes)and medium (wolves, reindeer, wolverines) animals dominate atthe site, but there are also bones of large mammals (bears, cavelions, horses and mammoths), showing that the prey spectrum ofhunters was wide. The total large number of animals remainssupports the suggestion that they were accumulated during rela-tively long human occupation of the site, perhaps months or evenyears.
The methods of hunting used by the Pavlovian inhabitants wereprobably also diversified and adapted to prey size. It is possible thatsmall animals (birds, hares, foxes) were hunted using knotted netsand arrows. Hunters transported to the site all parts of the killedanimals, including small, medium and large. The hunters broughtinto the camp not only carcasses of herbivores but also of carni-vores. At the site all stages of animals processing were carried outeskinning, dismembering and filleting. Some cut marks reflectdifferent human action: e.g., during skinning, a gambrel or rope hadbeen used to hang carcasses of small (foxes) and medium(wolverines, wolves, reindeer) mammals.
It seems obvious that reindeer, woolly mammoth, horse andhare were very significant taxa in the everyday life of the peoplefrom Pavlov I site. These animals provided both meat and rawmaterials. All parts of the bodies were used e the hide, the marrow,and the meat. The herbivore bones, ivory, and antlers were alsoused in the production of tools, personal ornaments, and art. Anextremely impressive feature of the Pavlov materials is the pres-ence of a large amount of carnivore and bird remains belonging todozens (wolves, tetraonids and ravens), and sometimes hundreds(foxes) of individuals. Birds were certainly a source of food butfeathers also may have been an important raw material. Because ofthe great number of individuals it is evident that carnivorese foxes,wolf, and wolverine e were significant components in the life ofPavlov I inhabitants. Carnivores were mainly the source of hidesand raw materials for the production of tools and decorations. Thepresence of cut marks that had been created undoubtedly duringdismembering and filleting of wolves and foxes, together withisotopic studies, confirm that carnivore meat was consumed by thePavlovian inhabitants.
Acknowledgements
Wewould like to thank Martin Oliva (Anthropos Institute, Brno)and Moravské zemské muzeum (Moravian Museum) for allowingus to study the material stored in Budi�sov castle. We thank alsoSabine Gaudzinski-Windheuser, Lutz Kindler, Gary Haynes and twoanonymous reviewers for their important and valuable remarkswhich improved the text. Gary Haynes also corrected the Englishtext. Sabine Gaudzinski-Windheuser and Lutz Kindler are thankedfor encouraging us to present the subject at a session (Homininsubsistence in the Old World during the Pleistocene and EarlyHolocene) they organized at the 11th ICAZ conference in Paris,France, 2010.
References
Absolon, K., Klíma, B., 1977. P�redmosti. Ein Mammut-Jagerplatz in Mahren. Arche-ologický ústav �CSAV v Brn�e, Brno.

P. Wojtal et al. / Quaternary International 252 (2012) 122e141140
Adovasio, J.M., Hyland, D.C., Soffer, O., 1997. Textiles and cordage: a preliminaryassessment. In: Svoboda, J. (Ed.), Pavlov I Northwest: The Upper PaleolithicBurial and Its Settlement Context, Dolní V�estonice Studies, vol. 4,pp. 403e424.
Adovasio, J., Soffer, O., Klima, B., 1996. Upper Palaeolithic fibre technology: inter-laced woven finds from Pavlov I, Czech Republic, c. 26 000 years ago. Antiquity70, 526e534.
Adovasio, J.M., Soffer, O., Hyland, D.C., 2005. Textiles and cordage. In: Svoboda, J.(Ed.), Pavlov I southeast. A Window into Gravettian Lifestyles, Dolní V�estoniceStudies, vol. 14, pp. 432e443.
Auguste, P., 1992. Étude Archéozoologique des grands mammifères du site Plèis-tocene Moyen de Biache-Saint-Vaast (Pas-de-Calais, France): apports Bio-stratigraphiques et Palethnographiques. L’Anthropologie 96, 49e70.
Bennet, J.L., 1999. Thermal alternation of buried bone. Journal of ArchaeologicalScience 26, 1e8.
Binford, L.R., 1981. Bones: Ancient Men and Modern Myths. Academic Press,New York.
Blasco, R., Rosell, J., Arsuaga, J.L., Bermúdez de Castro, J.M., Carbonell, E., 2010. Thehunted hunter: the capture of a lion (Panthera leo fossilis) at the Gran Dolinasite, Sierra de Atapuerca, Spain. Journal of Archaeological Science 37,2051e2060.
Boche�nski, Z.M., Tomek, T., Wilczy�nski, J., Svoboda, J., Wertz, K., Wojtal, P., 2009.Fowling during the gravettian: the avifauna of Pavlov I, the Czech Republic.Journal of Archaeological Science 36, 2655e2665.
Brühl, E., 2005. The bone, antler and ivory tools. Dolní V�estonice Studies. In:Svoboda, J. (Ed.), Pavlov I Southeast. A Window into Gravettian Lifestyles, vol.14, Brno, pp. 252e293.
Brugère, A., Fontana, L., Oliva, M., 2009. Mammoth procurement and exploitation atMilovice (Czech Republic) New data for the Moravian gravettian. In: Fontana, L.,Chauvière, F.-X., Bridault, A. (Eds.), In Search of Total Animal Exploitation. CaseStudies from the Upper Palaeolithic and Mesolithic. Proceedings of the XVthUISPP Congress, Session C61, vol. 42, Lisbon, 4e9 September 2006. BAR Inter-national Series 2040, pp. 45e69.
Buikstra, J.E., Swegle, M., 1989. Bone modification due to burning: experimentalevidence. In: Bonnichsen, R., Sorg, M.H. (Eds.), Bone Modification. Center for theStudy of the First Americans. University of Maine, Orono, pp. 247e258.
Cramp, S., Perrins, C.M., 1994. The Birds of the Western Palearctic. vol. VIII. OxfordUniversity Press, Oxford, New York.
d’Errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M., Tillier, A.-M., Soressi, M.,Bresson, F., Maureille, B., Nowell, A., Lakarra, J., Backwell, L., Julien, M., 2003.Archaeological evidence for the emergence of language, symbolism, andmusic e an alternative multidisciplinary perspective. Journal of World Prehis-tory 17 (1), 1e70.
Fladerer, F.A., 2001. Die Faunareste vom jungpaläolithischen Lagerplatz Krems-Wachtberg, Ausgrabung 1930. Jagdwild und Tierkörpernutzung an der Donauvor 27.000 Jahren. Mittelungen der Prähistorischen Kommision. Österreichi-sche Akademie der Wissenschaften. Philosophish-historiche Klasse, vol. 39,Wien.
Fladerer, F.A., Salcher, T., 2004. Faunal remains from the Krems-Hundssteig/Wachtberg gravettian site complex e a difference in research techniques and/or site function?. The Dolní V�estonice Studies In: Svoboda, J.A., Sedlá�cková, L.(Eds.), The Gravettian Along the Danube, vol. 11, Brno, pp. 100e115.
Gál, E., 2005. New data to the bird bone artefacts from Hungary and Romania. In:Luik, H., Choyke, A.M., Batey, C.E., L}ougas, L. (Eds.), From Hooves to Horns, fromMollusc to Mammoth. Manufacture and Use of Bone Artefacts from PrehistoricTimes to the Present. Proceedings of the 4th Meeting of the ICAZ Worked BoneResearch Group at Tallinn, 26the31st of August 2003. Muinasaja teadus, vol. 15,Tallinn, pp. 325e338.
Guthrie, D., Van Kolfschoten, T., 2000. Neither warm and most, nor cold and arid:the ecology of the mid upper palaeolithic. In: Roebroeks, W., Mussi, M.,Svoboda, J., Fennema, K. (Eds.), Hunters of the Golden Age. The Mid UpperPalaeolithic of Eurasia, 30,000e20,000 BP. Leiden University Press, Leiden,pp. 13e20.
Haynes, G., 1980. Evidence of carnivore gnawing on pleistocene and recentmammalian bones. Paleobiology 6 (3), 341e351.
Haynes, G., 1983. A guide for differentiating mammalian carnivore taxa responsiblefor gnaw damage to herbivore limb bones. Paleobiology 9 (2), 164e172.
Hladilová, �S, 2005. Tertiary fossils, especially molluscs. Dolní V�estonice Studies. In:Svoboda, J. (Ed.), Pavlov I Southeast. A Window into Gravettian Lifestyles, vol.14, Brno, pp. 374e390.
Hladilová, �S, Mikulá�s, R., 2005. Fossil shark tooth. A remarkable working tool fromPavlov I locality. Dolní V�estonice Studies. In: Svoboda, J. (Ed.), Pavlov I South-east. A Window into Gravettian Lifestyles, vol. 14, Brno, pp. 391e395.
Kaminská, L., Kozlowski, J.K., Svoboda, J. (Eds.), 2006. Pleistocene Environments andArchaeology of the Dzeravá skala Cave. Lesser Carpathians, Slovakia. Kraków.
Klein, R.G., Cruz-Uribe, K., 1984. The Analysis of Animal Bones from ArchaeologicalSites. Univ. of Chicago Press, Chicago.
Klima, B., 1994. Die Knochenidustrie, Zier- und Kunstgegenstände. ERAUL 66/ DolniV�estonice Studies. In: Svoboda, J. (Ed.), Pavlov I e Excavations 1952e53, vol. 2,Liege, pp. 97e159.
Klima, B., 2005. Excavations at Pavlov I e 1954 and 1956. In: Svoboda, J. (Ed.), PavlovI Southeast. A window into Gravettian lifestyles, Brno, pp. 17e24.
Koz1owski, J.K., 1986. The gravettian in central and eastern Europe. Advances inWorld Archeology 5 New York, 131e200.
Koz1owski, J.K. (Ed.), 1998, Complex of the Upper Palaeolithic Sites Near Moravany,Western Slovakia, vol. II. Moravany-Lopata II (excavations 1993e1996),Kraków.
Laroulandie, V., 2003. Exploitation des Oiseaux au Magdalénien en France: Etat deslieux. In: Costamagno, S., Laroulandie, V. (Eds.), Mode de vie au Magdalénien:Apports de l’archéozoologie/Zooarchaeological insights into Magdalenian life-ways. Actes du colloque 6.4 du XIVe Congrès du l’UISPP, Liège, Belgique 2e8septembre 2001. BAR n� 1144, Oxford: Royaume-Uni.
Lipecki, G., Wojtal, P., 1998. Mammal remains. In: Koz1owski, J.K. (Ed.), Complex ofUpper Palaeolihic Sites Near Moravany, Western Slovakia, vol. 2. Moravany-Lopata (Excavations 1993e1996). Institute of Archaeology, Jagellonian Univer-sity, Cracow/Archaeological Institute, Slovak Academy of Sciences, Nitra,pp. 103e126.
Lupo, K.D., Schmitt, D.N., 2002. Upper paleolithic net-hunting, small prey exploi-tation, and women’s work effort: a view from the ethnographic and ethno-archaeological record of the Congo Basin. Journal of Archaeological Method andTheory 9 (2), 147e179.
Lyman, L., 1994. Vertebrae Taphonomy. Cambridge University Press, Cambridge.Lyman, L., 2008. Quantitative Paleozoology. Cambridge University Press, Cambridge.Münzel, S.C., Conard, N.J., 2004. Cave bear hunting in the Hohle Fels, a cave site in
the AchValley, Swabian Jura. Revue de Paléobiologie 23, 877e885.Musil, R., 1955. Osteologický materiál z paleolitického sídli�st�e v Pavlov�e. Acta Acad.
sc. cechoslov. Basis Brunensis 27 (6), 279e320.Musil, R., 1959. Osteologický materiál z paleolitického sídli�st�e v Pavlov�e. �Cást II.
Anthropozoikum 8, 83e106. 1958.Musil, R., 1994. The fauna. ERAUL 66/ Dolni V�estonice Studies. In: Svoboda, J. (Ed.),
Pavlov I. Excavations 1952e53, vol. 2, Liege, pp. 183e209.Musil, R., 2003. The middle and upper palaeolithic game Suite in central and
Southeastern Europe. In: van Andel, T.H., Davies, W. (Eds.), Neanderthals andModern Humans in the European Landscape during the Last Glaciation:Archaeological Results of the Stage 3 Project. Mc. Donald Institute Monographs,Cambridge, pp. 167e190.
Musil, R., 2005a. Animal prey. Dolni V�estonice Studies. In: Svoboda, J. (Ed.), Pavlov ISoutheast. A Window into Gravettian Lifestyles, vol. 14, Brno, pp. 190e228.
Musil, R., 2005b. Jaro�sov II e Podvr�st’a. A faunal anomaly among Gravettian sites.Osteological material analysis. In: �Skrdla, P. (Ed.), The Upper Palaeolithic on theMiddle Course of the Morava River, Brno, pp. 203e216.
Nývltová Fi�sáková, M., 2007. Seasonality of gravettian sites based on study ofmammal’s dental cement microstructures. P�rehled výzkum�u 48, 13e23.
Novák, M., 2005. Pavlov I e Southeast: review of spatial distributions. DolniV�estonice Studies. In: Svoboda, J. (Ed.), Pavlov I Southeast. A Window into theGravettian Lifestyles, vol. 14, Brno, pp. 53e71.
Oliva, M., 2009. Geography, stratigraphy and dating. In: Oliva, M. (Ed.), Milovice:Site of the Mammoth People Below the Pavlov Hills: The Question of MammothBone Structures, Brno, pp. 22e24.
Olsen, S.L., Shipman, P., 1988. Surface modification on bone: trampling versusButchery. Journal of Archaeological Science 15, 535e553.
Pangau-Adam, M., Noske, R., 2010. Wildlife hunting and bird trade in northernPapua (Irian Jaya), Indonesia. In: Tidemann, S., Gosler, A. (Eds.), Ethno-orni-thology. Birds, Indigenous Peoples, Culture and Society. Earthscan, London,Washington DC, pp. 73e85.
Pryor, A.J.E, 2006. A Preliminary Isotopic Study of the Ecology of the MoravianLocale Around 30,000ya as Seen through the Site of Dolní Vĕstonice II.Undergraduate Dissertation (Unpublished). University of Cambridge.
Purnama, S., Indrawan, M., 2010. Entrapment of wetland birds: local customs andmethods of hunting, Indramayu, central java. In: Tidemann, S., Gosler, A. (Eds.),Ethno-ornithology. Birds, Indigenous Peoples, Culture and Society. Earthscan,London, Washington DC, pp. 67e72.
Roebroeks, W., Mussi, M., Svoboda, J., Fennema, K. (Eds.), 2000. Hunters of theGolden Age. The Mid Upper Palaeolithic of Eurasia, 30,000e20,000 B.P. LeidenUniversity.
Serjeantson, D., 2009. Birds. Cambridge Manuals in Archaeology. CambridgeUniversity Press, Cambridge, New York, Melbourne.
Schiegl, S., Goldberg, P., Pfretzschner, H-U., Conard, N.J., 2003. Paleolithic Burnt BoneHorizons from the Swabian Jura: Distinguishing between In Situ Fireplaces andDumping Areas. Geoarchaeology: An International Journal 18 (5), 541e565.
Shahack-Gross, R., Bar-Yosef, O., Weiner, S., 1997. Black-coloured bones in Hayonimcave, Israel: differentiating between burning and oxide staining. Journal ofArchaeological Science 24, 439e446.
Shipman, P., Rose, J., 1983. Early hominid hunting, butchering, and carcass-processing behaviors: approaches to the fossil record. Journal of Anthropolog-ical Archaeology 2, 57e98.
Shipman, P., Foster, G., Schoeninger, M., 1984. Burnt bones and teeth: an experi-mental study of color, morphology, crystal structure and shrinkage. Journal ofArchaeological Science 11, 307e325.
Soffer, O., 1985. The Upper Palaeolithic of the Central Russian Plain. Academic Press,Orlando.
Soffer, O., Vandiver, P., 1994. The ceramics. In: Svoboda, J. (Ed.), Pavlov I. Excavations1952e53, vol. 2, pp. 161e174. ERAUL 66/Dolni V�estonice Studies.
Soffer, O., Adavasio, J.M., 2004. Textiles and upper paleolithic lives. A focus on theperishable and the invisible. Dolni V�estonice Studies. In: The Gravettian alongthe Danube, vol. 11, Brno, 270e282.
Stiner, M., 1994. Honor Among Thieves: A Zooarchaeological Study of NeandertalEcology. Princeton University Press, Princeton.

P. Wojtal et al. / Quaternary International 252 (2012) 122e141 141
Stiner, M.C., Kuhn, S.L., Weiner, S., Bar-Yosef, O., 1995. Differential burning, recrys-tallization, and fragmentation of archaeological bone. Journal of ArchaeologicalScience 22, 223e237.
Sutcliffe, A.J., 1970. Spotted hyaena: crusher, gnawer, digester and collector of bones.Nature 227, 1110e1113.
Svoboda, J., 1991. Dolní V�estonice II e Western Slope, ERAUL, vol. 54. Liege.Svoboda, J. (Ed.), 1994a, Pavlov I. Excavations 1952e53, vol. 2 ERAUL 66/Dolni
V�estonice Studies.Svoboda, J., 1994b. Afterword. In: Svoboda, J. (Ed.), Pavlov I. Excavations 1952e53,
ERAUL 66/Dolni V�estonice Studies, vol. 2, pp. 211e226.Svoboda, J., 2005. Pavlov I e southeast. Location, stratigraphy, microstratigraphies,
and features. Dolni V�estonice Studies. In: Svoboda, J. (Ed.), Pavlov I Southeast. AWindow into Gravettian Lifestyles, vol. 14, Brno, pp. 25e52.
Svoboda, J., 2007. The Gravettian on the Middle Danube, Le Gravettien: entitésrégionales d’une paléoculture européenne. Table ronde - Les Eyzies - juillet2004. Paleo 19, 203e220.
Svoboda, J., 2008. Formation of the archaeological layers, structure of the site andfeatures. In: Svoboda, J. (Ed.), Petrkovice. On shouldred points and femalefigurines, Brno, pp. 31e45.
Svoboda, J., Králík, M., �Culíková, V., Hladilová, �S, Novák, M., Nývltová Fi�sáková, M.,Nývlt, D., Zelinková, M., 2009. Pavlov VI: an Upper Palaeolithic living unit.Antiquity 83, 282e295.
Svoboda, J., Lozek, V., Vlcek, E., 1996. Hunters between East and West. The Palae-olithic of Moravia. Plenum, New York, London.
Svoboda, J., Péan, S., Wojtal, P., 2005. Mammoth bone deposits and subsistencepractices during mid-upper palaeolithic in central Europe: three cases fromMoravia and Poland. Quaternary International 126e128, 209e221.
Théry-Parisot, I., 2002. Fuel management (bone and wood) during the lower auri-gnacian in the pataud rock shelter (Lower Palaeolithic Les Eyzies de Tayac,Dordogne, France). Contribution of experimentation. Journal of ArchaeologicalScience 29, 1415e1421.
Tillet, T., Binford, L.R., 2002. L’Homme et l’Ours. Actes du colloque d’Auberives-en-Royans du 4 au 6 novembre 1997. Université de Liége. ERAUL 100.
Trinkaus, E., Svoboda, J.A. (Eds.), 2006, Early Modern Human Evolution in CentralEurope: The People of Dolní V�estonice and Pavlov, Dolní V�estonice Studies, vol.12. Oxford University Press, New York.
Trinkaus, E., Svoboda, J.A., Wojtal, P., Nývltová Fi�sáková, M., Wilczy�nski, J., 2010.Human remains from the Moravian gravettian: morphology and taphonomy ofAdditional elements from Dolní V�estonice II and Pavlov I. International Journalof Osteoarchaeology 20, 645e669.
Valde-Nowak, P., 2003. Upper palaeolithic sequence. In: Valde-Nowak, P.,Nadachowski, A., Madeyska, T. (Eds.), Ob1azowa Cave. Human Activity, Stratig-raphy and Palaeoenvironment, Kraków, pp. 44e68.
Verpoorte, A., 2000. Pavlovian reflexes and the Pompeii premise: a spatial analysisof stone artefacts from Pavlov I (Moravia, Czech Republic). ArcheologickéRozhledy 52, 577e594.
Verpoorte, A., 2005. Lithic assemblage of Pavlov I south-central (1954, 1956, 1963,1964). Dolni V�estonice Studies. In: Svoboda, J. (Ed.), Pavlov I Southeast. AWindow into the Gravettian Lifestyles, vol. 14, Brno, pp.75e111.
Villa, P., Bon, F., Castel, J.C., 2002. Fuel, fire and fireplaces in the palaeolithic ofwestern Europe. Review of Archaeology 23, 33e42.
Villaverde, V., Aura, J.E., Barton, C., 1998. The upper paleolithic in MediterraneanSpain: a review of current evidence. Journal of World Prehistory 12, 121e198.
West, D.L., 1996. Kraków-Spadzista, unit E and unit F. Faunal remains. In: the upperpalaeolithic site Kraków-Spadzista. Units D, E and F (Excavations1986e1989).Folia Quaternaria 67, 21e34.
West, D.L., 2001. Analysis of the fauna recovered from the 1986/1987 excava-tions at DolnívV�estonice II, western slope. Památky Archeologické 92,98e123.
Wijngaarden-Bakker, L.H., 1997. The selection of bird bones for artefact produc-tion at Dutch neolithic sites. International Journal of Osteoarchaeology 7,339e345.
Wojtal, P., 2007. Zooarchaeological Studies of the Late Pleistocene Sites in Poland.Institute of Systematics and Evolution of Animals, Kraków.
Wojtal, P., Sobczyk, K., 2005. Man and woolly mammoth at the Kraków SpadzistaStreet (B)e taphonomyof the site. Journal of Archaeological Science 32,193e206.
Copyright © 2022 FDOKUMEN




















