
Glucocorticoids induce mitochondrial gene transcription in HepG2 cells

Hindawi Publishing CorporationBioMed Research InternationalVolume 2013 Article ID 459017 9 pageshttpdxdoiorg1011552013459017
Research ArticleEffects of Tamarindus indica Fruit Pulp Extract onAbundance of HepG2 Cell Lysate Proteins and Their PossibleConsequential Impact on Metabolism and Inflammation
Ursula R W Chong1 Puteri S Abdul-Rahman12 Azlina Abdul-Aziz12
Onn H Hashim12 and Sarni Mat-Junit12
1 Department of Molecular Medicine Faculty of Medicine University of Malaya 50603 Kuala Lumpur Malaysia2 University of Malaya Centre for Proteomics Research Medical Biotechnology Laboratory Faculty of MedicineUniversity of Malaya 50603 Kuala Lumpur Malaysia
Correspondence should be addressed to Sarni Mat-Junit sarniumedumy
Received 17 October 2013 Accepted 28 November 2013
Academic Editor Antoni Camins
Copyright copy 2013 Ursula R W Chong et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
The fruit pulp extract of Tamarindus indica has been reported for its antioxidant and hypolipidemic properties In this studythe methanol extract of T indica fruit pulp was investigated for its effects on the abundance of HepG2 cell lysate proteinsCell lysate was extracted from HepG2 cells grown in the absence and presence of the methanol extract of T indica fruit pulpApproximately 2500 spots were resolved using two-dimensional gel electrophoresis and the abundance of 20 cellular proteinswas found to be significantly reduced Among the proteins of reduced abundance fourteen including six proteins involvedin metabolism (including ethanolamine phosphate cytidylyltransferase) four mitochondrial proteins (including prohibitin andrespiratory chain proteins) and four proteins involved in translation and splicing were positively identified by mass spectrometryand database search The identified HepG2 altered abundance proteins when taken together and analyzed by Ingenuity PathwaysAnalysis (IPA) software are suggestive of the effects of T indica fruit pulp extract on metabolism and inflammation which aremodulated by LXRRXR In conclusion the methanol fruit pulp extract of T indica was shown to cause reduced abundance ofHepG2 mitochondrial metabolic and regulatory proteins involved in oxidative phosphorylation protein synthesis and cellularmetabolism
1 Introduction
Tamarindus indica or tamarind is a tropical fruit tree nativeto the African savannahs but it can now be found in manytropical countries It is categorized as a monospecific genusin the family of Leguminosae The sweet and sour taste of itsfruit pulp is used to add flavor to local cuisines Besides culi-nary tamarind is also used in traditionalmedicine as laxativediuretic antibacterial agents as well as in treatment of feverand malarial infections [1 2] Previous biochemical analyseshave demonstrated that extracts of T indica possess highantioxidant activities [3 4] In addition T indica extractshave also been shown to reduce the levels of blood cholesteroland triacylglycerol in hypercholesterolemic hamsters [3]and in humans [5] However the molecular mechanisms of
the anti-inflammatory and hypolipidemic effects of the fruitremain elusive
In our previous study the methanol extract of T indicafruit pulp was shown to alter the expression of more than athousand genes in HepG2 cells many of which are associatedwith lipid metabolism [6] Our recent study also showedthat the fruit pulp extract was able to alter the secretionof alpha enolase apolipoprotein A-1 transthyretin and rabGDP dissociation inhibitor beta from HepG2 cells whichmay account for the lipid-lowering effects of the fruit Theseeffects were hypothesized to occur via activation of LXRRXR [7]
The lipid-lowering properties of T indica fruit pulpare likely attributed to the presence of polyphenols in its
2 BioMed Research International
extract The methanol extract of T indica fruit pulp con-tains predominantly proanthocyanidins of various formsincluding (+)-catechin [4 8] and (minus)-epicatechin along withtaxifolin apigenin eriodictyol luteolin and naringenin [4]Proanthocyanidins from grape seeds have been shown tolower the levels of blood cholesterol and triacylglycerol inhamsters through excretion of bile [9] In addition otherpolyphenols like naringenin [10] and tea catechins [11] havealso been shown to have effects on transcriptional regulationof hepatic lipid metabolism in rats and humans
In this study the methanol extract of T indica fruit pulpwas further investigated for its effects on the abundanceof HepG2 cell lysate proteins by two-dimensional gel elec-trophoresis and densitometry Identification of HepG2 celllysate proteins that are significantly altered in abundancemayhelp to improve our understanding of themetabolic pathwaysandmolecular mechanisms that are affected by the fruit pulp
2 Materials and Methods
21 Preparation of Methanol Extract of the T indica FruitPulp T indica fruit pulps were collected at Universiti PutraMalaysia and the fruits were identified through comparisonwith a registered voucher specimen (KLU45976) deposited inthe Herbarium of Institute of Biological Sciences Universityof Malaya T indica fruit pulp extract was prepared aspreviously described [6] with slight modifications Brieflyripe fruit pulp of T indica was separated from the seedsair-dried and powdered The powdered fruit pulp (10 g) wasplaced in a conical flask and soaked in 200mL methanolat room temperature The mixture was then stirred witha magnetic stirrer for 1 h and kept in the dark for 24 hThe resulting extract was then filtered and dried in a rotaryevaporator and finally redissolved in 10 DMSO Sampleswere kept at minus20∘C until further analysis
22 Cell Culture Human hepatoma HepG2 cell line (ATCCManassas VA USA) was grown in a complete mediumconsisting of Dulbeccorsquos modified Eaglersquos medium (DMEM)supplemented with 5mM glucose 10 foetal bovine serum(FBS HyClone Australia) 037 (wv) sodium bicarbonateand 048 (wv) HEPES pH 74 in a CO
2humid incubation
chamber at 37∘C
23 Treatment of HepG2 Cells with T indica Fruit Pulp Extractand Preparation of Cell Lysate HepG2 cells were seeded ata density of 90 times 106 in a 75 cm2 flask for 18ndash24 h followedby extensive washing with phosphate-buffered saline (PBS)The cells were then incubated for 24 h in serum-free mediumin the presence of 002 DMSO (vehicle) as controls or afinal concentration of 60120583gmLmethanol extract of T indicafruit pulp The cells were then lysed in rehydration buffer(7Murea 2M thiourea 2wv CHAPS 05 vv IPG bufferorange G protease inhibitor) and protein concentration oflysate was determined using the Bradford assay kit (Bio-radHercules CA USA)
24 Two-Dimensional Gel Electrophoresis (2D-GE) Forty 120583gof cell lysate proteins were cleaned using the 2D clean-up kit(GE Piscataway USA) The resulting protein pellet was thenreconstituted in rehydration solution containing 7M urea2M thiourea 2 wv CHAPS 05 vv IPG buffer orange Gand protease inhibitor Immobiline pH gradient strip (13 cmnon-linear pH 3ndash10 GE Healthcare Uppsala Sweden) wasthen rehydrated in the reconstituted protein sample for18 h at room temperature First dimension separation wasperformed on an Ettan IPGphor III (GE Uppsala Sweden)ran on the following conditions (i) 500V 1 hr 10mins stepand hold (ii) 1000V 1 hr gradient (iii) 8000V 2 hrs 30minsgradient and (iv) 8000V 55mins step and hold The stripswere then equilibrated in SDS equilibration buffer containing6M urea 75mM Tris-HCl pH 88 293 vv glycerol 2wv SDS 0002 wv bromophenol blue and 1 wv dithio-threitol (DTT) for 15min followed by a second equilibrationusing the same buffer containing 45 wv iodoacetamideinstead of DTT for another 15min Second dimension sepa-ration was carried out on 120 polyacrylamide gel using theSE 600Ruby electrophoresis system (GEHealthcareUppsalaSweden) at a constant voltage of 50V and 40mAgel for20min and then switched to 500V and 40mAgel until thebromophenol bluemarker was 1mmaway from the bottomofthe gel Gels were silver-stained with PlusOne Silver StainingKit (GE Healthcare Uppsala Sweden) and scanned using theImageScanner III (GE Healthcare Uppsala Sweden)
25 Image and Data Analyses Gel images were analyzedusing the ImageMaster 2D Platinum V 70 software (GEHealthcare Uppsala Sweden) Briefly the 2D gel images weresubjected to spot detection and quantification in the differ-ential in-gel analyses module Protein spots were normalizedusing percentage of volume to minimize variations betweengels within the same group Statistically significance (119875 lt005 Studentrsquos t-test) and presence in all 6 gels were the twocriteria for acceptance of protein spots of altered abundanceSelected spots were filtered based on an average expressionlevel change of at least 15-fold
26 In-Gel Tryptic Digestion Protein spots were excisedmanually from 2D-GE gels and washed with 100mMNH4HCO3for 15min The gel plugs were then destained
twice with 15mM potassium ferricyanide50mM sodiumthiosulphate with shaking They were then reduced with10mM DTT at 60∘C for 30min and alkylated with 55mMiodoacetamide in the dark at room temperature for 20minThe plugs were later washed thrice with 500120583L of 50ACN50mM NH
4HCO3for 20min dehydrated with 100
ACN for 15min and dried using the SpeedVac The gel plugswere finally digested in 6 ng120583L trypsin (Pierce RockfordIL USA) in 50mM NH
4HCO3at 37∘C for at least 16 h
Peptide mixtures were extracted twice with 50 ACN and100 ACN respectively and finally concentrated using theSpeedvac until completely dry Dried peptides were then keptat minus20∘C or reconstituted with 10 120583L of 01 TFA prior todesalting using the Zip Tip C18 micropipette tips (MilliporeBillerica MA USA)
BioMed Research International 3
27 Mass Spectrometry and Database Search Trypsin digest-ed peptidemixturewas analyzed using anAppliedBiosystems4800 Plus MALDI-TOFTOF (Foster City CA USA) Thepeptide mixture was crystallized with alpha-cyano-4-hydroxycinnamic acid matrix solution (10mgmL 70 ACNin 01 (vv) TFA aqueous solution) and spotted onto aMALDI target (192-well) plate The MS results were auto-matically acquiredwith a trypsin autodigest exclusion list and20 most intense ions were selected for MSMS analysis Dataanalysis was carried out using the GPS Explorer software(Applied Biosystems CA USA) and database search usingthe in-house MASCOT program (Matrix Science LondonUK) Combined MS and MSMS searches were conductedwith the following settings Swiss-Prot database Homosapiens peptide tolerance at 200 ppm MSMS tolerance at04Da carbamidomethylation of cysteine (variable modifi-cation) and methionine oxidation (variable modifications)A protein is considered identified when a MASCOT scoreof higher than 55 and 119875 lt 005 were obtained from the MSanalysis
28 Functional Analyses Using Ingenuity Pathways Analysis(IPA) Software Functional analyses to predict networks thatare affected by the differentially expressed proteins werecarried out using Ingenuity Pathways Analysis (IPA) software(Ingenuity Systems httpwwwingenuitycom) Details ofthe proteins their quantitative expression values (fold changedifference of at least 15) and 119875 values (119875 lt 005) wereimported into the IPA software Each protein identifierwas mapped to its corresponding protein object and wasoverlaid onto a global molecular network developed frominformation contained in the Ingenuity Knowledge BaseNetwork predictions based on the protein input were gener-ated algorithmically by utilizing the information containedin the Ingenuity Knowledge Base Right-tailed Fischerrsquos exacttest was used to calculate a 119875 value indicating the probabilitythat each biological function assigned to the network is dueto chance alone
29 Validation of Proteins of Reduced Abundance Three ofthe altered proteins ethanolamine phosphate cytidylyltrans-ferase (PCYT2) and NADH dehydrogenase (ubiquinone) 1alpha subcomplex subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reductase complex core protein 2 (UQCRC2)were selected for validation using Western blotting HepG2cells (3 times 106) were treated with 60120583gmL T indica fruit pulpextract for 24 h The cells were then trypsinized and lysedwith RIPA buffer (150mM NaCl 50mM Tris-HCl pH 741mMEDTA 1 Triton X-100 1 sodium deoxycholate 01SDS) Total cell lysate proteins were quantified using BCAassay kit (Pierce Rockford IL USA) Forty micrograms ofcell lysate protein was separated on a 125 SDS-PAGE andtransferred onto a PVDF membrane with 045120583m pore size(Thermo Scientific IL USA) at 100V 110mA for 90 minutesThe blot was then blocked overnight and developed againstanti-PCYT2 (ab126142 rabbit polyclonal Abcam UK) anti-NDUFA10 (ab103026 rabbit polyclonal Abcam UK) anti-UQCRC2 (ab103616 rabbit polyclonal Abcam UK) and
Table 1 Average percentage of volume of spots
Spot ID Average percentage of volume plusmn SEMControl Treated
363 00273 plusmn 00016 00187 plusmn 00019398 00214 plusmn 00021 00134 plusmn 00020540 00128 plusmn 00019 00073 plusmn 00011590 00892 plusmn 00073 00609 plusmn 00039615 00240 plusmn 00023 00157 plusmn 00016637 00254 plusmn 00033 00156 plusmn 00011655 00499 plusmn 00033 00328 plusmn 00056657 00655 plusmn 00088 00360 plusmn 00035675 00660 plusmn 00067 00440 plusmn 00017722 00223 plusmn 00022 00144 plusmn 00014751 00420 plusmn 00038 00235 plusmn 00027765 00470 plusmn 00031 00315 plusmn 00040796 00162 plusmn 00011 00093 plusmn 00009900 00394 plusmn 00048 00206 plusmn 00044914 00279 plusmn 00015 00177 plusmn 000121084 00305 plusmn 00015 00209 plusmn 000171254 00154 plusmn 00018 00078 plusmn 000071355 00107 plusmn 00018 00054 plusmn 000101634 00746 plusmn 00125 00435 plusmn 000311649 00639 plusmn 00069 00416 plusmn 00032The protein spots show significant differences in their abundance (119875 lt 005)in T indica-treated and nontreated (control) HepG2 cells by more than 15-fold
anti-beta actin (ab8227 rabbit polyclonal Abcam UK) as theloading control using the WesternDot 625 Goat Anti-RabbitWestern Blot kit (Invitrogen Oregon USA) Quantificationof the band intensity was calculated using ImageJ software
3 Results
31 2D-GE Analysis of HepG2 Cell Lysate Proteins Analysisof HepG2 cell lysate by 2D-GE resolved more than 2500protein spots in each gel Figure 1 demonstrates typical 2D-GE resolved patterns of the cell lysate proteins from controlsand HepG2 cells treated with the fruit pulp extract of Tindica The percentage of volume contribution of each spotwas then determined using ImageMaster 2D Platinum V 70software and the fold change if any was acquired Proteinspots that were present in all gels (119899 = 6) and showedsignificant differences in their abundance (119875 lt 005) intreated and nontreated cells by more than 15-fold wereselected Based on these criteria the altered abundance of 20protein spots were detected when HepG2 cells were exposedto the fruit pulp extract of T indica (Table 1)
32 Identification of Altered Abundance HepG2 Cell LysateProteins Among the 20 protein spots that were significantlyreduced in abundance 14 were successfully identified bymass spectrometry and database search (Table 2) Six spots(protein spot ID 398 615 637 1254 1355 and 1649) wereconsidered not successfully identified as their scores werelower than the cut-off value for positive inclusion criteria
4 BioMed Research International
MW
(kD
a)
180120
40
80
50
12
25
20
10
pH 3ndash10 nonlinear
Control Treated
Control
Treated
Figure 1 A representative proteome map of HepG2 cell lysate Approximately 2500 spots per gel within the pH 3ndash10 range for cell lysate ofHepG2 cells were detected Twenty spots were altered in abundance (circled and labelled) and they were all significantly reduced in abundance(119875 lt 005)
The 14 identified proteins may be grouped according totheir biological processes using the UniProt Protein Knowl-edgebase (UniProtKB) NADH dehydrogenase (ubiquinone)1 alpha subcomplex subunit 10 ubiquinol-cytochrome-creductase complex core protein 2 andNADHdehydrogenase(ubiquinone) flavoprotein 1 were grouped under ldquomitochon-drial respiratory chainrdquo category Three proteins that iseukaryotic translation initiation factor 3 subunit 3 elonga-tion factor Tu and tyrosyl-tRNA synthetase are involvedin ldquoprotein synthesisrdquo Another six proteins were catego-rized under ldquometabolismrdquo with glyceraldehyde-3-phosphatedehydrogenase and GDP-L-fucose synthetase being involvedin carbohydrate metabolism GMP reductase 2 and UMPsynthase in nucleotide and nucleosidemetabolism S-methyl-5-thioadenosine phosphorylase in polyamine metabolic pro-cess and ethanolamine phosphate cytidylyltransferase inbiosynthesis of phospholipids Prohibitin on the other handwas categorized under ldquocell proliferation and differentiationrdquowhile heterogeneous nuclear ribonucleoprotein H3 does notbelong to any of the above groups and therefore categorizedas ldquoothersrdquo
33 Biological Processes and Pathway Interaction AnalysisThe cell lysate proteins of altered abundance when analyzedusing Ingenuity Pathways Analysis (IPA) software generateda single network on ldquoHereditary disorder metabolic diseasemolecular transportrdquo with a score of 48 (Table 3) Mitochon-drial dysfunction was ranked with the highest significance(119875 lt 365 times 10minus4) from a canonical pathway analysis When
the data was reanalyzed to include our earlier reported resultsof HepG2 proteins that were differentially secreted whenthe cells were exposed to the T indica fruit pulp extract[7] the software identified ldquoLipid Metabolism MolecularTransport and Small Molecule Biochemistryrdquo as top putativenetwork (score of 31) that links the proteins of alteredabundance with other interactomes and gluconeogenesis Ibecame the top canonical pathway (119875 lt 467times10minus4) (Table 3)Figure 2 shows a graphical representation of the predictedmolecular relationships between HepG2 proteins that werealtered in abundance in response to exposure to T indicafruit pulp extract Other than the lipid-related interactomesthe HepG2 proteins of altered abundance were shown to beassociated with inflammation-related molecules like tumournecrosis factor (TNF) and interleukin-1 beta (IL-1120573) Twotranscription regulators mediator of RNA polymerase IIItranscription subunit 30 (MED30) and transcription factorE2F1 (E2F1) were also shown to be interconnected with theproteins
34 Confirmation of the Effects of T indica Fruit Pulp Extracton HepG2 Proteins To validate the effects of the T indicafruit pulp extract on HepG2 proteins Western blotting wasperformed using antisera raised against the cellular proteinsIn view of the scarce amount of HepG2 cell lysate proteinextract that was generated in this study three proteins thatis ethanolamine phosphate cytidylyltransferase (PCYT2)NADH dehydrogenase (ubiquinone) 1 alpha subcomplex
BioMed Research International 5
Table2Listof
celllysateproteins
ofalteredabun
danceinTindica
fruitextract-tr
eatedcells
identifi
edby
MALD
I-TOFTO
FMSMS
Spot
no
Proteindescrip
tion
Accno
Score
pIM
w(kDa)
Av
FCa
Cov
bMatched
peptide
sequ
ences
Functio
nal
category
657
NADHdehydrogenase(ub
iquino
ne)1
alph
asub
complex
subu
nit10(C
I-42
kD)
O95299
194
8674073
minus18
44117
ndash12213ndash139140ndash
161253ndash261
290ndash
295
Mito
chon
drial
respira
tory
chain
765
Ubiqu
inol-cytochrom
e-cr
eductase
complex
core
protein2mito
chon
drial
precursorc
orep
rotein
IIP2
2695
537
8744841
minus15
2871ndash8
4163ndash183184ndash
196200ndash
217232ndash241
900
NADHdehydrogenase(ub
iquino
ne)
flavoprotein1mito
chon
drialprecursor
(CI-51kD
)P4
9821
275
8515079
minus19
3372ndash81153ndash159160ndash
174376ndash
386
441ndash44
9
796
Ethano
lamine-ph
osph
ate
cytid
ylyltransfe
rase
Q9944
7339
644
4381
minus17
38185ndash199262ndash271333ndash348
Phosph
olipid
biosynthesis
590
Glyceraldehyde-3-ph
osph
ate
dehydrogenase(GAPD
H)
P044
06192
8573603
minus15
18235ndash248310ndash
323
Carbo
hydrate
metabolism
722
GDP-L-fucose
synthetase
Q13630
280
6133587
minus15
4926ndash4
482ndash8890ndash107200ndash214291ndash297
307ndash320
655
GMPredu
ctase2
Q9P
2T1
386
679378
5minus15
5270ndash78178ndash189192ndash213277ndash286
278ndash286292ndash298292ndash306
Nucleotidea
ndnu
cleoside
metabolism
914
Urid
ine51015840
-mon
opho
sphatesynthase
(UMPsynthase)
P1117
2379
6815220
minus16
376ndash
2230ndash
41146
ndash155353ndash363389ndash4
05
460ndash
467469ndash
477
363
S-methyl-5
-thioadenosinep
hospho
rylase
(MTA
P)Q13126
253
675312
3minus15
4312ndash2972ndash8283ndash9
9100ndash
116134ndash
147
181ndash187272ndash282
Polyam
ine
metabolism
1634
Proh
ibitin
P35232
66227297
9minus17
21134ndash
143
Cellproliferation
anddifferentiatio
n
675
Eukaryotictransla
tioninitiationfactor
3subu
nit3
(elF3H
)O15372
279
609399
1minus15
4352ndash75242ndash249260ndash
265304ndash
313
314ndash
331
Protein
biosynthesis
751
Elon
gatio
nfactor
Tum
itochon
drial
precursor(EF
-Tu)
P49411
11472
64951minus18
29105ndash120239ndash
252
1084
Tyrosyl-tRN
Asynthetasecytop
lasm
ic(TyrRS
)P5
4577
96661591
1minus15
2185ndash93179ndash
189433ndash450
540
Heterogenou
snuclear
ribon
ucleop
rotein
H3(hnR
NPH3)
P31942
409
6373690
minus17
3956ndash6
785ndash9
098ndash104
206ndash222223ndash232
262ndash287288ndash301
Others
a Negativev
alue
signifiesd
own-regu
lationin
term
soffold-differencesA
llratio
sare
statisticallysig
nificantw
ith119875lt005(Studentrsquos119905-te
st)
b Coverageo
fthe
identifi
edsequ
ence
6 BioMed Research International
Table 3 Top network and canonical pathways generated by Ingenuity Pathways Analysis
Proteins subjectto analysis
Associated networkfunctions Scoreb Top canonical
pathway 119875 value
Cell lysateproteins
Hereditary disorder metabolicdisease molecular transport 48 Mitochondrial
Dysfunction 365119864 minus 04
Cell lysate andsecretoryproteinsa
Lipid metabolism moleculartransport small moleculeBiochemistry
31 Gluconeogenesis I 467119864 minus 04
aEarlier reported [7]
bA score of 2 or higher indicates at least a 99 confidence of not being generated by random chance and higher scores indicate a greater confidence
subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reduc-tase complex core protein 2 (UQCRC2) that represent thoseinvolved in the mitochondrial and metabolic activities wereselected Densitometry scanning of bands detected by theantisera showed that the abundance of PCYT2 NDUFA10and UQCRC2 was indeed lower in HepG2 cells treated withthe T indica fruit pulp extract compared to the controls withfold differences of ndash17 ndash15 and ndash15 respectively (Figure 3)
4 Discussion
In this study the abundance of 20 cell lysate proteins wasfound to be significantly reduced when HepG2 cells wereexposed to the T indica fruit pulp extract Fourteen of theproteins were identified by mass spectrometry and databasesearch and the reduced abundance of three of the HepG2proteins was subsequently validated by Western blottingDue to limited availability of antibodies and HepG2 celllysate protein extract validation was performed on thethree selective proteins that are representative of separatemitochondrial functions and metabolic activities
Among the identified HepG2 proteins three com-ponents of the mitochondrial respiratory chain namelyubiquinol-cytochrome-c reductase complex core protein 2(UQCRC2) NADH dehydrogenase (ubiquinone) 1 alphasubcomplex subunit 10 (NDUFA10) and NADH dehydroge-nase (ubiquinone) flavoprotein 1 (NDUFV1) were found tobe reduced in abundance when HepG2 cells were exposedto T indica fruit pulp extract UQCRC2 belongs to complexIII of the mitochondrial respiratory chain while NDUFA10andNDUFV1 are components of complex I Inmitochondriaonly complex I [12 13] and complex III [14] of the respiratorychain are known to produce reactive oxygen species (ROS)Hence the decreased amount of the three mitochondrialcomponents may indicate reduced production of free rad-icals although the functionality of the mitochondria is yetto be confirmed Nevertheless earlier studies in obese ratmuscles have shown that reduced levels of respiratory chaincomplex I and diminishedROS production that were inducedby chronic supplementation with grape seed proanthocyani-dins did not affect the function of the mitochondria [15]
Prohibitin is another mitochondrial protein that wasshown to be of reduced abundance in HepG2 cells exposedto T indica fruit pulp extract The protein that was initiallythought to be a negative regulator of cell proliferation has
been demonstrated to play a role in biogenesis and function ofmitochondria [16] In a study of the nematodeCaenorhabditiselegans prohibitin complex was shown to promote longevityby modulating mitochondrial function and fat metabolism[17] Deficiency of prohibitin prolongs the lifespan of Celegans with compromised mitochondrial function or fatmetabolism In this study depletion of prohibitin in HepG2cells upon exposure to the fruit pulp extract of T indicaappears to suggest a similar mechanism in an attempt toextend lifespan of the cells
In addition to the mitochondrial proteins the gly-colytic enzyme glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as well as GDP-L-fucose synthetase (TSTA3) alsoappeared to be down-regulated after treatment with T indicafruit pulp extract Oxidative stress is known to induce up-regulation of GAPDH levels [18 19] Hence the reducedamount of GAPDH in this case may possibly indicate astate of repressed oxidative stress This together with theearlier data on the reduced mitochondrial respiratory chainproteins may shed some light on molecular mechanismsinvolved in the well acclaimed antioxidant properties of Tindica [3 4]
The fruit pulp extract of T indica also appeared tocause decreased abundance of proteins involved in themetabolism of nucleic acids and polyamines in HepG2 cellsTo the best of our knowledge these have not been previ-ously reported and their rationale is not quite understoodOn the other hand decreased abundance of ethanolaminephosphate cytidylyltransferase (PCYT2) the rate-limitingenzymewhich catalyzes conversion of phosphoethanolamineto cytidylylphosphoethanolamine in the biosynthesis ofphosphatidylethanolamine in HepG2 cells exposed to thefruit extract of T indica may compromise availability ofthe phospholipid which stores arachidonic acid for theproduction of prostaglandinsThis could possibly explain theanti-inflammatory action ofT indica thatwas earlier reported[20]
In addition eukaryotic translation initiation factor 3 sub-unit 3 (eIF3H) tyrosyl-tRNA synthetase (YARS) elongationfactor Tu (EFTU) and heterogenous nuclear ribonucleopro-tein H3 (hnRNPH3) which are generally involved in proteinsynthesis and splicing also appeared to be down-regulated inHepG2 cells treated with the T indica extract These proteinsmay be key regulatory points of action of the fruit pulp extractof T indica Suppression of these proteins by the extract may
BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

2 BioMed Research International
extract The methanol extract of T indica fruit pulp con-tains predominantly proanthocyanidins of various formsincluding (+)-catechin [4 8] and (minus)-epicatechin along withtaxifolin apigenin eriodictyol luteolin and naringenin [4]Proanthocyanidins from grape seeds have been shown tolower the levels of blood cholesterol and triacylglycerol inhamsters through excretion of bile [9] In addition otherpolyphenols like naringenin [10] and tea catechins [11] havealso been shown to have effects on transcriptional regulationof hepatic lipid metabolism in rats and humans
In this study the methanol extract of T indica fruit pulpwas further investigated for its effects on the abundanceof HepG2 cell lysate proteins by two-dimensional gel elec-trophoresis and densitometry Identification of HepG2 celllysate proteins that are significantly altered in abundancemayhelp to improve our understanding of themetabolic pathwaysandmolecular mechanisms that are affected by the fruit pulp
2 Materials and Methods
21 Preparation of Methanol Extract of the T indica FruitPulp T indica fruit pulps were collected at Universiti PutraMalaysia and the fruits were identified through comparisonwith a registered voucher specimen (KLU45976) deposited inthe Herbarium of Institute of Biological Sciences Universityof Malaya T indica fruit pulp extract was prepared aspreviously described [6] with slight modifications Brieflyripe fruit pulp of T indica was separated from the seedsair-dried and powdered The powdered fruit pulp (10 g) wasplaced in a conical flask and soaked in 200mL methanolat room temperature The mixture was then stirred witha magnetic stirrer for 1 h and kept in the dark for 24 hThe resulting extract was then filtered and dried in a rotaryevaporator and finally redissolved in 10 DMSO Sampleswere kept at minus20∘C until further analysis
22 Cell Culture Human hepatoma HepG2 cell line (ATCCManassas VA USA) was grown in a complete mediumconsisting of Dulbeccorsquos modified Eaglersquos medium (DMEM)supplemented with 5mM glucose 10 foetal bovine serum(FBS HyClone Australia) 037 (wv) sodium bicarbonateand 048 (wv) HEPES pH 74 in a CO
2humid incubation
chamber at 37∘C
23 Treatment of HepG2 Cells with T indica Fruit Pulp Extractand Preparation of Cell Lysate HepG2 cells were seeded ata density of 90 times 106 in a 75 cm2 flask for 18ndash24 h followedby extensive washing with phosphate-buffered saline (PBS)The cells were then incubated for 24 h in serum-free mediumin the presence of 002 DMSO (vehicle) as controls or afinal concentration of 60120583gmLmethanol extract of T indicafruit pulp The cells were then lysed in rehydration buffer(7Murea 2M thiourea 2wv CHAPS 05 vv IPG bufferorange G protease inhibitor) and protein concentration oflysate was determined using the Bradford assay kit (Bio-radHercules CA USA)
24 Two-Dimensional Gel Electrophoresis (2D-GE) Forty 120583gof cell lysate proteins were cleaned using the 2D clean-up kit(GE Piscataway USA) The resulting protein pellet was thenreconstituted in rehydration solution containing 7M urea2M thiourea 2 wv CHAPS 05 vv IPG buffer orange Gand protease inhibitor Immobiline pH gradient strip (13 cmnon-linear pH 3ndash10 GE Healthcare Uppsala Sweden) wasthen rehydrated in the reconstituted protein sample for18 h at room temperature First dimension separation wasperformed on an Ettan IPGphor III (GE Uppsala Sweden)ran on the following conditions (i) 500V 1 hr 10mins stepand hold (ii) 1000V 1 hr gradient (iii) 8000V 2 hrs 30minsgradient and (iv) 8000V 55mins step and hold The stripswere then equilibrated in SDS equilibration buffer containing6M urea 75mM Tris-HCl pH 88 293 vv glycerol 2wv SDS 0002 wv bromophenol blue and 1 wv dithio-threitol (DTT) for 15min followed by a second equilibrationusing the same buffer containing 45 wv iodoacetamideinstead of DTT for another 15min Second dimension sepa-ration was carried out on 120 polyacrylamide gel using theSE 600Ruby electrophoresis system (GEHealthcareUppsalaSweden) at a constant voltage of 50V and 40mAgel for20min and then switched to 500V and 40mAgel until thebromophenol bluemarker was 1mmaway from the bottomofthe gel Gels were silver-stained with PlusOne Silver StainingKit (GE Healthcare Uppsala Sweden) and scanned using theImageScanner III (GE Healthcare Uppsala Sweden)
25 Image and Data Analyses Gel images were analyzedusing the ImageMaster 2D Platinum V 70 software (GEHealthcare Uppsala Sweden) Briefly the 2D gel images weresubjected to spot detection and quantification in the differ-ential in-gel analyses module Protein spots were normalizedusing percentage of volume to minimize variations betweengels within the same group Statistically significance (119875 lt005 Studentrsquos t-test) and presence in all 6 gels were the twocriteria for acceptance of protein spots of altered abundanceSelected spots were filtered based on an average expressionlevel change of at least 15-fold
26 In-Gel Tryptic Digestion Protein spots were excisedmanually from 2D-GE gels and washed with 100mMNH4HCO3for 15min The gel plugs were then destained
twice with 15mM potassium ferricyanide50mM sodiumthiosulphate with shaking They were then reduced with10mM DTT at 60∘C for 30min and alkylated with 55mMiodoacetamide in the dark at room temperature for 20minThe plugs were later washed thrice with 500120583L of 50ACN50mM NH
4HCO3for 20min dehydrated with 100
ACN for 15min and dried using the SpeedVac The gel plugswere finally digested in 6 ng120583L trypsin (Pierce RockfordIL USA) in 50mM NH
4HCO3at 37∘C for at least 16 h
Peptide mixtures were extracted twice with 50 ACN and100 ACN respectively and finally concentrated using theSpeedvac until completely dry Dried peptides were then keptat minus20∘C or reconstituted with 10 120583L of 01 TFA prior todesalting using the Zip Tip C18 micropipette tips (MilliporeBillerica MA USA)
BioMed Research International 3
27 Mass Spectrometry and Database Search Trypsin digest-ed peptidemixturewas analyzed using anAppliedBiosystems4800 Plus MALDI-TOFTOF (Foster City CA USA) Thepeptide mixture was crystallized with alpha-cyano-4-hydroxycinnamic acid matrix solution (10mgmL 70 ACNin 01 (vv) TFA aqueous solution) and spotted onto aMALDI target (192-well) plate The MS results were auto-matically acquiredwith a trypsin autodigest exclusion list and20 most intense ions were selected for MSMS analysis Dataanalysis was carried out using the GPS Explorer software(Applied Biosystems CA USA) and database search usingthe in-house MASCOT program (Matrix Science LondonUK) Combined MS and MSMS searches were conductedwith the following settings Swiss-Prot database Homosapiens peptide tolerance at 200 ppm MSMS tolerance at04Da carbamidomethylation of cysteine (variable modifi-cation) and methionine oxidation (variable modifications)A protein is considered identified when a MASCOT scoreof higher than 55 and 119875 lt 005 were obtained from the MSanalysis
28 Functional Analyses Using Ingenuity Pathways Analysis(IPA) Software Functional analyses to predict networks thatare affected by the differentially expressed proteins werecarried out using Ingenuity Pathways Analysis (IPA) software(Ingenuity Systems httpwwwingenuitycom) Details ofthe proteins their quantitative expression values (fold changedifference of at least 15) and 119875 values (119875 lt 005) wereimported into the IPA software Each protein identifierwas mapped to its corresponding protein object and wasoverlaid onto a global molecular network developed frominformation contained in the Ingenuity Knowledge BaseNetwork predictions based on the protein input were gener-ated algorithmically by utilizing the information containedin the Ingenuity Knowledge Base Right-tailed Fischerrsquos exacttest was used to calculate a 119875 value indicating the probabilitythat each biological function assigned to the network is dueto chance alone
29 Validation of Proteins of Reduced Abundance Three ofthe altered proteins ethanolamine phosphate cytidylyltrans-ferase (PCYT2) and NADH dehydrogenase (ubiquinone) 1alpha subcomplex subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reductase complex core protein 2 (UQCRC2)were selected for validation using Western blotting HepG2cells (3 times 106) were treated with 60120583gmL T indica fruit pulpextract for 24 h The cells were then trypsinized and lysedwith RIPA buffer (150mM NaCl 50mM Tris-HCl pH 741mMEDTA 1 Triton X-100 1 sodium deoxycholate 01SDS) Total cell lysate proteins were quantified using BCAassay kit (Pierce Rockford IL USA) Forty micrograms ofcell lysate protein was separated on a 125 SDS-PAGE andtransferred onto a PVDF membrane with 045120583m pore size(Thermo Scientific IL USA) at 100V 110mA for 90 minutesThe blot was then blocked overnight and developed againstanti-PCYT2 (ab126142 rabbit polyclonal Abcam UK) anti-NDUFA10 (ab103026 rabbit polyclonal Abcam UK) anti-UQCRC2 (ab103616 rabbit polyclonal Abcam UK) and
Table 1 Average percentage of volume of spots
Spot ID Average percentage of volume plusmn SEMControl Treated
363 00273 plusmn 00016 00187 plusmn 00019398 00214 plusmn 00021 00134 plusmn 00020540 00128 plusmn 00019 00073 plusmn 00011590 00892 plusmn 00073 00609 plusmn 00039615 00240 plusmn 00023 00157 plusmn 00016637 00254 plusmn 00033 00156 plusmn 00011655 00499 plusmn 00033 00328 plusmn 00056657 00655 plusmn 00088 00360 plusmn 00035675 00660 plusmn 00067 00440 plusmn 00017722 00223 plusmn 00022 00144 plusmn 00014751 00420 plusmn 00038 00235 plusmn 00027765 00470 plusmn 00031 00315 plusmn 00040796 00162 plusmn 00011 00093 plusmn 00009900 00394 plusmn 00048 00206 plusmn 00044914 00279 plusmn 00015 00177 plusmn 000121084 00305 plusmn 00015 00209 plusmn 000171254 00154 plusmn 00018 00078 plusmn 000071355 00107 plusmn 00018 00054 plusmn 000101634 00746 plusmn 00125 00435 plusmn 000311649 00639 plusmn 00069 00416 plusmn 00032The protein spots show significant differences in their abundance (119875 lt 005)in T indica-treated and nontreated (control) HepG2 cells by more than 15-fold
anti-beta actin (ab8227 rabbit polyclonal Abcam UK) as theloading control using the WesternDot 625 Goat Anti-RabbitWestern Blot kit (Invitrogen Oregon USA) Quantificationof the band intensity was calculated using ImageJ software
3 Results
31 2D-GE Analysis of HepG2 Cell Lysate Proteins Analysisof HepG2 cell lysate by 2D-GE resolved more than 2500protein spots in each gel Figure 1 demonstrates typical 2D-GE resolved patterns of the cell lysate proteins from controlsand HepG2 cells treated with the fruit pulp extract of Tindica The percentage of volume contribution of each spotwas then determined using ImageMaster 2D Platinum V 70software and the fold change if any was acquired Proteinspots that were present in all gels (119899 = 6) and showedsignificant differences in their abundance (119875 lt 005) intreated and nontreated cells by more than 15-fold wereselected Based on these criteria the altered abundance of 20protein spots were detected when HepG2 cells were exposedto the fruit pulp extract of T indica (Table 1)
32 Identification of Altered Abundance HepG2 Cell LysateProteins Among the 20 protein spots that were significantlyreduced in abundance 14 were successfully identified bymass spectrometry and database search (Table 2) Six spots(protein spot ID 398 615 637 1254 1355 and 1649) wereconsidered not successfully identified as their scores werelower than the cut-off value for positive inclusion criteria
4 BioMed Research International
MW
(kD
a)
180120
40
80
50
12
25
20
10
pH 3ndash10 nonlinear
Control Treated
Control
Treated
Figure 1 A representative proteome map of HepG2 cell lysate Approximately 2500 spots per gel within the pH 3ndash10 range for cell lysate ofHepG2 cells were detected Twenty spots were altered in abundance (circled and labelled) and they were all significantly reduced in abundance(119875 lt 005)
The 14 identified proteins may be grouped according totheir biological processes using the UniProt Protein Knowl-edgebase (UniProtKB) NADH dehydrogenase (ubiquinone)1 alpha subcomplex subunit 10 ubiquinol-cytochrome-creductase complex core protein 2 andNADHdehydrogenase(ubiquinone) flavoprotein 1 were grouped under ldquomitochon-drial respiratory chainrdquo category Three proteins that iseukaryotic translation initiation factor 3 subunit 3 elonga-tion factor Tu and tyrosyl-tRNA synthetase are involvedin ldquoprotein synthesisrdquo Another six proteins were catego-rized under ldquometabolismrdquo with glyceraldehyde-3-phosphatedehydrogenase and GDP-L-fucose synthetase being involvedin carbohydrate metabolism GMP reductase 2 and UMPsynthase in nucleotide and nucleosidemetabolism S-methyl-5-thioadenosine phosphorylase in polyamine metabolic pro-cess and ethanolamine phosphate cytidylyltransferase inbiosynthesis of phospholipids Prohibitin on the other handwas categorized under ldquocell proliferation and differentiationrdquowhile heterogeneous nuclear ribonucleoprotein H3 does notbelong to any of the above groups and therefore categorizedas ldquoothersrdquo
33 Biological Processes and Pathway Interaction AnalysisThe cell lysate proteins of altered abundance when analyzedusing Ingenuity Pathways Analysis (IPA) software generateda single network on ldquoHereditary disorder metabolic diseasemolecular transportrdquo with a score of 48 (Table 3) Mitochon-drial dysfunction was ranked with the highest significance(119875 lt 365 times 10minus4) from a canonical pathway analysis When
the data was reanalyzed to include our earlier reported resultsof HepG2 proteins that were differentially secreted whenthe cells were exposed to the T indica fruit pulp extract[7] the software identified ldquoLipid Metabolism MolecularTransport and Small Molecule Biochemistryrdquo as top putativenetwork (score of 31) that links the proteins of alteredabundance with other interactomes and gluconeogenesis Ibecame the top canonical pathway (119875 lt 467times10minus4) (Table 3)Figure 2 shows a graphical representation of the predictedmolecular relationships between HepG2 proteins that werealtered in abundance in response to exposure to T indicafruit pulp extract Other than the lipid-related interactomesthe HepG2 proteins of altered abundance were shown to beassociated with inflammation-related molecules like tumournecrosis factor (TNF) and interleukin-1 beta (IL-1120573) Twotranscription regulators mediator of RNA polymerase IIItranscription subunit 30 (MED30) and transcription factorE2F1 (E2F1) were also shown to be interconnected with theproteins
34 Confirmation of the Effects of T indica Fruit Pulp Extracton HepG2 Proteins To validate the effects of the T indicafruit pulp extract on HepG2 proteins Western blotting wasperformed using antisera raised against the cellular proteinsIn view of the scarce amount of HepG2 cell lysate proteinextract that was generated in this study three proteins thatis ethanolamine phosphate cytidylyltransferase (PCYT2)NADH dehydrogenase (ubiquinone) 1 alpha subcomplex
BioMed Research International 5
Table2Listof
celllysateproteins
ofalteredabun
danceinTindica
fruitextract-tr
eatedcells
identifi
edby
MALD
I-TOFTO
FMSMS
Spot
no
Proteindescrip
tion
Accno
Score
pIM
w(kDa)
Av
FCa
Cov
bMatched
peptide
sequ
ences
Functio
nal
category
657
NADHdehydrogenase(ub
iquino
ne)1
alph
asub
complex
subu
nit10(C
I-42
kD)
O95299
194
8674073
minus18
44117
ndash12213ndash139140ndash
161253ndash261
290ndash
295
Mito
chon
drial
respira
tory
chain
765
Ubiqu
inol-cytochrom
e-cr
eductase
complex
core
protein2mito
chon
drial
precursorc
orep
rotein
IIP2
2695
537
8744841
minus15
2871ndash8
4163ndash183184ndash
196200ndash
217232ndash241
900
NADHdehydrogenase(ub
iquino
ne)
flavoprotein1mito
chon
drialprecursor
(CI-51kD
)P4
9821
275
8515079
minus19
3372ndash81153ndash159160ndash
174376ndash
386
441ndash44
9
796
Ethano
lamine-ph
osph
ate
cytid
ylyltransfe
rase
Q9944
7339
644
4381
minus17
38185ndash199262ndash271333ndash348
Phosph
olipid
biosynthesis
590
Glyceraldehyde-3-ph
osph
ate
dehydrogenase(GAPD
H)
P044
06192
8573603
minus15
18235ndash248310ndash
323
Carbo
hydrate
metabolism
722
GDP-L-fucose
synthetase
Q13630
280
6133587
minus15
4926ndash4
482ndash8890ndash107200ndash214291ndash297
307ndash320
655
GMPredu
ctase2
Q9P
2T1
386
679378
5minus15
5270ndash78178ndash189192ndash213277ndash286
278ndash286292ndash298292ndash306
Nucleotidea
ndnu
cleoside
metabolism
914
Urid
ine51015840
-mon
opho
sphatesynthase
(UMPsynthase)
P1117
2379
6815220
minus16
376ndash
2230ndash
41146
ndash155353ndash363389ndash4
05
460ndash
467469ndash
477
363
S-methyl-5
-thioadenosinep
hospho
rylase
(MTA
P)Q13126
253
675312
3minus15
4312ndash2972ndash8283ndash9
9100ndash
116134ndash
147
181ndash187272ndash282
Polyam
ine
metabolism
1634
Proh
ibitin
P35232
66227297
9minus17
21134ndash
143
Cellproliferation
anddifferentiatio
n
675
Eukaryotictransla
tioninitiationfactor
3subu
nit3
(elF3H
)O15372
279
609399
1minus15
4352ndash75242ndash249260ndash
265304ndash
313
314ndash
331
Protein
biosynthesis
751
Elon
gatio
nfactor
Tum
itochon
drial
precursor(EF
-Tu)
P49411
11472
64951minus18
29105ndash120239ndash
252
1084
Tyrosyl-tRN
Asynthetasecytop
lasm
ic(TyrRS
)P5
4577
96661591
1minus15
2185ndash93179ndash
189433ndash450
540
Heterogenou
snuclear
ribon
ucleop
rotein
H3(hnR
NPH3)
P31942
409
6373690
minus17
3956ndash6
785ndash9
098ndash104
206ndash222223ndash232
262ndash287288ndash301
Others
a Negativev
alue
signifiesd
own-regu
lationin
term
soffold-differencesA
llratio
sare
statisticallysig
nificantw
ith119875lt005(Studentrsquos119905-te
st)
b Coverageo
fthe
identifi
edsequ
ence
6 BioMed Research International
Table 3 Top network and canonical pathways generated by Ingenuity Pathways Analysis
Proteins subjectto analysis
Associated networkfunctions Scoreb Top canonical
pathway 119875 value
Cell lysateproteins
Hereditary disorder metabolicdisease molecular transport 48 Mitochondrial
Dysfunction 365119864 minus 04
Cell lysate andsecretoryproteinsa
Lipid metabolism moleculartransport small moleculeBiochemistry
31 Gluconeogenesis I 467119864 minus 04
aEarlier reported [7]
bA score of 2 or higher indicates at least a 99 confidence of not being generated by random chance and higher scores indicate a greater confidence
subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reduc-tase complex core protein 2 (UQCRC2) that represent thoseinvolved in the mitochondrial and metabolic activities wereselected Densitometry scanning of bands detected by theantisera showed that the abundance of PCYT2 NDUFA10and UQCRC2 was indeed lower in HepG2 cells treated withthe T indica fruit pulp extract compared to the controls withfold differences of ndash17 ndash15 and ndash15 respectively (Figure 3)
4 Discussion
In this study the abundance of 20 cell lysate proteins wasfound to be significantly reduced when HepG2 cells wereexposed to the T indica fruit pulp extract Fourteen of theproteins were identified by mass spectrometry and databasesearch and the reduced abundance of three of the HepG2proteins was subsequently validated by Western blottingDue to limited availability of antibodies and HepG2 celllysate protein extract validation was performed on thethree selective proteins that are representative of separatemitochondrial functions and metabolic activities
Among the identified HepG2 proteins three com-ponents of the mitochondrial respiratory chain namelyubiquinol-cytochrome-c reductase complex core protein 2(UQCRC2) NADH dehydrogenase (ubiquinone) 1 alphasubcomplex subunit 10 (NDUFA10) and NADH dehydroge-nase (ubiquinone) flavoprotein 1 (NDUFV1) were found tobe reduced in abundance when HepG2 cells were exposedto T indica fruit pulp extract UQCRC2 belongs to complexIII of the mitochondrial respiratory chain while NDUFA10andNDUFV1 are components of complex I Inmitochondriaonly complex I [12 13] and complex III [14] of the respiratorychain are known to produce reactive oxygen species (ROS)Hence the decreased amount of the three mitochondrialcomponents may indicate reduced production of free rad-icals although the functionality of the mitochondria is yetto be confirmed Nevertheless earlier studies in obese ratmuscles have shown that reduced levels of respiratory chaincomplex I and diminishedROS production that were inducedby chronic supplementation with grape seed proanthocyani-dins did not affect the function of the mitochondria [15]
Prohibitin is another mitochondrial protein that wasshown to be of reduced abundance in HepG2 cells exposedto T indica fruit pulp extract The protein that was initiallythought to be a negative regulator of cell proliferation has
been demonstrated to play a role in biogenesis and function ofmitochondria [16] In a study of the nematodeCaenorhabditiselegans prohibitin complex was shown to promote longevityby modulating mitochondrial function and fat metabolism[17] Deficiency of prohibitin prolongs the lifespan of Celegans with compromised mitochondrial function or fatmetabolism In this study depletion of prohibitin in HepG2cells upon exposure to the fruit pulp extract of T indicaappears to suggest a similar mechanism in an attempt toextend lifespan of the cells
In addition to the mitochondrial proteins the gly-colytic enzyme glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as well as GDP-L-fucose synthetase (TSTA3) alsoappeared to be down-regulated after treatment with T indicafruit pulp extract Oxidative stress is known to induce up-regulation of GAPDH levels [18 19] Hence the reducedamount of GAPDH in this case may possibly indicate astate of repressed oxidative stress This together with theearlier data on the reduced mitochondrial respiratory chainproteins may shed some light on molecular mechanismsinvolved in the well acclaimed antioxidant properties of Tindica [3 4]
The fruit pulp extract of T indica also appeared tocause decreased abundance of proteins involved in themetabolism of nucleic acids and polyamines in HepG2 cellsTo the best of our knowledge these have not been previ-ously reported and their rationale is not quite understoodOn the other hand decreased abundance of ethanolaminephosphate cytidylyltransferase (PCYT2) the rate-limitingenzymewhich catalyzes conversion of phosphoethanolamineto cytidylylphosphoethanolamine in the biosynthesis ofphosphatidylethanolamine in HepG2 cells exposed to thefruit extract of T indica may compromise availability ofthe phospholipid which stores arachidonic acid for theproduction of prostaglandinsThis could possibly explain theanti-inflammatory action ofT indica thatwas earlier reported[20]
In addition eukaryotic translation initiation factor 3 sub-unit 3 (eIF3H) tyrosyl-tRNA synthetase (YARS) elongationfactor Tu (EFTU) and heterogenous nuclear ribonucleopro-tein H3 (hnRNPH3) which are generally involved in proteinsynthesis and splicing also appeared to be down-regulated inHepG2 cells treated with the T indica extract These proteinsmay be key regulatory points of action of the fruit pulp extractof T indica Suppression of these proteins by the extract may
BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 3
27 Mass Spectrometry and Database Search Trypsin digest-ed peptidemixturewas analyzed using anAppliedBiosystems4800 Plus MALDI-TOFTOF (Foster City CA USA) Thepeptide mixture was crystallized with alpha-cyano-4-hydroxycinnamic acid matrix solution (10mgmL 70 ACNin 01 (vv) TFA aqueous solution) and spotted onto aMALDI target (192-well) plate The MS results were auto-matically acquiredwith a trypsin autodigest exclusion list and20 most intense ions were selected for MSMS analysis Dataanalysis was carried out using the GPS Explorer software(Applied Biosystems CA USA) and database search usingthe in-house MASCOT program (Matrix Science LondonUK) Combined MS and MSMS searches were conductedwith the following settings Swiss-Prot database Homosapiens peptide tolerance at 200 ppm MSMS tolerance at04Da carbamidomethylation of cysteine (variable modifi-cation) and methionine oxidation (variable modifications)A protein is considered identified when a MASCOT scoreof higher than 55 and 119875 lt 005 were obtained from the MSanalysis
28 Functional Analyses Using Ingenuity Pathways Analysis(IPA) Software Functional analyses to predict networks thatare affected by the differentially expressed proteins werecarried out using Ingenuity Pathways Analysis (IPA) software(Ingenuity Systems httpwwwingenuitycom) Details ofthe proteins their quantitative expression values (fold changedifference of at least 15) and 119875 values (119875 lt 005) wereimported into the IPA software Each protein identifierwas mapped to its corresponding protein object and wasoverlaid onto a global molecular network developed frominformation contained in the Ingenuity Knowledge BaseNetwork predictions based on the protein input were gener-ated algorithmically by utilizing the information containedin the Ingenuity Knowledge Base Right-tailed Fischerrsquos exacttest was used to calculate a 119875 value indicating the probabilitythat each biological function assigned to the network is dueto chance alone
29 Validation of Proteins of Reduced Abundance Three ofthe altered proteins ethanolamine phosphate cytidylyltrans-ferase (PCYT2) and NADH dehydrogenase (ubiquinone) 1alpha subcomplex subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reductase complex core protein 2 (UQCRC2)were selected for validation using Western blotting HepG2cells (3 times 106) were treated with 60120583gmL T indica fruit pulpextract for 24 h The cells were then trypsinized and lysedwith RIPA buffer (150mM NaCl 50mM Tris-HCl pH 741mMEDTA 1 Triton X-100 1 sodium deoxycholate 01SDS) Total cell lysate proteins were quantified using BCAassay kit (Pierce Rockford IL USA) Forty micrograms ofcell lysate protein was separated on a 125 SDS-PAGE andtransferred onto a PVDF membrane with 045120583m pore size(Thermo Scientific IL USA) at 100V 110mA for 90 minutesThe blot was then blocked overnight and developed againstanti-PCYT2 (ab126142 rabbit polyclonal Abcam UK) anti-NDUFA10 (ab103026 rabbit polyclonal Abcam UK) anti-UQCRC2 (ab103616 rabbit polyclonal Abcam UK) and
Table 1 Average percentage of volume of spots
Spot ID Average percentage of volume plusmn SEMControl Treated
363 00273 plusmn 00016 00187 plusmn 00019398 00214 plusmn 00021 00134 plusmn 00020540 00128 plusmn 00019 00073 plusmn 00011590 00892 plusmn 00073 00609 plusmn 00039615 00240 plusmn 00023 00157 plusmn 00016637 00254 plusmn 00033 00156 plusmn 00011655 00499 plusmn 00033 00328 plusmn 00056657 00655 plusmn 00088 00360 plusmn 00035675 00660 plusmn 00067 00440 plusmn 00017722 00223 plusmn 00022 00144 plusmn 00014751 00420 plusmn 00038 00235 plusmn 00027765 00470 plusmn 00031 00315 plusmn 00040796 00162 plusmn 00011 00093 plusmn 00009900 00394 plusmn 00048 00206 plusmn 00044914 00279 plusmn 00015 00177 plusmn 000121084 00305 plusmn 00015 00209 plusmn 000171254 00154 plusmn 00018 00078 plusmn 000071355 00107 plusmn 00018 00054 plusmn 000101634 00746 plusmn 00125 00435 plusmn 000311649 00639 plusmn 00069 00416 plusmn 00032The protein spots show significant differences in their abundance (119875 lt 005)in T indica-treated and nontreated (control) HepG2 cells by more than 15-fold
anti-beta actin (ab8227 rabbit polyclonal Abcam UK) as theloading control using the WesternDot 625 Goat Anti-RabbitWestern Blot kit (Invitrogen Oregon USA) Quantificationof the band intensity was calculated using ImageJ software
3 Results
31 2D-GE Analysis of HepG2 Cell Lysate Proteins Analysisof HepG2 cell lysate by 2D-GE resolved more than 2500protein spots in each gel Figure 1 demonstrates typical 2D-GE resolved patterns of the cell lysate proteins from controlsand HepG2 cells treated with the fruit pulp extract of Tindica The percentage of volume contribution of each spotwas then determined using ImageMaster 2D Platinum V 70software and the fold change if any was acquired Proteinspots that were present in all gels (119899 = 6) and showedsignificant differences in their abundance (119875 lt 005) intreated and nontreated cells by more than 15-fold wereselected Based on these criteria the altered abundance of 20protein spots were detected when HepG2 cells were exposedto the fruit pulp extract of T indica (Table 1)
32 Identification of Altered Abundance HepG2 Cell LysateProteins Among the 20 protein spots that were significantlyreduced in abundance 14 were successfully identified bymass spectrometry and database search (Table 2) Six spots(protein spot ID 398 615 637 1254 1355 and 1649) wereconsidered not successfully identified as their scores werelower than the cut-off value for positive inclusion criteria
4 BioMed Research International
MW
(kD
a)
180120
40
80
50
12
25
20
10
pH 3ndash10 nonlinear
Control Treated
Control
Treated
Figure 1 A representative proteome map of HepG2 cell lysate Approximately 2500 spots per gel within the pH 3ndash10 range for cell lysate ofHepG2 cells were detected Twenty spots were altered in abundance (circled and labelled) and they were all significantly reduced in abundance(119875 lt 005)
The 14 identified proteins may be grouped according totheir biological processes using the UniProt Protein Knowl-edgebase (UniProtKB) NADH dehydrogenase (ubiquinone)1 alpha subcomplex subunit 10 ubiquinol-cytochrome-creductase complex core protein 2 andNADHdehydrogenase(ubiquinone) flavoprotein 1 were grouped under ldquomitochon-drial respiratory chainrdquo category Three proteins that iseukaryotic translation initiation factor 3 subunit 3 elonga-tion factor Tu and tyrosyl-tRNA synthetase are involvedin ldquoprotein synthesisrdquo Another six proteins were catego-rized under ldquometabolismrdquo with glyceraldehyde-3-phosphatedehydrogenase and GDP-L-fucose synthetase being involvedin carbohydrate metabolism GMP reductase 2 and UMPsynthase in nucleotide and nucleosidemetabolism S-methyl-5-thioadenosine phosphorylase in polyamine metabolic pro-cess and ethanolamine phosphate cytidylyltransferase inbiosynthesis of phospholipids Prohibitin on the other handwas categorized under ldquocell proliferation and differentiationrdquowhile heterogeneous nuclear ribonucleoprotein H3 does notbelong to any of the above groups and therefore categorizedas ldquoothersrdquo
33 Biological Processes and Pathway Interaction AnalysisThe cell lysate proteins of altered abundance when analyzedusing Ingenuity Pathways Analysis (IPA) software generateda single network on ldquoHereditary disorder metabolic diseasemolecular transportrdquo with a score of 48 (Table 3) Mitochon-drial dysfunction was ranked with the highest significance(119875 lt 365 times 10minus4) from a canonical pathway analysis When
the data was reanalyzed to include our earlier reported resultsof HepG2 proteins that were differentially secreted whenthe cells were exposed to the T indica fruit pulp extract[7] the software identified ldquoLipid Metabolism MolecularTransport and Small Molecule Biochemistryrdquo as top putativenetwork (score of 31) that links the proteins of alteredabundance with other interactomes and gluconeogenesis Ibecame the top canonical pathway (119875 lt 467times10minus4) (Table 3)Figure 2 shows a graphical representation of the predictedmolecular relationships between HepG2 proteins that werealtered in abundance in response to exposure to T indicafruit pulp extract Other than the lipid-related interactomesthe HepG2 proteins of altered abundance were shown to beassociated with inflammation-related molecules like tumournecrosis factor (TNF) and interleukin-1 beta (IL-1120573) Twotranscription regulators mediator of RNA polymerase IIItranscription subunit 30 (MED30) and transcription factorE2F1 (E2F1) were also shown to be interconnected with theproteins
34 Confirmation of the Effects of T indica Fruit Pulp Extracton HepG2 Proteins To validate the effects of the T indicafruit pulp extract on HepG2 proteins Western blotting wasperformed using antisera raised against the cellular proteinsIn view of the scarce amount of HepG2 cell lysate proteinextract that was generated in this study three proteins thatis ethanolamine phosphate cytidylyltransferase (PCYT2)NADH dehydrogenase (ubiquinone) 1 alpha subcomplex
BioMed Research International 5
Table2Listof
celllysateproteins
ofalteredabun
danceinTindica
fruitextract-tr
eatedcells
identifi
edby
MALD
I-TOFTO
FMSMS
Spot
no
Proteindescrip
tion
Accno
Score
pIM
w(kDa)
Av
FCa
Cov
bMatched
peptide
sequ
ences
Functio
nal
category
657
NADHdehydrogenase(ub
iquino
ne)1
alph
asub
complex
subu
nit10(C
I-42
kD)
O95299
194
8674073
minus18
44117
ndash12213ndash139140ndash
161253ndash261
290ndash
295
Mito
chon
drial
respira
tory
chain
765
Ubiqu
inol-cytochrom
e-cr
eductase
complex
core
protein2mito
chon
drial
precursorc
orep
rotein
IIP2
2695
537
8744841
minus15
2871ndash8
4163ndash183184ndash
196200ndash
217232ndash241
900
NADHdehydrogenase(ub
iquino
ne)
flavoprotein1mito
chon
drialprecursor
(CI-51kD
)P4
9821
275
8515079
minus19
3372ndash81153ndash159160ndash
174376ndash
386
441ndash44
9
796
Ethano
lamine-ph
osph
ate
cytid
ylyltransfe
rase
Q9944
7339
644
4381
minus17
38185ndash199262ndash271333ndash348
Phosph
olipid
biosynthesis
590
Glyceraldehyde-3-ph
osph
ate
dehydrogenase(GAPD
H)
P044
06192
8573603
minus15
18235ndash248310ndash
323
Carbo
hydrate
metabolism
722
GDP-L-fucose
synthetase
Q13630
280
6133587
minus15
4926ndash4
482ndash8890ndash107200ndash214291ndash297
307ndash320
655
GMPredu
ctase2
Q9P
2T1
386
679378
5minus15
5270ndash78178ndash189192ndash213277ndash286
278ndash286292ndash298292ndash306
Nucleotidea
ndnu
cleoside
metabolism
914
Urid
ine51015840
-mon
opho
sphatesynthase
(UMPsynthase)
P1117
2379
6815220
minus16
376ndash
2230ndash
41146
ndash155353ndash363389ndash4
05
460ndash
467469ndash
477
363
S-methyl-5
-thioadenosinep
hospho
rylase
(MTA
P)Q13126
253
675312
3minus15
4312ndash2972ndash8283ndash9
9100ndash
116134ndash
147
181ndash187272ndash282
Polyam
ine
metabolism
1634
Proh
ibitin
P35232
66227297
9minus17
21134ndash
143
Cellproliferation
anddifferentiatio
n
675
Eukaryotictransla
tioninitiationfactor
3subu
nit3
(elF3H
)O15372
279
609399
1minus15
4352ndash75242ndash249260ndash
265304ndash
313
314ndash
331
Protein
biosynthesis
751
Elon
gatio
nfactor
Tum
itochon
drial
precursor(EF
-Tu)
P49411
11472
64951minus18
29105ndash120239ndash
252
1084
Tyrosyl-tRN
Asynthetasecytop
lasm
ic(TyrRS
)P5
4577
96661591
1minus15
2185ndash93179ndash
189433ndash450
540
Heterogenou
snuclear
ribon
ucleop
rotein
H3(hnR
NPH3)
P31942
409
6373690
minus17
3956ndash6
785ndash9
098ndash104
206ndash222223ndash232
262ndash287288ndash301
Others
a Negativev
alue
signifiesd
own-regu
lationin
term
soffold-differencesA
llratio
sare
statisticallysig
nificantw
ith119875lt005(Studentrsquos119905-te
st)
b Coverageo
fthe
identifi
edsequ
ence
6 BioMed Research International
Table 3 Top network and canonical pathways generated by Ingenuity Pathways Analysis
Proteins subjectto analysis
Associated networkfunctions Scoreb Top canonical
pathway 119875 value
Cell lysateproteins
Hereditary disorder metabolicdisease molecular transport 48 Mitochondrial
Dysfunction 365119864 minus 04
Cell lysate andsecretoryproteinsa
Lipid metabolism moleculartransport small moleculeBiochemistry
31 Gluconeogenesis I 467119864 minus 04
aEarlier reported [7]
bA score of 2 or higher indicates at least a 99 confidence of not being generated by random chance and higher scores indicate a greater confidence
subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reduc-tase complex core protein 2 (UQCRC2) that represent thoseinvolved in the mitochondrial and metabolic activities wereselected Densitometry scanning of bands detected by theantisera showed that the abundance of PCYT2 NDUFA10and UQCRC2 was indeed lower in HepG2 cells treated withthe T indica fruit pulp extract compared to the controls withfold differences of ndash17 ndash15 and ndash15 respectively (Figure 3)
4 Discussion
In this study the abundance of 20 cell lysate proteins wasfound to be significantly reduced when HepG2 cells wereexposed to the T indica fruit pulp extract Fourteen of theproteins were identified by mass spectrometry and databasesearch and the reduced abundance of three of the HepG2proteins was subsequently validated by Western blottingDue to limited availability of antibodies and HepG2 celllysate protein extract validation was performed on thethree selective proteins that are representative of separatemitochondrial functions and metabolic activities
Among the identified HepG2 proteins three com-ponents of the mitochondrial respiratory chain namelyubiquinol-cytochrome-c reductase complex core protein 2(UQCRC2) NADH dehydrogenase (ubiquinone) 1 alphasubcomplex subunit 10 (NDUFA10) and NADH dehydroge-nase (ubiquinone) flavoprotein 1 (NDUFV1) were found tobe reduced in abundance when HepG2 cells were exposedto T indica fruit pulp extract UQCRC2 belongs to complexIII of the mitochondrial respiratory chain while NDUFA10andNDUFV1 are components of complex I Inmitochondriaonly complex I [12 13] and complex III [14] of the respiratorychain are known to produce reactive oxygen species (ROS)Hence the decreased amount of the three mitochondrialcomponents may indicate reduced production of free rad-icals although the functionality of the mitochondria is yetto be confirmed Nevertheless earlier studies in obese ratmuscles have shown that reduced levels of respiratory chaincomplex I and diminishedROS production that were inducedby chronic supplementation with grape seed proanthocyani-dins did not affect the function of the mitochondria [15]
Prohibitin is another mitochondrial protein that wasshown to be of reduced abundance in HepG2 cells exposedto T indica fruit pulp extract The protein that was initiallythought to be a negative regulator of cell proliferation has
been demonstrated to play a role in biogenesis and function ofmitochondria [16] In a study of the nematodeCaenorhabditiselegans prohibitin complex was shown to promote longevityby modulating mitochondrial function and fat metabolism[17] Deficiency of prohibitin prolongs the lifespan of Celegans with compromised mitochondrial function or fatmetabolism In this study depletion of prohibitin in HepG2cells upon exposure to the fruit pulp extract of T indicaappears to suggest a similar mechanism in an attempt toextend lifespan of the cells
In addition to the mitochondrial proteins the gly-colytic enzyme glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as well as GDP-L-fucose synthetase (TSTA3) alsoappeared to be down-regulated after treatment with T indicafruit pulp extract Oxidative stress is known to induce up-regulation of GAPDH levels [18 19] Hence the reducedamount of GAPDH in this case may possibly indicate astate of repressed oxidative stress This together with theearlier data on the reduced mitochondrial respiratory chainproteins may shed some light on molecular mechanismsinvolved in the well acclaimed antioxidant properties of Tindica [3 4]
The fruit pulp extract of T indica also appeared tocause decreased abundance of proteins involved in themetabolism of nucleic acids and polyamines in HepG2 cellsTo the best of our knowledge these have not been previ-ously reported and their rationale is not quite understoodOn the other hand decreased abundance of ethanolaminephosphate cytidylyltransferase (PCYT2) the rate-limitingenzymewhich catalyzes conversion of phosphoethanolamineto cytidylylphosphoethanolamine in the biosynthesis ofphosphatidylethanolamine in HepG2 cells exposed to thefruit extract of T indica may compromise availability ofthe phospholipid which stores arachidonic acid for theproduction of prostaglandinsThis could possibly explain theanti-inflammatory action ofT indica thatwas earlier reported[20]
In addition eukaryotic translation initiation factor 3 sub-unit 3 (eIF3H) tyrosyl-tRNA synthetase (YARS) elongationfactor Tu (EFTU) and heterogenous nuclear ribonucleopro-tein H3 (hnRNPH3) which are generally involved in proteinsynthesis and splicing also appeared to be down-regulated inHepG2 cells treated with the T indica extract These proteinsmay be key regulatory points of action of the fruit pulp extractof T indica Suppression of these proteins by the extract may
BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

4 BioMed Research International
MW
(kD
a)
180120
40
80
50
12
25
20
10
pH 3ndash10 nonlinear
Control Treated
Control
Treated
Figure 1 A representative proteome map of HepG2 cell lysate Approximately 2500 spots per gel within the pH 3ndash10 range for cell lysate ofHepG2 cells were detected Twenty spots were altered in abundance (circled and labelled) and they were all significantly reduced in abundance(119875 lt 005)
The 14 identified proteins may be grouped according totheir biological processes using the UniProt Protein Knowl-edgebase (UniProtKB) NADH dehydrogenase (ubiquinone)1 alpha subcomplex subunit 10 ubiquinol-cytochrome-creductase complex core protein 2 andNADHdehydrogenase(ubiquinone) flavoprotein 1 were grouped under ldquomitochon-drial respiratory chainrdquo category Three proteins that iseukaryotic translation initiation factor 3 subunit 3 elonga-tion factor Tu and tyrosyl-tRNA synthetase are involvedin ldquoprotein synthesisrdquo Another six proteins were catego-rized under ldquometabolismrdquo with glyceraldehyde-3-phosphatedehydrogenase and GDP-L-fucose synthetase being involvedin carbohydrate metabolism GMP reductase 2 and UMPsynthase in nucleotide and nucleosidemetabolism S-methyl-5-thioadenosine phosphorylase in polyamine metabolic pro-cess and ethanolamine phosphate cytidylyltransferase inbiosynthesis of phospholipids Prohibitin on the other handwas categorized under ldquocell proliferation and differentiationrdquowhile heterogeneous nuclear ribonucleoprotein H3 does notbelong to any of the above groups and therefore categorizedas ldquoothersrdquo
33 Biological Processes and Pathway Interaction AnalysisThe cell lysate proteins of altered abundance when analyzedusing Ingenuity Pathways Analysis (IPA) software generateda single network on ldquoHereditary disorder metabolic diseasemolecular transportrdquo with a score of 48 (Table 3) Mitochon-drial dysfunction was ranked with the highest significance(119875 lt 365 times 10minus4) from a canonical pathway analysis When
the data was reanalyzed to include our earlier reported resultsof HepG2 proteins that were differentially secreted whenthe cells were exposed to the T indica fruit pulp extract[7] the software identified ldquoLipid Metabolism MolecularTransport and Small Molecule Biochemistryrdquo as top putativenetwork (score of 31) that links the proteins of alteredabundance with other interactomes and gluconeogenesis Ibecame the top canonical pathway (119875 lt 467times10minus4) (Table 3)Figure 2 shows a graphical representation of the predictedmolecular relationships between HepG2 proteins that werealtered in abundance in response to exposure to T indicafruit pulp extract Other than the lipid-related interactomesthe HepG2 proteins of altered abundance were shown to beassociated with inflammation-related molecules like tumournecrosis factor (TNF) and interleukin-1 beta (IL-1120573) Twotranscription regulators mediator of RNA polymerase IIItranscription subunit 30 (MED30) and transcription factorE2F1 (E2F1) were also shown to be interconnected with theproteins
34 Confirmation of the Effects of T indica Fruit Pulp Extracton HepG2 Proteins To validate the effects of the T indicafruit pulp extract on HepG2 proteins Western blotting wasperformed using antisera raised against the cellular proteinsIn view of the scarce amount of HepG2 cell lysate proteinextract that was generated in this study three proteins thatis ethanolamine phosphate cytidylyltransferase (PCYT2)NADH dehydrogenase (ubiquinone) 1 alpha subcomplex
BioMed Research International 5
Table2Listof
celllysateproteins
ofalteredabun
danceinTindica
fruitextract-tr
eatedcells
identifi
edby
MALD
I-TOFTO
FMSMS
Spot
no
Proteindescrip
tion
Accno
Score
pIM
w(kDa)
Av
FCa
Cov
bMatched
peptide
sequ
ences
Functio
nal
category
657
NADHdehydrogenase(ub
iquino
ne)1
alph
asub
complex
subu
nit10(C
I-42
kD)
O95299
194
8674073
minus18
44117
ndash12213ndash139140ndash
161253ndash261
290ndash
295
Mito
chon
drial
respira
tory
chain
765
Ubiqu
inol-cytochrom
e-cr
eductase
complex
core
protein2mito
chon
drial
precursorc
orep
rotein
IIP2
2695
537
8744841
minus15
2871ndash8
4163ndash183184ndash
196200ndash
217232ndash241
900
NADHdehydrogenase(ub
iquino
ne)
flavoprotein1mito
chon
drialprecursor
(CI-51kD
)P4
9821
275
8515079
minus19
3372ndash81153ndash159160ndash
174376ndash
386
441ndash44
9
796
Ethano
lamine-ph
osph
ate
cytid
ylyltransfe
rase
Q9944
7339
644
4381
minus17
38185ndash199262ndash271333ndash348
Phosph
olipid
biosynthesis
590
Glyceraldehyde-3-ph
osph
ate
dehydrogenase(GAPD
H)
P044
06192
8573603
minus15
18235ndash248310ndash
323
Carbo
hydrate
metabolism
722
GDP-L-fucose
synthetase
Q13630
280
6133587
minus15
4926ndash4
482ndash8890ndash107200ndash214291ndash297
307ndash320
655
GMPredu
ctase2
Q9P
2T1
386
679378
5minus15
5270ndash78178ndash189192ndash213277ndash286
278ndash286292ndash298292ndash306
Nucleotidea
ndnu
cleoside
metabolism
914
Urid
ine51015840
-mon
opho
sphatesynthase
(UMPsynthase)
P1117
2379
6815220
minus16
376ndash
2230ndash
41146
ndash155353ndash363389ndash4
05
460ndash
467469ndash
477
363
S-methyl-5
-thioadenosinep
hospho
rylase
(MTA
P)Q13126
253
675312
3minus15
4312ndash2972ndash8283ndash9
9100ndash
116134ndash
147
181ndash187272ndash282
Polyam
ine
metabolism
1634
Proh
ibitin
P35232
66227297
9minus17
21134ndash
143
Cellproliferation
anddifferentiatio
n
675
Eukaryotictransla
tioninitiationfactor
3subu
nit3
(elF3H
)O15372
279
609399
1minus15
4352ndash75242ndash249260ndash
265304ndash
313
314ndash
331
Protein
biosynthesis
751
Elon
gatio
nfactor
Tum
itochon
drial
precursor(EF
-Tu)
P49411
11472
64951minus18
29105ndash120239ndash
252
1084
Tyrosyl-tRN
Asynthetasecytop
lasm
ic(TyrRS
)P5
4577
96661591
1minus15
2185ndash93179ndash
189433ndash450
540
Heterogenou
snuclear
ribon
ucleop
rotein
H3(hnR
NPH3)
P31942
409
6373690
minus17
3956ndash6
785ndash9
098ndash104
206ndash222223ndash232
262ndash287288ndash301
Others
a Negativev
alue
signifiesd
own-regu
lationin
term
soffold-differencesA
llratio
sare
statisticallysig
nificantw
ith119875lt005(Studentrsquos119905-te
st)
b Coverageo
fthe
identifi
edsequ
ence
6 BioMed Research International
Table 3 Top network and canonical pathways generated by Ingenuity Pathways Analysis
Proteins subjectto analysis
Associated networkfunctions Scoreb Top canonical
pathway 119875 value
Cell lysateproteins
Hereditary disorder metabolicdisease molecular transport 48 Mitochondrial
Dysfunction 365119864 minus 04
Cell lysate andsecretoryproteinsa
Lipid metabolism moleculartransport small moleculeBiochemistry
31 Gluconeogenesis I 467119864 minus 04
aEarlier reported [7]
bA score of 2 or higher indicates at least a 99 confidence of not being generated by random chance and higher scores indicate a greater confidence
subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reduc-tase complex core protein 2 (UQCRC2) that represent thoseinvolved in the mitochondrial and metabolic activities wereselected Densitometry scanning of bands detected by theantisera showed that the abundance of PCYT2 NDUFA10and UQCRC2 was indeed lower in HepG2 cells treated withthe T indica fruit pulp extract compared to the controls withfold differences of ndash17 ndash15 and ndash15 respectively (Figure 3)
4 Discussion
In this study the abundance of 20 cell lysate proteins wasfound to be significantly reduced when HepG2 cells wereexposed to the T indica fruit pulp extract Fourteen of theproteins were identified by mass spectrometry and databasesearch and the reduced abundance of three of the HepG2proteins was subsequently validated by Western blottingDue to limited availability of antibodies and HepG2 celllysate protein extract validation was performed on thethree selective proteins that are representative of separatemitochondrial functions and metabolic activities
Among the identified HepG2 proteins three com-ponents of the mitochondrial respiratory chain namelyubiquinol-cytochrome-c reductase complex core protein 2(UQCRC2) NADH dehydrogenase (ubiquinone) 1 alphasubcomplex subunit 10 (NDUFA10) and NADH dehydroge-nase (ubiquinone) flavoprotein 1 (NDUFV1) were found tobe reduced in abundance when HepG2 cells were exposedto T indica fruit pulp extract UQCRC2 belongs to complexIII of the mitochondrial respiratory chain while NDUFA10andNDUFV1 are components of complex I Inmitochondriaonly complex I [12 13] and complex III [14] of the respiratorychain are known to produce reactive oxygen species (ROS)Hence the decreased amount of the three mitochondrialcomponents may indicate reduced production of free rad-icals although the functionality of the mitochondria is yetto be confirmed Nevertheless earlier studies in obese ratmuscles have shown that reduced levels of respiratory chaincomplex I and diminishedROS production that were inducedby chronic supplementation with grape seed proanthocyani-dins did not affect the function of the mitochondria [15]
Prohibitin is another mitochondrial protein that wasshown to be of reduced abundance in HepG2 cells exposedto T indica fruit pulp extract The protein that was initiallythought to be a negative regulator of cell proliferation has
been demonstrated to play a role in biogenesis and function ofmitochondria [16] In a study of the nematodeCaenorhabditiselegans prohibitin complex was shown to promote longevityby modulating mitochondrial function and fat metabolism[17] Deficiency of prohibitin prolongs the lifespan of Celegans with compromised mitochondrial function or fatmetabolism In this study depletion of prohibitin in HepG2cells upon exposure to the fruit pulp extract of T indicaappears to suggest a similar mechanism in an attempt toextend lifespan of the cells
In addition to the mitochondrial proteins the gly-colytic enzyme glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as well as GDP-L-fucose synthetase (TSTA3) alsoappeared to be down-regulated after treatment with T indicafruit pulp extract Oxidative stress is known to induce up-regulation of GAPDH levels [18 19] Hence the reducedamount of GAPDH in this case may possibly indicate astate of repressed oxidative stress This together with theearlier data on the reduced mitochondrial respiratory chainproteins may shed some light on molecular mechanismsinvolved in the well acclaimed antioxidant properties of Tindica [3 4]
The fruit pulp extract of T indica also appeared tocause decreased abundance of proteins involved in themetabolism of nucleic acids and polyamines in HepG2 cellsTo the best of our knowledge these have not been previ-ously reported and their rationale is not quite understoodOn the other hand decreased abundance of ethanolaminephosphate cytidylyltransferase (PCYT2) the rate-limitingenzymewhich catalyzes conversion of phosphoethanolamineto cytidylylphosphoethanolamine in the biosynthesis ofphosphatidylethanolamine in HepG2 cells exposed to thefruit extract of T indica may compromise availability ofthe phospholipid which stores arachidonic acid for theproduction of prostaglandinsThis could possibly explain theanti-inflammatory action ofT indica thatwas earlier reported[20]
In addition eukaryotic translation initiation factor 3 sub-unit 3 (eIF3H) tyrosyl-tRNA synthetase (YARS) elongationfactor Tu (EFTU) and heterogenous nuclear ribonucleopro-tein H3 (hnRNPH3) which are generally involved in proteinsynthesis and splicing also appeared to be down-regulated inHepG2 cells treated with the T indica extract These proteinsmay be key regulatory points of action of the fruit pulp extractof T indica Suppression of these proteins by the extract may
BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
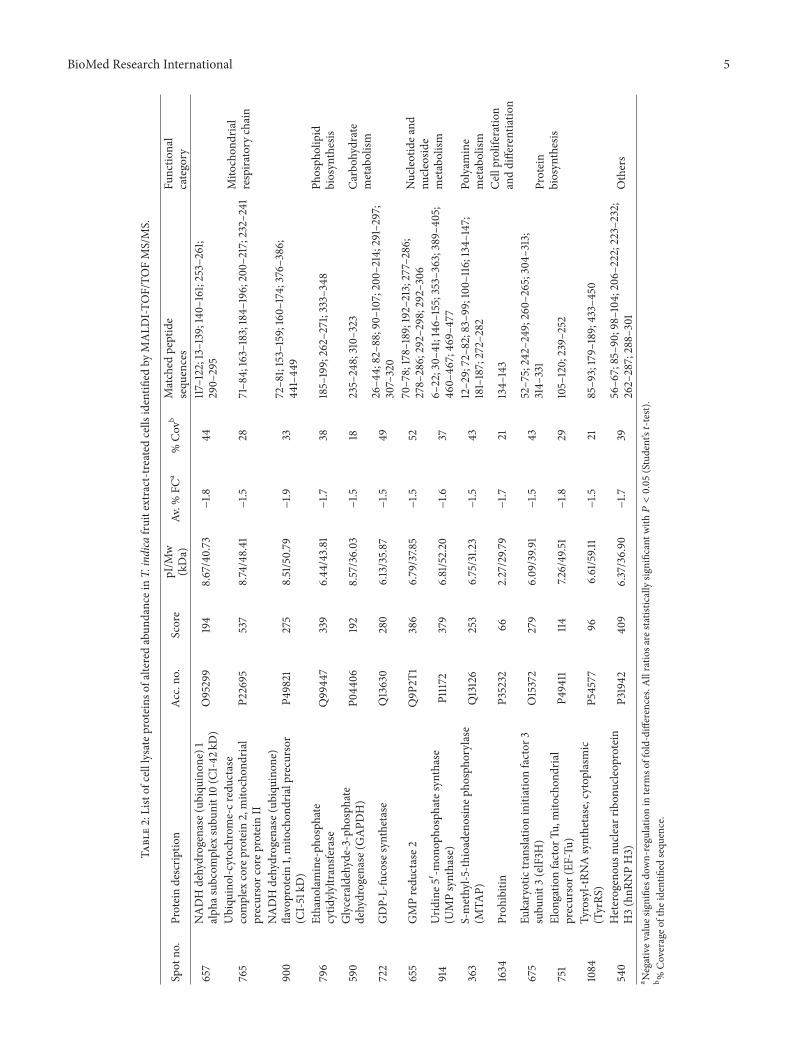
BioMed Research International 5
Table2Listof
celllysateproteins
ofalteredabun
danceinTindica
fruitextract-tr
eatedcells
identifi
edby
MALD
I-TOFTO
FMSMS
Spot
no
Proteindescrip
tion
Accno
Score
pIM
w(kDa)
Av
FCa
Cov
bMatched
peptide
sequ
ences
Functio
nal
category
657
NADHdehydrogenase(ub
iquino
ne)1
alph
asub
complex
subu
nit10(C
I-42
kD)
O95299
194
8674073
minus18
44117
ndash12213ndash139140ndash
161253ndash261
290ndash
295
Mito
chon
drial
respira
tory
chain
765
Ubiqu
inol-cytochrom
e-cr
eductase
complex
core
protein2mito
chon
drial
precursorc
orep
rotein
IIP2
2695
537
8744841
minus15
2871ndash8
4163ndash183184ndash
196200ndash
217232ndash241
900
NADHdehydrogenase(ub
iquino
ne)
flavoprotein1mito
chon
drialprecursor
(CI-51kD
)P4
9821
275
8515079
minus19
3372ndash81153ndash159160ndash
174376ndash
386
441ndash44
9
796
Ethano
lamine-ph
osph
ate
cytid
ylyltransfe
rase
Q9944
7339
644
4381
minus17
38185ndash199262ndash271333ndash348
Phosph
olipid
biosynthesis
590
Glyceraldehyde-3-ph
osph
ate
dehydrogenase(GAPD
H)
P044
06192
8573603
minus15
18235ndash248310ndash
323
Carbo
hydrate
metabolism
722
GDP-L-fucose
synthetase
Q13630
280
6133587
minus15
4926ndash4
482ndash8890ndash107200ndash214291ndash297
307ndash320
655
GMPredu
ctase2
Q9P
2T1
386
679378
5minus15
5270ndash78178ndash189192ndash213277ndash286
278ndash286292ndash298292ndash306
Nucleotidea
ndnu
cleoside
metabolism
914
Urid
ine51015840
-mon
opho
sphatesynthase
(UMPsynthase)
P1117
2379
6815220
minus16
376ndash
2230ndash
41146
ndash155353ndash363389ndash4
05
460ndash
467469ndash
477
363
S-methyl-5
-thioadenosinep
hospho
rylase
(MTA
P)Q13126
253
675312
3minus15
4312ndash2972ndash8283ndash9
9100ndash
116134ndash
147
181ndash187272ndash282
Polyam
ine
metabolism
1634
Proh
ibitin
P35232
66227297
9minus17
21134ndash
143
Cellproliferation
anddifferentiatio
n
675
Eukaryotictransla
tioninitiationfactor
3subu
nit3
(elF3H
)O15372
279
609399
1minus15
4352ndash75242ndash249260ndash
265304ndash
313
314ndash
331
Protein
biosynthesis
751
Elon
gatio
nfactor
Tum
itochon
drial
precursor(EF
-Tu)
P49411
11472
64951minus18
29105ndash120239ndash
252
1084
Tyrosyl-tRN
Asynthetasecytop
lasm
ic(TyrRS
)P5
4577
96661591
1minus15
2185ndash93179ndash
189433ndash450
540
Heterogenou
snuclear
ribon
ucleop
rotein
H3(hnR
NPH3)
P31942
409
6373690
minus17
3956ndash6
785ndash9
098ndash104
206ndash222223ndash232
262ndash287288ndash301
Others
a Negativev
alue
signifiesd
own-regu
lationin
term
soffold-differencesA
llratio
sare
statisticallysig
nificantw
ith119875lt005(Studentrsquos119905-te
st)
b Coverageo
fthe
identifi
edsequ
ence
6 BioMed Research International
Table 3 Top network and canonical pathways generated by Ingenuity Pathways Analysis
Proteins subjectto analysis
Associated networkfunctions Scoreb Top canonical
pathway 119875 value
Cell lysateproteins
Hereditary disorder metabolicdisease molecular transport 48 Mitochondrial
Dysfunction 365119864 minus 04
Cell lysate andsecretoryproteinsa
Lipid metabolism moleculartransport small moleculeBiochemistry
31 Gluconeogenesis I 467119864 minus 04
aEarlier reported [7]
bA score of 2 or higher indicates at least a 99 confidence of not being generated by random chance and higher scores indicate a greater confidence
subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reduc-tase complex core protein 2 (UQCRC2) that represent thoseinvolved in the mitochondrial and metabolic activities wereselected Densitometry scanning of bands detected by theantisera showed that the abundance of PCYT2 NDUFA10and UQCRC2 was indeed lower in HepG2 cells treated withthe T indica fruit pulp extract compared to the controls withfold differences of ndash17 ndash15 and ndash15 respectively (Figure 3)
4 Discussion
In this study the abundance of 20 cell lysate proteins wasfound to be significantly reduced when HepG2 cells wereexposed to the T indica fruit pulp extract Fourteen of theproteins were identified by mass spectrometry and databasesearch and the reduced abundance of three of the HepG2proteins was subsequently validated by Western blottingDue to limited availability of antibodies and HepG2 celllysate protein extract validation was performed on thethree selective proteins that are representative of separatemitochondrial functions and metabolic activities
Among the identified HepG2 proteins three com-ponents of the mitochondrial respiratory chain namelyubiquinol-cytochrome-c reductase complex core protein 2(UQCRC2) NADH dehydrogenase (ubiquinone) 1 alphasubcomplex subunit 10 (NDUFA10) and NADH dehydroge-nase (ubiquinone) flavoprotein 1 (NDUFV1) were found tobe reduced in abundance when HepG2 cells were exposedto T indica fruit pulp extract UQCRC2 belongs to complexIII of the mitochondrial respiratory chain while NDUFA10andNDUFV1 are components of complex I Inmitochondriaonly complex I [12 13] and complex III [14] of the respiratorychain are known to produce reactive oxygen species (ROS)Hence the decreased amount of the three mitochondrialcomponents may indicate reduced production of free rad-icals although the functionality of the mitochondria is yetto be confirmed Nevertheless earlier studies in obese ratmuscles have shown that reduced levels of respiratory chaincomplex I and diminishedROS production that were inducedby chronic supplementation with grape seed proanthocyani-dins did not affect the function of the mitochondria [15]
Prohibitin is another mitochondrial protein that wasshown to be of reduced abundance in HepG2 cells exposedto T indica fruit pulp extract The protein that was initiallythought to be a negative regulator of cell proliferation has
been demonstrated to play a role in biogenesis and function ofmitochondria [16] In a study of the nematodeCaenorhabditiselegans prohibitin complex was shown to promote longevityby modulating mitochondrial function and fat metabolism[17] Deficiency of prohibitin prolongs the lifespan of Celegans with compromised mitochondrial function or fatmetabolism In this study depletion of prohibitin in HepG2cells upon exposure to the fruit pulp extract of T indicaappears to suggest a similar mechanism in an attempt toextend lifespan of the cells
In addition to the mitochondrial proteins the gly-colytic enzyme glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as well as GDP-L-fucose synthetase (TSTA3) alsoappeared to be down-regulated after treatment with T indicafruit pulp extract Oxidative stress is known to induce up-regulation of GAPDH levels [18 19] Hence the reducedamount of GAPDH in this case may possibly indicate astate of repressed oxidative stress This together with theearlier data on the reduced mitochondrial respiratory chainproteins may shed some light on molecular mechanismsinvolved in the well acclaimed antioxidant properties of Tindica [3 4]
The fruit pulp extract of T indica also appeared tocause decreased abundance of proteins involved in themetabolism of nucleic acids and polyamines in HepG2 cellsTo the best of our knowledge these have not been previ-ously reported and their rationale is not quite understoodOn the other hand decreased abundance of ethanolaminephosphate cytidylyltransferase (PCYT2) the rate-limitingenzymewhich catalyzes conversion of phosphoethanolamineto cytidylylphosphoethanolamine in the biosynthesis ofphosphatidylethanolamine in HepG2 cells exposed to thefruit extract of T indica may compromise availability ofthe phospholipid which stores arachidonic acid for theproduction of prostaglandinsThis could possibly explain theanti-inflammatory action ofT indica thatwas earlier reported[20]
In addition eukaryotic translation initiation factor 3 sub-unit 3 (eIF3H) tyrosyl-tRNA synthetase (YARS) elongationfactor Tu (EFTU) and heterogenous nuclear ribonucleopro-tein H3 (hnRNPH3) which are generally involved in proteinsynthesis and splicing also appeared to be down-regulated inHepG2 cells treated with the T indica extract These proteinsmay be key regulatory points of action of the fruit pulp extractof T indica Suppression of these proteins by the extract may
BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

6 BioMed Research International
Table 3 Top network and canonical pathways generated by Ingenuity Pathways Analysis
Proteins subjectto analysis
Associated networkfunctions Scoreb Top canonical
pathway 119875 value
Cell lysateproteins
Hereditary disorder metabolicdisease molecular transport 48 Mitochondrial
Dysfunction 365119864 minus 04
Cell lysate andsecretoryproteinsa
Lipid metabolism moleculartransport small moleculeBiochemistry
31 Gluconeogenesis I 467119864 minus 04
aEarlier reported [7]
bA score of 2 or higher indicates at least a 99 confidence of not being generated by random chance and higher scores indicate a greater confidence
subunit 10 (NDUFA10) and ubiquinol-cytochrome-c reduc-tase complex core protein 2 (UQCRC2) that represent thoseinvolved in the mitochondrial and metabolic activities wereselected Densitometry scanning of bands detected by theantisera showed that the abundance of PCYT2 NDUFA10and UQCRC2 was indeed lower in HepG2 cells treated withthe T indica fruit pulp extract compared to the controls withfold differences of ndash17 ndash15 and ndash15 respectively (Figure 3)
4 Discussion
In this study the abundance of 20 cell lysate proteins wasfound to be significantly reduced when HepG2 cells wereexposed to the T indica fruit pulp extract Fourteen of theproteins were identified by mass spectrometry and databasesearch and the reduced abundance of three of the HepG2proteins was subsequently validated by Western blottingDue to limited availability of antibodies and HepG2 celllysate protein extract validation was performed on thethree selective proteins that are representative of separatemitochondrial functions and metabolic activities
Among the identified HepG2 proteins three com-ponents of the mitochondrial respiratory chain namelyubiquinol-cytochrome-c reductase complex core protein 2(UQCRC2) NADH dehydrogenase (ubiquinone) 1 alphasubcomplex subunit 10 (NDUFA10) and NADH dehydroge-nase (ubiquinone) flavoprotein 1 (NDUFV1) were found tobe reduced in abundance when HepG2 cells were exposedto T indica fruit pulp extract UQCRC2 belongs to complexIII of the mitochondrial respiratory chain while NDUFA10andNDUFV1 are components of complex I Inmitochondriaonly complex I [12 13] and complex III [14] of the respiratorychain are known to produce reactive oxygen species (ROS)Hence the decreased amount of the three mitochondrialcomponents may indicate reduced production of free rad-icals although the functionality of the mitochondria is yetto be confirmed Nevertheless earlier studies in obese ratmuscles have shown that reduced levels of respiratory chaincomplex I and diminishedROS production that were inducedby chronic supplementation with grape seed proanthocyani-dins did not affect the function of the mitochondria [15]
Prohibitin is another mitochondrial protein that wasshown to be of reduced abundance in HepG2 cells exposedto T indica fruit pulp extract The protein that was initiallythought to be a negative regulator of cell proliferation has
been demonstrated to play a role in biogenesis and function ofmitochondria [16] In a study of the nematodeCaenorhabditiselegans prohibitin complex was shown to promote longevityby modulating mitochondrial function and fat metabolism[17] Deficiency of prohibitin prolongs the lifespan of Celegans with compromised mitochondrial function or fatmetabolism In this study depletion of prohibitin in HepG2cells upon exposure to the fruit pulp extract of T indicaappears to suggest a similar mechanism in an attempt toextend lifespan of the cells
In addition to the mitochondrial proteins the gly-colytic enzyme glyceraldehyde 3-phosphate dehydrogenase(GAPDH) as well as GDP-L-fucose synthetase (TSTA3) alsoappeared to be down-regulated after treatment with T indicafruit pulp extract Oxidative stress is known to induce up-regulation of GAPDH levels [18 19] Hence the reducedamount of GAPDH in this case may possibly indicate astate of repressed oxidative stress This together with theearlier data on the reduced mitochondrial respiratory chainproteins may shed some light on molecular mechanismsinvolved in the well acclaimed antioxidant properties of Tindica [3 4]
The fruit pulp extract of T indica also appeared tocause decreased abundance of proteins involved in themetabolism of nucleic acids and polyamines in HepG2 cellsTo the best of our knowledge these have not been previ-ously reported and their rationale is not quite understoodOn the other hand decreased abundance of ethanolaminephosphate cytidylyltransferase (PCYT2) the rate-limitingenzymewhich catalyzes conversion of phosphoethanolamineto cytidylylphosphoethanolamine in the biosynthesis ofphosphatidylethanolamine in HepG2 cells exposed to thefruit extract of T indica may compromise availability ofthe phospholipid which stores arachidonic acid for theproduction of prostaglandinsThis could possibly explain theanti-inflammatory action ofT indica thatwas earlier reported[20]
In addition eukaryotic translation initiation factor 3 sub-unit 3 (eIF3H) tyrosyl-tRNA synthetase (YARS) elongationfactor Tu (EFTU) and heterogenous nuclear ribonucleopro-tein H3 (hnRNPH3) which are generally involved in proteinsynthesis and splicing also appeared to be down-regulated inHepG2 cells treated with the T indica extract These proteinsmay be key regulatory points of action of the fruit pulp extractof T indica Suppression of these proteins by the extract may
BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 7
Figure 2 IPA graphical representation of the molecular rela-tionships between HepG2 secreted and cytosolic proteins aftertreatment The network is displayed graphically as nodes (proteins)and edges (the biological relationships between the nodes) Nodesin red indicate up-regulated proteins while those in green representdown-regulated proteins Nodes without colors indicate unalteredexpression Various shapes of the nodes represent functional classof the proteins Edges are displayed with various labels that describethe nature of the relationship between the nodes TransthyretinTTR thyroglobulin TG interleukin-1 beta IL1B tumour necro-sis factor TNF apolipoprotein A-1 APOA1 apolipoprotein C-1 APOC1 lecithin cholesterol acyltransferase LCAT endothe-lial lipase LIPG haptoglobin HP phospholipase A2 PLA2G2Acholesterylester transfer protein CETP ATP-binding cassette trans-porter sub-family C member 9 ABCC9 ATP-sensitive inwardrectifier potassium channel 11 KCNJ11 ectonucleotide pyrophos-phatasephosphodiesterase family member 1 ENPP1 amyloid pre-cursor protein APP glyceraldehyde-3-phosphate dehydrogenaseGAPDH alpha enolase ENO1 adenylate kinase AK1 ubiquinol-cytochrome-c reductase complex core protein 2UQCRC2 xanthinedehydrogenase XDH cystathionine beta-synthase CBS methio-nine adenosyltransferase I alpha MAT1A rab GDP dissociationinhibitor beta GDI2 NLR (nucleotide-binding domain and leucinerich repeat containing family) family pyrin domain containing3 NLRP3 Vacuolar ATP synthase subunit E ATP6V1E1 elonga-tion factor Tu TUFM prohibitin PHB choline-phosphate cytidy-lyltransferase A PCYT1A uridine 51015840-monophosphate synthaseUMPS NADH dehydrogenase (ubiquinone) 1 alpha subcomplexsubunit 10 NDUFA10 NADH dehydrogenase (ubiquinone) flavo-protein 1 NDUFV1 mediator of RNA polymerase III transcriptionsubunit 30 MED30 transcription factor E2F1 E2F1 suppressor ofG2 allele of SKP1 homolog SUGT1
reflect the underlying epigenetic mechanism that ultimatelycaused the reduced expression of all the mitochondrial andmetabolic proteins and enzymes
Subjecting the altered abundance proteins to IPA anal-ysis generated a single network on ldquoHereditary disor-der metabolic disease molecular transportrdquo which rankedmitochondrial dysfunction with the highest significance
(119875 lt 365 times 10minus4) However ldquoLipid Metabolism MolecularTransport Small Molecule Biochemistryrdquo became the topnetwork involved when IPA was reanalyzed to include pro-teins that were previously shown to be differentially secretedby HepG2 cells treated with the same fruit extract [7] Thisnetwork was not generated in the earlier analysis as PCYT2was the sole cellular protein involved in lipidmetabolism thatwas affected whenHepG2 cells were exposed to T indica fruitpulp extract In our earlier IPA analysis of secreted proteins ofaltered abundance fromHepG2 exposed to the T indica fruitpulp extract a score of 9 was obtained [7] and this improvedto 31 when the data were reanalyzed to include cell lysateproteins of reduced abundance signifying markedly higherprobability In addition the IPA software also identifiedtumour necrosis factor (TNF) and interleukin-1 beta (IL-1120573)both of which are potent inflammatory mediators as inter-actomes in the network affected by T indica (Figure 2) Thisfurther supports our earlier speculation on the molecularmechanism involved in anti-inflammatory effects ofT indica
The lipid-lowering effects induced by plant polyphenolshave been reported by many In fact there is an upcomingtrend of research revealing the potentials of plant polyphenolsin regulating in vitro and in vivo metabolic processes Themethanolic extract of T indica fruit pulp contains proan-thocyanidins which constitutes more than 73 of its totalphenolic content [4] Proanthocyanidinmodulates activationof LXRs [9] which are oxysterol-activated nuclear receptorsthat control cholesterol homeostasis bymodifying expressionof genes involved in cholesterol absorption and efflux fromperipheral tissues This process is mediated through ABCA-1-mediated cholesterol efflux andABCG58-mediated choles-terol excretion and absorption Interestingly LXRs also reg-ulate genes essentially involved in inflammation (Figure 2)As speculated earlier the anti-inflammatory action of Tindica fruit pulpmay possibly occur by inhibition of synthesisof phospholipids and hence depleting the arachidonic acidreservoir for the generation of prostanoid inflammatorymediators
5 Conclusion
The methanol fruit pulp extract of T indica was shownto cause reduced abundance of HepG2 mitochondrialmetabolic and regulatory proteins involved in oxidativephosphorylation protein synthesis and cellular metabolismThe cellular proteins when taken together with the earlierHepG2 secreted proteins of altered abundance are suggestiveof the effects of the fruit pulp extract of T indica oninflammation and lipid metabolism which are modulated byLXRs
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
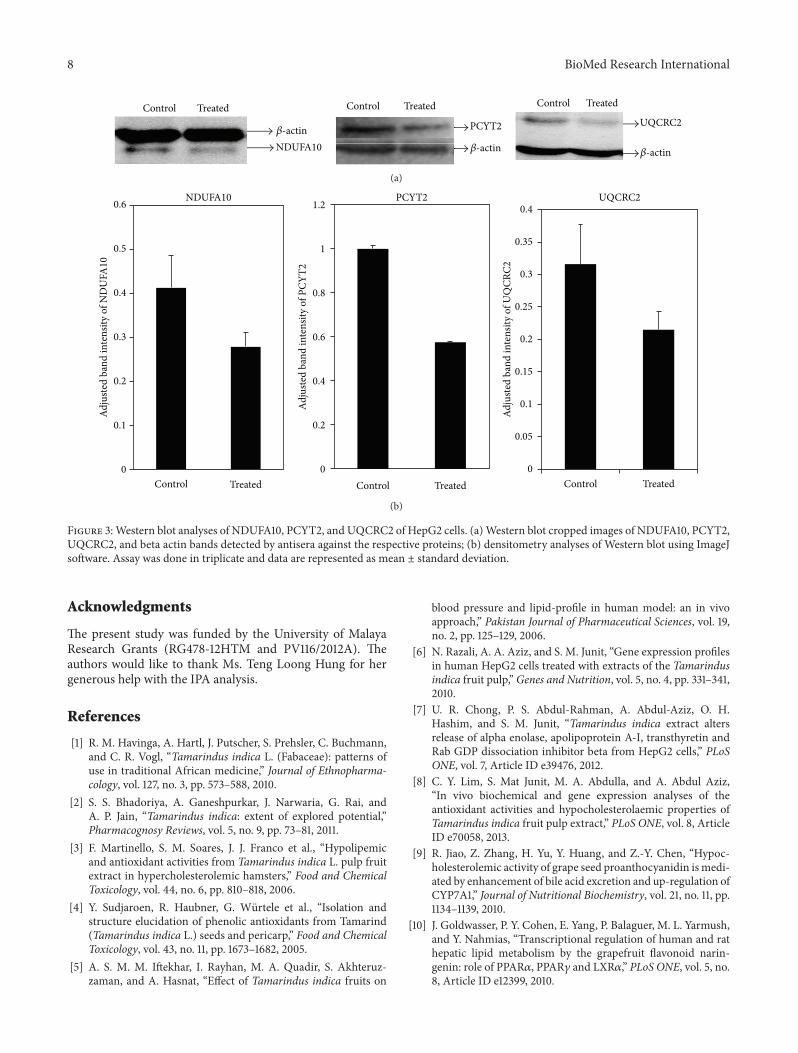
8 BioMed Research International
UQCRC2
Control Treated
120573-actin
120573-actin120573-actinNDUFA10
PCYT2Control ControlTreated Treated
(a)
0
01
02
03
04
05
06
Adju
sted
band
inte
nsity
of N
DU
FA10
NDUFA10
TreatedControl0
02
04
06
08
1
12
Adju
sted
band
inte
nsity
of P
CYT2
PCYT2
Control Treated0
005
01
015
02
025
03
035
04
Control Treated
Adju
sted
band
inte
nsity
of U
QCR
C2
UQCRC2
(b)
Figure 3Western blot analyses of NDUFA10 PCYT2 and UQCRC2 of HepG2 cells (a)Western blot cropped images of NDUFA10 PCYT2UQCRC2 and beta actin bands detected by antisera against the respective proteins (b) densitometry analyses of Western blot using ImageJsoftware Assay was done in triplicate and data are represented as mean plusmn standard deviation
Acknowledgments
The present study was funded by the University of MalayaResearch Grants (RG478-12HTM and PV1162012A) Theauthors would like to thank Ms Teng Loong Hung for hergenerous help with the IPA analysis
References
[1] R M Havinga A Hartl J Putscher S Prehsler C Buchmannand C R Vogl ldquoTamarindus indica L (Fabaceae) patterns ofuse in traditional African medicinerdquo Journal of Ethnopharma-cology vol 127 no 3 pp 573ndash588 2010
[2] S S Bhadoriya A Ganeshpurkar J Narwaria G Rai andA P Jain ldquoTamarindus indica extent of explored potentialrdquoPharmacognosy Reviews vol 5 no 9 pp 73ndash81 2011
[3] F Martinello S M Soares J J Franco et al ldquoHypolipemicand antioxidant activities from Tamarindus indica L pulp fruitextract in hypercholesterolemic hamstersrdquo Food and ChemicalToxicology vol 44 no 6 pp 810ndash818 2006
[4] Y Sudjaroen R Haubner G Wurtele et al ldquoIsolation andstructure elucidation of phenolic antioxidants from Tamarind(Tamarindus indica L) seeds and pericarprdquo Food and ChemicalToxicology vol 43 no 11 pp 1673ndash1682 2005
[5] A S M M Iftekhar I Rayhan M A Quadir S Akhteruz-zaman and A Hasnat ldquoEffect of Tamarindus indica fruits on
blood pressure and lipid-profile in human model an in vivoapproachrdquo Pakistan Journal of Pharmaceutical Sciences vol 19no 2 pp 125ndash129 2006
[6] N Razali A A Aziz and S M Junit ldquoGene expression profilesin human HepG2 cells treated with extracts of the Tamarindusindica fruit pulprdquoGenes and Nutrition vol 5 no 4 pp 331ndash3412010
[7] U R Chong P S Abdul-Rahman A Abdul-Aziz O HHashim and S M Junit ldquoTamarindus indica extract altersrelease of alpha enolase apolipoprotein A-I transthyretin andRab GDP dissociation inhibitor beta from HepG2 cellsrdquo PLoSONE vol 7 Article ID e39476 2012
[8] C Y Lim S Mat Junit M A Abdulla and A Abdul AzizldquoIn vivo biochemical and gene expression analyses of theantioxidant activities and hypocholesterolaemic properties ofTamarindus indica fruit pulp extractrdquo PLoS ONE vol 8 ArticleID e70058 2013
[9] R Jiao Z Zhang H Yu Y Huang and Z-Y Chen ldquoHypoc-holesterolemic activity of grape seed proanthocyanidin ismedi-ated by enhancement of bile acid excretion and up-regulation ofCYP7A1rdquo Journal of Nutritional Biochemistry vol 21 no 11 pp1134ndash1139 2010
[10] J Goldwasser P Y Cohen E Yang P Balaguer M L Yarmushand Y Nahmias ldquoTranscriptional regulation of human and rathepatic lipid metabolism by the grapefruit flavonoid narin-genin role of PPAR120572 PPAR120574 and LXR120572rdquo PLoS ONE vol 5 no8 Article ID e12399 2010
BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 9
[11] K Lee ldquoTransactivation of peroxisome proliferator-activatedreceptor alpha by green tea extractsrdquo Journal of VeterinaryScience vol 5 no 4 pp 325ndash330 2004
[12] G Barja and A Herrero ldquoLocalization at complex I andmechanism of the higher free radical production of brainnonsynaptic mitochondria in the short-lived rat than in thelongevous pigeonrdquo Journal of Bioenergetics and Biomembranesvol 30 no 3 pp 235ndash243 1998
[13] M L Genova B Ventura G Giuliano et al ldquoThe site ofproduction of superoxide radical inmitochondrial Complex I isnot a bound ubisemiquinone but presumably iron-sulfur clusterN2rdquo FEBS Letters vol 505 no 3 pp 364ndash368 2001
[14] A Boveris E Cadenas and A O M Stoppani ldquoRole ofubiquinone in the mitochondrial generation of hydrogen per-oxiderdquo Biochemical Journal vol 156 no 2 pp 435ndash444 1976
[15] D Pajuelo A Fernandez-Iglesias S Dıaz et al ldquoImprovementof mitochondrial function in muscle of genetically obese ratsafter chronic supplementation with proanthocyanidinsrdquo Jour-nal of Agricultural and Food Chemistry vol 59 no 15 pp 8491ndash8498 2011
[16] M Artal-Sanz and N Tavernarakis ldquoProhibitin and mitochon-drial biologyrdquo Trends in Endocrinology and Metabolism vol 20no 8 pp 394ndash401 2009
[17] M Artal-Sanz and N Tavernarakis ldquoProhibitin couples dia-pause signalling to mitochondrial metabolism during ageing inC elegansrdquo Nature vol 461 no 7265 pp 793ndash797 2009
[18] Y Ito P J Pagano K Tornheim P Brecher and R ACohen ldquoOxidative stress increases glyceraldehyde-3-phosphatedehydrogenase mRNA levels in isolated rabbit aortardquoAmericanJournal of Physiology Heart and Circulatory Physiology vol 270no 1 pp H81ndashH87 1996
[19] F deMarco E Bucaj C Foppoli et al ldquoOxidative stress inHPV-driven viral carcinogenesis redox proteomics analysis of HPV-16 dysplastic and neoplastic tissuesrdquo PLoS ONE vol 7 no 3Article ID e34366 2012
[20] V Rimbau C Cerdan R Vila and J Iglesias ldquoAntiinflamma-tory activity of some extracts from plants used in the tradi-tional medicine of north-African countries (II)rdquo PhytotherapyResearch vol 13 pp 128ndash132 1999
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of AddictionHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AnesthesiologyResearch and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Copyright © 2022 FDOKUMEN




















