
Effects of oriC relocation on control of replication initiation in Bacillus subtilis
13
Downloaded from www.microbiologyresearch.org by IP: 54.144.197.59 On: Fri, 22 Apr 2016 07:23:30 Effects of oriC relocation on control of replication initiation in Bacillus subtilis Shigeki Moriya, 1 Yoshikazu Kawai, 2 Sakiko Kaji, 3 Adrian Smith, 4 Elizabeth J. Harry 1 and Jeffery Errington 2 Correspondence Shigeki Moriya [email protected] 1 Institute for the Biotechnology of Infectious Diseases, University of Technology Sydney, PO Box 123, Broadway, NSW 2007, Australia 2 Institute for Cell and Molecular Biosciences, University of Newcastle, Framlington Place, Newcastle NE2 4HH, UK 3 Graduate School of Biological Sciences, Nara Institute of Science and Technology, 8916-5 Takayama, Ikoma, Nara 630-0192, Japan 4 Centenary Institute of Cancer Medicine and Cell Biology, University of Sydney, Locked Bag No. 6, Newtown, NSW 2042, Australia Received 22 April 2009 Revised 21 June 2009 Accepted 25 June 2009 In bacteria, DNA replication initiation is tightly regulated in order to coordinate chromosome replication with cell growth. In Escherichia coli, positive factors and negative regulatory mechanisms playing important roles in the strict control of DNA replication initiation have been reported. However, it remains unclear how bacterial cells recognize the right time for replication initiation during the cell cycle. In the Gram-positive bacterium Bacillus subtilis, much less is known about the regulation of replication initiation, specifically, regarding negative control mechanisms which ensure replication initiation only once per cell cycle. Here we report that replication initiation was greatly enhanced in strains that had the origin of replication (oriC) relocated to various loci on the chromosome. When oriC was relocated to new loci further than 250 kb counterclockwise from the native locus, replication initiation became asynchronous and earlier than in the wild-type cells. In two oriC-relocated strains (oriC at argG or pnbA, 2576 or 3006 on the 3606 chromosome map, respectively), DnaA levels were higher than in the wild-type but not enough to cause earlier initiation of replication. Our results suggest that the initiation capacity of replication is accumulated well before the actual time of initiation, and its release may be suppressed by a unique DNA structure formed near the native oriC locus. INTRODUCTION Replication of genomic DNA is indispensable for cell growth. In bacteria, replication begins at a fixed site (oriC) within the genome and proceeds bidirectionally. In order to coordinate replication with cell growth, the most important control is carried out at the initiation stage (von Meyenburg & Hansen, 1987). In fast-growing cells (generation time ,60 min), the time required for replication and cell division is longer than the generation time. To compensate for this potentially serious problem, cells start a new round of replication before previous rounds terminate in a process known as dichotomous replication (Yoshikawa & Wake, 1993). It is unclear how cells precisely recognize the right stage in the cell cycle to initiate replication. In Escherichia coli, several positive and negative factors appear to participate in the regulation of replication initiation. DnaA protein level is a positive factor, as increased levels of DnaA cause overinitiation and advance the initiation event to an earlier time in the cell cycle (Atlung & Hansen, 1993; Løbner-Olesen et al., 1989). The level of the ATP-bound form of DnaA, which is the active form for replication initiation, correlates with this property (Kurokawa et al., 1999). A recent finding has indicated that DiaA tetramers stimulate assembly of active ATP–DnaA– oriC initiation complex by directly binding to multiple DnaA molecules (Keyamura et al., 2007). Three negative mechanisms are known to inhibit extra initiations of DNA replication. The first one is sequestration of newly duplicated hemimethylated origins into the cell membrane by SeqA protein (Campbell & Kleckner, 1990; Lu et al., 1994; Ogden et al., 1988). Two others are inactivation of DnaA initiation protein by Hda and DnaN (b subunit of DNA polymerase III) (Katayama et al., 1998; Kato & Katayama, 2001) and sequestration of DnaA at the datA Abbreviations: DAPI, 49,6-diamidino-2-phenylindole; PLR, polar local- ization region. A supplementary table, listing nucleotide sequences of primers used in this study, is available with the online version of this paper. Microbiology (2009), 155, 3070–3082 DOI 10.1099/mic.0.030080-0 3070 030080 G 2009 SGM Printed in Great Britain
Transcript of Effects of oriC relocation on control of replication initiation in Bacillus subtilis

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
Effects of oriC relocation on control of replicationinitiation in Bacillus subtilis
Shigeki Moriya,1 Yoshikazu Kawai,2 Sakiko Kaji,3 Adrian Smith,4
Elizabeth J. Harry1 and Jeffery Errington2
Correspondence
Shigeki Moriya
1Institute for the Biotechnology of Infectious Diseases, University of Technology Sydney,PO Box 123, Broadway, NSW 2007, Australia
2Institute for Cell and Molecular Biosciences, University of Newcastle, Framlington Place,Newcastle NE2 4HH, UK
3Graduate School of Biological Sciences, Nara Institute of Science and Technology, 8916-5Takayama, Ikoma, Nara 630-0192, Japan
4Centenary Institute of Cancer Medicine and Cell Biology, University of Sydney, Locked Bag No. 6,Newtown, NSW 2042, Australia
Received 22 April 2009
Revised 21 June 2009
Accepted 25 June 2009
In bacteria, DNA replication initiation is tightly regulated in order to coordinate chromosome
replication with cell growth. In Escherichia coli, positive factors and negative regulatory
mechanisms playing important roles in the strict control of DNA replication initiation have been
reported. However, it remains unclear how bacterial cells recognize the right time for replication
initiation during the cell cycle. In the Gram-positive bacterium Bacillus subtilis, much less is
known about the regulation of replication initiation, specifically, regarding negative control
mechanisms which ensure replication initiation only once per cell cycle. Here we report that
replication initiation was greatly enhanced in strains that had the origin of replication (oriC)
relocated to various loci on the chromosome. When oriC was relocated to new loci further than
250 kb counterclockwise from the native locus, replication initiation became asynchronous and
earlier than in the wild-type cells. In two oriC-relocated strains (oriC at argG or pnbA, 2576 or
3006 on the 3606 chromosome map, respectively), DnaA levels were higher than in the wild-type
but not enough to cause earlier initiation of replication. Our results suggest that the initiation
capacity of replication is accumulated well before the actual time of initiation, and its release may
be suppressed by a unique DNA structure formed near the native oriC locus.
INTRODUCTION
Replication of genomic DNA is indispensable for cellgrowth. In bacteria, replication begins at a fixed site (oriC)within the genome and proceeds bidirectionally. In order tocoordinate replication with cell growth, the most importantcontrol is carried out at the initiation stage (von Meyenburg& Hansen, 1987). In fast-growing cells (generation time,60 min), the time required for replication and cell divisionis longer than the generation time. To compensate for thispotentially serious problem, cells start a new round ofreplication before previous rounds terminate in a processknown as dichotomous replication (Yoshikawa & Wake,1993). It is unclear how cells precisely recognize the rightstage in the cell cycle to initiate replication.
In Escherichia coli, several positive and negative factorsappear to participate in the regulation of replicationinitiation. DnaA protein level is a positive factor, asincreased levels of DnaA cause overinitiation and advancethe initiation event to an earlier time in the cell cycle(Atlung & Hansen, 1993; Løbner-Olesen et al., 1989). Thelevel of the ATP-bound form of DnaA, which is the activeform for replication initiation, correlates with this property(Kurokawa et al., 1999). A recent finding has indicated thatDiaA tetramers stimulate assembly of active ATP–DnaA–oriC initiation complex by directly binding to multipleDnaA molecules (Keyamura et al., 2007). Three negativemechanisms are known to inhibit extra initiations of DNAreplication. The first one is sequestration of newlyduplicated hemimethylated origins into the cell membraneby SeqA protein (Campbell & Kleckner, 1990; Lu et al.,1994; Ogden et al., 1988). Two others are inactivation ofDnaA initiation protein by Hda and DnaN (b subunit ofDNA polymerase III) (Katayama et al., 1998; Kato &Katayama, 2001) and sequestration of DnaA at the datA
Abbreviations: DAPI, 49,6-diamidino-2-phenylindole; PLR, polar local-ization region.
A supplementary table, listing nucleotide sequences of primers used inthis study, is available with the online version of this paper.
Microbiology (2009), 155, 3070–3082 DOI 10.1099/mic.0.030080-0
3070 030080 G 2009 SGM Printed in Great Britain

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
region on the chromosome (Kitagawa et al., 1998; Ogawaet al., 2002). Accordingly, in E. coli, initiation of replicationis tightly regulated by these factors and mechanisms so thatthe chromosome is replicated only once at the correct stagein every cell cycle.
DnaA is also essential for initiation of replication inBacillus subtilis (Moriya et al., 1990) and high levels ofDnaA protein advance the stage of replication initiation toearlier in the cell cycle (Ogura et al., 2001). It has not yetbeen examined whether both ATP- and ADP-bound formsof DnaA exist in B. subtilis cells (only ATP–DnaA is activefor replication initiation). As to negative factors, intrigu-ingly, there are no homologues of SeqA, Dam methylaseand Hda (Kunst et al., 1997). It is therefore unlikely thatthese two negative regulation mechanisms, sequestration ofnew origins into the cell membrane and inactivation ofDnaA by Hda, are acting in B. subtilis as they do in E. coli.However, a novel negative regulator of replicationinitiation, YabA, has been recently found in B. subtilis(Hayashi et al., 2005; Noirot-Gros et al., 2002). When theYabA level is reduced, replication begins at a decreased cellmass, whereas when the YabA level is increased, replicationinitiation is delayed. This protein does not have asignificant homology with E. coli Hda, but like this protein,it appears to interact with both DnaA and DnaN (slidingclamp of DNA polymerase III) (Noirot-Gros et al., 2002,2006). Very recently, it has been proposed that YabAspatially sequesters DnaA initiator molecules from oriC bybinding with the replisome during most of the cell cycle toprevent inappropriate reinitiation of replication. DnaAcolocalizes with replication origin regions only at early andlate stages of the cell cycle (Soufo et al., 2008). Intriguingly,it has also been very recently reported that Soj, which is a B.subtilis orthologue of ParA engaged in plasmid partition-ing, interacts with DnaA directly and controls its activity(Murray & Errington, 2008). Furthermore, although themechanism has not yet been determined, DNA replicationinitiates at a lower cell mass in pgcA disruption mutants(Weart et al., 2007).
Previously, we constructed an oriC-relocated strain of B.subtilis by transferring oriC from the native locus to argG,which is very far (~1200 kb) from the native oriC on thechromosome (Kadoya et al., 2002). Here we show by flowcytometry that in this oriC-relocated strain, synchrony ofreplication initiation is disrupted and initiation frequencyis enhanced. In order to examine the correlation betweenrelocation of oriC and the control of replication initiation,new oriC-relocated strains were constructed by transferringoriC to various loci on the chromosome. When oriC wasrelocated to loci further than 250 kb counterclockwisefrom the native locus, the strict control of replicationinitiation was disrupted, resulting in asynchronous ini-tiation and early initiation during the cell cycle. AlthoughDnaA levels were measurably higher in the oriC-relocatedstrains compared with the wild-type, this increase was notenough to cause the early initiation. Our results suggestthat the potential for replication initiation is accumulated
well before the actual time of initiation, and that release ofthis potential is suppressed by specific DNA structures inthe oriC region of the B. subtilis chromosome such as so-called ‘Spo0J domains’ (Breier & Grossman, 2007; Murrayet al., 2006), the polar localization region (PLR) (Wu &Errington, 2002) and other unknown higher-order DNAstructures.
METHODS
Plasmids. Nucleotide sequences of primers used in this study are
shown in Supplementary Table S1.
For pSM5022-proA9, pSM5022-9ykcC, pSM5022-9pnbA and
pSM5022-purA9, an internal DNA fragment of each gene was
amplified by PCR using the appropriate F and R primers shown in
Table S1 and cloned into the BamHI site of an oriC plasmid,
pSM5022 (Moriya et al., 1992). The lengths of each internal DNA
fragment amplified were as follows: proA9, 1.07 kb (Hassan et al.,
1997); 9ykcC, 621 bp; 9pnbA, 595 bp; purA9, 660 bp. For pSM5022-
9ermC9, an internal DNA fragment (1188 bp) of the ermC gene was
amplified from pMutinT3 (Moriya et al., 1998) with two primers,
erm-iF and erm-iR, and cloned into the BamHI site of pSM5022.
Strains. B. subtilis strains used in this study are listed in Table 1.
Strains in this table were constructed as follows.
For NIS6553, 59 and 39 halves of the iolJ gene were amplified by PCR
from B. subtilis wild-type chromosomal DNA with two primer sets,
-140kAF-AR and -140kBF-BR (Table S1), respectively. The ermC gene
was amplified from pMutinT3 plasmid DNA using two primers, ermF
and ermR (Table S1). Because -140kAR and -140kBF contain
sequences complementary to ermF and ermR, respectively, the two
iolJ DNA fragments amplified as above can be joined to the ermC gene
by PCR using the -140kAF-ermR and ermF-140kBR primer sets,
respectively. These two PCR fragments, iolJA-ermC and ermC-iolJB,
can be further joined to iolJB and iolJA PCR fragments, respectively,
by PCR with two primers, -140kAF and -140kBR. The resultant PCR
product containing iolJA-ermC-iolJB was used as donor DNA to
transform CRK6000. Erythromycin-resistant transformants were
selected and the insertion of the resistant gene into iolJ gene by
double crossover was confirmed by PCR. Subsequently, this strain,
NIS6503 (Table 1), was again transformed with DNA of plasmid
pSM5022-9ermC9. Chloramphenicol-resistant transformants were
selected and checked for erythromycin-sensitivity. The plasmid
DNA is integrated at the iolJ locus by a single crossover using the
homologous ermC genes for recombination. Proper integration was
confirmed by PCR. Finally, this strain, NIS6513, was again
transformed with chromosomal DNA of NIS6551 to introduce a
deletion of oriC at the native locus. Transformants were selected on a
plate containing erythromycin. The replacement of dnaA–dnaN
(oriC) with the bgaB–ermC construct was confirmed as indicated in
Fig. 1(d).
For NIS6555, SU6556, SU6557 and NIS6558, the strains were
constructed as indicated above for NIS6553, except that chromosomal
DNA of NIS6554 was used as donor followed by selection on a plate
containing spectinomycin to introduce the deletion of the native oriC
(Table 1). The ermR2 primer was used instead of ermR to construct
SU6556 and SU6557.
Escherichia coli, TC1963 [dnaA46 int(mini-R1) RB1-
dnaA : : lacZ,araD139 D(ara, leu)7697 DlacX74, galU, galK, strA]
(Andrup et al., 1988) was used as a cloning host throughout this
study.
Effects of oriC relocation in B. subtilis
http://mic.sgmjournals.org 3071

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
Growth conditions. B. subtilis cells were grown in a rich medium,
Antibiotic Medium 3 (Penassay medium; Becton Dickinson),
supplemented with adenine (20 mg ml21) and guanosine (20 mg
ml21) at 30 or 37 uC throughout this study. When necessary, various
concentrations of antibiotics were added: 7.5 mg chlorampheni-
col ml21, 1.0 mg erythromycin ml21, 40 mg spectinomycin ml21. As
addition of antibiotics reduced the growth rate slightly, cells were
grown without antibiotics for flow cytometry.
4§,6-Diamidino-2-phenylindole (DAPI) staining of cells. Cells
were collected at mid-exponential phase (OD600 0.2) and stained with
DAPI, as previously described (Hassan et al., 1997). Briefly, after
collection by centrifugation, cells were fixed in 70 % (v/v) ethanol at
4 uC overnight and spread on a glass slide coated with poly-L-lysine.
After drying, cells were stained with DAPI solution [1 mg ml21 in
50 % (v/v) glycerol] followed by observation under a BX50
fluorescence microscope (Olympus). DAPI-staining images of cells
were captured by a colour chilled 3CCD camera (C5810,
Hamamatsu) or a high-sensitivity digital black and white cooled
CCD camera (ORCA II-ER-1394, Hamamatsu). Cell length was
measured from 250–500 cells of each strain using MacScope or V-
Windows+ programs.
Flow cytometry. When the OD600 of the B. subtilis cell culture
reached 0.1–0.3, chloramphenicol was added to the culture at a
concentration of 200 mg ml21 in order to inhibit initiation of new
rounds of replication, and the cell culture was further incubated for
5 h to finish ongoing replication. Cells were collected and fixed as
described elsewhere (Løbner-Olesen et al., 1989). Genomic DNA
within cells was stained with a mixture of ethidium bromide and
mithramycin, and the number of genome equivalents in individual
cells, namely the number of replication origins which had existed in
individual exponential-phase cells, was estimated with a Bryte HS
flow cytometer (Bio-Rad Laboratories). Alternatively, the same
analysis was carried out using another flow cytometer, an LSR II
(BD Biosciences), after fixed cells had been stained with Hoechst
33258 and FITC, as described by Torheim et al. (2000).
Measurement of DNA and protein concentrations by colori-
metry. Measurement was carried out exactly as described previously
(Kadoya et al., 2002). Briefly, cells were collected by centrifugation
from 30 ml exponentially growing cell culture, and both nucleic acids
and proteins were extracted by a slightly modified Schneider method
(Kadoya et al., 2002). DNA and protein concentrations in the factions
were quantitatively determined by the Burton (1956) and Lowry
Table 1. B. subtilis strains used in this study
Arrows indicate transformation. Donor DNA and the recipient strain are shown at the left and right of arrows, respectively, with antibiotics and
markers for selection. Cp, chloramphenicol; Em, erythromycin; Sp, spectinomycin; Ems, erythromycin-sensitive; pro2, proline requirement; purA2,
adenine requirement. Names of strains temporarily made during construction are shown in parentheses after selection methods. Transformation of
B. subtilis cells was carried out as described elsewhere (Kunst et al., 1994). chr., chromosomal.
Strain Relevant genotype Reference or construction
CRK6000 purA16 metB5 hisA3 guaB Moriya et al. (1990)
NIS6451 CRK6000 argG : : pAH3 (oriC cat)
DoriC(dnaA-dnaN) : : bgaB-ermC
Kadoya et al. (2002)
NIS6251 CRK6000 proA : : pSM5022-9proA (oriC cat)
DoriC(dnaA-dnaN) : : bgaB-ermC
pSM5022-9proAACRK6000 (Cp, pro2) (NIS6250)
Linear form of pRK4 (Kadoya et al., 2002)ANIS6250 (Em)
NIS6261 CRK6000 ykcC : : pSM5022-9ykcC (oriC cat)
DoriC(dnaA-dnaN) : : bgaB-ermC
pSM5022-9ykcC and linear form of pRK4ACRK6000 (Cp, Em)
NIS6551 CRK6000 pnbA : : pSM5022-9pnbA (oriC cat)
DoriC(dnaA-dnaN) : : bgaB-ermC
pSM5022-9pnbA and linear form of pRK4ACRK6000 (Cp, Em)
NIS6552 CRK6000 purA : : pSM5022-9purA (oriC cat)
DoriC(dnaA-dnaN) : : bgaB-ermC
168 chr.ACRK6000 (purA+) (NIS6502) pSM5022-9purA and
linear form of pRK4ANIS6502 (Cp, purA2)
NIS6553 CRK6000 iolJ (ermC) : : pSM5022-9ermC9
DoriC(dnaA-dnaN) : : bgaB-ermC
iolJA-ermC-iolJB PCR productsACRK6000 (Em) (NIS6503)
pSM5022-9ermC9ANIS6503 (Cp, Ems) (NIS6513)
NIS6551 chr. DNAANIS6513 (Em)
NIS6554 CRK6000 iolJ (ermC) : : pSM5022-9erm9
DoriC(dnaA-dnaN) : : bgaB-spc
Linear form of pEm : : Spc (Lindow et al., 2002)ANIS6553 (Sp)
NIS6555 CRK6000 yxlA (ermC) : : pSM5022-9ermC9
DoriC(dnaA-dnaN) : : bgaB-spc
yxlAA-ermC-yxlAB PCR productsACRK6000 (Em) (NIS6505)
pSM5022-9ermC9ANIS6505 (Cp, Ems) (NIS6515)
NIS6554 chr. DNAANIS6515 (Cp, Sp)
SU6556 CRK6000 albF (ermC) : : pSM5022-9ermC9
DoriC(dnaA-dnaN) : : bgaB-spc
albEF-ermC-albG PCR productsACRK6000 (Em) (SU6506)
pSM5022-9ermC9ASU6506 (Cp, Ems) (SU6516)
NIS6554 chr. DNAASU6516 (Cp, Sp)
SU6557 CRK6000 ywoE (ermC) : : pSM5022-9ermC9
DoriC(dnaA-dnaN) : : bgaB-spc
ywoE-ermC-ywoF PCR productsACRK6000 (Em) (SU6507)
pSM5022-9ermC9ANIS6507 (Cp, Ems) (SU6517)
NIS6554 chr. DNAASU6517 (Cp, Sp)
NIS6558 CRK6000 yvyD (ermC) : : pSM5022-9ermC9
DoriC(dnaA-dnaN) : : bgaB-spc
yvyDA-ermC-yvyDB PCR productsACRK6000 (Em) (NIS6508)
pSM5022-9ermC9ANIS6508 (Cp, Ems) (NIS6518)
NIS6554 chr. DNAANIS6518 (Cp, Sp)
S. Moriya and others
3072 Microbiology 155

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
methods (Lowry et al., 1951), respectively. The DNA : protein ratio
was calculated from the total amounts of DNA and protein in the
fractions.
Western blotting. Cells were collected from 3 ml cell culture at mid-
exponential phase (OD600 0.2) and further treated for Western
blotting as described previously (Moriya et al., 1990). After proteins
in samples had been separated in an SDS-polyacrylamide gradient(10–20 %) gel by electrophoresis, they were blotted onto a Hybond-PPVDF membrane (GE Healthcare Life Sciences). DnaA protein signalswere detected with the ECL-Plus enhanced chemifluorescence system(GE Healthcare Life Sciences) as described previously (Ogura et al.,
Fig. 1. Flow cytometry and DAPI-staining images of oriC-relocated strains. (a) B. subtilis circular genome schematicallyrepresented as a map of 3606. Genes to which oriC has been relocated are indicated. (b) DAPI-staining images of wild-type(CRK6000) and three oriC-relocated strains, NIS6451 (oriC at argG), NIS6251 (oriC at proA) and NIS6261 (oriC at ykcC),which were exponentially grown in Penassay medium at 30 6C. Bar, 4 mm. (c) Flow cytometry of wild-type and NIS6451 oriC-relocated cells exponentially grown in the rich medium. (d) Schematic structures of the integration site (ykcC gene) of an oriC
plasmid and the native oriC site in the novel oriC-relocated strain NIS6261. (i) Structure of the integration site. Thin and thicklines indicate genomic and plasmid DNA, respectively. Thick and thin arrows represent genes and primers, respectively. Shadedarrows show complete and partial ykcC genes at the recombination site between genomic and plasmid DNA. (ii) Structures ofthe oriC region in the oriC-relocated (NIS6261, DoriC) and the wild-type strains. The thick line represents part of the pRK4sequence (Kadoya et al., 2002). ERV, restriction site for EcoRV. (iii) PCR products digested with EcoRV. The oriC region wasamplified by long-range PCR using Ex Taq DNA polymerase (Takara Bio) with p5 and p6 primers [shown in (ii)]. After digestionwith EcoRV, DNA fragments were separated in a 1 % agarose gel by electrophoresis and visualized by staining with ethidiumbromide. Lanes: M, standard markers; 1–2, independent clones of oriC-relocated strain NIS6261; w, wild-type strain.
Effects of oriC relocation in B. subtilis
http://mic.sgmjournals.org 3073

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
2001). The DnaA bands detected were scanned and the signal
intensity was quantitatively estimated using NIH image.
RESULTS
Control of replication initiation is defective in anoriC-relocated strain
We previously constructed an oriC-relocated strain inwhich the replication origin (oriC) was transferred from itsnative position (0u) to the argG locus at 257u of the B.subtilis chromosome (a circular map of 360u) (Kadoya etal., 2002). To examine the effects of relocating oriC oncontrol of replication initiation, the number of replicationorigins in individual cells was analysed by flow cytometry.B. subtilis cells generally tend to form chains, which canmake flow cytometry difficult to interpret. However, ourparent strain (CRK6000) does not form chains under usualcultivation conditions, and thus is suitable for flowcytometry (Ogura et al., 2001).
In the wild-type cells, synchrony of replication initiation isstrictly maintained. Under our cultivation conditions,either four or eight replication origins were detected inindividual wild-type cell particles (Fig. 1c). No peak wasdetected for cells with 16 genome equivalents (data notshown), which was consistent with no chain formation ofthis strain, as described above. Treating B. subtilis cells withcephalexin to inhibit cell division caused cell lysis, unlike inE. coli. This drug, therefore, was not added to B. subtiliscells to prepare cell samples for flow cytometry.Furthermore, as two different techniques, flow cytometry
and fluorescence in situ hybridization, appeared to show asimilar result for the number of replication origins per cellin exponentially grown cells (Kadoya et al., 2002), theprocedure used for flow cytometry in this study seemed toinhibit cell division in B. subtilis.
Flow cytometry revealed that many cells appeared to containsix origins in cells in which oriC was relocated to argG on theB. subtilis chromosome (Fig. 1a, c). The DNA replicationrate may be significantly slowed down in the oriC-relocatedstrain because of collision of the replication forks with head-on transcription on most of the chromosome (Wang et al.,2007a). Therefore, incubation of cells for 5 h after additionof chloramphenicol may not be long enough to completeongoing replication in this mutant for flow cytometry.However, in the DNA histogram of the mutant, cells withgenome equivalents between four and eight would distributerandomly, because replication forks do not completely stopat particular positions on the chromosome. Thus, detectionof a peak of cells with six genome equivalents in the DNAhistogram of this oriC-relocated strain suggests thatasynchronous initiation of replication was actually occurringin this mutant.
In addition, many cells in this oriC-relocated strain seemedto contain more than eight origins, suggesting thatreplication initiation was also enhanced. When individualcells from this strain were observed using phase-contrastmicroscopy they seemed to be longer than wild-type cells(Fig. 1b). Indeed, when the cell length was measured from500 cells, the average cell length of NIS6451 wasapproximately 1.2 mm longer than that of the wild-typecells (Table 2) (note that, as described above, our parent
Table 2. Distribution of cells in size, average cell length, growth rate and DNA : protein ratio of oriC-relocated strains used in thisstudy
Cells were grown in a rich medium (Penassay medium) at 30 uC. At mid-exponential phase, cells were fixed with 70 % ethanol and the cell length
was measured in 250–500 cells. Experiments 1 and 2 were carried out in different laboratories, and thus the distribution of cell length of the wild-
type strain probably reflects the variation of aeration from the use of different shaker incubators. –, Not measured/done.
Strain Distribution of cell
size (mm)
Average cell length (mm)
(relative to wild-type)
Growth rate*
(doublings per
hour)
DNA : protein
ratioD (relative to
wild-type)
Experiment 1 Experiment 2 Experiment 1 Experiment 2
Wild-type 2.36–7.26 2.00–6.03 4.27 (1.00) 3.41 (1.00) 2.00 0.041 (1.00)
oriC at 260 kb 2.33–6.97 – 4.29 (1.00) – 2.00 0.042 (1.02)
oriC at 2140 kb 2.72–7.79 – 4.49 (1.05) – 2.00 0.043 (1.05)
oriC at 2250 kb – 2.14–6.94 – 3.91 (1.15) 1.88 0.046 (1.12)
oriC at 2350 kb 2.76–10.67 – 5.37 (1.26) – 2.00 0.046 (1.12)
oriC at 2450 kb 2.03–15.59 – 5.71 (1.34) – 1.76 0.049 (1.20)
oriC at 2550 kb – 2.49–11.06 – 4.75 (1.39) 1.69 0.050 (1.22)
oriC at pnbA 2.44–10.64 – 5.34 (1.25) – 1.69 0.052 (1.27)
oriC at argG 1.94–11.26 – 5.43 (1.27) – – 0.048 (1.17)
*Doubling time was determined from the growth curve of each strain in Penassay medium at 37 uC by measuring OD600. Growth rate was
calculated by dividing one hour by the doubling time.
DThese values are averages from two or three independent experiments. For every strain, SD was less than 0.001.
S. Moriya and others
3074 Microbiology 155
Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
strain does not form chains at least under these cultivationconditions, and thus cell length was determined bymeasuring the distance between the two ends of individualcells). The oriC-relocated cells with more than eightreplication origins might be produced simply due to theincrease in cell length. If this were the case, only celldivision would be delayed and the balance between DNAreplication and mass increase would be still maintained inthe oriC-relocated strain as in the wild-type strain. Tocheck this possibility, the ratio of DNA to protein wasmeasured quantitatively using a chemical method. Theresults indicated that the ratio was significantly higher(0.048±0.001) in the oriC-relocated strain (NIS6451) thanin the wild-type (0.041±0.001). Thus, DNA replicationwas actually enhanced in this oriC-relocated strain.
Cells divide abnormally at a high frequency whenoriC is relocated near 117‡ of the genetic map
In order to estimate whether replication initiation is alwaysenhanced by relocation of oriC, we constructed anotheroriC-relocated strain, NIS6261, in which oriC was trans-ferred to the ykcC locus at 116u of the B. subtilis circulargenome map (Fig. 1a).
Integration of an oriC plasmid into the ykcC locus andreplacement of the native oriC with an unrelated sequencecontaining an antibiotic-resistance marker were simulta-neously performed through a single transformation usingtwo different donor DNA sources (Table 1). Integration ofthe plasmid into the ykcC locus in both chloramphenicol-and erythromycin-resistant transformants was confirmedby PCR using plasmid- and ykcC-specific primer sets, asshown in Fig. 1d(i) (p1–p2 and p3–p4 primer sets). PCRproducts with the expected lengths were detected in severaltransformants (data not shown). Subsequently, the replace-ment at the native oriC in the transformants was confirmedby PCR followed by digestion with EcoRV. If the oriC locuswas correctly replaced by the bgaB and ermC genes, twoEcoRV DNA fragments (2394 and 2640 bp) would bedetected instead of four fragments [Fig. 1d(ii)]. As shown inFig. 1d(iii), two candidates displayed the expected pattern.The results indicate that these transformants contain theoriC locus relocated into the ykcC locus.
Exponentially growing oriC-relocated cells were stainedwith DAPI to localize the DNA and observed using afluorescence microscope. Surprisingly, they were found tobe very small compared with the wild-type cells, probablydue to increased cell division (Fig. 1b). This was not due totransfer of oriC into the ykcC locus, because the samesmall-cell phenotype, including anucleate cells produced bypolar division, was also observed when oriC was transferredinto another locus close to ykcC, proA, at 118u (Fig. 1b),and when oriN, the replication origin of a plasmid pLS32,was integrated into the proA locus as a new origin ofchromosome replication (Moriya et al., 1997). Theseresults indicate that when DNA replication begins near117u on the B. subtilis chromosome, cells divide prema-
turely for unknown reasons (see Discussion). Because ofthe abnormal cell division events, these two oriC-relocatedstrains are unsuitable for analysis by flow cytometry.
Replication initiation is enhanced in another oriC-relocated strain
Another oriC-relocated strain was constructed to deter-mine whether replication initiation is always enhanced byrelocation of oriC. In the new oriC-relocated strain, oriCwas integrated downstream of the pnbA gene at 300u on theB. subtilis genome map (Fig. 1a). Both integration of oriCat the pnbA locus and replacement of oriC with theunrelated plasmid sequence were confirmed as describedabove.
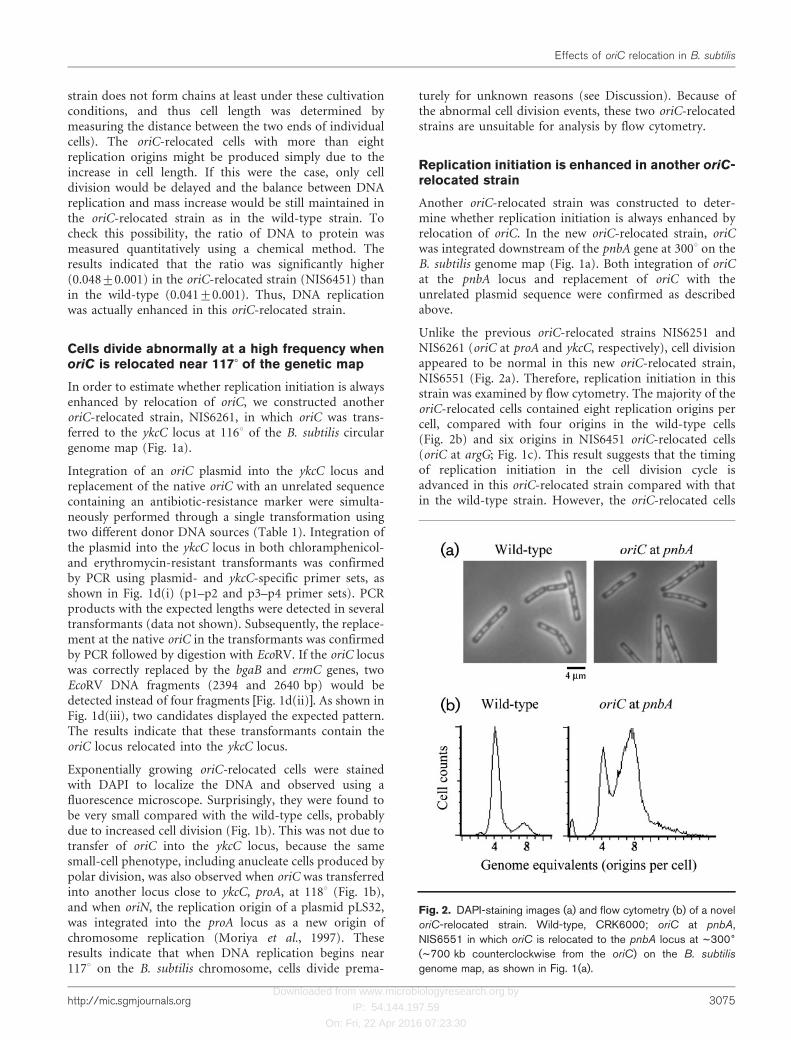
Unlike the previous oriC-relocated strains NIS6251 andNIS6261 (oriC at proA and ykcC, respectively), cell divisionappeared to be normal in this new oriC-relocated strain,NIS6551 (Fig. 2a). Therefore, replication initiation in thisstrain was examined by flow cytometry. The majority of theoriC-relocated cells contained eight replication origins percell, compared with four origins in the wild-type cells(Fig. 2b) and six origins in NIS6451 oriC-relocated cells(oriC at argG; Fig. 1c). This result suggests that the timingof replication initiation in the cell division cycle isadvanced in this oriC-relocated strain compared with thatin the wild-type strain. However, the oriC-relocated cells
Fig. 2. DAPI-staining images (a) and flow cytometry (b) of a noveloriC-relocated strain. Wild-type, CRK6000; oriC at pnbA,NIS6551 in which oriC is relocated to the pnbA locus at ~3006
(~700 kb counterclockwise from the oriC) on the B. subtilis
genome map, as shown in Fig. 1(a).
Effects of oriC relocation in B. subtilis
http://mic.sgmjournals.org 3075

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
were slightly longer than the wild-type cells (Table 2), andthis elongation could lead to an apparent increase in cellswith eight replication origins. To determine whether DNAreplication was enhanced in this oriC-relocated strain, theratio of DNA to protein was quantitatively measured. TheDNA : protein ratio was significantly higher (0.052±0.001)in this oriC-relocated strain, NIS6551, compared with thewild-type (0.041±0.001). These results indicate thatnormal control of replication initiation is disturbed andreplication initiation is upregulated in this new oriC-relocated strain.
DnaA protein levels in oriC-relocated strains weresimilar to those in the wild-type strain
Relocation of oriC alters the gene organization around oriCand could affect gene expression of the oriC-linked dnaA–dnaN operon. An increase in the level of DnaA caused byrelocation of oriC would advance timing of replicationinitiation during the cell division cycle, because over-production of DnaA protein advances the timing ofreplication initiation (Ogura et al., 2001). To test thispossibility, DnaA protein levels were compared betweenoriC-relocated and wild-type strains.
A series of dilutions was prepared for each sample andDnaA protein in the dilutions was detected by Westernblotting (Fig. 3b). As shown in Fig. 3(a), amounts of totalprotein were approximately equal between samples pre-pared from oriC-relocated strains and the wild-type.Densitometry of the DnaA bands revealed that DnaA levelsin NIS6451 (oriC at argG) and NIS6551 (oriC at pnbA)were 1.45±0.20 and 1.12±0.22 times the wild-type level,respectively. Thus, the DnaA level in NIS6451 wassignificantly higher than that in the wild-type but theDnaA level in another oriC-relocated stain, NIS6551, wassimilar to that in the wild-type strain. It has previouslybeen found that when the DnaA level is increased 1.5-fold,
only a slight advance is observed in replication initiation(Ogura et al., 2001). Therefore, the present results indicatethat the asynchronous and advanced initiation of replica-tion observed in oriC-relocated strains is not attributable toan increased cellular level of DnaA.
Flow cytometry of new oriC-relocated strains inwhich oriC was transferred to loci between thenative oriC and pnbA at ~100 kb intervals
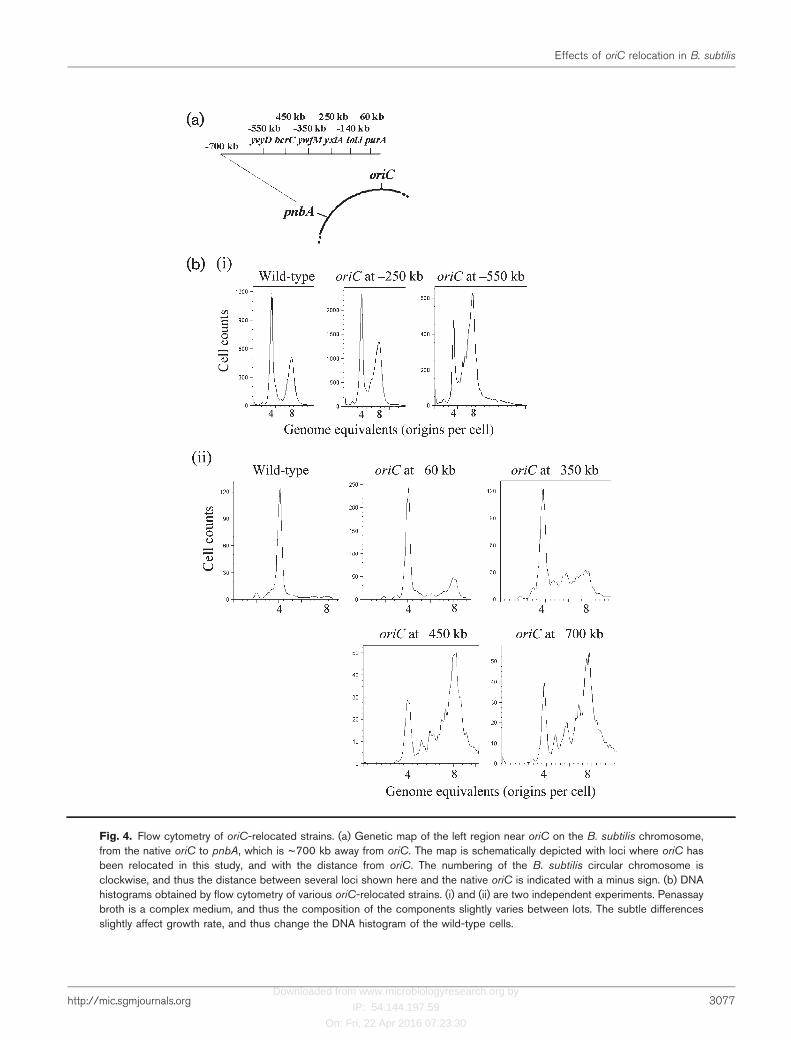
When oriC was relocated to two different loci (argG andpnbA) on the genome, both of which were very far from thenative oriC, strict control of replication initiation was lost.To maintain this strict control, some unique conformationof the chromosome may be formed near the native oriC, orperhaps special genes exist near the native oriC. In order totest this possibility, several new oriC-relocated strains wereconstructed by transferring oriC to new loci between thenative oriC and pnbA at ~100 kb intervals (Fig. 4a), andcontrol of replication initiation was analysed by flowcytometry.
As shown in Fig. 4b(ii), a DNA histogram obtained by flowcytometry from a new oriC-relocated strain in which oriCwas transferred to purA, ~60 kb away from the nativelocus, was nearly the same as that of wild-type cells, havingmainly cells with four origins. Furthermore, a DNAhistogram from another oriC-relocated strain in whichoriC was transferred to iolJ, ~140 kb away from the nativeone, was also very similar to that of the wild-type (data notshown). These results indicate that replication initiation isnormally controlled as long as oriC is relocated within a140 kb region between iolJ and oriC.
In contrast, when oriC was relocated to three loci, 450, 550and 700 kb away from the native locus, the majority of cellscontained eight replication origins per cell, compared withfour origins in wild-type cells [Fig. 4b(ii)]. Moreover,
Fig. 3. DnaA protein levels in oriC-relocatedstrains. (a) Total protein levels in samplesprepared for Western blotting from wild-type(w) and two oriC-relocated strains, NIS6451(oriC at argG) (lane 1) and NIS6551 (oriC atpnbA) (lane 2). Total proteins were extractedfrom exponentially growing cells as describedin Methods and visualized by staining withCoomassie brilliant blue. (b) Western blottingwith rabbit anti-DnaA antibody. Variousamounts of total protein were loaded in thelanes of an SDS-polyacrylamide gradient gel.Total protein at a similar concentration to thatin each lane of (a) was loaded in lane 10, andtotal protein corresponding to 2/5, 3/5 and 4/5of that in lane 10 was loaded in lanes 4, 6 and8, respectively.
S. Moriya and others
3076 Microbiology 155
Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
Fig. 4. Flow cytometry of oriC-relocated strains. (a) Genetic map of the left region near oriC on the B. subtilis chromosome,from the native oriC to pnbA, which is ~700 kb away from oriC. The map is schematically depicted with loci where oriC hasbeen relocated in this study, and with the distance from oriC. The numbering of the B. subtilis circular chromosome isclockwise, and thus the distance between several loci shown here and the native oriC is indicated with a minus sign. (b) DNAhistograms obtained by flow cytometry of various oriC-relocated strains. (i) and (ii) are two independent experiments. Penassaybroth is a complex medium, and thus the composition of the components slightly varies between lots. The subtle differencesslightly affect growth rate, and thus change the DNA histogram of the wild-type cells.
Effects of oriC relocation in B. subtilis
http://mic.sgmjournals.org 3077

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
asynchronous initiation of replication was commonlyobserved in these oriC-relocated strains.
When oriC was relocated to a locus ~350 kb away from thenative one, the oriC-relocated cells showed an intermediateDNA histogram by flow cytometry [Fig. 4b(ii)]. Themajority of the cells contained four replication originsper cell like wild-type cells, but asynchronous initiation wasdetected in this oriC-relocated strain [Fig. 4b(ii)]. A similarpattern was also observed in a DNA histogram obtainedfrom another oriC-relocated strain in which oriC wasrelocated to a locus 250 kb away from the native oriC(Fig. 4b).
The number of cells having eight origins was slightly higherin three oriC-relocated strains (oriC at 260, 2250 and2350 kb) than in the wild-type strain (Fig. 4b). Since cellswere slightly elongated by relocation of oriC (Table 2), thesmall increase in the number of cells that had initiatedreplication might simply be due to cell elongation. If thiswere the case, then the DNA : protein ratio would remainconstant in both oriC-relocated and wild-type strains.Indeed, the DNA : protein ratios in two oriC-relocatedstrains (oriC at 260 and 2140 kb) were nearly the same asin the wild-type strain (Table 2). In contrast, theDNA : protein ratio was significantly increased in twoother oriC-relocated strains (oriC at 2250 and 2350 kb)(Table 2). When oriC was relocated to further away fromits native locus (to 2450, 2550 kb and in pnbA), theDNA : protein ratio was further increased by about 20 %(Table 2). These results clearly indicated that replicationinitiation was actually upregulated in these oriC-relocatedstrains. In other words, the increase in the number of cellswith eight replication origins detected by flow cytometry inthe 2250 and 2350 kb oriC-relocated strains is not simplydue to cell elongation.
Replication initiation is partially inhibited indouble oriC strains
During the course of constructing oriC-relocated strains,double oriC strains can be temporarily created. We wereinterested to know how replication initiation is controlledin these double oriC strains. Using two double oriC strainsin which the extra oriC was located at 2250 or 2550 kb,the number of replication origins in the cells was examinedby flow cytometry.
The analysis revealed that the number of cells having eightorigins was remarkably reduced in the double oriC strainscompared with that in the wild-type strain (Fig. 5). Thischange was not due to reduction in cell size because theaverage cell lengths of the double oriC strains were almostthe same as that of the wild-type (0.99 and 0.88 relative tothe wild-type 1.00). These results indicate that replicationinitiation is slightly delayed in double oriC strains.Furthermore, a significant number of cells were foundbetween the peaks of representing four and eight origins ina DNA histogram from a double oriC strain (extra oriC at
2550 kb) (Fig. 5), suggesting that synchrony of replicationinitiation was also disturbed in the double oriC strain.
DISCUSSION
In this report we show that when oriC is relocated to argG,which is very far from the native oriC (257u on the 360umap), normal control of replication initiation is lost.Initiation became asynchronous and enhanced as theDNA : protein ratio increased in the oriC-relocated strain.Very recently, more frequent replication initiation has beenindependently observed in the same oriC-relocated strainusing genome-wide hybridization on a DNA microarray(Wang et al., 2007a). When oriC was relocated to otherloci, ykcC and proA, which are located very close to eachother on the chromosome (116 and 118u, respectively) andalso very far from the native oriC, cells were much shorterthan the wild-type and appeared to divide more frequently.This extra cell division may be caused by an increase in theFtsZ protein level in the oriC-relocated strains. The ftsZgene is located at ~200 kb downstream of proA (Kunstet al., 1997) and thus replication starts near ftsZ in thesestrains. This situation leads to an increase in ftsZ genedosage in these oriC-relocated cells compared with wild-type cells and may result in elevated levels of FtsZ in theoriC-relocated strains. Alternatively, rRNA synthesis maybe greatly reduced in the two oriC-relocated mutantsbecause transcription of rRNA operons clustered near 0u ofthe chromosome (Kunst et al., 1997) is inhibited bycollision with replication forks in these mutants. Thereduction of ribosomal synthesis decreases the rate of massincrease, resulting in production of small cells by celldivision at normal frequency. To examine this possibility,transcription of the rRNA cluster should be comparedbetween the mutants (oriC at proA and ykcC) and anotheroriC-relocated strain in which oriC is located on the leftarm of the chromosome (oriC at argG) and does notproduce small cells. Due to the abnormal small cellphenotype, replication initiation was not analysed by flow
Fig. 5. Flow cytometry of double oriC strains in which additionaloriCs are located at sites 250 and 550 kb left of the native oriC.
S. Moriya and others
3078 Microbiology 155

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
cytometry in these oriC-relocated strains. However, inanother oriC-relocated strain (oriC at pnbA, 300u), bothasynchronous and enhanced initiation of DNA replicationwere again observed. These results indicate that loss ofnormal regulation and enhanced initiation of replicationdo not only occur in a special strain, but are perhapscommon in many oriC-relocated strains and are notunique to oriC relocation at argG.
The abnormal regulation of replication initiation was notsimply due to an increase in the DnaA initiator protein levelwithin the cell. Although the DnaA levels were slightlyelevated (by about 1.1- and 1.5-fold in the two oriC-relocated strains), the increase was not enough to disrupt thetight regulation of replication initiation in B. subtilis (Oguraet al., 2001). However, the increase in replication initiationwas not observed when the dnaA-independent heterologousorigin oriN was used as the sole origin for chromosomereplication at argG and near the native oriC sites (Wang et al.,2007a). Thus, it still may be that overinitiation of replicationin the oriC-relocated strain is caused by the increased DnaAlevel. Nevertheless, it appears that some characteristic of theregion surrounding the native oriC locus (within 140 kb)contributes to the tight regulation of replication initiation,since control of replication initiation was disturbed whenoriC was relocated to loci further than 250 kb counter-clockwise from the native locus. When oriC was relocated toloci at either 2250 or 2350 kb (counterclockwise), controlof replication initiation was slightly disturbed andDNA : mass ratios increased by 10 % compared with thatof the wild-type. When relocated to loci further than 450 kbfrom the native locus, replication initiation was moreenhanced and DNA : protein ratios increased further byabout 20 %. Thus, two regions seem to participate in thetight control of replication initiation. One lies between 140and 250 kb and another between 350 and 450 kb counter-clockwise from the native oriC.
What mechanism could exist in this origin region toregulate replication initiation? A candidate may bemembrane binding of the origin region, similar to thatreported in E. coli. It has been demonstrated in B. subtilisthat the purA gene near the origin is enriched in themembrane fraction (Winston & Sueoka, 1980; Yamaguchi& Yoshikawa, 1977). A membrane-binding DNA sequencenear purA has been isolated from the membrane fractionand identified by membrane association abilities in vivoand in vitro (Sargent & Bennett, 1986; Sato et al., 1991).However, this sequence is located ~45 kb from oriC (Itayaet al., 1992). Moreover, counterparts of SeqA and Dammethyltransferase, which play essential roles in sequesteringnewly replicated origins to the cell membrane in E. coli, donot exist in B. subtilis (Kunst et al., 1997). Therefore, thenature of any DNA–membrane binding in B. subtilis maybe different from the sequestration of newly replicatedorigins into the cell membrane.
In B. subtilis, DNA replication is arrested at ~200 kbdownstream on either side of oriC during the stringent
response and premature initiation of replication aftertemperature shift-down of dnaB-ts initiation mutants(Henckes et al., 1989; Levine et al., 1991). The replicationterminator protein (RTP) has been proposed to beinvolved in the arrest of replication during the stringentresponse and the arrest sites have been mapped in detail(Autret et al., 1999; Levine et al., 1995), although thereplication arrest by the stringent response has been foundvery recently to occur throughout the chromosomeindependently of Rtp (Wang et al., 2007b). According tothe previous reports, approximately half of the replicationforks are arrested at the LSTer1 site between 294 and2104 kb and the remaining half at the LSTer2 site between2127 and 2135 kb. When oriC was relocated to the2140 kb locus, which is outside these arrest regions,significant effects on the control of replication initiationwere not observed. Thus, it is unlikely that the mechanismthat arrests replication forks during the stringent responseand premature initiation of replication is related to theregulation of replication initiation observed in this study.
A recent paper has shown that replication elongation isslowed down by head-on transcription in an extendedchromosomal region of an oriC-relocated strain (Wang etal., 2007a). In this case, oriC was relocated to argG, thesame as NIS6451 used in our study. The authors observed,by DNA microarray analysis, that replication initiation wasmore frequent in this mutant. This result appears to beconsistent with our result obtained by flow cytometry.However, DNA microarray analysis only shows the relativeratio of amounts of origin- and terminus-proximal DNA,thus the relative increase of origin-proximal DNA may beexplained by, for example, selective degradation of theterminus-proximal region of the chromosome, withoutaffecting control of replication initiation. Such degradationcould occur in cells by treatment with DNA-damagingagents or by collision of replication with transcription.Indeed, the same group detected a relative increase in theamount of origin-proximal DNA after treatment withmitomycin C (Goranov et al., 2006). If such degradation isoccurring, the DNA : protein ratio would be reducedcompared with that under normal conditions. Our resultsindicated that this was not the case for the oriC-relocatedstrains in this study, because the DNA : protein ratio wasactually higher in NIS6451 than in the wild-type.Therefore, our results confirm that replication initiationis actually enhanced in the oriC-relocated strain. Our studyalso showed that a small difference (~100 kb) in relocationof oriC, for instance between 2350 and 2450 kb, caused adrastic change in control of replication initiation. Thus, amechanism other than the slowing of replication elonga-tion may be an appropriate interpretation of the asyn-chronous and enhanced initiation of replication in oriC-relocated strains.
A special higher-order DNA structure may be formed onthe oriC region of the chromosome and such a structuremay be important for the regulation of replicationinitiation in B. subtilis. It has been suggested in E. coli
Effects of oriC relocation in B. subtilis
http://mic.sgmjournals.org 3079

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
that two large regions (~900 kb) near oriC and terC areorganized to form domains (Niki et al., 2000). Indeed, veryrecently, it has been shown that the terminus region(800 kb) of the E. coli chromosome is organized into amacrodomain and plays a role in chromosome segregation(Mercier et al., 2008). In B. subtilis, eight Spo0J-bindingsites are located in the origin-proximal region (~20 %) ofthe chromosome (Lin & Grossman, 1998), and indeed,Spo0J binds to these sites in vivo (Breier & Grossman, 2007;Murray et al., 2006). This binding is not just on the shortrecognition sequence but is spread over 10–20 kb (Breier &Grossman, 2007; Murray et al., 2006). It has been suggestedthat this large nucleoprotein structure (called ‘Spo0Jdomains’) is involved in organizing the oriC region ofthe chromosome (Murray et al., 2006). In B. subtilis, theDnaA protein level in the wild-type cells appears to have apotential which is enough to initiate replication once(Moriya et al., 1990). However, the potential is usuallysuppressed until it becomes the right time for firingorigins. The Spo0J domains in the oriC region may beinvolved in this suppression by inhibiting contact of freeDnaA molecules with replication origins bound to DnaA.Indeed, replication initiation is greatly enhanced in spo0Jnull mutants (Lee et al., 2003; Ogura et al., 2003), and arecent paper suggests that the stimulation of replicationinitiation in the mutant is perhaps caused by alteration ofDNA topology near oriC (Murray & Errington, 2008). InoriC-relocated strains in which oriC is moved to new locioutside the Spo0J domains, namely outside the suppressionregion, free DnaA molecules can attack the new oriC, andthus extra initiation of replication may occur. Six Spo0Jbinding sites are located on the left half of the B. subtilischromosome (Breier & Grossman, 2007). Four out of thesix binding sites are clustered within ~60–70 kb counter-clockwise from oriC, one ~300 kb apart from oriC and thelast one close to terC (Breier & Grossman, 2007). Controlof replication initiation was almost normal when oriC wasrelocated to 260 or 2140 kb and obviously disruptedwhen it was relocated to loci further than 250 kb. Thus,our results would be consistent with the model that theSpo0J binding site located at 334u (Breier & Grossman,2007) is important to form a higher-order structurebetween dispersed Spo0J domains.
The PLR which is essential to bring the oriC region close tothe cell pole in sporulating cells lies ~150–300 kb to the leftof oriC (Wu & Errington, 2002). This long sequence mayalso play a role in the control of replication initiation.However, when oriC was relocated to loci further than450 kb counterclockwise from the native locus, thefrequency of initiation of replication increased further.Therefore, our results also suggest that some DNAstructure outside the two regions (Spo0J domains andPLR) is involved in suppression of replication initiation.When oriC is moved over those DNA regions, allsuppression mediated by the regions would be lostcompletely and free DnaA molecules would make contactwith oriC without any hindrance, resulting in early and
asynchronous initiation of replication. If this is the case,the same abnormal control of replication initiation wouldalso be observed in double oriC mutants. Althoughasynchronous initiation was observed in these mutants,early initiation was not detected even when an extra oriCwas inserted at 2550 kb (Fig. 5). Rather, replicationinitiation was delayed compared with the wild-type. This isprobably due to the strong autorepression activity of DnaA(Ogura et al., 2001). The two dnaA promoters present inthese double oriC mutants would be completely repressedat the wild-type DnaA level. Since free DnaA moleculeswould be titrated by DnaA boxes in two oriCs (Moriya etal., 1988), there may be much fewer residual free DnaAmolecules in double oriC cells than in wild-type cells. Thislow level of free DnaA molecules may delay replicationinitiation in double oriC strains. Another possibility is thatoriC at the native position is dominant over translocatedoriC sequences and that the moderate asynchrony observedin double oriC strains is due to leaky and uncontrolledinitiation of replication at the translocated origin.
ACKNOWLEDGEMENTS
We thank Heath Murray for critical reading of the manuscript. This
study was partly supported by a grant from the UK Biotechnology
and Biological Sciences Research Council.
REFERENCES
Andrup, L., Atlung, T., Ogasawara, N., Yoshikawa, H. & Hansen, F. G.(1988). Interaction of the Bacillus subtilis DnaA-like protein with the
Escherichia coli DnaA protein. J Bacteriol 170, 1333–1338.
Atlung, T. & Hansen, F. G. (1993). Three distinct chromosome
replication states are induced by increasing concentrations of DnaA
protein in Escherichia coli. J Bacteriol 175, 6537–6545.
Autret, S., Levine, A., Vannier, F., Fujita, Y. & Seror, S. J. (1999). The
replication checkpoint control in Bacillus subtilis: identification of a
novel RTP-binding sequence essential for the replication fork arrest
after induction of the stringent response. Mol Microbiol 31, 1665–1679.
Breier, A. M. & Grossman, A. D. (2007). Whole-genome analysis of
the chromosome partitioning and sporulation protein Spo0J (ParB)
reveals spreading and origin-distal sites on the Bacillus subtilis
chromosome. Mol Microbiol 64, 703–718.
Burton, K. (1956). A study of the conditions and mechanism of the
diphenylamine reaction for the colorimetric estimation of DNA.
Biochem J 62, 315–323.
Campbell, J. L. & Kleckner, N. (1990). E. coli oriC and the dnaA gene
promoter are sequestered from dam methyltransferase following the
passage of the chromosomal replication fork. Cell 62, 967–979.
Goranov, A. I., Kuester-Schoeck, E., Wang, J. D. & Grossman, A. D.(2006). Characterization of the global transcriptional responses to
different types of DNA damage and disruption of replication in
Bacillus subtilis. J Bacteriol 188, 5595–5605.
Hassan, A. K. M., Moriya, S., Ogura, M., Tanaka, T., Kawamura, F. &Ogasawara, N. (1997). Suppression of initiation defects of chro-
mosome replication in Bacillus subtilis dnaA and oriC-deleted
mutants by integration of a plasmid replicon into the chromosomes.
J Bacteriol 179, 2494–2502.
S. Moriya and others
3080 Microbiology 155

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
Hayashi, M., Ogura, Y., Harry, E. J., Ogasawara, N. & Moriya, S.(2005). Bacillus subtilis YabA is involved in determining the timing
and synchrony of replication initiation. FEMS Microbiol Lett 247,
73–79.
Henckes, G., Harper, F., Levine, A., Vannier, F. & Seror, S. J. (1989).Over-replication of the origin region in the dnaB37 mutant of Bacillus
subtilis: post-initiation control of chromosomal replication. Proc Natl
Acad Sci U S A 86, 8660–8663.
Itaya, M., Laffan, J. J. & Sueoka, N. (1992). Physical distance between
the site of type II DNA binding to the membrane and oriC on the
Bacillus subtilis 168 chromosome. J Bacteriol 174, 5466–5470.
Kadoya, R., Hassan, A. K. M., Kasahara, Y., Ogasawara, N. &
Moriya, S. (2002). Two separate DNA sequences within oriC
participate in accurate chromosome segregation in Bacillus subtilis.
Mol Microbiol 45, 73–87.
Katayama, T., Kubota, T., Kurokawa, K., Crooke, E. & Sekimizu, K.(1998). The initiator function of DnaA protein is negatively regulated
by the sliding clamp of the E. coli chromosomal replicase. Cell 94,
61–71.
Kato, J. & Katayama, T. (2001). Hda, a novel DnaA-related protein,
regulates the replication cycle in Escherichia coli. EMBO J 20, 4253–
4262.
Keyamura, K., Fujikawa, N., Ishida, T., Ozaki, S., Su’etsugu, M.,Fujimitsu, K., Kagawa, W., Yokoyama, S., Kurumizaka, H. &Katayama, T. (2007). The interaction of DiaA and DnaA regulates
the replication cycle in E. coli by directly promoting ATP DnaA-
specific initiation complexes. Genes Dev 21, 2083–2099.
Kitagawa, R., Ozaki, T., Moriya, S. & Ogawa, T. (1998). Negative
control of replication initiation by a novel chromosomal locus
exhibiting exceptional affinity for Escherichia coli DnaA protein. Genes
Dev 12, 3032–3043.
Kunst, F., Msadek, T. & Rapoport, G. (1994). Signal transduction
network controlling degradative enzyme synthesis and competence in
Bacillus subtilis. In Regulation of Bacterial Differentiation, pp. 1–20.
Edited by P. J. Piggot, C. P. Moran, Jr & P. Youngman. Washington,
DC: American Society for Microbiology.
Kunst, F., Ogasawara, N., Moszer, I., Albertini, A. M., Alloni, G.,Azevedo, V., Bertero, M. G., Bessieres, P., Bolotin, A. & otherauthors (1997). The complete genome sequence of the Gram-positive
bacterium Bacillus subtilis. Nature 390, 249–256.
Kurokawa, K., Nishida, S., Emoto, A., Sekimizu, K. & Katayama, K.(1999). Replication cycle-coordinated change of the adenine nucleo-
tide-bound forms of DnaA protein in Escherichia coli. EMBO J 18,
6642–6652.
Lee, P. S., Lin, D. C., Moriya, S. & Grossman, A. D. (2003). Effects of
the chromosome partitioning protein Spo0J (ParB) on oriC
positioning and replication initiation in Bacillus subtilis. J Bacteriol
185, 1326–1337.
Levine, A., Vannier, F., Dehbi, M., Henckes, G. & Seror, S. J. (1991).
The stringent response blocks DNA replication outside the ori region
in Bacillus subtilis and at the origin in Escherichia coli. J Mol Biol 219,
605–613.
Levine, A., Autret, S. & Seror, S. J. (1995). A checkpoint involving
RTP, the replication terminator protein, arrests replication down-
stream of the origin during the stringent response in Bacillus subtilis.
Mol Microbiol 15, 287–295.
Lin, D. C. & Grossman, A. D. (1998). Identification and character-
ization of a bacterial chromosome partitioning site. Cell 92, 675–685.
Lindow, J. C., Kuwano, M., Moriya, S. & Grossman, A. D. (2002).Subcellular localization of the Bacillus subtilis structural maintenance
of chromosomes (SMC) protein. Mol Microbiol 46, 997–1009.
Løbner-Olesen, A., Skarstad, K., Hansen, F. G., von Meyenburg, K. &Boye, E. (1989). The DnaA protein determines the initiation mass ofEscherichia coli K-12. Cell 57, 881–889.
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. (1951). Proteinmeasurement with the Folin phenol reagent. J Biol Chem 193, 265–275.
Lu, M., Campbell, J. L., Boye, E. & Kleckner, N. (1994). SeqA: a negativemodulator of replication initiation in E. coli. Cell 77, 413–426.
Mercier, R., Petit, M.-A., Schbath, S., Robin, S., Karoui, M. E.,Boccard, F. & Espeli, O. (2008). The MatP/matS site-specific systemorganizes the terminus region of the E. coli chromosome into amacrodomain. Cell 135, 475–485.
Moriya, S., Fukuoka, T., Ogasawara, N. & Yoshikawa, H. (1988).Regulation of initiation of the chromosomal replication by DnaA-boxes in the origin region of the Bacillus subtilis chromosome. EMBOJ 7, 2911–2917.
Moriya, S., Kato, K., Yoshikawa, H. & Ogasawara, N. (1990). Isolationof a dnaA mutant of Bacillus subtilis defective in initiation ofreplication: amount of DnaA protein determines cells’ initiationpotential. EMBO J 9, 2905–2910.
Moriya, S., Atlung, T., Hansen, F. G., Yoshikawa, H. & Ogasawara, N.(1992). Cloning of an autonomously replicating sequence (ars) fromthe Bacillus subtilis chromosome. Mol Microbiol 6, 309–315.
Moriya, S., Hassan, A. K. M., Kadoya, R. & Ogasawara, N. (1997).Mechanism of anucleate cell production in the oriC-deleted mutantsof Bacillus subtilis. DNA Res 4, 115–126.
Moriya, S., Tsujikawa, E., Hassan, A. K. M., Asai, K., Kodama, T. &Ogasawara, N. (1998). A Bacillus subtilis gene-encoding proteinhomologous to eukaryotic SMC motor protein is necessary forchromosome partition. Mol Microbiol 29, 179–187.
Murray, H. & Errington, J. (2008). Dynamic control of the DNAreplication initiation protein DnaA by Soj/ParA. Cell 135, 74–84.
Murray, H., Ferreira, H. & Errington, J. (2006). The bacterialchromosome segregation protein Spo0J spreads along DNA fromparS nucleation sites. Mol Microbiol 61, 1352–1361.
Niki, H., Yamaichi, Y. & Hiraga, S. (2000). Dynamic organization ofchromosomal DNA in Escherichia coli. Genes Dev 14, 212–223.
Noirot-Gros, M.-F., Dervyn, E., Wu, L. J., Mervelet, P., Errington, J.,Ehrlich, S. D. & Noirot, P. (2002). An expanded view of bacterial DNAreplication. Proc Natl Acad Sci U S A 99, 8342–8347.
Noirot-Gros, M.-F., Velten, M., Yoshimura, M., McGovern, S.,Morimoto, T., Ehrlich, S. D., Ogasawara, N., Polard, P. & Noirot, P.(2006). Functional dissection of YabA, a negative regulator of DNAreplication initiation in Bacillus subtilis. Proc Natl Acad Sci U S A 103,2368–2373.
Ogawa, T., Yamada, Y., Kuroda, T., Kishi, T. & Moriya, S. (2002). ThedatA locus predominantly contributes to the initiator titrationmechanism in the control of replication initiation in Escherichia coli.Mol Microbiol 44, 1367–1375.
Ogden, G. B., Pratt, M. J. & Schaechter, M. (1988). The replicationorigin of the Escherichia coli chromosome binds to cell membranesonly when hemimethylated. Cell 54, 127–135.
Ogura, Y., Imai, Y., Ogasawara, N. & Moriya, S. (2001).Autoregulation of the dnaA–dnaN operon and effects of DnaAprotein levels on replication initiation in Bacillus subtilis. J Bacteriol183, 3833–3841.
Ogura, Y., Ogasawara, N., Harry, E. J. & Moriya, S. (2003). Increasingthe ratio of Soj to Spo0J promotes replication initiation in Bacillussubtilis. J Bacteriol 185, 6316–6324.
Sargent, M. G. & Bennett, M. F. (1986). Identification of a specificmembrane-particle-associated DNA sequence in Bacillus subtilis.J Bacteriol 166, 38–43.
Effects of oriC relocation in B. subtilis
http://mic.sgmjournals.org 3081

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Fri, 22 Apr 2016 07:23:30
Sato, Y., McCollum, M., McKenzie, T., Laffan, J., Zuberi, A. & Sueoka, N.(1991). In vitro type II binding of chromosomal DNA to membrane inBacillus subtilis. J Bacteriol 173, 7732–7735.
Soufo, C. D., Soufo, H. D. J., Noirot-Gros, M.-F., Steindorf, A., Noirot, P.& Graumann, P. L. (2008). Cell-cycle-dependent spatial sequestrationof the DnaA replication initiator protein in Bacillus subtilis. Dev Cell 15,935–941.
Torheim, N. K., Boye, E., Løbner-Olesen, A., Stokke, T. & Skarstad, K.(2000). The Escherichia coli SeqA protein destabilizes mutant DnaA204protein. Mol Microbiol 37, 629–638.
von Meyenburg, K. & Hansen, F. G. (1987). Regulation of chromosomereplication. In Escherichia coli and Salmonella typhimurium: Cellularand Molecular Biology, vol. 2, pp. 1555–1577. Edited by F. C. Neidhardt,J. L. Ingraham, K. B. Low, B. Magasanik, M. Schaechter & H. E.Umbarger. Washington, DC: American Society for Microbiology.
Wang, J. D., Berkmen, M. B. & Grossman, A. D. (2007a). Genome-wide coorientation of replication and transcription reduces adverseeffects on replication in Bacillus subtilis. Proc Natl Acad Sci U S A 104,5608–5613.
Wang, J. D., Sanders, G. M. & Grossman, A. D. (2007b). Nutritionalcontrol of elongation of DNA replication by (p)ppGpp. Cell 128, 865–875.
Weart, R. B., Lee, A. H., Chien, A.-C., Haeusser, D. P., Hill, N. S. &Levin, P. A. (2007). A metabolic sensor governing cell size in bacteria.
Cell 130, 335–347.
Winston, S. & Sueoka, N. (1980). DNA–membrane association is
necessary for initiation of chromosomal and plasmid replication in
Bacillus subtilis. Proc Natl Acad Sci U S A 77, 2834–2838.
Wu, L. J. & Errington, J. (2002). A large dispersed chromosomal region
required for chromosome segregation in sporulating cells of Bacillus
subtilis. EMBO J 21, 4001–4011.
Yamaguchi, K. & Yoshikawa, H. (1977). Chromosome–membrane
association in Bacillus subtilis. III. Isolation and characterization of a
DNA-protein complex carrying replication origin markers. J Mol Biol
110, 219–253.
Yoshikawa, H. & Wake, G. (1993). Initiation and termination of
chromosome replication. In Bacillus subtilis and other Gram-Positive
Bacteria: Biochemistry, Physiology, and Molecular Genetics, pp. 507–
528. Edited by A. L. Sonenshein, J. A. Hoch & R. Losick. Washington,
DC: American Society for Microbiology.
Edited by: L. Janniere
S. Moriya and others
3082 Microbiology 155




















