
Effects of nicotinic and muscarinic ligands on embryonic neurones of Periplaneta americana in...
14
Journal of Insect Physiology 44 (1998) 227–240 Effects of nicotinic and muscarinic ligands on embryonic neurones of Periplaneta americana in primary culture: a whole cell clamp study Isabelle Van Eyseren, Franc ¸ois Tiaho, Janine Le Guen, Jean Claude Guillet, Yves Pichon * Groupe de neurobiologie, Equipe C.R.M., UPRES-A CNRS 6026, Universite ´ de Rennes, Campus de Beaulieu, F-35042 Rennes-cedex, France Received 30 June 1997; received in revised form 25 September 1997 Abstract The pharmacological properties of acetylcholine (ACh) receptors of cultured neurones from embryonic cockroach brains were studied using the whole-cell configuration of the patch-clamp technique. More than 90% of the studied neurones responded to ACh by a monophasic inward current, the intensity of which varied from cell to cell. The sequence of potency of the five tested agonists was ACh > nicotine = carbamylcholine > suberyldicholine = oxotremorine. The dose–response relationship was complex, suggesting the existence of two populations of receptors: high-affinity receptors (extrapolated K d around 10 -7 M) and low-affinity receptors (extrapolated K d around 5 × 10 25 M). The current–voltage relationship of the induced current was linear between 2 80 and 2 40 mV and the extrapolated reversal potential was not significantly different from 0 mV. The sequence of decreasing potency of the antagonists of the ACh response was: methyllycaconitine > a-bungarotoxin > mecamylamine > curare > strychnine > bicucul- line > atropine > picrotoxin. These results show: (1) that, in embryonic brain neurones, the response to ACh corresponds to the opening of non-selective cationic channels; and (2) that the pharmacology of the ACh receptors is mainly but not solely nicotinic. The nature of the single events which underlie this response, as well as the structure of the channels (homo or hetero-oligomeric) remain to be investigated. 1998 Elsevier Science Ltd. All rights reserved. Keywords: ACh-receptors; Embryonic neurones; Periplaneta americana; Patch-clamp; Primary culture 1. Introduction The two main excitatory neurotransmitters are acetyl- choline (ACh) and glutamate. In vertebrates, ACh is mostly involved in fast transmission at the neuromuscu- lar junction whereas glutamate is responsible for fast transmission between neurones in the central nervous system (CNS). In insects, their respective role is inverted: ACh is the main excitatory neurotransmitter in the CNS whereas neuromuscular transmission is mediated by glutamate. The insect CNS contains large amounts of acetylcholine (see review by Pichon and Manaranche, 1985) and the CNS of insects is one of the richest sources of ACh receptors (AChRs) known today, besides the Torpedo electroplax. * Corresponding author. 0022–1910 /98 /$19.00 1998 Elsevier Science Ltd. All rights reserved. PII:S0022-1910(97)00165-0 A good deal of our present knowledge on cholinergic transmission in insects originates from studies on the American cockroach, Periplaneta americana L. (see Callec, 1974, 1985; Pichon, 1974). With the development of molecular biology and the cloning of a large number of membrane receptors and ionic channels, our understanding of the mode of action of receptor ligands has changed dramatically. Thus, it is now generally accepted that nicotinic receptors and muscarinic receptors belong to two distinct superfamilies with clearly different molecular structures: the former belong to the ligand-gated ion channel (ionotropic) superfamily, the latter to the second messenger linked receptor (metabotropic) superfamily (Pichon, 1993). To try to establish a correlation between the overall effects of the cholinergic ligands on the insect CNS and known or suspected properties of the ACh receptors, it was necessary to obtain more quantitative data, both at the single channel level and at the whole cell level.
-
Upload
univ-rennes1 -
Category
Documents
-
view
0 -
download
0
Transcript of Effects of nicotinic and muscarinic ligands on embryonic neurones of Periplaneta americana in...

Journal of Insect Physiology 44 (1998) 227–240
Effects of nicotinic and muscarinic ligands on embryonic neuronesof Periplaneta americanain primary culture: a whole cell clamp
study
Isabelle Van Eyseren, Franc¸ois Tiaho, Janine Le Guen, Jean Claude Guillet,Yves Pichon*
Groupe de neurobiologie, Equipe C.R.M., UPRES-A CNRS 6026, Universite´ de Rennes, Campus de Beaulieu, F-35042 Rennes-cedex, France
Received 30 June 1997; received in revised form 25 September 1997
Abstract
The pharmacological properties of acetylcholine (ACh) receptors of cultured neurones from embryonic cockroach brains werestudied using the whole-cell configuration of the patch-clamp technique. More than 90% of the studied neurones responded to AChby a monophasic inward current, the intensity of which varied from cell to cell. The sequence of potency of the five tested agonistswas ACh > nicotine= carbamylcholine > suberyldicholine= oxotremorine. The dose–response relationship was complex, suggestingthe existence of two populations of receptors: high-affinity receptors (extrapolatedKd around 10−7 M) and low-affinity receptors(extrapolatedKd around 5× 1025 M). The current–voltage relationship of the induced current was linear between2 80 and2 40 mV and the extrapolated reversal potential was not significantly different from 0 mV. The sequence of decreasing potencyof the antagonists of the ACh response was: methyllycaconitine >a-bungarotoxin > mecamylamine > curare > strychnine > bicucul-line > atropine > picrotoxin. These results show: (1) that, in embryonic brain neurones, the response to ACh corresponds to theopening of non-selective cationic channels; and (2) that the pharmacology of the ACh receptors is mainly but not solely nicotinic.The nature of the single events which underlie this response, as well as the structure of the channels (homo or hetero-oligomeric)remain to be investigated. 1998 Elsevier Science Ltd. All rights reserved.
Keywords:ACh-receptors; Embryonic neurones;Periplaneta americana; Patch-clamp; Primary culture
1. Introduction
The two main excitatory neurotransmitters are acetyl-choline (ACh) and glutamate. In vertebrates, ACh ismostly involved in fast transmission at the neuromuscu-lar junction whereas glutamate is responsible for fasttransmission between neurones in the central nervoussystem (CNS). In insects, their respective role isinverted: ACh is the main excitatory neurotransmitter inthe CNS whereas neuromuscular transmission ismediated by glutamate. The insect CNS contains largeamounts of acetylcholine (see review by Pichon andManaranche, 1985) and the CNS of insects is one of therichest sources of ACh receptors (AChRs) known today,besides theTorpedoelectroplax.
* Corresponding author.
0022–1910 /98 /$19.00 1998 Elsevier Science Ltd. All rights reserved.PII: S0022-1910 (97)00165-0
A good deal of our present knowledge on cholinergictransmission in insects originates from studies on theAmerican cockroach,Periplaneta americanaL. (seeCallec, 1974, 1985; Pichon, 1974).
With the development of molecular biology and thecloning of a large number of membrane receptors andionic channels, our understanding of the mode of actionof receptor ligands has changed dramatically. Thus, itis now generally accepted that nicotinic receptors andmuscarinic receptors belong to two distinct superfamilieswith clearly different molecular structures: the formerbelong to the ligand-gated ion channel (ionotropic)superfamily, the latter to the second messenger linkedreceptor (metabotropic) superfamily (Pichon, 1993). Totry to establish a correlation between the overall effectsof the cholinergic ligands on the insect CNS and knownor suspected properties of the ACh receptors, it wasnecessary to obtain more quantitative data, both at thesingle channel level and at the whole cell level.

228 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
Compared with the relative wealth of information con-cerning single channel activity in vertebrate muscles andnerve cells, very little is known about insect receptors.The first published work of a single channel study of theeffects of ACh (and CCh) was made on cultured cock-roach neurones (Beadle et al., 1985), it was soon fol-lowed by that of Hanke and Breer (1986), who describeda single channel activity induced by the activation, bysuberyl-dicholine, of a locust neuronal acetylcholinereceptor protein reconstituted in planar lipid bilayers,and that of Sattelle et al. (1986) on dissociated adultcockroach neurones. These first accounts were followedby a few other studies: studies on the house fly,Muscadomestica(Harrison et al., 1990; Leech and Sattelle,1992; Albert and Lingle, 1993), on the fruit fly,Droso-phila melanogaster(Albert and Lingle, 1993), on thelocust,Schistocerca gregaria(Albert and Lingle, 1993)and on cockroach brain neurones in culture (Beadle etal., 1989). Altogether, these studies reveal that, in most,if not all preparations, the macroscopic ACh current cor-responds to the opening of two major classes of ACh-activated channels with average conductances of 20–32pS for the small channels and 37–59 pS for the largechannels, depending upon the species and the experi-mental conditions (Beadle et al., 1989; Albert and Lin-gle, 1993). The current–voltage relation of these singlechannels was linear and their reversal potential was notfar from zero, suggesting that they are non-selective cat-ion channels. In that respect, insect ACh receptors do notdiffer from vertebrate neuronal nicotinic-ACh receptors.Since the two major studies in which these two popu-lations of receptors have been well characterized werecarried out on embryonic or larval preparations, it hasbeen suggested that the small conductance channelscould correspond to an embryonic form and the largeconductance channels to an adult form of the same pro-tein (Beadle et al., 1989).
Our understanding of the properties of the ACh-Rs ofthe insect CNS is still rudimentary. Thus, although mostsingle channel studies were carried out at low concen-trations of agonists (in themM range), it is reasonableto assume that some desensitization might have occurredduring the course of the experiments (see Albert and Lin-gle, 1993; p. 608). Under these conditions, one cannotexclude the possibility that single channel activity corre-sponding to the fast response of the neurones to AChmay have been missed. Furthermore, it is known that theconcentration of ACh in the synaptic clefts during therelease of the neurotransmitter is probably very high (inthe mM range), the biophysical and pharmacologicalproperties of the channels which are open at these con-centrations might well differ from those associated withthe activation of high-affinity receptors. In the above-mentioned studies, the dose–response curves have beenlimited to a very restricted range of concentrations. Fur-thermore, the technique which was used to apply the
agonists (iontophoresis or pressure application) did notallow precise kinetic studies to be carried out. Thepresent study has been devoted to the study of the whole-cell response of cultured brain neurones of embryoniccockroaches to controlled superfusion with a wide rangeof concentrations of cholinergic agonists in the absenceand in the presence of antagonists. This preparation hasbeen chosen for several reasons:
1. The cell somata have a simple geometry (a sphere)and the cell membrane is directly accessible to themolecules contained in the external medium due tothe absence of glia.
2. The age of the embryos used to start the culture aswell as the age of the culture are known and con-trolled.
3. The mean cell size is 12± 0.23mm and the neuritesare very thin (0.5–2mm), conditions under which areasonably good control of the membrane voltage canbe achieved, especially when the membrane currentsare small, as in the present experiments.
4. Seals can easily be formed between the neuronalmembrane and patch pipettes and kept for extendedperiods of time (up to 1 h).
2. Materials and methods
2.1. Cell culture
The egg cases which hold 14–16 embryos were col-lected every day from selected insect cages containinga restricted number of adult females and a few adultmales. They were then stored in an incubator at 28–29°Cin a humid atmosphere for 21–23 days before being usedfor the culture. The culture technique was derived fromthat of Chen and Levi-Montalcini (1970a, b), asdescribed by Beadle and Hicks (1985) and modified byAmar (1991). The egg cases were cleaned, sterilized inabsolute ethanol for 10 min, then cut longitudinally withsharp scissors. The heads of the embryos were thenremoved and stored in a small sterile vessel containingSchneider’s revisedDrosophila medium. The brainswere then removed from the head capsules and trans-ferred into a second vessel. When a sufficient numberof brains were isolated, they were subsequently trans-ferred into a glass tube where the cells were dissociatedby gentle mechanical trituration with a Pasteur pipette(no enzyme treatment was necessary) in a defined vol-ume of culture medium. The cultures were initiated in amedium (5 + 4) containing five parts of Schneider’srevisedDrosophila medium and four parts of Eagle’sbasal medium containing 100 iu/ml penicillin and 100mg/ml streptomycin complemented with 6 mg/mll-glu-tamine and 2.5mg/ml fungizone. Four drops of the cellsuspension were placed in the middle of a 50 mm outside

229I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
diameter Falcon light-seal plastic Petri dish coated withpoly-l-lysine and covered with a sterile glass coverslipsupported on the side by two small 1 mm thick glasspylons, forming a ‘hanging column’. After 45 min, theculture dishes were turned upside down, to separate theliving neurones which stick to the plastic from thedebris, and stored in an incubator at 29°C in a humidatmosphere. After 5–7 days, the first (4+ 5) culturemedium was replaced by a second (L+ G) medium madeup of equal parts of Leibovitz’s L-15 medium and Yunk-er’s modified Grace medium containing penicillin, strep-tomycin, glutamine and fungizone supplemented with10% foetal calf serum. This second medium wasrenewed every week. Whereas, in the first medium, thevast majority of the glial cells did not survive and littleneuronal growth was seen, the second medium enabledthe fast development of neuritic processes into a densenetwork which could survive up to 3 months. The mor-phology of the cell bodies remained basically the samethroughout the culture although neurones tended to formsmall aggregates. All culture media were obtained fromGibco–BRL, Cergy Pontoise, France.
2.2. Experimental procedure
Immediately before the experiments, the culturemedium was replaced by a solution containing 100 mMNaCl, 70 mM Tris–HCl, 3.1 mM KCl, 5 mM CaCl2, 4mM MgCl2, 10 mM Hepes buffer adjusted to pH 7.2using NaOH, and the Petri dish was placed directly ontothe stage of a Nikon TMS inverted microscope. The cellswere usually observed under phase contrast. All experi-ments were performed at room temperature (20–27°C).
The electrophysiological properties of the neuroneswere studied using the ‘whole-cell’ configuration of the‘patch-clamp’ technique (Hamill et al., 1981).
The patch electrodes were made using Borosilicate 1.5mm (Clark Electromedical) glass using aFlaming/Brown micropipette puller (Sutter InstrumentsCo., USA). They were filled with a solution containing90 mM K-gluconate, 20 mM KCl, 2 mM MgCl2, 10 mMEGTA, 3 mM ATP, 0.5 mM GTP and 10 mM Hepesbuffer adjusted to pH 7.3 using KOH and their resistanceranged from 2 to 10 MV. These electrodes were placedon the headstage of an Axopatch 1C amplifier (AxonInstruments, Forster City, CA, USA). The tip of the elec-trode was advanced towards the cell soma until a gentlecontact was established. A small negative pressure wasthen usually sufficient to create a gigaseal ( > 10 GV).The patch membrane was then destroyed by suction or,when necessary, by applying a short (0.1–5 ms) 1.5 Vpulse across the membrane. The resting membranepotential of the cell was noted under current-clamp, thenthe patch amplifier switched back to the voltage-clampmode at a holding membrane potential of2 60 mV.When the patch was stable for 1–2 min, the superfusion
system was turned on with normal saline in the superfus-ate. The superfusion was maintained at a constant ratethroughout the experiment.
The output of the patch amplifier was connected to aTektronix 5103-M storage oscilloscope (Tektronix,Beaverton, OR, USA) and to the input stages of a modi-fied Sony digital audio processor (PCM 701) connectedto a Sony Betamax video recorder (Sony Corporation,Japan). The current traces were filtered using a 80dB/decade 8 pole low pass Bessel filter set at 5 KHz.
In a limited number of experiments, a different set-up was used which included a Biologic RK-300 patchamplifier and a 80486 PC clone connected to the ampli-fier through a Labmaster (Scientific Solutions, USA)data acquisition board. In these experiments, the pClamp(Axon Instruments) 5.5 software was used for stimu-lation, data acquisition and analysis.
2.3. Superfusion system
To enable a quantitative analysis of the effects of thevarious agonists and antagonists, special care was takenin the design of the superfusion system. The superfusionapparatus consisted of a battery of six reservoirs locatedabout 50 cm above the preparation connected to 1 cmlong, 0.5 mm inner diameter glass tubing via plastic tub-ing of decreasing diameters. Each reservoir could beopened and closed independently. The flow rate of eachchannel was adjusted before each experiment to 300ml/min. The tip of the superfusion tubing was placed inthe bath at about 500mm from the studied neurone.Under these conditions, unstirred layer effects as well asturbulences were restricted to a minimum and the asym-metry in the concentrations between the exposed andsheltered side of the cell restricted to a minimum.Experiments in which the potassium concentration of theperfusing fluid was changed from 3.1 to 20 mM whilethe cell was voltage-clamped at2 60 mV indicated: (1)that the change in concentration around the cell (as esti-mated from the time course of the inward potassiumcurrent) was achieved within about 1 s (measured timeconstants of the on and off responses of 0.9 and 0.8 s,respectively); and (2) no tail of current or rebound fol-lowed the return to the low K+ solution, indicating thatthere was no turbulence. A quick calculation indicatedthat the speed of the flowing solution at the level of thecell was of about 1 mm/s. The antagonists were appliedbefore and during the superfusion of the agonists. In allcases, the level of the solution in the culture dish wasmaintained constant using a capillarity based suctionsystem.
2.4. Drugs
The following drugs were used in the experiments:acetylcholine (ACh), carbamylcholine (CCh), nicotine,
230 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
suberyl-dicholine (SCh), oxotremorine (OXO),a-bunga-rotoxin (a-Bgtx), curare, atropine sulphate, mecamylam-ine (MEC), methyllycaconitine (MLA), picrotoxin, bicu-culline and strychnine. Methyllycaconitine waspurchased from RBI (Research Chemical International).All the other molecules were purchased from Sigma.
2.5. Data analysis
The data were analysed off-line. They were digitizedon 12 bits on an 80286 PC clone using a Data Trans-lation (DT2801A) board at a 50 Hz sampling frequency.The resulting binary data files were examined using aspecially designed program (pat2s), which enabledsome quantification as well as the selection of the inter-esting parts of the recordings and their conversion intoASCII files directly compatible with all available dataanalysis software. In the present experiments, the Excel(Microsoft), Freelance Graphics (Lotus) and Sigmaplot(Jandel Scientific) programs were used.
3. Results
3.1. ACh-induced currents
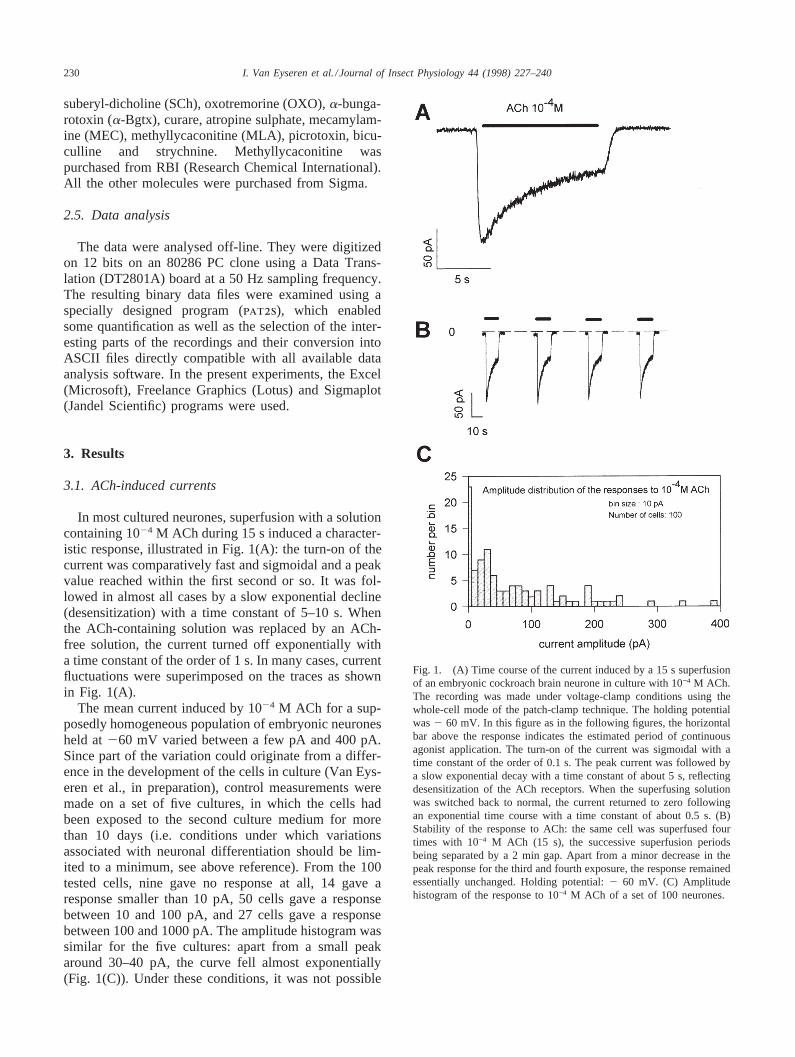
In most cultured neurones, superfusion with a solutioncontaining 1024 M ACh during 15 s induced a character-istic response, illustrated in Fig. 1(A): the turn-on of thecurrent was comparatively fast and sigmoidal and a peakvalue reached within the first second or so. It was fol-lowed in almost all cases by a slow exponential decline(desensitization) with a time constant of 5–10 s. Whenthe ACh-containing solution was replaced by an ACh-free solution, the current turned off exponentially witha time constant of the order of 1 s. In many cases, currentfluctuations were superimposed on the traces as shownin Fig. 1(A).
The mean current induced by 1024 M ACh for a sup-posedly homogeneous population of embryonic neuronesheld at260 mV varied between a few pA and 400 pA.Since part of the variation could originate from a differ-ence in the development of the cells in culture (Van Eys-eren et al., in preparation), control measurements weremade on a set of five cultures, in which the cells hadbeen exposed to the second culture medium for morethan 10 days (i.e. conditions under which variationsassociated with neuronal differentiation should be lim-ited to a minimum, see above reference). From the 100tested cells, nine gave no response at all, 14 gave aresponse smaller than 10 pA, 50 cells gave a responsebetween 10 and 100 pA, and 27 cells gave a responsebetween 100 and 1000 pA. The amplitude histogram wassimilar for the five cultures: apart from a small peakaround 30–40 pA, the curve fell almost exponentially(Fig. 1(C)). Under these conditions, it was not possible
Fig. 1. (A) Time course of the current induced by a 15 s superfusionof an embryonic cockroach brain neurone in culture with 10−4 M ACh.The recording was made under voltage-clamp conditions using thewhole-cell mode of the patch-clamp technique. The holding potentialwas2 60 mV. In this figure as in the following figures, the horizontalbar above the response indicates the estimated period of continuousagonist application. The turn-on of the current was sigmoı¨dal with atime constant of the order of 0.1 s. The peak current was followed bya slow exponential decay with a time constant of about 5 s, reflectingdesensitization of the ACh receptors. When the superfusing solutionwas switched back to normal, the current returned to zero followingan exponential time course with a time constant of about 0.5 s. (B)Stability of the response to ACh: the same cell was superfused fourtimes with 10−4 M ACh (15 s), the successive superfusion periodsbeing separated by a 2 min gap. Apart from a minor decrease in thepeak response for the third and fourth exposure, the response remainedessentially unchanged. Holding potential:2 60 mV. (C) Amplitudehistogram of the response to 10−4 M ACh of a set of 100 neurones.

231I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
to make a direct comparison between the responses ofdifferent neurones under different experimental con-ditions. Fortunately, it was found that in most cases theresponse of a given neurone to ACh (as well as to otheragonists and antagonists) remained stable for long per-iods (up to 30 min or more). Furthermore, under ourexperimental conditions, recovery from desensitizationwas relatively fast, as illustrated in Fig. 1(B). Underthese conditions, each individual cell could be used asits own control (most usually, the response to a 15 ssuperfusion with 1024 M ACh) before, during and afterthe experiment. This protocol has been used throughoutthe following experiments.
3.2. Effects of cholinergic agonists
The pharmacological profile of the culture neuroneswas determined using a selection of five cholinergicagonists: two purely nicotinic agonists, nicotine and sub-eryldicholine; two mixed agonists, ACh and carbamylch-oline; and one muscarinic agonist, oxotremorine.
Fig. 2(A) illustrates the response of one neurone tosuccessive superfusions with 1024 M concentrations ofeach of these five compounds. ACh gives a typicalresponse with a comparatively large desensitizing phase,nicotine gives a smaller response with a slightly differenttime course. The response to CCh is even smaller, andthe responses to SCh and oxotremorine are very small.This order of potency of the five agonists was typical ofall probed neurones, with the exception of three cellswhich gave an equally large response with ACh and nic-otine. In some cases, the response to oxotremorine wastoo small to be detected. This is illustrated in Fig. 2(B),which shows the mean peak current induced by 1024 Mconcentrations of the same five cholinergic agonists ona set of 37 cells from more than 10 day-old cultures. Fora holding potential of260 mV, the mean currents andSEM were 115.8± 14.5 pA for acetylcholine (ACh),42.7 ± 6.5 pA for nicotine (Nic), 42.4± 6.4 for carba-mylcholine (CCh), 9.56± 2.1 for suberyldicholine (SCh)and 9.3± 2.3 for oxotremorine (Oxo), respectively. Theresponse to ACh was significantly larger than theresponses to the other four agonists (P , 0.0004). Theresponses to nicotine and carbamylcholine were signifi-cantly larger than the responses to suberyldicholine andoxotremorine (P , 0.0005). There was no significantdifference between the responses to nicotine and carba-mylcholine nor between suberyldicholine and oxotremo-rine. From these experiments, the pharmacological pro-file of the embryonic neurones can be defined as follows:ACh > > nicotine= carbamylcholine > > suberyldicho-line = oxotremorine.
3.3. Current–voltage relationships and effects of Na-free calcium solutions
The current–voltage relationship of the response of thecultured neurones to ACh and its agonists was studied
Fig. 2. Effects of five cholinergic agonists on embryonic cockroachbrain neurones in culture. (A) Time course of the effects on one neu-rone of 15 s superfusions with 10−4 M acetylcholine (ACh), nicotine(Nic), carbamylcholine (CCh), suberyldicholine (SCh) and oxotremor-ine (Oxo). Holding potential:2 60 mV. (B) Mean peak currentinduced by 10−4 M concentrations of the same five cholinergic agonistson a set of 37 cells from more than 10 day-old cultures. Each cell wasprobed with the five agonists. The mean currents and SEM were 115.8± 14.5 pA for acetylcholine (ACh), 42.7± 6.5 pA for nicotine (Nic),42.4± 6.4 for carbamylcholine (CCh), 9.56± 2.1 for suberyldicholine(SCh) and 9.3± 2.3 for oxotremorine (Oxo), respectively. Theresponse to ACh was significantly larger than the responses to the otherfour agonists (P , 0.0004). The responses to nicotine and carbamylch-oline were significantly larger than the responses to suberyldicholineand oxotremorine (P , 0.0005). Holding potential:2 60 mV.
under steady-state and non-steady-state conditions. Inthe first case, the holding potential was changed beforeeach superfusion, and the size of the response plottedagainst the holding potential value. In the second case,the current–voltage relationship was established byapplying a ramp change in potential between2100 and+ 100 mV during the plateau phase of the response. Thecontribution of the voltage-dependent conductances andthe leak to the total current was measured using the sameramp protocol before and after the application of theagonist, and subtracted from the test current. The resultsobtained with the two techniques were essentially thesame. The current–voltage relationship was linear formembrane potential values between280 and240 mV,and the extrapolated reversal potential was not signifi-cantly different from 0 mV. This is illustrated in Fig.
232 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
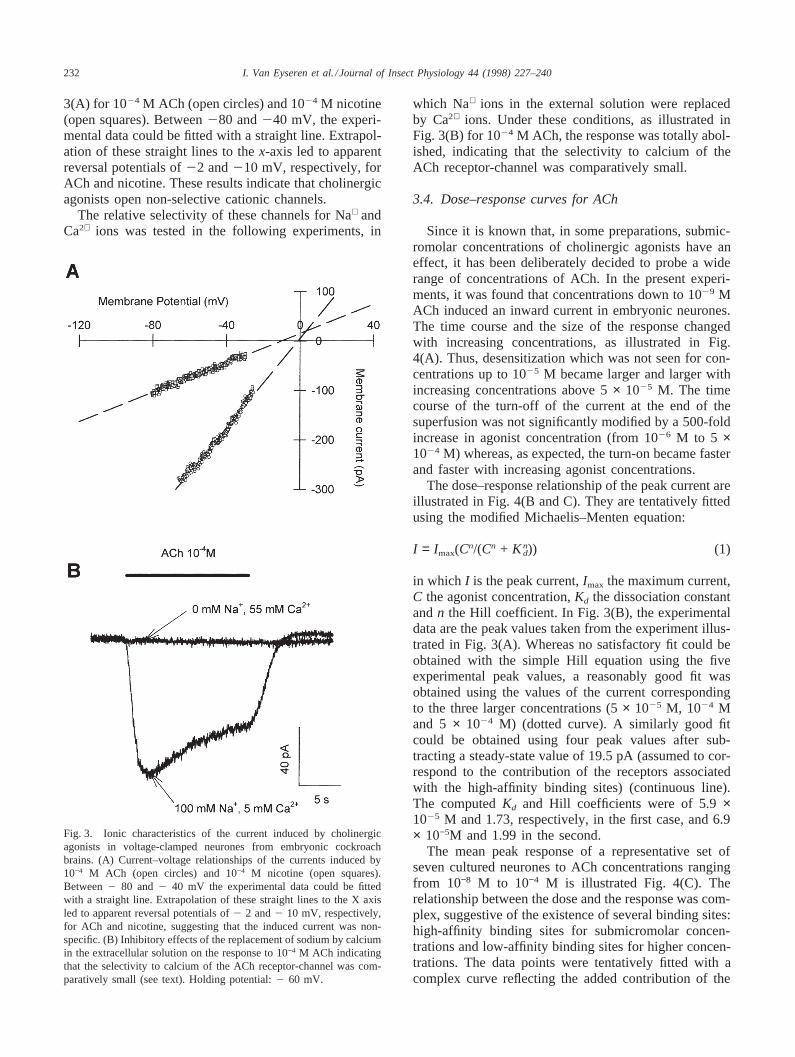
3(A) for 1024 M ACh (open circles) and 1024 M nicotine(open squares). Between280 and240 mV, the experi-mental data could be fitted with a straight line. Extrapol-ation of these straight lines to thex-axis led to apparentreversal potentials of22 and210 mV, respectively, forACh and nicotine. These results indicate that cholinergicagonists open non-selective cationic channels.
The relative selectivity of these channels for Na+ andCa2+ ions was tested in the following experiments, in
Fig. 3. Ionic characteristics of the current induced by cholinergicagonists in voltage-clamped neurones from embryonic cockroachbrains. (A) Current–voltage relationships of the currents induced by10−4 M ACh (open circles) and 10−4 M nicotine (open squares).Between2 80 and2 40 mV the experimental data could be fittedwith a straight line. Extrapolation of these straight lines to the X axisled to apparent reversal potentials of2 2 and2 10 mV, respectively,for ACh and nicotine, suggesting that the induced current was non-specific. (B) Inhibitory effects of the replacement of sodium by calciumin the extracellular solution on the response to 10−4 M ACh indicatingthat the selectivity to calcium of the ACh receptor-channel was com-paratively small (see text). Holding potential:2 60 mV.
which Na+ ions in the external solution were replacedby Ca2+ ions. Under these conditions, as illustrated inFig. 3(B) for 1024 M ACh, the response was totally abol-ished, indicating that the selectivity to calcium of theACh receptor-channel was comparatively small.
3.4. Dose–response curves for ACh
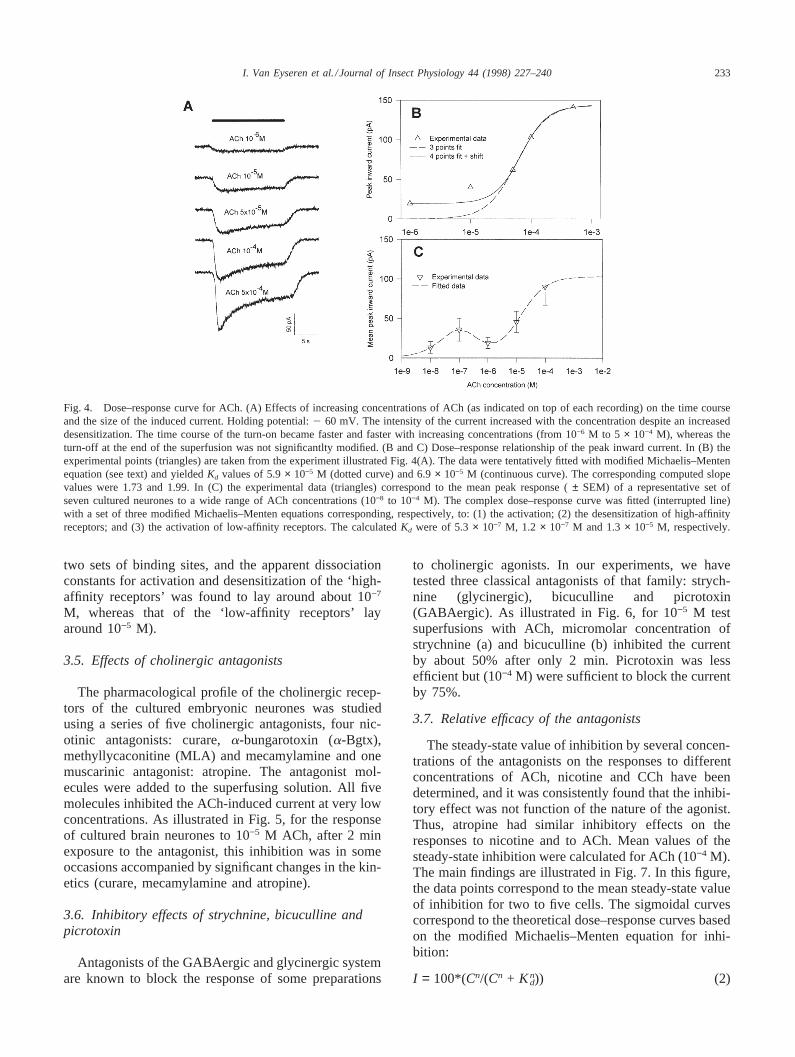
Since it is known that, in some preparations, submic-romolar concentrations of cholinergic agonists have aneffect, it has been deliberately decided to probe a widerange of concentrations of ACh. In the present experi-ments, it was found that concentrations down to 1029 MACh induced an inward current in embryonic neurones.The time course and the size of the response changedwith increasing concentrations, as illustrated in Fig.4(A). Thus, desensitization which was not seen for con-centrations up to 1025 M became larger and larger withincreasing concentrations above 5× 1025 M. The timecourse of the turn-off of the current at the end of thesuperfusion was not significantly modified by a 500-foldincrease in agonist concentration (from 1026 M to 5 ×1024 M) whereas, as expected, the turn-on became fasterand faster with increasing agonist concentrations.
The dose–response relationship of the peak current areillustrated in Fig. 4(B and C). They are tentatively fittedusing the modified Michaelis–Menten equation:
I = Imax(Cn/(Cn + Knd)) (1)
in which I is the peak current,Imax the maximum current,C the agonist concentration,Kd the dissociation constantandn the Hill coefficient. In Fig. 3(B), the experimentaldata are the peak values taken from the experiment illus-trated in Fig. 3(A). Whereas no satisfactory fit could beobtained with the simple Hill equation using the fiveexperimental peak values, a reasonably good fit wasobtained using the values of the current correspondingto the three larger concentrations (5× 1025 M, 1024 Mand 5 × 1024 M) (dotted curve). A similarly good fitcould be obtained using four peak values after sub-tracting a steady-state value of 19.5 pA (assumed to cor-respond to the contribution of the receptors associatedwith the high-affinity binding sites) (continuous line).The computedKd and Hill coefficients were of 5.9×1025 M and 1.73, respectively, in the first case, and 6.9× 10−5M and 1.99 in the second.
The mean peak response of a representative set ofseven cultured neurones to ACh concentrations rangingfrom 10−8 M to 10−4 M is illustrated Fig. 4(C). Therelationship between the dose and the response was com-plex, suggestive of the existence of several binding sites:high-affinity binding sites for submicromolar concen-trations and low-affinity binding sites for higher concen-trations. The data points were tentatively fitted with acomplex curve reflecting the added contribution of the
233I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
Fig. 4. Dose–response curve for ACh. (A) Effects of increasing concentrations of ACh (as indicated on top of each recording) on the time courseand the size of the induced current. Holding potential:2 60 mV. The intensity of the current increased with the concentration despite an increaseddesensitization. The time course of the turn-on became faster and faster with increasing concentrations (from 10−6 M to 5 × 10−4 M), whereas theturn-off at the end of the superfusion was not significantlty modified. (B and C) Dose–response relationship of the peak inward current. In (B) theexperimental points (triangles) are taken from the experiment illustrated Fig. 4(A). The data were tentatively fitted with modified Michaelis–Mentenequation (see text) and yieldedKd values of 5.9× 10−5 M (dotted curve) and 6.9× 10−5 M (continuous curve). The corresponding computed slopevalues were 1.73 and 1.99. In (C) the experimental data (triangles) correspond to the mean peak response (± SEM) of a representative set ofseven cultured neurones to a wide range of ACh concentrations (10−8 to 10−4 M). The complex dose–response curve was fitted (interrupted line)with a set of three modified Michaelis–Menten equations corresponding, respectively, to: (1) the activation; (2) the desensitization of high-affinityreceptors; and (3) the activation of low-affinity receptors. The calculatedKd were of 5.3× 10−7 M, 1.2 × 10−7 M and 1.3× 10−5 M, respectively.
two sets of binding sites, and the apparent dissociationconstants for activation and desensitization of the ‘high-affinity receptors’ was found to lay around about 10−7
M, whereas that of the ‘low-affinity receptors’ layaround 10−5 M).
3.5. Effects of cholinergic antagonists
The pharmacological profile of the cholinergic recep-tors of the cultured embryonic neurones was studiedusing a series of five cholinergic antagonists, four nic-otinic antagonists: curare,a-bungarotoxin (a-Bgtx),methyllycaconitine (MLA) and mecamylamine and onemuscarinic antagonist: atropine. The antagonist mol-ecules were added to the superfusing solution. All fivemolecules inhibited the ACh-induced current at very lowconcentrations. As illustrated in Fig. 5, for the responseof cultured brain neurones to 10−5 M ACh, after 2 minexposure to the antagonist, this inhibition was in someoccasions accompanied by significant changes in the kin-etics (curare, mecamylamine and atropine).
3.6. Inhibitory effects of strychnine, bicuculline andpicrotoxin
Antagonists of the GABAergic and glycinergic systemare known to block the response of some preparations
to cholinergic agonists. In our experiments, we havetested three classical antagonists of that family: strych-nine (glycinergic), bicuculline and picrotoxin(GABAergic). As illustrated in Fig. 6, for 10−5 M testsuperfusions with ACh, micromolar concentration ofstrychnine (a) and bicuculline (b) inhibited the currentby about 50% after only 2 min. Picrotoxin was lessefficient but (10−4 M) were sufficient to block the currentby 75%.
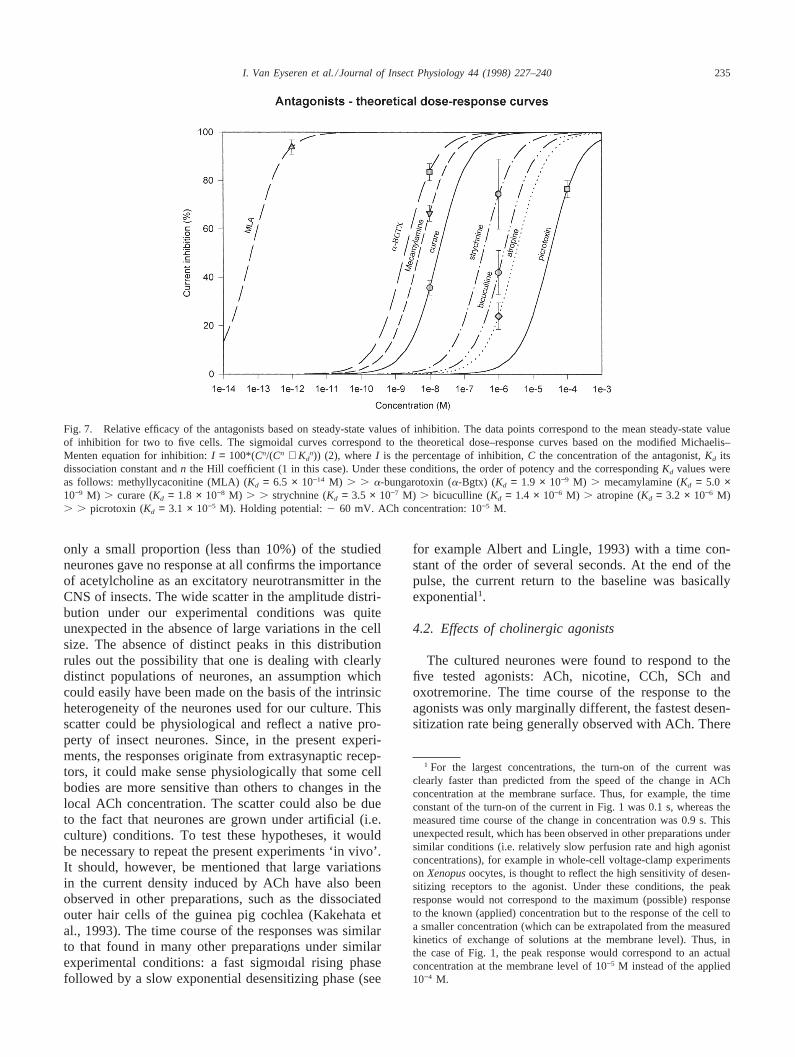
3.7. Relative efficacy of the antagonists
The steady-state value of inhibition by several concen-trations of the antagonists on the responses to differentconcentrations of ACh, nicotine and CCh have beendetermined, and it was consistently found that the inhibi-tory effect was not function of the nature of the agonist.Thus, atropine had similar inhibitory effects on theresponses to nicotine and to ACh. Mean values of thesteady-state inhibition were calculated for ACh (10−4 M).The main findings are illustrated in Fig. 7. In this figure,the data points correspond to the mean steady-state valueof inhibition for two to five cells. The sigmoidal curvescorrespond to the theoretical dose–response curves basedon the modified Michaelis–Menten equation for inhi-bition:
I = 100*(Cn/(Cn + Knd)) (2)

234 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
Fig. 5. Inhibitory effects of cholinergic antagonists (curare,a-bungarotoxin (a-Bgtx), methyllycaconitine (MLA), mecamylamine and atropine)on the response of cultured brain neurones to 10−5 M ACh. The antagonist molecules were added to the superfusing solution. The recordings weretaken 2 min after the addition of the inhibitory molecules. All molecules at the tested concentrations inhibited the ACh current at very lowconcentrations. This inhibition was on some occasions accompanied by changes in the kinetics (curare, mecamylamine and atropine). Holdingpotential:2 60 mV. The vertical scale (current intensity) should read 500 pA for the two recordings in (b) in which the current was exception-ally large.
Fig. 6. Inhibitory effects of glycine (strychnine) and GABAA (bicuculline and picrotoxin) antagonists on the response of cultured brain neuronesto 10−5 M ACh. The inhibitory molecules were added to the superfusing solution. The recordings were taken 2 min after the addition of theinhibitory molecules. Micromolar concentrations of strychnine and bicuculline inhibited the current by 50%. Higher concentrations of picrotoxin(10−4 M) were needed to block. Holding potential:2 60 mV. The 60 pA current scale applies for (a) and (b), the 120 pA for (c).
where I is the percentage of inhibition,C the concen-tration of the antagonist,Kd its dissociation constant andn the Hill coefficient. In the absence of detailed steady-state dose–response curves for all the antagonists, theHill coefficient (n) has been given a default value of 1(an assumption which does not change the order ofpotency but has a small effect on theKd value). Underthese conditions, the order of potency and the corre-spondingKd values were as follows: methyllycaconitine(MLA) ( Kd = 6.5 × 10−14 M) > > a-bungarotoxin (a-Bgtx) (Kd = 1.9 × 10−9 M) > mecamylamine (Kd = 5.0× 10−9 M) > curare (Kd = 1.8 × 10−8 M) > > strychnine(Kd = 3.5 × 10−7 M) > bicuculline (Kd = 1.4 × 10−6 M) >atropine (Kd = 3.2 × 10−6 M) > > picrotoxin (Kd = 3.1× 10−5 M).
4. Discussion
As expected, the present study provides a significantamount of quantitative information concerning theproperties of the cholinergic receptors in embryonic neu-rones ofP. americana.
4.1. Response to acetylcholine
This study clearly demonstrates that, as could be pre-dicted from earlier studies on the insect nervous system,cultured embryonic neurones possess functional recep-tors to ACh. This confirms the previous observations ofBeadle et al. (1982, 1984, 1989), Lees et al. (1983, 1985)and Amar (1991) on the same preparation. The fact that
235I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
Fig. 7. Relative efficacy of the antagonists based on steady-state values of inhibition. The data points correspond to the mean steady-state valueof inhibition for two to five cells. The sigmoidal curves correspond to the theoretical dose–response curves based on the modified Michaelis–Menten equation for inhibition:I = 100*(Cn/(Cn + Kd
n)) (2), whereI is the percentage of inhibition,C the concentration of the antagonist,Kd itsdissociation constant andn the Hill coefficient (1 in this case). Under these conditions, the order of potency and the correspondingKd values wereas follows: methyllycaconitine (MLA) (Kd = 6.5 × 10−14 M) . . a-bungarotoxin (a-Bgtx) (Kd = 1.9 × 10−9 M) . mecamylamine (Kd = 5.0 ×10−9 M) . curare (Kd = 1.8 × 10−8 M) . . strychnine (Kd = 3.5 × 10−7 M) . bicuculline (Kd = 1.4 × 10−6 M) . atropine (Kd = 3.2 × 10−6 M). . picrotoxin (Kd = 3.1 × 10−5 M). Holding potential:2 60 mV. ACh concentration: 10−5 M.
only a small proportion (less than 10%) of the studiedneurones gave no response at all confirms the importanceof acetylcholine as an excitatory neurotransmitter in theCNS of insects. The wide scatter in the amplitude distri-bution under our experimental conditions was quiteunexpected in the absence of large variations in the cellsize. The absence of distinct peaks in this distributionrules out the possibility that one is dealing with clearlydistinct populations of neurones, an assumption whichcould easily have been made on the basis of the intrinsicheterogeneity of the neurones used for our culture. Thisscatter could be physiological and reflect a native pro-perty of insect neurones. Since, in the present experi-ments, the responses originate from extrasynaptic recep-tors, it could make sense physiologically that some cellbodies are more sensitive than others to changes in thelocal ACh concentration. The scatter could also be dueto the fact that neurones are grown under artificial (i.e.culture) conditions. To test these hypotheses, it wouldbe necessary to repeat the present experiments ‘in vivo’.It should, however, be mentioned that large variationsin the current density induced by ACh have also beenobserved in other preparations, such as the dissociatedouter hair cells of the guinea pig cochlea (Kakehata etal., 1993). The time course of the responses was similarto that found in many other preparations under similarexperimental conditions: a fast sigmoı¨dal rising phasefollowed by a slow exponential desensitizing phase (see
for example Albert and Lingle, 1993) with a time con-stant of the order of several seconds. At the end of thepulse, the current return to the baseline was basicallyexponential1.
4.2. Effects of cholinergic agonists
The cultured neurones were found to respond to thefive tested agonists: ACh, nicotine, CCh, SCh andoxotremorine. The time course of the response to theagonists was only marginally different, the fastest desen-sitization rate being generally observed with ACh. There
1 For the largest concentrations, the turn-on of the current wasclearly faster than predicted from the speed of the change in AChconcentration at the membrane surface. Thus, for example, the timeconstant of the turn-on of the current in Fig. 1 was 0.1 s, whereas themeasured time course of the change in concentration was 0.9 s. Thisunexpected result, which has been observed in other preparations undersimilar conditions (i.e. relatively slow perfusion rate and high agonistconcentrations), for example in whole-cell voltage-clamp experimentson Xenopusoocytes, is thought to reflect the high sensitivity of desen-sitizing receptors to the agonist. Under these conditions, the peakresponse would not correspond to the maximum (possible) responseto the known (applied) concentration but to the response of the cell toa smaller concentration (which can be extrapolated from the measuredkinetics of exchange of solutions at the membrane level). Thus, inthe case of Fig. 1, the peak response would correspond to an actualconcentration at the membrane level of 10−5 M instead of the applied10−4 M.

236 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
was no indication of the existence of two distinct compo-nents analogous to those observed by Lapied et al.(1990) on isolated DUM neurones dissociated from thelast abdominal ganglion of the cockroach. The amplitudeof the response was nearly always larger for ACh thanfor the other agonists. The responses to suberyldicholineand oxotremorine were small, when detectable. Theresponses to nicotine and carbamylcholine were signifi-cantly smaller than those to ACh. This pharmacologicalprofile was basically similar for all tested neuronesdespite the above-mentioned large scatter in the ampli-tude of the responses. It differs only slightly from thatestimated by Albert and Lingle (1993) for larvalDroso-phila neurones (ACh = CCh > nicotine >suberyldicholine) but is very different from that derivedfrom microelectrode experiments on the desheathed lastabdominal ganglion of the cockroach by Kerkut et al.(1969a, b) (i.e.: nicotine > carbamylcholine > pilocar-pine > ACh). Although the experiments were done invery different experimental conditions, this differencecould be genuine and reflect either a difference betweenembryonic and adult neurones (the microelectrodeexperiments were made on adult cockroaches), a differ-ence between two populations of neurones under differ-ent experimental conditions, or the effects of glia on theexpression of the receptors (for ‘in situ’ recordings).
The observation that oxotremorine, which is con-sidered to be a typical ‘muscarinic’ agonist, can inducean inward current similar to that induced by suberyldich-oline, a typical ‘nicotinic’ agonist, is of particular interestand illustrates the now generally accepted view that theclassification of the cholinergic receptors into ‘nicotinic’and ‘muscarinic’ (and/or mixed) has to be re-examined.
4.3. Current–voltage relationship and ionic selectivity
The current–voltage relationship was linear for mem-brane potentials between2 80 and2 40 mV, and extra-polation of this linear portion to thex-axis gave anextrapolated reversal potential slightly more negativethan 0 mV. This finding, which is in agreement withprevious observations on a large number of preparations,indicates that the ionic channels activated by ACh andnicotine are non-selective. They also agree with the sin-gle channel data on this same preparation (Beadle et al.,1989). The fact that calcium cannot replace sodium sug-gests that the relative permeability to calcium of thechannels is low2. This conclusion is in apparent contra-diction with several recent reports, showing that theapplication of nicotine or ACh induces an increase in the
2 One cannot eliminate the possibility that high external calcium (55mM in our experiments) has some specific effect on the receptor itself,since it has been shown that neuronala7 nicotinic receptors possessdifferent categories of regulatory calcium binding sites (Galzi et al.,1996).
intraneuronal calcium concentration (Bicker and Kreissl,1994; Bicker, 1996; Grolleau et al., 1996). A plausibleexplanation would be that, as in the chick ciliary gang-lion (Pugh and Berg, 1994), the increase of intracellularcalcium is indirect and results from a two-step process:membrane depolarization (due to the activation of theACh receptors), which in turn induces the activation ofvoltage-gated calcium channels. Indeed, voltage-depen-dent calcium currents have been found in a variety ofinsect neuronal preparations including cockroach brainneurones in culture (Christensen et al., 1988; Amar andPichon, 1992; Tiaho et al., 1996) and cockroach DUMneurones (Wicher and Penzlin, 1994, 1997).
4.4. Dose–response curves
In the present experiments, acetylcholine was foundto induce detectable inward currents for a very widerange of concentrations (from 10−9 to 10−2 M). Thedose–response curves could be dissociated into twocomponents: a ‘high-affinity’ component correspondingto the concentrations ranging from 10−9 and 10−5 M anda ‘low-affinity’ component corresponding to concen-trations ranging from 10−5 to 10−2 M. The first phaseexhibited a maximum for 10−7 M and then dropped forhigher concentrations. The second phase was sigmoı¨dal.Before attempting to analyse these curves in more detail,it should be pointed out that the experimental curve,especially at high agonist concentrations, cannot be afaithful representation of the real dose–response relation-ship due to fast receptor desensitization (which tends toreduce the apparent peak value in a dose-dependentmanner) and to the comparatively slow agonist appli-cation rates which tend to shift the dose–response curvetowards higher concentrations (see above) and to reduceits slope. A similar observation was made by Albert andLingle (1993) for the ACh receptors ofDrosophila lar-val neurones.
In the present experiments, the analysis has beenrestricted to the upper part of the dose–response curve,which corresponds to the range of concentrations usedin most earlier studies (10−5 to 10−3 M). With somerestrictions, the experimental data could be fitted withsigmoidal curves with dissociation constants (Kd)approximating 5× 10−5 M and slopes approximating 2.These values compare well with those obtained byAlbert and Lingle (1993) for larvalDrosophilaneurones(1.7 × 10−5 M and 1.4). The value of the slope suggeststhat, as in other preparations, two molecules of ACh arenecessary to open one channel. The dissociation constantis of the same order of magnitude as that of nicotine onlocust nicotinic receptors (a-L1) expressed inXenopusoocytes (Marshall et al., 1990). On the other hand, ourresults are significantly different from those of Davidand Sattelle (1984) on the Df neurones of the cockroach.Under their experimental conditions, the neurones were

237I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
found to be more sensitive to nicotine than to ACh (evenin the presence of neostigmine) and carbamylcholine(about 100 times) and the dose–response curves had aslope of about 1.
4.5. Effects of cholinergic and non-cholinergicantagonists
The experiments presented in the last part of this paperclearly indicate that a variety of antagonists of the nACh-GABAA-glycine superfamily of receptors are powerfulblockers of the response of cultured embryonic cock-roach neurones to ACh and nicotine. Interestingly, as forthe agonists, inhibition was observed for ‘nicotinic’ aswell as ‘muscarinic’ antagonists, although, in this lastcase, (i.e. atropine with a calculatedKd of 3.2 × 10−6
M), the concentration needed to block the response wassignificantly higher than for the other tested ‘nicotinic’antagonists, which were all efficient in the nanomolar orsubnanomolar range of concentrations. The observationthat GABAA receptor antagonists bicuculline and picro-toxin block the response to ACh in these embryonic neu-rones is reminiscent of the observations of Liu et al.(1994) on the nicotinic receptors of cultured embryonicrat skeletal muscles, and those of Benson (1988, 1992)for isolated neuronal somata from the thoracic ganglionof the locust,Locusta migratoria. An inhibitory effectsof bicuculline on the cholinergic response of insect neu-rones has also been described by Buckingham et al.(1994) for the cell body of the giant interneurone 2 (GI2) in the sixth abdominal ganglion of the cockroach. Inthis last case, a 50% block of the ACh-induced currentwas obtained for 10−4 M bicuculline (i.e. 100 times morethan in the present experiments). Strychnine, a knownantagonist of the glycine receptors, has been shown toinhibit the effects of acetylcholine on the (atypical) nic-otinic receptors of the outer hair cells of the cochlea(Bartolami et al., 1993), as well as the ‘nicotinic’ (ACh1)response of neuronal somata isolated from locust tho-racic ganglia (Benson, 1992).
Compared with the previously published electrophysi-ological data, the present experiments indicate that theACh receptors of the embryonic cockroach brain neu-rones are very sensitive to a wide range of antagonists.Of particular interest are the sensitivities to methyllyca-conitine (where the calculatedKd: 6.5 × 10−14 M is inthe subpicomolar range of concentration) anda-bungar-otoxin (calculatedKd: 1.9 × 10−9 M). Similarly lowvalues have been obtained in rat brain and locust(Schistocerca gregaria) ganglia for the binding of125I-a-bungarotoxin (Macallan et al., 1988). One to 1000 fMconcentrations of methyllycaconitine were found todecrease the frequency of anatoxin-induced single chan-nel openings in foetal rat hippocampal neurones(Alkondon et al., 1992) and, in this same preparation,type IA nicotinic ACh receptors were found to be
blocked by 1 nM methyllycaconitine and 10 nMa-Bgtx(Alkondon and Albuquerque, 1993). A similarly highsensitivity to methyllycaconitine (IC50 in the picomolarrange) was observed for reconstituted homomerica7receptors inXenopusoocytes (Palma et al., 1996).
The ranking order of potency of the various antagon-ists in cockroach embryonic neurones (i.e. methyllyca-conitine (Kd = 6.5 × 10−14 M) > > a-bungarotoxin (a-Bgtx) (Kd = 1.9 × 10−9 M) > mecamylamine (Kd = 5.0× 10−9 M) > curare (Kd = 1.8 × 10−8 M) > > strychnine(Kd = 3.5 × 10−7 M) > bicuculline (Kd = 1.4 × 10−6 M) >atropine (Kd = 3.2 × 10−6 M) > > picrotoxin (Kd = 3.1× 10−5 M)) is almost identical, at least qualitatively, tothat reported by David and Sattelle (1984) for the Dfneurones of the same species (a-Bgtx > mecamylam-ine > d-tubocurarine= atropine) and consistent with thatreported by Benson (1992) for the ACh1 response ofthe locust (a-Bgtx > strychnine > mecamylamine > d-tubocurarine > picrotoxin= bicuculline= atropine). Withthe exception of strychnine which was 10 times moreefficient in the locust than in the embryonic cockroachneurones (2.3× 10−8 M against 3.5× 10−7 M) and picro-toxin which was equipotent in the two preparations (2.9× 10−5 M and 3.1 × 10−5 M), the cultured cockroachneurones were consistently 10 times more sensitive thanthe locust neurones. These differences which areunlikely to be artifactual (differences in the methodsused to calculate theKd (or IC50) values) could reflectspecies differences or differences between embryonicand adult neurones. An even larger difference in thevalues of concentrations can thus be observed betweenembryonic neurones and adult Df neurones of the samespecies where the IC50 were of 6.4× 10−8 M for a-Bgtx,2.5 × 10−6 M for mecamylamine, 8.0× 1025 M for d-tubocurarine and 1.0× 10−4 M for atropine, respectively(David and Sattelle, 1984). The sequence of inhibitionof the response to ACh of larvalDrosophila neurones(Albert and Lingle, 1993) is qualitatively similar to thatreported for the other insect preparations (the responseof the Drosophila neurones to 20mM ACh was abol-ished by 10−8 M a-Bgtx, inhibited by about 60% (at260 mV) by 10−5 M curare and by more than 50% by 2× 10−7 M mecamylamine).
4.6. Molecular properties of the receptors
As indicated in the Introduction, the purpose of thepresent experiments was to provide some new clues onthe molecular properties of the ACh receptors in a simplepreparation. The present experiments clearly show thatthese receptors are able to bind a wide variety of mol-ecules ranging from the ‘classical’ nicotinic agonists andantagonists to some ‘classical’ muscarinic agonists andGABAA and glycinergic antagonists. This result, whichconfirms some earlier observations, indicates that thebinding site(s) of the receptor(s) is much less specific

238 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
than previously thought. Although it has been shown thatthe membrane of these cultured neurones possesses twodistinct populations of ACh-activated channels withclearly different single channel conductances (Beadle etal., 1989), the time course of the whole-cell responseand its pharmacological profile does not enable a cleardistinction to be made between these two populations.It can be argued that the single channel data, which havebeen obtained in the cell-attached configuration, corre-spond to a small residual population of channels whichdo not desensitize during long exposures to the agonists.Single channel recordings with a fast superfusion systemin the outside–out configuration of the patch-clamp tech-nique are necessary to clarify this point. Another possi-bility would be to analyse the whole-cell response tovery low concentrations of agonists as shown by Albertand Lingle (1993) forDrosophila neurones or to usefluctuation analysis. Unfortunately, in both cases, theanalysis of the experimental data will be complicated bythe presence of clusters of channels (see Beadle et al.,1989) and the non-linearity of the response due to thedesensitization of the receptors.
Quite unexpectedly, despite the heterogeneous originof the neurones used to start the cultures, the responsesto the various agonists and antagonists were very homo-geneous and it was not possible to make a distinctionbetween pharmacologically distinct groups of neurones.On the other hand, the complex shape of the dose–response curve to ACh strongly suggests that these neu-rones possess at least two (and most probably three) dif-ferent types of binding sites with clearly different affin-ities. In the absence of data concerning the dose–response curve for the other agonists, it is premature toconclude but it can be tentatively proposed that the partof the dose–response curve between 10−9 and 10−5 Mcorresponds to a population of receptors on which AChacts as an agonist at the lower concentrations but almostcompletely desensitizes for the larger concentrations.The upper part of the dose–response curve (between1025 and 10−2 M) corresponds to the more ‘classical’low-affinity receptor exhibiting fast desensitization. Asfor most other ACh receptors, the slope of the dose–response curve for the low-affinity receptors is signifi-cantly higher than 1, suggesting that two molecules ofACh are needed to open one channel.
As discussed earlier, the speed of our perfusion sys-tem was too slow to enable a proper analysis of the kin-etics of the binding and unbinding reactions of the agon-ists on the receptor. A faster superfusion systemassociated with a cooling of the preparation and the useof smaller concentrations will enable us to overcomethis difficulty.
The molecular structure of the ACh receptors remainsto be elucidated. Their high sensitivity toa-Bgtx sug-gests that they belong to thea7–a9 family of receptors,which are known to form homomeric ACh receptors
characterized by fast desensitization and a high calciumpermeability and an unusual pharmacological profile(Elgoyhen et al., 1994; Gerzanich et al., 1994). Thiswould fit with the idea that embryonic insect receptorsare more primitive than vertebrate receptors. However,it has recently been shown that cDNAs isolated from thenervous system ofDrosophila ALS, Da2 and ARDcould not form functional receptors when expressed inXenopusoocytes unless a chickb2 structural subunitwas coinjected (Bertrand et al., 1994). When homo-oligomeric channels can be formed as in the case ofaL1(the locust a-like nicotinic receptor subunit) theirproperties are different from those of native receptors:they exhibit a very low sensitivity to nicotine, activatewith a very slow kinetics and show no tendency todesensitize (Amar et al., 1995). It therefore seems morereasonable to assume that, as in the vertebrate nervoussystem, the receptors are hetero-oligomeric structures(most probably pentamers) containinga andb subunits.
Acknowledgements
The authors wish to thank Dr J. Benson for his com-ments on an earlier versions of this paper. They wish toacknowledge D. De Saint Jan and X. Durant Castel fortheir contributions at different stages of the work and tothank P. Cadic for his technical assistance. I.V.E. wassupported by a research grant from the Re´gion Bretagne.
References
Albert, J.L., Lingle, C.J., 1993. Activation of nicotinic acetylcholinereceptors on culturedDrosophilaand other insect neurones. Journalof Physiology, London 463, 605–630.
Alkondon, M., Albuquerque, E.X., 1993. Diversity of nicotinic acetyl-choline receptors in rat hippocampal neurons. I. Pharmacologicaland functional evidence for distinct structural subtypes. Journal ofPharmacology and Experimental Therapeutics 265 (3), 1455–1473.
Alkondon, M., Pereira, E.F., Wonnacott, S., Albuquerque, E.X., 1992.Blockade of nicotinic currents in hippocampal neurons definesmethyllycaconitine as a potent and specific receptor antagonist.Molecular Pharmacology 41 (4), 802–808.
Amar, M. Etude biophysique et pharmacologique des canaux ioniquesde la membrane de neurones en culture d’un insecte, la blatte, Per-iplaneta americana. The`se de Doctorat de L’Universite´ Pierre etMarie Curie, 1991, pp. 1–452.
Amar, M., Pichon, Y., 1992. Calcium currents in cultured cockroachneurones. Journal of Physiology, London 446, 171P.
Amar, M., Thomas, P., Wonnacott, S., Lunt, G.G., 1995. A nicotinicacetylcholine receptor subunit from insect brain forms a non-desen-sitising homo-oligomeric nicotinic acetylcholine receptor whenexpressed inXenopus oocytes. Neuroscience Letters 199 (2),107–110.
Bartolami, S., Ripoll, C., Eybalin, M., 1993. Anticholinertgic effectsof strychnine on the cochlea do not involve muscarinic receptors.Neuroreport 4 (8), 1003–1006.
Beadle, D.J., Beadle, C.A., Pichon, Y., Shimahara, T. 1985. Patch-clamp and noise analysis studies of cholinergic properties of cul-

239I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
tured cockroach neurones.J. Physiol., London 371, 145P(Abstract).
Beadle, D.J., Hicks, D. Insect nerve cell culture, in: G.A. Kerkut andL.I. Gilbert Eds, Comprehensive Insect Physiology, Biochemistryand Pharmacology Pergamon Press, Oxford, 1985, pp. 181–211.
Beadle, D.J., Hicks, D., Middleton, C., 1982. Fine structure of neu-rones from embryonicPeriplaneta americanagrowing in long-termculture. Journal of Neurocytology 11, 611–626.
Beadle, D.J., Lees, G., Botham, R.P. Cholinergic neurones in neuronalcultures from Periplaneta americana, in: A.B. Borkovec, T.J. Kelly(Eds), Insect Neurochemistry and Neurophysiology New York: Ple-num Press. 1984 pp. 317–320.
Beadle, D.J., Horseman, G., Pichon, Y., Amar, M., Shimahara, T.,1989. Acetylcholine-activated ion channels in embryonic cock-roach neurones growing in culture. Journal of ExperimentalBiology 142, 337–355.
Benson, J.A., 1988. Bicuculline blocks the response to acetylcholineand nicotine but not to muscarine or GABA in isolated insect neu-ronal somata. Brain Research 458, 45–71.
Benson, J.A., 1992. Electrophysiological pharmacology of the nic-otinic and muscarinic cholinergic responses of isolated neuronalsomata from locust thoracic ganglia. Journal of ExperimentalBiology 170, 203–233.
Bertrand, D., Ballivet, M., Gomez, M., Bertrand, S., Phannavong, B.,Gundelfinger, E.D., 1994. Physiological properties of neuronal nic-otinic receptors reconstituted from the vertebrate beta 2 subunit andDrosophila alpha subunits. European Journal of Neurosciences 6,869–875.
Bicker, G., 1996. Transmitter-induced calcium signalling in culturedneurones of the insect brain. Journal of Neuroscience Methods 69,33–41.
Bicker, G., Kreissl, S., 1994. Calcium imaging reveals nicotinic acetyl-choline receptors on cultured mushroom body neurons. Journal ofNeurophysiology 71 (2), 808–810.
Buckingham, S.D., Hue, B., Sattelle, D.B., 1994. Actions of bicucul-line on cell body and neuropilar membranes of identified insectneurones. Journal of Experimental Biology 186, 235–244.
Callec, J.J. Synaptic transmission in the central nervous system ofinsects, in: J.E. Treherne (Ed.), Insect Neurobiology Amsterdam,North Holland, 1974, pp. 119–178.
Callec, J.J. Synaptic transmission in the central nervous system, in:G.A. Kerkut, L.I. Gilbert (Eds), Comprehensive Insect Physiology,Biochemistry and Pharmacology. Pergamon Press, Oxford, 1985,vol. 5, pp. 139–179.
Chen, J.S., Levi-Montalcini, R., 1970a. Axonal growth from insectneurones in glia-free cultures. Proceedings of the National Acad-emy of Sciences, USA 66, 32–39.
Chen, J.S., Levi-Montalcini, R., 1970b. Long-term cultures of dis-sociated nerve cells from the embryonic nervous system of thecockroachPeriplaneta americana. Archives Italiennes de Biologie108, 503–537.
Christensen, B.N., Larmet, Y., Shimahara, T., Beadle, D.J., Pichon,Y., 1988. Ionic currents in neurones cultured from embryonic cock-roach (Periplaneta americana) brains. Journal of ExperimentalBiology 135, 193–214.
David, J.A., Sattelle, D.B., 1984. Actions of cholinergic pharmacologi-cal agents on the cell body membrane of the fast coxal depressormotoneurone of the cockroach (Periplaneta americana). Journal ofExperimental Biology 108, 119–136.
Elgoyhen, A.B., Johnson, D.S., Boulder, J., Vetter, D.E., Heinemann,S., 1994. Alpha 9: an acetylcholine receptor with novel pharmaco-logical properties expressed in rat cochlear hair cells. Cell 79 (4),705–715.
Galzi, J.L., Bertrand, S., Corringer, P.J., Changeux, J.P., Bertrand, D.,1996. Identification of calcium binding sites that regulate potenti-ation of a neuronal nicotinic acetylcholine receptor. EMBO Journal15, 5824–5832.
Gerzanich, V., Anand, R., Lindstrom, J., 1994. Homomers of alpha 8and alpha 7 subunits of nicotinic receptors exhibit similar channelbut contrasting binding site properties. Molecular Pharmacology445 (2), 212–220.
Grolleau, F., Lapied, B., Buckingham, S.D., Mason, W.T., Sattelle,D.B., 1996. Nicotine increases [Ca2 + ]i and regulates electricalactivity in insect neurosecretory cells (DUM neurons) via an acetyl-choline receptor with ‘mixed’ nicotinic–muscarinic pharmacology.Neuroscience Letters 220 (2), 142–146.
Hamill, O.P., Marty, A., Neher, E., Sakmann, B., Sigworth, F., 1981.Improved patch-clamp techniques for high-resolution current rec-ording from cells and cell-free membrane patches. Pflu¨gersArchives 391, 85–100.
Harrison, J.B., Leech, C.A., Katz, J., Sattelle, D.B., 1990. Embryonicand adult neurones of the housefly (Musca domestica) in culture.Tissue and Cell 22, 337–347.
Hanke, W., Breer, H., 1986. Channel properties of an insect neuronalacetylcholine receptor protein reconstituted in planar lipid bilayers.Nature, London 321, 171–174.
Kakehata, S., Nakagawa, T., Takasaka, T., Akaike, N., 1993. Cellularmechanism of acetylcholine-induced response in dissociated outerhair cells of guinea-pig cochlea. Journal of Physiology, London463, 227–244.
Kerkut, G.A., Pitman, R.M., Walker, R.J., 1969a. Sensitivity of neu-rones of the insect central nervous system to iontophoreticallyapplied acetylcholine or GABA. Nature 222, 1075–1076.
Kerkut, G.A., Pitman, R.M., Walker, R.J., 1969b. Iontophoretic appli-cation of acetylcholine and GABA onto insect central neurones.Comparative Biochemistry and Physiology 31, 611–633.
Lapied, B., Le Corronc, H., Hue, B., 1990. Sensitive nicotinic andmixed nicotinic–muscarinic receptors in insect neurosecretory cells.Brain Research 533, 132–136.
Leech, C.A., Sattelle, D.B., 1992. Multiple conductances of neuronalnicotinic acetylcholine receptors. Neuropharmacology 31, 501–507.
Lees, G., Beadle, D.J., Botham, R.P., 1983. Cholinergic receptors oncultured neurones from the central nervous system of embryoniccockroaches. Brain Research 288, 49–59.
Lees, G., Beadle, D.J., Botham, R.P., Kelly, J.S., 1985. Excitableproperties of insect neurones in culture: a developmental study.Journal of Insect Physiology 31, 135–144.
Liu, Q.Y., Dunlap, V., Barker, J.L., 1994. Gamma-aminobutyric acidtype A receptor antagonists picrotoxin and bicuculline alter acetyl-choline channel kinetics in cultured embryonic rat skeletal muscle.Molecular Pharmacology 46, 1197–1203.
Macallan, D.R., Lunt, G.G., Wonnacott, S., Swanson, K.L., Rapoport,H., Albuquerque, E.X., 1988. Methyllycaconitine and (+)-anatoxin-a differentiate between nicotinic receptors in vertebrate and invert-ebrate nervous systems. FEBS Letters 226 (2), 357–363.
Marshall, J., Buckingham, S.D., Shingai, R., Lunt, G.G., Goosey,M.W., Darlison, M.G., Sattelle, D.B., Barnard, E.A., 1990.Sequence and functional expression of a singlea-subunit of aninsect nicotinic acetylcholine receptor. EMBO Journal 9 (13),4391–4398.
Palma, E., Bertrand, S., Binzoni, T., Bertrand, D., 1996. Neuronal nic-otinic alpha 7 receptor excpressed inXenopusoocytes presents fiveputative binding sites for methyllycaconitine. Journal of Physi-ology, London 491, 151–161.
Pichon, Y., Manaranche, R. Biochemistry of the nervous system. InG.A. Kerkut and L.I. Gilbert (Eds), Comprehensive Insect Physi-ology, Biochemistry and Pharmacology, Pergamon Press, Oxford.1985, vol. 10, pp. 417–450.
Pichon, Y. The pharmacology of the insect nervous system. In M.Rockstein (Ed.), The Physiology of Insecta. Academic Press, NewYork, 1974, pp. 101–174.
Pichon, Y. Comparative Molecular Neurobiology. Birkau¨ser Verlag,Basel, 1993, pp. 1–433.

240 I. Van Eyseren et al. / Journal of Insect Physiology 44 (1998) 227–240
Pugh, P.C., Berg, D.K., 1994. Neuronal acetylcholine receptors thatbind alpha-bungarotoxin mediate neurite retraction in a calcium-dependent manner. Journal of Neurosciences 14 (2), 889–896.
Sattelle, D.B., Sun, Y.A., Wu, C.F., 1986. Neuronal acetylcholinereceptor: patch-clamp recording of single channel properties fromdissociated insect neurones. IRCS Medical Sciences 14, 65–66.
Tiaho, F., Charrier, N., Le Guen, J., Pichon, Y., 1996. Calcium currentin developing neurones from cockroach brains in primary culture.Biology of the Cell 86, 194.
Wicher, D., Penzlin, H., 1994. Ca2 + currents in cockroach neurones:properties and modulation by neurohormone D. Neuroreport 5 (9),1023–1026.
Wicher, D., Penzlin, H., 1997. Ca2 + currents in central insect neurons:electrophysiological and pharmacological properties. Journal ofNeurophysiology 77 (1), 186–199.
















![Mapping muscarinic receptors in human and baboon brain using [N-11C-methyl]-benztropine](https://static.fdokumen.com/doc/165x107/6344f35df474639c9b049d90/mapping-muscarinic-receptors-in-human-and-baboon-brain-using-n-11c-methyl-benztropine.jpg)



