
Fish in the Filipino Diet Fish & Fish products Fish & Fish products
Upload
independentCategory
view
2download
0
Effects of Fish Oil on the Central Nervous System: A NewPotential Antidepressant?
K. NALIWAIKOa, R.L.F. ARAUJOa, R.V. DA FONSECAa, J.C. CASTILHOa, R. ANDREATINIa, M.I. BELLISSIMOa,B.H. OLIVEIRAb, E.F. MARTINSc, R. CURIc, L.C. FERNANDESd and A.C. FERRAZa,*
aLaboratorio de Fisiologia e Farmacologia do Sistema Nervoso Central, Departamento de Fisiologia e Farmacologia, Universidade Federal do Parana,81.531-990 -Curitiba, PR, Brazil; bLaboratorio de Produtos Naturais, Departamento de Quımica, Universidade Federal do Parana, Curitiba, PR, Brazil;cLaboratorio Fisiologia Celular, Departamento de Fisiologia e Biofısica, ICB/ USP, Sao Paulo, SP, Brazil; dLaboratorio de Metabolismo Celular,Departamento de Fisiologia, Universidade Federal do Parana, Curitiba, PR, Brazil
(Received 17 January 2004; Revised 4 March 2004; In final form 28 March 2004)
In the last 100 years major depression has increasedworldwide. In this study we provided coconut fat (CF,rich in saturated fatty acids) or fish oil (FO, rich in n-3polyunsaturated fatty acids) to female rats throughoutpregnancy and lactation and then to their offspring post-weaning and examined lipid brain profile and thepossible effect of FO as antidepressant agent in theoffspring in adulthood (F1). Rats were submitted toforced swimming test, elevated plus maze, Morris watermaze and open field. Peroxidation rate in the cerebralcortex and hippocampus were measured. Docosahexa-enoic acid (DHA) concentration in dam’s milk, eicosa-pentaenoic acid (EPA) and DHA concentration inhippocampus and cerebral cortex from F1 rats FOsupplemented increased significantly when comparedto control (C) and CF rats. Arachidonic acid/EPA ratio inthe cerebral cortex and hippocampus decreased in ratssubmitted to forced swimming test. Peroxidation ratewere not different between the groups. Immobility timein the forced swimming test in FO group was reducedðp < 0:01Þ when compared to C and CF rats. We concludethat lifelong intake of FO was able to induce anantidepressant effect with EPA and DHA concentrationincreased in the cerebral cortex and hippocampus.
Keywords: Cerebral function; Depression; Fish oil; Forcedswimming; Polyunsaturated fatty acids
INTRODUCTION
The brain has high concentration of complex lipids,which will determine the structural and functionalproperties of its cellular and subcellular membranes(Farooqui et al., 2000; Das, 2003). Docosahexaenoic acid
(22:6n-3, DHA) and arachidonic acid (20:4n-6, AA)are the most important polyunsaturated fatty acids(PUFAs) in neuronal membranes, but the major PUFAspresent is usually DHA (Broadhurst et al., 2002).The precursors of these PUFAs are a-linolenic acid(18:3n-3, LNA) and linoleic acid (18:2n-6, LA),respectively, which cannot be synthesized de novo bymammals, and are therefore, considered as essentialfatty acids. DHA and AA are incorporated by cellmembranes during development and adult age, withbrain and retina requesting more DHA than othertissues (Jimenez et al., 1997). DHA plays animportant role on brain metabolism influencingcognitive performance, visual acuity, neurotransmit-ters metabolism and neural development (Jimenezet al., 1997; Carlson and Neuringer, 1999; Chalon et al.,2001). Long-term deficiency in LNA leads to changesin brain fatty acid composition causing loss oflearning ability (Innis, 2000) and during neuronaldevelopment could be responsible for memoryimpairment or neurological disease in adult(Moriguchi et al., 2000; Youdim et al., 2000). Modifi-cations of the brain membrane fatty acid (FA)composition occur with supplementation of variousdietary oils. If the lipid profiles in diet are not suitableto maintain cerebral needs, the cells will replace DHAby other PUFAs, changing brain functions (Fernstrom,1999; Carlson, 2001).
Amount and type of FA in the diet has beenreported to be involved with some disease develop-ment (Simopoulos, 2002). In fact, modern Westerndiet in the past 100 years has changed significantly,
ISSN 1028-415X print/ISSN 1476-8305 online q 2004 Taylor & Francis Ltd
DOI: 10.1080/10284150410001704525
*Corresponding author. Tel.: þ55-41-361-1722. Fax: þ55-41-266-2042. E-mail: [email protected]
Nutritional Neuroscience, Volume 7 Number 2 (April 2004), pp. 91–99

which lowered the ratio of omega-3 to omega-6 toabout 1:25 (Simopoulos et al., 1999). The correctbalance of omega-3 and omega-6 fatty acids withinphospholipids is ultimately essential for normalneuronal function. In addition, during the last 100years, the prevalence of major depression hasincreased worldwide (Hibbeln, 1998). There areevidences that alterations in FA metabolism and thecomposition of phospholipids in the serum andmembranes are involved in the development ofmajor depression (Hibbeln et al., 1996; Maes et al.,1999). Epidemiological studies have shown thatplaces with higher rates of omega-3 consumption,where fish is a core of the diet, have lower rates ofdepressive disorders (Tanskanen et al., 2001).In countries such as Japan and Germany, prevalencerates of depression are 0.12 and 5%, respectively. Thishas been associated with the annual fish consump-tion diet which is close to 150 lbs per person in Japanand less than 30 lbs per person in Germany (Hibbeln,1998). This author found a correlation of fishconsumption in different countries with the preva-lence of major depression showing a highlysignificant decrease of depression with increasingfish consumption. In addition, fish oil (FO) has beenused as mood stabilizer or adjunctive therapy indepression, bipolar disorders and schizophrenia(Stoll et al., 1999; Nemets et al., 2002).
Most previous studies investigating the effect ofdietary fatty acids on major depression have usedhuman beings (Peet and Horrobin, 2002; Puri et al.,2002). There are few reports on the effect of dietaryfatty acids in laboratory animals on majordepression. These studies have used young adultanimals fed for a short period before or afterinduction of the depression (Hillakivi-Clarke et al.,1996; Raygada et al., 1998). A more likely scenario isthat a dietary pattern will be lifelong and may existfrom conception. We are not aware of anystudies that have investigated the effect of lifelongconsumption of particular fatty acids on suscepti-bility to depression in adulthood. Therefore, inthis study we provided coconut fat (CF, rich insaturated fatty acids) or FO (rich in n-3 PUFA) tofemale rats throughout pregnancy and lactation andthen to their offspring post-weaning and examinedthe possible effect as antidepressant agent in theoffspring in adulthood.
MATERIAL AND METHODS
Animals and Study Design
All studies involving animals were approved by theUniversity Federal of Parana Committee of AnimalWelfare. Female Wistar rats adults were kept in atemperature-controlled room ð22 ^ 28CÞ on a 12/12 h
light/dark cycle (lights on 07:00 h). The animals weredivided into three groups. One received a normalchow diet (C) ad libitum, and the other two were fedchow but were also supplemented orally with eitherCF or FO, kindly donated by Herbarium Foundation(Curitiba, PR, Brazil). The FO used was a mixedmarine triacylglycerol preparation containing 180 geicosapentaenoic acid (EPA) and 120 g DHA per kg(Table I). The oils were supplemented at a level of3 g/kg body weight per day and were provided as asingle bolus using a pipette during 15 days, whenthey were 90 days old. Then they were mated withmale Wistar rats that had been fed on normallaboratory chow. Throughout gestation and lactationthe females continued to receive CF or FO, accordingto the same supplementation protocol. After wean-ing (age 21 days) the male offspring (F1) received thesame diet and supplementation as their mothers,until adult age (90 days), when they were submittedto different behavior tests as described below.
Forced Swimming Test
The procedure was done as described by Beijaminiet al. (1998), a modified method of Porsolt et al. (1978).Rats were placed in glass aquarium ð20 £ 20 £
40 cm3Þ containing 15 cm deep water ð24 ^ 18CÞ for15 min followed by a 5 min retest session 24 h later.Immobility time was recorded during the testsession. The rat was judged immobile wherever itstopped swimming and remained floating in thewater, with its head just above water level. The waterwas changed for each rat after test session to avoidinfluence of alarm substance (Abel and Belitzke,1990). Following the test, the animals were dried in awarm enclosure.
Morris Water Maze
A black circular tank ð160 cm diameter £ 50 cm deepÞwas filled with water ð22 ^ 18CÞ to a depth of 30 cm.A transparent acrylic platform ð10 £ 10 cm2Þ waslocated in one of four quadrants, 1 cm below
TABLE I Fatty acid composition (%) of the diets given to rats
Fatty acids Fish oil Coconut fat
Fatty acid composition (g/100 g total fatty acids)Lauric acid 12:0 0.50 61.04Miristic acid 14:0 11.45 18.93Palmitic acid 16:0 14.19 5.57Estearic acid 18:0 3.16 2.62Oleic acid 18:1 (n-9) 8.69 7.04Linoleic acid 18:2 (n-6) 0.20 2.86Linolenic acid 18:3 (n-3) 1.74 0.50Araquidonic acid 20:4 (n-6) 12.11 –Eicosapentaenoic acid 20:5 (n-3) 26.01 1.42Docosahexaenoic acid 22:6 (n-3) 20.19 –
K. NALIWAIKO et al.92

the water surface. All tests were recorded by a videocamera system located in the roof (Morris, 1984).
Place Learning
During six consecutive days the rats were submittedto four trials per day. For each trial, the rat wasplaced in water facing the tank wall at one of fourpossible starting locations (North, South, East orWest). In the room, there were extra-maze cued,including posters on the walls, lights and theexperimenter, who always stood at the sameposition. The rat used these cued to develop aspatial map and locate the hidden platform. Thelatency to find the platform was recorded. If the ratdid not find the platform after 60 s, it was gently puton the platform and placed there for 20 s. After that,the rat was placed in a cage out side the water mazefor 30 s and then placed in a new start point. Thisprotocol was repeated for six days. On the seventhday, the platform was removed from the tank anddid the probe trial. The rat was placed in thequadrant opposite to that the platform and it swamduring the 60 s.
Working-memory
One week after the probe trial, animals weresubmitted to working-memory test during fiveconsecutive days, with four trials per day (DosSantos, 1999). The rat was placed in the tank facingthe wall in one of four start points. Latency wasrecorded in each trial. If the rat did not find theplatform after 60 s, it was gently put on the platformand placed there for 30 s. After that, rat was placedin a new start point. The hidden platform wasplaced in a different quadrant each day.
Elevated Plus Maze
The elevated plus maze consists of two oppositeopen arms and two opposite enclose arms ð45 £
15 cm2Þ: The arms are connected by a central squareð15 £ 15 cm2Þ; forming a plus shape. The maze iselevated 70 cm above the floor in a room. This testhas been widely used to measure anxiety orexploratory behavior (File and Zangrossi, 1993; Fileand Gonzales, 1996). The rat was placed in thecentral square and its behavior was observed for5 min.
Open-field
The open-field apparatus consists in a circular arena(1 m diameter), the floor is divided into central andperipheral units (Kelly, 1993). The rat was placedindividually in the center of the open-field. Theanimal’s behavior was evaluated during 5 min
and locomotion (number of units crossed) wasrecorded. After that, the open-field was washedwith water–alcohol (10%) solution, to avoid bias dueto odors or residues left by the rats tested earlier.
Fatty Acid Composition in the Diet, Milk, CerebralCortex and Hippocampus
Total lipids were extracted from diet, dam’s milk andfrom cerebral cortex and hippocampus tissue of F1generation using chloroform–methanol (2:1 vol/vol)according to Folch et al. (1957). Fatty acids werederivatized with 4-bromomethyl-7-coumarin andthen separated on a Shimadzu LC-10A highperformance liquid chromatograph using an octa-decylsilica column (25 cm £ 4:6 mm i.d.; particle size5mm). Fatty acids were resolved isocratically using amobile phase of acetonitrile–water (gradient from77:23 to 90:10 vol/vol) at a flow rate of 1 ml/min.Fatty acid derivatives were detected by fluorescence(325-nm excitation; 395-nm emission).
Determination of the Products of LipidPeroxidation
Measurement of Stable Products of LipidPeroxidation by Thiobarbituric Acid Assay
Aldehyde products of lipid peroxidation weremeasured using thiobarbituric acid (TBA), followingmethod described by Brown and Kelly (1996). Briefly,brain tissue was extracted in phosphate buffer(0.1 M), pH 7.4. Then, 0.5 ml was mixed with 0.5 mlTBA (1% in NaOH 50 mM), 0.5 ml of HCl 25% and0.045 ml of butylated hydroxytoluene (2% in metha-nol). The samples were then heated in a boiling waterbath for 10 min. After cooling, 1.5 ml of butanol wasadded to the mixture and centrifuged at 3000 rpm(Eppendorf 5810 R) for 20 min and the absorbance ofthe supernatant was read at 532 nm. The concen-tration of malondialdehyde (MDA) was calculatedfrom TBA-reactive material using the extinctioncoefficient of 1:56 £ 105 M21 cm21 given for theMDA-TBA adduct.
Lipid Hydroperoxide Measurement
The products of lipid peroxidation were measured aslipid hydroperoxides following the method describedby Jiang et al. (1991). Briefly, brain tissue washomogenized in phosphate buffer (0.1 M), pH 7.4.One hundred microliters was added to 900ml of thefollowing reaction mixtures and incubated for 30 minat room temperature prior to measurement at560 nm: 100mM of xylenol orange, 25 mM of H2SO4,250mM of FeSO4, and 4 mM of butylated hydroxy-toluene, in 90% (v/v) methanol. The reactionis essentially complete after 15 min and the color is
ANTIDEPRESSANT EFFECT OF FISH OIL 93
stable overnight at room temperature. The extinctioncoefficient of the Fe3þ-xylenol orange complex ð1:5 £
104 M21 cm21Þ was used to calculate oxidation offerrous by hydroperoxide.
Statistical Analysis
Data are presented as mean ^ SEM or median ^
semi-interquartile range when indicated. Kruskall–Wallis ANOVA followed by post-hoc Dunn testevaluated the data of the forced swimming test.The place of learning and working memory in theMorris water maze were examined by two-wayanalysis of variance (ANOVA) followed by Duncantest. Elevated plus maze, open field, fatty acidscomposition and products of lipid peroxidationwere analyzed by one-way ANOVA followedby post-hoc Newman – Keuls. Differences wereconsidered statistically significant when p , 0:05:
RESULTS
Fatty Acid Composition
Milk
The fatty acid composition of the DHA in the dam’smilk differed according to diet (Table II). Dam’s milkfrom rats fed with FO contained higher proportionsof DHA than those from rats fed the control diet orsupplemented with CF ðp , 0:001Þ: Unsaturarionindex in FO was about 10% higher when comparedto C and CF groups.
Cerebral Cortex
The fatty acid composition of cerebral cortex lipidsalso differed according to diet (Table III). Cerebral
cortex from animals supplemented with FO con-tained higher proportions of EPA and DHA thanthose from rats fed the control diet or supplementedwith CF ðp , 0:001Þ: The AA – EPA ratio ofcerebral cortex lipids decreased in the orderC (0.35) . CF (0.25) . FO (0.09). The proportion ofsaturated fat (except 14:0) and linoleic acid weresignificantly lower in FO group when comparedto C and CF animals. Unsaturation index inFO group was approximately 2.4-fold greater thanC and CF groups.
Hippocampus
The fatty acid composition of hippocampus lipidsalso differed according to diet (Table IV). Hippo-campus from animals supplemented with FOcontained higher proportions of EPA and DHA
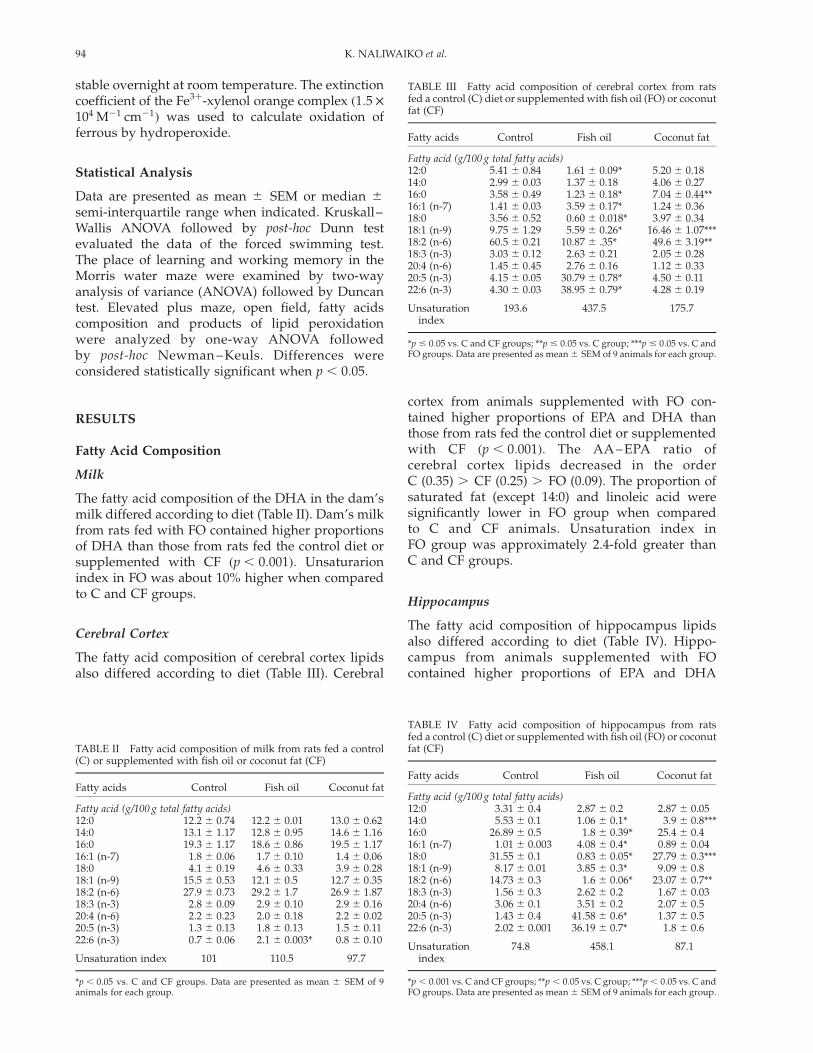
TABLE II Fatty acid composition of milk from rats fed a control(C) or supplemented with fish oil or coconut fat (CF)
Fatty acids Control Fish oil Coconut fat
Fatty acid (g/100 g total fatty acids)12:0 12.2 ^ 0.74 12.2 ^ 0.01 13.0 ^ 0.6214:0 13.1 ^ 1.17 12.8 ^ 0.95 14.6 ^ 1.1616:0 19.3 ^ 1.17 18.6 ^ 0.86 19.5 ^ 1.1716:1 (n-7) 1.8 ^ 0.06 1.7 ^ 0.10 1.4 ^ 0.0618:0 4.1 ^ 0.19 4.6 ^ 0.33 3.9 ^ 0.2818:1 (n-9) 15.5 ^ 0.53 12.1 ^ 0.5 12.7 ^ 0.3518:2 (n-6) 27.9 ^ 0.73 29.2 ^ 1.7 26.9 ^ 1.8718:3 (n-3) 2.8 ^ 0.09 2.9 ^ 0.10 2.9 ^ 0.1620:4 (n-6) 2.2 ^ 0.23 2.0 ^ 0.18 2.2 ^ 0.0220:5 (n-3) 1.3 ^ 0.13 1.8 ^ 0.13 1.5 ^ 0.1122:6 (n-3) 0.7 ^ 0.06 2.1 ^ 0.003* 0.8 ^ 0.10
Unsaturation index 101 110.5 97.7
*p , 0:05 vs. C and CF groups. Data are presented as mean ^ SEM of 9animals for each group.
TABLE III Fatty acid composition of cerebral cortex from ratsfed a control (C) diet or supplemented with fish oil (FO) or coconutfat (CF)
Fatty acids Control Fish oil Coconut fat
Fatty acid (g/100 g total fatty acids)12:0 5.41 ^ 0.84 1.61 ^ 0.09* 5.20 ^ 0.1814:0 2.99 ^ 0.03 1.37 ^ 0.18 4.06 ^ 0.2716:0 3.58 ^ 0.49 1.23 ^ 0.18* 7.04 ^ 0.44**16:1 (n-7) 1.41 ^ 0.03 3.59 ^ 0.17* 1.24 ^ 0.3618:0 3.56 ^ 0.52 0.60 ^ 0.018* 3.97 ^ 0.3418:1 (n-9) 9.75 ^ 1.29 5.59 ^ 0.26* 16.46 ^ 1.07***18:2 (n-6) 60.5 ^ 0.21 10.87 ^ .35* 49.6 ^ 3.19**18:3 (n-3) 3.03 ^ 0.12 2.63 ^ 0.21 2.05 ^ 0.2820:4 (n-6) 1.45 ^ 0.45 2.76 ^ 0.16 1.12 ^ 0.3320:5 (n-3) 4.15 ^ 0.05 30.79 ^ 0.78* 4.50 ^ 0.1122:6 (n-3) 4.30 ^ 0.03 38.95 ^ 0.79* 4.28 ^ 0.19
Unsaturationindex
193.6 437.5 175.7
*p # 0:05 vs. C and CF groups; **p # 0:05 vs. C group; ***p # 0:05 vs. C andFO groups. Data are presented as mean ^ SEM of 9 animals for each group.
TABLE IV Fatty acid composition of hippocampus from ratsfed a control (C) diet or supplemented with fish oil (FO) or coconutfat (CF)
Fatty acids Control Fish oil Coconut fat
Fatty acid (g/100 g total fatty acids)12:0 3.31 ^ 0.4 2.87 ^ 0.2 2.87 ^ 0.0514:0 5.53 ^ 0.1 1.06 ^ 0.1* 3.9 ^ 0.8***16:0 26.89 ^ 0.5 1.8 ^ 0.39* 25.4 ^ 0.416:1 (n-7) 1.01 ^ 0.003 4.08 ^ 0.4* 0.89 ^ 0.0418:0 31.55 ^ 0.1 0.83 ^ 0.05* 27.79 ^ 0.3***18:1 (n-9) 8.17 ^ 0.01 3.85 ^ 0.3* 9.09 ^ 0.818:2 (n-6) 14.73 ^ 0.3 1.6 ^ 0.06* 23.07 ^ 0.7**18:3 (n-3) 1.56 ^ 0.3 2.62 ^ 0.2 1.67 ^ 0.0320:4 (n-6) 3.06 ^ 0.1 3.51 ^ 0.2 2.07 ^ 0.520:5 (n-3) 1.43 ^ 0.4 41.58 ^ 0.6* 1.37 ^ 0.522:6 (n-3) 2.02 ^ 0.001 36.19 ^ 0.7* 1.8 ^ 0.6
Unsaturationindex
74.8 458.1 87.1
*p , 0:001 vs. C and CF groups; **p , 0:05 vs. C group; ***p , 0:05 vs. C andFO groups. Data are presented as mean ^ SEM of 9 animals for each group.
K. NALIWAIKO et al.94
than those from rats fed the control diet orsupplemented with CF ðp , 0:001Þ: The AA-EPAratio of hippocampus lipids decreased in the order C(2.13) . CF (1.15) . FO (0.08). The proportion ofsaturated fat (except 12:0) and linoleic acidwere significantly lower in FO group whencompared to C and CF animals. Unsaturation indexin FO group was approximately 5.6-fold greater thanC and CF groups.
Determination of the Products of LipidPeroxidation
The TBARs and lipid hydroperoxides content in thecerebral cortex and hippocampus was not differentbetween the groups ðp . 0:05Þ; which was about 5.0and 4.5mmol/mg protein and 133 and 117 mmol/mgprotein, respectively.
Forced Swimming Test
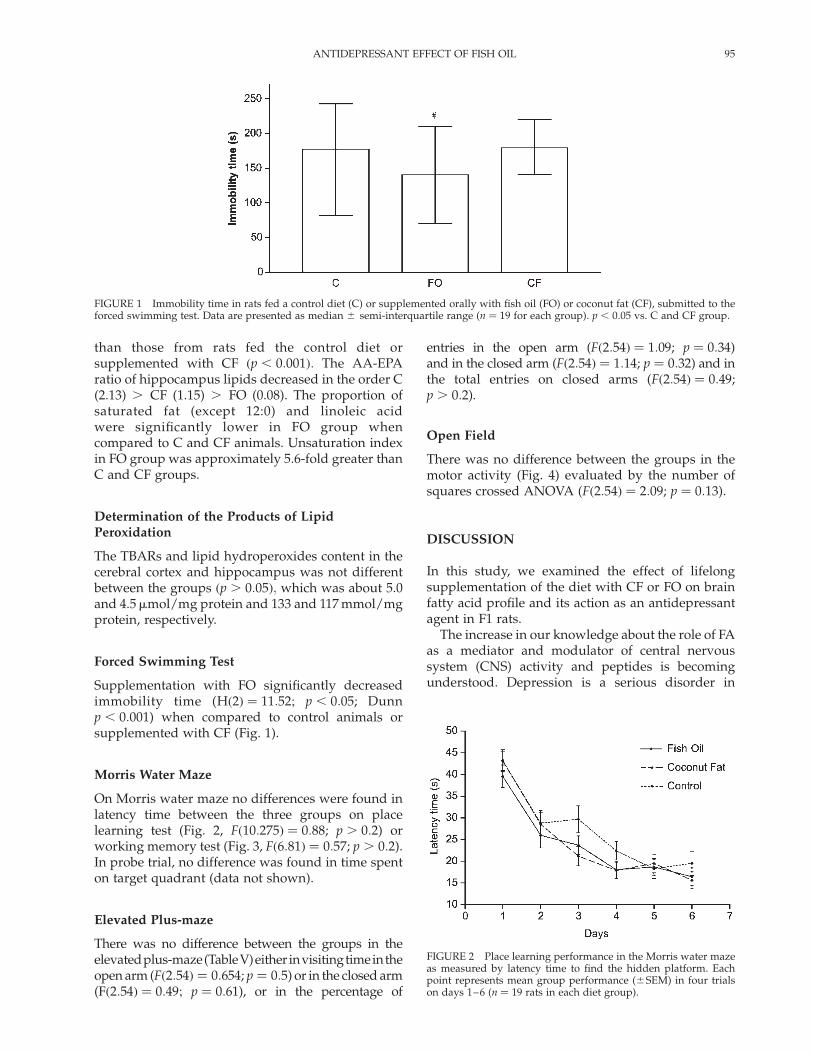
Supplementation with FO significantly decreasedimmobility time (Hð2Þ ¼ 11:52; p , 0:05; Dunnp , 0:001) when compared to control animals orsupplemented with CF (Fig. 1).
Morris Water Maze
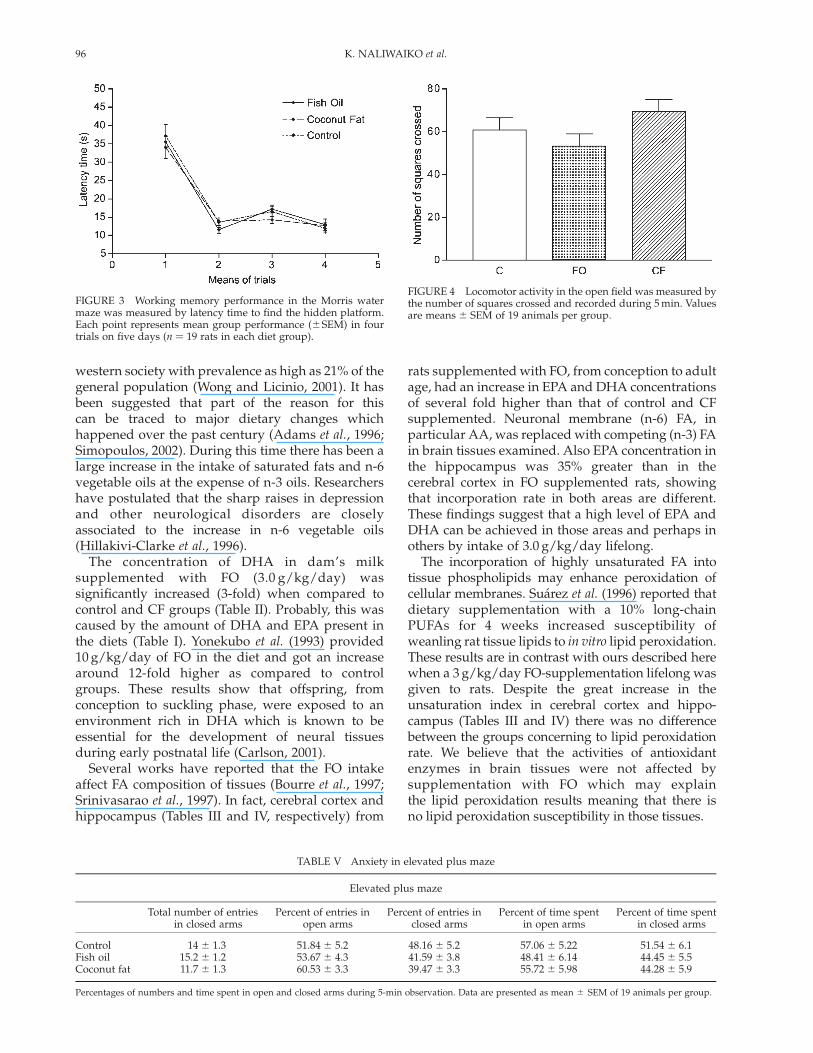
On Morris water maze no differences were found inlatency time between the three groups on placelearning test (Fig. 2, Fð10:275Þ ¼ 0:88; p . 0:2) orworking memory test (Fig. 3, Fð6:81Þ ¼ 0:57; p . 0:2).In probe trial, no difference was found in time spenton target quadrant (data not shown).
Elevated Plus-maze
There was no difference between the groups in theelevatedplus-maze(TableV)either invisitingtimeintheopen arm (Fð2:54Þ ¼ 0:654; p ¼ 0:5) or in the closed arm(Fð2:54Þ ¼ 0:49; p ¼ 0:61), or in the percentage of
entries in the open arm (Fð2:54Þ ¼ 1:09; p ¼ 0:34)and in the closed arm (Fð2:54Þ ¼ 1:14; p ¼ 0:32) and inthe total entries on closed arms (Fð2:54Þ ¼ 0:49;p . 0:2).
Open Field
There was no difference between the groups in themotor activity (Fig. 4) evaluated by the number ofsquares crossed ANOVA (Fð2:54Þ ¼ 2:09; p ¼ 0:13).
DISCUSSION
In this study, we examined the effect of lifelongsupplementation of the diet with CF or FO on brainfatty acid profile and its action as an antidepressantagent in F1 rats.
The increase in our knowledge about the role of FAas a mediator and modulator of central nervoussystem (CNS) activity and peptides is becomingunderstood. Depression is a serious disorder in
FIGURE 1 Immobility time in rats fed a control diet (C) or supplemented orally with fish oil (FO) or coconut fat (CF), submitted to theforced swimming test. Data are presented as median ^ semi-interquartile range (n ¼ 19 for each group). p , 0:05 vs. C and CF group.
FIGURE 2 Place learning performance in the Morris water mazeas measured by latency time to find the hidden platform. Eachpoint represents mean group performance (^SEM) in four trialson days 1–6 (n ¼ 19 rats in each diet group).
ANTIDEPRESSANT EFFECT OF FISH OIL 95
western society with prevalence as high as 21% of thegeneral population (Wong and Licinio, 2001). It hasbeen suggested that part of the reason for thiscan be traced to major dietary changes whichhappened over the past century (Adams et al., 1996;Simopoulos, 2002). During this time there has been alarge increase in the intake of saturated fats and n-6vegetable oils at the expense of n-3 oils. Researchershave postulated that the sharp raises in depressionand other neurological disorders are closelyassociated to the increase in n-6 vegetable oils(Hillakivi-Clarke et al., 1996).
The concentration of DHA in dam’s milksupplemented with FO (3.0 g/kg/day) wassignificantly increased (3-fold) when compared tocontrol and CF groups (Table II). Probably, this wascaused by the amount of DHA and EPA present inthe diets (Table I). Yonekubo et al. (1993) provided10 g/kg/day of FO in the diet and got an increasearound 12-fold higher as compared to controlgroups. These results show that offspring, fromconception to suckling phase, were exposed to anenvironment rich in DHA which is known to beessential for the development of neural tissuesduring early postnatal life (Carlson, 2001).
Several works have reported that the FO intakeaffect FA composition of tissues (Bourre et al., 1997;Srinivasarao et al., 1997). In fact, cerebral cortex andhippocampus (Tables III and IV, respectively) from
rats supplemented with FO, from conception to adultage, had an increase in EPA and DHA concentrationsof several fold higher than that of control and CFsupplemented. Neuronal membrane (n-6) FA, inparticular AA, was replaced with competing (n-3) FAin brain tissues examined. Also EPA concentration inthe hippocampus was 35% greater than in thecerebral cortex in FO supplemented rats, showingthat incorporation rate in both areas are different.These findings suggest that a high level of EPA andDHA can be achieved in those areas and perhaps inothers by intake of 3.0 g/kg/day lifelong.
The incorporation of highly unsaturated FA intotissue phospholipids may enhance peroxidation ofcellular membranes. Suarez et al. (1996) reported thatdietary supplementation with a 10% long-chainPUFAs for 4 weeks increased susceptibility ofweanling rat tissue lipids to in vitro lipid peroxidation.These results are in contrast with ours described herewhen a 3 g/kg/day FO-supplementation lifelong wasgiven to rats. Despite the great increase in theunsaturation index in cerebral cortex and hippo-campus (Tables III and IV) there was no differencebetween the groups concerning to lipid peroxidationrate. We believe that the activities of antioxidantenzymes in brain tissues were not affected bysupplementation with FO which may explainthe lipid peroxidation results meaning that there isno lipid peroxidation susceptibility in those tissues.
FIGURE 3 Working memory performance in the Morris watermaze was measured by latency time to find the hidden platform.Each point represents mean group performance (^SEM) in fourtrials on five days (n ¼ 19 rats in each diet group).
TABLE V Anxiety in elevated plus maze
Elevated plus maze
Total number of entriesin closed arms
Percent of entries inopen arms
Percent of entries inclosed arms
Percent of time spentin open arms
Percent of time spentin closed arms
Control 14 ^ 1.3 51.84 ^ 5.2 48.16 ^ 5.2 57.06 ^ 5.22 51.54 ^ 6.1Fish oil 15.2 ^ 1.2 53.67 ^ 4.3 41.59 ^ 3.8 48.41 ^ 6.14 44.45 ^ 5.5Coconut fat 11.7 ^ 1.3 60.53 ^ 3.3 39.47 ^ 3.3 55.72 ^ 5.98 44.28 ^ 5.9
Percentages of numbers and time spent in open and closed arms during 5-min observation. Data are presented as mean ^ SEM of 19 animals per group.
FIGURE 4 Locomotor activity in the open field was measured bythe number of squares crossed and recorded during 5 min. Valuesare means ^ SEM of 19 animals per group.
K. NALIWAIKO et al.96

Many studies in animal models of depressioninvestigating drugs treatment and criteria for theirevaluation have been established, but few studieshave approached alteration in the diet and depression(Wainwright et al., 1994; Raygada et al., 1998).
Our results show that rats chronically sup-plemented with FO for one generation decreasedsignificantly immobility time when compared tocontrol and CF groups suggesting that FO was ableto cause an antidepressant effect in rats (Fig. 1). Theforced swimming test is used to evaluate anantidepressant effect of a substance preclinically(Porsolt et al., 1978). However, this test alone can notbe used to exclude other factors that could cause afalse positive test. Hence we submit the rats to tasksto assess possible behavioral alterations such asanxiety, motor activity and learning which couldinterfere in the results found in forced swimmingtest. Anxiety can trigger the behavior despair.Animals with increased motor activity would showa reduced immobility time. An impairment learningmeans that the animals will not remember thetraining session which happens 24 h earlier, leadingto increased mobility and consequently reducedimmobility time. In the open field (Fig. 4), watermaze (Figs. 2 and 3) and elevated plus maze tests(Table V) all groups present the same behavior,showing that FO supplementation has no effect onthis behaviors. Other studies found the same result,which corroborate our data (Wainwright et al., 1994;Wainright et al., 1999; Arterburn et al., 2000; Carrieet al., 2000; Wilde et al., 2002). These behavior testswere important because they give support to suggestthat the results shown in forced swimming test werecaused by FO effect upon CNS.
Cerebral cortex and hippocampus are areasinvolved in learning and depression, respectively(Spedding et al., 2003). Morphologic and morpho-metric analysis of the hippocampus in depressedpatients have revealed structural alterations (Sheline2000). FO supplementation increased significantlyEPA and DHA concentration in both areas. Work inhumans reported that lower concentration of longchain n-3 PUFAs in cellular membranes is associatedwith the appearance of depression and that n-3PUFAs supplementation would be efficient toalleviate the depression symptoms (Hibbeln andSalen, 1995; Edwards et al., 1998; Peet et al., 1998).Alterations in phospholipids and cholesterol of thecell membranes in the brain may induce changes inthe fluidity/viscosity and consequently, in manyneurotransmitter systems thought to be related todepression development, e.g. serotonin (Engelberg,1992; Yadid et al., 2000) and noradrenaline (Anandand Charney, 2000). There are findings that AA–EPAratio in serum cholesteryl and phospholipids arehigher in major depression subjects (Adams et al.,1996; Maes et al., 1996). Our results show that
the AA–EPA ratio in the cerebral cortex andhippocampus (Tables III and IV, respectively) wasdecreased in rats submitted to forced swimming test(Fig. 1).
The mechanisms by which FO induce antidepres-sant effect are unknown and were not investigatedhere. However, we suggest that FO may increasenumber of serotonin receptors and the level ofreceptor binding because it has been shown thata-linolenic acid (18:3 n-3), the parent acid of EPA andDHA, is able to do it (Bergelson, 1995; Rego andOliveira, 1995; Delion et al., 1996;). In addition, onemay take into consideration the content of DHA andits relation to the molecular activity of Na^K^
ATPase in the membrane bilayer. Up to 60% ofenergy consumption in the brain is linked to theNa^K^ ATPase enzyme. This may representa fundamental relationship, underlying metabolicactivity, but it may also represent the link betweenreduced levels of PUFAs and neurological dysfunc-tion as the majority of energy consumption in thebrain is linked to the Na^K^ ATPase enzyme (Turneret al., 2003). Both hypotheses must be tested.
The results presented herein led us to concludethat the lifelong intake of FO was able to induce anantidepressant effect with EPA and DHA concen-tration increased in the cerebral cortex andhippocampus.
Acknowledgements
This work was supported by a grant from Pronex-CNPq and FUNPAR.
References
Abel, E.L. and Bilitzke, J. (1990) “A possible alarm substance inthe forced swimming test”, Physiology and Behavior 48,233–239.
Adams, P.B., Lawson, S., Sanigorski, A. and Sinclair, A.J. (1996)“Arachidonic acid to eicosapentaenoic acid ratio in bloodcorrelates positively with clinical symptoms of depression”,Lipids 31, S157–S161.
Anand, A. and Charney, D.S. (2000) “Norepinephine dysfunctionin depression”, Journal of Clinical Psychiatry 61(10), 16–24.
Aronson, W.J., Glapsy, J.A., Reddy, S.T., Reese, D., Heber, D. andBagga, D. (2001) “Modulation of omega-3/omega-6 poly-unsaturated ratios with dietary fish oils in men with protatecancer”, Urology 58, 283–288.
Arterburn, L.M., Boswell, K.D., Koskelo, E.K., Kassner, S.L., Kelly, C.and Kyle, D.J. (2000) “A combined subchornic (90-day)toxicity and neurotoxicity study of a single-cell source ofdocosahexaenoic acid triglyceride (DHASCOw)”, Food andChemical Toxicology 38, 35–49.
Beijamini, V., Skalisz, L.L., Joca, S.R.L. and Andreatini, R. (1998)“The effect of oxcarbazepine on behavioral despair andlearned helplessness”, European Journal of Pharmacology 347,23–27.
Bergelson, L.D. (1995) “Dynamic lipid heterogeneity and receptorevents”, Molecular Membrane Biology 12, 125–129.
Bourre, J-M.E., Dumont, O.L., Piciotti, M.J., Clement, M.E. andDurand, G.A. (1997) “Comparison of vegetable and fish oil inthe provision of N-3 polyunsaturated fatty acids for nervoustissue and selected organs”, Nutritional Biochemistry 8,472–478.
ANTIDEPRESSANT EFFECT OF FISH OIL 97

Broadhurst, C.L., Wang, Y., Crawford, M.A., Cunnane, S.C.,Parkington, J.E. and Schimidt, W.F. (2002) “Brain-specificlipids from marine, lacustrine, or terrestrial food resources:potential impact on early African Homo sapiens”, ComparativeBiochemistry and Physiology part b 131, 653–673.
Brown, R.K. and Kelly, F.J. (1996) “Peroxides and other products”,In: Punchard, N.A. and Kelly, F.J., eds, Free Radicals (OxfordUniversity Press Inc., New York), pp 119–131.
Carlson, S.E. (2001) “Docosahexainoic acid and arachidonic acid ininfant development”, Seminars in Neonatology 6, 437–449.
Carlson, S.E. and Neuringer, M. (1999) “Polyunsaturated fatty acidstatus and neurodevelopment: a summary and criticalanalysis of the literature”, Lipids 34, 171–178.
Carrie, I., Clement, M., de Javel, D., Frances, H. and Bourre, J-M.(2000) “Phospholipid supplementation reverses behavioraland biochemical alterations induced by n-3 polyunsaturatedfatty acid deficiency in mice”, Journal of Lipid Research 41,473–480.
Chalon, S., Vancassel, S., Zimmer, L., Guilloteau, D. and Durand, G.(2001) “Polyunsaturated fatty acids and cerebral function: focuson monoaminergic neurotransmission”, Lipids 36, 937–944.
Das, U.N. (2003) “Long-chain polyunsaturated fatty acids in thegrowth and development of the brain and memory”, Nutrition19, 62–65.
Delion, S., Chalon, S., Guilloteau, D., Besnard, J.C. and Durand, G.(1996) “a-linolenic acid dietary deficiency alters age-relatedchanges of dopaminergic and serotoninergic neurotrans-mission in the rat frontal cortex”, Journal of Neurochemistry 66,1582–1591.
Dos Santos, A.M.G. (1999) “Aprendizagem e memoria no labirintoaquatico de Morris”, In: Xavier, G.F., ed., Tecnicas para o estudodo sistema nervoso (Pleiade, Sao Paulo) 1, pp 135–158.
Edwards, R., Peet, M., Shay, J. and Horrobin, D. (1998) “Omega-3polyunsaturated fatty acid levels in the diet and in red bloodcell membranes of depressed patients”, Journal of AffectiveDisorders 48, 149–155.
Engelberg, H. (1992) “Low serum cholesterol and suicide”, Lancet339, 727–729.
Farooqui, A.A., Horrocks, L.A. and Farooqui, T. (2000) “Glycero-phospholipids in brain: their metabolism, incorporation intomembranes, function, and involvement in neurologicaldisorders”, Chemistry and Physics of Lipids 106, 1–29.
Fernstrom, J.D. (1999) “Effects of dietary polyunsaturated fattyacids on neuronal function”, Lipids 34, 161–169.
File, S.E. and Gonzales, L.E. (1996) “Anxiolytic effects in the plus-maze of 5-HT1A receptor ligands in dorsal raphe and ventralhippocampus”, Pharmacology Biochemistry and Behavior 54,123–128.
File, S.E., Zangrossi, H., Jr., Viana, M. and Graeff, F.G. (1993) “Trial2 in the elevated plus-maze: a different form of fear?”,Psycopharmacology (berl.) 111, 491–494.
Folch, J., Lees, M. and Stanley, G.H.S. (1957) “A simple method forthe isolation and purification of total lipid from animaltissues”, Journal of Biological Biochemistry 226, 497.
Hibbeln, J.R. (1998) “Fish consumption and major depression”,Lancet 351, 1213.
Hibbeln, J.R. and Salen, N., Jr. (1995) “Dietary polyunsaturatedfatty acids and depression: when cholesterol does not satisfy”,American Journal of Clinical Nutrition 62, 1–9.
Hibbeln, J.R., Umnau, J.C., George, D.T. and Salem, N., Jr. (1996)“Do plasma polyunsaturates predict hostility and violence?”,World Review of Nutritional Dietetics 82, 175–186.
Hillakivi-Clarke, I., Onojafe, I. and Cho, E. (1996) “High-fat dietinduces aggressive behavior in male mice and rats”, LifeScience 58, 1653–1660.
Innis, S.M. (2000) “The role of dietary n-6 and n-3 fatty acids inthe developing brain”, Developmental Neuroscience 22,474–480.
Jiang, Z-Y., Woollard, A.C.S. and Wolff, S.P. (1991) “Lipidhydroperoxide measurement by oxidation of Fe2þ in thepresence of xylenol orange. Comparison with the TBA assayand an iodometric method”, Lipids 26, 853–856.
Jimenez, J., Boza, J., Suarez, M.D. and Gil, A. (1997) “The effect of aformula supplemented with n-3 and n-6 long-chain poly-unsaturated fatty acids on plasma phospholipid, livermicrossomal, retinal, and brain fatty acid composition inneonatal piglets”, Nutritional Biochemistry 8, 217–223.
Kelly, A.E. (1993) “Locomotor activity and exploration”, In: Sahgal,A., ed., Behavioral Neuroscience: a practical approach (OxfordUniversity Press, Oxford) 2, pp 1–21.
Maes, M., Smith, R., Christophe, A., Cosyns, P., Desnyder, R.and Meltzer, H. (1996) “Fatty acid composition in majordepression: decreased v-3 fractions in cholesteryl esters andincreased C20: 4v6 / C20:5v3 ratio in cholesteryl estersand phospholipids”, Journal of Affective Disorders 38, 35–46.
Maes, M., Christophe, A., Delange, J., Altamura, C., Neels, H. andMeltzer, H.Y. (1999) “Lowered omega-3 polyunsaturated fattyacids in serum phospholipids and cholesteryl esters ofdepressed patients”, Psychiatry Research 85, 275–291.
Maidment, I.D. (2000) “Are fish oil an effective therapy in mentalillness—an analysis of the data”, Acta Psychiatrica Scandinavica102, 3–11.
Moriguchi, T., Greiner, R.S. and Salem, N., Jr. (2000) “Behavioraldeficits associated with dietary induction of decreased braindocosahexaienoic acid concentration”, Journal of Neurochem-istry 75, 2563–2573.
Morris, R.G.M. (1984) “Developments of a water-maze procedurefor studying spatial learning in the rat”, Journal of NeuroscienceMethods 11, 47–60.
Nemets, B., Stahl, Z. and Belmaker, R.H. (2002) “Addition ofomega-3 fatty acid to maintenance medication treatment forrecurrent unipolar depressive disorder”, American Journal ofPsychiatry 159, 477–479.
Peet, M. and Horrobin, D.F. (2002) “A dose-ranging study of theeffects of eicosapentaenoate in patients with ongoingdepression despite apparently adequate treatment withstandard drugs”, Archives of General Psychiatry 59, 913–919.
Peet, M., Murphy, B., Shay, J. and Horobin, D. (1998) “Depletion ofomega-3 fatty acids levels in red blood cell membranes ofdepressive patients”, Society of Biological Psychiatry 43,315–319.
Porsolt, R.D., Anton, G., Blavet, N. and Jalfre, M. (1978)“Behavioral despair in rats: a new model sensitive toantidepressant treatments”, European Journal of Pharmacology47, 379–391.
Puri, B.K., Counsell, S.J., Richardson, A.J. and Horrobin, D.F.(2002) “Eicosapentaenoic acid in treatment-resistantdepression”, Archives of General Psychiatry 59, 91–92.
Raygada, M., Cho, E. and Hilakivi-Clarke, L. (1998) “Highmaternal intake of polyunsaturated fatty acids duringpregnancy in mice alters offsprings’ aggressive behavior,immobility in the swimming test, locomotor activity andbrain protein kinase C activity”, Journal of Nutrition 128,2505–2511.
Rego, A.C. and Oliveira, C.R. (1995) “Influence of lipid peroxi-dation on [3H] ketanserin binding to 5-HT2 prefrontal cortexreceptors”, Neurochemistry International 27, 489–496.
Sheline, Y.I. (2000) “3D MRI studies of neuroanatomic changes inunipolar major depression: the role of stress and medicalcomorbidity”, Biological Psychiatry 48, 791–800.
Simopoulos, A.P. (2002) “The importance of the ratio of omega-6/omega-3 essential fatty acids”, Biomedicine and Pharmaco-therapy 56, 365–379.
Simopoulos, A.P., Leaf, A. and Salem, N., Jr. (1999) “Essentialityof and recommended dietary intakes for omega-6 andomega-3 fatty acids”, Annals of Nutrition and Metabolism 43,127–130.
Spedding, M., Neau, I. and Harsing, L. (2003) “Brain plasticity andpathology in psychiatric disease: sites of action for potentialtherapy”, Current Opinion in Pharmacology 3, 33–40.
Srinivasarao, P., Vajreswari, A., Suryaprakash, P., Rupalatha, M.and Narayanareddy, K. (1997) “Lipid composition and fattyacid profile of myelin and synaptosomal membranes of ratbrain in response to the consumption of different fats”, TheJournal of Nutritional Biochemistry 8, 527–534.
Stoll, A.L., Locke, C.A., Marangel, L.B. and Severus, W.E. (1999)“Omega-3 fatty acids and bipolar disorder: a review”,Prostaglandins, Leukotrienes and Essential Fatty Acids 60,329–337.
Suarez, A., Faus, M.J. and Gil, A. (1996) “Dietary supplementationwith long-chain polyunsaturated fatty acids increasessusceptibility of weanling rat tissue lipids to in vitro lipidperoxidation”, The Journal of Nutritional Biochemistry 7,252–260.
K. NALIWAIKO et al.98

Tanskanen, A., Hibbeln, J.R., Tuomilehto, J., Uutela, A., Viinamaki,H., Lehtonen, J. and Vartiainen, E. (2001) “Fish consumptionand depressive symptoms in the general population inFinland”, Psychiatric Services 52, 529–531.
Turner, N., Else, P.L. and Hulbert, A.J. (2003) “Docosahexaenoicacid (DHA) content of membranes determines molecularactivity of the sodium pump: implications for diseases statesand metabolism”, Naturwissenschaften 90, 521–523.
Wainwright, P.E., Hunag, Y.S., Coscina, D.V., Levesque, S. andMcCutcheon, D. (1994) “Brain and behavioral effects ofdietary n-3 deficiency in mice: a three generation study”,Developmental Psychobiology 27(7), 467–487.
Wainright, P.E., Xing, H.C., Ward, G.R., Huang, Y.S., Bobik, H.,Auestad, N. and Montalto, M. (1999) “Water maze perform-ance is unaffected in artificially reared rats fed dietssupplemented with arachidonic acid and docosahexaienoicacid”, Journal of Nutrition 129, 1079–1089.
Wilde, M.C., Farkas, E., Gerrits, M., Kiliaan, A.J. and Luiten, P.G.M.(2002) “The effect of n-3 polyunsaturated fatty acid-richdiets on cognitive and cerebrovascular parameters inchronic cerebral hypoperfusion”, Brain Research 974, 166–173.
Wong, M.L. and Licinio, J. (2001) “Research and treatmentapproaches to depression”, Nature Reviews Neuroscience 2,343–351.
Yadid, G., Nakash, R., Deri, I., Grin, T., Kinor, N., Gispan, I. andZangen, A. (2000) “Elucidation of the neurobiology ofdepression: insights from a novel genetic animal model”,Progress in Neurobiology 62, 353–378.
Yonekubo, A., Honda, S., Okano, M., Takahashi, K. and Yamamoto,Y. (1993) “Dietary fish oil alters rat milk composition of fetaland neonatal rats”, Journal of Nutrition 123, 1703–1708.
Youdim, K.A., Martin, A. and Joseph, A.A. (2000) “Essentialfatty acids and the brain: possible health implications”,International Journal of Developmental Neuroscience 18, 383–399.
ANTIDEPRESSANT EFFECT OF FISH OIL 99

Copyright © 2022 FDOKUMEN




















